Embed Size (px)
Citation preview

Dreidimensionale Struktur von Proteinen (Voet Kapitel 7)
1. Sekundärstruktur2. Faserproteine3. Globuläre Proteine4. Protein Stabilisierung5. Quartärstruktur
U.Albrecht BC1

4. Protein-Stabilisierung
Proteine sind unter physiologischen Bedingungen stabil.Stabilisierungsenergie kommt von nichtkovalenten Kräften:
A. elektrostatische WechselwirkungenB. Wasserstoff-BrückenC. hydrophobe Kräfte
Proteinstruktur = Gleichgewicht von sich ausgelichenden Kräften
A. Elektrostatische Kräfte
Moleküle = Ansammlung von elektrisch geladenen PartikelnCoulomb-Gesetz:
U = k q1 q2
D rU = Anziehungskraftq1 q2 = elektrische Ladungenk = KonstanteD = Dielektrizitätskonstante Medium (=1 in Vakuum)r = Abstand zwischen Ladungen
Elektrostat. Kräfte wirken über grosseRäume schwierig Beitrag zur Stabilitätzu quantifizieren
U.Albrecht BC1

Ionische Wechselwirkungen sind stark, tragen aber nicht sehr zur Proteinstab. bei
Ionenpaare Salzbrücken
müssen jedoch solvatisiert werden. Solvatationsenergie ist etwa gleich wie die freieEnergie einer Ionenbrücken Bildung in nicht solvatisiertem Zustand. Netto Energieist 0. Beitrag von Ionenbrücken zur Stabilisierung von Proteinen gering.
U.Albrecht BC1

Dipol - Dipol - Wechselwirkungen sind schwach, tragen jedoch zur Stabilisierung bei
Nichtkovalente Anziehung zwischen elektr.neutralen Molekülen van der Waals Kräfte
Carbonyl- und Amidgruppen besitzen perma-nentes Dipolmoment = viel schwächereLadungen als Ionen. Anziehung nimmt mit r ab.
In Protein sind diese Dipole oft geordnetadditiv im Proteininneren (D niedrig) signifikante Stabilisierung
3
Diese Kräfte spielen nur bei kontaktierendenGruppen eine Rolle. Proportional zu r Bei grosser Anzahl interatomarer Kontakte wichtig zur Festlegung der Konformation
6
-0.3 kJ/mol
-9.3 kJ/mol
0.5 nm
U.Albrecht BC1

B. Wasserstoffbrücken-Bindungen
Donor - H . . . Akzeptor
N-H . . . O=C
H-Brücken stärker gerichtet als van derWaals Kräfte.
0.27 - 0.31 nm
-12 bis -30 kJ/mol
Protein ungefaltet, Akzeptor = Wasserd.h. Netto kein Energiegewinn gefaltet vs.ungefaltet.
Wenn Protein gefaltet würde ohne H-Brückenwürde freie Energie verloren gehen, d.h.H-Brücken bilden sich aus
U.Albrecht BC1

C. Hydrophobe Kräfte
Als hydrophoben Effekt bezeichnet man ein Phänomen das den Kontakt unpolarerSubstanzen mit Wasser minimiert.
Polare Lösungsmittel (DMSO, DMF) denaturieren Proteine
Orientierung von Wassermolekülen in Umgebung eines unpolaren gelösten Stoffes
Wassermoleküle überziehen unpolaren Stoffund maximieren ihre H-Brücken.
geordnete Struktur des Wassers isttreibende Kraft bei der Proteinfaltung
Solvatation von unpolaren Gruppen Verringerung des Wasservolumens
U.Albrecht BC1

Einschlussverbindung (Clathrat). Clathrate sind kristalline Komplexe unpolarerVerbindungen mit Wasser. Polyederkäfig bildet sich aus.
U.Albrecht BC1

Hydrophatie-Index = Mass für Hydrophobizität. Für jede AS anders. Negative Werte = hydrophile AS, positive Werte = hydrophobe AS
Hydrophatie-Plot des Chymotrypsinogens
externe Regionen
interne Regionen
U.Albrecht BC1

D. Disulfid-Bindungen (S-S)
Disulfid-Bindungen sind kovalente Bindungen welche die DreidimensionaleStruktur stabilisieren.
Cytoplasma hat reduziernden chemischen Charakter wo S-S Bindungen nicht stabilisierend wirken.
Extrazelluläre Umgebung ist oxidierend, deshalb haben Proteine die einen extrazellulären Bestimmungsort haben Disulfid-Brücken zur Stabilisierung
Im Labor werden Disulfid-Brücken mit reduzierenden Agenzien wie β−Mercaptoethanol geöffnet.
U.Albrecht BC1

E. Protein-Denaturierung
Native Proteinstruktur = kooperative Wechsel-wirkungen jede partielle Entfaltung desta-bilisiert Reststruktur
Temperatur beeiflusst kooperative WW starkTemp. wo Struktur zur Hälfte denaturiert=Schmelztemperatur TM. Thermophile Bakterien haben sehr hohe TM z.B. Taq Polymerase
pH-Variation ändert Ionisierung der AS (H-Brücken)
Detergenzien (z.B. Seifen) stören hydrophobe WW
wasserlösl. org. Substanzen stören hydrophobe WW (z.B. ethylenglycol, Saccharose)
optische Rotation von RNAse A als Funktionder Temperatur. Tm ist Wendepunkt der Kurve
U.Albrecht BC1

Einfluss von Salzen auf Proteine
Einige Salze stabilisieren andere de-stabilisieren Proteinstruktur
zum Aussalzen von Proteinen werdenvor allem stabilisierende Salze ver-wedet
Ionen die Protein-Denaturierung fördern bezeichnet man als chaotrop
Schmelztemperatur von RNAse A in Abhängig-keit von den Konzentrationen verschiedener Salze
Auch Harnstoff und das GuanidiniumIon denaturieren Proteine bei einer Konzentration von 5 M bis 10 M
U.Albrecht BC1

Dreidimensionale Struktur von Proteinen (Voet Kapitel 7)
1. Sekundärstruktur2. Faserproteine3. Globuläre Proteine4. Protein Stabilisierung5. Quartärstruktur
U.Albrecht BC1

5. Quartärstruktur
Polypeptid-Untereinheiten können in einer spezifischen Geometrie assoziieren.Die räumliche Anordnung dieser Untereinheiten wird als Quartärstruktur bezeichent.
Vorteile: Grosse Proteine können aus untereinheiten Bausatzmässig zusammengebaut werden. Effizienter als Verlängerung der Polypeptidkette. Auf Genom die Gene der Untereinheiten verteilt. Regulationsmöglichkeit beim Aufbau von
Komplexen.
4 Unterein- heiten =Protomere
U.Albrecht BC1

Symmetrie in Proteinen
In oligomeren Proteinen sind Protomeresymmetrisch angeordnet
Protomere formieren sich um einen Punkt Proteine nur rotationssym-metrisch. Keine Inversions- oder Spiegel-symmetrien, sonst müssten L-AS zuD-AS werden.
z.B. Virusproteine
U.Albrecht BC1

Präalbumin-Dimer entlang einer zweizähligen Achse (rot). Jedes Protomer auszwei 4 strängigen β−Faltblättern. Zwei dieser Dimere assoziieren Rücken an Rückenund bilden ein Tetramer mit D2-Symmetrie.
U.Albrecht BC1
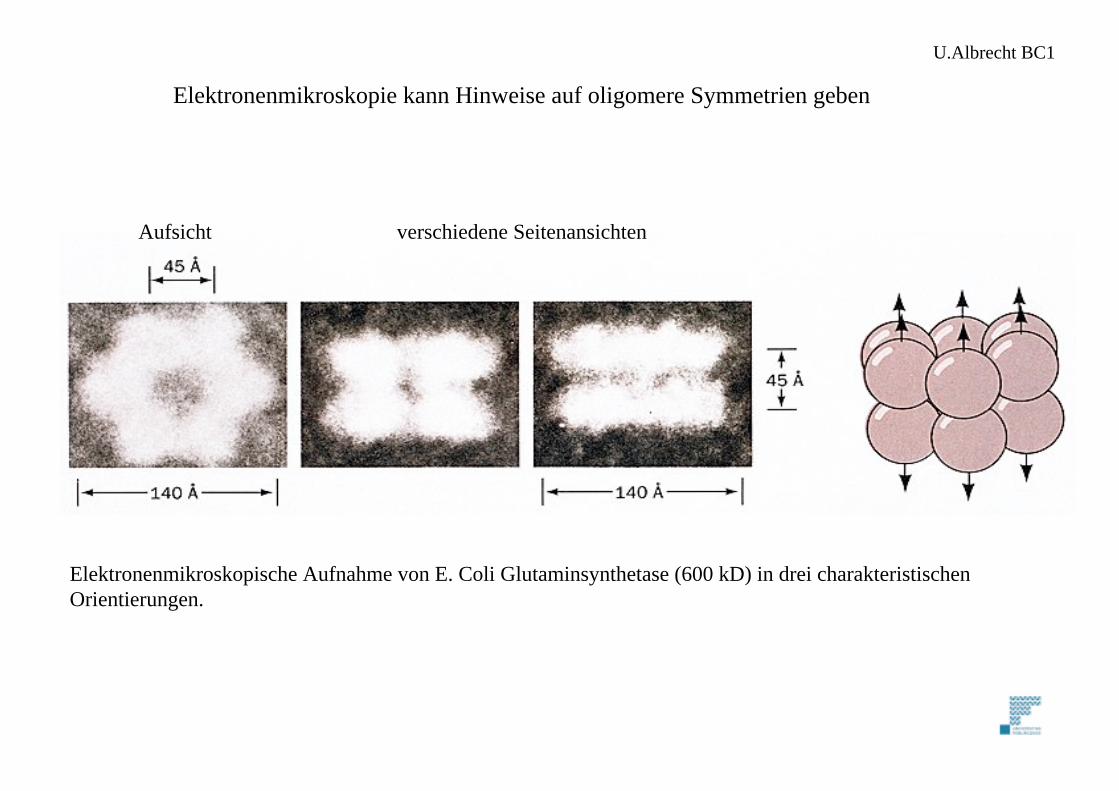
Elektronenmikroskopische Aufnahme von E. Coli Glutaminsynthetase (600 kD) in drei charakteristischenOrientierungen.
Aufsicht verschiedene Seitenansichten
Elektronenmikroskopie kann Hinweise auf oligomere Symmetrien geben
U.Albrecht BC1

Helicale Symmetrie
Einheit anden Endenleicht andereUmgebung
Umgebung für Einheiten quasiäquivalent
Viele strukturelle Proteine bildenFasern mit helicaler Symmetrie z.B. Muskel
U.Albrecht BC1

Bestimmung der Zusammensetzung der Untereinheiten
Hybridmoleküle liefern Information überQuartärstruktur
Lactatdehydrogenase (LDH) zwei Arten:M-Typ (Skelettmuskulatur)H-Typ (Herzmuskulatur)
wiederhohltes Einfrieren liefert 5 Isozymesichtbar durch elektrophoret. Auftrennung
Wenn Untereinheiten gleich, Veränderung einer UE durch chemische Modifikation, welche die Mobilität verändert.So wurde Quartärstruktur von E. ColiAspartat-Transcarbomoylase bestimmt
U.Albrecht BC1
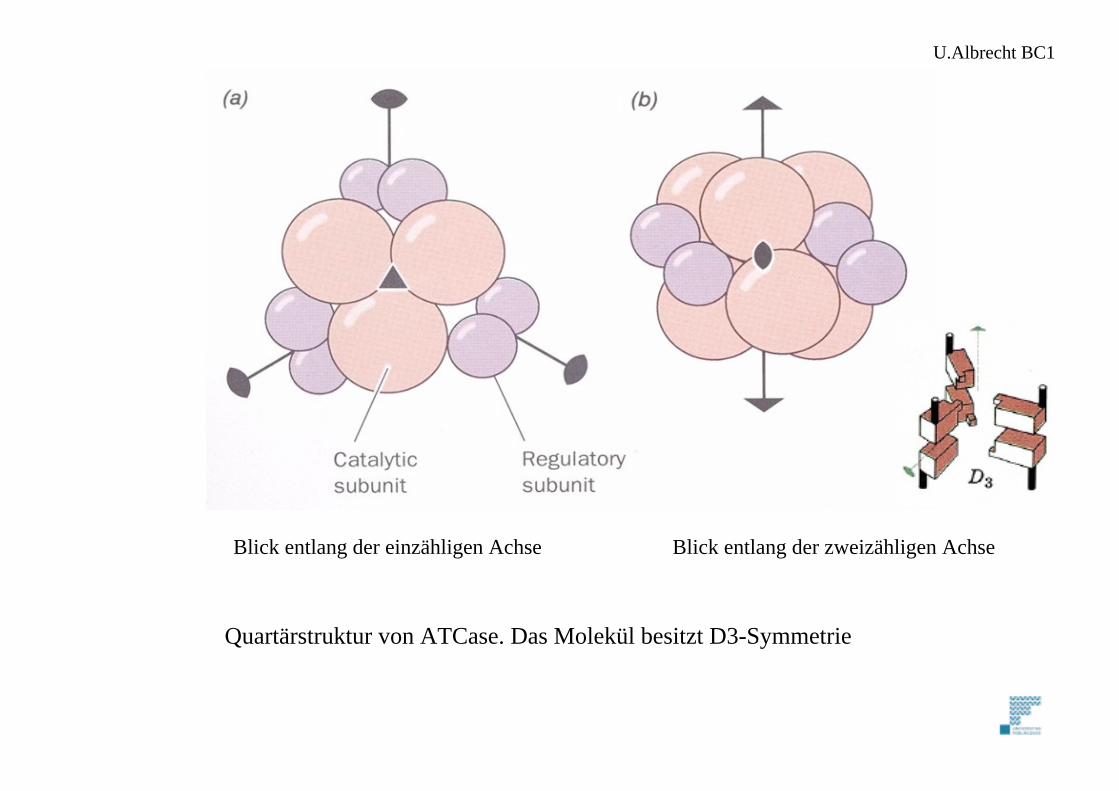
Quartärstruktur von ATCase. Das Molekül besitzt D3-Symmetrie
Blick entlang der einzähligen Achse Blick entlang der zweizähligen Achse
U.Albrecht BC1

Quervernetzende Agentien stabilisieren Oligomere
Vernknüpfung zweier Lys Seitenketten
Vergleich der Protomer Massemit der Masse nach Verknüpfungergibt Anzahl Untereinheiten.
Länge des Cross-Linkers kannAufschluss über die Abständezwischen Untereinheiten geben
5C
8C
U.Albrecht BC1