Embed Size (px)
Citation preview

1
REVISED MANUSCRIPT (Ver. 3) 1
2
Submitted to Appl. Environ. Microbiol. 3
(Biotechnology section) 4
5
Construction and application of a functional library of cytochrome P450 6
monooxygenases from the filamentous fungus Aspergillus oryzae 7
8
Running title: Functional screening of Aspergillus oryzae cytochrome P450 9
10
11
KHM Nazmul Hussain Nazir1, Hirofumi Ichinose
1*, and Hiroyuki Wariishi
1, 2, 3 12
13
14
1Faculty of Agriculture, Kyushu University, 6-10-1 Hakozaki, Higashi-ku, 15
Fukuoka 812-8581, Japan. 16
2Bio-Architecture Center, Kyushu University, 6-10-1 Hakozaki, Higashi-ku, 17
Fukuoka 812-8581, Japan. 18
3Innovation Center for Medical Redox Navigation, Kyushu University, 6-10-1 Hakozaki, 19
Higashi-ku, 20
Fukuoka 812-8581, Japan. 21
22
23
*Corresponding author: Hirofumi Ichinose 24
Faculty of Agriculture, Kyushu University 25
6-10-1 Hakozaki, Fukuoka 812-8581 Japan 26
Tel & Fax, +81-92-642-2994; E-mail, [email protected] 27
Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.02491-10 AEM Accepts, published online ahead of print on 4 March 2011
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

2
ABSTRACT 1
A functional library of cytochrome P450 monooxygenases from Aspergillus oryzae 2
(AoCYPs) was constructed, in which 121 isoforms were co-expressed with yeast 3
NADPH-cytochrome P450 oxidoreductase in Saccharomyces cerevisiae. Using this 4
functional library, novel catalytic functions of AoCYPs, such as catalytic potentials of 5
CYP57B3 against genistein, were elucidated for the first time. Comprehensive 6
functional screening promises rapid characterization of catalytic potentials and utility of 7
AoCYPs. 8
9
Key words: Cytochrome P450; Aspergillus oryzae; catalytic function; genistein; 10
functional screening 11
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

3
Cytochromes P450 (CYPs) constitute a large superfamily of heme-containing 1
monooxygenases, that are distributed among a wide variety of organisms. An 2
evolutionary history of extensive divergence from a common ancestor is implied (14, 20, 3
28). The majority of CYPs are highly specialized and play crucial roles in meeting the 4
metabolic requirements of particular organisms. They are involved in secondary 5
metabolic pathways such as detoxification of xenobiotics and synthesis of secondary 6
metabolites (9, 30). Over the last few years, the sequence database of CYPs has greatly 7
enlarged, and continues to increase (27, 31). A compilation of CYP sequences would 8
facilitate understanding of metabolic diversity and evolutionary history among living 9
organisms. In addition to their biological importance, the catalytic functions of CYPs 10
are of great interest to those involved in biotechnology (4, 5, 8, 13, 16, 42). Biocatalytic 11
regiospecific and stereospecific oxidations by CYPs attract much attention in industry 12
because (i) use of biocatalysts allows reduction in the number of tedious steps such as 13
blocking and deblocking that are common in conventional chemical synthesis, (ii) a 14
single product without by-product formation reduces the cost of downstream 15
purification steps, and (iii) enantiomeric mixtures (rather than racemic mixtures) are 16
strictly required in some fields such as the pharmaceutical industry (12, 33, 39, 42). 17
Therefore, a rational comprehensive approach towards elucidating catalytic potentials 18
and utilities of CYPs is required to develop bioindustrial applications (8, 19, 34). 19
A wide variety of filamentous fungi are used to produce economically valuable 20
consumer items. The filamentous fungus Aspergillus oryzae is one of the most widely 21
utilized microorganisms. It has been used for more than 1000 years in Japanese 22
fermentation industries to produce indigenous products such as sake (rice wine), miso 23
(soybean paste), and shoyu (soy sauce). Besides traditional fermentation technologies, 24
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

4
much recent research into A. oryzae has focused on production of recombinant enzymes 1
and primary and secondary metabolites (1, 40). The whole genome of A. oryzae has 2
been sequenced and is available to the public (21). Genomic data have potential to 3
provide a more thorough understanding of metabolic diversity and capability of A. 4
oryzae. Recently, we identified 155 genes including 13 putative pseudogenes of A. 5
oryzae CYPs (AoCYPs), and cloned 121 full-length cDNAs encoding an open reading 6
frame (25). Molecular and functional diversity of AoCYPs is presumably important to 7
the metabolic capability of this fungus; however, biological functions and catalytic 8
potentials remain obscure. A rational approach to find and exploit catalytic potentials of 9
various CYP enzymes would facilitate a myriad of biocatalytic processes. In this study, 10
we have aimed to develop a functional screening system for AoCYPs and to increase 11
understanding of their functional information. 12
Heterologous expression of AoCYPs. Using the isolated cDNAs, we attempted 13
to express recombinant enzymes for a functional screening system of AoCYPs. For 14
catalytic activity, CYPs must be associated with electron transfer systems providing 15
reducing equivalents. Thus, systematic production of both CYPs and their redox 16
partner(s) would be important to facilitate comprehensive functional screening (19, 34). 17
In this study, we employed S. cerevisiae as a host strain for heterologous expression 18
because (i) a yeast expression system seemed appropriate to obtain recombinant 19
eukaryotic CYP without genetic engineering of cDNA, (ii) S. cerevisiae has only three 20
CYP genes and their catalytic functions are well characterized (26), and (iii) S. 21
cerevisiae is potentially useful for industrial applications as well as basic research. In 22
addition, cytochrome P450 oxidoreductase (CPR) from S. cerevisiae is capable of 23
donating reducing equivalents for various CYPs. It has been shown that a wide variety 24
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

5
of eukaryotic CYPs including Aspergillus spp. could be functional with CPR from S. 1
cerevisiae (3, 23, 24, 32). Presumably, eukaryotic CYPs can exhibit catalytic activities 2
with a heterologous CPR. Therefore, we constructed a yeast co-expression system of 3
AoCYP and yeast CPR using pGYR vector (24, 35, 36; see also the supplemental 4
material and methods). To elucidate heterologous expression of AoCYPs, we analyzed 5
the CO-difference spectrum of S. cerevisiae cells harboring the expression plasmid (29). 6
Figure 1 depicts a CO-difference spectrum of CYP628C1 expressed in S. cerevisiae. No 7
peak was observed around 450 nm from a transformant harboring a plasmid without an 8
AoCYP, indicating that the recombinant AoCYP enzyme was successfully expressed in 9
S. cerevisiae in an active form. These results also implied that endogenous CYPs of S. 10
cerevisiae did not interfere with spectroscopic analysis of recombinant AoCYPs due to 11
their low levels of expression in this host strain (2). We have confirmed significant 12
expression of 84 AoCYPs based upon CO-difference spectra. 13
Comprehensive functional screening of AoCYPs. To develop a functional 14
screening system, each transformant was separately inoculated into 0.5 mL culture 15
medium and grown in 96 DeepWell plates (supplemental materials and methods). 16
Transformants accommodated in the 96-well plates were easily replicated and used for 17
further experiments. To validate potential utility of the screening system, we initiated a 18
functional survey using 7-ethoxycoumarin as a tentative model substrate. After 2 days 19
incubation, significant product formation was achieved by S. cerevisiae harboring 20
expression plasmids of CYP57B3, CYP62C2, CYP68Q1, CYP531E1, CYP620G1, 21
CYP620H1, CYP620H3, CYP620H9, CYP628C1, CYP675A2, CYP5061B5, 22
CYP5078A5, and CYP5080B2. The product was identified as 7-hydroxycoumarin by 23
comparison of the retention time on HPLC with an authentic standard (Fig. 2). 24
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

6
Moreover, several AoCYPs such as 5061B5 yielded minor unidentified product(s), in 1
addition to the formation of 7-hydroxycoumarin (Fig. 2). Although further information 2
should aim to elucidate their biochemical and biophysical properties, it can be 3
hypothesized that the natural substrate(s) of these AoCYPs might be structurally related 4
to coumarin, such as polyketide derivatives. It is thus a good example that 5
non-target-driven screening would provide new insight into the fascinating biology and 6
metabolic processes of A. oryzae. Interestingly, catalytic conversion of 7
7-ethoxycoumarin to 7-hydroxycoumarin was clearly demonstrated by the transformant 8
harboring CYP5080B2 expression plasmid; nevertheless, we could not confirm its 9
expression by CO-difference spectrum. Such observation was also found from several 10
AoCYPs using different substrates. Combining the results of spectroscopic analysis and 11
bioconversion, we could conclude that at least 92 AoCYPs were functionalized in S. 12
cerevisiae. These results highlight the potential utility of our screening system. 13
Using the functional screening system, we further carried out targeted functional 14
screening of AoCYP to produce value-added isoflavonoids. Genistein is one of the 15
major isoflavonoids found in soybean. Because of its biological and pharmacological 16
activities, genistein derivatives are likely to have various useful applications (6, 10, 11, 17
15, 17, 41). When genistein was utilized as a substrate, significant product formations 18
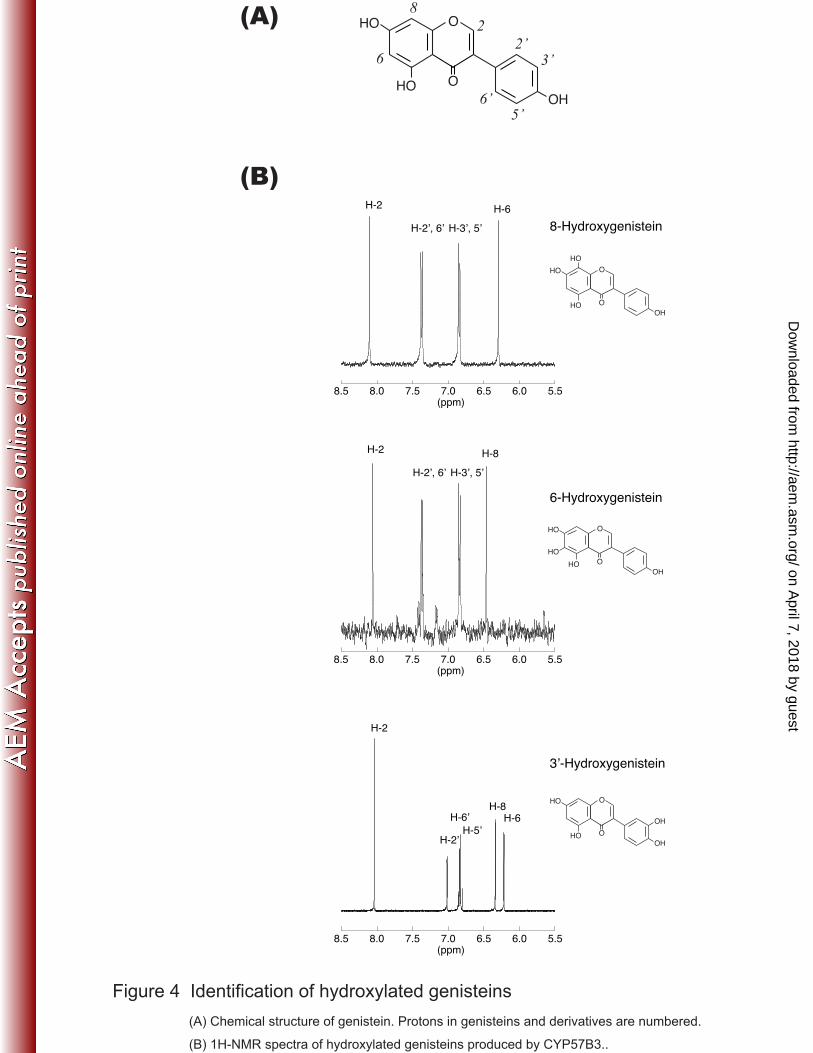
were achieved by S. cerevisiae expressing CYP57B3 (Fig. 3). Incorporation of a 19
hydroxyl group was clearly demonstrated by LC-ESI-MS analysis (data not shown). 20
Based upon 1H-NMR spectroscopic analysis, products were identified as 8-hydroxy-, 21
6-hydroxy-, and 3’-hydroxygenistein (Fig. 4). To the best of our knowledge, this is the 22
first report describing catalytic activities of CYP57B3 against genistein and suggesting 23
its possible roles in production of hydroxygenistein during soybean fermentation 24
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

7
processes (6, 11). Based on sequence comparison, CYP57B3 seems phylogenetically 1
close to a CYP from the plant pathogen Nectria haematococca; a fungus involved in 2
biodegradation of phytoalexin pisatin (22). Although it is still unclear whether 3
CYP57B3 exhibits catalytic activities against pisatin, it may play a biological role in the 4
metabolism of phytoalexins with isoflavone skeletons. Biological and pharmacological 5
potentials of the products have also been reported. 8-Hydroxygenistein shows much 6
stronger antioxidative activity than genistein (11) and exhibits antiproliferative activity 7
against cancer cells (15). 3’-Hydroxygenistein, known as orobol, exhibits unique 8
pharmacological activities such as enhancing sensitivity of human ovarian carcinoma 9
cells against the anti-cancer drug paclitaxel (17), and inhibiting human 10
immunodeficiency virus 1 integrase (41). Although hydroxylated genistein can be 11
isolated from natural products including fermented products, these natural compounds 12
are limited in supply and problematic for practical utilization. Because the synthesis of 13
isoflavones remains an important object (10, 37), it would be of great interest to utilize 14
CYP57B3 for production of value-added rare isoflavonoids from genistein. However, 15
further research efforts should aim to improve conversion efficiency for practical 16
application. It has been reported that reaction efficiency can be influenced by redox 17
partner(s) such that native combination of CYP-CPR from a same organism shows 18
slightly better functionality than heterologous combination and several monooxygenase 19
reactions are coupled with cytochrome b5 (7, 18, 30, 38). Further investigation and 20
possible re-engineering of CYP57B3 and its redox partner(s) should facilitate 21
systematic understanding for the biotechnology sector. 22
Through functional screening, the novel activities of AoCYPs were experimentally 23
explored using a series of compounds such as flavonoids, terpenoids, steroids, and 24
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

8
pharmaceutical chemicals (Table). To the best of our knowledge, this is the first report 1
experimentally demonstrating functional diversity of Aspergillus CYP. The functional 2
screening system promises rapid and comprehensive characterization of the catalytic 3
potential of AoCYPs, which would further provide novel insight into the biology and 4
biotechnology. In this study, we have focused on the catalytic capability of CYP57B3 to 5
produce value-added hydroxylated products from genistein. However, the screening 6
system should be applicable for a myriad of compounds, and facilitate investigation in 7
industrial and biological fields. Thorough understanding of AoCYP functions will open 8
the door for advanced fungal biology and biotechnology. 9
10
This research was supported in part by a Noda Institute for Scientific Research 11
(NIRS) Research Grant (to H. I.). 12
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

9
REFERENCES 1
1. Abe, K., K. Gomi, F. Hasegawa, and M. Machida. 2006. Impact of Aspergillus 2
oryzae genomics on industrial production of metabolites. Mycopathologia 3
162:143–153. 4
2. Akiyoshi-Shibata, M., E. Usui, T. Sakaki, Y. Yabusaki, M. Noshiro, K. Okuda, 5
and H. Ohkawa. 1991. Expression of rat liver vitamin D3 25-hydroxylase cDNA in 6
Saccharomyces cerevisiae. FEBS Lett. 280:367-70. 7
3. Artigot, M. P., N. Loiseau, J. Laffitte, L. Mas-Reguieg, S. Tadrist, I. P. Oswald, 8
and O. Puel. 2009. Molecular cloning and functional characterization of two 9
CYP619 cytochrome P450s involved in biosynthesis of patulin in Aspergillus 10
clavatus. Microbiol. 155:1738-1747. 11
4. Bernhardt, R. 2006. Cytochromes P450 as versatile biocatalysts. J. Biotechnol. 12
124:128-145. 13
5. Chang, M. C. Y., R. A. Eachus, W. Trieu, D.-K. Ro, and J. D. Keasling. 2007. 14
Engineering Escherichia coli for production of functionalized terpenoids using plant 15
P450s. Nat. Chem. Biol. 3:274-277. 16
6. Chang, T.-S., H.-Y. Ding, S. S.-K. Tai, and C.-Y. Wu. 2007. Metabolism of the soy 17
isoflavones daidzein and genistein by fungi used in the preparation of various 18
fermented soybean foods. Biosci. Biotechnol. Biochem. 71:1330-1333. 19
7. Chemler, J. A., C. G. Lim, J. L. Daiss, and M. A. G. Koffas. 2010. A versatile 20
microbial system for biosynthesis of novel polyphenols with altered estrogen 21
receptor binding activity. Chem. Biol. 17:392-401. 22
8. Chigu, N. L., S. Hirosue, C. Nakamura, H. Teramoto, H. Ichinose, and H. 23
Wariishi. 2010. Cytochrome P450 monooxygenases involved in anthracene 24
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

10
metabolism by the white-rot basidiomycete Phanerochaete chrysosporium. Appl. 1
Microbiol. Biotechnol. 87:1907-1916. 2
9. Demain, A. L., and A. Fang. 2000. The natural functions of secondary metabolites. 3
Adv. Biochem. Eng. Biotechnol. 69:1–39. 4
10. Dixon, R. A., and D. Ferreira. Genistein. 2002. Phytochem. 60:205-211. 5
11. Esaki, H., H. Onozaki, Y. Morinitsu, S. Kawakishi, and T. Osawa. 1998. Potent 6
antioxidant isoflavones isolated from soybeans fermented with Aspergillus saitoi. 7
Biosci. Biotechnol. Biochem. 62:740-746. 8
12. Gavrilescu, M., and Y. Chisti. 2005. Biotechnology—a sustainable alternative for 9
chemical industry. Biotechnol. Adv. 23:471-479. 10
13. Gillam, E. M. J. 2008. Engineering cytochrome P450 enzymes. Chem. Res. 11
Toxicol. 21:220-231. 12
14. Gotoh, O. 1993. Evolution and differentiation of P450 genes. p. 255-272, In Omura, 13
T. Ishimura, Y. Fujii-Kuriyama, Y. (ed.), Cytochrome P-450 2nd
ed.. Kodansha, 14
Tokyo. 15
15. Hirota, A., S. Taki, S. Kawaii, M. Yano, and N. Abe. 2000. 16
1,1-Diphenyl-2-picrylhydrazyl radical-scavenging compounds from soybean miso 17
and antiproliferative activity of isoflavones from soybean miso toward the cancer 18
cell lines. Biosci. Biotech. Biochem. 64:1038-1040. 19
16. Ichinose, H., H. Wariishi, and H. Tanaka. 1999. Biotransformation of recalcitrant 20
4-methyldibenzothiophene to water-extractable products using lignin-degrading 21
basidiomycete Coriolus versicolor. Biotechnol. Prog. 15:706–714. 22
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

11
17. Isonishi, S., M. Saitou, M. Saitou, M. Yasuda, and T. Tanaka. 2007. Differential 1
regulation of the cytotoxicity activity of paclitaxel by orobol and platelet derived 2
growth factor in human ovarian carcinoma cells. Oncol. Rep. 18:195-201. 3
18. Lamb, D. C., D. E. Kelly, N. J. Manning, M. A. Kaderbhai, and S. L. Kelly. 4
1999. Biodiversity of the P450 catalytic cycle: yeast cytochrome b5/NADH 5
cytochrome b5 reductase complex efficiently drives the entire sterol 6
14-demethylation (CYP51) reaction. FEBS Lett. 462:283-288. 7
19. Lamb, D. C., T. Skaug, H. L. Song, C. J. Jackson, L. M. Podust, M. R. 8
Waterman, D. B. Kell, D. E. Kelly, and S. L. Kelly. 2002. The cytochrome P450 9
complement (CYPome) of Streptomyces coelicolor A3(2). J. Biol. Chem. 10
277:24000-24005. 11
20. Lewis, D. F. L., E. Watson, and B. G. Lake. 1998. Evolution of the cytochrome 12
P450 superfamily: sequence alignments and pharmacogenetics. Mutat. Res. 13
410:245-270. 14
21. Machida, M., K, Asai, M. Sano, T. Tanaka, T. Kumagai, G. Terai, K. 15
Kusumoto, T. Arima, O. Akita, Y. Kashiwagi, K. Abe, K. Gomi, H. Horiuchi, K. 16
Kitamoto, T. Kobayashi, M. Takeuchi, D. W. Denning, J. E. Galagan, W. C. 17
Nierman, J. Yu, D. B. Archer, J. W. Bennett, D. Bhatnagar, T. E. Cleveland, N. 18
D. Fedorova, O. Gotoh, H. Horikawa, A. Hosoyama, M. Ichinomiya, R. 19
Igarashi, K. Iwashita, P. R. Juvvadi, M. Kato, Y. Kato, T. Kin, A. Kokubun, H. 20
Maeda, N. Maeyama, J. Maruyama, H. Nagasaki, T. Nakajima, K. Oda, K. 21
Okada, I. Paulsen, K. Sakamoto, T. Sawano, M. Takahashi, K. Takase, Y. 22
Terabayashi, J. R. Wortman, O. Yamada, Y. Yamagata, H. Anazawa, Y. Hata, 23
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

12
Y. Koide, T. Komori, Y. Koyama, T. Minetoki, S. Suharnan, A. Tanaka, K. 1
Isono, S. Kuhara, N. Ogasawara, and H. Kikuchi. 2005. Genome sequencing and 2
analysis of Aspergillus oryzae. Nature 438:1157–1161. 3
22. Maloney, A. P., and H. D. VanEtten. 1994. A gene from the fungal plant pathogen 4
Nectria haematococca that encodes the phytoalexin-detoxifying enzyme pisatin 5
demethylase defines a new cytochrome P450 family. Mol. Gen. Genet. 6
243:506-514. 7
23. Martel, C. M., J. E. Parker, A. G. Warrilow, N. J. Rolley, S. L. Kelly, and D. E. 8
Kelly. 2010. Complementation of a Saccharomyces cerevisiae ERG11/CYP51 9
(sterol 14a-demethylase) doxycycline-regulated mutant and screening of the azole 10
sensitivity of Aspergillus fumigatus isoenzymes CYP51A and CYP51B. Antimicrob. 11
Agents Chemother. 54:4920-4923. 12
24. Murakami, H., Y. Yabusaki, T. Sakaki, M. Shibata, and H. Ohkawa. 1990. 13
Expression of cloned yeast NADPH-cytochrome P450 reductase gene in 14
Saccharomyces cerevisiae. J. Biochem. 108:859-865. 15
25. Nazir, K. H. M. N. H., H. Ichinose,
and H. Wariishi. 2010. Molecular 16
characterization and isolation of cytochrome P450 genes from the filamentous 17
fungus Aspergillus oryzae. Arch. Microbiol. 192: 395-408. 18
26. Nelson, D. R. 1999. Cytochrome P450 and the individuality of species. Arch. 19
Biochem. Biophys. 369:1-10. 20
27. Nelson, D. R. 2009. The cytochrome p450 homepage. Hum. Genomics. 4:59-65. 21
28. Nelson, D. R., T. Kamataki, D. J. Waxman, F. P. Guengerich, R. W. Estabrook, 22
R. Feyereisen, F. J. Gonzalez, M. J. Coon, I. C. Gunsalus, O. Gotoh, K. Okuda, 23
and D. W. Nebert. 1993. The P450 superfamily-update on new sequences, gene 24
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

13
mapping, accession numbers, early trivial names of enzymes, and nomenclature. 1
DNA Cell Biol. 12:1-51. 2
29. Omura, T. and R. Sato. 1964. The carbon monoxide-binding pigment of liver 3
microsomes, II. Solubilization, purification, and properties. J. Biol. Chem. 4
239:2379-2385. 5
30. Ortiz de Montellano, P. R. 2005. Cytochrome P450: structure, mechanism, and 6
biochemistry, 3rd ed. Kluwer Academic/Plenum, New York. 7
31. Park, J., B. Park, K. Jung, S. Jang, K. Yu, J. Choi, S. Kong, J. Park, S. Kim, H. 8
Kim, S. Kim, J. F. Kim, J. E. Blair, K. Lee, S. Kang, and Y. H. Lee. 2008. 9
CFGP: a web-based, comparative fungal genomics platform. Nucleic Acids Res. 36: 10
D562–D571. 11
32. Renaud, J. P., M. A. Peyronneau, P. Urban, G. Truan, C. Cullin, D. Pompon, 12
Ph. Beaune, and D. Mansuy. 1993. Recombinant yeast in drug metabolism. 13
Toxicol. 82:39-52. 14
33. Ro, D. K., E. M. Paradise, M. Ouellet, K. J. Fisher, K. L. Newman, J. M. 15
Ndungu, K. A. Ho, R. A. Eachus, T. S. Ham, J. Kirby, M. C. Chang, S. T. 16
Withers, Y. Shiba, R. Sarpong, and J. D. Keasling. 2006. Production of the 17
antimalarial drug precursor artemisinic acid in engineered yeast. Nature. 18
440:940-943. 19
34. Sabbadin, F., R. Hyde, A. Robin, E. M. Hilgarth, M. Delenne, S. Flitsch, N. 20
Turner, G. Grogan, and N. C. Bruce. 2010. LICRED: a versatile drop-in vector 21
for rapid generation of redox-self-sufficient cytochrome P450s. Chembiochem 22
11:987-994. 23
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

14
35. Sakaki, T., M. Akiyoshi-Shibata, Y. Yabusaki, and H. Ohkawa. 1992. 1
Organella-targeted expression of rat liver cytochrome P450c27 in yeast: genetically 2
engineered alteration of mitochondrial P450 into a microsomal form creates a novel 3
functional electron transport chain. J. Biol. Chem. 267:16497-16502. 4
36. Sakaki, T., R. Shinkyo, T. Takita, M. Ohta, K. Inouye. 2002. Biodegradation of 5
polychlorinated dibenzo-p-dioxins by recombinant yeast expressing rat CYP1A 6
subfamily. Arch. Biochem. Biophys. 410:91-98. 7
37. Sato S., K. Hiroe, T. Kumazawa, and J.-I. Onodera. 2006. Total synthesis of two 8
isoflavone C-glycosides: genistein and orobol 8-C-beta-D-glucopyranosides. 9
Carbohydr. Res. 341:1091-1095. 10
38. Schenkman, J. B., and I. Jansson. 2003. The many roles of cytochrome b5. 11
Pharmacol. Ther. 97:139-152. 12
39. Schmid, A., J. S. Dordick, B. Hauer, A. Hauer, M. Wubbolts, and B. Witholt. 13
2001 Industrial biocatalysis today and tomorrow. Nature 409:258-268 14
40. Tailor, M. J., and T. Richardson. 1979. Applications of microbial enzymes in 15
food systems and in biotechnology. Adv. Appl. Microbiol. 25:7–35. 16
41. Tewtrakul, S., S. Subhadhirasakul, S. Cheenpracha, and C. Karalai. 2007. 17
HIV-1 protease and HIV-1 integrase inhibitory substance from Eclipta prostrata. 18
Phytother. Res. 21:1092-1095. 19
42. Urlacher, V. B., and S. Eiben. 2006. Cytochrome P450 monooxygenases: 20
perspectives for synthetic application. Trends. Biotechnol. 24:324-330. 21
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

15
FIGURE LEGENDS 1
Figure 1 CO-difference spectrum of CYP628C1 expressed in S. cerevisiae 2
3
Figure 2 Screening of AoCYPs catalyzing 7-ethoxycoumarin conversions 4
Metabolic products observed from S. cerevisiae expressing no AoCYP (control), 5
CYP620H9, CYP5080B2, and CYP5061B5, analyzed by HPLC at 320 nm. Arrows (in 6
CYP5061B5) indicate unidentified products. The chromatogram for CYP5061B5 is 7
plotted as a 10-fold expanded scale. 8
9
Figure 3 HPLC analysis of genistein conversion catalyzed by CYP57B3 10
Metabolic products observed from S. cerevisiae expressing no AoCYP (control) and 11
CYP57B3 (CYP57B3), analyzed by HPLC at 260 nm. 8HOG, 6HOG, 3’HOG indicate 12
8-hydroxy-, 6-hydroxy-, and 3’-hydroxygenistein, respectively. 13
14
Figure 4 Identification of hydroxylated genisteins 15
(A) Chemical structure of genistein. Protons in genisteins and derivatives are numbered. 16
(B) 1H-NMR spectra of hydroxylated genisteins produced by CYP57B3. 17
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

Table Catalytic Potentials of AoCYPs against various compounds
Substrate AoCYP
7-Ethoxycoumarin CYP57B3, CYP62C2, CYP68Q1, CYP531E1, CYP620G1, CYP620H1, CYP620H3, CYP620H9,
CYP628C1, CYP675A2, CYP5061B5, CYP5078A5, CYP5080B2.
Genistein CYP57B3
Naringenin(1)
CYP57B3, CYP62C2
Testosterone(2)
CYP65T4, CYP547C3, CYP595B1, CYP5061B5
Dehydroabietic acid(3)
CYP5061B5
Diclofenac(4)
CYP65AD1, CYP68Q1, CYP65AF1
(1) HPLC analysis of naringenin conversion are shown in Figure S2.
(2) HPLC analysis of testosterone conversion are shown in Figure S3.
(3) HPLC analysis of dehydroabietic aicd conversion are shown in Figure S4.
(4) HPLC analysis of diclofenac conversion are shown in Figure S5.
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

Wavelength (nm)
400 450 500
A = 0.005
Ab
so
rba
nce
Figure 1 CO-difference spectrum of CYP628C1 expressed in S. cerevisiae
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

1.0
Retention Time (min)
Inte
nsity (
32
0 n
m)
Control
2.0 2.5 3.0 3.51.5
CYP620H9
CYP5080B2
CYP5061B5
(x10)
7-e
tho
xyco
um
arin
7-h
yd
roxyco
um
arin
7-h
yd
roxyco
um
arin
7-h
yd
roxyco
um
arin
Figure 2 Screening of AoCYPs catalyzing 7-ethoxycoumarin conversions
Metabolic products observed from S. cerevisiae expressing no AoCYP (control), CYP620H9, CYP5080B2, and
CYP5061B5, analyzed by HPLC at 320 nm. Arrows (in CYP5061B5) indicate unidentified products.
The chromatogram for CYP5061B5 is plotted as a 10-fold expanded scale.
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from

1.5 2.0 2.5 3.0
Retention Time (min)
Inte
nsity (
26
0 n
m)
Ge
nis
tein
Ge
nis
tein
8H
OG
6H
OG
3’
HO
G
Control
CYP57B3
Figure 3 HPLC analysis of genistein conversion catalyzed by CYP57B3
Metabolic products observed from S. cerevisiae expressing no AoCYP (control) and CYP57B3 (CYP57B3),
analyzed by HPLC at 260 nm. 8HOG, 6HOG, 3’ HOG indicate 8-hydroxy-, 6-hydroxy-, and
3’ -hydroxygenistein, respectively.
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from
O
O
OHHO
HO8
6
2
2’
3’
6’
5’
8.5 8.0 7.5 7.0 6.5 6.0 5.5(ppm)
H-2
H-2’H-5’
H-8H-6H-6’
3’-Hydroxygenistein
O
O
OHHO
HO
OH
8.5 8.0 7.5 7.0 6.5 6.0 5.5(ppm)
H-2
H-2’, 6’ H-3’, 5’
H-6
8-Hydroxygenistein
O
O
OHHO
HO
HO
8.5 8.0 7.5 7.0 6.5 6.0 5.5(ppm)
H-2
H-2’, 6’ H-3’, 5’
H-8
6-Hydroxygenistein
O
O
OHHO
HO
HO
(A)
(B)
Figure 4 Identification of hydroxylated genisteins
(A) Chemical structure of genistein. Protons in genisteins and derivatives are numbered.
(B) 1H-NMR spectra of hydroxylated genisteins produced by CYP57B3..
on April 7, 2018 by guest
http://aem.asm
.org/D
ownloaded from





























