Embed Size (px)
Citation preview

1
Cell Surface Heparan Sulfate Proteoglycans Potentiate Chordin’s Antagonism of
BMP signaling and Participate in Cellular Uptake of Chordin*
Reema Jasuja‡§, Benjamin L. Allen¶§||, William N. Pappano§||**,
Alan C. Rapraeger‡¶‡‡§§, and Daniel S. Greenspan‡¶**‡‡§§
Programs in ‡Molecular and Cellular Pharmacology and ¶Cellular and Molecular
Biology, **Department of Biomolecular Chemistry, and ‡‡Department of Pathology and
Laboratory Medicine, University of Wisconsin
1300 University Avenue, Madison, WI, 53706, USA
§These authors contributed equally to this work.
||Predoctoral fellows of the American Heart Association.
§§To whom correspondence should be addressed: Department of Pathology and
Laboratory Medicine, University of Wisconsin, 1300 University Avenue, Madison, WI,
53706
D.S.G. Tel.: 608-262-4676; Fax: 608-262-6691; E-mail: [email protected].
JBC Papers in Press. Published on September 20, 2004 as Manuscript M408129200
Copyright 2004 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2
A.C.R. Tel.: 608-262-7577; Fax: 608-265-3301; E-mail: [email protected]
*This work was supported by National Institutes of Health Grants AR47746, GM63471
(to D.S.G.) and GM48850 (to A.C.R.)
Running Title:
HSPGs Regulate Chordin Function, Uptake and Diffusion
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3
SUMMARY
Signaling by bone morphogenetic proteins (BMPs) plays central roles in early
embryonic patterning, organogenesis and homeostasis in a broad range of species.
Chordin, an extracellular antagonist of BMP signaling, is thought to readily diffuse in
tissues, thus forming gradients of BMP inhibition that result in reciprocal gradients of
BMP signaling. The latter determine cell fates along the embryonic dorsoventral axis.
The secreted protein Twisted Gastrulation is thought to help shape BMP signaling
gradients by acting as a cofactor that enhances Chordin’s inhibition of BMP signaling.
Here we demonstrate that mammalian Chordin binds heparin with an affinity similar to
that of factors known to functionally interact with heparan sulfate proteoglycans (HSPGs)
in tissues. We further demonstrate that Chordin binding in mouse embryonic tissues is
dependent upon its interaction with cell surface HSPGs and that Chordin binds to cell
surface HSPGs (e.g. syndecans), but not to basement membranes containing the HSPG
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4
perlecan. Surprising, mammalian twisted gastrulation does not bind heparin unless
prebound to Chordin and/or BMP-4, although Drosophila Twisted gastrulation has been
reported to bind heparin on its own. Importantly, results are also presented which
indicate that Chordin-HSPG interactions strongly potentiate the antagonism of BMP
signaling by Chordin and are necessary to the retention and uptake of Chordin by cells.
These data and others regarding Chordin diffusion have implications for the paradigm of
how Chordin is thought to regulate BMP signaling in the extracellular space and how
gradients of BMP signaling are formed.
Transforming growth factor-β1 (TGF-β)-like bone morphogenetic proteins
(BMPs) were first isolated from osteogenic extracts of bone, and some BMPs are capable
of inducing ectopic bone formation (1-3). In addition to roles in bone and cartilage
formation, BMPs are involved in morphogenesis of various soft tissues and organs during
development, and play central roles in dorsoventral patterning in early embryogenesis (4).
Dorsoventral patterning in Xenopus requires BMPs 2, 4, and 7 for specification of ventral
mesodermal and ectodermal cell fates (5-7), while specification of dorsal mesoderm
(Spemann’s organizer) and neurectoderm requires inhibition of BMP signaling. Dorsal
inhibition of BMP signaling is achieved by secreted BMP antagonists such as Chordin
and Noggin, which are expressed in Spemann’s organizer (for a review, see Ref. 8). In
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5
Drosophila, dorsoventral patterning of the embryonic axis is mediated by BMP-2/4
homologue Decapentaplegic (DPP) (9) and BMP-7 homologue Screw (SCW) (10), while
Chordin is represented by the homologue Short gastrulation (SOG) (11, 12). Drosophila
homologues do not exist for the other vertebrate extracellular antagonists of BMP
signaling, such as Noggin and the Dan Family of BMP inhibitors (13). In Drosophila,
DPP is expressed dorsally, whereas SOG is expressed ventrolaterally. Secreted SOG is
thought to inhibit signaling by binding DPP in an extracellular latent complex (14), and it
appears that SOG diffuses to form a concentration gradient, resulting in a reciprocal
gradient of DPP signaling, that in turn determines cell fates along the dorsoventral axis
(14, 15). While a similar model has been generally accepted for vertebrate systems (16),
there are differences. First, in addition to the SOG orthologue Chordin, vertebrates
possess various other extracellular BMP antagonists. Second, it remains unclear to what
extent diffusion of Chordin over long distances is involved in vertebrate patterning (17).
Specifically, although high levels of overexpressed recombinant Chordin appear capable
of diffusing to directly inhibit BMP over long distances, there is evidence suggesting that
endogenous Chordin may employ a cell relay mechanism to effect long-range BMP
inhibition (17).
In Drosophila, gene products Tolloid (TLD) and Twisted gastrulation (TSG) help
shape the gradient of DPP signaling that forms the dorsoventral axis, through interactions
with SOG and DPP. Specifically, the metalloproteinase TLD cleaves SOG to release
DPP from SOG-DPP complexes (14), whereas TSG increases the avidity with which
SOG binds DPP, thus enhancing SOG’s antagonism of DPP signaling (18). In mammals,
the TLD-related proteinases BMP-1 and mammalian Tolloid-like 1 (mTLL-1) process
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6
Chordin in vivo (19, 20), whereas overexpression of various TLD-related proteinases in
Xenopus and zebrafish can counteract the dorsalizing effects of overexpressed Chordin
(9, 21-23). Vertebrates also possess homologues of TSG (Tsg) (24-26), which bind
Chordin and the DPP orthologue BMP-4 in a ternary complex (24-26), thereby enhancing
antagonism of BMP signaling (25, 26). TSG homologues may also be able to act as
agonists of BMP signaling under certain conditions (24).
Heparan sulfate proteoglycans (HSPGs), are abundant cell surface and ECM
molecules consisting of protein cores to which heparan sulfate (HS) chains are attached.
Cell surface protein cores include the syndecans (27), CD44 (V3) (28, 29), and
betaglycan (30), all of which have transmembrane domains; and the glypicans, which
bind plasma membranes via glycosyl phosphatidylinositol linkages (31). In both
interstitial and basement membrane ECM, perlecan is the primary core protein for HS
attachment (32, 33). HSPGs are implicated in diverse functions that include cell binding
to the ECM and to various growth factors (32). Secreted growth factors that interact with
HSPGs include, but are not limited to fibroblast growth factors (FGFs), the epidermal
growth factor and platelet-derived growth factor families, TGF-β1 and 2, and hepatocyte
growth factor (34). HSPGs have been shown to interact with various developmentally
important ligands including Wnt family members (34) and TGF-β-like BMPs (35, 36). In
regard to the latter, the Drosophila glypican dally is required for normal DPP signaling
(36), whereas HS may be capable of modulating BMP-2 activities in cell culture and
embryonic chick limb bud assays (35, 37, 38). Recently, it has been shown that Noggin
binds to HSPGs (39).
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7
Here, we demonstrate that Chordin binds heparin-Sepharose with elution profiles
similar to those of proteins, such as FGFs, that are known to functionally interact with
HSPGs in tissues. Surprisingly, we show that mammalian Tsg does not bind heparin, in
contrast to Drosophila TSG (40), but that it will bind heparin if complexed to Chordin
and/or Tsg. We show that Chordin binds embryonic tissues in a manner that is dependent
upon its interactions with tissue HSPGs, and that Chordin binds cell surface HSPGs, and
specifically cell surface syndecans, but that it does not detectably bind basement
membranes containing the major HSPG perlecan. Importantly, results presented herein
indicate that cell surface HSPGs strongly potentiate Chordin’s antagonism of BMP
signaling, effect the retention of Chordin at cell surfaces, and are necessary for the
cellular uptake of Chordin. Implications of the various data for the modulation of BMP
signaling and the formation of BMP signaling gradients are discussed.
EXPERIMENTAL PROCEDURES
Production of Recombinant Proteins- Flag-tagged mouse Chordin, Flag-tagged
Chordin fragments, and Protein C-tagged mouse Tsg were expressed and purified as
described previously (19, 25). Concentrations of BMP-4 (R&D Systems), Flag-tagged
mouse Chordin, Flag-tagged Chordin fragments, and Protein C-tagged mouse Tsg were
calculated by comparing intensities of Coomassie blue-stained bands to protein standards
of known concentrations.
Heparin-Sepharose Chromatography - A heparin-Sepharose slurry (CL-6B,
Amersham Pharmacia) was pelleted and resuspended in a double volume of phosphate
buffered saline (PBS), containing 1 mg/ml bovine serum albumin (BSA), and 100 µl were
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8
added to separate equimolar amounts of BMP-4, Chordin or Tsg, or to equimolar
combinations of these proteins that had been preincubated together for 30 min at 37 oC.
Heparin-Sepharose protein mixtures were incubated for 2 h at 4°C, then added to a 0.8 x
4 cm column and the flow through was collected. The column was washed with 500 µl
PBS containing 1 mg/ml BSA, and stepwise elution was performed using 75 µl aliquots
of PBS containing increasing concentrations of NaCl. Elution was followed by addition
of 4 x SDS-PAGE sample buffer, with 5% β-mercaptoethanol, to fractions and
electrophoresis on 4-15% acrylamide gradient gels (BioRad). Immunoblotting by
electrotransfer to polyvinylidene diflouride membranes, incubations of blots with
antibodies, and washes were performed as described previously (41). Rabbit polyclonal
antibody raised against the peptide EPPALPIRSEKEPLPVRGA (20), corresponding to
residues 30-48 at the N-terminus of the published mouse Chordin sequence (42);
monoclonal anti-BMP-4 antibody (R&D Systems); monoclonal anti-protein C-peroxidase
(Roche Applied Science), for detection of protein C epitope-tagged mouse Tsg;
biotinylated monoclonal anti-Flag (Sigma), streptavidin-horseradish peroxidase conjugate
and secondary antibodies (Amersham) were all used at a concentration of 1:5000.
Cell Binding Assays and Immunofluorescence - Raji cells stably transfected with
syndecan-1 (Raji-S1) (43), Raji cells similarly transfected with syndecan-4 (Raji-S4) and
parental Raji cells, cultured as previously described (44), and 10t1/2 cells, cultured in
Dulbecco’s modified Eagle’s medium (DMEM) and 10% fetal bovine serum (FBS)
(Hyclone), were fixed by incubation with 4% paraformaldehyde/PBS for 1 h. Fixed cells
were then incubated with/without 60 nM Chordin in Roswell Park Memorial Institute
(RPMI) medium, 10% FBS for 1h at room temperature. Three 5 min washes with PBS
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

9
were followed by incubation with anti-Chordin antibody (described above) at a
concentration of 1:200, with rat anti-syndecan-1 monoclonal antibody 281.2 at a
concentration of 1 µg/ml, or with rabbit polyclonal anti-syndecan 4 antibody (see below)
at a concentration of 1:300 for 1h. This was followed by three 5 min washes with PBS
and then incubation with Alexa 546-conjugated goat anti-rabbit antibody or Alexa 488-
conjugated donkey anti-rat antibody (Molecular Probes) at 1:1000 for 30 min, followed
by a final three 5 min washes with PBS. Anti-syndecan 4 rabbit polyclonal antibody was
made using a syndecan-4 exodomain-GST fusion protein. The antibody was passed over
a GST affinity column, leaving only syndecan 4-specific antibody. Transfected M2-10B4
cells were fixed as above, but not incubated 1 h at room temperature prior to addition of
anti-Chordin antibody.
For experiments in which Chordin-transfected and untransfected cells were co-
cultured, transfected M2-10B4 cells were labeled with CellTrackerTM Green CMFDA (5-
chloromethylfluorescein diacetate, Molecular Probes) as per manufacturer’s protocol.
The labeled/transfected cells were plated in droplets of medium and allowed to adhere
and spread for 5 h, such that they formed colonies. After 5 h, non-transfected cells in
sufficient medium to cover the bottom of the dish were plated, such that they adhered and
spread in areas surrounding the colonies of chordin-transfected cells. Cells were co-
cultured for an additional 24 h and were then fixed and immunostained, as described
above.
In Situ HS Binding Assays - In situ binding to HS was performed essentially as described
previously (45). Frozen tissue sections, cut to a thickness of 5 µm, were air dried, fixed
in 4% paraformaldehyde on ice, treated with 2 M NaCl to remove exogenous HS binding
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

10
molecules and then blocked overnight in RPMI + 10% FBS at 4°C. The following day,
sections were incubated with 60 nM Chordin for 1 h and, after three washes with PBS,
bound Chordin was detected using rabbit anti-Chordin antibody (described above) at a
concentration of 1:200 and Alexa 546-conjugated goat ant-rabbit antibody. Serial
sections were incubated with rabbit anti-perlecan antibodies (kind gift of J. Hassell,
University of South Florida) at 1:1000 and detected with Alexa 546-conjugated goat anti-
rabbit antibody at 1:1000
Removal of endogenous HS was accomplished by treatment of tissue sections or
cells with 0.006 IU/ml heparinase I and heparinase III (heparin lyases) (Seikagaku
America) for 2 h at 37°C, followed by addition of fresh enzyme for an additional 2 h.
Characterization of the distribution of total HS in tissues was with antibody mAb3G10
(Seikagaku America), which detects unsaturated glucuronate residues remaining on core
proteins following heparinase III treatment (46). Sections were incubated with mAb3G10
at 1:200 dilution in RPMI + 10% fetal calf serum followed by 1:300 Cy3-conjugated
donkey anti-mouse secondary antibody (Molecular Probes). Staining of sections without
prior heparinase III treatment showed no 3G10 staining (data not shown).
Western Blot Analysis - Cell lysate collection and subsequent Western blot
analysis were performed as described previously (47). Briefly, 10t1/2 cells were lysed in
cold PBS containing 1% Triton-X-100, 0.1% SDS, 5mM EDTA, 1mM PMSF, 1µg/ml
leupeptin and 2µg/ml aprotinin, and the cell layer was then scraped into this solution.
Following removal of the insoluble fraction by centrifugation, total protein levels were
determined by BCA assay. 100 µg of total protein was then precipitated overnight in 2.5
volumes of methanol at -20°C. Pellets were then resuspended in heparinase buffer (50
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

11
mM HEPES (pH 6.5), 50 mM NaOAc, 150 mM NaCl, 5 mM CaCl2) and incubated for 2
h at 37°C with 0.0001 units heparinase I (IBEX), 0.0001 heparinase III (IBEX), and 0.005
units Chondroitin ABC lyase (Sigma). Samples were resolved by SDS-PAGE,
transferred to Immobilon-P PVDF (Millipore), fixed with 0.05% glutaraldehyde for 30
min, and blocked overnight at 4°C in TBS containing 3% BSA. HSPGs were detected
with mAb3G10, followed by incubation with AP-conjugated secondary antibodies.
Proteins were visualized with ECF detection reagent (Amersham) and scanned on a
Storm phosphorimager (Molecular Dynamics).
Alkaline Phosphatase (AP), BMP Signaling Assay - M2-10B4 cells were
transfected either with empty pcDNA3.1 vector (Invitrogen), or with the same vector
containing cDNA coding sequences for murine Chordin with a COOH-terminal Flag
epitope, using lipofectAMINE (Life Technologies). The Chordin expression vector was
constructed by excising the Chordin-Flag insert from a previously described pCEP-based
vector (19) and inserting it between the pcDNA3.1 AflII and NotI sites. Transfected M2-
10B4 cells were selected with G418, resistant colonies were ring-cloned, and one clonal
line derived from empty vector-transfected cells, and one of the clonal lines expressing
highest levels of recombinant Chordin were used for the AP assays. AP assays were
performed essentially as described in Zebboudj et al. (48). Briefly, cells were washed
twice with PBS and grown for 24 hrs in either DMEM, 10% FBS or in low sulfate
DMEM (49) containing 10% dialysed FBS and 50 mM chlorate. Cells were then
trypsinized and plated in triplicate into 96 well plates at 10,000 cells per well in DMEM,
10% FBS or in low sulfate DMEM containing 10% dialyzed FBS and 50 mM chlorate.
After 16 h, cells were washed once with PBS and then placed in 200 µl of DMEM, 10%
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

12
FBS or 200 µl low sulfate DMEM with 10% dialysed FBS and 50 mM chlorate;
containing either 6 nM BMP-4 (dissolved in 4 mM HCl and 0.1% BSA) or 4 mM HCl
containing 0.1% BSA. AP activity was determined 48 h later. Cells were washed once
with PBS and lysed by freeze-thawing twice in 50 µl of 0.2% NP-40, 1 mM MgCl2. 150
µl of AP buffer [5 mg p-nitrophenyl Phosphate dissolved in 2.5 ml Sigma 221 Alkaline
Buffer Solution (Sigma, St. Louis, MO) diluted 1:2 with water] was added to each well
and plates were incubated for 1 h at 37°C. AP activity was then measured by absorbance
readings taken at 405 nm using a Universal Microplate Reader (Bio-Tek Instruments,
Winooski VT).
RESULTS
Chordin and BMP-4, but not Tsg Bind Heparin In Vitro - To gain insight into the
potential of Chordin to bind HS in vivo, purified recombinant murine Chordin was
incubated with heparin-Sepharose, and the mixture was placed in a column and eluted
with a step gradient of 0.15-2.0 M NaCl. As can be seen (Fig. 1A), Chordin binds
heparin, with the majority of bound Chordin eluting at 0.55-1.60 M NaCl, and some
requiring elution with 2.0 M NaCl or boiling in SDS. It has previously been shown, that
TGF-β-like BMPs (e.g. BMP-2, Ref. 35), bind heparin with high affinity. Here, BMP-4
is shown to bind heparin with an elution profile similar to that of Chordin (Fig. 1A).
Murine Tsg did not bind heparin under these conditions, as the vast majority was found in
the flow through and wash (Fig. 1A). This result was surprising, as Drosophila TSG has
been reported to be a heparin-binding protein (40).
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

13
Chordin binds BMP-4 and Tsg, and can bind the two proteins simultaneously in a
ternary complex (24-26). To determine whether binding to Tsg might decrease the
affinity of Chordin and/or BMP-4 for heparin, or whether binding of Tsg to the other two
proteins might increase its retention on heparin, the three proteins were pre-incubated
together prior to incubation with heparin-Sepharose. As can be seen (Fig. 1B) in the
presence of Chordin and BMP-4, some portion of the input Tsg is retained on heparin and
is eluted at fairly high concentrations of NaCl, similar to those at which Chordin and
BMP-4 are themselves eluted. As only some portion of input Tsg is expected to bind
Chordin and/or BMP-4 during pre-incubation, it seems likely that the portion retained on
heparin-Sepharose represents that portion of Tsg that is bound to Chordin and/or BMP-4.
Tsg separately pre-incubated with either Chordin or BMP-4 is retained on heparin-
Sepharose in amounts and with affinities similar to Tsg pre-incubated with the two
proteins together (data not shown). Thus, binding to Chordin and/or BMP-4 is
necessary/sufficient for Tsg binding to heparin.
The extracellular BMP antagonist Noggin binds heparin predominantly via a short
segment of sequence that includes a stretch of basic amino acid residues (39). Similarly,
BMP-2 binds heparin predominantly via a stretch of basic amino acids near its NH2-
terminus (35), and the highly similar BMP-4 most probably binds heparin via an NH2-
terminal region even more basic than that of BMP-2. Full-length Chordin contains 4
cysteine-rich (CR) domains that are thought to be involved in BMP binding (42, 50) (Fig.
2A). To determine whether Chordin might also bind heparin via a site localized to a
relatively small portion of the protein, recombinant versions of each of the three CR-
containing fragments that are generated via cleavage of Chordin by BMP-1/Tolloid-like
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

14
metalloproteinases (19) were prepared, as described (25), and were separately incubated
with heparin-Sepharose. As can be seen (Fig. 2B), the NH2-terminal CR1-containing
fragment, the middle CR2/CR3-containing fragment, and the COOH-terminal CR4-
containing fragment all bind heparin, with elution occurring at relatively high
concentrations of NaCl. Some of the CR1-containing fragment, and large amounts of the
CR4-containing fragment are also found in the flow through and wash, perhaps reflecting
improper folding by some fraction of these recombinant fragments. Nevertheless,
binding by all three fragments suggests that binding of Chordin to heparin is via more
than one site, consistent with the relatively even distribution of basic residues in the
Chordin sequence (42, 50).
Chordin Binds Tissues in an HSPG-dependent Manner - Next, we directly tested
the ability of Chordin to bind endogenous HSPGs in embryonic tissues. It has previously
been shown that proteins which bind HSPGs in vivo can, when added exogenously to
tissue sections, bind in a manner dependent upon interactions with endogenous HSPGs,
and that different endogenous HSPGs show selectivity for specific ligands (44, 45). To
determine whether exogenous Chordin might selectively bind tissues in an HSPG-
dependent fashion, recombinant Chordin was incubated on frozen sections from 17 dpc
mouse embryos. In such assays, Chordin was found to bind tissues in a manner
dependent upon interactions with endogenous HS, since binding was blocked by
pretreatment of tissues with heparin lyases (heparinases I and III) (Figs. 3-5). In contrast,
pre-treatment of tissue sections with chondroitin ABC lyase, which removes the side
chains of endogenous chondroitin sulfate proteoglycans, had no effect on Chordin binding
(data not shown).
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

15
In skin, a relatively weak HS-dependent signal was observed for Chordin bound to
epidermis and hair follicles (Fig. 3A). However, mast cells, which served as internal
controls for Chordin binding in skin, since these cells produce heparin and contain high
levels of heparin stored in α-granules, showed relatively high signal levels for Chordin
binding. This latter result is consistent with the probability that binding of Chordin to
tissues is via the same types of interactions by which it binds heparin-Sepharose in vitro.
Interestingly, Chordin did not detectably bind the HSPG-rich basement membrane at the
dermal-epidermal junction.
In the thorax, signal for Chordin binding was at particularly high levels in
developing ribs (Fig. 3B) and lung (Fig. 4A). Interestingly, in situ hybidization has
previously shown Chordin to be expressed at high levels in both these tissues (51). In rib
and lung, the tissue distribution of HS is demonstrated by use of the antibody mAb3G10,
which binds to residual HS stubs that remain on endogenous HSPG core proteins after
heparin lyase treatment (46). As can be seen (Figs. 3B and 4A), use of mAb3G10 shows
both ribs and lung to be abundant in endogenous HS. Moreover, binding of Chordin to
serial sections shows signal for Chordin to co-distribute with the placement of HS in
these tissues. In addition, heparin lyase treatment destroys Chordin signal in ribs and
lung, demonstrating binding of Chordin to be dependent on interactions with endogenous
HS in these tissues. Signal detected in skeletal muscles adjacent to the ribs, even after
heparin lyase treatment (Fig. 3B) is not due to Chordin binding, but is instead due to non-
specific binding of the primary antibody. A salient feature of Chordin binding in lung
was the absence of Chordin signal in the roughly circular areas of high HS content that
correspond to basement membranes surrounding the bronchioles (Fig. 4A). This is
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

16
illustrated more clearly in Fig. 4B, in which Chordin signal is seen to be absent from
basement membranes surrounding bronchioles and blood vessels, but present in
bronchiole epithelial cells (Fig. 4B, asterisk) and to smooth muscle cells surrounding both
bronchioles and blood vessels. The HS-rich circular areas that show no signal for
Chordin are confirmed as basement membranes by detection with antibodies to the
basement membrane-specific HSPG perlecan. Additionally, at high magnification (Fig.
4C) intense and punctate patterns of Chordin signal are seen in cells encircling airway
passages, while signal is absent from basement membranes adjacent to those cells. In a
similar fashion, signal for Chordin is clearly detectable in cells, but absent from basement
membranes in two additional tissues in which relatively high levels of Chordin signal
were detected: gut (Fig. 5A) and brain (Fig. 5B). A table is presented that summarizes
the distribution of Chordin binding and background signals in tissues that were evaluated
(Table 1).
Chordin Binds Cell Surface HSPG’s - Since signal for bound Chordin in tissues was
associated with cellular, rather than basement membrane HSPG (see above), we more
closely examined the ability of Chordin to bind cell surface HSPGs. Towards this end,
Chordin was incubated with cultured 10t1/2 mouse embryo cells, which have previously
been employed in culture-based assays of Chordin function (16). As can be seen (Fig.
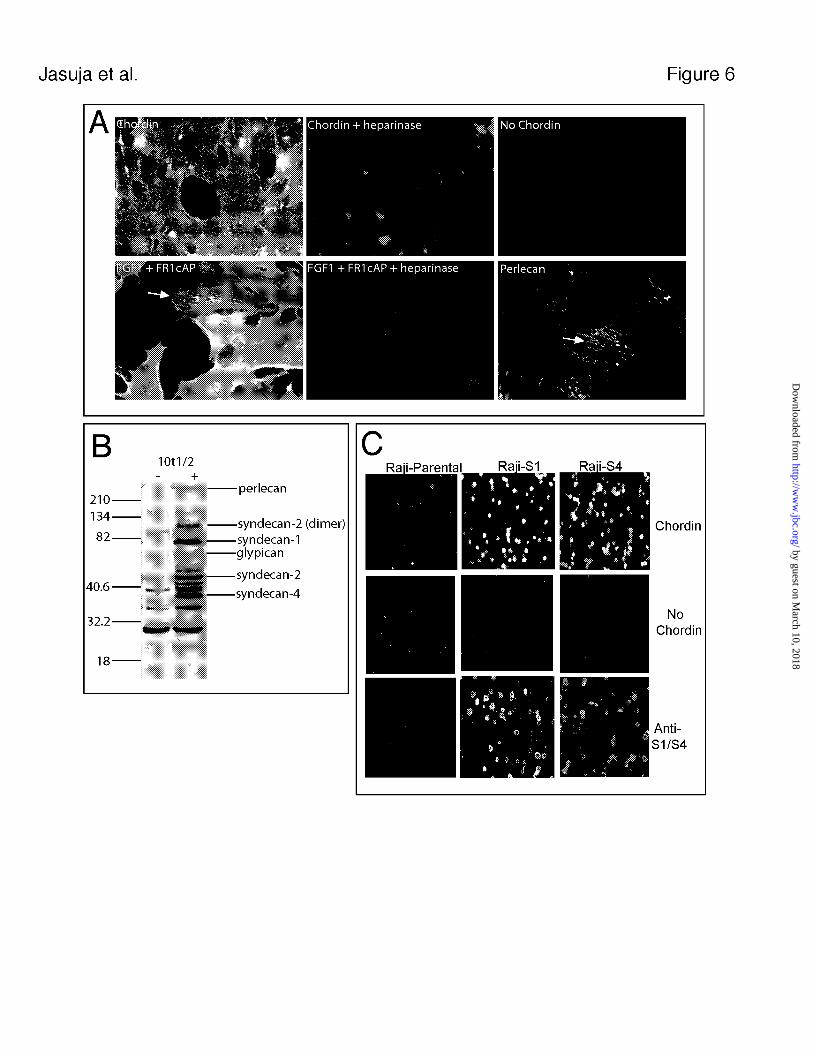
6A), Chordin binds 10t1/2 cell surfaces in an HS-dependent fashion. Such binding also
demonstrated the same selectivity for HSPGs as that observed in vivo, since Chordin
failed to co-localize with perlecan in any of the 10t1/2 cultures examined (see Fig. 6A).
Such selectivity contrasted with the binding in similar cultures of FGF1, a well-
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

17
characterized HS-binding growth factor, which not only bound 10t1/2 cell surface HS,
but which co-localized with perlecan as well (Fig. 6A).
To determine the profile of HSPG core proteins associated with 10t1/2 cells, a
Western blot of heparin lyase-treated cell layers was performed, using mAb3G10. This
approach showed 10t1/2 cells to contain syndecans 1, 2 and 4, and glypicans (Fig. 6B).
To begin characterizing which cell surface HSPGs might be involved in binding, Chordin
was incubated with cultured Raji cells, a lymphoblastoid cell line with little or no
expression of endogenous cell surface proteoglycans, or with Raji-S1 cells, a stable line
of Raji cells transfected with an expression vector for syndecan-1 (43). As shown in Fig.
6C, although Chordin did not bind to parental Raji cells, it avidly bound to Raji-S1 cells.
Similar results were obtained with Raji cells transfected with an expression vector for
syndecan-4. Thus, Chordin binds cell surface HSPGs, but this binding does not appear to
be specific to HS chains attached to a single type of HSPG core protein.
Cell Surface HSPGs are Powerful Regulators of Chordin’s Antagonism of BMP
Signaling and of Cellular Uptake of Chordin - To determine whether interactions with
HSPGs might affect Chordin’s ability to antagonize BMP signaling, we transfected M2-
10B4 murine marrow stromal cells with a Chordin expression vector, and compared the
ability of exogenously added BMP-4 to induce osteoblastic differentiation of these
precursor cells in standard culture medium (DMEM) or under conditions in which
sulfation of HSs is blocked (low sulfate DMEM containing chlorate, see Ref. 52).
Interestingly, Chordin was detected only in the conditioned medium of transfected M2-
10B4 cells cultured in low sulfate DMEM and Chlorate (Fig. 7A), consistent with the
possibillity that secreted Chordin is trapped at the cell surface by HSPGs under standard
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

18
growth conditions. In fact, immunofluorescent staining showed transfected M2-10B4
cells grown under normal culture conditions to be associated with high levels of signal for
Chordin, including high levels of intracellular Chordin, whereas transfected M2-10B4
cells in low sulfate medium and chlorate were essentially devoid of signal for associated
Chordin.
Levels of BMP signaling, as reflected by levels of osteoblastic differentiation
induced in M2-10B4 cells by addition of exogenous BMP-4, were determined by
measuring induced levels of osteoblast-specific alkaline phosphatase activity.
Importantly, whereas the ability of BMP-4 per se to induce osteoblastic differentiation
was unaffected by low sulfate, chlorate treatment, the ability of Chordin to inhibit BMP
signaling was totally abrogated under the same conditions (Fig. 7B). Thus, HSPGs
appear to play an essential role in the ability of Chordin to modulate BMP signaling. It
should be noted that in control experiments in which 10 mM sulfate was added to
chlorate-treated cultures, at the same time as chlorate, chlorate treatment had no effects
on either the association of Chordin with cells, as detected by immunofluorescence, or on
the ability of Chordin to inhibit BMP-signaling, in assays such as that used in Fig. 7B
(data not shown). Since addition of 10 mM sulfate allows sulfation of HSPGs to occur in
the presence of the concentrations of chlorate used here (52), such controls show the
effects of chlorate/low sulfation to occur via sulfation alone, and not via some
unanticipated effect of chlorate.
The above experiments show HSPGs to be involved in retaining Chordin at the
cell surface and in cellular uptake of Chordin. Such processes should profoundly affect
the diffusion of Chordin. In fact, the release of Chordin into the medium of cultures in
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

19
which sulfation is blocked (Fig. 7A) shows the degree to which HSPGs can affect release
of Chordin beyond the cell surface. However, since it is possible that diffusion of
Chordin may occur directly from the surface of a Chordin-producing cell to the surfaces
of adjacent cells, we fluorescently tagged transfected, Chordin-producing M2-10B4 cells,
co-cultured these with non-transfected M2-10B4 cells, and then used immunofluorescent
staining to determine whether Chordin was only associated with Chordin-producing cells,
or with adjacent non-producers as well. As can be seen (Fig. 8), detectable Chordin was
associated only with Chordin-producing cells. Together, results from Figs. 7 and 8 thus
indicate that HSPGs profoundly limit the diffusion of Chordin from the cell surface to the
extracellular space and from one cell surface to another.
DISCUSSION
Here we have demonstrated that mammalian Chordin binds heparin in vitro with a
high affinity matching those of factors, such as the FGFs, which have previously been
shown to bind and functionally interact with HSPGs in tissues. Moreover, we show that
Chordin binds tissues in a manner dependent upon its ability to bind HSPGs. This
binding seems limited to cell surface HSPGs, as basement membranes rich in the HSPG
perlecan are not detectably bound. The binding to cell surface, but not basement
membrane HSPGs denotes specificity in the interactions of Chordin with HSPGs. thus
indicating that charge alone is not the sole determinant of Chordin-HSPG binding. We
have also shown that Chordin is capable of binding both syndecans 1 and 4. The latter
result indicates that Chordin-HS binding is not limited to a single type of HSPG core
protein. Rather, Chordin binding may be specific to a certain type of HS chain which, in
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

20
turn may attached to a variety of core proteins. In this regard, it is of interest to note that
binding of FGFs to tissue HSPGs has shown to vary in response to cell type/tissue-
specific differences in HS sulfation patterns (44). Thus, similar differences in HS chains
may determine the degree to which Chordin binds different cells and tissues. The issues
of which spectrum of cell surface core proteins and what types of HS side chains may be
involved in determining Chordin binding remain to be resolved.
Interestingly, tissues to which exogenously added Chordin binds most strongly in
an HSPG-dependent way (e.g. cells lining the air passageways of the lungs, developing
bone, gut, and brain), are those in which we have previously shown Chordin RNA to be
expressed at highest levels in development and in the adult (19, 25, 51). Thus, higher
Chordin levels may normally be found localized to the surfaces of cells that secrete
Chordin.
Cell surface HSPGs are integrally involved in modulating the signaling of various growth
factors, including FGFs (52, 53), hepatocyte growth factor (54, 55), heparan sulfate-
binding epidermal growth factor (56, 57), and vascular endothelial growth factor (58, 59).
In addition, the Drosophila glypican dally is required for normal DPP signaling (36),
whereas heparin appears able to modulate signaling by BMP-2 in embryonic chick limb
bud assays (35). Here we demonstrate that cell surface HSPGs bind Chordin and that
such interactions serve to powerfully enhance the ability of Chordin to antagonize BMP
signaling. Specifically, the ability of Chordin to antagonize BMP signaling is essentially
abrogated in M2-10B4 cells via blocking of HSPG sulfation. Although a postulated role
for HS binding is the protection of proteins from digestion by endogenous proteinases
(60), we have found that heparin does not appreciably protect Chordin from in vitro
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

21
cleavage by BMP-12, a proteinase responsible for the in vivo proteolysis of Chordin in
mammals (20). Thus, a likely interpretation of the various data is that cell surface HSPGs
act to potentiate Chordin’s antagonism of BMP signaling by concentrating Chordin at the
cell surface, where it is optimally placed to limit the binding of BMP-2/4 to cognate cell
surface receptors. Interestingly, chlorate did not affect levels of BMP signaling in M2-
10B4 cells in the absence of Chordin (see Fig. 7B). Thus, despite the ability of BMP-4 to
bind heparin, sulfated HSPGs may not regulate BMP signaling, in at least some cell
types, in the absence of exogenous inhibitors such as Chordin.
Blockage of HSPG sulfation not only abrogated Chordin’s inhibition of BMP-2/4
signaling, but also dropped the high levels of intracellular Chordin in transfected M2-
10B4 cells down to basal levels. The most straightforward interpretation of this effect is
that intracellular Chordin in transfected M2-10B4 cells is predominantly Chordin that has
been previously secreted, and subsequently endocytosed via a mechanism dependent upon
cell surface HSPGs. Previously, Srinivasan et al. (15) have demonstrated that uptake of
SOG by cells in vivo is a dynamin-dependent mechanism involved in limiting
extracellular accumulation of SOG dorsally, thus acting to maintain a proper SOG
gradient in Drosophila embryos. Our results, with the SOG orthologue Chordin, are
consistent with the roles that HSPGs have been reported to play in this type of uptake and
degradation of various extracellular proteins (34), including the BMP antagonist
Follistatin (61). It will be of interest to determine whether HSPG facilitates not only the
uptake and degradation of Chordin, but of Chordin-BMP2/4 complexes as well, or
perhaps complexes of Chordin, bound to BMP-2/4 and their associated type I and II
receptors. HSPG potentiation of BMP-2/4 and receptor uptake and degradation would
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

22
constitute another mechanism whereby HSPG might serve to potentiate the inibition of
BMP signaling, although the data of Fig. 7 shows that such a mechanism would be
dependent on the presence of Chordin. Chordin bound to cell surface HSPGs might well
have relatively frequent interactions with BMP-2/4-receptor complexes, as both would be
limited to two dimensional diffusion within the same plane.
Larrain et al. (62) have reported that Chordin binds to cell surfaces via integrin-
α3. It seems quite possible that Chordin may bind both cell surface HSPGs and other cell
surface proteins, such as integrins, and no data in the current report are contrary to that
possibility. However, Larrain et al. also report that bound Chordin appears to co-localize
with structures reminiscent of focal-adhesion plaques and with actin filaments underlying
the cell surface. Although we did find that Chordin bound nonspecifically to intracellular
filaments in permeabilized 10t1/2 cells or to patches of filaments in partially
permeabilized 10t1/2 cells, we did not see Chordin localization to adhesion plaques when
permeabilization was avoided (data not shown). Thus, perhaps due to differences in
technique, we cannot confirm the distribution of cell surface Chordin binding reported in
the previous study.
In addition to its roles in potentiating the antagonism of BMP signaling by
Chordin and cellular uptake of Chordin, another outcome of the strong interactions
between Chordin and cell surface HSPGs appears to be limiting the free diffusion of
Chordin. Thus, the combined effects of cell surface HSPGs on Chordin would be
expected to powerfully contribute to shaping and stabilizing fields of BMP inhibition in
vivo, with concomitant shaping and stabilizing of reciprocal fields of BMP signaling.
Such interactions may operate in both formation of the embryonic dorsoventral axis, and
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

23
in organogenesis. In regard to the latter, our finding of a correlation between those
tissues which produce and those tissues which bind the highest levels of Chordin suggests
that such binding may help localize peak levels of Chordin to those areas that synthesize
the protein, and in this way sharpen the fields and structures of presumptive organs. Our
finding of cell surface HSPG- Chordin binding is of interest in the context of the findings
of Blitz et al. (17), who have noted that although high levels of overexpressed
recombinant Chordin appear capable of diffusing to directly inhibit BMP over long
distances, lower levels of endogenous Chordin may directly inhibit BMP signaling over a
short range, propagated over longer distances via a cell relay mechanism. Thus,
endogenous Chordin may act only locally in tissues when expressed at levels low enough
to be completely bound by the surface HSPGs of Chordin-producing cells, but via long
range diffusion when produced at levels that overwhelm the binding capacities of HSPGs
on the surfaces of Chordin-producing cells.
Surprisingly, we did not detect binding of mammalian Tsg to heparin-Sepharose,
despite previous work showing Drosophila TSG to be a heparin-binding protein (40).
Nevertheless, the present Tsg result is consistent with a previous finding that vertebrate
Tsg appears to be a highly diffusible molecule in vivo (26). Chang et al. (26) have found
that despite its role in antagonizing BMP signaling, Tsg is incapable of inducing
secondary dorsal axes in Xenopus embryos unless tethered to a recombinant
transmembrane domain. This suggests that tethering is necessary to achieve local Tsg
concentrations high enough to induce secondary dorsal axes. Based on previous findings
that vertebrate Tsg, Chordin and BMP-2/4 form a ternary complex (24-26), and on the
present finding that Tsg only binds heparin in the presence of Chordin and/or BMP-2/4,
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

24
we suggest that establishment of high local concentrations of Tsg in vivo may be
influenced by tethering of Tsg to cell surface HSPGs via Chordin. The system whereby
fields of BMP signaling are shaped by SOG/Chordin and TSG/Tsg is remarkably
conserved in arthropods and vertebrates, although various differences do exist (14, 19,
21). Differences in ability of TSG/Tsg to bind heparin may mark one point of divergence
of the system in widely diverged species.
REFERENCES
1. Wozney, J.M., Rosen, V., Celeste, A.J., Mitsock, L.M., Whitters, M.J., Kriz, R.W.,
Hewick, R.M., and Wang, E.A. (1988) Science 242, 1528-1534
2. Celeste, A.J., Iannazzi, J.A., Taylor, R.C., Hewick, R.M., Rosen, V., Wang, E.A., and
Wozney, J.M. (1990) Proc. Natl. Acad. Sci. U.S.A. 87, 9843-9847
3. Wang, E.A., Rosen, V., D’Alessandro, J.S., Bauduy, M., Cordes, P., Harada, T., Israel,
D.I., Hewick, R.M., Kerns, K.M., LaPan, P., Luxenberg, D.P., McQuaid, D.,
Moutsatsos, I.K., Nove, J., and Wozney, J.M. (1990) Proc. Natl. Acad. Sci. U.S.A. 87,
2220-2224
4. Hogan, B.L. (1996) Genes Dev. 10, 1580-1594
5. Graff, J.M., Thies, R.S., Song, J.J., Celeste, A.J., and Melton, D.A. (1994) Cell 79,
169
-179
6. Suzuki, A., Thies, R.S., Yamaji, N., Song, J.J., Wozney, J.M., Murakami, K., and
Ueno,
N. (1994) Proc. Natl. Acad. Sci. U.S.A. 91, 10255-10259
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

25
7. Hawley, S.H., Wünnenberg-Stapleton, K., Hashimoto, C., Laurent, M.N., Watabe, T.,
Blumberg, B.W., and Cho, K.W. (1995) Genes Dev. 9, 2923-2935
8. Cho, K.W.Y., and Blitz, I.L. (1998) Curr. Op. Gen. Dev. 8, 443-449
9. Padgett, R.W., St. Johnston, R.D., and Gelbart, W.M. (1987) Nature 325, 81-84
10. Arora, K., Levine, M.S., and O’Connor, M.B. (1994) Genes Dev. 8, 2588-2601
11. Francois, V., Sollway, M., O’Neill, J.W., Emery, J., and Bier, E. (1994) Genes Dev. 8,
2602-2616
12. Francois, V., and Bier, E. (1995) Cell 80, 19-20
13. Massagué, J., and Chen, Y.-G. (2000) Genes & Dev. 14, 627-644
14. Marqués, G., Musacchio, M., Shimell, J.J., Wünnenberg-Stapleton, K., Cho, K.W.Y.,
and O’Connor, M.B. (1997) Cell 91, 417-426
15. Srinivasan, S., Rashka, K.E., and Bier, E. (2002) Dev. Cell 2, 91-101
16. Piccolo, S., Sasai, Y., Lu, B., and De Robertis, E.M. (1996) Cell 86, 589-598
17. Blitz, I.L., Shimmi, O., Wünnenberg-Stapleton, K., O’Connor, M.B., and Cho,
K.W.Y.,
(2000) Dev. Biol. 223, 120-138
18. Ross, J.J., Shimmi, O., Vilmos, P., Petryk, A., Kim, H., Gaudenz, K., Hermanson, S.,
Ekker, S.C., O’Connor, M.B., and Marsh, J.L. (2001) Nature 410, 479-483
19. Scott, I.C., Blitz, I.L., Pappano, W.N., Imamura, Y., Clark, T.G., Steiglitz, B.M.,
Thomas,
C.L., Maas, S.A., Takahara, K., Cho, K.W.Y., and Greenspan, D.S. (1999) Dev. Biol.
213, 283-300
20. Pappano, W.N., Steiglitz, B.M., Scott, I.C., Keene, D.R., and Greenspan, D.S. (2003)
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

26
Mol. Cell. Biol. 23, 4428-4438
21. Piccolo, S., Agius, E., Lu, B., Goodman, S., Dale, L., and De Robertis, E.M. (1997)
Cell
91, 407-416
22. Blader, P., Rastegar, S., Fisher, N., and Strähle, U. (1997) Science 278, 1937-1940
23. Wardle, F.C., Welch, J.V., and Dale, L. (1999) Mech. Dev. 86, 75-85
24. Oelgeschläger, M., Larrain, J., Geissert, and De Robertis, E.M. (2000) Nature 405,
757-
763
25. Scott, I.C., Blitz, I.L., Pappano, W.N., Maas, S.A., Cho, K.W.Y., and Greenspan, D.S.
(2001) Nature 410, 475-478
26. Chang, C., Holtzman, D.A., Chau, S., Chickering, T., Woolf, E.A., Holmgren, L.M.,
Bodorova, J., Gearing, D.P., Holmes, and Brivanlou, A.H. (2001) Nature 410, 483-
487
27. Zimmermann, P., and David, G. (1999) FASEB J. 13, S91-S100
28. Bennett, K.L., Jackson, D.G., Simon, J.C., Tanczos, E., Peach, R., Modrell, B.,
Stamenkovic, I., Plowman, G., and Aruffo, A. (1995) J. Cell Biol. 128, 687-698
29. Jackson, D.G., Bell, J.I., Dickinson, R., Timans, J., Shields, J., and Whittle, N. (1995)
J.
Cell Biol. 128, 673-685
30. Andres, J.L., Stanley, K., Cheifetz, S., and Massague, J. (1989) J. Cell Biol.
109, 3137
-3145.
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

27
31. Filmus, J. (2001) Glycobiology 11, 19R-23R
32. Noonan, D.M., Fulle, A., Valente, P., Cai, S., Horigan, E., Sasaki, M., Yamada, Y.,
and
Hassell, J.R. (1991) J. Biol. Chem. 266, 22939-22947
33. Iozzo, R.V. (1998) Annu. Rev. Biochem. 67, 609-652
34. Bernfield, M., Gotte, M., Park, P.W., Reizes, O., Fitzgerald, M.L., Lincecum, J., and
Zako, M. (1999) Annu. Rev. Biochem. 68, 729-777
35. Ruppert, R., Hoffmann, E., and Sebald, W. (1996) Eur. J. Biochem. 237, 295-302
36. Jackson, S.M., Nakato, H., Sugiura, M., Jannuzi, A., Oakes, R., Kaluza, V., Golden,
C.,
and Selleck, S.B. (1997) Development 124, 4113-4120
37. Irie, A., Habuchi, H., Kimata, K., and Sanai, Y. (2003) Biochem. Biophys. Res.
Commun. 308, 858-865
38. Takada, T., Katagiri, T., Ifuku, M., Morimura, N., Kobayashi, M., Hasegawa, K.,
Ogamo,
A., and Kamijo, R. (2003) J. Biol. Chem. 278, 43229-43235
39. Paine-Saunders, S., Viviano, B.L., Economides, A.N., and Saunders, S. (2002) J.
Biol.
Chem. 277, 2089-2096
40. Mason, E.D., Williams, S., Grotendorst, G.R., and Marsh, J.L. (1997) Mech. Dev. 64,
61-75
41. Lee, S., Solow-Cordero, D.E., Kessler, E., Takahara, K., and Greenspan, D.S. (1997)
J.
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

28
Biol. Chem. 272, 19059-19066
42. Pappano, W.N., Scott, I.C., Clark, T.G., Eddy, R.L., Shows, T.B., and Greenspan,
D.S.
(1998) Genomics 52, 236-239
43. Lebakken, C.S., and Rapraeger, A.C. (1996) J. Cell Biol. 132, 1209-1221
44. Allen, B.L., Filla, M.S., and Rapraeger, A.C. (2001) J. Cell Biol. 155, 845-857
45. Friedl, A., Chang, Z., Tierney, A., and Rapraeger, A.C. (1997) Am. J. Pathol. 150,
1443
-1455
46. David, G., Bai, X.M., Van Der Schueren, B., Cassiman, J.-J., and Van Den Berghe,
H.
(1992) J. Cell Biol. 119, 961-975
47. Burbach, B.J., Friedl, A., Mundhenke, C., and Rapraeger, A.C. (2003) Matrix Biol.
22,
163-177
48. Zebboudj, A.F., Imura, M., and Boström, K., (2002) J. Biol. Chem. 277, 4388-4394
49. Rapraeger, A.C., Guimond, S., Krufka, A., and Olwin, B.B., (1994) Methods
Enzymol.
245, 219-240
50. Sasai, Y., Lu, B., Steinbeisser, H., Geissert, D., Gont, L.K., and De Robertis, E.M.
(1994) Cell 79, 779-790
51. Scott, I.C., Steiglitz, B.M., Clark, T.G., Pappano, W.N., and Greenspan, D.S. (2000)
Dev. Dyn. 217, 449-456
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

29
52. Rapraeger, A.C., Krufka, A., and Olwin, B.B. (1991) Science 252, 1705-1708
53. Yayon, A., Klagsbrun, M., Esko, J.D., Leder, P., and Ornitz, D.M. (1991) Cell 64,
841-
848
54. Sakata, H., Stahl, S.J., Taylor, W.G., Rosenberg, J.M., Sakaguchi, K., Wingfield,
P.T.,
and Rubin, J.S. (1997) J. Biol. Chem. 272, 9457-9463
55. Sergeant, N., Lyon, M., Rudland, P.S., Fernig, D.G., and Delehedde, M. (2000) J.
Biol.
Chem. 275, 17094-17099
56. Kleeff, J., Ishiwata, T., Kumbasar, A., Friess, H., Buchler, M.W., Lander, A.D., and
Korc, M. (1998) J. Clin. Invest. 102, 1662-1673
57. Paria, B.C., Elenius, K., Klagsbrun, M., and Dey, S.K. (1999) Development 126,
1997-
2005
58. Cohen, T., Gitay-Goren, H., Sharon, R., Shibuya, M., Lalaban, R., Levi, B.Z., and
Neufeld, G. (1995) J. Biol. Chem. 270, 11322-11326
59. Gengrinovitch, S., Berman, B., David, G., Witte, L., Neufeld, G., and Ron, D. (1999)
J.
Biol. Chem. 274, 10816-10822
60. Gospodarowicz, D., and Cheng, J. (1986) J. Cell Physiol. 128, 475-484
61. Hashimoto, O., Nkamura, T., Shoji, H., Shimasaki, S., Hayashi, Y., and Sugino, H.
(1997) J. Biol. Chem. 272, 13835-13842
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

30
62. Larrain, J., Brown, C., and De Robertis, E.M. (2003) EMBO Rep. 4, 813-818
1The abbreviations used are: BMP, bone morphogenetic protein; DPP,
decapentaplegic; SCW, Screw; SOG, Short gastrulation; TLD, Tolloid; TSG, Drosophila
Twisted gastrulation; mTLL-1, mammalian Tolloid-like 1; Tsg, vertebrate Twisted
gastrulation; HSPG, heparan sulfate proteoglycan; ECM, extracellular matrix; HS,
heparan sulfate; PBS, phosphate buffered saline; BSA, bovine serum albumin; PAGE,
polyacrylamide gel electrophoresis; DMEM, Dulbecco’s modified Eagle’s medium; FBS,
fetal bovine serum; RPMI, Roswell Park Memorial Institute; AP, alkaline phosphatase.
2 W. N. Pappano and D. S. Greenspan, Unpublished observations.
FIG. 1. Heparin binds Chordin and BMP-4, but not Tsg. Western blot analyses using
monoclonal antibodies specific for BMP-4, the C-terminal Flag-epitope of Chordin, or
the C-terminal protein C epitope of Tsg were used to identify fractions containing the
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

31
various proteins following elution from a heparin-Sepharose column at increasing
concentrations of NaCl. Chordin, BMP-4, and Tsg were either separately added to
heparin-Sepharose (A) or pre-incubated together for 30 min at 37ºC before addition to
heparin-Sepharose (B). In the final lanes of A and B, material remaining on beads
subsequent to elution with 2.0 M NaCl was removed by boiling beads in SDS-PAGE
sample buffer. Longer exposures of the Tsg Western blot in A verified that no Tsg bound
to heparin-Sepharose at higher NaCl concentrations.
FIG. 2. Chordin cleavage fragments containing cysteine-rich domains interact with
heparin. (A) Schematic diagram of full-length Chordin, including four cysteine-rich
(CR) domains, which are thought to be required for interaction with BMPs. Arrows
indicate defined sites of cleavage by BMP-1/Tolloid-like proteases. (B) Recombinant,
Flag-tagged, CR-containing cleavage products were incubated with heparin-Sepharose,
followed by elution at increasing NaCl concentrations. SDS-PAGE was performed on the
elution products, followed by Western blot analysis with monoclonal anti-Flag
antibodies.
FIG. 3. Chordin binds HS in the epidermis and rib of 17 dpc mouse embryos.
Exogenous Chordin was bound to 17 dpc mouse embryo sagittal sections and was
detected with Chordin specific antibodies, as described in Experimental Procedures. (A)
Sections from 17 dpc mouse skin demonstrate Chordin binding to HS in the epidermis as
well as in hair follicles, as denoted by the arrows (left panel). Chordin also interacts with
heparin in mast cells. In the absence of exogenous Chordin addition, only background
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

32
levels of signal are detected (right panel). (B) Chordin interacts with HS in the
developing rib (upper left panel). Total HS is identified with mAb3G10 (lower left
panel). Pre-treatment of sections with heparin lyases abrogates Chordin binding (lower
right panel). No endogenous Chordin is detected in the developing rib, although some
non-specific binding to muscle cells is detected (see arrow, upper right panel). Ep,
epidermis; Fo, follicle; Ma, Mast cell; Mu, Muscle.
FIG. 4. Chordin interacts with cell-associated, but not basement membrane HS in
17 dpc mouse lung. Exogenous Chordin was bound to 17 dpc mouse embryo sagittal
sections and was detected with Chordin specific antibodies, as described in Experimental
Procedures. (A) Exogenously added Chordin interacts with HS in the lung (upper left
panel), although endogenous Chordin protein is not detected (upper right panel).
mAb3G10 detection of total HS (lower left panel). Heparin lyase pre-treatment prevents
Chordin binding to lung tissue (lower right panel). (B) Chordin interacts with epithelial
HS (marked by asterisk) in the bronchioles of 17 dpc mice (upper left panel). No signal
is detected in the absence of exogenous Chordin addition (upper right panel, asterisk
marks position of epithelial cells). mAb3G10 identified HS both on cell surfaces of the
epithelia (marked by asterisk) as well as in basement membranes (lower left panel), while
anti-perlecan antibodies stained basement membranes (lower right panel), but not
epithelial cells (marked by asterisk). (C) Chordin binds at high levels to specific cells
within the bronchioles of 17 dpc mice. Higher magnification inspection of Chordin
binding to HS reveals intense punctate localization to specific cells lining bronchioles
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

33
(see arrows, left panels). No signal is detected in the absence of exogenous Chordin
(right panels), or after pre-treatment with heparin lyases (middle panels).
FIG. 5. Interaction of Chordin with cell-associated HS in E17 mouse gut and brain.
Exogenous Chordin was bound to 17 dpc mouse embryo sagittal sections and was
detected with Chordin specific antibodies, as described in Experimental Procedures.
Chordin interacts with cell-associated HS in both gut (A) and brain (B) of 17 dpc mouse
embryos (upper left panels). Only background levels of signal are detected without
Chordin addition (upper right panels). Total HS (lower left panels). Perlecan (lower
right panels). Arrows indicate epithelia lining the lumen of the gut. BV, blood vessel.
FIG. 6. Chordin interacts with cell surface HSPGs on 10t1/2 fibroblasts. (A) Fixed
monolayers of 10t1/2 fibroblasts were incubated with 60 nM Chordin (upper left panel) or
pre-treated with heparinases I and III followed by incubation with 60 nM Chordin (upper
middle panel). Cells were also incubated with anti-Chordin antibody in the absence of
Chordin addition (upper right panel). 10t1/2 cells were also incubated with 30 nM FGF1
and 100 nM of a fusion protein consisting of the extracellular portion of the FGF-receptor
1 fused to alkaline phosphatase (FR1cAP) following no treatment (lower left panel), or
treatment with heparinases I and III (lower middle panel). Arrows in the lower right
panel indicates ECM HSPG distribution following staining with anti-perlecan antibody.
Arrows in lower left panel indicate FGF-FR1cAP binding to HS associated with perlecan.
(B) Identification of 10t1/2 HSPG core protein expression profile following no treatment
(-) or treatment with heparinases I and III (+). (C) Binding of Chordin to Raji parental
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

34
cells (upper left panel) or Raji cells expressing recombinant syndecan-1 (Raji-S1) (upper
middle panel) or recombinant syndecan-4 (Raji-S4) (upper right panel) following
incubation with 60 nM Chordin. Cells were also incubated with anti-Chordin antibody
alone (middle panels) or with anti-S1 antibody (lower left and middle panels) or anti-S4
antibody (lower right panel). Although not shown, Raji parental cells showed no signal
when incubated with anti-S4 antibody.
FIG. 7. HSPGs participate in cellular binding and uptake of Chordin and potentiate
Chordin’s antagonism of BMP signaling. (A) M2-10B4 cells transfected either with an
empty vector, or with a Chordin expression vector were grown in normal DMEM or in
low sulfate (LS) DMEM containing chlorate. Conditioned media were then subjected to
Western blot analysis for the presence of Chordin (left panel). Cell layers were subjected
to immunofluorescent staining with anti-Chordin antibody (right panel). Insets show
fields of cells at lower magnification, whereas higher magnification shows details of
cellular staining. (B) M2-10B4 cells transfected either with an empty vector or a Chordin
expression construct (Chordin) were incubated in DMEM or in low sulfate DMEM
containing 50 mM chlorate, in the presence (+) or absence (-) of BMP-4. After 48 hrs,
alkaline phosphatase activity, as a measure of BMP signaling, was quantified by reading
the absorbance @ 405nm, and is given in arbitrary units.
FIG. 8. Chordin does not readily diffuse between surfaces of adjacent cells.
Chordin-expressing M2-10B4 cells were tagged with green fluorescent dye, co-cultured
with non-tagged, non-Chordin producing M2-10B4 cells, and immunofluorescent staining
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

35
was then performed using anti-Chordin primary antibodies and secondary antibodies
tagged with Alexa 546. A Shows the green dye-tagged Chordin-producing cells in the co-
cultures, B shows the immunostaining for Chordin, and C shows merging of the two
signals (yellow). D is a phase-contrast photomicrograph of the same fields of cells shown
in A-C. Insets show fields of cells at lower magnification, whereas higher magnification
shows details of cellular staining.
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Jasuja et al. Table I
Table I
Summary of exogenous Chordin’s interaction with HS on various
mouse embryo tissues
HS site 1° + 2° Abs alone +60 nM Chordin
Skin - +Skeletal Muscle + +Mast Cells - +Gut - ++Rib - ++Lung - ++Liver - +Submandibular Gland - +Atrium - +Ventricle - +Artery - +Vein - -Brain Capillary - -Brain - ++
Sectioned embryos were either treated with primary anti-Chordin and secondary antibodies alone (1° + 2° Abs alone), as a control, or with 60 nM exogenous Chordin followed by 1° + 2° Abs (+60 nM Chordin). Chordin-HS interactions are denoted by + or ++, reflecting the relative binding intensities. Background signal could be detected in skeletal muscle when slides were treated with 1° + 2° Abs alone, possibly as a result of antibody trapping, as no Chordin protein or other distinct bands could be detected on
Western blots of homogenized muscle samples using these antibodies2.
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from

GreenspanReema Jasuja, Benjamin L. Allen, William N. Pappano, Alan C. Rapraeger and Daniel S.
signaling and are necessary for cellular uptake of chordinCell surface heparan sulfate proteoglycans potentiate chordin's antagonism of BMP
published online September 20, 2004J. Biol. Chem.
10.1074/jbc.M408129200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on March 10, 2018
http://ww
w.jbc.org/
Dow
nloaded from