Embed Size (px)
Citation preview

PL-ISSN 0015-5497 (print), ISSN1734-9168 (online) FoliaBiologica (Kraków), vol. 61 (2013),No 1-2Ó Institute of Systematics andEvolution ofAnimals, PAS,Kraków, 2013 doi:10.3409/fb61_1-2.79
Comparison of the Effect of "1- and "2-Adrenoceptor
Agonists and Antagonists on Muscle Contractility
of the Rabbit Abdominal Aorta in vitro
Jan GNUS, Agnieszka RUSIECKA, Albert CZERSKI, Wojciech ZAWADZKI, Wojciech WITKIEWICZ,and Willy HAUZER
Accepted November 22, 2012
GNUS J., RUSIECKA A., CZERSKI A., ZAWADZKI W., WITKIEWICZ W., HAUZEr W. 2013.Comparison of the effect of "1- and "2-adrenoceptor agonists and antagonists on musclecontractility of the rabbit abdominal aorta in vitro. Folia Biologica (Kraków) 61: 79-85.The aim of the study was to demonstrate the effect of selected agonists and antagonists ofá-adrenergic receptors on muscle contractility of the rabbit abdominal aorta in vitro withparticular emphasis on á2-adrenergic receptor subtypes. The study was conducted on 30 NewZealand breed rabbits from which specimens of the abdominal aorta were collected. Thesections were set up in an automatic water bath in a Krebs-Henseleit buffer at 37oC. Theexperiments showed that á1-adrenergic receptors played the main role in the contractileresponse of the rabbit abdominal aorta. Stimulation of á1-adrenergic receptor by administrationof phenylephrine resulted in an increase in smooth muscle tonus of the rabbit abdominal aortaby an average of 4.75mN. The reaction after stimulation of á2-adrenergic receptors by similardoses of their agonists was much weaker. Prolonged tissue response time and time needed toreach maximum tonus for á2-adrenergic receptor agonists were observed. The obtainedresults confirm the thesis that the á1-adrenergic receptor is the most important factor controllingthe contractility of the rabbit abdominal aorta, but the á2-adrenergic receptor is also involvedin maintaining muscle tissue tonus.Key words: Abdominal aorta, adrenergic receptors, contractility, in vitro studies, rabbit.Jan GNUS, Wojciech WITKIEWICZ, Willy HAUZER, Regional Specialist Hospital in Wroc³aw,Research and Development Centre, Kamieñskiego 73a, 51-124 Wroc³aw, Poland.E-mail: [email protected]
[email protected]@gmail.pl
Agnieszka RUSIECKA, Albert CZERSKI,Wojciech ZAWADZKI, Department of Animal Physiol-ogy and Biostructure, Institute of Animal Physiology, Wroc³aw University of Environmentaland Life Sciences, K. C. Norwida 31, 50-375 Wroc³aw, Poland.E-mail: [email protected]
[email protected]@up.wroc.pl
Activation of the sympathetic system plays animportant role in the regulation of vascular resis-tance (OSWALD et al. 1983). It is generally as-sumed that the postsynaptic cooperation of á1- andá2-adrenergic receptors in blood vessels is an im-portant element of lumen regulation (DOCHERTYet al. 1979; DREW et al. 1979).
The á1-adrenergic receptors are stimulating re-ceptors responsible for regulation of many bio-logical processes; they are heterogeneous, i.e. theyhave different subtypes. There are 3 subtypes ofthe á1-adrenergic receptor: á1A, á1B, á1D (HIEBLE et al.1995; HIEBLE & RUFFOLO 1996; DOCHERTY 1998;BRODDE & MICHEL 1999; LANGER 1999).
The á1-adrenergic receptor is connected withcalcium signalling, which means that its stimula-
tion leads to an increased concentration of intracel-lular calcium ions (SMITH et al. 1997). Stimulationof the á1-adrenergic receptor causes an intracellu-lar influx of Ca2+ ions by opening calcium chan-nels sensitive to 1.4-dihydropyridine, while theá1B-adrenergic receptor releases intracellular Ca2+(HAN et al. 1987; MINNEMAN 1988; SUZUKI et al.1990). The á1-adrenergic receptors are present inthe brain (all subtypes), smooth muscles (á1A- andá1D-adrenergic), liver (á1A- and á1D-adrenergic),heart and prostate (á1A-adrenergic).
The á2-adrenergic receptors are involved in theregulation of many physiological processes in-cluding the cardiovascular system. They belong tothe family of G proteins that inhibit adenylyl cy-clase and thus lower the intracellular concentra-tion of cAMP.

According to the current classification adoptedby the International Union of Pharmacology Sub-committee on Nomenclature for Adrenoceptors,there are 3 subtypes of the á2-adrenergic receptor:á2A, á2B and á2C. This classification is the result ofadopting pharmacological criteria and using mo-lecular cloning techniques (BYLUND et al. 1988;LOMASNEY et al. 1991; HEIN et al. 1995; BYLUND1998). An á2D subtype was also distinguished, butnow it is assumed to be the same as the á2A recep-tor, though identified in other animal species.
The á2-adrenergic receptor subtypes are involvedin the regulation of distinct physiological func-tions. Existing pharmacological and genetic stud-ies indicate that the á2A subtype is responsible formost of the reactions typical for á2-adrenergic re-ceptors, including adjustment of anti-hypertensivemechanisms (ALTMAN 1999). In addition, it playsa central role in presynaptic noradrenaline release,increases the local blood flow in the prefrontal cor-tex and is responsible for working memory per-formance (MACMILLAN et al. 1996; ALTMAN 1999;AVERY et al. 2000). á2A/D and á2C receptors arealso associated with Attention Deficit Hyperactiv-ity Disorder (CHO et al. 2008). Moreover, activa-tion of á2-adrenoceptors affects digestive systemmotility, especially activation of the á2A subtype,which is probably responsible for its functioning.á2 receptors are also involved in the regulation ofuterine contractile activity. A study conducted invivo on rat uterus sections showed that á2A and á2Csubtypes were responsible for reducing the con-tractile response after administration of norepi-nephrine in the final days of pregnancy (GASPARet al. 2007).
Compared to the á2A subtype, the typical func-tions of the á2B and á2C subtypes are less known. Incontrast to the presynaptic localisation of á2A andá2C subtypes, á2B receptors are postsynaptic(DREW et al. 1979; PHILIPP et al. 2002). In addi-tion, the á2B subtype mediates the induction ofuterine contractions in pregnant rats, unlike á2A andá2C subtypes, which inhibit uterine contractions.
The á2C-adrenoreceptor subtype is localised mainlyin the central nervous system. Its role in hemody-namic processes has not been fully understood yet;it seems to mediate vein contraction (GAVIN et al.1997), but it was also found in the arterial smoothmuscle (CHOTANI et al. 2000). It participates in theregulation of the cerebral cortex, and activation ofthese receptors in a depressive mood may be bene-ficial for the treatment of neuropsychiatric disor-ders and diseases (GYIRES et al. 2009).
Summing up, separate locations within the bodymay indicate that á2-adrenoceptor subtypes (á2A,á2B, á2C) are involved in various biological pro-cesses and have different regulatory functions.
Material and Methods
The study was approved by the Local II EthicalReview Board, approval no. 89/2010.
The study was conducted on 30 New Zealandbreed rabbits, weighing 3-4 kg, from which speci-mens of the abdominal aorta were collected.
The experimental animals were euthanized byadministrating an intravenous solution of pento-barbital (prep. Morbital), and immediately afterdeath, 4-5 cm long specimens of the abdominalaorta were collected and cut into 1.5 cm long sec-tions. The sections were cleared from fat and con-nective tissue. The aorta diameter in a New Zealandbreed rabbit weighing 3-4 kg was on average 4 to 6 mm.The sections were set up in an automatic waterbath in four 20 ml chambers. The samples wereplaced horizontally by threading safil 4.0 surgicalthread inside the aorta lumen and mounted so thatchanges in the area of their transverse sectionswere registered.
All the samples were stretched to a tension of 5mN. The time required to balance the record wasdetermined experimentally at 40 minutes. TheKrebs-Henseleit buffer was used and the incuba-tion environment contained: NaCl – 118 mM; KCl– 4.7 mM; CaCl2 – 2.5 mM; MgSO4 – 1.6 mM;NaHCO3– 24.3 mM; KH2PO4– 1.18 mM; glucose– 5.6 mM (ECKERT 2000). Incubation of the sec-tions was carried out at a temperature of 37°C in agaseous mixture of oxygen and carbon dioxideused in the following proportions: 95% of O2 and5% of CO2, in order to obtain a pH value of 7.3-7.5.The aortic contractions were registered with iso-meric transducers (Letica Scientific Instruments)combined with bridge amplifiers (BridgeAmp,ADInstruments, Australia) and a 4-channel dataacquisition system (PowerLab/400, ADInstru-ments) connected to a Macintosh computer. Thespontaneous contractile activity of the aortic mus-cle was recorded for 30 minutes. Afterwards, theagonists and the antagonists of the adrenergic re-ceptors were introduced into the incubation cham-bers with an isolated section. The agonists werealways added to the chambers 40 minutes after theantagonists.
The following chemical substances were added:phenylephrine- á1-adrenergic receptor agonist(Sigma-Aldrich), xylazine- á2-adrenergic receptoragonist (Sigma-Aldrich), oxymetazoline- á2A-a-drenergic receptor agonist (Sigma-Aldrich), B-HT933- selective agonist of the á2 receptor (Sigma-Aldrich), phentolamine- á1-adrenergic receptorantagonist (Sigma-Aldrich), yohimbine- á2 recep-tor antagonist (Sigma-Aldrich) and RX821002-selective antagonist of the á2 receptor (Sigma-Aldrich).
J. GNUS et al.80

Doses of the preparations were established ex-perimentally by 10-fold dilutions. When definingthe experimental dose, we introduced the prepara-tion into the incubation chamber, starting with thehighest dilution and not washing the chamber be-tween the administration of subsequent doses(which led to the accumulation of the doses), untila visible effect was obtained on the chart. The ex-perimental dose was defined as the lowest concen-tration of preparation triggering the required effectand confirmed in a few subsequent experiments.
We evaluated the results by analysing thestrength of contractions expressed in mN (contrac-tility amplitude), time to tissue reaction (from ad-ministration to a visible effect) and time to maximaltonus. The results of the tests were processed withthe use of Microsoft Office Excel 2000 and analysedstatistically with Student’s t-test and a single-factor analysis of variance (ANOVA) for inde-pendent variables.
Results
The in vitro lifetime of rabbit abdominal aortasections amounted to 6-8 hours with no statisti-cally significant difference in contractile activity.After 30 minutes of control recording (duringwhich the strength of the muscle tone stabilisedand the metabolic activity of the tissue rose due totemperature and oxygenation increase), agonistsand antagonists of the adrenergic receptors wereadded to the incubation chamber, in a system of ac-cumulated doses, starting from the lowest concen-tration.
The results were processed with the strength ofcontractions expressed in mN (contractility ampli-tude), time to tissue reaction (from administrationto visible effect) and time to maximal tonus. De-tailed reactions of the rabbit abdominal aorta arepresented in the graphs: contractility amplitude(Figs 1, 2, 3), time to tissue reaction (Figs 4, 5, 6)and maximal tonus (Figs 7, 8).
Phenylephrine increased the smooth muscle to-nus in the rabbit abdominal aorta by an average4.75 mN at a concentration of 100 FM. In the caseof xylazine, a maximum increase in the muscle to-nus was observed at a concentration of 100 FM,and this amounted to 1.18 mN. In relation to themaximum response induced by the administrationof phenylephrine at a concentration of 100FM, themaximum muscle tonus resulting from the admini-stration of xylazine at the same concentration wasalmost four times smaller and equalled 24.8% ofthe initial tonus induced by phenylephrine. The ad-ministration of B-HT 933 at a concentration of 100FM caused a tonus increase constituting only 12%of the tonus recorded after the administration of
Effect of " Adrenoceptor stimulation on Aorta Contractility 81
Fig. 1. Relationship between the increase of rabbit abdominalaorta muscle tonus stimulated by selected á-adrenoceptoragonists and the agonist concentration, (*P#0.001,**P#0.01, ***P#0.05).
Fig. 2. Relationship between the increase of rabbit abdominalaorta muscle tonus stimulated by selected á2-adrenoceptoragonists and the agonist concentration. á2-receptors werepreviously blocked by 2·10-7M yohimbine administered tothe incubation chamber, (*P#0.001, **P#0.01, ***P#0.05).
Fig. 3. Relationship between tonus decrease (relaxation ofmuscle) of the rabbit abdominal aorta stimulated by selectedá1- and á2-adrenoceptor antagonists and the antagonistconcentration (*P#0.001, **P#0.01, ***P#0.05).
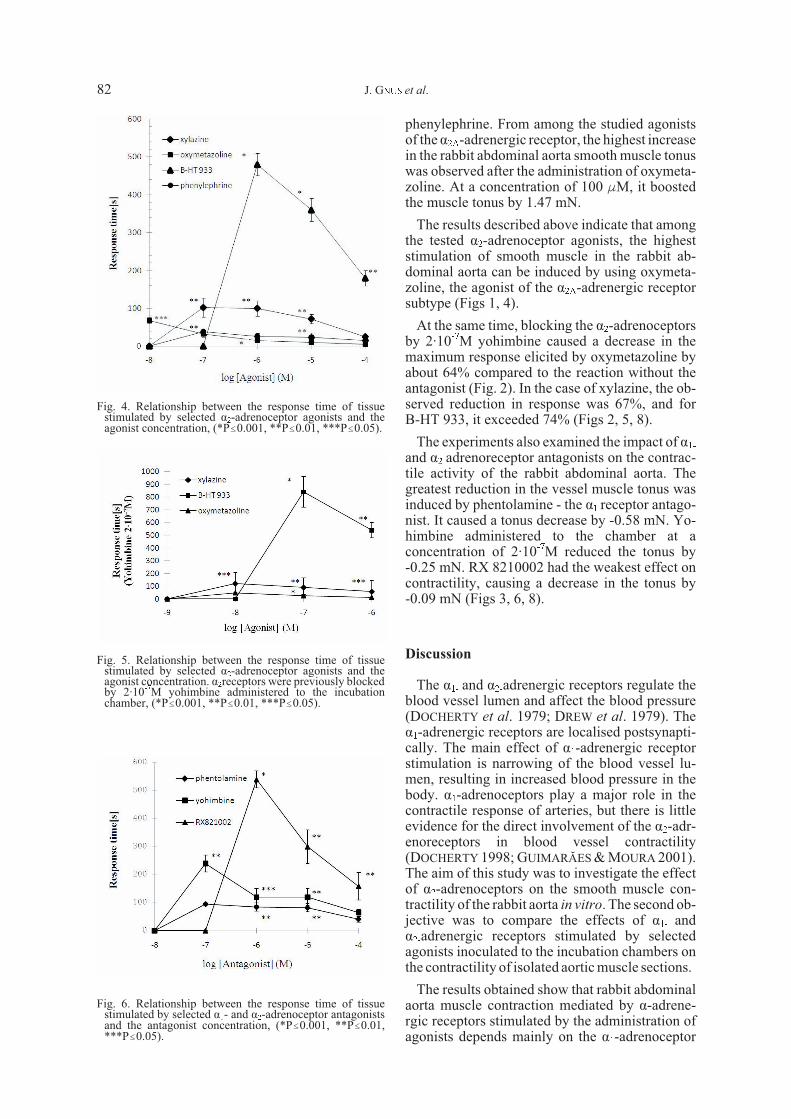
phenylephrine. From among the studied agonistsof the á2A-adrenergic receptor, the highest increasein the rabbit abdominal aorta smooth muscle tonuswas observed after the administration of oxymeta-zoline. At a concentration of 100 FM, it boostedthe muscle tonus by 1.47 mN.
The results described above indicate that amongthe tested á2-adrenoceptor agonists, the higheststimulation of smooth muscle in the rabbit ab-dominal aorta can be induced by using oxymeta-zoline, the agonist of the á2A-adrenergic receptorsubtype (Figs 1, 4).
At the same time, blocking the á2-adrenoceptorsby 2·10-7M yohimbine caused a decrease in themaximum response elicited by oxymetazoline byabout 64% compared to the reaction without theantagonist (Fig. 2). In the case of xylazine, the ob-served reduction in response was 67%, and forB-HT 933, it exceeded 74% (Figs 2, 5, 8).
The experiments also examined the impact of á1-and á2-adrenoreceptor antagonists on the contrac-tile activity of the rabbit abdominal aorta. Thegreatest reduction in the vessel muscle tonus wasinduced by phentolamine - the á1 receptor antago-nist. It caused a tonus decrease by -0.58 mN. Yo-himbine administered to the chamber at aconcentration of 2·10-7M reduced the tonus by-0.25 mN. RX 8210002 had the weakest effect oncontractility, causing a decrease in the tonus by-0.09 mN (Figs 3, 6, 8).
Discussion
The á1- and á2-adrenergic receptors regulate theblood vessel lumen and affect the blood pressure(DOCHERTY et al. 1979; DREW et al. 1979). Theá1-adrenergic receptors are localised postsynapti-cally. The main effect of á1-adrenergic receptorstimulation is narrowing of the blood vessel lu-men, resulting in increased blood pressure in thebody. á1-adrenoceptors play a major role in thecontractile response of arteries, but there is littleevidence for the direct involvement of the á2-adr-enoreceptors in blood vessel contractility(DOCHERTY 1998; GUIMARÃES & MOURA 2001).The aim of this study was to investigate the effectof á2-adrenoceptors on the smooth muscle con-tractility of the rabbit aorta in vitro. The second ob-jective was to compare the effects of á1- andá2-adrenergic receptors stimulated by selectedagonists inoculated to the incubation chambers onthe contractility of isolated aortic muscle sections.
The results obtained show that rabbit abdominalaorta muscle contraction mediated by á-adrene-rgic receptors stimulated by the administration ofagonists depends mainly on the á1-adrenoceptor
Fig. 4. Relationship between the response time of tissuestimulated by selected á2-adrenoceptor agonists and theagonist concentration, (*P#0.001, **P#0.01, ***P#0.05).
Fig. 5. Relationship between the response time of tissuestimulated by selected á2-adrenoceptor agonists and theagonist concentration. á2receptors were previously blockedby 2·10-7M yohimbine administered to the incubationchamber, (*P#0.001, **P#0.01, ***P#0.05).
Fig. 6. Relationship between the response time of tissuestimulated by selected á1- and á2-adrenoceptor antagonistsand the antagonist concentration, (*P#0.001, **P#0.01,***P#0.05).
J. GNUS et al.82
(Figs 1, 4). The administration of phenylephrine tothe incubation chamber evoked a much strongermuscle response than the administration of a simi-lar concentration of á2-adrenergic receptor ago-nists (Figs 1, 4). The response observed after thestimulation of the á2-adreneceptors was muchweaker. Compared to the maximum response in-duced by phenylephrine, the maximum contractileresponse induced by oxymetazoline representedapproximately 31%, and by B-HT 933, 12% of theactivity resulting from the stimulation of the á1-a-drenoceptors by phenylephrine (Fig. 1).
The largest increase in contractile activity in-duced by oxymetazoline may be due to the fact thatamong three á2-adrenoceptor subtypes, the á2A re-ceptors are the most involved in the norepi-nephrine-dependent contraction of blood vessels.At the same time, the concentration of á2A recep-tors in the blood vessels, both veins and arteries, ismuch higher than the concentration of other sub-types of adrenergic receptors. The dominant roleof the á2A subtype in the regulation of the cardio-vascular system has also been demonstrated in ex-
periments based on the deletion of genes encodingthe á2A-adrenoceptor subtype (BREDE et al. 2002).The deletion led to an increase in blood pressure,increased heart rate and susceptibility to heart fail-ure (BREDE et al. 2002). However, in tissues ex-pressing the á2A subtype, the presence of theá2-adrenoceptor agonists induced the narrowing ofblood vessels (MACMILLAN et al. 1996). Oxy-metazoline is not fully selective towards á2A-adr-enoreceptors, and it is also a partial agonist ofá1A-adrenoreceptors (MINNEMAN & ESBENSHADE1994) and 5-HT2A receptors (LACHNIT et al. 1997).Therefore, the aortic reaction to oksymetazolineshould be compared to that induced by UK14304,a full agonist of the á2-adrenergic receptors.
Stimulation of the receptors by xylazine induceda slightly weaker reaction than using oxymeta-zoline. The weakest contractility was evoked byB-HT 933. At the same time, after blocking the ad-renergic receptors, the greatest reduction in con-traction response was observed for B-HT 933.
An interesting reaction was observed after theadministration of yohimbine into the chamber(Figs 1, 2). With the exception of BHT-933, thereis no rightward shift of the curves in the presenceof yohimbine, but only a depression of the maxi-mal response. In numerous studies performed inother groups, yohimbine behaved as a competitiveantagonist of á2-adrenergic mediated contractileresponses. Our antagonism experiments demon-strated that the constrictor response to oxymeta-zoline and xylazine was inhibited by yohimbine ina non-competitive fashion, although the non-competitive antagonism by yohimbine is quite un-like the response of the rabbit artery to those drugs.The reason for this discrepancy is unclear and fur-ther studies will be needed in order to resolve thisphenomenon.
The second issue is that phentolamine, yo-himbine and RX82002 seem to be inverse agonistsin the rabbit abdominal aorta. This is explained bythe constitutive mobilisation of intracellular Ca2+and the inhibition of cAMP production. This coulddetermine the increase in the number of receptorsthat are spontaneously active in the absence of anagonist (LEFKOWITZ et al. 1993; BLACK &SHANKLEY1995; BONDet al. 1995; MILLIGANet al.1995). The antagonists bind to the receptor andchange it from an activated state to an inactivatedone or bind to inactive receptors and disrupt theequilibrium between the two states (active and in-active) in favour of the inactive state (SCHUTZ &FREISSMUTH 1992). New evidence suggests atwo-state model, in which the receptors are inequilibrium between the inactive conformationand a spontaneously active conformation that cou-ples to the G protein in the absence of the ligand.The first state can be fixed by inverse agonists
Fig. 7. Relationship between the time to reach maximumresponse of the tissue stimulated by selectedá2-adrenoceptor agonists and the agonist concentration.á2-receptors were previously blocked by 2·10-7 Myohimbine administered to the incubation chamber,(*P#0.001, **P#0.01, ***P#0.05).
Fig. 8. Relationship between the time to reach maximumresponse of the tissue stimulated by selected á1- andá2-adrenoceptor antagonists and the antagonistconcentration, (*P#0.001, **P#0.01, ***P#0.05).
Effect of " Adrenoceptor stimulation on Aorta Contractility 83

(BOND et al. 1995). The present work shows thatphentolamine, yohimbine and RX82002 do not actas antagonists but as inverse agonists; they inhibitthe increase in the resting tone in the absence of theagonist. In the case of yohimbine and RX82002,similar results were obtained by other authors(WADE et al. 2001). They concluded that severalfactors, such as the type of assay, receptor sub-types, cell type and local cellular G protein con-centrations, may affect constitutive receptoractivity.
The effects of the agonists and antagonists of theadrenergic receptors on the time to tissue reactionand the time necessary to reach the peak responseof the rabbit aorta were assessed in the same study.The quickest reaction was noticed after the ad-ministration of oxymetazoline and phenylephrineto the incubation chamber, whereas the slowest tis-sue response was induced by 933rd B-HT. A simi-lar relationship can be observed after blocking theaortic sections with yohimbine. The quickest aor-tic contractile response was recorded after the ad-ministration of oxymetazoline, and the slowestwas in the case of 933rd B-HT. Phenylephrine andoxymetazoline are similarly effective in triggeringthe contractile response of the investigated sec-tions. This may be due to a non-specific binding ofoxymetazoline to á1-adrenergic receptors, particu-larly at higher concentrations of the agonist.Moreover, the reaction time after the administra-tion of B-HT 933, a specific agonist of the á2-adre-nergic receptors, was approximately 10 timeshigher than for oksymetazoline. A similar situa-tion was found for antagonists; the time to tissuereaction after the administration of phentolaminewas about 7 times shorter than for RX821002, a se-lective antagonist of á2-adrenergic receptors. Thisdata indicates that the quickness of the rabbit ab-dominal aorta contractile reaction depends mainlyon the á1-adrenergic receptors, but later on, it isfurther enhanced by the activation of the á2-adre-nergic receptors.
Previous in vitro studies suggest that á1 recep-tors constitute the main factors involved innorepinephrine-dependent contractions in the ratand mouse mesenteric artery (HUSSAIN et al.2000). Other studies concerning the effect of á1-a-drenoceptor subtypes on the contractility of therabbit abdominal aorta show that the contractile re-sponse is mainly conditioned by postsynapticá1A/á1D subtypes (ABOUD et al. 1993; GUI-MARÃES & MOURA et al. 2001; GNUS et al. 2012).The available literature contains little evidence onthe direct effect of á2-adrenoceptors on the arterialcontractile response. It is currently assumed that aclear influence of the á2-adrenoceptors on vaso-constriction occurs in veins but not in arteries(GÖRNEMANN et al. 2007). Originally, this phe-
nomenon was described in pulmonary circulationin dogs (OHLSTEIN et al. 1989). Further studies re-vealed that the murine mesenteric vein was moresensitive to the vasoconstriction activity of theá-adrenoceptor agonists than the mesenteric ar-tery. It was also discovered that á-adrenergic re-ceptors in the veins were more resistant toagonist-induced desensitisation. The greater reac-tivity and resistance of á-adrenergic receptors todesensitisation may be important for blood pres-sure regulation. Studies conducted on the arteriesand veins in mice suggest the existence of differentcontraction mechanisms associated with the acti-vation of á-adrenoreceptors. The á1 receptors me-diate the contraction response in both veins andarteries, whereas the á2 receptors operate in theveins. Such differences in the function and phar-macology of the adrenoreceptors account for thedifferences in reactivity between arteries and veins.
Some articles show that the role of the á2-adren-oceptors in the blood vessel contractile response isbased on co-activation of both á1- and á2-adreno-ceptors (BYLUND et al.1988).
In summary, the á1-adrenergic receptors play amajor role in the regulation of rabbit aorta contrac-tility, but the á2-adrenergic receptors also affectthe tonus of the blood vessel muscular layer.
Acknowledgment
This publication is part of the project “Wrovasc– Integrated Cardiovascular Centre”, co-financedby the European Regional Development Fund,within the Innovative Economy Operational Pro-gramme, 2007-2013.“European Funds – for thedevelopment of innovative economy”
References
ABOUDR., SHAFIIM., DOCHERTY J.R. 1993. Investigation ofthe subtypes of á1-adrenoceptor mediating contractions ofrat aorta, vas deferens and spleen. Br. J. Pharmacol. 109:80-87.
ALTMAN J. D., TRENDELENBURG A. U., MACMILLAN L.,BERNSTEIN D., LIMBIRD L., STARKE K. 1999. Abnormalregulation of the sympathetic nervous system in alpha 2a-adrenergic receptor knockout mice. Mol. Pharmacol. 56:154-161.
AVERY R. A., FRANOWICZ J. S., STUDHOLME C., VANDYCKC. H., ARNSTEN A. F. 2000. The alpha 2a-adrenoreceptoragonist, guanfacine, increases regional cerebral blood flowin dorsolateral prefrontal cortex of monkeys performing aspatial working memory task. Neuropsychopharmacol. 23:240-249.
BLACK J. V., SHANKLEY N. P. 1995. Inverse agonists ex-posed. Nature 374: 214-215.
BOND R. A., LEFF P., JOHNSON T. D., MILANO C. A., ROCK-MANH. A., MCMINNT. R., APPARSUNDARAM S., HYEKM.F., KENAKIN T. P., ALLEN L. F., LEFKOWITZ R. J. 1995.Physiological effects of inverse agonists in transgenic mice
J. GNUS et al.84

with myocardial overexpression of the b2-adrenoceptor. Na-ture 374: 272-276.
BREDEM., WIESMANN F., JAHNSR., HADAMEK K., ARNOLTC., NEUBAUER S. 2002. Feedback inhibition of catechola-mine release by two different alpha 2-adrenoceptor subtypesprevents progression of heart failure. Circulation 106:2491-2496.
BRODDE O. E., MICHEL M. C. 1999. Adrenergic and muscar-inic receptors in the human heart. Pharmacol. Rev. 51: 651-90.
BYLUND D. B., RAY-PRENGER C., MURPHY T. J.1988. Alpha2a- and alpha 2b-adrenergic receptor subtypes: antagonistbinding in tissues and cell lines containing only one subtype.J. Pharmacol. Exp. Ther. 245: 600-607.
BYLUNDD. B. 1998. Subtypes of alpha 2-adrenoceptors: phar-macological and molecular biological evidence converge.Trends Pharmacol. Sci. 9: 356-361.
CHO S. C., KIM J. W., KIM B. N., HWANG J. W., PARK M.,KIM S. A.2008. Possible association of the alpha 2A-adrene-rgic receptor gene with response time receptor gene with re-sponse time disorders. Am. J. Med. Genet. B.Neuropsychiatr. Genet. 5: 957-963
CHOTANI M. A., FLAVAHAN S., MITRA S., DAUNT D., FLA-VAHAN N. A. 2000. Silent alpha (2c)-adrenergic receptorsenable cold-induced vasoconstriction in cutaneous arteries.Am. J. Physiol. Heart Circ. Physiol. 278: 1075-1083.
DOCHERTY J. R., MCGRATH J. C. 1979. Inhibition of sympa-thetic transmission in rat heart byclonidine: the roles ofstimulation frequency, endogenous feedback andnoradrenalinere-uptake. Naunyn Schmiedebergs Arch.Pharmacol. 309: 225-233.
DOCHERTY J. R. 1998. Subtypes of functional alpha 1- and al-pha 2-adrenoceptors. Eur. J. Pharmacol. 361: 1-15.
DREW G. M., WHITING S. B. 1979. Evidence for two distincttypes of postsynaptic alpha adrenoceptor in vascular smoothmuscle in vivo. Br. J. Pharmacol. 67: 207-15.
ECKERT R. E., KARSTEN A. J., UTZ J., ZIEGLER M. 2000.Regulation of renal artery smooth muscle tone by alpha1-adrenoceptors: role of voltage-gated calcium channels andintracellular calcium stores. Urol Res. 28:122-127.
GASPAR R., GAL A., GALIK M., DUCZA E., MINORICS R.,KOLAROVSZKI-SIPICZKI Z. 2007. Different roles of alpha2-adrenoceptor subtypes in non-pregnant and late-pregnantuterine contractility in vitro in the rat. Neurochem. Int. 51:311-318.
GAVIN K. T., COLGAN M. P., MOORE D., SHANIK G.,DOCHERTY J. R.1997. Alpha 2c-adrenoceptors mediate con-tractile responses to noradrenaline in the human saphenousvein. Naunyn Schmiedebergs Arch. Pharmacol. 355:406-411.
GNUS J., CZERSKIA., ZAWADZKIW., WITKIEWICZ W., HAU-ZERW., RUSIECKAA., FERENCS. 2012. in vitro contractilityof normal and aneurismal abdominal aorta muscle coat sec-tions in human and animal material. Folia Biol. (Kraków)60: 71-77.
GÖRNEMANN T., VON WENCKSTERN H., KLEUSER B.,VILLÜNC. M., CENTURIÜND., JÄHNICHENS.2007. Charac-terisation of the postjunctional alpha 2c-adrenoreceptor me-diating vasoconstrictor to UK 14304 in porcine pulmonaryveins. Br. J. Pharmacol. 151: 186-194.
GUIMARÃES S., MOURA D. 2001. Vascular adrenoreceptors:an update. Pharmacol. Rev. 53: 319-356.
GYIRES K., ZÀDORI Z. S., TÖRÖK T., MÀTYUS P. 2009. á2-adrenoreceptor subtypes-mediated physiological, pharma-cological actions. Neurchem. Internat. 55: 447-453.
HANC., ABEL P. W., MINNEMAN K. P. 1987. ál-adrenoceptorsubtypes linked to different mechanisms for increasing intra-cellular Ca2+ in smooth muscle. Nature 329: 333-335.
HEIN L., KOBILKA B. K. 1995. Adrenergic receptor signaltransduction and regulation. Neuropharmacology 34: 357-366.
HIEBLE J. P., BONDINELL W. E., RUFFOLO R. J. 1995. á- andâ-adrenoceptors: from the gene to the clinic. Molecular biol-ogy and adrenoceptor subclassifcation. J. Med. Chem. 38:3415-3444.
HIEBLE J. P., RUFFOLO J. R. 1996. Subclassification and no-menclature of á1- and á2-adrenoceptors. Prog. Drug. Res. 47:81-130.
HUSSAIN M. B., MARSHALL I. 2000. á1– Adrenoreceptorsubtypes mediating contractions of the rat mesentericarter-ies. Eur. J. Pharmacol. 395: 69-76.
LACHNIT W. G., TRAN A. M., CLARKE D. E., FORD A. P.1997. Pharmacological characterization of an a1A-adrenoceptor mediating contractile responses to noradrena-line in isolated caudal artery of rat. Br. J. Pharmacol.120:819-826.
LANGER S. Z. 1999. History and nomenclature of á1-adren-oceptors. Eur. Urol. 36: 2-6.
LEFKOVITZ R. J., COTECCHIA S., SAMANA P., COSTA T.1993. Constitutive activity of receptors coupled to guaninenucleotide regulatory proteins. Trends Pharmacol. Sci.14:303-307.
LOMASNEY J. W., COTECCHIA S., LORENZW., LEUNGW. Y.,SCHWINN D. A., YANG-FENG T. L.1991. Molecular cloningand expression of the cDNA for the alpha 1A-adrenergic re-ceptor. The gene for which is located on human chromosome5. J. Biol. Chem. 266: 6365-6369.
MACMILLAN L. B., HEIN L., SMITH M. S., PIASCIK M. T.,LIMBIRD L. E. 1996. Central hypotensive effects of thealpha2a-adrenergic receptor subtype. Science 273: 801-803.
MILLIGAN G., BOND R. A., LEE M. 1995. Inverse agonism:pharmacological curiosity or potential therapeutic strategy?.Trends Pharmacol. Sci. 16: 10-13.
MINNEMAN K. 1988. Alpha 1- adrenergic receptor subtypes.Inositol phosphates and sources of cell calcium. Pharmacol.Rev. 40: 87-93.
MINNEMAN K. P., ESBENSHADE T. A. 1994. Alpha 1-adrenergic receptor subtypes. Annu. Rev. Pharmacol. Toxi-col. 34: 117-133.
OHLSTEIN E. H., HOROHONICH S., SHEBUSKI R. J., RUFFOLOR. R.1989. Localization and characterization of alpha-2 ad-renoreceptors in the isolated canine pulmonary vein. J. Phar-macol. Exp. Ther. 248: 233-239.
OSSWALD W., GUIMARAES S. 1983. Adrenergic mechanismsin blood vessels: morphological and pharmacological as-pects. Rev. Physiol. Biochem. Pharmacol. 96: 53-122.
PHILIPP M., BREDE M., HEIN L. 2002. Physiological signifi-cance of alpha(2)-adrenergic receptor subtype diversity: onereceptor is not enough. Am. J. Physiol. Regul. Integr. Comp.Physiol. 283: 287-295.
SCHUTZ W., FREISSMUTH M. 1992. Reverse intrinsic activityof antagonists on G protein-coupled receptors. Trends Phar-macol. Sci. 13: 376-379.
SMITH K., MACmILLAN J. B., MCGRATH J. C. 1997. Investi-gations of á1-adrenoceptor subtype mediating vasoconstric-tion in rabbit cutaneous resistance arteries. Br. J. Pharmacol.122: 825-832.
SUZUKI E., TSUJIMOTO G., TAMURA K., HASHIMOTO K.1990. Two pharmacologically distinct alpha 1-adrenoreceptor subtypes In the contraction of rabbit aorta:each subtype couples with a different Ca2+signaling mecha-nism and plays a different physiological role. Mol. Pharma-col. 38: 725-730.
WADE S. M., LAN K., MOORE D. J., NEUBIG R. R. 2001. In-verse agonist activity at the alpha(2A)-adrenergic receptor.Mol. Pharmacol. 59: 532-542.
Effect of " Adrenoceptor stimulation on Aorta Contractility 85























![Adrenoceptor Agents [Compatibility Mode]](https://img.dokumen.tips/doc/110x75/577d26cc1a28ab4e1ea236f9/adrenoceptor-agents-compatibility-mode.jpg)





