Embed Size (px)
Citation preview

발간등록번호: 11-1480592-001055-01
NIBR201530101
이산화탄소 생물전환을 위한 자생 유용
미세조류 발굴 1차년도
Investigation of Indigenous Useful
Microalgae for CO2 Bioconversion 1
2015
국립생물자원관National Institute of Biological Resources

이산화탄소 생물전환을 위한 자생 유용
미세조류 발굴 1차년도
생물자원활용부 유용자원분석과
김지형, 신나래, 배창환
Investigation of Indigenous Useful
Microalgae for CO2 Bioconversion 1
Ji Hyung Kim, Narae Shin, Chang-Hwan Bae
Biological and Genetic Resources Assessment Division
Biological Resources Utilization Department
National Institute of Biological Resources
2015
국립생물자원관National Institute of Biological Resources

소속 연구분야 담당자
유용자원분석과연구사업 총괄
미세조류 확보 및 분석
김지형
(총괄연구책임자)
미세조류 확보 배창환
미세조류 분리·동정 및 배양 신나래
미생물자원과 미세조류 형태동정 김진희
미세조류 형태동정 윤지혜
한국해양과학기술원 미세조류 성분분석 강도형
미세조류 성분분석 허수진
미세조류 성분분석 오철홍
미세조류 최적 배양조건 확립 김태호
미세조류 최적 배양조건 확립 최우석
미세조류 최적 배양조건 확립 이경아
서울대학교 미세조류 배양 박세창
미세조류 배양 윤새길
미세조류 배양 김현중
참여 연구진

- i -
요 약 문
1. 제 목
이산화탄소 생물전환을 위한 자생 유용 미세조류 발굴 1차년도
2. 연구목적
이산화탄소 생물전환이 가능한 자생 미세조류의 유용성분 및 배양능 평가를
통하여 CO2 저감 및 식품, 의약품의 소재로 활용 가능한 유용 미세조류주를 발굴
하여, 환경정화 및 기능성 생물소재로써의 활용도를 평가
3. 연구내용 및 방법
가. 수생태계 유래 자생 미세조류 배양체 확보
1) 수계 환경시료내미세조류 순수분리 및 증식배지 내 분리주배양가능성확인
2) 배양 가능 미세조류의 형태학적 분류․동정
3) 유전자 염기서열 기반 분자생물학적 분류․동정
나. 자생 미세조류 유용성 검증
1) 미세조류 바이오매스 확보
2) 미세조류 일반성분 분석 및 평가
3) 미세조류 세부성분 분석 및 평가
다. 유용 미세조류 선별 및 배양기법 탐색
1) 유용성 검증결과 기반 유용 미세조류주 선별
2) 성장률 기반 최적 배양조건 탐색
4. 연구결과
가. 수생태계 유래 자생 미세조류 배양체 확보(10주)
1) 배양 가능 자생 미세조류 10주 배양체 확보
가) 다양한 수계 환경(담수, 기수 및 해수) 시료 내 미세조류 세포 분리 통한
50주 이상의 미세조류 세포 순수 분리
나) 증식배지(BG-11, f/2, BBM, Zarrouk's medium 등) 내 배양능 검정
통한 미세조류주 10주 배양체 확보

- ii -
2) 미세조류 배양체 10주의 형태학적 분류․동정 결과, 남조류(Cyanophyta)
5주 및 녹조류(Chlorophyta) 5주로 판별
가) 남조류: Section III(Oscillatoriales) 4주 및 Section II(Pleurocapsales) 1주
나) 녹조류: Chlorophyceae 강(class) 5주
3) 유전자 염기서열 기반 분류․동정 결과, 16S rRNA 염기서열 결과 기반
남조류 4속 및 18S rRNA/ITS 염기서열 결과 기반 녹조류 5속 판별
가) 남조류: Arthrospira 속 2주, Leptolyngbya 속 1주, Myxosarcina 속 1주,
Lyngbya 속 1주
나) 녹조류: Chlamydomonas 속 1주, Haematococcus 속 1주, Lobosphaera 속 1주,
Scenedesmus 속 1주, Chlorella 속 1주
나. 자생 미세조류 유용성 검증(10주)
1) 미세조류 바이오매스 확보(10주)
가) 5L급 진탕배양을 통한 미세조류 바이오매스(건조중량 10g 이상) 확보
2) 미세조류 일반성분 분석 및 평가(10주)
가) 미세조류주 조단백, 조지방, 조탄수화물, 회분 및 수분 함량 분석 및 평가
3) 미세조류 세부성분 분석 및 평가(10주)
가) 미세조류주 아미노산, 지방산 및 당 조성 분석 및 평가
다. 유용 미세조류 선별 및 배양기법 탐색(3주)
1) 유용성 검증결과 기반 유용 미세조류주 선별(남조류 2주, 녹조류 1주)
가) 남조류: Leptolyngbya 속; 조단백 함량 55% 이상, 고도 성장능, 무독성
Lyngbya 속; 조단백 함량 70% 이상, 고도 성장능, 거대 구조
나) 녹조류: Lobosphaera 속; 특이 불포화지방산(Arachidonic acid) 생산
2) 배양조건(pH, 온도 및 염도) 변화에 따른 성장률 기반 최적 배양조건 탐색(3주)
가) 남조류: Leptolyngbya 속; pH(5-9), 온도(25-35°C), 염도(0-30psu)
Lyngbya 속; pH(5-9), 온도(25-35°C), 염도(0-30psu)
나) 녹조류: Lobosphaera 속; pH(6-8), 온도(25-30°C), 염도(0-20psu)
5. 연구결과의 활용에 대한 건의
가. 발굴된 자생 유용 미세조류는 향후 탄소 배출권 확보를 위한 친환경적 CO2
저감방안 개발 및 유용 천연물 소재 확보를 위한 기반자료로 활용
나. 자생 미세조류 발굴을 통하여 국가 생물다양성을 확보하며, 유용성이 검증된
미세조류는 향후 산업재산권 확보를 통한 국가 고유 생물자원으로 활용

- iii -
목 차
요약문 ····················································································································· i
목차 ······················································································································ iii
표 목차 ················································································································ iv
그림 목차 ·············································································································v
Abstract ·············································································································vii
Ⅰ. 서론 ·················································································································1
Ⅱ. 연구방법 ·······································································································11
1. 수생태계 유래 자생 미세조류 배양체 확보 ·······································13
2. 자생 미세조류 유용성 검증 ···································································18
3. 유용 미세조류 선별 및 배양기법 탐색 ···············································22
III. 연구결과 ·······································································································25
1. 수생태계 유래 자생 미세조류 배양체 확보 ·······································27
2. 자생 미세조류 유용성 검증 ···································································61
3. 유용 미세조류 선별 및 배양기법 탐색 ···············································75
IV. 고찰 ··············································································································87
V. 참고문헌 ········································································································98
부록 I. 미세조류 배양체 염기서열 ·····························································101
부록 II. 미세조류 배양용 영양배지 조성 ··················································115

- iv -
표 목 차
Ⅰ. 서론
표 I-1. 유용성이 검증된 미세조류(남조류 및 녹조류) 현황 ································7
표 I-2. 녹조류(Chlorella) 및 남조류(Spirulina)의 아미노산 함량 비교 예 ······ 9
Ⅱ. 연구방법
표 II-1. 염기서열 분석용 Oligonucleotide primer 정보 ······································17
표 II-2. 유리 아미노산 조성 분석조건 ····································································20
표 II-3. 유리 당 조성 분석조건 ················································································21
표 II-4. 유용 미세조류주 선정조건 및 산정기준 ··················································22
Ⅲ. 연구결과
표 III-1. 분리된 자생 미세조류주(NIBRUIM) 현황 ·············································27
표 III-2. 자생 미세조류주 배양조건 및 최종 생산량 ···········································61
표 III-3. 자생 남조류 일반성분 분석 결과 ·····························································62
표 III-4. 자생 녹조류 일반성분 분석 결과 ·····························································64
표 III-5. 자생 남조류 당분석 결과 ···········································································73
표 III-6. 자생 녹조류 당분석 결과 ···········································································74
표 III-7. 유용성 검증결과 기반 자생 남조류주 선정 결과 ·································75
표 III-8. 유용성 검증결과 기반 자생 녹조류주 선정 결과 ·································77

- v -
그 림 목 차
Ⅰ. 서론
그림 I-1. 남조류(Spirulina)의 생산·성분분석 및 활용성 분석 ··························8
Ⅱ. 연구방법
Ⅲ. 연구결과
그림 III-1. Arthrospira sp. NIBRIUM-1의 형태 분석 ·······································28
그림 III-2. Arthrospira sp. NIBRIUM-1의 계통분류도 ·····································30
그림 III-3. Arthrospira sp. NIBRIUM-2의 형태 분석 ········································31
그림 III-4. Arthrospira sp. NIBRIUM-2의 계통분류도 ·····································33
그림 III-5. Leptolyngbya sp. NIBRIUM-3의 형태 분석 ·····································34
그림 III-6. Leptolyngbya sp. NIBRIUM-3의 계통분류도·····································37
그림 III-7. Myxosarcina sp. NIBRIUM-4의 형태 분석 ······································38
그림 III-8. Myxosarcina sp. NIBRIUM-4의 계통분류도 ···································40
그림 III-9. Lyngbya sp. NIBRIUM-5의 형태 분석 ·············································41
그림 III-10. Lyngbya sp. NIBRIUM-5의 계통분류도 ·········································43
그림 III-11. Chlamydomonas sp. NIBRIUM-6의 형태 분석 ······························44
그림 III-12. Chlamydomonas sp. NIBRIUM-6의 계통분류도 ···························47
그림 III-13. Haematococcus sp. NIBRIUM-7의 형태 분석 ······························48
그림 III-14. Haematococcus sp. NIBRIUM-7의 계통분류도 ····························51
그림 III-15. Lobosphaera sp. NIBRIUM-8의 형태 분석 ····································52
그림 III-16. Lobosphaera sp. NIBRIUM-8의 계통분류도 ··································54
그림 III-17. Scenedesmus sp. NIBRIUM-9의 형태 분석 ··································55
그림 III-18. Scenedesmus sp. NIBRIUM-9의 계통분류도 ·································57
그림 III-19. Chlorella sp. NIBRIUM-10의 형태 분석 ········································58
그림 III-20. Chlorella sp. NIBRIUM-10의 계통분류도 ······································60
그림 III-21. 자생 남조류 5주의 일반성분 분석결과················································62
그림 III-22. 자생 녹조류 5주의 일반성분 분석결과················································64
그림 III-23. 자생 남조류 5주의 아미노산 조성 분석결과·······································66
그림 III-24. 자생 녹조류 5주의 아미노산 조성 분석결과·······································68
그림 III-25. 자생 남조류 5주의 세부 지방산 조성 분석결과·································69

- vi -
그림 III-26. 자생 남조류 5주의 지방산 함량 비교결과 ·······································70
그림 III-27. 자생 녹조류 5주의 세부 지방산 조성 분석결과·································71
그림 III-28. 자생 녹조류 5주의 지방산 함량 비교결과··········································72
그림 III-29. Leptolyngbya sp. NIBRIUM-3의 최적 배양조건 결과 ················81
그림 III-30. Lyngbya sp. NIBRIUM-5의 최적 배양조건 결과 ·························83
그림 III-31. Lobosphaera sp. NIBRIUM-8의 최적 배양조건 결과 ··················85

Abstract
Recently, microalgae(including cyanobacteria) hold promise as producers of
sustainable biomass for CO2 fixations as well as for the production of
biofuels and other biomaterials. However, the selection of microalgal strains
with efficient and robust production of desirable resources remains
challenging due to low productivity and potential toxicity. In this study, we
isolated several indigenous microalgal strains from various aquatic regions of
Korea including freshwater, estuarine water and seawater, and evaluated their
potential for the production of valuable bioresources. From the several
collected microalgal isolates, 10 mass-culturable microalgal strains were
preliminarily screened and identified. Based on the morphological
classifications, 4 and 1 strains were respectively identified as cyanobacteria
belong to section III(Oscillatoriales) and section II(Pleurocapsales), whereas 5
strains were identified as eukaryotic microalgae belong to Chlorophyceae.
Molecular phylogenetic analyses revealed that the 5 cyanobacterial strains
were finally classified into the genus Arthrospira(2 strains), Leptolyngbya(1
strain), Myxosarcina(1 strain) and Lyngbya(1 strain). Moreover, the 5
eukaryotic microalgal isolates were classified into the genus Chlamydomonas(1
strain), Haematococcus(1 strain), Lobosphaera(1 strain), Scenedesmus(1 strain)
and Chlorella(1 strain). To determine the usefulness of the microalgal strains,
the proportions of the microalgal cellular components(Ash, carbohydrate, lipid,
moisture and protein) including the detailed composition of amino acids,
monosaccharides and fatty acids were examined, respectively. Among the 10
selected culturable microalgal isolates, 2 strains of cyanobacteria(Leptolyngbya
and Lyngbya) with high protein composition and high biomass productivity,
and 1 strain of chlorophyta(Lobosphaera) with valuable fatty acids
composition were chosen for further culturability tests in various
environmental conditions. And finally, the optimum growth conditions for the
selected 3 microalgal strains were obtained. Based on these results, it can be
suggested that the newly isolated 3 indigenous microalgal strains will have
strong potential as a microalgal bioresource due to its valuable cellular
components and/or biomass productivity compared with other reported
microalgal species from other countries.

- 1 -
Ⅰ. 서 론


- 3 -
I. 서론
1. 연구의 필요성
가. 배경
1) 국가 고유 생물자원 발굴 및 생물주권 확보
생물자원은 배양가능한 생물체와 복제 가능한 생물체의 부분, 배양 불가능한
생물체 등의 분자, 생리, 유전자 및 구조적 정보까지 포함하여 실제적 활용가치가
있는 실물자원으로 정의한다. 이러한 생물자원의 범위는 인간유래자원, 동물, 식물,
미생물(균류, 미세조류, 원핵생물, 원생생물, 바이러스 등)로 의료, 농림수산,
식품, 향장, 에너지, 환경, 화학산업 등 다양한 분야의 원천소재로 사용된다.
1992년 생물다양성 협약의 발효로 인해 생물자원은 인류공동의 자산
이라는 개념에서 보유국의 주권적 권리라는 개념으로 전환되었으며, 2010년
10월 “유전자원의 접근 및 이익 공유(Access to Genetic Resources and
Benefit Sharing)에 관한 나고야 의정서” 채택으로 생물유전자원을 이용하기
위해서는 유전자원 제공국의 사전승인을 구해야 하며, 이용방법 및 정당한
이익 공유 조건에 관한 상호합의를 할 것으로 규정하고 있다. 이에 따라, 자국의
생물자원을 발굴하여 생물주권의 근거를 확보하고, 유용한 생물자원을 선점
하기 위한 국가 간의 경쟁이 치열해지고 있다. 이러한 시점에서 자생생물의
발굴 뿐 만 아니라 그 유전적 근거를 확보하고, 해외 혹은 근연 분류군과의
비교분석을 통한 우리나라 생물자원의 특이성 규명을 위한 연구는 나고야 의정서
발효 뒤 본격적인 국가 간 생물자원 전쟁 국면에서 자생생물에 대한 주권을
지키기 위한 근거자료로 필수불가결한 요소라 할 수 있다.
대한민국은 기후 및 지리적 특성상 좁은 면적에도 불구하고 많은 생물종을
가지고 있는 생물자원의 보고로, 총 10만종 정도가 존재하는 것으로 추산되며,
2014년까지 약 42,000종이 발굴․보고되었다. 그러나 이러한 생물자원의 발굴
관련 연구는 대부분 야생 동․식물로 한정되어 있으며, 최근 전 세계적으로
활용성 연구가 강화되고 있는 미생물 자원과 관련되어서는 아직까지 체계적
발굴 및 활용성 연구가 특히 미진하다. 따라서 자생 미생물 자원 발굴에 대한
국가적인 연구가 시급한 실정이며, 더불어 활용성이 있는 미생물 유래 유전
자원 확보․관리 및 향후 특정 미생물종의 보존 및 관리 정책을 수립하는데

- 4 -
필요한 다양한 정보를 확보할 필요가 있다. 특히, 유전다양성은 변화무쌍한
환경에 유연하게 적응하기 위한 자생 미생물종의 적응성과 밀접히 관련되어,
특정 유용종의 관리정책 수립을 위해서는 자생 미생물 군집 변화 모니터링
및 유전적 다양성 분석이 필수적으로 수행될 필요가 있다.
2) 이산화탄소 배출에 따른 기후변화
현재 우리가 사용하고 있는 화석연료는 유기물의 장기적 순환과정을 통해
생성된 산물이며, 탄소순환에 의해 화석연료를 사용하고 있다. 기존 자연 상태
에서 배출되던 탄소화합물은 산업혁명 이후 급격한 화석연료의 사용으로 현재
연간 70-80억 톤이 생성되고 있으며, 이중 약 30억 톤이 광합성 생물에 의해
고정되고 있는 반면 매년 30-40억 톤의 탄소가 대기 중으로 방출되고 있다.
미국 해양대기청(NOAA)은 지난 40만년 동안 180-300 ppm사이를 반복하던
지구 대기 중 이산화탄소 농도가 최근 50년간 무려 100 ppm 가까이 급격하게
상승하여 현재는 400 ppm에 근접하고 있다고 보고하고 있다. 전 지구적 화석
연료 소비증가에 따라 배출되는 이산화탄소의 양도 꾸준히 증가할 것으로 예상
되며, 인류의 산업 활동에 의한 온실가스의 증가는 지구에서 방사되는 열을
재흡수함으로써 대기 온도를 상승시켜 21세기 말 극지의 온도는 현재보다
7-8°C 이상 상승할 것으로 예측된다(Peterson & Baringer, 2008).
재생 가능한 바이오연료는 화석연료에 비해 상대적으로 적은 양의 온실가스를
배출하며, 바이오연료의 원료식물들은 대기 중 이산화탄소를 이용하여 광합성
을 하고 산소를 생산하므로 탄소 중립인 풍력, 조력, 지열, 태양열/태양광 등
의 다른 신·재생에너지보다 유리한 특성을 지닌다. 특히 현재 연구되고 있는
신재생에너지 부문 중 수송 분야는 액체연료가 당분간 사용되어야 할 것으로
판단되어 바이오연료의 중요성은 더욱 부각되고 있다. 현 시점에서 재생가능
하고 지속가능한 미세조류 유래 바이오연료 자원은 이산화탄소 배출량 감축과
같은 환경 문제에 대한 해결방안을 기대할 수 있는 대체 자원으로 주목받고
있다.
3) 곡물가격 상승 및 식량부족
20세기부터 지속되어 온 선진국의 육상작물 이용 바이오연료 개발은 상기
기술한 여러 장점과 중요성에도 불구하고, 기후변화에 의한 곡물가격 상승과

- 5 -
식량가격 폭등 및 개발도상국과 후진국의 식량공급 부족을 초래하고 있다.
육상식물의 바이오연료 원료 수급에 의한 국제 곡물가격 상승은 바이오연료를
생산하기 위해 꾸준히 증가하고 있는 식용작물 생산과 밀접한 관계가 있다.
예시로 브라질 및 미국의 원유가격 상승에 따른 바이오에탄올 생산 증대로
인해 사탕수수와 옥수수가 많이 사용되면서, 국제 식량가격이 폭등하고 있다.
결론적으로 많은 연구자들과 환경관련 단체들은 비식용작물을 이용한
바이오연료 생산이 현재의 화석연료고갈에 의한 고유가 문제와 기후변화에
따른 환경보전 문제를 동시에 해결할 수 있는 환경 친화적 대안으로 제시한다.
4) 화석연료 고갈
비산유국인 대한민국의 경우, 지난 수년간 에너지 수입액은 급증하였으며
이에 따른 에너지 수입 의존도는 97%에 달한다(2004년 496억 달러⇒2013년
1787억 달러 수입). 이러한 에너지 수요의 급격한 증가는 전 세계적인 추세로,
이는 지난 30여 년간 배럴당 10-40달러에서 상대적으로 안정되었던 국제 유가가
최근 10여 년 사이 급등한 것에서 확인할 수 있다(2013 에너지통계연보, 2013).
2008년 7월 이후부터 현재까지 유가의 상승세가 세계 소비시장의 감소와 셰일
가스의 개발로 다소 감소하였으나, 추후 가격 재상승이 예상되고 있는 실정이다.
급격한 화석연료 사용량의 증가로 인해 세계 모든 지역의 원유매장량이 급격
하게 줄어들고 있을 뿐만 아니라 산유국에의한생산량조절에따라원유가 상승은
가속화될 전망이다.
국가적 전기수요 및 산업 발전에 따라 석유, 석탄, 천연가스 등 화석에너지
소비량은 앞으로도 계속 증가할 것으로 전망되고 있지만, 고갈되어 가는 화석
연료 때문에 이들을 대체할 만한 고유 생물자원 유래 신재생연료의 개발이 시급한
것이 현실이다.
나. 기술적 측면
1) 미세조류(Microalgae)의 정의
미세조류(Microalgae)는 ‘물에 떠다니는 방랑자’로 풀이되는 플랑크톤
(Plankton) 중 식물성플랑크톤으로분류되는분류군으로한정된다. 미세조류는 대부분
광합성 색소를 가지고 독립영양생활을 하며, 서식 장소에 따라서 담수조류(淡水

- 6 -
藻類), 기수조류(汽水藻類) 및 해조류(海藻類) 등으로 구분한다. 일반적으로 우리가
일컫는 미세조류는 분류학적으로는 보통 9개의 문(門)으로 나누어지며, 원핵생물에
속하는 남조류(藍藻類: Cyanophyta)와 진핵생물에 속하는 규조류(硅藻類:
Bacillariophyta) 및 녹조류(綠藻類: Chlorophyta) 등으로 대표된다. 이러한 미세
조류는 지구상에서 가장 오래된 생물이며, 35억년 이전에 생성된 스트로마톨라이트
(stromatolite)로 불리는 화석에서 남조류가 발견되기도 한다. 따라서 미세조류는
최초의 광합성 능력을 지닌 미생물로써 담수계 및 해양 내 먹이사슬의 기초생물로
알려지고 있으며, 광합성에 의한 단위면적당 생산량이 육상작물에 비해 훨씬 높고,
유용 물질들을 함유하고 있는 것으로 보고된다. 이러한 미세조류는 대기 및 수중의
이산화탄소를 탄소원으로 이용하는 광합성을 통해 산소를 방출하며 단백질,
탄수화물, 지질 및 비타민 등 유기양분을 합성하고 이산화탄소 고정화 작용으로
기후변화에 영향을 미치는 이산화탄소를 대량으로 저감할 수 있는 활용성을
지니고 있다.
2) 미세조류(Microalgae)의 활용성
20세기 중반부터 미세조류는 ①수생 생태계의 기초 생산자로써 생태계의
생물 다양성 및 건강성을 확보하기 위해, ②동·식물 및 기타 미생물로부터 생산
불가능한 다양한 유용 천연물질 및 화합물 생산을 위해, ③바이오매스 기반 바이오
디젤 및 바이오에탄올 등의 신재생 바이오에너지 생산을 위한 생물 플랫폼
(Platform)으로써 활발히 연구가 진행되고 있다. 이중 다량의 단백질, 탄수화물
및 지질을 포함하고 있으며 비타민(프로비타민 A, 비타민 E, 티아민, 코발라민,
비오틴), 불포화 지방산, 철분, 베타카로틴, 클로로필 등의 유용물질을 생산하는
것으로 알려진 Botryococcus, Chlamydomonas, Chlorella, Dunaliella,
Haematococcus, Spirulina (Arthrospira), Nostoc 속 등의 미세조류는 활발히
연구되고 있다(Cohen, 1986)(표 I-1). 먼저 Botryococcus, Chlorella 및
Haematococcus 속 등의 진핵 녹조류는 바이오디젤로 전환 가능한 다량의 지질
(특히 triacylglycerol) 및 바이오에탄올 발효과정의 원료가 될 수 있는 다량의
탄수화물(특히 glucose)을 함유하고 있어 바이오에너지를 생산하기 위한 제3세
대 바이오매스로 연구되고 있으며, 기존의 곡물·식물유 기반 1세대 바이오매스
및 목질계 기반 2세대 바이오매스를 대체할 수 있는 가능성을 지닌다. 또한 베타
카로틴 및 클로로필 등의 색소물질을 비롯한 다양한 유용물질을 생산하여 건강
식품, 수산양식용 사료, 의약품, 화장품 생산 등을 위한 생물자원으로 중요성을
인정받고 있다.

- 7 -
종명 특성 유용물질 생산 및 활용 Reference
남조류(Cyanophyta)
Spirulinamaxima
수확용이(거대 코일형),
배양 온도 높음, 고탄소
배지 요구
건강식품 (단백질, 지질,
비타민, 미네랄), 양식사료 및
바이오에너지
Qiang &
Richmond, 1996
Spirulinaplatenesis
수확용이(거대 코일형),
배양 온도 높음,
고탄소 배지 요구
건강식품 (단백질, 지질,
비타민, 미네랄), 양식사료 및
바이오에너지 생산
Qiang &
Richmond, 1996
Lyngbya sp.수확용이(거대 선형),
온도 적응성 높음
Dolostatin 계열 항암물질
생산
Luesch et al.
2002
Nostoc sp.수확용이(선형),
온도 적응성 높음
건강식품 (단백질, 지질,
미네랄) 및 양식사료 Facciola, 1998
Porphyridiumaerugineum
수확용이(선형),
온도 적응성 높음
색소 (피코에리틴), 다당류,
지방산 등 유용물질 생산Cohen &
Cohen, 1991
Leptolyngbyasp.
수확용이(선형), 온도,
pH 및 염도 적응성 높음
고단백 및 고지질 함량으로
식품첨가물 가능Kim et al.,
2015
녹조류(Chlorophyta)
Botryococcusbraunii
고지질 함량(>50%),
대량배양 난해
미세조류 바이오디젤 생산용
모델 균주
Baba et al.,
2012
Chlamydomonasreinhardtii
전체 유전체 분석 완료,
분자생물학적 모델
생물, 대량배양 가능
미세조류 바이오디젤 생산용
모델 균주, 양식사료 생산
Kim & Choi,
2014
Chlorellavulgaris
온도 및 염도 적응성
높음, 대량배양 가능
건강식품 (단백질, 지질,
비타민, 미네랄), 양식사료 및
바이오에너지 생산
Kim & Choi,
2014
Closteriummoniliferum
방사성 물질 흡수능유해물질 검정용, 중금속 및
방사능 물질 흡수 가능
Krejici et al.,
2011
Dunaliellasalina
염도 적응성 높음,
대량배양 가능
베타카로틴 등 색소물질
생산용, 바이오디젤 생산용
Borowitzka et
al., 1990
Haematococcuspluvialis
아스타잔틴 생산능,
대량배양 성공
아스타잔틴 등 색소물질
생산용, 바이오디젤 생산용
Kim & Choi,
2014
Scenedesmussp.
온도 적응성 높음,
대량배양 용이
바이오에너지 및 양식사료
생산용
Kim & Choi,
2014
표 I-1. 유용성이 검증된 미세조류(남조류 및 녹조류) 현황
다음으로 원핵생물인 남조류는 다른 진핵조류와는 달리 세포벽에 다당류가
결여되어 있어 인간을 비롯한 동물이 이를 섭취할 경우 쉽게 소화할 수 있다고
알려져 있으며(Richmond & Preiss 1980), 세포 내 단백질의 함량이 매우 높아

- 8 -
건강식품 및 동물 사료원으로 오래전부터 사용되어 왔다. 실제로 Spirulina(스피
룰리나) 속 남조류의 경우, 아프리카의 챠드(Chad) 호수와 멕시코 텍스코코
(Texcoco) 호수와 같은 열대지역 알칼리성 호수의 수면에서 왕성하게 자생하며
16세기 아즈텍, 마야인들의 주식이었다는 기록이 전해진 것으로 보아 식용으로
사용된 오랜 역사를 가지고 있다. 스피룰리나는 인체에 필요한 5대 영양소 및
생명체 유지를 위한 영양소가 다량 함유되어 있어, WHO에서 완전식품, 수퍼
푸드 등으로 칭하여 의료, 건강, 기아해방 프로그램 등에 이용하고 있으며,
NASA에서는 우주식량으로도 개발하여 사용 중인 모델 남조류이다(그림 I-1,
표 I-2). 또한 Nostoc 속 남조류(N. commune, N. flagelliforme, N.
punctiforme) 역시 다양한 방법으로 식재료에 이용되고 있다(Facciola, 1998).
남조류 유래 광합성 색소물질인 C-Phycocyanin은 강력한 항산화물질로서 자유
라디칼 손상감소 및 종양세포 사멸유도 효과(Subhashini et al. 2004)와 더불어
항알레르기, 면역력 증진 등의 특성을 지니고 있어(Nemoto-Kawamura et al.
2004), 식품 및 화장품 등의 천연 색소물질로 첨가되고 있다. 또한 남조류는
Herpes virus, HIV 및 Influenza virus 등에 대한 항바이러스 작용을 하는 다당류
(Schaeffer and Krylov, 2000) 및 Dolostatin과 같은 항암물질(Luesch et al.
2002) 등을 생산하는 것으로 보고되고 있다.
그림 I-1. 남조류(Spirulina or Arthrospira)의 생산·성분분석 및 활용성 분석

- 9 -
아미노산 종류 Spirulina Chlorella 콩 쇠고기 달걀 공미리
이소류신
(Isoleucine)3.3~3.9 3.9 1.8 0.93 0.67 0.83
류신
(Leucine)5.9~6.5 6.01 2.7 1.7 1.08 1.28
리신
(Lysine)2.6~3.3 3.6 2.58 1.76 0.89 1.95
메티오닌
(Methionine)1.3~2.0 0.61 0.48 0.43 0.4 0.58
시스틴
(Cystine)0.5~0.7 0.48 0.48 0.23 0.35 0.38
페닐알라닌
(Phenyl alanine)2.6~3.3 3 3 0.86 0.65 0.61
티로신
(Tyrosine)2.6~3.3 2.53 2.53 0.68 0.49 0.61
트레오닌
(Threonine)3.0~3.6 2.3 2.3 0.86 0.59 0.99
트립토판
(Tryptophan)1.0~1.6 0.59 0.59 0.25 0.2 0.3
발린
(Valine)4.0~4.6 3.3 3.3 1.05 0.83 1.02
표 I-2. 녹조류(Chlorella) 및 남조류(Spirulina)의 아미노산 함량 비교 예
다. 연구의 필요성
1) 자생 유용 미세조류 확보의 필요성
상기와 같은 결과를 토대로 남조류와 녹조류를 포함하는 미세조류는 이산화
탄소 저감, 바이오에너지 생산 및 천연 기능성 물질 생산 등이 가능한 생물 플랫폼
(Platform)으로써 그 연구가치 및 효용성이 충분하다고 할 수 있다. 우리나라의
미세조류 연구의 경우, 환경부의 ⌜자생생물 발굴사업⌟ 및 ⌜녹조 방제 사업
등을 비롯하여 미래부의 ⌜차세대바이오매스 연구단 사업⌟ 및 해수부의 ⌜미세조류의 해양 배양을 통한 바이오디젤 생산 기술개발 사업⌟ 등을 통하여 미세
조류의 활용성을 증진시키기 위한 연구를 진행하고 있다. 그러나 환경부 사업은
미세조류 활용방안에 대한 연구가 부족하며, 타 부처의 사업은 유용 미세조류
발굴과 관련된 연구가 부족한 실정이다.
상기 제시된 Chlorella, Haematococcus, Nostoc 및 Spirulina(Arthrospira)
속 등의 미세조류는 효율적인 단백질 및 유용물질을 포함한 바이오매스 공급원

- 10 -
으로써 그 중요성을 인정받고 있으나, 대부분 국제적으로 특허 미생물로 등록
되어 국내에서의 산업적 적용에 문제점을 노출하고 있다. 또한 현재 전 세계적
으로 상업적인 미세조류 배양은 몇몇 국제 기업에 의해서 독점되고 있으며, 국내
에서 이러한 국제 기업을 따라잡기 위해서는 국제적 특허분쟁의 소지가 없는
자생 유용 미세조류를 이용한 바이오매스 생산 기술 개발이 시급한 실정이다.
이러한 이유로 본 연구는 ①배양이 가능한 자생 미세조류를 확보하여 ②유용
성을 탐색하고 ③유용성이 검증된 미세조류의 최적 배양조건을 탐색하는 것을
그 목표로 한다. 본 연구를 통하여 수입 미세조류 종을 대체할 수 있는 자생
미세조류 종을 발굴하여 국내 미세조류 관련 산업의 기반을 확충하며, 미세
조류와 관련된 국가 생물 다양성 관련 연구에 기반자료를 제공한다.

- 11 -
II. 연 구 방 법


- 13 -
Ⅱ. 연구방법
1. 수생태계 유래 자생 미세조류 배양체 확보
가. 연구방법
1) 시료 채집 및 미세조류 분리
본 연구에 사용된 미세조류 배양체는 담수 및 기수의 다양한 수생 서식환경
(수생생물배양장, 저수지, 강, 하구둑, 갯벌, 조간대 지역등)에서 부유성 플랑크톤을
중심으로 채집하였다. 채집은 망목 크기가 20㎛인 플랑크톤 네트로 채집수를 농축
시킨 후, 채수병에 담아 실험실로 옮겨졌다. 다양한 수체로부터 채집한 시료로부터
우점종을 모세관법 혹은 한천배지(BBM, BG-11 medium, Zarrouk's medium, f/2
medium 영양배지) 도말법 등을 이용하여 분리하였다. 분리한 배양체는 분류군에
따라 적합한 배지(BBM, BG-11 medium, Zarrouk's medium, f/2 medium; 배지
조성표 부록 II 기재)에서 100.0 μmol/m2/s, pH 7.0, 25±2 °C, 12L:12D 명암주기
하에서 보존 배양 한 후 증식에 이용하였다.
2) 배양 가능 미세조류의 형태학적 분류·동정
배양체의 정확한 종 동정을 위하여 광학현미경(Eclipse 80i; Nikon Co.)을
이용하여 분리된 미세조류 세포의 형태를 관찰하였고, 카메라(DXM 1200C;
Nikon Co.)를 이용하여 사진을 촬영하였다. 운동성 미세조류의 경우 관찰 전
포르말린 수용액(formaldehyde, 10% 최종 농도 v/v)으로 고정되었다. 미세조류
세포의 크기는 이미지분석기(NIS-Elements BR 3.0; Nikon Co.)로 측정하였다.
특정 미세조류의 초미세구조 분석을 위해 주사전자현미경(Scanning electron
microscope)과 투과전자현미경(Transmission electron microscope) 검경을 진행
하였다. 주사전자현미경 검경을 위하여 미세조류 세포를 2.5% glutaraldehyde가
녹아있는 0.1 M cacodylate buffer로 고정시키고, 2% osmium tetroxide으로
후고정시켰다. 고정된 미세조류 세포를 단계적으로(30%, 50%, 70%, 80%, 90%,
95% 및 100%) 에탄올을 이용하여 탈수시켰다. 탈수된 미세조류 시료는 100%
hexamethyldisilaza로 건조시킨 후, aluminum stub에 탑재하여 sputter
coater(SCD050; BAL-TEC)를 이용하여 90초 동안 백금으로 코팅하였다. 투과

- 14 -
전자현미경 검경을 위해 고정 과정부터 세척, 후고정, 탈수과정까지 주사전자
현미경 전처리 과정과 동일하게 수행한 후, Spurr’s resin(Spurr, 1969)에 최종
고정시켰다. 고정된 미세조류 샘플은 ultra-microtome(EM UC7; Leica
Microsystems)을 이용하여 70nm에서 절편되었고, 2% uranyl acetate와 lead
citrate로 염색한 후 투과전자현미경(JEM1010; JEOL Ltd.) 분석을 수행하였다.
3) 유전자 염기서열 기반 분자생물학적 분류·동정
가) 남조류(Cyanophyta)의 분자생물학적 분류·동정
남조류 배양체의 게놈 DNA는 제조업자의 설명서에 따라 DNeasy®Blood
and Tissue kit(Qiagen)을 이용하여 추출되었으며, 추출된 DNA는 PCR 주형
DNA로 이용되었다. universal 27F/1492R 프라이머 및 16S1407F/23S30R 프라이머를
이용하여 16S rRNA 및 16S-23S intergenic spacer(ITS) 영역들이 각각 증폭
되었으며, 염기서열은 ABI 377 Sequencer(Applied Biosystems)를 이용하여
Macrogen Inc.(Korea)에서 분석되었다. 확보된 16S rRNA와 16S-23S ITS 염기
서열들은 National Center for Biotechnology Information(NCBI)의 BLAST
search(http://www.ncbi.nlm.nih.gov/BLAST)를 이용하여 유사도(similarity)
검색을 통하여 분석되었다.
남조류 배양체의 16S rRNA 염기서열은 GenBank 데이터베이스에서 이용
가능한 일부 군들(taxa)을 포함하는 AlgaeBase(http://www.algaebase.org/) 내
유사 속(genera) 내의 분류학상 받아들여지는 종의 해당 염기서열들과 비교하여
CLUSTALX v.1.83(Thompson et al., 1997), BioEdit Sequence Alignment Editor
v. 7.1.0.3(Hall, 1999) 및 Muscle v.3.6(Edgar. 2004)을 이용하여 정렬되었다. 모호
하게 정렬된 위치들은 Gblocks 0.91b(Castresana. 2000) 프로그램의 “allowed
gap positions” 항목 내 “with half”를 이용하여 제거되었다. 해당 남조류 배양체
의 계통분석은 MEGA v.5.0.5(Tamura et al., 2011) 프로그램 내의
Jukes-Cantor distances matrix 기반 Neighbour-joining(NJ) 방법을 이용하여
1,000 부트스트랩(bootstrap) 반복검증(replicates) 수행을 통해 계통도를 구성
하였다.
Maximum-likelihood(ML) 계통도 구성의 경우, jModeltest v.0.1.1(Posada,
2008)에 의하여 제안된 모델을 기반으로 PHYML v.3.0(Guindon & Gascuel,
2003) 프로그램을 이용하여 1,000 부트스트랩(bootstrap) 반복검증(replicates)을

- 15 -
통해 계통도를 구성하였다.
Clade credibility 계통도 구성의 경우, jModeltest v.0.1.1(Posada, 2008)에
의하여 제안된 모델을 기반으로 MrBayes v.3.1.2(Huelsenbeck & Ronquist,
2001) 프로그램 내 Bayesian inference 방법을 이용하여 3개의 병렬 실행
(parallel runs)으로 분석되었다. 분석 과정에서는 100 세대마다 샘플이 수집되었고,
실행들 간의 split frequencies의 표준 편차가 0.05 이하가 될 때까지 분석이 진행
되었다. 상기 3종의 계통도 분석들에 있어 계통도의 외집단(outgroup)으로
Gloeobacter violaceus PCC 7421(GenBank accession No. AF132790) 혹은
Escherichia coli K12(GenBank accession No. AE000460)가 사용되었다.
남조류 배양체의 16S-23S ITS 영역은 GenBank로부터 이용 가능한 유사
속(genera) 내 다른 종 균주 유래 대표적인 염기서열들과 비교하여 정렬되었다.
상기 정렬된 16S-23S ITS 영역 염기서열들은 Muscle v3.6(Edgar. 2004) 프로그램을
이용하여 정렬되었으며, MEGA v.5.0.5(Tamura et al., 2011)를 이용하여 1,000
부트스트랩(bootstrap) 반복검증(replicates) 수행을 통해 Maximum
parsimony(MP) 및 NJ 분석법을 통하여 계통도가 구성되었다. MP 계통도는 10
개 복제물의 무작위 삽입 서열에 의하여 구성된 초기 계통도와 함께
Neighbour-interchange heuristic search (MP search level 1) 및 갭 혹은 손실
염기서열 데이터들의 complete deletion option을 이용하여구성되었다. NJ 계통도는
손실된 염기서열 데이터들에 대한 complete deletion option 및 Jukes- Cantor
distances matrix를 이용하여 구성되었다. 계통도 분석들에 있어 계통도의 외집단
(outgroup)으로 유사 속(genara)의 16S-23S ITS 염기서열 영역들이 선택적으로
이용되었다.
나) 녹조류(Chlorophyta)의 분자생물학적 분류·동정
녹조류 배양체의 게놈 DNA는 Plant DNA isolation reagent(Takara)를
이용하여 추출하였다. 분리된 게놈 DNA를 PCR 주형으로 이용해 18S rRNA
또는 D1/D2 region(ITS1-5.8S rRNA-ITS2-28S rRNA, 이후 ITS region으로
언급)을 포함하는 18S-28S ITS region을 증폭시켰다. 18S rRNA 유전자는
NS1, NS3 및 NS8 프라이머를 이용하여 증폭시켰고, ITS region은 ITS1, ITS4
및 LR3R 프라이머를 이용하여 증폭시켰다. 증폭된 PCR 산물은 pGEM®-T
Easy Vector(Promega)를 이용하여 클로닝 되었고, 증폭된 유전자 서열을 분석
하기 위해 ABI 377 Sequencer(Applied Biosystems)를 이용하여 Macrogen
Inc.(Korea)에서 분석되었다. 확보된 18S rRNA와 18S-28S ITS region 염기

- 16 -
서열들은 NCBI의 BLAST search(http://www.ncbi.nlm.nih.gov/BLAST)를
이용하여 유사도(similarity) 검색을 통하여 분석되었다.
녹조류 배양체의 18S rRNA 염기서열은 GenBank 데이터베이스에서 이용
가능한 일부 군들(taxa)을 포함하는 AlgaeBase(http://www.algaebase.org/) 내
유사 속(genera) 내의 분류학상 받아들여지는 종의 해당 염기서열들과 비교하여
CLUSTALX v.1.83(Thompson et al., 1997), BioEdit Sequence Alignment
Editor v. 7.1.0.3(Hall, 1999) 및 Muscle v.3.6(Edgar. 2004)을 이용하여 정렬되었다.
모호하게 정렬된 위치들은 Gblocks 0.91b(Castresana. 2000) 프로그램의
“allowed gap positions” 항목 내 “with half”를 이용하여 제거되었다. 해당 녹조류
배양체의 계통분석은 MEGA v.5.0.5(Tamura et al., 2011) 프로그램 내의
Jukes-Cantor distances matrix 기반 NJ 계통도 방법을 이용하여 1,000 부트스트랩
(bootstrap) 반복검증(replicates) 수행을 통해 계통도를 구성하였다. ML 계통도
구성의 경우, jModeltest v.0.1.1(Posada, 2008)에 의하여 제안된 모델을 기반으로
MEGA v.5.0.5(Tamura et al., 2011) 프로그램을 이용하여 1,000 부트스트랩
(bootstrap) 반복검증(replicates)을 통해 계통도를 구성하였다.
녹조류 배양체의 18S-28S ITS region은 GenBank로부터 이용 가능한 유사
속(genera) 내 다른 종 균주 유래 대표적인 염기서열들과 비교하여 정렬되었다.
상기 정렬된 18S-28S ITS 영역 염기서열들은 CLUSTALX v.1.83(Thompson
et al., 1997), BioEdit Sequence Alignment Editor v.7.1.0.3(Hall, 1999) 및
Muscle v.3.6(Edgar. 2004)을 이용하여 정렬되었으며, MEGA v.5.0.5(Tamura et
al., 2011)를 이용하여 1,000 부트스트랩(bootstrap) 반복검증(replicates) 수행을
통해 MP 및 NJ 분석법을 통하여 계통도가 구성되었다. MP 계통도는 10개 복제
물의 무작위 삽입 서열에 의하여 구성된 초기 계통도와 함께 Neighbour-
interchange heuristic search(MP search level 1) 및 갭 혹은 손실 염기서열
데이터들의 complete deletion option을 이용하여 구성되었다. NJ 계통도는 손실된
염기서열 데이터들에 대한 complete deletion option 및 Jukes-Cantor distances
matrix를 이용하여 구성되었다. 계통도 분석들에 있어 계통도의 외집단
(outgroup)으로 유사 속(genara)의 18S-28S ITS 영역 염기서열들이 선택적으로
이용되었다.

- 17 -
표 II-1. 염기서열 분석용 Oligonucleotide primer 정보
유전자 Primer 명 염기서열(5'-3') 참고문헌
16S
rRNA
27F AGAGTTTGATCMTGGCTCAGLane et al. 1991
1492R TACGGYTACCTTGTTACGACTT
16S-23S
ITS
region
16S1407F TGTACACACCGCCCGTCTaton et al., 2003
23S30R CTTCGCCTCTGTGTGCCTAGGT
18S
rRNA
NS1 GTAGTCATATGCTTGTCTC
White et al, 1990NS3 GCAAGTCTGGTGCCAGCAGCC
NS8 TCCGCAGGTTCACCTACGGA
18S-28S
ITS
region
ITS1 TCCGTAGGTGAACCTGCGG
White et al, 1990ITS4 TCCTCCGCTTATTGATATGC
LR3R GTCTTGAAACACGGACC

- 18 -
2. 자생 미세조류 유용성 검증
가. 연구방법
1) 미세조류 배양체의 성분 분석용 바이오매스 확보
확보된 10주의 남조류 및 녹조류 배양체의 바이오매스 확보를 위하여, 증식기의
각 미세조류주의 순수 배양체를 BBM, BG-11, f/2, 혹은 Zarrouk's 영양배지(5L
급)에 접종하여 100.0 μmol/m2/s, pH 7.0, 25 ± 2°C, 12L:12D 명암주기 하에서
증균 배양을 지속하였다. 미세조류 증식률은 Sedgwick-Rafter chamber를 사용
하여 2일 또는 4일 간격으로 측정하였으며, 정체기 또는 쇠퇴기에 도달할 때까지
최소 21일 최대 35일 동안 배양하였다. 선형 남조류의 경우 10 mL의 배양액을
2일 또는 4일 간격으로 GF/C 필터(Watmann)로 여과한 후 105℃에서 24시간
건조한 후 그 질량의 변화를 측정하여 성장률(P)을 결정하였다.
P = (X1-X0)/(t1-t0)
P, 바이오매스 생산량(g/L/day); X1 및 X0, t1 과 t0 시점의 바이오매스 건중량
미세조류 배양체는 플랑크톤 네트(망목 크기 20㎛) 혹은 원심분리법(7000
rpm, 20분)으로 수확되었으며, 확보된 바이오매스는 -70°C에서 급속 냉각한 후 동결
건조되어 분말형태로 암소에 건조된 상태로 보관하였다.
2) 미세조류 배양체 바이오매스의 일반성분 분석 및 평가
미세조류 일반성분(회분, 조탄수화물, 조지질, 수분 및 조단백)은 Official
Methods of Analysis of the Association of Official Analytical
Chemists(AOAC)에 따라 결정되었다(AOAC. 1990).
가) 수분 함량 측정 :
동결 건조된 미세조류 분말을 각 실험구 별 0.3 g 씩 선 계량된 실험용 도가니에
넣어 105℃로 설정된 건조기(Oven)에서 24 시간 건조시킨 후, 실온에서 방냉한
뒤, 데시케이터(dessicator)에서 약 30분간 방냉하였다. 건조된 도가니의 무게를
측정 후, 그 차이를 통해 수분의 함량을 측정하였다.

- 19 -
나) 무기질 함량 측정 :
수분 함량 측정 뒤, 무게를 측정한 도가니를 600℃로 설정된 회화로에서 24
시간 건조하였다. 실온에서 방냉한 뒤, 데시케이터에서 30분간 방냉한 후, 도가니의
무게를 측정하여 이를 회분의 함량으로 측정하였다.
다) 조단백 함량 측정 :
조단백질 함량은 Kjeldhal 방법(official method 976.05)으로 측정하였다. 먼저
동결 건조된 미세조류 분말을 각 실험구 별 0.3 g 씩 실험용 유리관에 분주한
후, 전처리를 하기 위해 황산(H2SO4) 12 ml과 촉매제를 넣어 1시간 반응을 유지
한다. 반응 종료 시, 30분간 식힌 후 KjeltecTM8200 system을 이용하여 조단백질
함량을 분석하였다.
라) 조지질 함량 측정 :
조지질 함량은 Soxhlet 방법(official method 920.39)으로 측정하였다. 먼저
동결 건조된 미세조류 분말을 각 실험구 별 50 mg을 15 mL conical tube에 분주
후, Chloroform:MeOH(1:2, v/v) 용액을 3 mL 첨가하였다. 건조된 샘플의 세포
파쇄를 위해 초음파분쇄(sonication) 과정을 거친 후, 5분간 원심분리(3000 rpm)
하여 샘플의 상등액을 따로 모은 후, Chloroform:MeOH(2:1, v/v) 용액을 3 mL
넣어 한 번 더 초음파 분쇄과정 후, 원심분리(3000 rpm)하여 샘플의 상등액 만을
취하였다. 이러한 과정을 상등액의 색이 투명해질 때까지 반복하였다. 최종 수집
된 샘플 상등액의 층분리를 위하여 9%의 NaCl을 전체 상등액 양의 50% 만큼
넣고 섞어준 후(Voltexing), 4℃ 냉장고 암소에서 1시간 보관한다. 5분간 원심
분리(3000 rpm)하여 층 분리가 완료되면 하층 용액을 회수하여 계량된 유리
vial에 분주하였다. 분주된 샘플은 농축기를 통해 유기용매를 휘발시킨 후 유리
vial의 무게를 측정하여, 전후 vial의 무게 차이를 통해 지질의 함량을 분석하였다.
마) 조탄수화물 함량 측정 :
총 조탄수화물 함량은 전체 구성성분을 100으로 했을 때, 수분, 회분, 단백질,
지질의 합을 뺀 양(by difference)으로 계량하였다.
3) 미세조류 배양체 바이오매스의 세부성분 분석 및 평가
가) 아미노산 조성 측정 :

- 20 -
미세조류 배양체 세포 유래 유리아미노산은 Kim et al. (2015)의 방법으로 분석을
진행하였다. 요약하자면, 동결 건조된 미세조류 시료를 6N HCl로 산가수분해한
후 0.2 ㎛ syringe filter로 여과하여 dilution buffer로 희석하였다. 미세조류 세포
에서 유리된 아미노산은 cation-exchange column으로 분리한 후 Ninhydrin과
반응시켜 Hydrindantin을 생성시켜 Amino acid Analyzer (Sykam S433;
Sykam)로 분석하였다. 기기분석 관련 정보는 표 II-2에 제시되었다.
표 II-2. 유리 아미노산 조성 분석조건
구분 세부사항
Instrument Amino Acid Analyzer, Membrapure, Germany
Eluent Membrapure Ready to use Buffers A, B, C, D, E and F
Reagent Membrapure "Reagent for Derivatization"
Column Narrow bore, stainless steel, 125 mm, ID 3 mm
Method Postcolumn - derivatization with Ninhydrin
Standard Amino Acid Standard Solution from Sigma (C/N AAS18)
Acquisition &
AnalysisaminoPeak S/W
Injection volume 20μL
나) 지방산 조성 측정 :
지방산 측정을 위해 Bligh and Dyer procedure(Bligh & Dyer, 1959) 방법으로
지질을 추출하였고, 추출된 지질에 0.5N KOH/MeOH를 넣고 70°C에서 30분간
가열한 후, 혼합 유기용매 Hexane:Diethyl ether(9:1, v/v)로 알코올과 스테롤을
분리, 추출하였다. 추출하고 남은 용액에 염산을 넣어 KOH를 침전시킨 후, 혼합
유기용매 Hexane:Diethyl ether(9:1, v/v)로 지방산을 추출하였다. 추출된 지방산에
BF3/MeOH를 첨가하고, 70°C에서 30분간 가열하여 지방산 메틸에스테르(Fatty
Acid Methyl Ester, FAME)로 유도한 후, 추출 전에 첨가된 internal standard인
n-Nonadecanoic acid와 비교하여 gas chromatocraphy-flame ionization
detector(GC-FID; Agilent 7890A)로 정량 분석하였다. 지방산의 분리, 분석에는
GC-FID에 ZB5MS column을 사용하였다. 적용된 온도조건은 시료 주입구

- 21 -
250°C, column 오븐은 50°C에서 120°C까지 10°C/분, 120°C에서 300°C까지 4°C/분
으로 설정되었고, 이동상 가스는 헬륨을 사용하였다. 각 지방산의 구조(탄소수,
포화도, 이중결합 위치)는 GC-mass spectrometry detector(GC-MSD)를 이용하여
70eV에서 원자질량 50-700 amu 범위에서 수행되었다. 지방산 성분 중 불포화
지방산의 탄소 이중결합 위치는 상업용 지방산 standard 또는 FAME을 FAME
acid picolinyl ester로 전환하여 GC-MSD로 확인하였다(Kim et al., 2015).
다) 단당 조성 측정 :
미세조류 세포의 단당 조성을 측정하기 위하여 Kim et al. (2015)의 방법으로
분석을 진행하였다. 요약하자면, 건조시료를 2M TFA(Trifluoroacetic acid) 산
가수분해 후 여과(0.2 ㎛) 및 희석하였고, 가수분해 된 당 성분 및 함량 측정은
high-performance anion-exchange chromatography coupled with a pulsed
amperometric detection system(HPAEC-PAD; Dionex Corp.)을 이용하여 분석
하였다. 기기분석 관련 정보는 표 II-3에 제시되었다.
표 II-3. 유리당 조성 분석조건
구분 세부사항
Instrument* HPAEC-PAD system, Dionex, USA
Eluent 18 mM NaOH
Column CarboPacTM PA1
Flow rate 1.0 mL/min
Injection volume 15μL
* LOD(Limit of Detection) = 10 pmole

- 22 -
특성 세부특성 조건/가중치
배양
특성
배양용이성기수 담수 해수
3 2 1
수확용이성플랑크톤 네트 원심분리
2 1
신규성 균주 신규성국외 활용 미보고 국내 활용 미보고 국내 활용 보고
3 2 1
일반성분
특성
조단백 함량>50% <50%
1 0
조지방 함량>20% <20%
1 0
조탄수화물
함량
>20% <20%
1 0
세부성분
특성
유용 지방산유 무
1 0
필수
아미노산
>50% <50%
1 0
유용 단당유 무
1 0
3. 유용 미세조류 선별 및 배양기법 탐색
가. 연구방법
1) 유용성 검증결과 기반 유용 미세조류주 선별
유용 미세조류주의 선별은 표 II-4와 같은 선정조건 및 산정기준으로 진행
되었다. 배양특성과 관련하여 ① 배양의 용이성은 염도조건으로 산정되어 타
미생물에 의한 오염이 적은 정도로 기수⇒담수⇒해수의 순으로 나열하였으며,
② 수확의 용이성은 대량배양을 감안하여 수확이 용이한 플랑크톤 네트⇒원심
분리 순으로 산정하였다. 미세조류주 활용 신규성과 관련하여서는 ①국외 활용
미보고, ②국내 활용 미보고, ③국내 활용 보고로 가중치를 산정하였다. 일반성분
특성과 관련하여 ①조단백 함량은 50% 이상 및 이하로 구분하였고, ②조지방 및 ③
조탄수화물 함량은 20% 이상 및 이하로 구분하여 산정되었다. 세부성분 특성
역시 ①유용 지방산의 유무, ②필수 아미노산(함량 50% 이상 및 이하로 구분),
③유용 단당의 유무로 구분하여 산정되었다. 각 미세조류 배양체의 특성을 이에
대입하여 점수를 산정하였고, 최종적으로 3주의 미세조류주를 선정하였다.
표 II-4. 유용 미세조류주 선정조건 및 산정기준

- 23 -
2) 성장률 기반 최적 배양조건 탐색
유용 미세조류주로 선별된 3주의 대수증식기의 남조류 및 녹조류 배양체를
준비하여 각 미세조류주의 순수 배양체를 다양한 염분, pH 및 온도 조건의
BG-11, 혹은 Zarrouk's 영양배지에 접종하여 100.0 μmol/m2/s, 12L:12D 명암
주기 하에서 28일 동안 폭기 방식의 증균 배양을 지속하였다. 실험에 사용된 염분의
농도는 0-100 psu(10 psu씩 증감), pH는 2-13(1씩 증감), 수온은 4-35°C(4, 15,
25, 30, 35°C)로 21일간 배양하였다. 미세조류 증식률은 Sedgwick-Rafter
chamber를 사용하여 2일 또는 4일 간격으로 측정하였으며, 선형 남조류의 경우
10 mL의 배양액을 2일 또는 4일 간격으로 GF/C 필터(Watmann)로 여과한 후
105°C에서 24시간 건조한 후 그 질량의 변화를 측정하여 성장 여부를 결정하였다.


- 25 -
III. 연 구 결 과


- 27 -
시료명 분리장소(GPS) 배지/배양능
NIBRUIM-1충남, 금강 하구둑
(36°01'48.0"N 126°45'27.4"E)
BBM BG-11 Zarrouk's f/2
✕ △ O ✕NIBRUIM-2
제주, 서귀포
(33°15'45.6"N 126°37'30.9"E)
BBM BG-11 Zarrouk's f/2
✕ △ O ✕NIBRUIM-3
경기, 안산
(37°17'14.8"N 126°50'00.8"E)
BBM BG-11 Zarrouk's f/2
O O O O
NIBRUIM-4제주, 구좌읍
(37°17'14.8"N 126°50'00.8"E)
BBM BG-11 Zarrouk's f/2
△ O △ O
NIBRUIM-5전북, 금강 하구둑
(36°01'24.9"N 126°45'59.3"E)
BBM BG-11 Zarrouk's f/2
△ △ O △
NIBRUIM-6경기, 안산
(37°17'04.8"N 126°50'03.3"E)
BBM BG-11 Zarrouk's f/2
O O ✕ ✕NIBRUIM-7
경기, 안산
(37°17'04.8"N 126°50'03.3"E)
BBM BG-11 Zarrouk's f/2
O O ✕ ✕NIBRUIM-8
충남, 태안
(36°53'35.0"N 126°19'40.8"E)
BBM BG-11 Zarrouk's f/2
O O ✕ O
NIBRUIM-9제주, 구좌읍
(33°32'35.5"N 126°48'38.4"E)
BBM BG-11 Zarrouk's f/2
O O ✕ ✕NIBRUIM-10
제주, 구좌읍
(33°32'35.5"N 126°48'38.4"E)
BBM BG-11 Zarrouk's f/2
O O ✕ ✕
III. 연구결과
1. 수생태계 유래 자생 미세조류 배양체 확보
가. 연구결과
1) 시료 채집 및 미세조류 분리
채집한 미세조류 시료 중 50주 이상의 미세조류 세포체를 모세관법으로 분리
하여 BBM, BG-11 medium, Zarrouk's medium, f/2 medium 등의 액체배지에
접종한 후 배양능이 있는 종을 선별하였으며, 이와 별도로 미세조류 시료를 정치
농축한 후 각각의 한천배지에 직접 도말하여 마찬가지로 배양능이 있는 종을
선별하였다. 이러한 선별과정을 통하여 배양이 용이한 미세조류주를 선정하였으며,
선정된 미세조류주의 분리 및 배지 관련 자료는 표 III-1에 제시하였다.
표 III-1. 분리된 자생 미세조류주(NIBRUIM) 현황
(※ O: 정상적 증식, △: 느린 증식 or 현상유지, X: 무증식 or 사멸)

- 28 -
2) 배양 가능 미세조류의 형태학적 및 분자생물학적 분류·동정
배양체의 색깔과 세포 특성을 기준으로 남조류(Cyanophyta)와 녹조류
(Chlorophyta)로 분류하였으며, 각 연구대상종의 형태적 특징은 다음과 같다.
가) 남조류(Cyanophyta)
(1) NIBRIUM-1
그림 III-1. Arthrospira sp. NIBRIUM-1의 형태 분석. 충청남도 금강하구
둑 주변에서 채집되어(A), 순수 분리된(B) Section III(Oscillatoriales) 남조
류 NIBRIUM-1(Scale bar=100 μm).
(가) 생태․형태적 특징[Cyanophyta Section III, Oscillatoriales]
본 미세조류주는 금강 하구둑 부근 기수계에서 분리되었으며, 유속이 적은
갈대숲 근처에서 채집되었다. 채집 당시 군체 및 독립 형태 모두 발견되었으며,
주로 다른 선형 남조류와 함께 군체를 다수 형성하고 있었다. 채집 당시 수온
(21.9°C), 조도(416.9 μmol photon/m2/s), 염도(3.9 psu) 및 pH(8.3) 등의 환경
조건이 측정되었다.
광학현미경 상 (그림 III-1) 분석에서 관찰된 NIBRIUM-1은 가지 없는
(non-branched) 단일 사상체(trichome)로 이루어져 있으며, 정치 액상 배양에서
전형적으로 50-100 개의 디스크(disc) 형상의 개별 세포들로 구성되고, 운동성
사상체의 형태는 대부분 물결(wavy)이나 구부러진(bent) 형태로 사상체의 길이는
0.3-3.0 mm까지 다양한 크기로 측정되었다(그림 III-1). 상기 사상체들에서는
sheath, heterocysts, akinetes, nodules 또는 necridic cell 등은 발견되지 않았다.
각각의 세포들은 길이가 5.2-8.8 μm, 지름이 0.8-1.1 μm였으며, 사상체 내 세포

- 29 -
간 cross-wall의 constriction은 불명확 하였다. 정단 세포(apical cell)들은 내피막
(calyptra) 없이 뭉툭한 원뿔 형상이었다. 이러한 형태적 특성을 통하여
NIBRIUM-1은 기수에 서식하는 Section III(Oscillatoriales) 남조류로 분류되었다.
(나) 분자생물학적 분류[Oscillatoriales, Arthrospira(Sitzenberger ex Gomont, 1892)]
Section III(Oscillatoriales)로 분류된 NIBRIUM-1 남조류의 분류학상 위치를
결정하기 위하여, 획득된 1421 bp의 16S rRNA 서열을 BLASTN 검색을 이용
하여 남조류의 GenBank 데이터베이스와 비교하였다. 그 결과 유사 서열들의
대부분이 Arthrospira(일반적으로 Spirulina로 명칭되는 남조류류) 속(genera)에
속하는 균주 유래라는 것이 밝혀졌다. BLAST 검색에 의하여 확인된 가장
유사한 균주는 Arthrospira platensis PCC 7345(GenBank accession No.
NR125711)로, 99.1%의 서열 동일성(identity)을 갖는다. 또한 현재 분류학상으로
받아들여지는 Arthrospira 속 내의 종들과 매우 높은 유사도를 나타내었다.
이러한 유사도 결과를 기반으로 남조류 NIBRIUM-1의 정확한 계통분류학적
위치를 확인하기 위하여 Arthrospira 속을 포함하는 Oscillatoriales 내 일부
매우 유사한 균주(strain)를 포함하는 남조류 유래 24개 데이터 세트의 16S
rRNA 서열을 이용하여 NJ 및 ML 계통분류학적 분석을 수행하였다. 상기 두
가지의 계통분석 결과 Oscillatoriales 내에서 Arthrospira 속은 모두 독립된 그룹을
형성하였으며, 해당 그룹 내에 NIBRIUM-1이 포함되는 것을 확인하였다(그림
III-2). 현재 Arthrospira 속은 maxima, platensis 등 총 24종이 분류학상으로
인정되고 있으며(http://www.algaebase.org/), 종간 분류는 매우 난해하여
Arthrospira 속 내의 종에 대한 재분류가 필요한 것으로 인정되고 있다.
따라서 본 연구에서 분리된 NIBRIUM-1 남조류는 A. maxima와 A.
platensis 간의 명확한 분류가 인정되지 않은 관계로 Arthrospira 속으로만 분류
되었으며, 최종적으로 Arthrospira sp. NIBRIUM-1로 명명되었다.

- 30 -
그림 III-2. Arthrospira sp. NIBRIUM-1의 계통분류도. Section
III(Oscillatoriales) 남조류 NIBRIUM-1의 16S rRNA 유전자를 이용한 ML
계통분류 분석이며, Outgroup으로 Escherichia coli K12(GenBank accession
No. AE000460)를 사용함.

- 31 -
(2) NIBRIUM-2
그림 III-3. Arthrospira sp. NIBRIUM-2의 형태 분석. 제주특별자치도 서귀
포시 쇠소깍 주변에서 채집되어(A), 순수 분리된(B) Section III(Oscillatoriales)
남조류 NIBRIUM-2(Scale bar=20 μm).
(가) 생태․형태적 특징[Cyanophyta Section III, Oscillatoriales]
본 미세조류주는 제주특별자치도 서귀포시 쇠소깍 부근 기수계에서 분리
되었으며, 유속이 적은 수변에서 확인되었다. 채집 당시 군체 및 독립 형태
모두 발견되었으며, 주로 다른 선형 남조류와 함께 군체를 다수 형성하고
있었다. 채집 당시 수온(20.1°C), 조도(108.6 μmol photon/m2/s), 염도(1.7 psu)
및 pH(8.0) 등의 환경조건이 측정되었다.
광학현미경 상 분석에서 관찰된 NIBRIUM-2은 가지 없는(non-branched)
단일 사상체(trichome)로 이루어져 있으며, 정치 액상 배양에서 전형적으로
50-200 개의 디스크(disc) 형상의 개별 세포들로 구성되고, 사상체의 형태는
대부분 또아리나 스프링(coil) 형태로 사상체의 길이는 0.6-2.6 mm까지 다양한
크기로 측정되었다(그림 III-3). NIBRIUM-2 남조류주 역시 운동성을 나타
내었으나, NIBRIUM-1주와 차별적으로 빛 조건 하의 회전운동에 한정되었다.
상기 사상체들에서는 sheath, heterocysts, akinetes, nodules 또는 necridic cell
등은 발견되지 않았다. 각각의 세포들은 길이가 4.2-9.2 μm, 지름이 0.8-1.4 μm
였으며, 사상체 내 세포 간 cross-wall의 constriction은 NIBRIUM-1에 비하여
명확하였다. 정단 세포(apical cell)들은 내피막(calyptra) 없이 뭉툭한 원뿔형상
이었다. 이러한 형태적 특성을 통하여 NIBRIUM-2은 기수에 서식하는 Section
III(Oscillatoriales) 남조류로 분류되었다.

- 32 -
(나) 분자생물학적 분류[Oscillatoriales, Arthrospira(Sitzenberger ex Gomont, 1892)]
Section III(Oscillatoriales)로 분류된 NIBRIUM-1 남조류의 분류학상 위치를
결정하기 위하여, 획득된 1423 bp의 16S rRNA 서열을 BLASTN 검색을 이용
하여 남조류의 GenBank 데이터베이스와 비교하였다. 그 결과 유사한 서열들의
대부분이 Arthrospira(일반적으로 Spirulina로 명칭되는 남조류류) 속(genera)에
속하는 균주 유래라는 것이 밝혀졌다. BLAST 검색에 의하여 확인된 가장
유사한 균주는 Arthrospira platensis PCC 7345(GenBank accession No.
NR125711)로, 99.5%의 서열 동일성(identity)을 갖는다. 또한 NIBRIUM-1과
마찬가지로 현재 분류학상으로 받아들여지는 Arthrospira 속 내의 종들과 매우
높은 유사도를 나타내었다. 그러나 Arthrospira 속으로 분류된 NIBRIUM-1
남조류주와의 상호 16S rRNA 비교 결과 NIBRIUM-2 남조류주는 98.7%의
동일성만을 나타내었으며, 형태학적으로도 군체 모양의 차이를 나타내었다.
이를 통하여 NIBRIUM-1과 NIBRIUM-2는 서로 다른 남조류주로 판정되었다.
이러한 유사도 결과를 기반으로 남조류 NIBRIUM-2의 정확한 계통분류학적
위치를 확인하기 위하여 Arthrospira 속을 포함하는 Oscillatoriales 내 일부
매우 유사한 균주(strain)를 포함하는 남조류 유래 24개 데이터 세트의 16S
rRNA 서열을 이용하여 NJ 및 ML 계통분류학적 분석을 수행하였다. 상기 두
가지의 계통분석 결과 Oscillatoriales 내에서 Arthrospira 속은 모두 독립된
그룹을 형성하였으며, 해당 그룹 내에 NIBRIUM-2이 포함되는 것을 확인하였다
(그림 III-4). 현재 Arthrospira 속은 maxima, platensis 등 총 24종이 분류학상
으로 인정되고 있으며(http://www.algaebase.org/), 종간 분류는 매우 난해하여
Arthrospira 속 내의 종에 대한 재분류가 필요한 것으로 인정되고 있다.
따라서 본 연구에서 분리된 NIBRIUM-2 남조류는 A. maxima와 A.
platensis 간의 명확한 분류가 인정되지 않은 관계로 Arthrospira 속으로만 분류
되었으며, 최종적으로 Arthrospira sp. NIBRIUM-2로 명명되었다.

- 33 -
그림 III-4. Arthrospira sp. NIBRIUM-2의 계통분류도. Section
III(Oscillatoriales) 남조류 NIBRIUM-2의 16S rRNA 유전자를 이용한 ML
계통분류 분석이며, Outgroup으로 Escherichia coli K12(GenBank accession
No. AE000460)를 사용함.

- 34 -
(3) NIBRIUM-3
그림 III-5. Leptolyngbya sp. NIBRIUM-3의 형태 분석. 경기도 안산시
Spirulina 배양장에서 채집되어(A), 순수 분리된 Section III(Oscillatoriales)
남조류 NIBRIUM-3의 광학현미경(B), 주사전자현미경(C) 및 투과전자현미
경(D) 검경 결과.
(가) 생태․형태적 특징[Cyanophyta Section III, Oscillatoriales]
본 미세조류주는 경기도 안산시 Spirulina 배양장에서 분리되었으며 채집
당시 군체 및 독립 형태 모두 발견되었다. 채집지 수온(21.1°C), 조도(416.9 μ
mol photon/m2/s), 염도(13.9 psu) 및 pH(10.3) 등의 환경조건이 측정되었다.
광학현미경 상 (그림 III-5) 분석에서 관찰된 NIBRIUM-3은 운동성이
없는 non-branched 단일 사상체(trichome)로, 50-100 개의 통(barrel) 형상의
세포들로 구성된 사상체는 일반적으로 곧거나(straight) 또는 구부러져(bent)
있다. 상기 사상체들에서 sheath, gas vacuole, heterocysts, akinetes, nodules
또는 necridic cell은 발견되지 않았다. 각각의 세포들은 길이가 0.9-2.2 μm,
지름이 0.8-1.1 μm였으며, 사상체 내 세포 간 cross-walls에 명확한
constriction을 나타내었다. 정단(apical) 세포들은 내피막(calyptra) 없이
뭉툭한 원뿔 형상이었다. 또한 저광 또는 최적 광 조건에서 사상체의 뿌리혹

- 35 -
또는 뿌리혹-유사 구조들을 형성이 관찰되지 않았다. 이러한 형태적 특성을
통하여 NIBRIUM-3은 기수에 서식하는 Section III(Oscillatoriales) 남조류로
분류되었다.
(나) 분자생물학적 분류[Oscillatoriales, Leptolyngbya(Anagnostidis & Komárek, 1988)]
Section III(Oscillatoriales)로 분류된 NIBRIUM-3 남조류의 분류학상 위치를
결정하기 위하여, 획득된 1467 bp의 16S rRNA, 489 bp의 16S-23S ITS 서열을
BLASTN 검색을 이용하여 남조류의 GenBank 데이터베이스와 비교하였다.
비교 결과 유사한 서열들의 대부분이 Leptolyngbya 및 Nodosilinea 속(genera)
에 속하는 균주 유래라는 것이 밝혀졌다. BLAST 검색에 의하여 확인된 가장
유사한 2개 균주들은 Nodosilinea sp. 0BB32S02(GenBank accession No.
AJ639894) 및 Nodosilinea sp. 0BB19S12(GenBank accession No. AJ639895)
로, 각각 97.5% 서열 동일성(identity)을 갖는다. 상기 분리물은 Nodosilinea
속의 종 N. nodulosa(Li & Brand) Perkerson & Casamatta UTEX
2910(GenBank accession No. EF122600)에 대하여 96.7% 16S rRNA 서열
유사성(similarity)을 보였다. 게다가, Leptolyngbya 속에서 현재 분류학상으로
받아들여지는 종들 중 가장 유사한 균주는 L. margaretheana 1T12(GenBank
accession No. FR798934) 및 L. antarctica ANT.LACV6.1(GenBank accession
No. AY493589)로, 이들은 각각 97.2% 및 96.4%의 서열 동일성을 가진다.
그러나 NIBRIUM-3은 Leptolyngbya sensu stricto 군에 속하는 Leptolyngbya
속의 표준균주인 L. boryana(Gomont) PCC 73110(GenBank accession No.
X84810)와 단지 87.7%의 16S rRNA 서열 유사성을 나타냈다.
따라서 남조류 NIBRIUM-3의 정확한 계통분류학적 위치를 확인하기 위하여
Oscillatoriales 또는 Pseudanabaenales로부터 유사 남조류주를 포함하는
Leptolyngbya 및 Nodosilinea 속으로부터 유래한 71개 데이터 세트의 16S
rRNA 서열을 이용하여 자세한 계통분류학적 분석을 수행하였다. 그 계통분기
신뢰성 계통도(clade credibility tree)(그림 III-6a)는 Leptolyngbya 속이 다계통
발생이며(polyphyletic), Leptolyngbya sensu stricto를 포함하는 그룹 I 및
Nodosilinea 및 Leptolyngbya 속 균주들과 마찬가지로 Pseudanabaenales에
속하는 다른 속들을 포함하는 그룹 II과 같은 2개의 주요 그룹으로 구분됨을
확인하였다(100% posterior value). 이중 Leptolyngbya 그룹 II로 구분된
NIBRIUM-3은 Leptolyngbya 그룹 II에 속하며, India에서 분리된
Leptolyngbya sp. N62DM와 가장 밀접하게 관련됨을 확인하였다. 유사한
결과가 또한 ML와 NJ 방법들을 이용하여 분석된 계통분류도에서 확인되었다.

- 36 -
그러나 상기 계통수 분석에서 그룹 II의 Leptolyngbya 속으로부터
Nodosilinea 속의 균주를 명확하게 구분하지는 못하였다.
NIBRIUM-3의 16S-23S ITS 영역의 489bp 염기서열을 BLAST 검색을
이용하여 GenBank 데이터베이스와 비교하였으나, 정렬 시 다수의 미정렬
부분이 발견되었다. 16S-23S ITS 서열 역시 90% 이상의 query coverage를
갖는 유사한 염기서열들의 대부분이 Nodosilinea 및 Leptolyngbya 속으로부터
유래한 것이며, Nodosilinea 속에 속하는 상기 균주들은 16S rRNA 서열 비교
에서 나타난 바와 같이, Leptolyngbya 속 유래 남조류주들보다 상대적으로 더
높은 염기서열의 동일성들을 보였다. BLAST 검색에 의하여 확인된 가장
유사한 종은 84.6% 동일성을 갖는 N. bijugata Kovacik 1986/5a(GenBank
accession No. EU528669)였으며, L. antarctica ANT.LACV6.1(GenBank
accession No. AY493632)는 Leptolyngbya 속에서 가장 높은 동일성(80.6%)을
보였다. MP 방법을 이용한 ITS 영역 염기서열의 계통분류학적 분석 결과,
NIBRIUM-3이 N. epilithica Kovacik 1990/52(GenBank accession No.
HM018679), Nodosilinea sp. FI2-2HA2(GenBank accession No. HM018678)
및 N. nodulosa PCC 7104(GenBank accession No. AY768381)와 독립된
cluster를 이루고 있다는 것을 밝혔으나, NIBRIUM-3은 유사 남조류주들과
분리된 lineage를 형성하였고(그림 III-6b), 또한 유사한 계통수 topology가
NJ 분석 시에도 확인되었다.
따라서 NIBRIUM-1의 형태학적 특징이 분류에 있어 중요한 특성으로
간주되어, 저광 조건 하(< 4 μmol photons/m2/s) 사상체들 내 뿌리혹들 또는
뿌리혹-유사 구조들의 형성들은 Leptolyngbya 속으로부터 Nodosilinea 속을
구별하는데 있어 주요 특징이었던 이전 연구(Perkerson et al., 2011)에 따라,
사상체에서 뿌리혹 또는 뿌리혹-유사 구조들을 형성하지 않은 NIBRIUM-3
남조류주는 Nodosilinea 속에서 배제되었다. 그러므로 형태학적 및 분자적
특징들에 기초할 때, 새롭게 분리된 남조류 NIBRIUM-3은 Leptolyngbya 속에
속하는 것으로 분류되었으며, 최종적으로 Leptolyngbya sp. NIBRIUM-3으로
표기하였다.

- 37 -
그림 III-6. Leptolyngbya sp. NIBRIUM-3의 계통분류도. Section
III(Oscillatoriales) 남조류 NIBRIUM-3의 16S rRNA 유전자를 이용한 Clade
credibility 계통분류(a) 및 16S-23S ITS 유전자를 이용한 MP 계통분류(b) 분석이며,
Outgroup으로 Gloeobacter violaceus PCC 7421(GenBank accession No. AF132790)
및 Oscillatoria sp. PCC 9240 (GenBank accession No. EF061079)를 사용함.

- 38 -
(4) NIBRIUM-4
그림 III-7. Myxosarcina sp. NIBRIUM-4의 형태 분석. 제주특별자치도
제주시 구좌읍 미세조류 배양장에서 채집되어 순수 분리된 Section
II(Pleurocapsales) 남조류 NIBRIUM-4의 광학현미경(A), 주사전자현미경(B)
및 투과전자현미경(C, D) 검경 결과.
(가) 생태․형태적 특징[Cyanophyta Section II, Pleurocapsales]
본 미세조류주는 제주특별자치도 제주시 구좌읍 미세조류 배양장에서
분리되었으며 채집 당시 군체 및 독립 형태 모두 발견되었다. 채집지 수온
(24.1°C), 조도(206.2 μmol photon/m2/s), 염도(10.1 psu) 및 pH(8.3) 등의 환경
조건이 측정되었다.
형태학적 분석(그림 III-7)에서 남조류 세포들은 직경 3-5 µm 크기로
둥근 정육면체부터 구 모양까지 다양한 형태를 보였고, 황갈색의 남조류 세포는
4-64개의 집합된 비극성 blastoparenchymatous세포로 분열하여 증식하였다.
분열 중 세포체의 둘레는 뚜렷한 합입점으로 구성되었고, 영양세포는 불규칙
적으로 3 방향으로 이분열에 의해 분열되거나, Baeocyte 생성을 통해 증식
하였다. 이러한 형태적 특성을 통하여 NIBRIUM-4는 기수에 서식하는

- 39 -
Section II(Pleurocapsales) 남조류로 분류되었다.
(나) 분자생물학적 분류[Pleurocapsales, Myxosarcina(H.Printz, 1921)]
Section II(Pleurocapsales)로 분류된 NIBRIUM-4 남조류의 분류학상 위치
를 결정하기 위하여, 획득된 1123 bp의 16S rRNA 서열을 BLASTN 검색을
이용하여 남조류의 GenBank 데이터베이스와 비교하였다. 비교 결과, 유사한
남조류는 Myxosarcina 속, Chroococcidiopsis 속과 Dermocapella 속에 포함
되어 있는 종들로 판별되었다(그림 III-8). 그 중, 가장 유사한 4개의 종은
Myxosrcina sp. LEGE 06146(GenBank accession No. HQ832897),
Chroococcidiopsis sp. CCMP1489(GenBank accession No. AJ344556),
Dermocarpella incrassata PCC7326(GenBank accession No. AJ344559) 및
Chroococcidiopsis sp. CCMP2623(GenBank accession No. JF810074)으로
각각 98.0%, 97.6%, 97.5% 및 97.5% 서열의 유사도를 나타냈다. 16S rRNA
염기서열 유사도 상 NIBRIUM-4는 Myxosarcina 속에 가장 가깝지만
Chroococcidiopsis 및 Dermocapella 속들과 구분되지 않았다. 따라서 분리된
종의 정확한 계통학적 위치를 결정하기 위해 증식 단계에서 baeocytes나
nanocytes 형태를 갖는 남조류의 16S rRNA를 이용하여 ML 계통분류법으로
계통수를 재구성하였다. 계통학적 분석 결과, NIBRIUM-4 남조류는 Myxosarcina
sp. LEGE 06146, Chroococcidiopsis sp. CCMP1489와 Chroococcidiopsis sp.
CCMP2623와 동일군을 이루었고, 반면에 D. incrassata SAG 29.84 및 Myxosarcina
sp. PCC 7312와 뚜렷한 다른 그룹을 형성하였다. 이러한 결과를 바탕으로 현재
Myxosarcina 속은 다계통군이고, Chroococcidiopsis 속 및 Dermocapella 속과
명확히 구분되지 않는 것으로 판단되었다.
가장 높은 16S rRNA 서열 유사도를 보였던 3개의 속 (Myxosarcina,
Chroococcidiopsis 및 Dermocarpella) 중에서 Myxosarcina 속과
Chroococcidiopsis 속은 번식단계 동안 nanocytes를 형성하는 반면,
Dermocarpella 속은 baeocytes를 형성하는 것으로 알려져 있다 (Komárek &
Anagnostidis 1986). 이러한 nanocytes의 운동성은 Chroococcidiopsis 속과
Myxosarcina 속간의 분류에서 가장 중요한 특징으로, Myxosarcina 속은
Chroococcidiopsis 속과 다르게 특이한 운동성 있는 nanocytes를 가지는 것으로
알려져 있다 (Waterbury and Stanier 1978). 이와 같은 결과를 토대로 본
연구에서 분리한 군체 형태의 남조류 NIBRIUM-4는 운동성 있는 nanocytes를
형성하였으며 계통분류 상 Myxosarcina 속의 다른 분리주들과 높은 유사도를
나타내었기에, Myxosarcina sp. NIBRIUM-4로 명명되었다.

- 40 -
그림 III-8. Myxosarcina sp. NIBRIUM-4의 계통분류도. Section
II(Pleurocapsales) 남조류 NIBRIUM-4의 16S rRNA 유전자를 이용한 ML
계통분류 분석이며, Outgroup으로 Gloeobacter violaceus PCC 7421(GenBank
accession No. AF132790)를 사용함.

- 41 -
(5) NIBRIUM-5
그림 III-9. Lyngbya sp. NIBRIUM-5의 형태 분석. 전라북도 금강 하구둑
부근에서 채집되어(A), 순수 분리된 Section III(Oscillatoriales) 남조류
NIBRIUM-5의 광학현미경(B, C) 검경 결과.
(가) 생태․형태적 특징[Cyanophyta Section III, Oscillatoriales]
본 미세조류주는 전라북도 금강 하구둑 부근 기수계에서 분리되었으며,
유속이 적은 갈대숲 부근에서 확인되었다. 채집 당시 독립체로 주로 수표면에서
부유 상태로 발견되었다. 채집 당시 수온(20.1°C), 조도(108.6 μmol
photon/m2/s), 염도(1.7 psu) 및 pH(8.0) 등의 환경조건이 측정되었다.
광학현미경(그림 III-5) 분석에서 관찰된 NIBRIUM-5는 non-branched 단일
사상체(trichome)로 이루어져 있으며, 50-200 개의 디스크(disc) 형상의 개별
세포들로 구성된 사상체는 0.5-2.1 mm까지 다양한 크기로 측정되었다, 운동성
사상체는 대부분 곧거나(straight) 또는 구부러진(bent) 형태로 증식하였으며,
sheath, heterocysts, akinetes, nodules 또는 necridic cell 등은 발견되지 않았다.
각각의 세포들은 길이가 4.0-9.1 μm, 지름이 0.6-1.1 μm였으며, 사상체 내 세포
간 cross-wall의 constriction은 명확하였다. 정단 세포(apical cell)들은 내피막
(calyptra) 없이 뭉툭한 원뿔 형상이었다. 이러한 형태적 특성을 통하여

- 42 -
NIBRIUM-5은 기수에 서식하는 Section III(Oscillatoriales) 남조류로 분류되었다.
(나) 분자생물학적 분류[Oscillatoriales, Lyngbya(Agardh ex Gomont, 1892)]
Section III(Oscillatoriales)로 분류된 NIBRIUM-5 남조류의 분류학상 위치
를 결정하기 위하여, 획득된 1420 bp의 16S rRNA 서열을 BLASTN 검색을
이용하여 남조류의 GenBank 데이터베이스와 비교하였다. 그 결과 유사한
서열들의 대부분이 Arthrospira(일반적으로 Spirulina로 명칭되는 남조류류)
속 및 Lyngbya 속에 속하는 균주 유래라는 것이 밝혀졌다. BLAST 검색에
의하여 확인된 가장 유사한 균주는 Arthrospira platensis PCC 7345(GenBank
accession No. NR125711)로, 99.6%의 서열 동일성을 갖는다. 또한
NIBRIUM-1, 2와 마찬가지로 현재 분류학상으로 받아들여지는 Arthrospira
속 내의 종들과 매우 높은 유사도를 나타내었다. 그러나 Arthrospira 속으로
분류된 NIBRIUM-1, 2 남조류주와의 형태학적 비교 결과, 사상체의 형태가
코일이나 물결형이 아닌 직선형으로 일반적으로 알려진 Arthrospira 속 형태와
차이를 나타내었다. 또한 형태학적으로 유사한 것으로 판정되는 Lyngbya 속
남조류주들과 유사도 비교를 통해 Lyngbya aestuarii PCC 7419(GenBank
accession No. NR114680)와 95.1%의 유사도로 가장 유사한 것으로 나타났다.
이러한 유사도 결과를 기반으로 남조류 NIBRIUM-5의 정확한 계통분류학
적 위치를 확인하기 위하여 Arthrospira 속 및 Lyngbya 속을 포함하는
Oscillatoriales 내 일부 유사균주를 포함하는 남조류 유래 24개 데이터 세트의
16S rRNA 서열을 이용하여 NJ 및 ML 계통분류학적 분석을 수행하였다. 상기
두 가지의 계통분석 결과 Oscillatoriales 내에서 Arthrospira 속과 Lyngbya 속은
독립된 그룹을 형성하지 못하였으며, 해당 그룹 내에 NIBRIUM-5가 포함되는
것을 확인하였다(그림 III-10). NIBRIUM-1 및 2와 관련하여 상기 언급하였던
것처럼 현재 Arthrospira 속은 maxima, platensis 등 총 24종이 분류학상으로
인정되고 있으며(http://www.algaebase.org/), 종간 분류는 매우 난해하여
Arthrospira 속 내의 종에 대한 재분류가 필요한 것으로 인정되고 있다. 또한
본 계통분류학적 분석을 통하여 Lyngbya 속이 별도로 Arthrospira 속에서 분리
되지 않아 NIBRIUM-5은 명확히 동정되지 못하였다. 그러나 형태적 특징만
으로도 본 남조류주는 Arthrospira 속에서 배제되어야 할 것으로 판명되어,
따라서 NIBRIUM-5 남조류주는 Lyngbya 속으로만 분류되어 최종적으로
Lyngbya sp. NIBRIUM-5로 명명되었다.

- 43 -
그림 III-10. Lyngbya sp. NIBRIUM-5의 계통분류도. Section
III(Oscillatoriales) 남조류 NIBRIUM-5의 16S rRNA 유전자를 이용한 ML
계통분류 분석이며, Outgroup으로 Escherichia coli K12(GenBank accession
No. AE000460)를 사용함.

- 44 -
나) 녹조류(Chlorophyta)
(1) NIBRIUM-6
그림 III-11. Chlamydomonas sp. NIBRIUM-6의 형태 분석. 경기도 안산시
미세조류 배양장에서 채집되어 순수 분리된 쌍편모 녹조류 NIBRIUM-6의
광학현미경(A, B), 주사전자현미경(C) 및 투과전자현미경(D) 검경 결과.
편모는 각각 검정 및 흰색 화살표로 표지됨.
(가) 생태․형태적 특징[Chlorophyta, Chlorophyceae, Chlamydomonadales]
본 미세조류주는 경기도 안산시 미세조류 배양장에서 분리되었으며 채집
당시 편모를 지닌 형태로 발견되었다. 채집지 수온(16.1°C), 조도(113.9 μmol
photon/m2/s), 염도(0.0 psu) 및 pH(7.3) 등의 환경조건이 측정되었다.
광학현미경 분석 결과, 한 쌍의 편모를 지닌 타원형 영양세포 (직경 4-6
µm), 접합자 (직경 11-14 µm) 또는 팔멜라 세포 (직경 4-7 µm)들이 관찰되었
다. 영양세포단계에서는 난형의 피레노이드가 엽록체에 의해 둘러싸여 있었고,
기저체에 붙어있는 두 개의 편모 (길이 8-10 µm, 너비 0.2 µm)가 관찰되었다.
초미세구조를 분석한 결과, 핵, 골지체, 엽록체, 피레노이드, 미토콘드리아,

- 45 -
전분과립과 지질과립 등 세포 소기관이 관찰되었다. 세포 중앙에 위치한 핵은
골지체에 둘러싸여 있었고, 세포의 가장자리에 위치한 컵 모양의 엽록체는
세포 내 기질의 대부분을 차지하고 있었다. 또한 타원형의 피레노이드는 큰 전분
과립에 의해 둘러싸여 있었다. 무성 생식기 동안, 영양세포는 2개의 유주자로
나뉘어졌고, 팔멜라 단계에서 포자낭은 각 세포벽으로 에워싸인 4개의 유주자를
지니고 있다. 여러 개의 상대적으로 큰 전분과립은 분열시기에 세포 내에서
관찰된 반면, 지질 입자는 대부분 영양세포의 세포질에서 관찰되었다. 또한,
미세조류 편모 구조 관찰 시, 축사의 단면구조에서 “9+2” 구조(Ringo, 1967)로
잘 알려져 있는 9개의 이중 미세소관과 중심부에 2개의 중심미세소관이 관찰
되었다. 한 쌍의 편모는 각각 세포질에서 편모뿌리와 미세소관, 섬유성 구성성분에
의해 고정되어 있는 기저체에 연결되어 있었다. 이러한 형태적 특성을 통하여
NIBRIUM-6는 담수에 서식하는 쌍편모 녹조류로 분류되었다.
(나) 분자생물학적 분류[Chlamydomonadales, Chlamydomonas(Ehrenberg, 1833)]
Chlorophyceae(Chlamydomonadales)로 분류된 NIBRIUM-6 녹조류의 분류학
상 위치를 결정하기 위하여, 획득된 1717 bp의 18S rRNA와 626 bp의 ITS
region 서열을 BLASTN 검색을 이용하여 녹조류의 GenBank 데이터베이스와
비교하였다. 18S rRNA염기서열에 대한 BLASTN 결과에 따르면,
Chlamydomonas baca CCAP 11/77(GenBank accession No. FR865613)가
97.7%의 가장 높은 유사성을 보였고, 그 다음으로 C. sphagnophila CCAP
11/31(GenBank accession No. FR865573)는 97.4%, C. mexicana CCAP 11/55A
(GenBank accession No. FR865592)는 97.2 %, C. rapa SAG 48.72 (GenBank
accession No. U70790)는 97.0%의 유사성을 보였다. Chlamydomonas 속의 대표
종인 C. reinhardtii CCAP 11/32C (GenBank accession No. FR865575)와는
95.9%의 유사성을 보였다. 또한, GenBank에서 Chlamydomonas 속에 속하는 종의
ITS region 염기서열을 비교한 결과, C. baca CCAP 11/77 (GenBank accession
No. FR865613)가 84.2%의 유사성을 보였고, C. rapa f. vasta CCAP 11/73
(GenBank accession No. FR865611)는 83.9%, C. mexicana CCAP 11/55A
(GenBank accession No. FR865592)는 83.5%의 유사성을 보였다. 18S rRNA 결과와
비슷하게 Chlamydomonas의 ITS region 염기서열 비교 결과, Chlamydomonas
속의 대표종인 C. reinhardtii CCAP 11/32C (GenBank accession No.
FR865575)과 80.2%의 유사성을 보였다.
이러한 유사도 결과를 기반으로 녹조류 NIBRIUM-6의 정확한 계통분류학적
위치를 확인하기 위하여 Chlamydomonas 속 내 일부 유사균주를 포함하는 25

- 46 -
개 데이터 세트의 18 rRNA 서열을 이용하여 NJ 및 ML 계통분류학적 분석을
수행하였다. 상기 두 가지의 계통분석 결과 Chlamydomonas 속 내에서 크게 3
개의 독립적인 그룹으로 나뉘었으며, 그 중 그룹 II 내에 NIBRIUM-6가 포함
되는 것을 확인하였다(그림 III-12). 또한 ITS region 염기서열을 이용한 MP
계통분석 결과, Chlamydomonas 속은 2개의 독립적인 그룹으로 나뉘어 18S
rRNA 계통분석 시 유사한 종으로 판별된 결과와 유사하게 나타났다. 현재
Chlamydomonas 속은 총 589종이 분류학상으로 인정되고 있으며
(http://www.algaebase.org/), 종간 분류는 매우 난해하여 Chlamydomonas 속
내 종에 대한 재분류가 필요한 것으로 인정되고 있다. 이러한 결과를 바탕으로
새롭게 분리된 단일세포의 녹조류는 Chlamydomonas 속으로 분류되었고,
최종적으로 Chlamydomonas sp. NIBRIUM-6으로 명명하였다.

- 47 -
그림 III-12. Chlamydomonas sp. NIBRIUM-6의 계통분류도. Chlamydomonas
속으로 분류된 녹조류 NIBRIUM-6의 18S rRNA 유전자를 이용한 ML 계통분류(a)
및 18S-28S ITS 유전자를 이용한 MP 계통분류(b) 분석이며, Outgroup으로
Scenedesmus acutus UTEX 72(GenBank accession No. AJ249508)를 사용함.

- 48 -
(2) NIBRIUM-7
그림 III-13. Haematococcus sp. NIBRIUM-7의 형태 분석. 경기도 안산시
미세조류 배양장에서 채집되어 순수 분리된 쌍편모 녹조류 NIBRIUM-7
영양세포의 광학현미경(A, B) 및 아스타잔틴 축적 cyst의 광학현미경(C, D)
및 투과전자현미경(E, F) 검경 결과. 편모는 흰색 화살표로 표지되었으며,
Sheath는 검은 화살표로 표지됨.

- 49 -
(가) 생태․형태적 특징[Chlorophyta, Chlorophyceae, Volvocales]
본 미세조류주는 경기도 안산시 미세조류 배양장에서 분리되었으며 채집
당시 붉은 색의 아스타잔틴이 농축된 cyst 형태로 발견되었다. 채집지 수온
(16.1°C), 조도(113.9 μmol photon/m2/s), 염도(0.0 psu) 및 pH(7.3) 등의 환경
조건이 측정되었다.
광학현미경 분석 결과, 한 쌍의 편모를 지닌 타원형 영양세포 (직경
15-26 µm), 접합자 또는 cyst 세포 (직경 22-44 µm)들이 관찰되었다. 영양
세포 단계에서는 원형의 피레노이드가 엽록체에 의해 둘러싸여 있었고, 기저체에
붙어있는 두 개의 편모 (길이 9-11 µm, 너비 0.2 µm)가 관찰되었다. 초미세
구조를 분석한 결과, 핵, 골지체, 엽록체, 피레노이드, 미토콘드리아, 전분 및
지질과립 등의 세포 소기관이 관찰되었으며, 붉은 색의 cyst 및 접합자 세포
에서는 아스타잔틴(Astaxanthin) 과립이 특이적으로 발견되었다. 이러한 아스타
잔틴 과립은 cyst 단계로 갈수록 세포 중앙에서부터 농축이 되는 것으로 확인
되었으며, 해당 단계의 세포는 지질과립 역시 다른 단계의 세포에 비하여
다수 포함하고 있는 것으로 확인할 수 있었다. 원형의 피레노이드는 큰 전분
과립에 의해 둘러싸여 있었으며 이러한 전분과립은 상대적으로 많은 세포질
부분을 차지하고 있었다. 또한, 미세조류 편모 구조 관찰 시, 축사의 단면구조
에서 “9+2” 구조(Ringo, 1967)로 잘 알려져 있는 9개의 이중 미세소관과 중심부에
2개의 중심미세소관이 관찰되었다. 이러한 형태적 특성을 통하여 NIBRIUM-7는
아스타잔틴을 생산하는 담수에 서식하는 쌍편모 녹조류로 분류되었다.
(나) 분자생물학적 분류[Volvocales, Haematococcus(Flotow, 1844)]
Chlorophyceae(Volvocales)로 분류된 NIBRIUM-7 녹조류의 분류학상
위치를 결정하기 위하여, 획득된 1748 bp의 18S rRNA와 1102 bp의 rbcL
유전자 서열을 BLASTN 검색을 이용하여 녹조류의 GenBank 데이터베이스와
비교하였다. 18S rRNA염기서열에 대한 BLASTN 결과에 따르면, 최근
Haematococcus pluvialis와 동명처리된 H. lacustris NIES-144(GenBank
accession No. AB360747)가 99.9%의 가장 높은 유사성을 보였고, 그 다음
으로 H. pluvialis SAG 34-1b(GenBank accession No. AF159369)는 99.6%,
H. zimbabwiensis SAG 26.96(GenBank accession No. AB360748)는 95.6%의
유사성을 보였다. 또한 rbcL 유전자의 경우, H. lacustris UTEX 16(GenBank
accession No. EF113446)가 96.2%의 가장 높은 유사성을 나타내었으며, H.
pluvialis CCAP 34/7(GenBank accession No. FJ438476)는 96.0%, H.
zimbabwiensis SAG 26.96(GenBank accession No. AB360757)는 85.8%의

- 50 -
유사성을 나타내었다.
이러한 유사도 결과를 기반으로 녹조류 NIBRIUM-7의 정확한 계통분류학적
위치를 확인하기 위하여 Haematococcus 속 내 일부 유사균주 및
Chlorogonium 속 등의 유사 속을 포함하는 12개 데이터 세트의 18 rRNA 서열
및 10개 데이터 세트의 rbcL 유전자를 이용하여 Maximum clade credibility
tree 계통분류학적 분석을 각각 수행하였다. 상기 두 가지의 계통분석 결과 H.
pluvialis와 NIBRIUM-7이 매우 유사한 것을 확인하였다(그림 III-14). 현재
Haematococcus 속은 총 4종이 분류학상으로 인정되고 있으며
(http://www.algaebase.org/), 속내 종에 대한 재분류가 필요한 것으로 인정되고
있다. 이러한 결과를 바탕으로 새롭게 분리된 단일세포의 편모 녹조류는
Haematococcus 속으로 분류되었고, 최종적으로 Haematococcus sp.
NIBRIUM-7로 명명하였다.

- 51 -
그림 III-14. Haematococcus sp. NIBRIUM-7의 계통분류도.
Haematococcus 속으로 분류된 녹조류 NIBRIUM-7의 18S rRNA 유전자를
이용한 Maximum clade credibility 계통분류(a) 및 rbcL 유전자를 이용한 MP
계통분류(b) 분석이며, Outgroup으로 Pleurastrum insigne SAG 30.93(GenBank
accession No. EF113464)를 사용함.

- 52 -
(3) NIBRIUM-8
그림 III-15. Lobosphaera sp. NIBRIUM-8의 형태 분석. 충청남도 태안의
조간대 지역에서 채집되어 순수 분리된 구형 녹조류 NIBRIUM-8의 광학
현미경(A, B), 주사전자현미경(C) 및 투과전자현미경(D) 검경 결과.
(가) 생태․형태적 특징[Chlorophyta, Trebouxiophyceae, Trebouxiales]
본 미세조류주는 충청남도 태안의 조간대 지역에서 분리되었으며 채집
당시 선형 남조류와 함께 군체로 발견되었다. 채집지 수온(17.4°C), 조도(214.5
μmol photon/m2/s), 염도(31.2 psu) 및 pH(8.0) 등의 환경조건이 측정되었다.
광학현미경 분석 결과, 구형 영양세포 (직경 9-12 µm)는 투명한 다당류로
추정되는 세포 외 물질에 함입되어 군체를 형성한 상태로 관찰되었다. 영양세포
단계에서는 난형의 피레노이드가 엽록체에 의해 둘러싸여 있었으며, 난할을 통한
영양세포 내 분열로 자생포자의 형성이 관찰되었다. 초미세구조를 분석한 결과,
핵, 골지체, 엽록체, 피레노이드, 미토콘드리아, 전분과립과 지질과립 등 세포
소기관이 관찰되었다. 또한 타원형의 피레노이드는 큰 전분 과립에 의해 둘러
싸여 있었다. 무성 생식기 동안, 영양세포는 4-16 개의 딸세포로 나뉘어졌고,
상대적으로 큰 전분과립은 분열시기에 세포 내에서 관찰된 반면, 지질 입자는 대부분
영양세포의 세포질에서 관찰되었다. 이러한 형태적 특성을 통하여 NIBRIUM-8는

- 53 -
기수에 서식하는 원형 녹조류로 분류되었다.
(나) 분자생물학적 분류[Trebouxiales, Lobosphaera(Reisigl, 1964)]
Trebouxiophyceae(Trebouxiales)로 분류된 NIBRIUM-8 녹조류의 분류학
상 위치를 결정하기 위하여, 획득된 1627 bp의 18S rRNA 유전자 서열을
BLASTN 검색을 이용하여 녹조류의 GenBank 데이터베이스와 비교하였다.
18S rRNA염기서열에 대한 BLASTN 결과에 따르면, Lobosphaera incisa
SAG 2466(GenBank accession No. KM020046) 및 Parietochloris ovoidea
ACKU 177-03(GenBank accession No. EU878374)와 99.7%의 가장 높은 유사성을
보였고, Myrmecia 및 Neocystis 등의 유사속들과 98% 이상의 유사도를 나타
내었다. 또한 ITS region의 경우, L. incisa SAG 2466(GenBank accession
No. KM020046)가 93.1%의 가장 높은 유사성을 나타내었으며, Chlorella 및
Micractinium 등의 Chlorellales 내 다양한 속들과 83% 이상의 유사성을 나타
내었다.
이러한 유사도 결과를 기반으로 녹조류 NIBRIUM-8의 정확한 계통분류학적
위치를 확인하기 위하여 Lobosphaera, Myrmecia 및 Parietochloris 속 내
유사균주를 비롯한 Chlorella 및 Micractinium 등 Chlorellales 내 다양한 속들을
포함하는 25개 데이터 세트의 18 rRNA 서열을 이용하여 NJ, ML 계통분류학적
분석을 수행하였다. 상기 계통분석 결과 Lobosphaera 및 Parietochloris 속으로
이루어진 그룹 내에 NIBRIUM-8이 포함되는 것을 확인하였다(그림 III-16).
또한 MP 방법을 이용한 ITS region의 계통분석 시, 18S rRNA 유전자를 이용한
계통분류와 유사한 결과를 도출하는 것을 확인하였다. 현재 Lobosphaera 속은
총 3종이 분류학상으로 인정되고 있으나(http://www.algaebase.org/),
Parietochloris(Watanabe & Floyd, 1990) 및 Myrmecia(Printz, 1920) 등의
유사 속과 재분류의 필요성이 제기되고 있다. 이러한 결과를 바탕으로 새롭게
분리된 단일세포의 녹조류는 Lobosphaera 속으로 분류되었고, 최종적으로
Lobosphaera sp. NIBRIUM-8로 명명하였다.

- 54 -
그림 III-16. Lobosphaera sp. NIBRIUM-8의 계통분류도. Lobosphaera 속
으로 분류된 녹조류 NIBRIUM-8의 18S rRNA 유전자를 이용한 ML 계통분류(a)
및 ITS region을 이용한 MP 계통분류(b) 분석이며, Outgroup으로 Trentepohlia
sp. UTEX 1227(GenBank accession No. AY052569)를 사용함.

- 55 -
(4) NIBRIUM-9
그림 III-17. Scenedesmus sp. NIBRIUM-9의 형태 분석. 제주특별자치도
제주시 구좌읍 미세조류 배양장에서 채집되어 순수 분리된 구형 녹조류
NIBRIUM-9 자생 군체의 광학현미경(A, B) 검경 결과.
(가) 생태․형태적 특징[Chlorophyta, Chlorophyceae, Sphaeropleales]
본 미세조류주는 제주특별자치도 제주시 구좌읍 미세조류 배양장에서
분리되었으며 채집 당시 자생 군체의 형태로 발견되었다. 채집지 수온
(24.1°C), 조도(206.2 μmol photon/m2/s), 염도(10.1 psu) 및 pH(8.3) 등의 환경
조건이 측정되었다.
광학현미경 분석 결과, 뗏목 형태의 타원형 영양세포(직경 9-11 µm)가
나란히 측면으로 배열하였으며, 2-16개의 세포가 연결된 형태의 자생 군체로
증식하였다. 세포는 피레노이드를 갖는 하나의 엽록체를 가지며 단핵으로 구성
되어 있었다. 생식 과정 동안 유주자가 형성되지 않았으며, 모세포는 세포벽을
붕괴하지 않은 상태에서 편모를 갖지 않는 세포들로 나뉜 후 나란히 측면으로
배열되었다. 세포벽의 붕괴는 연속된 분열을 통하여 최소 4개 이상의 핵이 형성
된 이후에 시작되는 것이 관찰되었다. 타원형의 양 끝 부분에는 가시 모양의
구조물이 관찰되지 않았다. 이러한 형태적 특성을 통하여 NIBRIUM-9는 기수에
서식하는 뗏목 형태의 가시가 없는 녹조류로 분류되었다.
(나) 분자생물학적 분류[Sphaeropleales, Scenedesmus(Meyen, 1829)]
Chlorophyceae(Sphaeropleales)로 분류된 NIBRIUM-9 녹조류의 분류학상
위치를 결정하기 위하여, 획득된 1611 bp의 18S rRNA 유전자 서열을
BLASTN 검색을 이용하여 녹조류의 GenBank 데이터베이스와 비교하였다.

- 56 -
18S rRNA염기서열에 대한 BLASTN 결과에 따르면, Scenedesmus obliquus
UTEX1450(GenBank accession No. AJ249515)가 99.7%의 가장 높은 유사성을
보였고, 다음으로 S. acuminatus Hegewald 1986-2(GenBank accession No.
AB037088)는 99.6%, S. deserticola BCP-SNI-2(GenBank accession No.
AY510462)는 99.6%의 유사성을 보였다.
이러한 유사도 결과를 기반으로 녹조류 NIBRIUM-9의 정확한 계통분류학적
위치를 확인하기 위하여 Scenedesmus 속 내 유사균주 및 Coelastrum 속 등의
유사 속을 포함하는 25개 데이터 세트의 18 rRNA 서열을 이용하여 NJ, ML
계통분류학적 분석을 수행하였다. 상기 계통분석 결과 Scenedesmus 속으로
이루어진 그룹 내에 NIBRIUM-9가 포함되는 것을 확인하였다(그림 III-18).
현재 Scenedesmus 속은 총 102종이 분류학상으로 인정되고 있으나
(http://www.algaebase.org/), Acutodesmus(Tsarenko, 2001) 및 Desmodesmus
(Friedl & Hegewald, 1999) 등의 속과 재분류의 필요성이 제기되고 있다. 이러한
결과를 바탕으로 새롭게 분리된 단일세포의 녹조류는 Scenedesmus 속으로
분류되었고, 최종적으로 Scenedesmus sp. NIBRIUM-9로 명명하였다.

- 57 -
그림 III-18. Scenedesmus sp. NIBRIUM-9의 계통분류도. Scenedesmus 속
으로 분류된 녹조류 NIBRIUM-9의 18S rRNA 유전자를 이용한 ML 계통분류
분석이며, Outgroup으로 Trentepohlia sp. UTEX 1227(GenBank accession No.
AY052569)를 사용함.

- 58 -
(5) NIBRIUM-10
그림 III-19. Chlorella sp. NIBRIUM-10의 형태 분석. 제주특별자치도
제주시 구좌읍 미세조류 배양장에서 채집되어 순수 분리된 구형 녹조류
NIBRIUM-10 자생 군체의 광학현미경(A, B) 검경 결과.
(가) 생태․형태적 특징[Chlorophyta, Trebouxiophyceae, Chlorellales]
본 미세조류주는 제주특별자치도 제주시 구좌읍 미세조류 배양장에서
분리되었으며 채집 당시 자생 군체의 형태로 발견되었다. 채집지 수온
(24.1°C), 조도(206.2 μmol photon/m2/s), 염도(10.1 psu) 및 pH(8.3) 등의 환경
조건이 측정되었다.
광학현미경 분석 결과, 구형 영양세포 (직경 4-6 µm)는 투명한 다당류로
추정되는 세포 외 물질에 함입되어 군체를 형성한 상태로 관찰되었다. 영양
세포 단계에서는 난형의 피레노이드가 엽록체에 의해 둘러싸여 있었으며, 난할을
통한 영양세포 내 분열로 자생포자의 형성이 관찰되었다. 타원형의 피레노이드는
큰 전분 과립에 의해 둘러싸여 있었다. 타원체 모양의 형태인 자생포자는 하나의
밋밋한 층으로 이루어진 세포벽을 가지는 것으로 확인되었다. 무성 생식기
동안, 영양세포는 2-8 개의 딸세포로 나뉘어졌고, 상대적으로 큰 전분과립은
분열시기에 세포 내에서 관찰된 반면, 지질 입자는 대부분 영양세포의 세포질
에서 관찰되었다. 이러한 형태적 특성을 통하여 NIBRIUM-10은 기수에 서식하는
원형 녹조류로 분류되었다.
(나) 분자생물학적 분류[Chlorellales, Chlorella(Beijerinck, 1890)]
Trebouxiophyceae(Chlorellales)로 분류된 NIBRIUM-10 녹조류의 분류학
상 위치를 결정하기 위하여, 획득된 1631 bp의 18S rRNA 유전자 서열을

- 59 -
BLASTN 검색을 이용하여 녹조류의 GenBank 데이터베이스와 비교하였다.
18S rRNA염기서열에 대한 BLASTN 결과에 따르면, Chlorella sorokiniana
SAG 211-8k(GenBank accession No. X62441)가 99.8%의 가장 높은 유사성을
보였고, 다음으로 C. lobophora Andreyeva 750-I(GenBank accession No.
X63504)는 99.3%, C. vulgaris SAG 211-11b(GenBank accession No. X13688)는
99.3%의 유사성을 보였다.
이러한 유사도 결과를 기반으로 녹조류 NIBRIUM-10의 정확한 계통분류학적
위치를 확인하기 위하여 Chlorellales 과 내외의 다양한 속을 포함하는 38개
데이터 세트의 18 rRNA 서열을 이용하여 NJ, ML 계통분류학적 분석을 수행
하였다. 상기 계통분석 결과 Chlorella 속으로 이루어진 그룹 내에
NIBRIUM-10이 포함되는 것을 확인하였다(그림 III-20). 현재 Chlorella 속은
총 43종이 분류학상으로 인정되고 있으며(http://www.algaebase.org/),
Chlorellales 과 내에 다양한 속이 인정되고 있다. 이러한 결과를 바탕으로 새롭게
분리된 단일세포의 녹조류는 Chlorella 속으로 분류되었고, 최종적으로
Chlorella sp. NIBRIUM-10으로 명명하였다.

- 60 -
그림 III-20. Chlorella sp. NIBRIUM-10의 계통분류도. Chlorella 속으로
분류된 녹조류 NIBRIUM-10의 18S rRNA 유전자를 이용한 ML 계통분류 분석
이며, Outgroup으로 Ulva curvata(GenBank accession No. AF189078)를 사용함.

- 61 -
미세조류주 배양배지
배양 조건Productivity
(건중량/5L)pH염도(psu)
온도(°C)
광도*배양기간
(days)
NIBRIUM-1Zarrouk's
medium8.2 12 25.0 80 25 ≈4.4g
NIBRIUM-2Zarrouk's
medium8.2 12 25.0 80 25 ≈4.0g
NIBRIUM-3Zarrouk's
medium8.2 12 25.0 80 25 ≈3.8g
NIBRIUM-4f/2
medium8.0 33 25.0 80 60 ≈2.2g
NIBRIUM-5Zarrouk's
medium8.2 12 25.0 80 25 ≈3.9g
NIBRIUM-6BG-11
medium7.2 0.0 25.0 80 25 ≈1.3g
NIBRIUM-7BG-11
medium7.2 0.0 25.0 80 20 ≈1.2g
NIBRIUM-8BG-11
medium7.2 0.0 25.0 80 28 ≈1.5g
NIBRIUM-9BG-11
medium7.2 0.0 25.0 80 21 ≈3.0g
NIBRIUM-10BG-11
medium7.2 0.0 25.0 80 21 ≈2.8g
2. 자생 미세조류 유용성 검증
가. 연구결과
1) 미세조류 배양체의 성분 분석용 바이오매스 확보
동정이 완료된 10주의 미세조류 배양체의 바이오매스 확보를 위하여 5L급
폭기 액체배양법을 통하여 배양을 진행하였다. 배양 시 영양배지는 순수분리에
사용된 동일한 영양염 조성을 사용하였으며, 각 미세조류주의 배양조건, 배양
기간 및 최종 생산량(g/5L)은 표 III-2와 같다.
표 III-2. 자생 미세조류주 배양조건 및 최종 생산량
(* μmol photons/m2/s)

- 62 -
남조류성분 함량(%)
회분(Ash)
다당류(Carbohydrate)
조지질(Lipid)
수분(Moisture)
조단백(Protein)
NIBRIUM-1 20.3 ± 0.3 17.8 ± 1.2 9.1 ± 0.5 2.2 ± 0.1 50.7 ± 0.3
NIBRIUM-2 10.9 ± 0.8 30.8 ± 1.6 11.6 ± 0.2 3.8 ± 0.1 42.9 ± 0.5
NIBRIUM-3 10.2 ± 0.1 24.5 ± 0.8 11.4 ± 0.5 1.3 ± 0.1 52.6 ± 0.1
NIBRIUM-4 27.7 ± 0.5 31.5 ± 1.1 18.8 ± 0.4 2.7 ± 0.1 19.4 ± 0.1
NIBRIUM-5 7.8 ± 1.0 1.2 ± 3.9 18.8 ± 2.1 1.6 ± 0.8 70.5 ± 0.1
2) 미세조류 배양체 바이오매스의 일반성분 분석 및 평가
가) 남조류 일반성분 분석 및 평가
연구방법에 제시된 바와 같이 동정이 완료된 5주의 남조류 배양체의 일반성분
(회분, 조탄수화물, 조지질, 수분 및 조단백) 분석을 Official Methods of Analysis
of the Association of Official Analytical Chemists(AOAC)에 따라 진행하였다.
5L급 폭기 배양조에서 수확된 남조류 바이오매스 10g씩을 분석에 사용하였으며,
분석결과는 표 III-3에 제시되었다.
표 III-3. 자생 남조류 일반성분 분석 결과
그림 III-21. 자생 남조류 5주의 일반성분 분석결과. Lyngbya 속으로 분류된
NIBRIUM-5의 경우 Arthrospira 속의 결과를 뛰어넘는 70% 이상의 조단백
함량을 지니는 것으로 확인됨.

- 63 -
분석결과 확보된 사상체형의 자생 남조류주(NIBRIUM-1, 2, 3, 5)는 전체
성분 중 50% 이상이 조단백으로 분석되었다. 일반적으로 Arthrospira 속에
포함되는 남조류는 대부분 높은 조단백 함량을 갖는 것으로 보고되고 있으며,
이는 본 연구에서 분석된 Arthrospira 속 NIBRIUM-1 및 2 남조류주에 있어
유사한 경향을 나타내었다. 흥미로운 것은 Arthrospira 속이 아닌 NIBRIUM-3
및 5 남조류주의 경우, 오히려 해당 속의 조단백 함량을 상회하는 것으로 나타난
점이다. 특히 NIBRIUM-5 남조류주의 경우 70% 이상이 조단백으로 구성되어
있는 것으로 나타났으며(그림 III-21), 이는 사료나 식품으로써의 가능성을
뒷받침한다.
상기와 같은 사상체형의 남조류주와 달리 구형의 남조류주 NIBRIUM-4의
경우, 회분의 함량이 27% 이상 나타났으며, 이는 f/2 medium 제작 시 사용된
해수 내에 포함된 염이 일정 부분 기여를 한 것으로 추정된다. 본 남조류주의
경우 다당류와 지질의 함량이 다른 남조류주에 비하여 상대적으로 높으며, 조
단백의 함량은 상대적으로 낮은 것으로 분석되었다. 이는 난할을 통해 분열하는
것으로 관찰된 남조류주의 특성 상 세포 내질 외의 세포 간 혹은 세포 외질이
분열 상태의 모세포에서 많은 것으로 추정되며, 이러한 부분을 다당류 중합체가
차지하여 상기와 같은 분석결과를 나타낸 것으로 파악된다. 또한 조지질의 경우
NIBRIUM-4 및 5 남조류주가 타 남조류주에 비하여 유의적으로 높은 것을
확인할 수 있었다. 이를 통하여 식품이나 사료로 활용성이 높은 것으로 이미 보고된
Arthrospira 속 NIBRIUM-1 및 2 남조류주 이외에, Leptolyngbya와 Lyngbya
속으로 각각 동정된 NIBRIUM-3 및 5 남조류주 역시 사료나 식품으로써 효용성이
있을 것으로 추정된다.
나) 녹조류 일반성분 분석 및 평가
연구방법에 제시된 바와 같이 동정이 완료된 5주의 녹조류 배양체의 일반성분
(회분, 조탄수화물, 조지질, 수분 및 조단백) 분석을 Official Methods of Analysis
of the Association of Official Analytical Chemists(AOAC)에 따라 진행하였다.
5L급 폭기 배양조에서 수확된 녹조류 바이오매스 6g씩을 분석에 사용하였으며,
분석결과는 표 III-4에 제시되었다.

- 64 -
녹조류주성분 함량(%)
회분(Ash)
다당류(Carbohydrate)
조지질(Lipid)
수분(Moisture)
조단백(Protein)
NIBRIUM-6 5.2 ± 0.2 16.8 ± 1.1 20.6 ± 0.8 3.9 ± 0.1 53.5 ± 0.8
NIBRIUM-7 4.9 ± 0.8 48.8 ± 1.8 22.2 ± 0.9 3.1 ± 0.1 21.1 ± 0.2
NIBRIUM-8 6.2 ± 0.0 28.1 ± 0.6 20.4 ± 0.3 1.9 ± 0.0 43.5 ± 0.3
NIBRIUM-9 5.4 ± 0.4 23.7 ± 3.5 25.0 ± 2.7 2.0 ± 0.1 43.9 ± 0.5
NIBRIUM-10 4.7 ± 0.8 29.4 ± 4.6 22.8 ± 2.6 2.4 ± 0.7 40.7 ± 0.8
표 III-4. 자생 녹조류 일반성분 분석 결과
그림 III-22. 자생 녹조류 5주의 일반성분 분석결과. Haematococcus 속으로
분류된 NIBRIUM-7의 경우 타 녹조류주와 달리 낮은 조단백 함량 및 높은 다당류
함량을 특징적으로 나타내는 것이 확인됨.
분석결과 확보된 녹조류 5주의 경우, 먼저 강(class)에 따른 그룹화
(Chlorophyceae: NIBRIUM-6, 7, 9; Trebouxiophyceae: NIBRIUM-8, 10)를 통해
일반성분의 추세를 분석하였다. 분석결과, 동일한 녹조류 강(class)에 포함되는
배양체의 일반성분은 경향성을 나타내지 않는 것을 확인할 수 있었다. 다음으로
목(order)에 따른 구분(Chlamydomonadales: NIBRIUM-6; Volvocales:
NIBRIUM-7; Trebouxiales: NIBRIUM-8; Sphaeropleales: NIBRIUM-9;
Chlorellales: NIBRIUM-10)을 통하여 일반성분 결과를 비교·분석하였다. 분석
결과, Volvocales 목(order)의 Haematococcus sp. NIBRIUM-7 녹조류주만이

- 65 -
특이적으로 낮은 조단백 함량 및 높은 다당류 함량을 나타내었다. 그러나 타 목
(order)의 녹조류주는 조지방 20% 이상 및 조단백 40% 이상의 유사한 세포 일반
성분 함량을 지니는 것으로 분석되었다(표 III-4; 그림 III-22). 이와 같은 분석
결과를 통하여 금번 확보한 녹조류는 사료·식품으로 사용 가능한 점을 우선
확인하였으며, 다음으로 20% 이상의 조지방 함량을 지닌 점을 이용한 바이오
디젤 생산 녹조류주로써의 활용 가능성을 확인할 수 있었다. 최근 미세조류 유래
다당류를 발효하여 바이오에탄올을 생산하는 것이 연구되고 있으나, 아직까지는
바이오매스 대량 생산에 있어서 문제점이 제기되고 있는 실정으로 다당류 성분
함량과 관련된 분석은 그 판단을 유보하고자 한다. 다당류 함량이 높은 것으로
분석된 Haematococcus sp. NIBRIUM-7의 경우 항산화능이 강한 고부가가치
물질로 알려진 Astaxanthin을 대량 생산할 수 있는 활용성이 높은 녹조류주로
가능성이 매우 높다고 할 수 있다. 현재 Astaxanthin은 대부분이 인공적으로
합성된 형태로 전량 수입되고 있으며, 따라서 본 물질을 생산할 수 있는 자생
미세조류를 확보한 것은 매우 고무적인 성과라 할 것이다.
상기와 같은 분석결과를 통해서 금번 배양체 바이오매스의 세부성분 분석은
현재 활용성이 입증된 조단백 및 조지질과 관련된 아미노산 및 지방산 조성
분석에 초점을 맞추어 진행하였다.

- 66 -
3) 미세조류 배양체 바이오매스의 세부성분 분석 및 평가
가) 미세조류 유래 유리 아미노산 분석 및 평가
(1) 남조류 유래 아미노산 분석 및 평가
연구방법에 제시된 바와 같이 동정이 완료된 5주의 남조류 배양체의 세포 내
유리 아미노산 분석을 Amino acid Analyzer (Sykam S433; Sykam)로 분석하였다.
5L급 폭기 배양조에서 수확된 남조류 바이오매스 1g씩을 분석에 사용하였으며,
분석결과는 그림 III-23에 제시되었다.
그림 III-23. 자생 남조류 5주의 아미노산 조성(%) 분석결과.

- 67 -
분석결과 확보된 자생 남조류주는 형태에 따라 상이한 아미노산 조성을
나타내었다. 사상체형과 구형 모두 공통적으로 Aspartic-acid, Glutamic acid,
Alanine, Leucine 등을 높은 비율로 함유하는 것으로 나타났으나, Tyrosine,
Arginine, Prolin 등의 함량에 있어 극명한 차이를 나타내었다. 구형 남조류주인
Myxosarcina sp. NIBRIUM-4의 경우, Tyrosine, Arginine 및 Prolin이 검출되지
않았으나, 사상체형의 남조류주 4주는 높은 비율로 해당 아미노산을 함유하는
것으로 확인되었다. 또한 Myxosarcina sp. NIBRIUM-4는 Valine, Histidine 및
Lysine을 사상체형 남조류주보다 상대적으로 높은 비율로 함유하는 것을 특징
으로 하였다. Cystine은 5주의 남조류주 모두 검출되지 않았으며, Ammonia는
Lyngbya sp. NIBRIUM-5 남조류주에서만 확인되지 않았다. 이러한 결과에
기반하여, 남조류 세포의 아미노산 조성은 추후 형태적 분류에 있어 또 하나의
분류 key로써 사용될 수 있는 가능성이 제시된다.
(2) 녹조류 유래 아미노산 분석 및 평가
연구방법에 제시된 바와 같이 동정이 완료된 5주의 녹조류 배양체의 세포 내
유리 아미노산 분석을 Amino acid Analyzer (Sykam S433; Sykam)로 분석하였다.
5L급 폭기 배양조에서 수확된 녹조류 바이오매스 1g씩을 분석에 사용하였으며,
분석결과는 그림 III-24에 제시되었다.
분석결과 확보된 자생 녹조류주는 앞서 제시되었던 남조류주의 아미노산
분석결과와 마찬가지로 Aspartic-acid, Glutamic acid, Alanine, Leucine 등을
높은 비율로 함유하는 것으로 확인되었다. 이에 반하여 Cystine은 5주 모두
검출되지 않았으며, Tyrosine은 매우 낮은 함량으로만 측정되었다. 특이적으로
Chlamydomonas sp. NIBRIUM-6, Haematococcus sp. NIBRIUM-7 및
Lobosphaera sp. NIBRIUM-8 녹조류주는 Methionine이 전혀 검출되지 않았다.
또한 Haematococcus sp. NIBRIUM-7은 Arginine 및 Prolin이 검출되지 않았
으며, Lobosphaera sp. NIBRIUM-8는 Prolin의 함량이 다른 3종의 녹조류에
비하여 특이적으로 낮았다. Scenedesmus sp. NIBRIUM-9와 Chlorella sp.
NIBRIUM-10은 특이적으로 다른 3주와 비교하여 Phenylalanine과 Prolin의
함량이 상대적으로 높았으며, Histidine과 Lysine의 함량은 상대적으로 낮음을
확인하였다. 본 연구는 BG-11 영양배지 상에서의 바이오매스 아미노산 조성
분석으로 한계점을 지니며, 추후 다른 배양조건에서의 아미노산 조성을 확인하여,
배양조건 별 아미노산 함량의 변화를 모니터할 필요가 제기된다.

- 68 -
그림 III-24. 자생 녹조류 5주의 아미노산 조성(%) 분석결과.

- 69 -
나) 미세조류 유래 지방산 분석 및 평가
(1) 남조류 유래 지방산 분석 및 평가
연구방법에 제시된 바와 같이 동정이 완료된 5주의 남조류 배양체 유래 지방
산 조성은 GC-mass spectrometry detector(GC-MSD)를 이용하여 분석하였다.
5L급 폭기 배양조에서 수확된 남조류 바이오매스 1g씩을 분석에 사용하였으며,
분석결과는 그림 III-25 및 그림 III-26에 제시되었다.
그림 III-25. 자생 남조류 5주의 세부 지방산 조성(%) 분석결과.
분석결과 Arthrospira 속 2주(NIBRIUM-1 및 2)는 매우 유사한 지방산 조성을

- 70 -
나타내었다. 먼저 포화지방산 C16:0 및 C18:0 등이 전체 지방산 함량 중 60% 이상을
차지하였으며, 단일불포화지방산은 C16:1 및 C18:1 계열이 전체 지방산 함량의
18% 이상을, 또한 다중불포화지방산은 C18:2 계열이 전체 지방산 함량의 15%
이상으로 측정되었다. 이러한 결과를 통하여 Arthrospira 속 남조류 2주는 사료 및
식품 등의 용도로 유용성을 지니는 것을 확인할 수 있었다. 다음으로 선형 사상체
구조를 지닌 Leptolyngbya 속 1주(NIBRIUM-3) 및 Lyngbya 속 1주(NIBRIUM-5)
는 상기 Arthrospira 속 2주와 상이한 양상을 나타내었다. 먼저 Leptolyngbya sp.
NIBRIUM-3의 경우 포화지방산인 C16:0 및 C18:0 함량이 26.6%로 상대적으로
낮았으며, 이에 반하여 단일불포화지방산인 C16:1 및 C18:1 함량이 전체 지방산의
약 40%를 차지하였다. Lyngbya sp. NIBRIUM-3의 경우, 포화지방산인 C16:0과
다중불포화지방산인 C18:3 계열이 전체 지방산의 약 46% 및 45%를 차지하는 반면,
단일불포화지방산은 매우 낮은 비율로 검출되었다. 이러한 결과를 통하여
Leptolyngbya 속 1주(NIBRIUM-3) 및 Lyngbya 속 1주(NIBRIUM-5)는 높은 불포화
지방산 함량으로 식품을 비롯한 바이오디젤 생산 등에 있어 유용성을 지니는 것을
확인하였다. 마지막으로 구형 군체를 형성하는 Myxosarcina sp. NIBRIUM-4는
C16:0이 대부분을 차지하는 포화지방산과 C16:1 계열이 대부분을 차지하는 단일
불포화지방산만 검출되었으며, 다중불포화지방산은 확인되지 않았다. 이러한 지방산
조성은 기존에 알려진 Section II Myxosarcina 속의 지방산 함량과 매우 유사한
것으로, 추후 지방산 조성이 Section II 남조류의 속 구분을 위한 마커로 사용될 수
있는 가능성을 내포한다.
그림 III-26. 자생 남조류 5주의 포화지방산(SFAs), 단일불포화지방산
(MUFAs) 및 다중불포화지방산(PUFAs)의 함량 비교결과.

- 71 -
(2) 녹조류 유래 지방산 분석 및 평가
연구방법에 제시된 바와 같이 동정이 완료된 5주의 녹조류 배양체 유래 지방산
조성은 GC-mass spectrometry detector(GC-MSD)를 이용하여 분석하였다. 5L급
폭기 배양조에서 수확된 녹조류 바이오매스 1g씩을 분석에 사용하였으며, 분석
결과는 그림 III-27 및 그림 III-28에 제시되었다.
그림 III-27. 자생 녹조류 5주의 세부 지방산 조성(%) 분석결과.
분석결과 녹조류의 경우 남조류와 달리 포화지방산의 함량이 상대적으로
낮은(30% 미만) 것으로 분석되었다. 또한 Chlamydomonas sp. NIBRIUM-6

- 72 -
녹조류주를 제외하고 모두 다중불포화지방산의 함량이 50%를 초과하였으며,
Haematococcus sp. NIBRIUM-7의 경우 다중불포화지방산 함량이 80%에
육박하는 것으로 분석되었다. 녹조류 유래 포화지방산과 다중불포화지방산은
영양학적 가치가 매우 높은 것으로 알려져 있으며, 따라서 금번 확보된 녹조류
5주의 경우 사료 및 식품으로써의 유용성이 있는 것으로 추측된다. 추후 세포
독성과 관련된 분석을 통하여 사료 및 식품으로써의 활용성을 평가할 필요가
제기된다. Chlamydomonas sp. NIBRIUM-6의 경우 C18:1 계열의 단일불포화
지방산의 함량이 타 녹조류주에 비하여 매우 높았으며, 이는 추후 녹조류
유래 바이오디젤 생산 시 다중불포화지방산이 많은 타 녹조류주에 비해 점도
및 융점이 상대적으로 낮아 바이오디젤로써의 효용성을 극대화시킬 수 있을
것으로 기대된다.
그림 III-28. 자생 녹조류 5주의 포화지방산(SFAs), 단일불포화지방산
(MUFAs) 및 다중불포화지방산(PUFAs)의 함량 비교결과.

- 73 -
남조류주당성분 함량(%)
Fuc Rha Ara Gal Glc Man Xyl
NIBRIUM-1 2.0 5.5 0.2 9.1 80.3 0.5 4.5
NIBRIUM-2 3.1 5.4 0.3 8.9 79.4 0.5 4.4
NIBRIUM-3 5.3 2.3 0.2 8.9 75.9 5.8 1.7
NIBRIUM-4 2.0 1.4 10.1 7.0 46.9 16.6 16.0
NIBRIUM-5 4.9 2.2 0.3 9.0 75.9 5.5 2.2
다) 미세조류 유래 당류 분석 및 평가
(1) 남조류 유래 당류 분석 및 평가
연구방법에 제시된 바와 같이 동정이 완료된 5주의 남조류 배양체 유래 당류
조성 분석은 high-performance anion-exchange chromatography coupled with
a pulsed amperometric detection system(HPAEC-PAD; Dionex Corp.)을 이용
하여 분석하였다. 5L급 폭기 배양조에서 수확된 남조류 바이오매스 1g씩을 분석에
사용하였으며, 분석결과는 표 III-5에 제시되었다.
표 III-5. 자생 남조류 당분석 결과
(Fuc: Fucose; Rha: Rhamnose; Ara: Arabinose; Gal: Galactose; Glu: Glucose; Man:
Mannose; Xyl: Xylose)
분석결과 사상체형의 남조류의 당 성분은 대부분 Glucose로 구성되어 있는
것으로 확인되었으며, Arthrospira 속은 Leptolyngbya 및 Lyngbya 속과 달리
Rhamnose와 Xylose의 함량이 상대적으로 높았으며 Mannose의 함량은 상대적
으로 낮았다. 이에 반하여 구형 군체를 형성하는 Myxosarcina sp.
NIBRIUM-4의 경우, glucose의 함량이 50% 미만인 반면 Arabinose와
Mannose 및 Xylose의 함량이 타 사상체형 남조류에 비하여 높은 경향을
나타내었다.
(2) 녹조류 유래 당류 분석 및 평가
연구방법에 제시된 바와 같이 동정이 완료된 5주의 녹조류 배양체 유래 당류
조성 분석은 high-performance anion-exchange chromatography coupled with

- 74 -
녹조류주당성분 함량(%)
Fuc Rha Ara Gal Glc Man Xyl
NIBRIUM-6 2.0 11.5 11.7 21.4 49.7 2.9 0.7
NIBRIUM-7 3.2 1.7 1.1 3.9 46.0 40.9 3.2
NIBRIUM-8 3.3 7.1 0.8 32.2 25.1 19.4 12.1
NIBRIUM-9 3.0 8.2 1.0 31.1 25.4 14.4 16.9
NIBRIUM-10 3.1 7.7 1.2 30.9 25.6 20.3 11.2
a pulsed amperometric detection system(HPAEC-PAD; Dionex Corp.)을 이용
하여 분석하였다. 5L급 폭기 배양조에서 수확된 녹조류 바이오매스 1g씩을 분석에
사용하였으며, 분석결과는 표 III-9에 제시되었다.
표 III-6. 자생 녹조류 당분석 결과
(Fuc: Fucose; Rha: Rhamnose; Ara: Arabinose; Gal: Galactose; Glu: Glucose; Man:
Mannose; Xyl: Xylose)
분석결과 녹조류주는 당 분석을 위한 가수분해 과정에서 분해가 완전히
되지 않는 문제점을 노출하였다. 현재 식물 유래 당 분석은 대부분 산 및
효소 가수분해 방법을 이용하나, 해조류 및 녹조류의 경우 세포벽의 구조가
일반 식물과 상이한 관계로 가수분해가 완벽히 되지 않는다. 따라서 본 결과
는 100% 가수분해가 진행된 당 분석 결과가 아닌, 일정량 이상 가수분해 된
결과만 제시되었다. 녹조류 당 성분 분석을 위한 가수분해법 연구는 추후
반드시 진행되어야 할 부분이다.

- 75 -
특성 세부특성조건/가중치
NIBRIUM-1 NIBRIUM-2 NIBRIUM-3 NIBRIUM-4 NIBRIUM-5
배양
특성
배양용이성 3 3 3 1 3
수확용이성 2 2 2 1 2
신규성 균주 신규성 1 1 3 3 3
일반성분
특성
조단백 함량 1 1 1 0 1
조지방 함량 0 0 0 0 0
조탄수화물
함량0 1 1 1 0
세부성분
특성
유용 지방산 1 1 1 1 1
필수
아미노산1 1 1 0 1
유용 단당 1 1 1 0 1
합계 10 11 12 7 12
3. 유용 미세조류 선별 및 배양기법 탐색
가. 연구결과
1) 유용성 검증결과 기반 유용 미세조류주 선별
가) 유용 남조류주 평가 및 선정
연구 방법에 제시된 평가 방식에 따라서 i)배양특성, ii)신규성, iii)일반
성분 특성, iv)세부성분 특성 등의 평가항목으로 확보된 자생 남조류주의 활용도를
측정하였다(표 III-7).
표 III-7. 유용성 검증결과 기반 자생 남조류주 선정 결과
먼저 Arthrospira sp. NIBRIUM-1 및 2 남조류주의 경우, 배양용이성과
수확용이성에 있어서 장점을 지니고 있었으며, 일반성분(조단백, 조지방 및 조
탄수화물 함량) 및 세부성분(아미노산, 지방산 및 단당) 등의 성분분석 결과

- 76 -
역시 충분한 활용성을 갖는 것으로 확인되었다. 그러나 균주의 신규성에 있어
국내·외 활용 사례가 다수 보고되어 있는 관계로 산업재산권 확보 등의 산업화를
위한 기반자료 확보 시 제한이 있을 것으로 파악되어 타 남조류주에 비하여
상대적으로 낮은 가중치를 책정하였다.
다음으로 Myxosarcina sp. NIBRIUM-4의 경우, 군체를 형성하기는 하지만
수확이 용이하지 않으며, 또한 해수의 염도 조건에서 배양이 되는 관계로
타 남조류주에 비하여 상대적으로 낮은 가중치가 책정되었다. 또한 배양과
관련하여 증식률이 낮은 관계로 바이오매스의 대량생산이 난해하여, 추후 배양
법에 대한 추가 연구가 필요한 것으로 추정된다. 일반성분(조단백, 조지방 및
조탄수화물 함량) 및 세부성분(아미노산, 지방산 및 단당) 등의 성분분석 결과
역시 군체를 형성하는 관계로 다당류가 상대적으로 높은 비율로 측정되어 다른
유용성분의 함량은 타 남조류주에 비해 낮은 것으로 파악되었다. 그러나 균주의
신규성 측면에 있어서 아직까지 해당 속의 남조류의 국·내외 활용 보고가 없어
추후 산업재산권 확보 등의 산업화를 위한 기반자료 확보가 용이할 것으로 파악
되어 타 남조류주에 비하여 상대적으로 높은 가중치를 책정하였다.
마지막으로 선형 사상체형 남조류인 Leptolyngbya sp. NIBRIUM-3 및
Lyngbya sp. NIBRIUM-5의 경우, 담수, 기수 및 해수 조건에서 모두 배양이
가능한 것으로 파악되었으며 플랑크톤 네트로 수확이 가능한 특성을 통해,
배양특성 부분에 있어 높은 가중치를 책정하였다. 또한 일반성분(조단백, 조지방
및 조탄수화물 함량) 및 세부성분(아미노산, 지방산 및 단당) 등의 성분분석
결과, 타 남조류주보다 상대적으로 높은 조단백 함량 혹은 조탄수화물 함량을
나타내어 식품 및 사료로써 활용성이 매우 높을 것으로 추정되어, 성분특성
역시 높은 가중치를 책정하였다. 마지막으로 균주의 신규성에 있어서 아직까지
해당 2 속의 남조류의 국·내외 활용 보고가 없어 추후 산업재산권 확보 등의
산업화를 위한 기반자료 확보가 용이할 것으로 파악되어 타 남조류주에 비해
상대적으로 높은 가중치를 책정하였다.
상기와 같은 유용성 검증결과를 기반으로 5종의 남조류주의 잠재적인
활용도를 평가하였으며, 평가결과 Leptolyngbya sp. NIBRIUM-3(가중치 12)=
Lyngbya sp. NIBRIUM-5(가중치 12) > Arthrospira sp. NIBRIUM-2(가중치
11) > Arthrospira sp. NIBRIUM-1(가중치 10) > Myxosarcina sp.
NIBRIUM-4(가중치 7) 순서로 나열되었다. 이와 같은 결과를 토대로 상위 2개의
남조류주인 Leptolyngbya sp. NIBRIUM-3 및 Lyngbya sp. NIBRIUM-5를 최
종적으로 선정하였으며, 선정된 2종의 남조류주는 추후 최적 배양조건 탐색을
위한 연구가 진행되었다.

- 77 -
특성 세부특성조건/가중치
NIBRIUM-6 NIBRIUM-7 NIBRIUM-8 NIBRIUM-9 NIBRIUM-10
배양
특성
배양용이성 2 2 3 2 2
수확용이성 1 1 1 1 1
신규성 균주 신규성 1 2 3 1 1
일반성분
특성
조단백 함량 1 0 0 0 0
조지방 함량 1 1 1 1 1
조탄수화물
함량0 1 1 1 1
세부성분
특성
유용 지방산 1 1 1 1 1
필수
아미노산1 1 1 1 1
유용 단당 1 1 1 1 1
합계 9 10 12 9 9
나) 유용 녹조류주 평가 및 선정
연구 방법에 제시된 평가 방식에 따라서 i)배양특성, ii)신규성, iii)일반
성분 특성, iv)세부성분 특성 등의 평가항목으로 확보된 자생 남조류주의 활용도를
측정하였다(표 III-8).
표 III-8. 유용성 검증결과 기반 자생 녹조류주 선정 결과
먼저 Chlamydomonas sp. NIBRIUM-6의 경우, 담수에서만 배양이 가능
하였으며, 수확 시 원심분리를 통하여서만 바이오매스 확보가 가능하였다. 일반
성분(조단백, 조지방 및 조탄수화물 함량) 및 세부성분(아미노산, 지방산 및
단당) 등의 성분분석 결과, 조단백 및 조지방 함량이 높아 가중치를 부여
하였으며 단일 불포화지방산, 필수 아미노산 및 유용 단당의 함량이 높아
가중치가 부여되었다. 그러나 균주의 신규성 측면에 있어 Chlamydomonas 속
녹조류는 전체 유전체가 분석되어 분자생물학적으로 깊이 연구되고 있으며,
응용생물학적으로도 국내·외 활용 사례가 다수 보고되어 있는 관계로 산업

- 78 -
재산권 확보 등의 산업화를 위한 기반자료 확보 시 제한이 있을 것으로 파악
되어 타 녹조류주에 비하여 상대적으로 낮은 가중치를 책정하였다.
다음으로 Haematococcus sp. NIBRIUM-7의 경우, 담수에서만 배양이
가능하였으며, 수확 시 원심분리를 통하여서만 바이오매스 확보가 가능하였다.
일반성분(조단백, 조지방 및 조탄수화물 함량) 및 세부성분(아미노산, 지방산
및 단당) 등의 성분분석 결과, 조지방 및 조탄수화물 함량을 비롯하여 다중
불포화지방산, 필수 아미노산 및 유용 단당 함량 등의 세부 성분 역시 상대적
으로 높아 가중치를 책정하였다. 균주의 신규성 측면에 있어 Haematococcus
속 녹조류는 Chlamydomonas 속 녹조류와 마찬가지로 유전체가 분석되어
분자생물학적으로 깊이 연구되고 있으며, 응용생물학적으로도 국내·외 활용
사례가 다수 보고되어 있는 관계로 산업재산권 확보 등의 산업화를 위한 기반
자료 확보 시 제한이 있을 것으로 파악되었으나 아직까지 국내 자생종을 확보한
보고가 없어 해당 녹조류주에 가중치를 책정하였다.
Lobosphaera sp. NIBRIUM-8의 경우, 담수, 기수 및 해수에서 모두 배양이
가능하였으며, 수확 시 원심분리를 통하여서만 바이오매스 확보가 가능하였다.
일반성분(조단백, 조지방 및 조탄수화물 함량) 및 세부성분(아미노산, 지방산
및 단당) 등의 성분분석 결과, 조지방 및 조탄수화물 함량을 비롯하여 단일
불포화지방산, 필수 아미노산 및 유용 단당 함량 등의 세부 성분 역시 상대적
으로 높아 가중치를 책정하였다. 균주의 신규성 측면에 있어 Lobosphaera 속
녹조류는 아직까지 해당 속의 국·내외 활용 보고가 없어 추후 산업재산권
확보 등의 산업화를 위한 기반자료 확보가 용이할 것으로 파악되어 타 녹조류
주에 비하여 상대적으로 높은 가중치를 책정하였다.
Scenedesmus sp. NIBRIUM-9의 경우, 담수에서만 배양이 가능하였으며,
수확 시 원심분리를 통하여서만 바이오매스 확보가 가능하였다. 일반성분
(조단백, 조지방 및 조탄수화물 함량) 및 세부성분(아미노산, 지방산 및 단당)
등의 성분분석 결과, 조단백 함량을 제외한 조지방, 조탄수화물 및 지방산,
아미노산, 단당류 등의 함량이 상대적으로 모두 높은 것으로 파악되어 가중치를
책정하였다. 균주의 신규성 측면에 있어 Scenedesmus 속 녹조류는 유전체가
분석되어 분자생물학적으로 깊이 연구되고 있으며, 응용생물학적으로도 국내·
외 활용 사례가 다수 보고되어 있는 관계로 산업재산권 확보 등의 산업화를
위한 기반자료 확보 시 제한이 있을 것으로 파악되어 타 녹조류주에 비하여
상대적으로 낮은 가중치를 책정하였다.
Chlorella sp. NIBRIUM-10의 경우, 담수에서만 배양이 가능하였으며,
수확 시 원심분리를 통하여서만 바이오매스 확보가 가능하였다. 일반성분

- 79 -
(조단백, 조지방 및 조탄수화물 함량) 및 세부성분(아미노산, 지방산 및 단당)
등의 성분분석 결과, Scenedesmus sp. NIBRIUM-9의 경우와 유사하게 조단백
함량을 제외한 조지방, 조탄수화물 및 지방산, 아미노산, 단당류 등의 함량이
상대적으로 모두 높은 것으로 파악되어 가중치를 책정하였다. Chlorella 속
녹조류는 유전체가 분석되어 분자생물학적으로 깊이 연구되고 있으며, 응용
생물학적으로도 식품 및 수산용 먹이생물로써 국내·외 활용 사례가 다수 보고
되어 있는 관계로 산업재산권 확보 등의 산업화를 위한 기반자료 확보 시
제한이 있을 것으로 파악되어 타 녹조류주에 비하여 상대적으로 낮은 가중치를
책정하였다.
상기와 같은 유용성 검증결과를 기반으로 5종의 녹조류주의 잠재적인
활용도를 평가하였으며, 평가결과 Lobosphaera sp. NIBRIUM-8(가중치 12) >
Haematococcus sp. NIBRIUM-7(가중치 10) > Chlamydomonas sp.
NIBRIUM-6(가중치 9) = Scenedesmus sp. NIBRIUM-9(가중치 9) =
Chlorella sp. NIBRIUM-10(가중치 9) 순서로 나열되었다. 이와 같은 결과를
토대로 상위 1개의 녹조류주인 Lobosphaera sp. NIBRIUM-8을 최종적으로
선정하였으며, 선정된 1종의 녹조류주는 추후 최적 배양조건 탐색을 위한
연구가 진행되었다.

- 80 -
2) 성장률 기반 최적 배양조건 탐색
가) 선정된 유용 남조류주 2주의 최적 배양조건 탐색
(1) Leptolyngbya sp. NIBRIUM-3 최적 배양조건 탐색
유용성 검증결과를 기반으로 선정된 사상체형 남조류 Leptolyngbya sp.
NIBRIUM-3의 최적 배양조건 탐색을 위하여 순수 배양체를 다양한 염분, pH
및 온도 조건의 Zarrouk's 영양배지에 접종하여 100.0 μmol/m2/s, 12L:12D 명암
주기 하에서 28일 동안 폭기 방식의 증균 배양을 지속하였다. 실험에 사용된
염분의 농도는 0-100psu(0, 10, 20, 30, 50, 70, 90, 100psu), pH는 2-13(1씩 증감),
수온은 4-35°C(4, 15, 25, 30, 35°C)로 21일간 배양하였다. 각 환경조건별 증식률은
10mL의 배양액을 2일 또는 4일 간격으로 GF/C 필터(Watmann)로 여과한 후
105°C에서 24시간 건조한 후 그 질량의 변화를 측정하여 성장 여부를 결정
하였다. 최종 결과는 배양 시작 후 28일이 되는 시점에서 일괄적으로 측정
되었으며 그 결과는 그림 III-29에 제시되었다.
사상체 형태의 남조류는 일반적으로 다양한 극한 환경조건에서 생존이
가능한 것으로 알려져 있으며, 본 연구에서 선정된 Leptolyngbya sp.
NIBRIUM-3 역시 일반적인 해수의 염도인 30-35psu를 뛰어넘어 염도조건
100psu까지 배양이 가능한 것으로 파악되었다. 그러나 바이오매스 생산량을
중심으로 적정 염도범위를 선정한 결과 대량 배양을 위한 적정 염도조건은
0-30psu로 확인되었다. 또한 pH의 변화에도 본 남조류주는 2-13까지 배양이
가능한 것으로 파악되었으나 바이오매스 생산량을 중심으로 적정 염도범위를
선정한 결과 대량 배양을 위한 적정 pH 조건은 pH 5-9로 확인되었다. 마지막
으로 온도 변화에 따른 바이오매스 생산량의 변화를 측정한 결과 4-35°C까지
배양이 가능한 것으로 측정되었으나, 바이오매스 생산량을 중심으로 적정
온도범위를 선정한 결과 대량 배양을 위한 적정 온도조건은 25-35°C로 확인
되었다. 상기와 같은 결과를 토대로 Leptolyngbya sp. NIBRIUM-3 바이오
매스 대량생산을 위한 적정 환경조건은 pH 5-9, 염도 0-30psu 및 온도
25-35°C로 확인되어 추후 관련 연구는 이와 같은 배양조건에서 진행될 것이다.

- 81 -
그림 III-29. Leptolyngbya sp. NIBRIUM-3의 최적 배양조건 결과. a)온도
조건별, b)염도 조건별, c)pH 조건별 남조류주의 바이오매스 생산량 변화.

- 82 -
(2) Lyngbya sp. NIBRIUM-5 최적 배양조건 탐색
유용성 검증결과를 기반으로 선정된 사상체형 남조류 Lyngbya sp.
NIBRIUM-5의 최적 배양조건 탐색을 위하여 순수 배양체를 다양한 염분, pH
및 온도 조건의 Zarrouk's 영양배지에 접종하여 100.0 μmol/m2/s, 12L:12D 명암
주기 하에서 28일 동안 폭기 방식의 증균 배양을 지속하였다. 실험에 사용된
염분의 농도는 0-100psu(0, 10, 20, 30, 50, 70, 90, 100psu), pH는 2-13(1씩 증
감), 수온은 4-35°C(4, 15, 25, 30, 35°C)로 21일간 배양하였다. 각 환경조건별
증식률은 10mL의 배양액을 2일 또는 4일 간격으로 GF/C 필터(Watmann)로
여과한 후 105°C에서 24시간 건조한 후 그 질량의 변화를 측정하여 성장 여부를
결정하였다. 최종 결과는 배양 시작 후 28일이 되는 시점에서 일괄적으로
측정되었으며 그 결과는 그림 III-30에 제시되었다.
사상체 형태의 남조류는 일반적으로 다양한 극한 환경조건에서 생존이
가능한 것으로 알려져 있으나, Lyngbya sp. NIBRIUM-5 남조류주는
Leptolyngbya sp. NIBRIUM-3에 비하여 환경 적응도가 낮은 것으로 분석
되었다. 본 연구에서 선정된 Lyngbya sp. NIBRIUM-5 남조류주는 일반적인
해수의 염도인 30-35psu를 뛰어넘어 염도조건 100psu까지 생존 및 증식이
가능한 것으로 파악되었으나, 바이오매스 생산량을 중심으로 적정 염도범위를
선정한 결과 대량 배양을 위한 적정 염도조건은 0-30psu로 확인되었다. 또한
pH의 변화에도 본 남조류주는 2-13까지 배양이 가능한 것으로 파악되었으나
바이오매스 생산량을 중심으로 적정 염도범위를 선정한 결과 대량 배양을
위한 적정 pH 조건은 pH 5-9로 확인되었다. 마지막으로 온도 변화에 따른 바이오
매스 생산량의 변화를 측정한 결과 4-35°C까지 배양이 가능한 것으로 측정
되었으나, 바이오매스 생산량을 중심으로 적정 온도범위를 선정한 결과 대량
배양을 위한 적정 온도조건은 25-35°C로 확인되었다. 상기와 같은 결과를
토대로 Lyngbya sp. NIBRIUM-5 바이오매스 대량생산을 위한 적정 환경조건은
Leptolyngbya sp. NIBRIUM-3과 동일하게 pH 5-9, 염도 0-30psu 및 온도
25-35°C로 확인되어 추후 관련 연구는 이와 같은 배양조건에서 진행될 것이다.
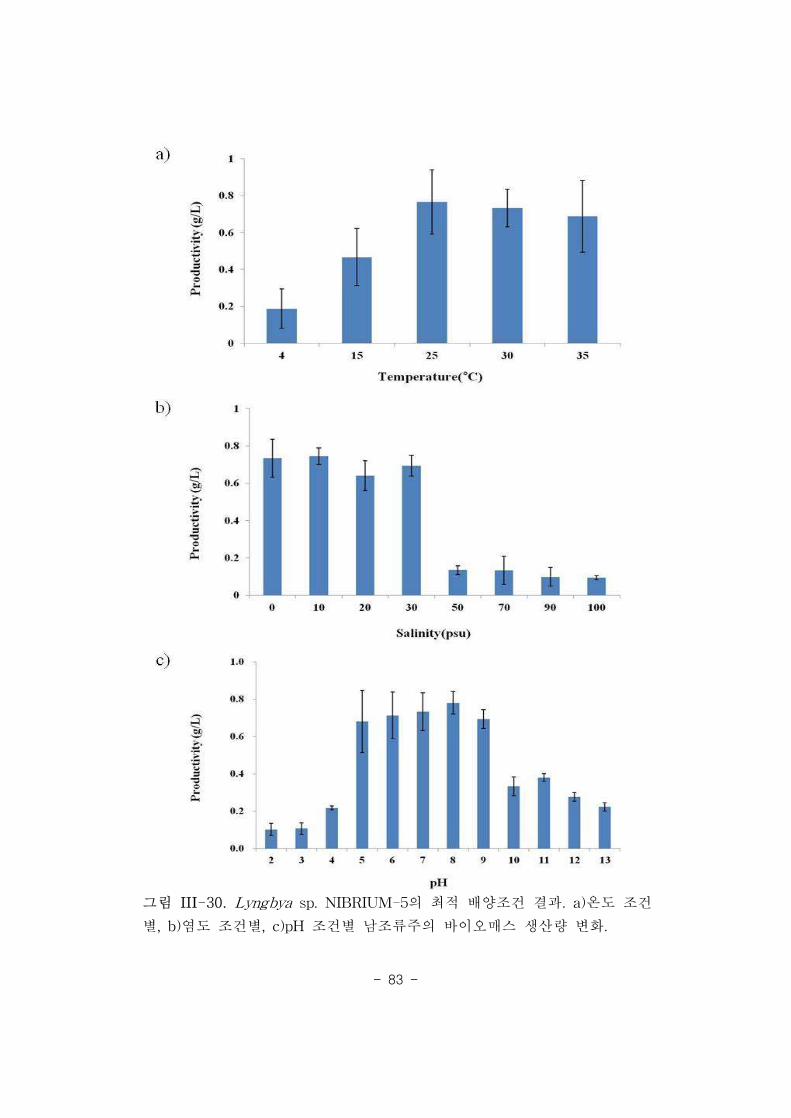
- 83 -
그림 III-30. Lyngbya sp. NIBRIUM-5의 최적 배양조건 결과. a)온도 조건
별, b)염도 조건별, c)pH 조건별 남조류주의 바이오매스 생산량 변화.

- 84 -
나) 선정된 유용 녹조류주 1주의 최적 배양조건 탐색
(1) Lobosphaera sp. NIBRIUM-8 최적 배양조건 탐색
유용성 검증결과를 기반으로 선정된 구형 녹조류 Lobosphaera sp.
NIBRIUM-8의 최적 배양조건 탐색을 위하여 순수 배양체를 다양한 염분, pH
및 온도 조건의 BG-11 영양배지에 접종하여 100.0 μmol/m2/s, 12L:12D 명암
주기 하에서 28일 동안 폭기 방식의 증균 배양을 지속하였다. 실험에 사용된
염분의 농도는 0-100psu(0, 10, 20, 30, 50, 70, 90, 100psu), pH는 2-13(1씩 증
감), 수온은 4-35°C(4, 15, 25, 30, 35°C)로 21일간 배양하였다. 본 녹조류의
증식률은 SR chamber를 사용하여 직접 세포 수 계량법으로 측정을 진행하려고
하였으나 배양 조건 내에서 주로 무성생식을 통한 딸세포 분열을 하는 것으로
관찰되어, 각 환경조건별 증식률은 10mL의 배양액을 2일 또는 4일 간격으로
GF/C 필터(Watmann)로 여과한 후 105°C에서 24시간 건조한 후 그 질량의
변화를 측정하여 성장 여부를 결정하였다. 최종 결과는 배양 시작 후 28일이
되는 시점에서 일괄적으로 측정되었으며 그 결과는 그림 III-31에 제시되었다.
구형의 녹조류는 일반적으로 남조류에 비하여 환경의 변화에 민감한 것으로
알려져 있으며, 본 연구에서 선정된 Lobosphaera sp. NIBRIUM-8 녹조류주
역시 상기 2주의 남조류에 비하여 환경 적응도가 낮은 것으로 확인되었다.
본 연구에서 Lobosphaera sp. NIBRIUM-8 녹조류주는 일반적인 해수의 염도인
30-35psu까지 생존 및 증식이 가능한 것으로 파악되었으며, 이를 초과하는
염도조건에서는 증식이 거의 이루어지지 않은 것으로 파악되었다. 또한 염도
조건 70psu를 초과하는 환경에서는 녹조류 세포가 사멸하였다. 따라서 바이오
매스 생산량을 중심으로 적정 염도범위를 선정한 결과 대량 배양을 위한 적정
염도조건은 0-20psu로 확인되었다. 또한 배양조건의 pH의 변화에 있어 본
녹조류주는 5-10까지 배양이 가능한 것으로 파악되었으나 바이오매스 생산량을
중심으로 적정 염도범위를 선정한 결과 대량 배양을 위한 적정 pH 조건은
pH 6-8로 확인되었다. 마지막으로 온도 변화에 따른 바이오매스 생산량의 변화를
측정한 결과 4-35°C까지 배양이 가능한 것으로 측정되었으나, 바이오매스
생산량을 중심으로 적정 온도범위를 선정한 결과 대량 배양을 위한 적정 온도
조건은 25-30°C로 확인되었다. 상기와 같은 결과를 토대로 Lobosphaera sp.
NIBRIUM-8 바이오매스 대량생산을 위한 적정 환경조건은 pH 6-8, 염도
0-20psu 및 온도 25-30°C로 확인되어 추후 관련 연구는 이와 같은 배양조건
에서 진행될 것이다.

- 85 -
그림 III-31. Lobosphaera sp. NIBRIUM-8의 최적 배양조건 결과. a)온도
조건별, b)염도 조건별, c)pH 조건별 남조류주의 바이오매스 생산량 변화.


- 87 -
IV. 고 찰


- 89 -
IV. 고찰
최근 급속한 산업화로 인한 대기 중 온실가스 농도 증가에 의해 초래되고
있는 지구 온난화로 다양한 해결방안이 요구되고 있다. 이러한 대기 중 온실
가스 농도 증가는 대부분 이산화탄소의 증가에 기인한 것으로 알려져 있으며,
이산화탄소를 포집할 수 있는 광합성 식물을 이용한 이산화탄소 흡수가 그
대안으로 제기되고 있다. 그러나 식물을 이용한 이산화탄소 흡수 방안은 현실적
으로 폭발적인 전 세계 인구증가와 맞물려 밀림을 비롯한 녹지가 파괴되어
가는 상황에서 그 실효성에 대한 의문이 제기되고 있다. 이러한 상황에 있어
최근 미세조류를 이용한 이산화탄소 저감 방안이 대안으로 제시되고 있다.
미세조류는 남조류와 녹조류 등을 포함하는 광합성 식물 플랑크톤으로 이산화
탄소 저감, 바이오에너지 생산 및 천연 기능성 물질 생산 등이 가능한 생물
플랫폼(Platform)으로써 그 연구가치 및 효용성을 인정받고 있다. 실제
Chlorella 및 Spirulina(Arthrospira) 속 등의 미세조류는 효율적인 단백질 및
유용물질을 포함한 바이오매스 공급원으로써 그 중요성을 인정받고 있으며,
Dunaliella 및 Botryococcus 속 등의 미세조류는 바이오디젤 생산을 위한
지질 공급원으로써 대량배양 및 지질 추출과 관련된 심화연구가 전 세계적으로
진행되고 있는 실정이다. 그러나 이러한 미세조류 관련 연구는 대부분 국제적
으로 특허권이 이미 확보되어 있는 미세조류주를 이용하여 진행되고 있으며,
따라서 이러한 특허 미세조류를 국내에서 산업적으로 적용하는데 있어 문제점을
노출하고 있다. 또한 현재 전 세계적으로 상업적인 미세조류 배양은 몇몇
국제 기업에 의해서 독점되고 있으며, 국내에서 이러한 국제 기업을 따라잡기
위해서는 국제적 특허분쟁의 소지가 없는 자생 유용 미세조류를 이용한 바이오
매스 생산 기술 개발이 시급한 실정이다. 이러한 이유로 대한민국 자생 미세
조류를 발굴하여 유용성 및 배양능을 분석하는 연구를 통하여 추후 미세조류와
관련된 국가 경쟁력을 확보를 도모하고자 “이산화탄소 생물전환을 위한 자생
유용 미세조류 발굴_1차년도” 사업을 추진하였다. 당해년도 연구 사업은 크게
①배양 가능한 자생 미세조류 확보 ②미세조류 배양체의 유용성 탐색 ③유용
미세조류의 최적 배양조건을 탐색을 그 목표로 한다. 본 연구를 통하여 수입
미세조류 종을 대체할 수 있는 자생 미세조류 종을 발굴하여 국내 미세조류
관련 산업의 기반을 확충하며, 미세조류와 관련된 국가 생물 다양성 관련
연구에 기반자료를 제공하고자 하였다.

- 90 -
먼저 자생 미세조류 확보와 관련하여 다양한 국내 수생 서식환경(수생생물
배양장, 저수지, 강, 하구둑, 갯벌, 조간대 지역 등)에서 부유성 플랑크톤을
중심으로 채집하였으며, 다양한 영양배지(BBM, BG-11 medium, Zarrouk's
medium, f/2 medium 등)에서 배양이 가능한 종을 우선적으로 선정하였다.
현재 미세조류 배양과 관련하여 다양한 증식용 영양배지가 개발되고 있으나,
실제 배양이 가능한 종은 아직까지 1%에 미치지 못하는 것으로 알려져 있다.
현재까지 미세조류 증식과 관련하여 탄소원(Carbon source), 질소원(Nitrogen
source) 및 인원(Phosphate source) 등의 주요영양인자 외 금속원(trace metal
source)과 비타민원(Vitamin source)등의 미량 인자들이 관련되어 있는 것으로
알려져 있으나, 각 분류군 및 종별로 영양배지에 대한 감수성이 상이한 것으로
보고되고 있다. 따라서 자생종을 발굴하여 배양법을 확립하는 것보다는 기존의
영양배지에 배양이 가능한 종을 선정하는 것이 현 시점에서 자생 유용 미세조류를
이용한 산업재산권 확보와 관련되어 좀 더 실질적인 성과를 낼 수 있을 것으로
추정되어 연구가 진행되었다. 다양한 수생환경에서 50주 이상의 미세조류를
모세관법 및 직접도말법 등을 통하여 분리하였고, 이 중 배양이 용이한 미세
조류주를 선정한 결과 남조류 5주(NIBRIUM-1, 2, 3, 4, 5)와 녹조류 5주
(NIBRIUM-6, 7, 8, 9, 10)의 총 10주의 배양이 용이한 균주가 선정되었다.
남조류주 5주의 경우, 형태학적 및 분자생물학적 동정을 통하여 Arthrospira
속 2주(NIBRIUM-1, 2), Leptolyngbya 속 1주(NIBRIUM-3), Myxosarcina 속 1
주(NIBRIUM-4), Lyngbya 속 1주(NIBRIUM-5)로 판명이 되었다.
Arthrospira 속의 경우, 현재까지 가장 많이 연구된 남조류 속으로써 높은
조단백 함량 및 빠른 생장속도로 전 세계적으로 ‘슈퍼푸드’로 각광받고 있다.
남아메리카와 아프리카 지역의 알칼리성 호수 지역에서 발견되는 Arthrospira
속은 이전부터 원주민의 식량 자원으로써 간주되어 왔으나, 우리나라의 경우
문헌에서 발견되지 않는 것으로 보아 식용으로 사용되지는 않은 것으로 파악
된다. 우리나라의 경우 Arthrospira 속 남조류 관련 연구는 대부분 국내 자생
주가 아닌 외래종으로 진행되고 있으며, 국내 자생주의 보고는 미미한 실정이
었다. Arthrospira 속 남조류는 일반적으로 기수 서식을 하는 것으로 알려져
있으며, 따라서 본 연구과제 수행 중 해당 속 남조류는 금강 하구둑 및 제주
서귀포 부근 기수 지역 등에서 관찰이 되었다. 타 지역의 기수 시료에서도
관찰이 되었으나 영양배지 내 증식이 원활하지 않아 2주의 Arthrospira 속을
확보하는데 그쳤다. 일반적으로 Arthrospira 속 남조류는 종 간 유전자 유사도가

- 91 -
매우 높으며, 현재 종 간 재분류가 필요한 것으로 보고되고 있다. 본 연구에서
확보된 Arthrospira 속 NIBRIUM-1 및 2 남조류주의 산업재산권 확보와 관련
하여 16S rRNA 염기서열 분석을 통하여 동정을 진행하였고, 분석 결과 역시
기존에 보고된 국내·외 Arthrospira 속 남조류주와 매우 유사한 것으로 판명
되었다. 따라서 추후 Arthrospira 속 NIBRIUM-1 및 2 남조류주의 산업
재산권을 확보하기 위해서는 포괄적인 전체 유전체 분석이 선행되어야 할 것이다.
유용성 분석을 위한 일반성분(조단백, 조지방 및 조탄수화물 함량) 및 세부
성분(아미노산, 지방산 및 단당) 등의 성분분석 결과 역시 기존에 보고된
국내·외 Arthrospira 속 남조류주와 매우 유사한 것으로 판명되었다. 그러나
50%가 넘어가는 조단백 함량 및 뛰어난 증식률은 아직까지 Arthrospira 속
NIBRIUM-1 및 2 남조류주의 강력한 경쟁력을 부여하고 있다. 그러나 균주의
신규성에 있어서 Arthrospira 속 남조류의 국내·외 활용 사례가 다수 보고
되어 있는 관계로 산업재산권 확보 등의 산업화를 위한 기반자료 확보 시
제한이 있을 것으로 파악되어 상기 언급한 바와 같이 추후 전체 유전체 분석
등을 통한 균주 특이성을 확립할 필요가 제기된다.
Myxosarcina 속의 경우, 군체를 형성하는 구형 남조류로 현재 Section II
로 분류되어 있으며, 우리나라의 경우 문헌에서 발견되지 않는 것으로 보아
미기록종으로 추정된다. Myxosarcina 속 남조류는 다양한 환경에 서식을
하는 것으로 알려져 있으며, 본 연구과제 수행 중 해당 속 남조류는 제주
구좌읍 부근 기수 지역에서 관찰이 되었으며, 타 지역 시료에서는 관찰되지
않았다. Myxosarcina 속 NIBRIUM-3은 일반적으로 알려진 이분법과 달리
특이하게 난할을 통하여 16~32개의 딸세포로 분열을 통해 혹은 Baeocyte
생성을 통해 증식하였으며, 증식영양배지 내 증식이 다른 남조류주에 비하여
매우 느렸다. 일반적으로 Myxosarcina 속 남조류는 Chroococcidiopsis 속 및
Dermocapella 속 간 유전자 유사도가 매우 높으며, 현재 종 간 재분류가 필요한
것으로 보고되고 있다. Myxosarcina 속은 현재 국내·외 활용성 연구가 거의
이루어지지 않았으며, 생태학적 연구 역시 거의 이루어지지 않은 상태이다.
따라서 본 연구에서 확보된 Myxosarcina 속 NIBRIUM-3 남조류주의 산업
재산권 확보와 관련하여 16S rRNA 염기서열 분석을 통하여 동정을 진행하였고,
분석 결과 역시 기존에 보고된 국내·외 Myxosarcina 속 남조류주와 상이한
것으로 판명되었다. 유용성 분석을 위한 일반성분(조단백, 조지방 및 조탄수화물
함량) 및 세부성분(아미노산, 지방산 및 단당) 등의 성분분석 결과 기존에 보고된
국외 Myxosarcina 속 남조류주와 매우 상이한 것으로 판명되었다. 그러나
난할 증식으로 인한 높은 다당류 함량 및 낮은 조단백 함량, 매우 낮은 증식률

- 92 -
등은 아직까지 Myxosarcina 속 NIBRIUM-3 남조류주가 해결해야 할 문제점
이지만 매우 높은 단일불포화지방산의 함량은 바이오에너지 생산을 위한 본
남조류주의 강력한 경쟁력을 부여하고 있다. 균주의 신규성 측면에 있어서
아직까지 해당 속의 남조류의 국내·외 활용 보고가 없어 추후 산업재산권
확보 등의 산업화를 위한 기반자료 확보가 용이할 것으로 추정된다. 더불어
추후 전체 유전체 분석 등을 통한 균주 특이성을 확립할 필요가 제기된다.
선형 사상체형 남조류인 Leptolyngbya 속 NIBRIUM-3 및 Lyngbya 속
NIBRIUM-5의 경우, 기존 우리나라 문헌에 기재되어 있는 것으로 파악된다.
상기 2종의 남조류주는 담수, 기수 및 해수 조건에서 모두 배양이 가능한 것으로
확인되었으며, 플랑크톤 네트로 수확이 가능한 특성을 통해 배양특성 부분에
있어 추후 활용성에 강점을 갖는다. 또한 바이오매스 생산량 역시 현재까지
가장 높은 바이오매스 생산량을 갖는 것으로 알려진 Arthrospira 속에 뒤지지
않으며, 뛰어난 환경 적응 능력(pH, 온도, 염도 등)을 갖는 것으로 확인되었다.
Leptolyngbya 및 Lyngbya 속은 현재 국내·외 활용성 연구가 거의 이루어지지
않았으며, 생태학적 연구 역시 거의 이루어지지 않은 상태이다. 따라서 본
연구에서 확보된 Leptolyngbya 속 NIBRIUM-3 및 Lyngbya 속 NIBRIUM-5
남조류주의 산업재산권 확보와 관련하여 16S rRNA 염기서열 분석을 통하여
동정을 진행하였고, 분석 결과 역시 기존에 보고된 국내·외 Leptolyngbya 및
Lyngbya 속 남조류주와 상이한 것으로 판명되었다. 유용성 분석을 위한 일반
성분(조단백, 조지방 및 조탄수화물 함량) 및 세부성분(아미노산, 지방산 및
단당) 등의 성분분석 결과 기존에 보고된 국내·외 Arthrospira 속 남조류주와
매우 유사하며, 조단백 함량 및 증식률은 Arthrospira 속을 오히려 능가하는
것으로 파악되어 해당 남조류주의 강력한 경쟁력을 부여하고 있다. 실제로
Lyngbya 속 NIBRIUM-5의 경우, 세포 내 조단백 함량이 70%를 상회하는 것으로
측정되었으며, 이는 지금까지 알려진 남조류의 조단백 함량 중 가장 높은
수치로 파악이 되고 있다. 이러한 특성은 아직까지 세포독성에 대한 연구가
이루어지지 않아 단정을 내릴 수는 없지만, 추후 기능성 식품 및 사료로써
강력한 경쟁력을 가질 것이다. 균주의 신규성 측면에 있어서 아직까지 해당
속의 남조류의 국내·외 활용 보고가 없어 추후 산업재산권 확보 등의 산업화를
위한 기반자료 확보가 용이할 것으로 추정된다. 더불어 추후 전체 유전체
분석 및 세포독성 연구 등을 통한 균주 특이성을 확립할 필요가 제기된다.
녹조류주 5주의 경우, 형태학적 및 분자생물학적 동정을 통하여

- 93 -
Chlamydomonas 속 1주(NIBRIUM-6), Haematococcus 속 1주(NIBRIUM-7),
Lobosphaera 속 1주(NIBRIUM-8), Scenedesmus 속 1주(NIBRIUM-9),
Chlorella 속 1주(NIBRIUM-10)로 판명이 되었다.
Chlamydomonas 속의 경우, 현재까지 가장 많이 연구된 녹조류 속으로써
분자생물학적 및 생태학적 연구가 깊이 진행되어 있다. 다양한 수계환경부터
극지의 빙산까지 발견되는 Chlamydomonas 속은 우리나라에서도 여러 건의
발견이 보고된다. 그러나 우리나라의 경우 Chlamydomonas 속 녹조류 관련
연구는 대부분 국내 자생주가 아닌 외래종으로 진행되고 있으며, 국내 자생주의
유용성 및 분자생물학적 분석 등의 보고는 미미한 실정이다. Chlamydomonas
속 녹조류는 일반적으로 담수 서식으로 알려져 있으며, 따라서 본 연구과제
수행 중 해당 속 녹조류는 유주자 형태로 경기도 안산을 포함한 다양한 담수
지역 등에서 관찰이 되었다. 일반적으로 Chlamydomonas 속 녹조류는 종 간
유전자 유사도가 매우 높으며, 현재 종 간 재분류가 필요한 것으로 보고되고
있다. 본 연구에서 확보된 Chlamydomonas 속 NIBRIUM-6 녹조류주의 산업
재산권 확보와 관련하여 18S rRNA 및 18S-28S 염기서열 분석을 통하여
동정을 진행하였고, 분석 결과 기존에 보고된 국내·외 Chlamydomonas 속
녹조류와 유사한 것으로 판명되었다. 추후 Chlamydomonas 속 NIBRIUM-6
녹조류주의 산업재산권을 확보하기 위해서는 포괄적인 전체 유전체 분석이
선행되어야 할 것이다. 유용성 분석을 위한 일반성분(조단백, 조지방 및 조탄수화물
함량) 및 세부성분(아미노산, 지방산 및 단당) 등의 성분분석 결과 기존에 보고
된 국내·외 Chlamydomonas 속 녹조류주와 상당히 유사한 것으로 판명되었다.
그러나 Chlamydomonas 속 NIBRIUM-6의 매우 높은 단일불포화지방산의
함량은 바이오에너지 생산을 위한 본 녹조류주의 강력한 경쟁력을 부여하고
있다. 그러나 균주의 신규성에 있어서 Chlamydomonas 속 녹조류의 국내·외
활용 사례가 다수 보고되어 있는 관계로 산업재산권 확보 등의 산업화를 위한
기반자료 확보 시 제한이 있을 것으로 파악되어 상기 언급한 바와 같이 추후
전체 유전체 분석 등을 통한 균주 특이성을 확립할 필요가 제기된다.
Haematococcus 속의 경우, 현재까지 가장 많이 연구된 녹조류 속 중
하나로써 분자생물학적 및 생리학적 연구가 깊이 진행되어 있다. 이는
Haematococcus가 생산하는 Astaxanthin이라는 색소물질에 기인하는데, 이 색소는
강력한 항산화능을 지닌 것으로 알려져 기능성 화장품 및 식품 등 다양한
활용도를 갖고 있다. Haematococcus 속 녹조류는 다양한 수계환경에서 발견
되는데 일반적으로 ‘조류(鳥類)의 목욕지‘에서 자주 분리되는 것으로 알려져

- 94 -
있으며, 아직까지 우리나라에는 보고된 기록이 없어 미기록종으로 추정된다.
Haematococcus 속 녹조류 관련 연구는 전세계적으로 활발히 이루어지고
있으며, 우리나라의 경우 Haematococcus 관련 연구는 전부 외래종으로 진행
되고 있어 국내 자생주의 유용성 및 분자생물학적 분석 등의 보고는 전무한
실정이다. Haematococcus 속 녹조류는 일반적으로 담수 서식으로 알려져
있으며, 따라서 본 연구과제 수행 중 해당 속 녹조류는 휴면포자 형태로 경기도
안산 담수 지역에서 관찰되었다. 현재 Haematococcus 속 녹조류는 종 간
유전자 유사도가 매우 높으며, 현재 종 간 재분류가 필요한 것으로 보고되고
있다. 본 연구에서 확보된 Haematococcus 속 NIBRIUM-7 녹조류주의 산업
재산권 확보와 관련하여 18S rRNA 및 rbcL 염기서열 분석을 통하여 동정을
진행하였고, 분석 결과 기존에 보고된 국외 Haematococcus 속 녹조류와 유사한
것으로 판명되었다. 추후 Haematococcus 속 NIBRIUM-7 녹조류주의 산업
재산권을 확보하기 위해서는 포괄적인 전체 유전체 분석이 선행되어야 할 것
이다. 유용성 분석을 위한 일반성분(조단백, 조지방 및 조탄수화물 함량) 및
세부성분(아미노산, 지방산 및 단당) 등의 성분분석 결과 기존에 보고된 국외
Haematococcus 속 녹조류와 상당히 유사한 것으로 판명되었다. 그러나
Haematococcus 속 NIBRIUM-7의 매우 높은 다중불포화지방산의 함량은
기능성 식품 및 화장품 등의 생산을 위한 본 녹조류주의 강력한 경쟁력을
부여하고 있다. 균주의 신규성에 있어서 Haematococcus 속 녹조류의 국외 활용
사례가 다수 보고되어 있는 관계로 산업재산권 확보 등의 산업화를 위한 기반
자료 확보 시 제한이 있을 것으로 파악되어 상기 언급한 바와 같이 추후 전체
유전체 분석 등을 통한 균주 특이성을 확립할 필요가 제기된다.
Lobosphaera 속 NIBRIUM-8의 경우, 상기 2종의 녹조류와 달리 거의
알려지지 않았으며 또한 기존 우리나라에 기록되지 않은 종으로 파악된다. 본
녹조류주는 담수, 기수 및 해수 조건에서 모두 배양이 가능한 것으로 확인되어
배양특성 부분에 있어 추후 활용성에 강점을 갖는다. 또한 바이오매스 생산량
역시 타 녹조류주에 뒤지지 않으며, 비교적 뛰어난 환경 적응 능력(pH, 온도,
염도 등)을 갖는 것으로 확인되었다. Lobosphaera 속은 현재 국내·외 활용성
연구가 거의 이루어지지 않았으며, 생태학적 연구 역시 거의 이루어지지 않은
상태이다. 따라서 본 연구에서 확보된 Lobosphaera 속 NIBRIUM-8 녹조류주의
산업재산권 확보와 관련하여 18S rRNA 및 18S-28S ITS 염기서열 분석을
통하여 동정을 진행하였고, 분석 결과 역시 기존에 보고된 국외 Lobosphaera
속과 상이한 것으로 판명되었다. 유용성 분석을 위한 일반성분(조단백, 조지방
및 조탄수화물 함량) 및 세부성분(아미노산, 지방산 및 단당) 등의 성분분석

- 95 -
결과 기존에 보고된 국외 Lobosphaera 속 녹조류주와 매우 유사하여 기능성
식품으로써 잠재력을 갖는다. 또한 ω6 계열의 특이 지방산을 상당량 생산하는
것으로 측정되었으며, 이는 지금까지 알려진 녹조류의 생산량 중 가장 높은
수치로 파악이 되고 있다. 이러한 특성은 아직까지 세포독성에 대한 연구가
이루어지지 않아 단정을 내릴 수는 없지만, 추후 기능성 식품 및 사료로써
강력한 경쟁력을 가질 것이다. 균주의 신규성 측면에 있어서 아직까지 해당
속의 녹조류의 국내·외 활용 보고가 없어 추후 산업재산권 확보 등의 산업화를
위한 기반자료 확보가 용이할 것으로 추정된다. 더불어 추후 전체 유전체
분석 및 세포독성 연구 등을 통한 균주 특이성을 확립할 필요가 제기된다.
Scenedesmus 속의 경우, 현재까지 가장 많이 연구된 녹조류 속으로써
생태학적 연구가 깊이 진행되어 있다. 다양한 수계환경에서 발견되는
Scenedesmus 속은 우리나라에서도 다수의 보고가 존재한다. Scenedesmus 속
녹조류 관련 연구는 최근 바이오에너지 생산과 결부되고 있으나, 해당 연구와
관련된 균주는 대부분 국내 자생주가 아닌 외래종으로 진행되고 있으며 국내
자생주의 유용성 및 분자생물학적 분석 등의 보고는 미미한 실정이다.
Scenedesmus 속 녹조류는 일반적으로 담수 서식으로 알려져 있으며, 따라서
본 연구과제 수행 중 해당 속 녹조류는 뗏목 형태의 군체로 제주도를 포함한
다양한 국내 담수 지역 등에서 관찰이 되었다. Scenedesmus 속 녹조류는 종
간 유전자 유사도가 매우 높아, 본 연구에서 확보된 Scenedesmus 속
NIBRIUM-9 녹조류주의 산업재산권 확보와 관련하여 18S rRNA 염기서열 분석을
통하여 동정을 진행하였고, 분석 결과 기존에 보고된 국내·외 Scenedesmus
속 녹조류와 유사한 것으로 판명되었다. 유용성 분석을 위한 일반성분(조단백,
조지방 및 조탄수화물 함량) 및 세부성분(아미노산, 지방산 및 단당) 등의 성분
분석 결과 기존에 보고된 국내·외 Scenedesmus 속 녹조류주와 상당히 유사한
것으로 판명되었다. 그러나 Scenedesmus 속 NIBRIUM-9의 상당히 높은 조지방
함량은 바이오에너지 생산을 위한 본 녹조류주의 강력한 경쟁력을 부여하고
있다. 그러나 균주의 신규성에 있어서 Scenedesmus 속 녹조류의 국내·외 활용
사례가 다수 보고되어 있는 관계로 산업재산권 확보 등의 산업화를 위한 기반
자료 확보 시 제한이 있을 것으로 파악되어 상기 언급한 바와 같이 추후 전체
유전체 분석 등을 통한 균주 특이성을 확립할 필요가 제기된다.
마지막으로 Chlorella 속의 경우, 현재까지 가장 많이 연구된 녹조류 속
중 하나로써 식품학적 효용성에 대한 연구가 깊이 진행되어 있다. 다양한
수계환경에서 발견되는 Chlorella 속은 우리나라에서도 다수 보고되며, 현재

- 96 -
어류 양식을 위한 먹이생물의 사료 및 기능성 식품 등으로 활발히 사용되고
있다. Chlorella 속 녹조류 관련 연구는 최근 바이오에너지 생산 등과도 결부
되고 있으나, 해당 연구와 관련된 균주는 대부분 국내 자생주가 아닌 외래종
으로 진행되고 있으며 국내 자생주의 유용성 분석 등의 보고는 미미한 실정이다.
Chlorella 속 녹조류는 일반적으로 담수, 기수 및 해수 서식으로 알려져 있으며,
따라서 본 연구과제 수행 중 해당 속 녹조류는 군체 형태로 제주도를 포함한
다양한 국내 수계 지역에서 관찰되었다. Chlorella 속 녹조류는 종 간 유전자
유사도가 매우 높아, 본 연구에서 확보된 Chlorella 속 NIBRIUM-10 녹조류주의
산업재산권 확보와 관련하여 18S rRNA 염기서열 분석을 통하여 동정을 진행
하였고, 분석 결과 기존에 보고된 국내·외 Chlorella 속 녹조류와 유사한 것으로
판명되었다. 유용성 분석을 위한 일반성분(조단백, 조지방 및 조탄수화물 함량)
및 세부성분(아미노산, 지방산 및 단당) 등의 성분분석 결과 기존에 보고된
국내·외 Chlorella 속 녹조류주와 상당히 유사한 것으로 판명되었다. 따라서
균주의 신규성에 있어 Chlorella 속 녹조류의 국내·외 활용 사례가 다수 보고
되어 있는 관계로 산업재산권 확보 등의 산업화를 위한 기반자료 확보 시
제한이 있을 것으로 파악되어 상기 언급한 바와 같이 추후 전체 유전체 분석
등을 통한 균주 특이성을 확립할 필요가 제기된다.
본 사업을 통해 확보된 10종의 자생 미세조류의 세포 성분 및 배양능 등의
유용성 관련 정보는 국내 자생 미세조류종의 보전 복원이나 경제적 가치가 높은
종 등 자생 생물자원의 과학적 관리 정책 마련 및 지속가능한 이용을 위한
기초 자료로 사용될 것이다. 특히, 미세조류는 광합성을 통하여 이산화탄소를
생물전환 방식으로 유기탄소 및 산소로 전환하는 능력을 보유하고 있으며, 체내
지질 및 당류 축적을 통한 바이오디젤 및 바이오에탄올 생산을 가능케 한다.
현 시점에서 재생가능하고 지속가능한 미세조류 유래 바이오연료 자원은 이산화
탄소 배출량 감축과 같은 환경 문제에 대한 해결방안을 기대할 수 있는 대체
자원으로 주목받고 있으며 미래 환경 개선을 위한 친환경적 접근 방안으로
활용이 가능할 것이다. 또한 차세대 기능성 식품 및 사료 생산을 위한 고단백,
고열량의 바이오매스로써도 활용성을 충분히 가질 것이다. 최근 국가 자생종의
유전체 정보 자체가 주요한 정보자원으로 여겨져, 전 세계적으로 미세조류의
유전체 해독을 위한 협력 연구가 가속되는 만큼, 우리나라 자생 유용 미세
조류의 유전체 해독 및 비교분석을 통한 기능유전체 연구는 추후 최우선 수행
되어야 할 과제이다. 따라서 “이산화탄소 생물전환을 위한 자생 미세조류

- 97 -
발굴”과 관련된 연구는 1차년도에서 선정된 유용 미세조류 3종(Leptolyngbya
sp. NIBRIUM-3, Lyngbya sp. NIBRIUM-5 및 Lobosphaera sp.
NIBRIUM-7)의 전체 유전체 및 유용물질 생산과 관련된 기능 유전체 분석과
관련된 부분에 초점을 맞추고자 한다. 이러한 연구를 통해 획득한 정보를
유전정보시스템(WIGIS)을 통해 체계적으로 관리하고, 관련 학계 및 산업계에
서비스할 수 있을 것이다.

- 98 -
V. 참고문헌
에너지경제연구원. 2013. 2013 에너지통계연보. 산업통상자원부. 8pp.
AOAC. 1990. Official Methods of Analysis of the Association of Official
Analytical Chemists, 15th edn. Association of Official Analytical
Chemists. Washington DC.
Baba M, Kikuta F, Suzuki I, Watanabe MM, Shiraiwa Y. 2012. Wavelength
specificity of growth, photosynthesis, and hydrocarbon production in
the oil-producing green alga Botryococcus braunii. Bioresour. Technol.109: 266-270.
Bligh EG, Dyer WJ. 1959. A rapid method of total lipid extraction and
purification. Can. J. Biochem. Physiol. 37: 911-917.
Borowitzka MA, Borowitzka LJ, Kessly D, 1990. Effects of salinity increase
on cartenoid accumulation in the green alga Dunaliella salina. J. Appl.Phycol. 2: 119-129.
Castresana J. 2000. Selection of conserved blocks from multiple alignments
for their use in phylogenetic analysis. Mol. Biol. Evol. 17: 540-552.
Cohen Z. 1986. Products from microalgae. In: A. Richmond (Eds), CRC
Handbook of Microalgal Mass Culture. CRC Press, Boca Raton, FL,
pp. 42: l-454.
Cohen Z, Cohen S. 1991. Preparation of eicosapentaenoic acid(EPA)
concentrate from Porphyridium cruentum. J. Am. Oil Chem. Soc., 68:16-19.
Edgar RC. 2004. MUSCLE: multiple sequence alignment with high accuracy
and high throughput. Nucleic. Acids Res. 32: 1792-1797.
Facciola S. 1998. Cornucopia II: A Sourcebook of Edible Plants. 2nd ed.
Kampong Publications, Vista, California.
Guindon S, Gascuel O. 2003. A simple, fast, and accurate algorithm to estimate
large phylogenies by maximum likelihood. Syst. Biol. 52: 696-704.
Hall TA. 1999. BioEdit: a user-friendly biological sequence alignment editor
and analysis. Department of Microbiology. North Carolina State
University, Raleigh, USA.

- 99 -
Huelsenbeck JP, Ronquist F. 2001. MRBAYES: Bayesian inference of
phylogenetic trees. Bioinformatics 17: 754-755.
Kim JH, et al., 2015. Isolation and characterization of Leptolyngbya sp.KIOST-1, a basophilic and euryhaline filamentous cyanobacterium
from an open paddle-wheel raceway Arthrospira culture pond inKorea. J. Appl. Microbiol. 119: 1597-1612.
Kim DG, Choi YE, 2014. Microalgae cultivation using LED light. K. Chem.
Eng. Res. 52: 8-16.
Komárek J, Anagnostidis K. 1986. Modern approach to the classification
system of cyanophytes, 2. Chroococcales. Arch. Hydrobiol. Algolog.
Stud. 43: 157-226.
Krejici M, et al., 2011. Selective sequestration of strontium in desmid green
algae by biogenic coprecipitation with barite. Chem. Sus. Chem. 4:
470-473.
Lane DJ, Pace B, Olsen GJ, Stahl DA, Sogin ML, Pace NR. 1985. Rapid
determination of 16S ribosomal RNA sequences for phylogenetic
analyses. Proc. Natl. Acad. Sci., U.S.A. 82: 6955-6959.
Luesch H, Williams PG, Yoshida WY, Moore RE, Paul VJ. 2002.
Ulongamides A-F, new beta-amino acid-containing cyclodepsipeptides
from palauan collections of the marine cyanobacterium Lyngbya sp. J.Nat. Prod. 65: 996-1000.
Nemoto-Kawamura C, Hirahashi T, Nagai T, Yamada H, Katoh T,
Hayashi O. 2004. Phycocyanin enhances secretary IgA antibody
response and suppresses allergic IgE antibody response in mice
immunized with antigen-entrapped biodegradable microparticles. J.
Nutr. Sci. Vitaminol. 50: 129-136.
Perkerson III RB, Johansen JR, Kovácik L, Brand J, Kaštovský J,
Casamatta DA. 2011. A unique pseudanabaenalean(cyanobacteria)
genus Nodosilinea gen. nov. based on morphological and moleculardata. J. Phycol 47: 1397-1412.
Peterson TC, Baringer MS. 2009. State of the climate in 2008. Bull. Amer.
Meteor. Soc., 90: S1-S196.
Posada D. 2008. jModelTest: phylogenetic model averaging. Mol. Biol. Evol.
25: 1253-1256.

- 100 -
Qiang H, Richmond A. 1996. Productivity and photosynthetic efficiency of
Spirulina platens is as affected by light intensity, algal density and rateof mixing in a flat plate photobioreactor. J. Appl. Phycol. 8: 139-145.
Richmond A, Preiss K. 1980. The biotechnology of algaculture. Interdiscip.
Sci. Rev. 5: 60-70.
Ringo DL. 1967. Flagellar motion and fine structure of the flagellar
apparatus in Chlamydomonas. J. Cell Biol. 33: 543-571.
Schaeffer DJ, Krylov VS. 2001. Anti-HIV Activity of Extracts and
compounds from algae and cyanobacteria. Department of Veterinary
Biosciences, University of Illinois, Urbana-Champaign, USA.
Spurr AR. 1969. A low-viscosity epoxy resin embedding medium for
electron microscopy. J. Ultrastruct. Res. 26: 31-43.
Subhashini J, Mahipal SV, Reddy MC, Mallikarjuna RM, Rachamallu A,
Reddanna P. 2004. Molecular mechanisms in c-phycocyanin induced
apoptosis in human chronic myeloid leukaemia cell line-k562.
Biochem. Pharm. 68:.453-462.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011.
MEGA5: molecular evolutionary genetics analysis using maximum
likelihood, evolutionary distance, and maximum parsimony methods.
Mol. Biol. Evol. 28: 2731-2739.
Taton A, Grubisic S, Brambilla E, De Wit R, Wilmotte A. 2003.
Cyanobacterial diversity in natural and artificial microbial mats of
lake Fryxell (McMurdo dry valleys, Antarctica): a morphological and
molecular approach. Appl. Environ. Microbiol. 69: 5157-5169.
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. 1997.
The ClustalX windows interface: Flexible strategies for multiple
sequence alignment aided by quality analysis tools. Nucleic. Acids.
Res. 24: 4876-4882.
Waterbury JB, Stanier RY. 1978. Patterns of growth and development in
pleurocapsalean cyanobacteria. Bacteriol. Rev. 42: 2-44.
White TJ, Bruns T, Lee S, Taylor JW. 1990. Amplification and direct
sequencing of fungal ribosomal RNA genes for phylogenetics. pp.
315-322 In: PCR Protocols: A Guide to Methods and Applications,
eds. Innis MA, Gelfand DH, Sninsky JJ, White TJ. Academic Press,
Inc., New York.

- 101 -
부록 I. 미세조류 배양체 염기서열
1. Cyanophyta
>Arthrospira sp._NIBRIUM-1. 16S ribosomal RNA gene, partial sequence (1421bp)
ATCCTAGCTCAGGATGAAAGCTGGCGGTCTGCTTAACACATGCAAGTCGAAC
GGGCTCTTCGGAGCTAGTGGCGGACGGGTGAGTAACACGTGAGAATCTGGCT
CCCGGTCGGGGACAACAGAGGGAAACTTCTGCTAATCCCGGATGAGCCGAAA
GGTAAAAGATTTATCGCCGGGAGATGAGCTCGCGTCTGATTAGCTAGTTGGT
GAGGTAAAGGCTCACCAAGGCGACGATCAGTAGCTGGTCTGAGAGGATGATC
AGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGG
GAATTTTCCGCAATGGGCGCAAGCCTGACGGAGCAAGACCGCGTGGGGGAGG
AAGGCTCTTGGGTTGTAAACCCCTTTTCTCAAGGAAGAACACAATGACGGTA
CTTGAGGAATAAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAG
GAGGCAAGCGTTATCCGGAATGATTGGGCGTAAAGCGTCCGTAGGTGGCTGT
TCAAGTCTGCTGTCAAAGACAGTGGCTTAACTACTGAAAGGCAGTGGAAACT
GAACAGCTAGAGTACGGTAGGGGCAGAGGGAATTCCCGGTGTAGCGGTGAAA
TGCGTAGATATCGGGAAGAACACCGGTGGCGAAAGCGCTCTGCTGGGCCGTA
ACTGACACTGAGGGACGAAAGCTAGGGGAGCGAATGGGATTAGATACCCCAG
TAGTCCTAGCCGTAAACGATGGAAACTAGGTGTAGCCTGTATCGACCCGGGC
TGTGCCGAAGCTAACGCGTTAAGTTTCCCGCCTGGGGAGTACGCACGCAAGTG
TGAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGTATGTGGTT
TAATTCGATGCAACGCGAAGAACCTTACCAGGGCTTGACATGTCCGGAATCT
TGGTGAAAGCCGAGAGTGCCTTCGGGAGCCGGAACACAGGTGGTGCATGGCT
GTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACC
CTCGTCCTTAGTTGCCATCATTCAGTTGGGCACTTTAGGGAGACTGCCGGTG
ACAAACCGGAGGAAGGTGGGGATGACGTCAAGTCATCATGCCCCTTACGTCC
TGGGCTACACACGTACTACAATGGGGGGGACAAAGGGTAGCCAAGACGCGAG
TCTGAGCCAATCCCGTAAACCTCTCCTCAGTTCAGATTGCAGGCTGCAACTCG
CCTGCATGAAGGAGGAATCGCTAGTAATCGCAGGTCAGCATACTGCGGTGAA
TCCGTTCCCGGGCCTTGTACACACCGCCCGTCACACCATGGAAGTTAGCCACG
CCGGAAGTCGTTACTCTAACCGTTCGCGGAGGAGGATGCCGCAGGCAGGGCTG
ATAACTCGGG

- 102 -
>Arthrospira sp._NIBRIUM-2. 16S ribosomal RNA gene, partial sequence (1423bp)
GGGGGTCTGCTTAACACATGCAAGCCGAACGGGCTCTTCACAGCTAGTGGCGG
ACGGGTGAGTAACACGTGAGAATCTGGCTCCCGGTCGGGACAACAGAGGGAA
ACTTCTGCTAATCCCGGATGAGCCGAAAGGTAAAAGATTTATCGCCGGGAGA
TGAGCTCGCGTCTGATTAGCTAGTTGGTGAGGTAAAGGCTCACCAAGGCGAC
GATCAGTAGCTGGTCTGAGAGGATGATCAGCCACACTGGGACTGAGACACGG
CCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGNAAGC
TGACGGAGCAAGACCGCGTGGGGGAGGAAGGCTCTTGGGTNGTAAACCCCTT
TTCTCAAGGAAGAACACAATGACGGTACTTGAGGAATAAGCCTCGGCTAACT
CCGTGCCAGCAGCCGCGGTAATACGGAGGAGGCAAGCGTTATCCGGAATGAT
TGGGCGTAAAGCGTCCGTAGGTGGCAGTTCAAGTCTGCTGTCAAAGACAGTA
GCTCAACTACTGAAAGGCAGTGGAAACTGAACAGCTAGAGTACGGTAGGGGC
AGAGGGAATTCCCGGTGTAGCGGTGAAATGCGTAGATATCGGGAAGAACACC
GGTGGCGAAAGCGCTCTGCTGGGCCGTAACTGACACTGAGGGACGAAAGCTA
GGGGAGCGAATGGGATTAGATACCCCAGTAGTCCTAGCCGTAAACGATGGAA
ACTAGGTGTAGCCTGTATCGACCCGGGCTGTGCCGAAGCTAACGCGTTAAGT
TTCCCGCCTGGGGAGTACGCACGCAAGTGTGAAACTCAAAGGAATTGACGGG
GGCCCGCACAAGCGGTGGAGTATGTGGTTTAATTCGATGCAACGCGAAGAAC
CTTACCAGGGCTTGACATGTCCGGAATCTTGGTGAAAGCCGAGAGTGCCTTC
GGGAGCCGGAACACAGGTGGTGCATGGCTGTCGTCAGCTCGTGTCGTGAGATG
TTGGGTTAAGTCCCGCAACGAGCGCAACCCTCGTCCTTAGTTGCCATCATTCA
GTTGGGCACTTTAGGGAGACTGCCGGTCACAAACCGGAGGAAGGTGGGGATG
ACGTCAAGTCATCATGCCCCTTACGTCCTGGGCTACACACGTACTACAATGGG
GGGGACAAAGGGTAGACAAGACGCGAGTCTGAGCCAATCCCGTAAACCTCTC
CTCAGTTCAGATTGCAGGCTGCAACTCGCCTGCATGAAGGAGGAATCGCTAG
TAATCGCAGGTCAGCATACTGCGGTGAATCCGTTCCCGGGCCTTGTACACACC
GCCCGTCACACCATGGAAGTTAGCCACGCCCGAATTCGTTACTCTAACCGTTC
GCGGAGGACGATGCCGAAGGCAGGGCTGATGACTGGGGTGAAGTCGTAACAA
GTTAGCCGTACC

- 103 -
>Leptolyngbya sp._NIBRIUM-3. 16S ribosomal RNA gene, partial sequence;
16S-23S ribosomal RNA intergenic spacer, complete sequence; and 23S ribosomal
RNA gene, partial sequence (1991bp)
AGGCTCAGGATGAACGCTGGCGGCGTGCTTAACACATGCAAGTCGAACGGGA
TCTTCGGATCTAGTGGCGGACGGGTGAGTAACGCGTGAGGATCTGCCCTTAG
GATGGGGACAACCACTGGAAACGGTGGCTAATACCCAATGTGCCGAGAGGTG
AAACATTAATGGCCTGAGGATGAACTCGCGTCTGATTAGCTAGTTGGTGAGG
TAAGGGCTCACCAAGGCGACGATCAGTAGCTGGTCTAAGAGGATGATCAGCC
ACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAAT
TTTCCGCAATGGGCGCAAGCCTGACGGAGCAACGCCGCGTGAGGGAGGAAGGC
CTTAGGGTTGTAAACCTCTTTTCTCTGGGAAGAAGACCTGACGGTACCAGAG
GAATAAGCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAAGACGGAGGAGGCA
AGCGTTATCCGGAATTATTGGGCGTAAAGCGTCCGCAGGCGGTTTTTTAAGT
CTGTTGTCAAAGGTCACAGCTCAACTGTGGATCGGCAATGGAAACTGGGGAA
CTTGAGTGTGGTAGGGGTAGAGGGAATTCCCGGTGTAGCGGTGAAATGCGTA
GATATCGGGAAGAACACCAGTGGCGAAGGCGCTCTACTGGGCCACAACTGAC
GCTGAGGGACGAAAGCTAGGGGAGCGAAAGGGATTAGATACCCCTGTAGTCC
TAGCTGTAAACGATGGATACTAGGTGTTGGACGTATCGACCCGTGCAGTACC
GTAGCTAACGCGTTAAGTATCCCGCCTGGGGAGTACGCACGCAAGTGTGAAA
CTCAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGGATGTGGTTTAATT
CGATGCAACGCGAAGAACCTTACCAAGGCTTGACATGTCGCGAATCTCTGAG
AGATCAGAGAGTGCCTTCGGGAGCGCGAACACAGGTGGTGCATGGCTGTCGT
CAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCACGT
TTTTAGTTGCCAGCATTCAGTTGGGCACTCTAAAGAGACTGCCGGGGACAAC
TCGGAGGAAGGTGTGGACGACGTCAAGTCATCATGCCCCTTACGTCTTGGGC
TACACACGTCCTACAATGCTACAGACAGAGGGCAGCGAGCGCGCGAGTGCAAG
CAAATCCCATAAACTGTGGCTCAGTTCAGATTGCAGGCTGCAACTCGCCTGC
ATGAAGGCGGAATCGCTAGTAATCGCCGGTCAGCATACGGCGGTGAATACGT
TCCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTTGGCCACGCCCGA
AGTCGTTACTCTAACCGTTCGCGGAGGAGGGCGCCGAAGGCAGGGCTGATGAC
TGGGGTGAAGTCGTAACAAGGTAGCCGTACCGGAAGGTGTGGCTGGATCACC
TCCTTTTAGGGAGACCTACCCTTGATAACTCCGAACCATTGTTATTAGGAGT
ATCGAGCGGTCACTCTAGGTCGGTCAGGACTTTGAGTCGAGTTTCTAGACTA
TGGTTGAGGTTTTGTTTAACCAACAGAACCCGGGCTATTAGCTCAGGTGGTT
AGAGCGCACCCCTGATAAGGGTGAGGTCCCTGGTTCGAGTCCAGGATGGCCCA
CTGCGTGGGGGTTTAGCTCAGTTGGTAGAGCGCCTGCTTTGCAAGCAGGATG

- 104 -
TCAGCGGTTCGAGTCCGCTAACCTCCACCACGAAAGGACGGATTGAATTCAGC
ACAGTATGTGTTTCTTGAATTATCAAGAGGATACATAGTCTGCTGGGTGAG
AGCCCGGCTAAGAACCTTGACAACTGAATAGTGAGAGAAGTGTAAGGTAGTA
CACACAGAACATTCAAAGTTCGTTGAGAGAAGAGTGAATTTTGTTGGGTAA
CACCAATGAAAGGAAGGTCAAGCTACAACGGGCTCACGGTGGATACCTAGGC
ACACAGAG

- 105 -
>Myxosarcina sp._NIBRIUM-4. 16S ribosomal RNA gene, partial sequence
(1123bp)
GCGGCCGATTAGCTAGTTGGTGGGGTAAGAGCCTACCAAGGCAGCGATCGGT
AGCTGGTCTGAGAGGATGAGCAGCCACACTGGGACTGAGACACGGCCCAGACT
CCTACGGGAGGCAGCAGTGGGGAATTTTCCGCAATGGGCGAAAGCCTGACGG
AGCAATACCGCGTGAGGGAGGAAGGCTTTTGGGTTGTAAACCTCTTTTGTTA
AGGAAGAATCAAGTGACGGTACTTAACGAATCAGCATCGGCTAACTCCGTGC
CAGCAGCCGCGGTAATACGGAGGATGCAAGCGTTATCCGGAATCATTGGGCG
TAAAGCGTCCGCAGGTGGTTCATCAAGTCTGCTGTTAAAGACAGAAGCTCAA
CTTCTGAAAGGCGGTGGAAACTGGAGAACTAGAGTACGGTAGGGGTCGAGGG
AATTCCCAGTGTAGCGGTGAAATGCGTAGAGATTGGGAAGAACACCAGTGGC
GAAGGCGCTCGACTGGGCCGTAACTGACACTCAGGGACGAAAGCTAGGGTAG
CGAAAGGGATTAGATACCCCTGTAGTCCTAGCGGTAAACGATGAACACTAGG
CGTTGCTTGTATCGACCCGAGCAGTGCCGTAGCCAACGCGTTAAGTGTTCCGC
CTGGGGAGTACGCACGCAAGTGTGAAACTCAAAGGAATTGACGGGGGCCCGC
ACAAGCGGTGGAGTATGTGGTTTAATTCGATGCAACGCGAAGAACCTTACCA
GGACTTGACATCCCAAGAACCCAGCGTAATGGCTGGGGTGCCTTAGGGAACT
TGGAGACAGGTGGTGCATGGCTGTCGTCAGCTCGTGTCGTGAGATGTTGGGT
TAAGTCCCGCAACGAGCGCAACCCTCGTTCTTAGTTGCCAGCATTAAGTTGG
GCACTTTAGGAAGACTGCCGGTGACAAACCGGAGGAAGGTGGGGATGACGTC
AAGTCAGCATGCCCCTTACGTTCTGGGCTACACACGTACTACAATGGTTGGG
ACAAAGGGCAGCGAACTCGCGAGAGTAAGCGAATCTCAGCAAACCCAGCCTC
AGTTCAGATTGCAGGCTGCAACTCGCCTGCATGAAGGAGGAATCGCTAGTAA
TCGCAGGTCAGCATACGGCGGTGAATCCG

- 106 -
>Lyngbya sp._NIBRIUM-5. 16S ribosomal RNA gene, partial sequence (1420bp)
GATGAACGCTGGCGGTCTGCTTAACACATGCAAGTCGAACGGGCTTCTTCGG
AGCTAGTGGCGGACGGGTGAGTAACACGTGAGAATCTGGCTCCCGGTCGGGG
ACAACAGAGGGAAACTTCTGCTAATCCCGGATGAGCCGAAAGGTAAAAGATT
TATCGCCGGGAGATGAGCTCGCGTCTGATTAGCTAGTTGGTGAGGTAAAGGC
TCACCAAGGCGACGATCAGTAGCTGGTCTGAGAGGATGATCAGCCACACTGG
GACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTCCGC
AATGGGCGCAAGCCTGACGGAGCAAGACCGCGTGGGGGAGGAAGGCTCTTGGG
TTGTAAACCCCTTTTCTCAAGGAAGAACACAATGACGGTACTTGAGGAATAA
GCCTCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGAGGCAAGCGTT
ATCCGGAATGATTGGGCGTAAAGCGTCCGTAGGTGGCAGTTCAAGTCTGCTG
TCAAAGACAGTAGCTCAACTACTGAAAGGCAGTGGAAACTGAACAGCTAGAG
TACGGTAGGGGCAGAGGGAATTCCCGGTGTAGCGGTGAAATGCGTAGATATC
GGGAAGAACACCGGTGGCGAAAGCGCTCTGCTGGGCCGTAACTGACACTGAGG
GACGAAAGCTAGGGGAGCGAATGGGATTAGATACCCCAGTAGTCCTAGCCGT
AAACGATGGAAACTAGGTGTAGCCTGTATCGACCCGGGCTGTGCCGAAGCTA
ACGCGTTAAGTTTCCCGCCTGGGGAGTACGCACGCAAGTGTGAAACTCAAAG
GAATTGACGGGGGCCCGCACAAGCGGTGGAGTATGTGGTTTAATTCGATGCA
ACGCGAAGAACCTTACCAGGGCTTGACATGTCCGGAATCTTGGTGAAAGCCG
AGAGTGCCTTCGGGAGCCGGAACACAGGTGGTGCATGGCTGTCGTCAGCTCGT
GTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTCGTCCTTAGTT
GCCATCATTCAGTTGGGCACTTTAGGGAGACTGCCGGTGACAAACCGGAGGA
AGGTGGGGATGACGTCAAGTCATCATGCCCCTTACGTCCTGGGCTACACACGT
ACTACAATGGGGGGGACAAAGGGTAGCCAAGACGCGAGTCTGAGCCAATCCC
GTAAACCTCTCCTCAGTTCAGATTGCAGGCTGCAACTCGCCTGCATGAAGGA
GGAATCGCTAGTAATCGCAGGTCAGCATACTGCGGTGAATCCGTTCCCGGGCC
TTGTACACACCGCCCGTCACACCATGGAAGTTAGCCACGCCCGAAGTCGTTAC
TCTAACCGTTCGCGGAGGAGGATGCCGAAGGCAGGGCTGATGACTGGGGTGA
AGTCGTA

- 107 -
2. Chlorophyta
>Chlamydomonas sp._NIBRIUM-6. 18S ribosomal RNA gene, partial sequence;
internal transcribed spacer 1, 5.8S ribosomal RNA gene, and internal transcribed
spacer 2, complete sequence; and 28S ribosomal RNA gene, partial sequence
(2655bp)
GGAACAGTCTGATCTAGTATAACTGCTTATACTGTGAAACTGCGAATGGCTC
ATTAAATCAGTTATAGTTTATTTGATGGTACTTACTACTTGGATAACCGTA
GTAATTCTAGAGCTAATACATGCGTAAATCCCGACTTCTGGAAGGGACGTAT
TTATTAGATAAAAGGCCAGCCGGGCTCTGCCCGACCTGCGGTGAATCATGAT
AACTTCACGAATCGCACGCCCTTGCGGCGGCGATGTTTCATTCAAATTTCTGC
CCTATCAACTTTCGATGGTAGGATAGAGGCCTACCATGGTGGTAACGGGTGA
CGGAGGATTAGGGTTCGATTCCGGAGAGGGAGCCTGAGAGATGGCTACCACA
TCCAAGGAAGGCAGCAGGCGCGCAAATTACCCAATCCCGACACGGGGAGGTAG
TGACAATAAATAACAATACCGGGCGCAACGCGTCTGGTAATTGGAATGAGTA
CAATTTAAATCCCTTAACGAGGATCCATTGGAGGGCAAGTCTGGTGCCAGCA
GCCGCGGTAATTCCAGCTCCAATAGCGTATATTTAAGTTGTTGCAGTTAAAA
AGCTCGTAGTTGGATTTCGGGTGGGTTGGCGCGGTCCGCCTCTGGTGTGCACT
GCGTCGGTCCACCTTCCTGCCGGGGACGGGCTCCTGGGCTTCACTGTTCGGGA
CTCGGAGTCGGCGTTGTTACTTTGAGTAAATTAGAGTGTTCAAAGCAAGCCT
ACGCTCTGAATATGTTAGCATGGAATAACGCGATAGGACTCTGGCCTATTCG
TTGGTCTGTGGGACTGGAGTAATGATTAAGAGGGACAGTCGGGGGCATTCGT
ATTCCGTTGTCAGAGGTGAAATTCTTGGATTTACGGAAGACGAACTACTGCG
AAAGCATTTGCCAAGGATGTTTTCATTAATCAAGAACGAAAGTTGGGGGCTC
GAAGACGATTAGATACCGTCGTAGTCTCAACCATAAACGATGCCGACTAGGG
ATCGGCGGGTGTTTCATTGATGACCCCGCCGGCACCTTATGAGAAATCAAAG
TTTTTGGGTTCCGGGGGGAGTATGGTCGCAAGGCTGAAACTTAAAGGAATTG
ACGGAAGGGCACCACCAGGCGTGGAGCCTGCGGCTTAATTTGACTCAACACGG
GAAAACTTACCAGGTCCAGACACGGGAAGGATTGACAGATTGAGAGCTCTTT
CTTGATTCTGTGGGTGGTGGTGCATGGCCGTTCTTAGTTGGTGGGTTGCCTT
GTCAGGTTGATTCCGGTAACGAACGAGACCTCAGCCTGCTAAATAGTCAGCA
TCGCACCTGCGGTGCGCAGACTTCTTAGAGGGACTATTGGCGTTTAGCCAAT
GGAAGTATGAGGCGATAACAGGTCTGTGATGCCCTTAGATGTTCTGGGCCGC
ACGCGCGCTACACTGACGCGATCAACGAGCCTATCCTTGGCCGAGAGGCCCGG
GTAATCTTGAAAACCGCGTCGTGATGGGGATAGATTATTGCAATTATTAGTC
TTCAACGAGGAATGCCTAGTAAGCGCGAGTCATCAGCTCGCGTTGATTACGT

- 108 -
CCCTGCCCTTTGTACACACCGCCCGTCGCTCCTACCGATTGGGTGTGCTGGTG
AAATGCTCGGATTCGTCCTTGGTGGGGCAACCCGCTTTGGATGGAGAAGTTC
ATTAAACCCTCCCACCTAGAGGAAGGAGAAGTCGTAACAAGGTTTCCGTAGG
TGAACCTGCGGAAGGATCATTGAATCTATCACAATCCACAACCACTGCGAAC
CGTCGAAATGTTGGCCCCCCGCTGACTCGGAGGGCGGCGAAAGCCCCCTCCGA
TGATGGGCCTCTTGCCTCCGGGCGAGGGGTCGGTCTTTATCCAACCAACCAAC
ACCAAACCATAACCTCTAAAGCGAGCTTGATTGATCGGCTTAGAGCCAATCG
ATCTACTAAACCAAGACAACTCTCAACAACGGATATCTTGGCTCTCGCAACG
ATGAAGAACGCAGCGAAATGCGATACGTAGTGTGAATTGCAGAAATACGTGA
ACCATCGAATCTTTGAACGCATATTGCGCTCGAGCCTTCGGGCAAGAGCATG
TCTGCCTCAGCGTCGCTCTAAATCCTCATCAACACTACTATCCTACACGTAGG
ATGGTTCGAGTTTGATGGAACTGGCTTTCCCAAACCATTCACTTGGATTGGG
TCGGCTGAAGAGCAGAGGTTGATGCATGGACCCGCTATGGGCCTCGGCTAGGT
AGGCAACTCGTTTGCTCATGCTTTAGCTGTTGGCTTGGGACTGTGCTTCGAC
CCAAACCAAGGAATGCCGAAGTTTGCCTTCGGCATAACTTTCTGCTTTCGAC
CTGAGCTCAGGCAAGAACACCCGCTGAACTTAAGCATATCAATAAGCGGAGG
AAAAGAAAATAACAATGATTGCCCTAGTAACGGCGAGTGAAGCGGCAATAGC
CCAACTTGAAAATCTCCCATTGGAGAATTGTAGTCTATAGAAGTAGCGTCTG
AGGCGAGCGGAGCTCAAGTCCACTGGAAAGTGGCGTCAGAGAGGGTGAGAAC
CCCGTCGGCTCCGTCTTTGTCTCTCCACGATGCGCTTTCCACGAGTCGGGTTG
TTTGGGAATGCAGCCCTAATTTGGTGGTAAATCCCATCTAAGGC

- 109 -
>Haematococcus sp._NIBRIUM-7. 18S ribosomal RNA gene, partial sequence (1777bp)
TACCTGGTTGATCCTGCCAGTAGTCATATGCTTGTCTCAAAGATTAAGCCAT
GCATGTCTAAGTATAAACTGCTTATACGGTGAAACTGCGAATGGCTCATTAA
ATCAGTTATAGTTTATTTGATGGTACTTTACTCGGATAACCGTAGTAATTC
TAGAGCTAATACGTGCGTATATCCCGACTTCTGGAAGGGACGTATTTATTAG
ATAAAAGGCCAGCCGGGCTTGCCCGACCTATGGCGAATCATGATAACTTCAC
GAATCGCACGGCCTTGCGCCGGCGATGTTTCATTCAAATTTCCGCCCTATCAA
CTTTCGATGGTAGGATAGAGGCCTACCATGGTGGTAACGGGTGACGGAGGAT
TAGGGTTCGATTCCGGAGAGGGAGCCTGAGAAACGGCTACCACATCCAAGGA
AGGCAGCAGGCGCGCAAATTACCCAATCCCGACACGGGGAGGTAGTGACAAT
AAATAACAATACCGGGCATCAATGTCTGGTAATTGGAATGAGAACAATTTA
AATCCCTTAACGAGTATCCATTGGAGGGCAAGTCTGGTGCCAGCAGCCGCGGT
AATTCCAGCTCCAATAGCGTATATTTAAGTTGTTGCAGTTAAAAAGCTCGTA
GTTGGATTTCGGGTGGGTTCCAGCGGTCTGCCTCTGGTATGTACTGCTGTGGC
CTACCTTTCTGCCGGGGACGTGTTCCTGGGCTTCATTGTCCGGGACTCGAATT
CGGCGAGGATACTTTGAGTAAAACAGCGTGTTCAAAGCAAGCCTACGCTCTG
AATGCATTAGCATGGAATATCACGATAGGACTCTGGCCTATCTTGTTGGTCT
GTAGGACCGGAGTAATGATTAAGAGGGACAGTCGGGGGCATTCGTATTTCAT
TGTCAGAGGTGAAATTCTTGGATTTATGAAAGACGAACTTCTGCGAAAGCA
TTTGCCAAGGATGTTTTCATTAATCAAGAACGAAAGTTGGGGGCTCGAAGAC
GATTAGATACCGTCGTAGTCTCAACCATAAACGATGCCGACTAGGGATTGGC
AGGTGTTTTATTGATGACCCTGCCAGCACCTTATGAGAAATCAAAGTTTTTG
GGTTCCGGGGGGAGTATGGTCGCAAGGCTGAAACTTAAAGGAATTGACGGAA
GGGCACCACCAGGCGTGGAGCCTGCGGCTTAATTTGACTCAACACGGGAAAAC
TTACCAGGTCCAGACACGGGAAGGATTGACAGATTGAGAGCTCTTTCTTGAT
TCTGTGGGTGGTGGTGCATGGCCGTTCTTAGTTGGTGGGTTGCCTTGTCAGG
TTGATTCCGGTAACGAACGAGACCTCAGCCTGCTAAATAGTCAAGCGTACCT
TGGTACGCGCCTGACTTCTTAGAGGGACTATTGACGTTTAGTCAGTGGAAGT
GTGAGGCAATAACAGGTCTGTGATGCCCTTAGATGTTCTGGGCCGCACGCGCG
CTACACTGATGCATTCAGCGAGCCTATCCTTGGCCGAGAGGTCCGGGTAATCT
TTGAAACTGCATCGTGATGGGGATAGATTATTGCAATTATTAGTCTTCAACG
AGGAATGCCTAGTAAGCGCGAGTCATCAGCTCGCGTTGATTACGTCCCTGCCC
TTTGTACACACCGCCCGTCGCTCCTACCGATTGGGTGTGCTGGTGAAGTGTTC
GGATTGACTTCAGCGGTGGGCAACCTCTGCTGTCGTTGAGAAGATCATTAAA
CCCTCCCACCTAGAGGAAGGAGAAGTCGTAACAAGGTCTCCGTAGGTGAACC
TGC

- 110 -
>Haematococcus sp._NIBRIUM-7. ribulose-1,5-bisphosphate carboxylase/oxygenase
large subunit gene, partial cds; chloroplast (1102bp)
GCGGGTGTTAAAGATTATCGTTTAACGTATTATACACCAGATTACGTTGTT
AGAGATACTGATATTTTAGCAGCGTTCCGTATGACACCACAACCAGGTGTAC
CACCAGAAGAATGTGGTGCAGCGGTAGCAGCTGAATCATCAACAGGTACTTG
GACAACAGTATGGACAGATGGTTTAACAAGTTTAGACCGTTACAAAGGTAG
ATGCTACGACATCGAGCCAGTACCCGGCGAGGACAATCAATACATCGCTTAC
GTAGCCTACTCTATCGATTTATTTGAAGAAGGTTCTGTAACTAACATGTTT
ACTTCAATTGTAGGGAACGTTTTTGGGTTTAAAGCATTACGTGCATTACGTT
TAGAAGATTTACGTATTCCACCAGCATATGTAAAAACATTCTCAGGACCTCC
ACACGGTATTCAAGTTGAACGTGACAAAATTAACAAATACGGCCGTGGATTA
TTAGGCTGTACGATTAAACCGAAATTAGGACTTTCAGCAAAAAATTATGGA
CGTGCCGTTTACGAATGTTTACGCGGTGGTCTAGACTTTACAAAAGATGACG
AAAACGTAACTTCACAACCTTTTATGCGCTGGAGAGACCGTTTCTTATTCGT
AGCTGAAGCTATTTACAAAGCACAATCAGAAACAGGAGAGGTTAAAGGCCAT
TATTTGAACGCAACTGCTGGTACAGCAGAAGAAATGCTAAAACGTGCACAAT
GTGCTAAGGAATTAGGTGTGCCCATTATCATGCATGATTATTTAACAGGTGG
TTTTACATCAAATACTTCATTAGCACATTTCTGTCGTGATAATGGTTTATT
ATTACACATTCACCGTGCTATGCATGCTGTTATCGACCGCCAAAGAAACCAT
GGTATTCATTTCCGTGTATTAGCGAAAGCTTTACGTATGTCAGGTGGTGACC
ACTTACACTCTGGAACAGTTGTAGGTAAATTAGAAGGTGAACGTGAAGTAA
CTTTAGGTTTCGTAGACTTAATGCGTGATGATTACATTGAAAAAGATCGTA
GTCGTGGTATTTACTTTACACAAGATTGGTGTTCAATGCCAGGTGTAATGCC
AGTAGCTTCAGGTGGTA

- 111 -
>Lobosphaera sp._NIBRIUM-8. 18S ribosomal RNA gene, partial sequence
(1627bp)
GCGAATGGCTCATTAAATCAGTTATAGTTTATTTGATGGTACCTTACTACTC
GGATAACCGTAGTAATTCTAGAGCTAATACGTGCGTAAATCCCGACTTCTGG
AAGGGACGTATTTATTAGATAAAAGGCCGACCGGGCTCTGCCCGACTCGCGG
TGAATCATGATAACTTCACGAATCGCATGGCCTCTGTGCCGGCGATGTTTCA
TTCAAATTTCTGCCCTATCAACTTTCGATGGTAGGATAGAGGCCTACCATGG
TGGTAACGGGTGACGGAGGATTAGGGTTCGATTCCGGAGAGGGAGCCTGAGA
AACGGCTACCACATCCAAGGAAGGCAGCAGGCGCGCAAATTACCCAATCCTGA
TTCAGGGAGGTAGTGACAATAAATAACAATACCGGGCTTTTTCAAGTCTGGT
AATTGGAATGAGTACAATCTAAATCCCTTAACGAGGATCAATTGGAGGGCA
AGTCTGGTGCCAGCAGCCGCGGTAATTCCAGCTCCAATAGCGTATATTTAAG
TTGTTGCAGTTAAAAAGCTCGTAGTTGGATTTCGGGTGGGCGATGTCGGTCC
GCCGTTTCGGTGTGCACTGGCAGCGCCTACCTTGCTGCCGGGGACGGGCTCCT
GGGCTTAACTGTCCGGGACCCGGAGTCGGCGAGGTTACTTTGAGTAAATTAG
AGTGTTCAAAGCAGGCCTACGCTCTGAATACATTAGCATGGAATAACACGAT
AGGACTCTGGCCTATCTTGTTGGTCTGTGGGACCGGAGTAATGATTAAGAGG
GACAGTCGGGGGCATTCGTATTTCATTGTCAGAGGTGAAATTCTTGGATTTA
TGAAAGACGAACTTCTGCGAAAGCATTTGCCAAGGATGTTTTCATTAATCAA
GAACGAAAGTTGGGGGCTCGAAGACGATTAGATACCGTCCTAGTCTCAACCA
TAAACGATGCCGACTAGGGATTGGAGGGTGTTTATTGATGACCCCTCCAGCA
CCTTATGAGAAATCAAAGTTTTTGGGTTCCGGGGGGAGTATGGTCGCAAGGC
TGAAACTTAAAGGAATTGACGGAAGGGCACCACCAGGCGTGGAGCCTGCGGC
TTAATTTGACTCAACACGGGAAAACTTACCAGGTCCAGACATAGTGAGGATT
GACAGATTGAGAGCTCTTTCTTGATTCTATGGGTGGTGGTGCATGGCCGTTC
TTAGTTGGTGGGTTGCCTTGTCAGGTTGATTCCGGTAACGAACGAGACCTCA
GCCTGCTAAATAGTCACGGTTGGTTCTTCCAGCCGGCCGACTTCTTAGAGGGA
CTATTGGCGACTAGCCAATGGAAGTGTGAGGCAATAACAGGTCTGTGATGCC
CTTAGATGTTCTGGGCCGCACGCGCGCTACACTGATGCATTCAACGAGCCTAG
CCTGGACCGAGAGGTTCGGGTAATCTTTGAAACTGCATCGTGATGGGGATAG
ATTATTGCAATTATTAATCTTCAACGAGGAATGCCTAGTAAGCGCGAGTCAT
CAACTCGCGTTGATTACGTCCATGCCCTTTGTACACACCGCCCGTCGTTCCTA
CCGATTGGGTGTGCTGGTGAAGCTTTCGGATTGGCGACTGCTGACGGTTCCCC
GCCCGCTGTC

- 112 -
>Lobosphaera sp._NIBRIUM-8. internal transcribed spacer 1, partial sequence; 5.8S
ribosomal RNA gene and internal transcribed spacer 2, complete sequence; and
28S ribosomal RNA gene, partial sequence (1157bp)
CGCTCAATCTCCTCGATTGCTGCCGGGTGCTCCGGGCGCTGGCCCGGCTGCGGC
CCCAGGCTCACCTTCCCCCCGGGGGAGGCCCTGGGGCCCGCATGTCGGGGTCCA
ACCCTGCTTAACCAGCAGGCGACGGTCCAGCGGGCGGGGAGGGCTCGGTTCCT
CTTTCGGGAGGTCCGGCCTGACCGCTGACCAAAACAACCTTTCCACCCAAAAC
CATCCTCTGAAGCTTGCGGAGACCGGCCTGGCCGGCTCCTTCAAACCAAAGAC
AACTCTCAACAACGGATATCTAGGCTCCCGTAACGATGAAGAACGCAGCGAA
ATGCGATACGTAGTGTGAATTGCAGAATTCCGTGAACCATCGAATCTTTGAA
CGCATATTGCGCTCGAGGCTTCGGCCGAGAGCATGTCTGCCTCAGCGTCGGTC
CATCCCCTCACACCCTCCCTTTGTTTTGCAAATGGGCTGGGCCGTGGATCTGG
CCGTGTCGGCTGCCCCGTGTGGCCGAGTCGGCTGAAGCTCAGAGGCTTGAGCA
AGCGCCCATTCAACCGGGCATCAACTTGGTGGGTGGCCTTCGGGCTGCGAAGC
CGTTGATGGCCGGGGGACTTGCTGGGGGCCTGCAGGAACACCGCCCCCCGACA
GGAGGGCTGGTGGCGACTCACCTTTCGACCTGAGATCAGGCAAGAACACCCGC
TGAACTTAAGCATATCACTAAGCGGAGGAAAAGAAACTAACCAGGATTCCCC
TAGTAGCGGCGAGCGAACCGGGAAGAGCCCAACTTGAAAATCTCCAGGCCTT
GCCTGGCGAATTGTAGTCTATAGAAGCGCCCTCTGCGGCAGGTCGCGCCCAAG
TCCCCTGGGAGGGGGCGTCAGAGAGGGTGAGAACCCCGTCGGCGCGCGCCTGT
CCGCTCCACGAGGCGCTTTCGCAGAGTCGGGTTGTTTGGGAATGCAGCCCAAA
GCAGGTGGTAAATCCCATCTAAGGCTAAATACTGACGGGAGACCGATAGCGA
ACAAGTACCGTTGAGGGAAAGATGAAAAGAACTTTGAAAAGAGAGTTAAAA
GTGCTTGAAATTGTTGAAGGGGGAAGAGATTGGAAGCCTCGGGTGCCCCAAG
GCTTATGCCGGCCTCACGGCCCGCTGCATGTGCTGGGTGCTGGTCAACAT

- 113 -
>Scenedesmus sp._NIBRIUM-9. 18S ribosomal RNA gene, partial sequence
(1611bp)
CAGTTATAGTTTATTTGGTGGTACCTTACTACTCGGATAACCGTAGTAATTC
TAGAGCTAATACGTGCGTAAATCCCGACTTCTGGAAGGGACGTATATATTAG
ATAAAAGGCCGACCGAGCTTTGCTCGACCCGCGGTGAATCATGATATCTTCA
CGAAGCGCATGGCCTTGTGCCGGCGCTGTTCCATTCAAATTTCTGCCCTATCA
ACTTTCGATGGTAGGATAGAGGCCTACCATGGTGGTAACGGGTGACGGAGGA
TTAGGGTTCGATTCCGGAGAGGGAGCCTGAGAAACGGCTACCACATCCAAGG
AAGGCAGCAGGCGCGCAAATTACCCAATCCTGATACGGGGAGGTAGTGACAA
TAAATAACAATACCGGGCATTTTATGTCTGGTAATTGGAATGAGTACAATC
TAAATCCCTTAACGAGGATCCATTGGAGGGCAAGTCTGGTGCCAGCAGCCGC
GGTAATTCCAGCTCCAATAGCGTATATTTAAGTTGTTGCAGTTAAAAAGCTC
GTAGTTGGATTTCGGGTGGGTTCTAGCGGTCCGCCTATGGTGAGTACTGCTA
TGGCCTTCCTTTCTGTCGGGGACGGGCTTCTGGGCTTCACTGTCCGGGACTCG
GAGTCGACGTGGTTACTTTGAGTAAATTAGAGTGTTCAAAGCAGGCTTACGC
CAGAATACTTTAGCATGGAATAACACGATAGGACTCTGGCCTATCTTGTTGG
TCTGTAGGACCGGAGTAATGATTAAGAGGGACAGTCGGGGGCATTCGTATTT
CATTGTCAGAGGTGAAATTCTTGGATTTATGAAAGACGAACTACTGCGAAA
GCATTGCCAAGGATGTTTTCATTAATCAAGAACGAAAGTTGGGGGCTCGAAG
ACGATTAGATACCGTCGTAGTCTCAACCATAAACGATGCGAACTAGGGATTG
GCGAATGTTTTTTTAATGACTTCGCCAGCACCTTATGAGAAATCAAAGTTTT
TGGGTTCCGGGGGGAGTATGGTCGCAAGGCTGAAACTTAAAGGAATTGACGG
AAGGGCACCACCAGGCGTGGAGCCTGCGGCTTAATTTGACTCAACACGGGAA
AACTTACCAGGTCCAGACATAGTGAGGATTGACAGATTGAGAGCTCTTTCTT
GATTCTATGGGTGGTGGTGCATGGCCGTTCTTAGTTGGTGGGTTGCCTTGTC
AGGTTGATTCCGGTAACGAACGAGACCTCAGCCTGCTAAATAGTCTCAGTTG
CTTTTTGCAGCTGGCTGACTTCTTAGAGGGACTATTGGCGTTTAGTCAATGG
AAGTATGAGGCAATAACAGGTCTGTGATGCCCTTAGATGTTCTGGGCCGCAC
GCGCGCTACACTGATGCATTCAACAAGCCTATCCTTGACCGAAGGGTCTGGGT
AATCTTTGAAACTGCATCGTGATGGGGATAGATTATTGCAATTATTAGTCT
TCAACGAGGAATGCCTAGTAAGCGCAAGTCATCAGCTTGCGTTGATTACGTC
CCTGCCCTTTGTACACACCGCCCGTCGCTCCTACCGATTGGGTGTGCTGGTGA
AGTGTTCGGATTGGCAGCTTAGGGTGGCAACACCTCAGGTCTGCCGAGAA

- 114 -
>Chlorella sp._NIBRIUM-10. 18S ribosomal RNA gene, partial sequence (1631bp)
ACTGCGAATGGCTCATTAAATCAGTTATAGTTTATTTGATGGTACCTACTAC
TCGGATACCCGTAGTAAATCTAGAGCTAATACGTGCGTAAATCCCGACTTCT
GGAAGGGACGTATTTATTAGATAAAAGGCCGACCGGGCTCTGCCCGACTCGC
GGTGAATCATGATAACTTCACGAATCGCATGGCCTTGCGCCGGCGATGTTTC
ATTCAAATTTCTGCCCTATCAACTTTCGATGGTAGGATAGAGGCCTACCATG
GTGGTAACGGGTGACGGAGGATTAGGGTTCGATTCCGGAGAGGGAGCCTGAG
AAACGGCTACCACATCCAAGGAAGGCAGCAGGCGCGCAAATTACCCAATCCTG
ACACAGGGAGGTAGTGACAATAAATAACAATACTGGGCCTTTTCAGGTCTGG
TAATTGGAATGAGTACAATCTAAACCCCTTAACGAGGATCAATTGGAGGGCA
AGTCTGGTGCCAGCAGCCGCGGTAATTCCAGCTCCAATAGCGTATATTTAAG
TTGCTGCAGTTAAAAAGCTCGTAGTTGGATTTCGGGTGGGGCCTGCCGGTCC
CGCCGTTTCGGTGTGCACTGGCAGGGCCCACCTTGTTGCCCGGGACGGGCTCC
TGGGCTTCACTGTCCGGGACTCGGAGTCGGCGCTGTTACTTTGAGTAAATTA
GAGTGTTCAAAGCAGGCCTACGCTCTGAATACATTAGCATGGAATAACACGA
TAGGACTCTGGCCTATCCTGTTGGTCTGTAGGACCGGAGTAATGATTAAGAG
GGACAGTCGGGGGCATTCGTATTTCATTGTCAGAGGTGAAATTCTTGGATTT
ATGAAAGACGAACTACTGCGAAAGCATTTGCCAAGGATGTTTTCATTAATCA
AGAACGAAAGTTGGGGGCTCGAAGACGATTAGATACCGTCCTAGTCTCAACC
ATAAACGATGCCGACTAGGGATCGGCGGATGTTTCTTCGATGACTCCGCCGGC
ACCTTATGAGAAATCAAAGTTTTTGGGTTCCGGGGGGAGTATGGTCGCAAGG
CTGAAACTTAAAGGAATTGACGGAAGGGCACCACCAGGCGTGGAGCCTGCGG
CTTAATTTGACTCAACACGGGAAAACTTACCAGGTCCAGACATAGTGAGGAT
TGACAGATTGAGAGCTCTTTCTTGATTCTATGGGTGGTGGTGCATGGCCGTT
CTTAGTTGGTGGGTTGCCTTGTCAGGTTGATTCCGGTAACGAACGAGACCTC
AGCCTGCTAAATAGTCACGGTTGGCTCGCCAGCCGGCGGACTTCTTAGAGGGA
CTATTGGCGACTAGCCAATGGAAGCATGAGGCAATAACAGGTCTGTGATGCC
CTTAGATGTTCTGGGCCGCACGCGCGCTACACTGATGCATTCAACGAGCCTAG
CCTTGGCCGAGAGGCCCGGGTAATCTTTGAAACTGCATCGTGATGGGGATAG
ATTATTGCAATTATTAATCTTCAACGAGGAATGCCTAGTAAGCGCAAGTCAT
CAGCTTGCGTTGATTACGTCCCTGCCCTTTGTACACACCGCCCGTCGCTCCTA
CCGATTGGGTGTGCTGGTGAAGTGTTCGGATTGGCGACCGGGGGCGGTCTCCG
CTCTCGGCCGCC

- 115 -
NO. Component AmountStock Solution
Concentration
Final
Concentration
1NaNO3
(Fisher BP360-500)10 mL/L 30 g/200 mL dH2O 17.6 mM
2K2HPO4
(Sigma P 3786)10 mL/L 0.8 g/200 mL dH2O 0.23 mM
3MgSO4·7H2O
(Sigma 230391)10 mL/L 1.5 g/200 mL dH2O 0.3 mM
4CaCl2·2H2O
(Sigma C-3881)10 mL/L 0.72 g/200 mL dH2O 0.24 mM
5Citric Acid·H2O
(Fisher A 104)10 mL/L 0.12 g/200 mL dH2O 0.031 mM
6 Ferric Ammonium Citrate 10 mL/L 0.12 g/200 mL dH2O 0.021 mM
7Na2EDTA·2H2O
(Sigma ED255)10 mL/L 0.02 g/200 mL dH2O 0.0027 mM
부록 II. 미세조류 배양용 영양배지 조성
1. BG-11 Medium
Directions (1L Total)
<액체배지>
1. dH20 약 900ml에 BG-11 Trace metal Solution을 제외한 9가지 Components를 계속 교
반하면서 차례대로 넣는다.
2. dH2O로 total volume을 1L로 맞춘다.
3. 배지를 호일로 덮고, 멸균한다.
4. 배지를 식힌 후 냉장 보관한다.
<고체배지>
1. dH2O 약 400mL 계속 교반하면서 순서대로 9 components를 추가한다.
2. dH2O로 total volume을 500ml로 맞춘다.
3. 별도의 용기에 dH2O 500mL, Agar 15g을 넣는다.
4. 두 Solution을 호일로 덮고, 멸균한다.
5. 두 Solution을 항온 수조에 넣고 40-50°C로 식힌다.
6. Agar solution에 살균한 component 10을 넣고 잘 섞는다.
7. Agar, liquid solution을 합치고 잘 섞는다. Agar가 잘 응고되는 것에 주의해야한다.
8. 배지를 식힌 후 냉장 보관한다.

- 116 -
8Na2CO3
(Baker 3604)10 mL/L 0.4 g/200 mL dH2O 0.19 mM
9*BG-11 Trace metal
Solution1 mL/L (see following recipe)
10
Sodium Thiosulfate
Pentahydrate
(agar media only, sterile)
(Baker 3946)
1 mL/L 49.8 g/200 mL dH2O 1 mM
NO. Component AmountStock Solution
Concentration
Final
Concentration
1H3BO3
(Baker 0084)2.86 g/L 46 mM
2MnCl2•4H2O
(Baker 25401.81 g/L 9 mM
3ZnSO4•7H2O
(Sigma Z 0251)0.22 g/L 0.77 mM
4Na2MoO4•2H2O
(J.T. Baker 3764)0.39 g/L 1.6 mM
5CuSO4•5H2O
(MCIB 3M11)0.079 g/L 0.3 mM
6Co(NO3)2•6H2O
(Mallinckroft 4544)49.4 mg/L 0.17 mM
*BG-11 Trace Metals Solution Recipe
Directions (1L Total)
1. dH2O 약 900mL을 계속 교반하면서 순서대로 Components를 추가한다.
2. dH2O로 total volume을 1L로 맞춘다.
3. 냉장 보관한다.

- 117 -
NO. Component AmountStock Solution
Concentration
Final
Concentration
1NaNO3
(Fisher BP360-500)1 mL 7.5 g/100 mL dH20 880 µM
2NaH2PO4·H2O
(MCIB 742)1 mL 0.5 g/100 mL dH20 36 µM
3Na2SiO3·9H2O
(Sigma 307815)1 mL 3 g/100 mL dH20 106 µM
4 *Trace Metals Solution 1 mL/L (see following recipe)
5 *Vitamin B12 1 mL/L (see following recipe)
6 *Biotin Vitamin Solution 1 mL/L (see following recipe)
7*Thiamine Vitamin
Solution1 mL/L (see following recipe)
NO. Component AmountStock Solution
Concentration
Final
Concentration
1ZnSO4•7H2O
(Sigma Z 0251)23 mg/L 0.08 µM
2MnSO4•H2O
(Sigma M8179)152 mg/L 0.9 µM
3Na2MoO4•2H2O
(J.T. Baker 3764)7.3 mg/L 0.03 µM
2. f/2 Medium
Directions (1L Total)
1. 살균되지 않은 해수(30-35ppt) 약 950mL을 계속 교반하면서 순서대로 Components를 추가
한다.(vitamin 제외)
2. 살균되지 않은 해수로 total volume을 1L로 맞춘다.
3. 배지를 호일로 덮고 멸균한다.
4. 식으면 멸균된 Vitamin을 추가한다.
5. 냉장 보관한다.
*Trace Metals Solution Recipe
Directions (1L Total)
1. dH2O 약 950mL에 열을 가하고 교반을 시작한다.
2. 교반하고 있는 물이 끓는점에 도달하면, 나열된 순서대로 Components를 추가한다.
3. dH2O로 total volume을 1L로 맞춘다.
4. 냉장 보관한다.

- 118 -
4CoSO4•7H2O
(Baker 1696)14 mg/L 0.05 µM
5CuCl2•2H2O
(Fisher C-455)6.8 mg/L 0.04 µM
6Fe(NH4)2(SO4)2•6H2O
(Sigma F-1513)4.6 g/L 11.7 µM
7Na2EDTA•2H2O
(Sigma ED255)4.4 g/L 11.7 µM
NO. Component AmountStock Solution
Concentration
Final
Concentration
1HEPES buffer pH 7.8
(Sigma H-3375)
2.4 g/200 mL
dH2O
2
Vitamin B12
(cyanocobalamin,
(Sigma V-6629)
0.027 g/200 mL
dH2O
NO. Component AmountStock Solution
Concentration
Final
Concentration
1HEPES buffer pH 7.8
(Sigma H-3375)2.4 g/200 mL dH2O
2Biotin
(Sigma B-4639)
0.005 g/200 mL
dH2O
*Vitamin B12 Recipe
Directions (200mL Total)
1. 200mL HEPES buffer (50mM)을 준비한다.
2. pH를 7.8로 맞추다.
3. 완전히 녹을 때까지 기다린 후 Vitamin B12(0.1mM)을 추가한다.
4. Sterilize by 0.45 µm Millipore filter. 냉동실 어두운 곳에 보관한다.
*Biotin Vitamin Solution Recipe
Directions (200mL Total)
1. 200mL HEPES buffer (50mM)을 준비한다.
2. pH를 7.8로 맞춘다.
3. 완전히 녹을 때까지 기다린 후 biotin(0.1mM)을 추가한다.
4. Sterilize by 0.45 µm Millipore filter. 냉동실 어두운 곳에 보관한다.

- 119 -
NO. Component AmountStock Solution
Concentration
Final
Concentration
1HEPES buffer pH 7.8
(Sigma H-3375)
H-3375) 2.4
g/200 mL dH2O
2Thiamine
(Sigma T-1270
0.067 g/200 mL
dH2O
*Thiamine Vitamin Solution Recipe
Directions (50mL Total)
1. 50mL HEPES buffer (50mM)을 준비한다.
2. pH로 7.8로 맞춘다.
3. 완전히 녹을 때까지 기다린 후 Thiamine(1mM)을 추가한다.
4. Sterilize by 0.45 µm Millipore filter. 냉동실 어두운 곳에 보관한다.

- 120 -
NO. ComponentStock Solution
Concentrationml/Litre
1 KH2PO4 8.75 g/500 ml 10 ml
2 CaCl2.2H2O 1.25 g/500 ml 10 ml
3 MgSO4.7H2O 3.75 g/500 ml 10 ml
4 NaNO3 12.5 g/500 ml 10 ml
5 K2HPO4 3.75 g/500 ml 10 ml
6 NaCl 1.25 g/500 ml 10 ml
7Na2EDTA
KOH
10 g/L
6.2 g/L1 ml
8FeSO4.7H2O
H2SO4 (conc.)
4.98 g/L
1 ml/L1 ml
9 *Trace Metal Solution (see following recipe) 1 ml
10 H3BO3 5.75 g/500 ml 0.7 ml
NO. Component g/Liter
1 H3BO3 2.86 g
2 MnCl2.4H2O 1.81 g
3 ZnSO4.7H2O 0.222 g
4 Na MoO4.5H2O 0.390 g
5 CuSO4.5H2O 0.079 g
6 Co(NO3)2.6H2O 0.0494 g
3. Modified BOLD BASAL MEDIUM (BBM)
Directions (1L Total)
1. dH20 약 900ml에 BBM Trace metal Solution을 제외한 9가지 Components를 계속 교반
하면서 차례대로 넣는다.
2. dH2O로 total volume을 1L로 맞춘다.
3. 배지를 호일로 덮고, 멸균한다.
4. 배지를 식힌 후 냉장 보관한다.
이 배지는 고농축이고, 많은 녹조류에 사용된다.
*Trace Metal Solution

- 121 -
상기의 Components를 녹이고, 다음 목록에 추가하기 전에 Trace metal solution을 따로 녹여야한다.
배지의 PH는 6.8로 NaOH나 HCl로 조정하고, 멸균한다.
*Reference: Stein, J. (ED.) Handbook of Phycological methods. Culture methods
and growth measurements. Cambridge University Press. 448 pp.

- 122 -
Componentstock solution
(g-L⁻1 dH2O)Quantity used
Medium (M)
Concentration 1n
Final Medium (M)
Na2EDTA 2H2O - 0.8g 2.15×10⁻6FeSO4 7H2O - 0.7g 2.52×10⁻6ZnSO4 7H2O 1 1ml 3.48×10⁻9MnSO4 7H2O 2 1ml 8.97×10⁻9H3BO3 10 1ml 1.62×10⁻7Co(NO3)2 6H2O 1 1ml 3.44×10⁻9Na2MoO4 2H2O 1 1ml 4.13×10⁻9CuSO4 5H2O 0.005 1ml 2.00×10⁻11
4. Modified Spirulina Medium(Aiba and Ogawa 1977, Schlosser 1994)
Directions (1L Total)
1. 두 개의 Solution은 분리해서 멸균하고 저온의 무균상태로 섞는다.
2. Solution Ⅰand Ⅱ을 준비한다.; Solution Ⅱ은 trace metal stock solution 1ml을 포함한다.
3. 그러나 vitamin stock solution은 trace metal stock solution을 포함하지 않는다.
4. Solution Ⅰand Ⅱ는 각각 멸균하고, 식으면 무균상태로 섞는다.
5. Vitamin Solution 1ml을 추가한다.
Component
(g-L⁻¹ dH2O) stock solutionQuantity
used
Concentration 1n Final
Medium (M)
Solution Ⅰ 500ml - -
NaHCO3 - 17.64g 1.62×10⁻¹K2HPO4 - 0.50g 2.87×10⁻3Solution Ⅱ 500ml - -
NaNO3 - 2.5g 2.97×10⁻2K2SO4 - 1.0g 5.74×10⁻3NaCl - 1.0g 1.71×10⁻2MgSO4 7H2O - 0.2g 8.11×10⁻4CaCl2 2H2O - 0.04g 2.72×10⁻4FeSO4 7H2O - 0.01g 3.60×10⁻5Na2EDTA 2H2O - 0.08g 2.15×10⁻4*Trace metal Solution (see following recipe) 1ml
*Vitamins solution (see following recipe) 1ml
(1) 0.4g EDTA and 0.7g FeSO47H2O in 100ml dH2O
(2) 0.4g EDTA and remaning element in 900ml dH2O
*Trace metal solution
dH20 900ml에 EDTA를 녹이고, 나머지 Components를 녹인다. total volume을 1L로 맞춘다.
*Vitamin solution

- 123 -
Componentstock solution
(g-L⁻1 dH2O)Quantity used
Medium (M)
Concentration 1n
Final Medium (M)
cyancobalamin (vitamin B12) - 5mg 3.69×10⁻9
dH20 1L에 cyancobalamin을 녹인다.
필터링을 통한 멸균 및 냉동 보관이 필요하다.
*Reference
Robert A. Andersen, Algal culturing Techniques, Phycological society of America






























