Embed Size (px)
DESCRIPTION
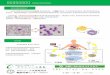
第八章 原核细胞的基因表达调控. 第六节 翻译调控. 转录调控 : 基本上在开始阶段进行,这是为了避免不必要的转录产物的产生所需消耗的能量,不过在伸长阶段没有调控,而在结束阶段则又有调控。. 基因表达的调控. 初级转录产物只有在被加工后所形成的 mRNA 的组成及其机能才被确定,同时 RNA 的二级结构在调控中也非常重要. 翻译调控也同转录调控类似. repressor. 调节因子结合部位. N N N N N N N N N N N A U G N N N N N N N N N N N N. mRNA. 核糖体结合部位. (一)自身调控. 1. 机制. - PowerPoint PPT Presentation
Citation preview

第八章 原核细胞的基因表达调控第八章 原核细胞的基因表达调控
基因表达的调控
转录调控 : 基本上在开始阶段进行,这是为了避免不必要的转录产物的产生所需消耗的能量,不过在伸长阶段没有调控,而在结束阶段则又有调控。初级转录产物只有在被加工后所形成的mRNA 的组成及其机能才被确定,同时RNA 的二级结构在调控中也非常重要
翻译调控也同转录调控类似
第六节 翻译调控

调节因子结合部位
N N N N N N N N N N N A U G N N N N N N N N N N N N
核糖体结合部位
repressor
mRNA
(一)自身调控1. 机制

2. 主要阻遏蛋白的作用部位
阻遏蛋白 靶基因 作用部位
R17 外壳蛋白 R17 replicase
核糖体结合部位的回文结构T4 Reg A 蛋白
初期 T4 mRNA 翻译开始密码在内的序列T4 DNA 聚合酶
T4 DNA 聚合酶
Shine-Dalgarno 序列T4 p32 基因 32 单链 5’前导序列

一个顺反子的翻译需要前一个顺反子翻译所引起的二级结构变化来调控

3. 核糖体蛋白的主要结构
操纵子 基因与蛋白 调节因子
str rps L rps G fus A tuf AS12 S7 EF-G EF-Tu S7
spc rplN rplX rplE rpsN rpsH rplF rplR rpsE rplD rpmO secY-XL14 L24 L5 S14 S8 L6 L18 S5 L30 L15 Y X S8
S10rpsJ rplC rplB rplD rplW rplS rplV rpsC rpsQ rplP rpmC
S10 L3 L2 L4 L23 S19 L22 S3 S17 L16 L29L4
αrpsM rpsK rpsD rpoA rplQ
S13 S11 S4 α L17
L1L11rplK rplA
L11 L1
S4
rif rplJ rplL rpoB rpoCL10 L7 β β’
L10

4. 核糖体蛋白操纵子的自身调控机能
r-protein
rRNA
rRNA 被合成后, r蛋白就与之结合
游离 r 蛋白不存在时,r 蛋白 mRNA 的翻译继续进行
rRNA 的量减少后,游离 r 蛋白蓄积
一个 r 蛋白与mRNA 结合抑制翻译

1.r 蛋白的量对应着细胞的繁殖速度。 rRNA 的量的调控,
使得细胞全部的核糖体组成部分的量能够得到调控2. 由这些操纵子编码的其它蛋白质与 r 蛋白的翻译程度
独立,是以其固有的速度进行合成。

自身调控是限制自身合成的系统,与依存其它因子进行调控对比,后者是阻遏蛋白和操作子的结合能力由外来性低分子物质的浓度进行调控,但是自身调控时,被调控蛋白的自身浓度直接影响着该蛋白的合成。

第七节 RNA 结构调控模型 RNARNA 在外界环境因素的驱使下,或借助碱基互补配对形成多在外界环境因素的驱使下,或借助碱基互补配对形成多
种空间结构,控制转录终止和种空间结构,控制转录终止和 RNARNA 分子的降解;或通过分子分子的降解;或通过分子间相互作用,导致其他间相互作用,导致其他 RNARNA 分子的灭活。分子的灭活。
(( 一一 )) 终止子工作原理:终止子工作原理: 转录终止反应必须破坏转录终止反应必须破坏 RNARNA 链与链与 DNADNA 链之间的所有氢键,链之间的所有氢键,
以便以便 DNADNA 双螺旋结构复原。终止信号存在于已转录出的双螺旋结构复原。终止信号存在于已转录出的RNARNA 序列中,即终止子结构。序列中,即终止子结构。
本征终止子本征终止子 intrinsic terminatorintrinsic terminator :发夹结构和大约:发夹结构和大约 66 个个UU 组成的尾部结构组成的尾部结构
发夹结构导致转录延缓,为最后终止创造有利条件,处于停止发夹结构导致转录延缓,为最后终止创造有利条件,处于停止状态的状态的 RNARNA 聚合酶遇到转录终止信号,即从模板链上解离下聚合酶遇到转录终止信号,即从模板链上解离下来。来。
ρ ρ 因子依赖型终止子:直接识别终止位点上游因子依赖型终止子:直接识别终止位点上游 5050 ~~ 9090 碱基碱基区域,该区域含丰富的区域,该区域含丰富的 CC 碱基,但碱基,但 GG 碱基贫乏。 碱基贫乏。 ρ ρ 因子通常因子通常与终止子上游特异位点结合,沿与终止子上游特异位点结合,沿 RNARNA 链向下游移动,其速度链向下游移动,其速度快于快于 RNARNA 聚合酶速度,赶上延宕的聚合酶速度,赶上延宕的 RNARNA 聚合酶,利用其聚合酶,利用其ATPATP 酶活性,释放能量,解开酶活性,释放能量,解开 DNADNA -- RNARNA杂合双链, 杂合双链, RNARNA 聚合酶也解离下来。聚合酶也解离下来。

可变性二级结构调控衰减作用

TRAP is activated by tryptophan and binds to trp mRNA. This allows the termination hairpin to form, with the result that RNA polymerase terminates, and the genes are not expressed. In the absence of tryptophan, TRAP does not bind, and the mRNA adopts a structure that prevents the terminator hairpin from forming.

Under normal conditions (in the presence of tryptophan) transcription terminates before the anti-TRAP gene. When tryptophan is absent, uncharged tRNATrp base pairs with the anti-TRAP mRNA, preventing formation of the terminator hairpin, thus causing expression of anti-TRAP.

Termination can be controlled via changes in RNA secondary structure that are determined by ribosome movement.

The trp operon consists of five contiguous structural genes preceded by a control region that includes a promoter, operator, leader peptide coding region, and attenuator.

An attenuator controls the progression of RNA polymerase into the trp genes. RNA polymerase initiates at the promoter and then proceeds to position 90, where it pauses before proceeding to the attenuator at position 140. In the absence of tryptophan, the polymerase continues into the structural genes (trpE starts at +163). In the presence of tryptophan there is ~90% probability of termination to release the 140-base leader RNA.

Antisense RNA can be generated by reversing the orientation of a gene with respect to its promoter, and can anneal with the wild-type transcript to form duplex RNA.

A protein that binds to a single-stranded region in a target RNA could be excluded by a regulator RNA that forms a duplex in this region.

By binding to a target RNA to form a duplex region, a regulator RNA may create a site that is attacked by a nuclease.

The secondary structure formed by base pairing between two regions of the target RNA may be prevented from forming by base pairing with a regulator RNA. In this example, the ability of the 3 end of the RNA to pair with the 5 end is prevented by the regulator.