Embed Size (px)
DESCRIPTION
Тема реферата "ФУНКЦИОНАЛЬНАЯ И РЕГЕНЕРАТИВНАЯ АКТИВНОСТЬ НЕЙРОНОВ ПОСЛЕ КРИОКОНСЕРВАЦИИ ИЗОЛИРОВАННОГО МОЗГА МОЛЛЮСКА LYMNAEA STAGNALIS L."
Citation preview

На правах рукописи
Ивличева Наталья Александровна
ФУНКЦИОНАЛЬНАЯ И РЕГЕНЕРАТИВНАЯ АКТИВНОСТЬ НЕЙРОНОВ
ПОСЛЕ КРИОКОНСЕРВАЦИИ ИЗОЛИРОВАННОГО МОЗГА МОЛЛЮСКА LYMNAEA STAGNALIS L.
03.01.02 - биофизика
АВТОРЕФЕРАТ диссертации на соискание ученой степени
кандидата биологических наук
ПУЩИНО – 2013

Работа выполнена в Федеральном государственном бюджетном учреждении науки Институте биофизики клетки Российской академии наук и в Пущинском государственном естественно-научном институте
Научный руководитель: кандидат биологических наук Гахова Эдит Николаевна Официальные оппоненты:
Буданцев Аркадий Юстианович - доктор биологических наук, профессор, Федеральное государственное бюджетное учреждение науки Институт теоретической и экспериментальной биофизики РАН, заведующий лабораторией функциональной гистохимии
Межевикина Людмила Михайловна - доктор биологических наук, Федеральное государственное бюджетное учреждение науки Институт биофизики клетки РАН, ведущий научный сотрудник лаборатории механизмов рецепции
Ведущая организация: Федеральное государственное бюджетное учреждение науки Институт биологии развития им. Н.К. Кольцова Российской академии наук
Защита диссертации состоится «21» февраля 2013 г. в 14 ч. 00 мин. на заседании совета по защите докторских и кандидатских диссертаций Д 002.038.01 при Федеральном государственном бюджетном Учреждении науки Институте биофизики клетки Российской академии наук по адресу: 142290, Московская область, г. Пущино, ул. Институтская, д. 3 С диссертацией можно ознакомиться в Центральной библиотеке НЦБИ РАН по адресу: 142290, Московская область, г. Пущино, ул. Институтская, 3. Автореферат разослан « 21» января 2013 г. Ученый секретарь диссертационного совета кандидат биологических наук Смолихина Татьяна Ивановна

3
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Длительное сохранение жизнеспособной нервной ткани в замороженном состоянии (криоконсервация в жидком азоте, при -196°С) является современной и актуальной проблемой в связи с развитием в последние годы методов трансплантологии в нейромедицине и, соответственно, необходимостью создания запаса нейронального материала в криобанках.
В настоящее время криоконсервация стала обычной процедурой для длительного сохранения многих типов клеток животных, таких как клетки крови, клетки костного мозга, сперматозоиды, клеточные культуры, и др. Успешные протоколы криоконсервирования в основном разработаны для однородных клеточных суспензий (Фуллер и др., 2003; Meryman, 2007; Choi, Bischof , 2010; Seth, 2012). Изолированные ткани плохо переносят низкотемпературную криоконсервацию, а для целых органов эти методы пока не разработаны. Клетки в составе ткани, создавая определенную архитектуру функциональной единицы, более плотно упакованы, чем в клеточных суспензиях, с которыми исследователи чаще всего имеют дело. Гетерогенный клеточный состав нейрональной ткани (разного типа нейроны и глиальные клетки) и экстраклеточная структура, - все вместе могут создавать проблемы при попытке экстраполировать протоколы криоконсервирования с клеточных суспензий на кусочки органов, ткани, нейросферы (Fagy et al., 2004; Pegg, 2007; Paynter, 2008). Одна из таких проблем заключается в том, что экстраклеточные структуры и межклеточные связи могут быть не менее важными объектами криоконсервирования, чем сами клетки, особенно, если они плотно упакованы в структуре ткани (Acker et al., 2000; Müller-Schweinitzer, 2009; Pegg, 2010). Поэтому в настоящее время являются особенно актуальными исследования механизмов криоповреждений и криозащиты нервной ткани как неделимой интегрированной структуры.
В этой связи в качестве модельных объектов интерес представляют нервные системы беспозвоночных, как например, мозг моллюска Lymnaea stagnalis L., который представляет собой гетерогенную ткань, включающую полифункциональные нейроны разного размера и обширные вненейрональные области, по многим свойствам сходные с аналогичными структурами позвоночных. Хорошо отлаженные способы регистрации электрической активности нейронов, разработанные методы клеточной и органной культуры (Гелетюк и др., 1970; Костенко и др. 1972; Wong et al., 1981; Ивличева, Гахова, 2007), а также доступность объекта для исследователей в любое время года, делают мозг моллюска привлекательной моделью для исследования в области криобиологии (Гахова и др., 1989; Дмитриева, 2004; Ивличева, Гахова, 2007). Изучение особенностей криоповреждений и криозащиты мозга моллюска как целой интегральной структуры позволит определить области криоповреждений, прогнозировать степень восстановления и оценить возможности режимов криоконсервации и создания оптимальных криозащитных сред.
Цель и основные задачи исследования. Цель данной работы заключалась в изучении последствий криоконсервации на функциональную и регенеративную способность зрелых нейронов на примере криоконсервирования изолированного мозга моллюска Lymnaea stagnalis L.
В соответствии с целью были поставлены и решены следующие задачи:
1. Изучить электрическую активность нейронов переживающего мозга моллюска в органной культуре без криоконсервации и после криоконсервации: мембранные потенциалы (МП), генерацию потенциалов действия (ПД) и постсинаптических потенциалов (ПСП), рецепторные ответы на медиаторы.

4
2. Провести поиск синаптических связей между нейронами переживающего мозга моллюска в органной культуре после криоконсервации. 3. Исследовать в клеточной культуре выживаемость и регенеративные способности нейронов, выделенных из замороженно-оттаянного мозга моллюска. 4. Выявить характер протективного действия (крио- или нейро- протекторное) пептида Thr-Ser-Lys-Tyr (TSKY) выделенного из мозга зимоспящих сусликов Spermophillus undulates. Определить его влияние на выживаемость и регенеративные способности нейронов в культуре на разных этапах криоконсервирования. 5. Проанализировать влияние пептида TSKY на кристаллизацию и формирование микрочастиц льда в криозащитных растворах в процессе замораживания до температуры жидкого азота.
Научная новизна. Важными достижениями работы являются результаты по изучению электрической активности нейронов переживающего мозга моллюска в органной культуре до и после криоконсервации, поскольку замораживанию подвергался интактный орган, состоящий из разнородной популяции дифференцированных клеток со сложными межклеточными связями и синаптическими структурами. Впервые показано, что на фоне сохранения целостности основной массы нейронов криоповреждения были связаны с межнейрональными областями, а именно с аксонами и синаптическими структурами. Нейроны, выделенные из криоконсервированных ганглиев, как показано в культуре in vitro, обладали способностью к регенерации нервных отростков и образованию контактов. Впервые показано, что пептид TSKY, выделенный из мозга зимоспящих животных, может участвовать в восстановительных процессах в период оттаивания и оказывать нейропротекторное, положительное воздействие, на жизнеспособность нейронов после криоконсервации, а не криопротекторное на этапе замораживания. Криомикроскопические исследования подтверждают отсутствие у пептида TSKY криопротекторных свойств, так он не влиял на характер кристаллизации и формирование микрочастиц льда при замораживании растворов.
Научно-практическое значение работы. Результаты работы вносят вклад в изучение механизмов криопровреждений и криозащиты тканей (и целых органов) животных при криоконсервации. Полученные знания о роли нейропротекторных факторов при гипотермии и при глубоком замораживании изолированных нейрональных тканей позволит продолжить поиск новых эффективных и безопасных способов криоконсервации нервных клеток и ткани. Результаты проведенных исследований могут быть использованы для модификации и разработки криозащитных сред, повышающих жизнеспособность, нейрональной ткани, нейросфер, кусочков мозга.
Конкурсная поддержка. РФФИ грант №10-04-01319-а; грант конкурса УМНИК, гос. контракт № 6639р/9106 от 02.03.09 и № 8229р/12633 от 06.07.10; АФГИР (США, CRDF) проект RUB 2-010001-PU-05; р2004Наукоград-а № 04-04-9730.
Апробация диссертации. Основные материалы исследований были представлены на 8-й, 9-й, 11-й, и 13-й пущинской школе-конференции молодых ученых «Биология – наука XXI века» (Пущино 2004, 2005, 2007, 2009гг.), на Всероссийской конференции молодых исследователей «Физиология и медицина» (Санкт-Петербург, 2005), VIII (Kazan, 2006) и IX (St. Petersburg, 2009) East European Conference of the International Society for Invertebrate Neurobiology «Simpler Nervous Systems», XIХ Международной конференции-совещании «Консервация генетических ресурсов» (Пущино, 2008), III Всероссийском Конгрессе с международным участием студентов и аспирантов-биологов «Симбиоз-Россия 2010», Школе-конференции «Инновационные подходы в изучении биосистем» (Нижний Новгород,

5
2010), Международной конференции «Рецепторы и внутриклеточная сигнализация» (Пущино, 2011), VII и VIII Международном междисциплинарном Конгрессе «Нейронаука для медицины и психологии» (Судак, Украина, 2011, 2012), 14th International Hibernation Symposium «Living in a seasonal world: Thermoregulatory and metabolic adaptations» (Semmering, Austria, 2012 ), IV Съезде биофизиков России (Нижний Новгород, 2012).
Публикации. Основные результаты диссертации представлены в 20 печатных работах, из них 5 в
рецензируемых журналах, рекомендуемых ВАК; 1 – глава в книге; 2 – в сборнике; 12 тезисов.
Структура и объем диссертации. Диссертационная работа изложена на 93 страницах машинописного текста с
использованием 26 рисунков 2 таблиц и включает: введение, обзор литературы, материалы и методы исследования, полученные экспериментальные данные и их обсуждение, заключение и выводы. Список литературы содержит 159 ссылок.
Список принятых сокращений. МП – мембранный потенциал; ПД – потенциал действия; ВПСП – возбуждающий постсинаптический потенциал; ТПСП – тормозной постсинаптический потенциал; НС – нервная система (мозг) моллюска; МКЧЛ – микрочастица льда; L-15 – среда для культивирования (Leibowitz); КСНП – культуральная среда для нейронов прудовика; TSKY – пептид Thr-Ser-Lys-Tyr; ДМСО – диметилсульфоксид.
МАТЕРИАЛЫ И МЕТОДЫ
Объект. Эксперименты проводили на ганглиях окологлоточного кольца (мозге) пресноводного моллюска Lymnaea stagnalis. Моллюсков собирали с весны до осени в естественных водоемах обитания. Собранных в природе прудовиков содержали в аквариумах при 4°С (перед экспериментом, их выдерживали сутки при комнатной температуре, 18-20°С). Кроме этого, моллюсков выращивали из оплодотворённой икры и содержали в аквариумах при 18-20°С.
Криоконсервация: метод замораживания-оттаивания изолированного мозга моллюска. Выделение мозга проводили при комнатной температуре. В качестве криопротектора использовали 2М диметилсульфоксид (ДМСО). Замораживали ганглии в парах жидкого азота со скоростью 400°С/мин. Материал сохраняли в жидком азоте от 24 ч. до 2 лет. Оттаивание осуществляли на водяной бане при 22-24°С со скоростью 500°С/мин. После оттаивания и постепенного отмывания криопротектора для изучения восстановления структуры и функции нейронов мозг помещали в физиологический раствор (мМ): 80 NaCl, 1.6 KCl; 1 MgCl2, 2 CaCl2, 7.5 pH и инкубировали при 4-6°С до 15-18 час, но не менее 1.5-2 час (Гахова и др, 1989).
Метод органотипического культивирования мозга моллюсков. Изолированный мозг моллюска (ганглии), предварительно освободив от толстой соединительно-тканной оболочки, помещали в 2-4 мл питательной среды, применяемой при культивировании изолированных нейронов. Ганглии находились в среде, не прикрепляясь к поверхности стекла. Флаконы хранили в темноте при 4-6°С. Смену среды проводили по мере того, как происходило ее закисление (от 2-3 до 6-9 сут). Обеззараживание осуществляли каждый раз при смене среды в растворе, содержащим 50 мкг/мл гентамицина. Культивирование ганглиев моллюска после криконсервации проводили таким же образом. Регистрацию электрических характеристик нейронов ганглиев проводили через каждые 2-3 сут. на протяжении 16 сут. культивирования. Срок культивирования в экспериментах составлял от 3-х до 16 сут.

6
Культивирование изолированных нервных клеток моллюсков осуществляли по методу, разработанному д.б.н. М.А. Костенко для нервных клеток моллюсков (Костенко, 1972, 1982). Сразу после изолирования мозг выдерживали при 4-6°С от 2 до 18 час в растворе, используемом для длительного переживания нервной ткани моллюсков (мM): 90 NaCl, 5 KCl, 2 CaCl2, 1.5 MgCl2, 7.8 pH (Костенко, 1985). Изолированное кольцо ганглиев подвергали ферментативной обработке (0.35% проназы на физиологическом растворе), отмывали от фермента и механическим образом (многократным пипетированием) выделяли клетки (Костенко и др., 1982). Культивирование осуществляли при комнатной температуре на питательной среде с антибиотиком (гентамицин 20 мкг/мл): 90 NaCl, 5 KCl, 2 CaCl2, 1.5 MgCl2, 0.25 trisHCl, без добавления сыворотки, но содержащей 20% L-15 (Sigma, США), 7.6-7.9 рН.
Жизнеспособность нейронов в клеточной культуре оценивали с помощью светового микроскопа МБИ-13, микроскопа Carl Zeiss Axiovert 40 CFL с настройками по Хоффману, и Olympus IX-70 по следующим морфологическим критериям: в норме нейроны имеют чёткие контуры и равномерную структуру; повреждённые нейроны имеют ярко окрашенное и/или диффузно окрашенное ядро и цитоплазму. Дегенерировавшие нейроны легко определялись по явным признакам лизиса, вакуолизации цитоплазмы или темно-бурой окраске пигмента. Регенерационную активность оценивали по доле клеток, сформировавших отростки в первые 24 ч. культивирования, от общего числа прикрепившихся к стеклу нейронов. Целостность клеточных мембран нейронов оценивали по проницаемости для витального красителя (метиленовый синий). Культивировали при 20°С. Контролем служили клетки животных, мозг которых не подвергался криоконсервации.
Жизнеспособность нейронов в интактном мозге моллюска оценивали методом прижизненного окрашивания Live/Dead. Для прижизненного окрашивания использовали Live/Dead Cell Double Staining Kit: (Sigma, США L-3224, Molecular Probes®) (Vasudevan et al., 2008). Компонент А – кальцеин АМ (λ ex/em ~ 495/~ 515 nm); компонент В – этидийгомодимер-1 (λ ex/em ~ 528/~ 617 nm). Образцы инкубировали 30 мин при 20-22°С в физиологическом растворе, содержащим 4мМ компонента А, тестирующего внутриклеточную эстеразную активность и 3мМ компонента В, тестирующего целостность клеточной мембраны. Флуоресценцию нейронов регистрировали на микроскопе Leica CTR DM 6000B при увеличении 2.5х, 20х и 40х.
Метод внутриклеточного отведения мембранных потенциалов. Вырезанное кольцо окологлоточных ганглиев вместе с буккальными ганглиями обрабатывали смесью ферментов (от 5 до 25 мин): проназа Е (Sigma, США) – 2мг/мл, коллагеназа (Sigma, США) – 2 мг/мл, трипсин – 2 мг/мл, с последующей отмывкой физиологическим раствором для моллюсков. Состав раствора (мМ): 50 NaCl, 1.6 KCl; 4 CaCl2, 1.5 MgCl2, 10 trisHCl, 7.4 pH. Затем ганглии помещали в рабочую камеру объёмом 1.5 мл без протока. Мембранный потенциал нейрона отводили внутриклеточным стеклянным микроэлектродом R=20-60 МОм, заполненным KCI (0.08-3 М) на усилитель производства фирмы Микромед и через аналого-цифровой преобразователь записывали на компьютер IBM PC 486DX4-S. Отведение осуществляли от нескольких нейронов, используя до 5-ти каналов одновременно. В электрофизиологических экспериментах было использовано более 60 изолированных нервных колец, и проанализировано свыше 300 записей нейронной активности мозга как после замораживания-оттаивания, так и не прошедших криоконсервацию. Внутриклеточное отведение мембранных потенциалов осуществляли от наиболее крупных идентифицированных нейронов: ПБ4, ПБ4-кл, ЛБ4, ЛБ4-кл, ППеД1, ЛПеД1, нейронов ППеА-кластера, ЛПеА-кл, ВД1, ВД4, ВА-кл. Серотонин (5-гидрокси-L-триптамин, 5-НТ) (Sigma, США) вносили в камеру микропипеткой или с протоком.

7
Обработка мозга пептидом TSKY. Пептид TSKY был выделен из мозга зимоспящих сусликов Spermophillus undulatus, затем синтезирован в лаборатории химии пептидов ФГБУН ИБХ РАН (Ziganshin et al, 1994). Изолированное окологлоточное нервное кольцо прудовика предварительно инкубировали 60 мин в растворе питательной среды (20% L-15) (Sigma, США) с добавлением пептида TSKY в концентрациях (1·10-7 М, 1·10-6 М и 1·10-5 М) при 22-24°С и при 4-6°С. Нейроны выделяли из ганглиев, как описано выше, затем культивировали при 22-24°С в КСНП. В контроле предварительную инкубацию мозга проводили в растворе без пептида. При криоконсервации мозга прудовика, пептид TSKY добавляли в раствор криопротектора и в отмывающий раствор в концентрации 1·10 -5М.
Криомикроскопия. Криопротектирующие растворы различной концентрации охлаждали в камере Фукса-Розенталя (объем образца 3.2 мкл, толщина слоя 200 мкм), помещённой в специально разработанную пенопластовую камеру, сначала в течение 15 мин в парах жидкого азота, затем после достижения -130°С в жидкий азот, со скоростью охлаждения от -5 до -25°С/мин. Температуру образцов измеряли непосредственно в замораживаемом растворе с помощью электронного термометра АТТ-2006 (Aktakom, Taiwan) с микротермопарой медь-константан (Copper/Constantan microthermocouple) и диаметром 0.1 мм. Наблюдения проводили в отражённом свете при помощи криомикроскопа Orthoplan (Zeiss, Germany) с использованием объектива Planachromat 4x/0.10 (Андреев и др., 2008). Фоторегистрацию изображений криомикрочастиц растворов осуществляли при температуре жидкого азота (-196°C) (Андреев и др., 2009). Видеорегистрацию проводили при помощи видеокамеры НВ-35 (Россия).
Методы оценки ошибок измерений и программное обеспечение. Для записи сигналов нервных клеток была использована программа Spike-C3 (автор Воронцов Д.Д.). Все данные представлены как средние значения и их стандартные ошибки. При проверке статистической достоверности различий между группами сравнения использовали непарный t-критерий Стьюдента для малых выборок.
ОСНОВНЫЕ РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
1. Выживаемость нейронов в клеточной культуре после криоконсервации мозга с использованием ДМСО и криомикроскопия протектирующих растворов
Изучение криоконсервации изолированного мозга моллюска с использованием разных концентраций ДМСО (1М, 2М и 3М) показало, что лучшая сохранность нейронов обеспечивается ДМСО в концентрации 2М (рис. 1). В этом варианте эксперимента в клеточной культуре выживало 78% нейронов, изолированных из заморожено-оттаянного мозга, в то время как с использованием других концентраций ДМСО (1М и 3М) живыми были 38% и 3% нейронов, соответственно (p < 0.001).
Известно, что сохранность клеток при замораживании и хранении в жидком азоте в первую очередь зависит от механических повреждений, связанных с характером кристаллизации, а также с размерами и формой микрочастиц льда (МКЧЛ). Крупные кристаллы и МКЧЛ, имеющие формы с острыми углами, как правило, являются причиной механических повреждений клеток. Одно из основных свойств криопротекторов – это изменение характера кристаллизации растворов. Криомикроскопический анализ растворов с разной концентрацией ДМСО показал, что при замораживании раствора с 2М ДМСО размеры МКЧЛ (при -196°С) не превышали 50 мкм (от 10 до 50 мкм) и имели преимущественно овальную форму, без острых углов, со сглаженными границами, или форму «розочек».

8
Рис. 1. Зависимость количества живых нейронов в культуре, выделенных из мозга после криоконсервации, от концентрации ДМСО, % * - достоверная разница между числом живых нейронов в контроле и после криоконсервации мозга моллюска с разными концентрациями ДМСО, р < 0.001.
В то же время, при замораживании растворов с 1М ДМСО МКЧЛ, образовавшиеся при температурах ниже -140°С, имели размеры от 25 до 100 мкм, а с 3М ДМСО – преобладали МКЧЛ размером более 50 мкм. Крупные МКЧЛ имели неправильную форму с четкими ровными границами, перемежающимися с острыми углами.
Таким образом, можно предположить, что лучшая сохранность нейронов при замораживании мозга с 2М ДМСО связана с формированием при этой концентрации используемого криопротектора менее травматичных для клеток МКЧЛ. Поэтому в наших опытах в качестве криопротектора мы использовали 2М ДМСО.
2. Морфофункциональное состояние нейронов в ганглиях моллюска после криоконсервации в жидком азоте (тестирование жизнеспособности с применением
витального красителя kit Live/Dead)
Для изучения последствий криоконсервации на изолированный мозг моллюска, как комплексной многоклеточной системы, прежде всего, было проведено тестирование на определение живых и мертвых нейронов в оттаянных интактных ганглиях. Для этой цели использовали 2-х компонентный флуоресцентный витальный краситель kit Live/Dead (Molecular Probes®). Флуоресценция клеток в зеленом диапазоне спектра (λ ex/em ~495/~515 nm) по всей поверхности ганглиев свидетельствовала о том, что окрашенные нейроны были живыми. Кроме этого, зеленая флуоресценция была тестом на наличие эстеразной активности в телах нейронов (Vasudevan et al., 2008). Нервные отростки, наблюдаемые на поверхности ганглиев (под ганглионарной оболочкой) также флуоресцировали зеленым, что говорит о сохранении эстеразной активности не только в телах нейронов, но и в нейрональных отростках.
Следующим шагом было изучение физиологической сохранности (или восстановления) не отдельных нейронов, а всей системы мозга после криоконсервации. Мы изучили электрические характеристики нейронов в системе интактного мозга до и после замораживания-оттаивания так как электрическая активность является основным показателем физиологической активности нервной системы.
3. Электрические характеристики нейронов переживающего мозга моллюска в органной культуре
Ранее было показано, что после криоконсервирования изолированного мозга моллюска Lymnaea stagnalis нейроны восстанавливают электрические характеристики (Гахова и др., 1989; Чекурова и др., 1987; Чекурова, 1992). Следует подчеркнуть, эти данные были получены на нейронах, изолированных из заморожено-оттаянного мозга моллюска. Процедура выделения нейронов из стромы ганглиев включает ферментативную и механическую обработки, которые тоже могут вызывать повреждения. Отделить эти повреждения от тех, которые являются результатом физико-химических процессов,

9
сопровождающих процесс криоконсервации, оказалось невозможным. Кроме этого, неизвестно распространяются ли эти повреждающие эффекты в процессе замораживания-оттаивания мозга на связи между нейронами в ганглиях (контакты, рецепторные структуры). Ответы на эти вопросы можно было получить при изучении электрофизиологических свойств нейронов в составе ганглиев мозга в переживающей органной культуре, а не на изолированных нейронах. Для этого мы исследовали переживание изолированного мозга моллюска в культуре in vitro в течение 16 сут культивирования до и после замораживания.
3.1. Электрические характеристики нейронов ганглиев моллюска в органной культуре без криоконсервации
Микроэлектродным методом отведения биопотенциалов было проанализировано 66 клеток в 22 нервных системах (НС). Для анализа электрических характеристик МП и ПД выбирали нейроны в области расположения идентифицированных нейронов (рис. 2): в буккальных ганглиях – клетки в области локализации нейронов Б1, Б2, Б3, Б4; в висцеральном ганглии – в области нейронов ВД1 – ВД4 и А-кластера; в педальных ганглиях – нейроны в области А- кластера и клетки Д1; в правом и левом париетальных ганглиях – группы клеток с вентральной стороны ганглиев; в церебральных ганглиях – в области нейрона Ц1.
Рис. 2. Схема расположения идентифицированных нейронов, электрические показатели которых использовали для оценки жизнеспособности мозга Lymnaea stagnalis (вид с дорзальной стороны).
Отмечены нейроны ЛБ1, ЛБ2, ЛБ3, ПБ4 в левом и правом буккальных ганглиях; ВА-кл - группа нейронов в области А-кластера и нейроны ВД1 и ВД4 в висцеральном ганглии; ПеА-кл - группа нейронов в области А-кластера и нейроны ЛПеД1 и ППеД1 в левом и правом педальных ганглиях; ЛЦеЦ1 ПЦеЦ1 нейроны в левом и правом церебральных ганглиях (обозначения и классификация нейронов приведены согласно работам Benjamin & Winlow, 1981; Syed et al, 1991). Овалами обозначены идентифицированные нейроны.
Из 66-ти исследованных нейронов электрические характеристики зарегистрировали у 57 нейронов (86.3%). В первые сутки органотипического культивирования МП нейронов варьировали в зависимости от конкретной клетки от -40.8 до -61.5 мВ (n = 56), амплитуды ПД – от 56.6 до 93.7 мВ; частота генерации ПД (одиночных и в пачках) варьировала от 23.4 до 164.6 импульсов в минуту, что соответствует частоте генерации от 0.39 до 2.7 Гц.
При длительном органном культивировании мозга моллюска у 17 из 57 активных нейронов (29.8%) были зарегистрированы постсинаптические потенциалы от 4 до 10 мВ. Важно отметить, что в органной культуре мозга моллюска были выявлены синаптические связи как в первые, так и в последующие 16 сут культивирования. Например, на 12 сут культивирования в ответ на деполяризацию (+1нА, 0,5 мсек) нейрона ППеД1 нейрон ВА-кл отвечал тормозными постсинаптическими потенциалами.
Кроме того, мы получили ответы клеток на аппликацию серотонина (3·10-5 М, 4·10-6 М), что свидетельствует о сохранности рецепторной активности нейронов мозга. Серотонин

10
(3·10-5 М) вызывал угнетение спонтанной генерации ПД нейрона ППеД1 правого педального ганглия и активацию электрической активности у нейрона А-кластера левого педального ганглия (рис. 6А).
Таким образом, на полученной модели переживающей органной культуры in vitro в течение 16 сут культивирования мы наблюдали функционирование изолированного мозга моллюска. Данная модель использовалась нами далее в качестве контроля для изучения последствий криоконсервации на уровне интегрированной НС.
3.2. Изучение электрических характеристик нейронов в органной культуре после криоконсервации изолированного мозга моллюска
Ранее было показано, что электрические характеристики нейронов восстанавливаются приблизительно через 2-2.5 ч после оттаивания мозга моллюска и инкубирования его в физиологическом растворе при 4-8°С (Гахова и др., 1989). В нашей работе регистрацию электрических параметров нейронов проводили в первые сутки, не ранее, чем через 8 ч после оттаивания криоконсервированного мозга, отмывания его от криопротектора и инкубирования в питательной среде.
Рис. 3. Генерация потенциалов действия нейронов после криоконсервации в первые сутки культивирования.
Нейрон ВД4 генерирует ПД. а) и б) – паттерны пачечной активности 2-х нейронов правого париетального (ППа) ганглия; б) – генерация ПД и ВПСП нейронов ППа.
В переживающем заморожено-оттаянном мозге прудовика было изучено 57 нейронов из 20 НС. Из них у 47 нейронов (84%) были зарегистрированы электрические характеристики.
Рис. 4. Периодическая пачечная активность ЛБ4 и ЛБ3 буккального ганглия и ПСП нейрона ЛБ4-кл заморожено-оттаянного мозга прудовика в органной культуре, 12-16 сут.

11
На рис. 3 даны примеры записи электрической активности нейронов Д4 висцерального ганглия и двух нейронов из группы клеток с вентральной стороны правого париетального ганглия мозга прудовика в первые сутки культивирования после оттаивания. В первые сутки органотипического культивирования МП разных нейронов в ганглиях варьировал от -41 до - 55.7 мВ (n = 44), амплитуды ПД – от -76.9 до -92.3 мВ; частота генерации ПД (одиночных и в пачках) варьировала от 4.5 до 72.5 импульсов в минуту, что соответствует от 0.74 до 1.2 Гц. При культивировании мозга моллюска у 19 из 47 нейронов (40.4%) были зарегистрированы постсинаптические потенциалы от 4 до 10 мB (рис. 3, рис. 4). Таким образом, количество живых функционально активных клеток не отличается в ганглиях до и после криоконсервации. Однако в разных опытах при сравнении идентичных пар нейронов в разных препаратах (n = 12, по 2 - 3 пары нейронов в каждом препарате) выявляются отличия между контрольными и криоконсервированными клетками. На рис. 5 даны в качестве примера иллюстрации электрической активности двух пар нейронов: ППеД1 - на 5 и 10 сут культивирования, и нейрона ВД4 - на 16-е сут органного культивирования.
Рис. 5. Электрическая активность нейронов в органной культуре на 5, 10 и 16 сут: ППеД1 и ВД4 без криоконсервации (контр) и аналогичных нейронов после криоконсервации (крио-).
Нейрон Д1, который, как известно, является дофаминэргическим интернейроном, обеспечивающим запуск респираторного ритма (Syed et al., 1990), после криоконсервации не проявляет типичной для него спонтанной электрической активности, но в то же время у него наблюдается ПСП-активность такая же, как в контрольных образцах. С другой стороны, интернейрон Д4 висцерального ганглия, который также входит в состав генератора респираторного ритма, обеспечивая работу пневмостома моллюска (Syed, Winlow, 1991b), после замораживания-оттаивания на 16 сут культивирования не только сохраняет спайковую активность, но и увеличивает частоту генерации ПД, а также проявляет пачечную импульсную активность в отличие от идентичного нейрона в контроле.
При культивировании заморожено-оттаянного мозга моллюска удалось зарегистрировать ответы нейронов на серотонин. На рис. 6Б приведены примеры, где серотонин в концентрации (3·10-5 М, 4·10-6 М) вызывал снижение частоты генерации ПД нейрона ППеД1 и одного из нейронов А-кластера левого педального ганглия, а в некоторых случаях ингибировал генерацию ПД. В отличие от контроля ответная реакция криоконсервированных нейронов на серотонин, как правило, имела более длительный временной интервал.

12
Рис. 6. Реакция нейронов мозга моллюска (10 сутки культивирования) на добавление серотонина. А) – мозг без криоконсервации, Б) – мозг после оттаивания. Маркированы начало подачи серотонина (+5-НТ) в проток и начало подачи отмывающего физраствора (-5-НТ). Серотонин (3·10-5 М) вызывал угнетение спонтанной генерации ПД дофаминового нейрона ППеД1 и нейрона ППеА-кл.
Далее мы анализировали возможность сохранения синаптических связей между нейронами, локализованными в разных ганглиях. С этой целью мы изучили связь между нейроном ППеД1 (педальный ганглий) и нейронами из висцерального ганглия (ВД1, ВД4, А-кластера). Известно, что эти нейроны являются основой функционирования нейрональной сети, отвечающей за респираторное и пищевое поведение моллюска (Syed et al., 1991).
После криоконсервации синаптические связи между нейронами выявить не удалось (рис. 7Б), в отличие от контроля (рис. 7А). Анализировали возможность связи в каждом из 20 замороженно-оттаянных НС. Как можно видеть (рис. 7Б), электрически вызванная деполяризация нейрона ППеД1 (+02 нА, 40 мсек) не вызывала синаптических ответов нейронов висцерального ганглия. Также, не выявлен ответ нейрона ППеД1 и нейрона ВД4 на электрически вызванную деполяризацию (+0,4 нА, 40 мсек) нейрона из ВА-кл. В тоже время, отсутствовали импульсные ответы нейрона из ВА-кл на вызванную деполяризацию (+0,1 нА, 40 мсек) нейрона ППеД1.
Таким образом, после криоконсервации ганглиев на фоне электрически активных нейронов (наличие МП, ПД и реакции на аппликацию серотонина) обнаружено отсутствие функциональных синаптических связей между ними. Это могло быть следствием повреждений, вызванных либо физическими воздействиями, сопровождающими процесс криоконсервации, либо токсическим влиянием ДМСО. Известно, что криопротекторы, в том числе ДМСО, обладают не только криозащитными свойствами, но могут оказывать токсическое действие на клетки и клеточные системы (Fahy, 2010).

13
Рис. 7. Пример тестирования синаптической связи в мозге моллюска в первые сутки органотипического культивирования до- и после- криоконсервации.
Чтобы исключить повреждающий эффект ДМСО на нервные отростки и синаптические структуры было изучено влияние ДМСО на наличие синаптической связи между нейронами ППеД1 и ВД4 в образцах, не подвергавшихся криоконсервации. На рис. 8 видно, что 2М ДМСО не влиял на функционирование синаптической связи: электрическая стимуляция (+1нА, 0,5 мсек) нейрона ППеД1 вызывала ВПСП ответ нейрона ВД4.
Рис. 8. Пример функционирования синаптической связи в мозге моллюска после обработки 2М ДМСО (без криоконсервации).
Таким образом, отсутствие синаптических связей в заморожено-оттаянных ганглиях было зафиксировано в первые сутки органотипического культивирования. Связи не восстанавливались в течение 16 сут культивирования. Такой характер повреждения возможен на уровне нейропиля в ганглиях или в комиссурах, соединяющих ганглии. Отсутствие связей при увеличении сроков культивирования может говорить о том, что для регенерации отростков и восстановления синаптических структур после криоконсервации требуется более 16 сут, либо свидетельствовать о необратимости повреждений.
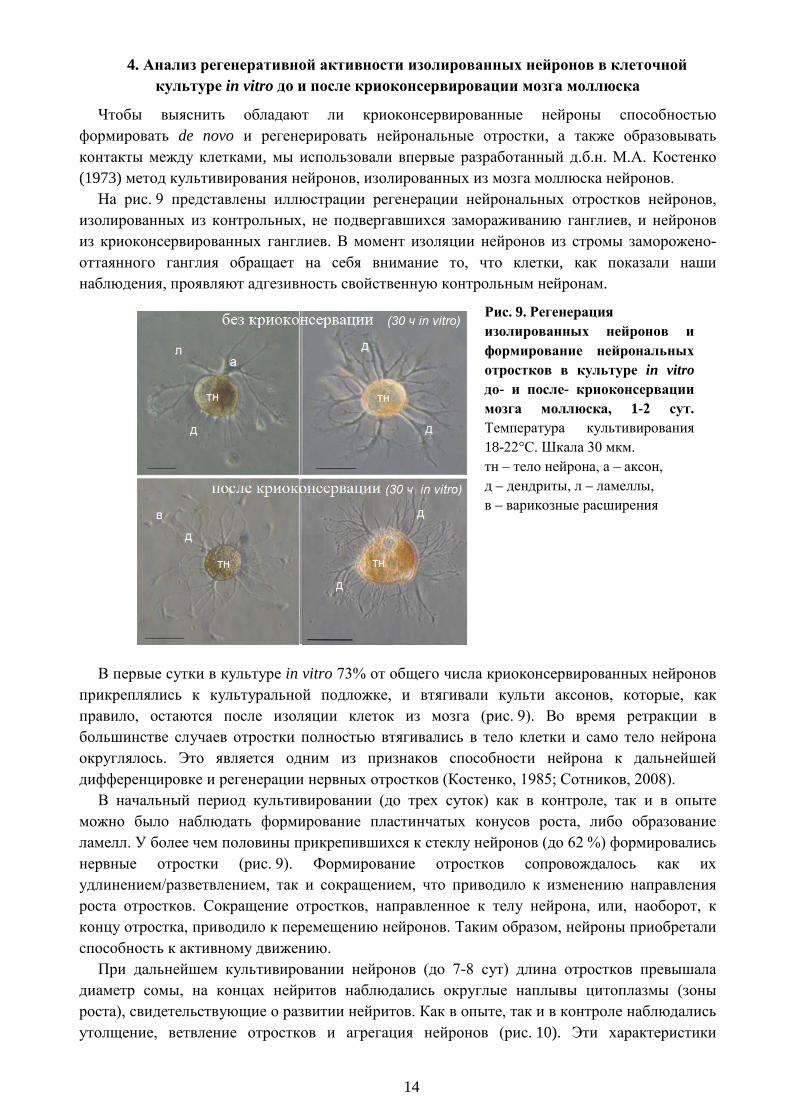
14
4. Анализ регенеративной активности изолированных нейронов в клеточной культуре in vitro до и после криоконсервировации мозга моллюска
Чтобы выяснить обладают ли криоконсервированные нейроны способностью формировать de novo и регенерировать нейрональные отростки, а также образовывать контакты между клетками, мы использовали впервые разработанный д.б.н. М.А. Костенко (1973) метод культивирования нейронов, изолированных из мозга моллюска нейронов.
На рис. 9 представлены иллюстрации регенерации нейрональных отростков нейронов, изолированных из контрольных, не подвергавшихся замораживанию ганглиев, и нейронов из криоконсервированных ганглиев. В момент изоляции нейронов из стромы заморожено-оттаянного ганглия обращает на себя внимание то, что клетки, как показали наши наблюдения, проявляют адгезивность свойственную контрольным нейронам.
Рис. 9. Регенерация изолированных нейронов и формирование нейрональных отростков в культуре in vitro до- и после- криоконсервации мозга моллюска, 1-2 сут. Температура культивирования 18-22°С. Шкала 30 мкм. тн – тело нейрона, а – аксон, д – дендриты, л – ламеллы, в – варикозные расширения
В первые сутки в культуре in vitro 73% от общего числа криоконсервированных нейронов прикреплялись к культуральной подложке, и втягивали культи аксонов, которые, как правило, остаются после изоляции клеток из мозга (рис. 9). Во время ретракции в большинстве случаев отростки полностью втягивались в тело клетки и само тело нейрона округлялось. Это является одним из признаков способности нейрона к дальнейшей дифференцировке и регенерации нервных отростков (Костенко, 1985; Сотников, 2008).
В начальный период культивировании (до трех суток) как в контроле, так и в опыте можно было наблюдать формирование пластинчатых конусов роста, либо образование ламелл. У более чем половины прикрепившихся к стеклу нейронов (до 62 %) формировались нервные отростки (рис. 9). Формирование отростков сопровождалось как их удлинением/разветвлением, так и сокращением, что приводило к изменению направления роста отростков. Сокращение отростков, направленное к телу нейрона, или, наоборот, к концу отростка, приводило к перемещению нейронов. Таким образом, нейроны приобретали способность к активному движению.
При дальнейшем культивировании нейронов (до 7-8 сут) длина отростков превышала диаметр сомы, на концах нейритов наблюдались округлые наплывы цитоплазмы (зоны роста), свидетельствующие о развитии нейритов. Как в опыте, так и в контроле наблюдались утолщение, ветвление отростков и агрегация нейронов (рис. 10). Эти характеристики

15
поведения нейронов в культуре, выделенных из заморожено-оттаянных ганглиев, не отличаются от контрольных и соответствуют описаниям культуры нейронов моллюсков, данных Костенко (1985) и Сотниковым (2008).
Без криоконсервации
После криоконсервации
Рис. 10. Регенерация нервных отростков и формирование контактов в клеточной культуре (от 3 сут) до и после криоконсервации мозга моллюска. а) – образование ламелл и филлоподий; б) – формирование отростков из «невтянувшегося» аксона; в) – образование ламеллоподий, формирование межнейронных контактов и агрегация нейронов в группы; г) – агрегация нейронов в группу, рост отростков и аутозамыкание нейрональных отростков; д) – образование нейрональных контактов, агрегация нейронов в группы. Стрелками отмечены участки образования нейрональных контактов. тн - тело нейрона, д – дендриты, а – аксоны, ф – филлоподии, л – ламеллы, к – контакты, аг – агрегация нейронов, о – отростки нейрона.
При культивировании более 7-8 суток на фоне интенсивного разрастания отростков и их ветвления наблюдали образование контактов между отростками соседних клеток. Важно отметить, что стремление нейронов устанавливать контакты было свойственно всем культивируемым нейронам, которые регенерировали отростки и находились на расстоянии 150 мкм друг от друга как в опыте, там и в контроле. Если у нейрона не были сформированы нейриты, в большинстве случаев он не контактировал с другими нейронами, а отростки, которые подходили от соседних клеток «обходили» данный нейрон.
Дифференцированные униполярные нейроны моллюска в культуре в результате регенерации превращались в мультиполярные. В живой культуре нейронов, выделенных как из интактного мозга не прошедшего криоконсервацию, так и из прошедшего криоконсервацию мозга, наблюдались постоянные перемещения нейронов вследствие сокращения или удлинения нейритов и их объединения с другими нейронами в группы.
Таким образом, мы показали, что нейроны, выделенные из криоконсервированных ганглиев моллюска, сохраняют регенерационную способность. В культуре in vitro они

16
постоянно формируют отростки, которые растут, ветвятся и образуют контакты друг с другом (рис. 10). На основании этих данных можно предположить, что отсутствие синаптических связей в заморожено-оттаянных ганглиях является следствием криоповреждений нервных отростков и структуры синапсов, находящихся в нейропиле и комиссурах.
Следующий этап нашей работы был направлен на поиск нейро- и криозащитных средств с целью максимального снижения криоповреждений вызванных криоконсервировацией органа и/или для повышения регенеративных свойств ткани.
5. Влияние пептида TSKY на жизнеспособность изолированных нейронов
Для оптимизации процесса криоконсервации изолированного мозга был выбран путь поиска нейропротекторных добавок, которые могли бы снижать уровень метаболизма нейронов, т.к. в гипометаболическом состоянии клетки более устойчивы к различным стрессовым факторам. Мы предположили, что одним из таких веществ может быть нейропептид TSKY, выделенный из мозга зимоспящих сусликов Spermophillus undulates (Ziganshin et al., 1994; Kramarova et al., 1996, Игнатьев и др., 1995).
5.1. Эффекты пептида TSKY в зависимости от температуры при предварительной инкубации ганглиев моллюска (22-24°С и 4-6°С)
Дозо-зависимые эффекты пептида TSKY при инкубации мозга моллюска при 22–24°С После инкубации мозга моллюска при температуре 22–24°С в растворе с различными
концентрациями пептида TSKY доля живых нейронов при культивировании увеличивалась по сравнению с контролем в среднем на 72.2% при концентрации пептида 1·10-5 М, на 54.4% - при 1·10-6 М и на 57.2% - при 1·10-7 М (рис. 11А). При этих же условиях доля нейронов с отростками через 24 ч культивирования снижалась по сравнению с контролем и в среднем составляла 30.8% при концентрации пептида 1·10-5 М, 29.17% - при 1·10-6 М и 13.3% - при 1·10-7 М (рис. 12А). Дозо-зависимые эффекты пептида TSKY при инкубации мозга моллюска при 4–6°С
После инкубации мозга моллюска при пониженной температуре (4–6°С) в растворе с различными концентрациями пептида TSKY доля живых нейронов при культивировании значительно увеличилась по сравнению с контролем в 3,5 раза при 1·10-5 М, в 3,2 раза при 1·10-6 М и в 2 раза при 1·10-7 М (рис. 11Б).
Рис. 11. Количество живых нейронов после культивирования в течение 24 ч в КСНП. Предварительная инкубация изолированного мозга моллюска при 22–24°С (А) и 4–6°С (Б) в течение 60 мин. (*), *(*), *** – разница между количеством живых нейронов в опыте с инкубацией в растворе без пептида (контроль) и после инкубации с пептидом в различных концентрациях TSKY; p < 0.1, p < 0.025; p < 0.0001, соответственно; непарный t-критерий Стьюдента.

17
Инкубация мозга моллюска при пониженной температуре (4–6°С) в растворе с различными концентрациями пептида TSKY не приводила к значимому изменению количества клеток с формирующимися отростками в первые сутки культивирования, и доля клеток с отростками составила 48% при 1·10-5 М, 24% - при 1·10-6 М и 29% - при 1·10-7 М (рис. 12Б). Однако необходимо отметить, что в данных условиях сохранялась тенденция к снижению формирования отростков. В то же время, количество нейронов, формирующих отростки в первые сутки культивирования после предварительной обработки пептидом TSKY в различных концентрациях, было достоверно ниже, чем в контроле при 22–24°С (рис. 12А). Нами не было обнаружено достоверных отличий между контрольными и опытными экспериментами после обработки пептидом при низкой температуре, 4–6°С.
Рис. 12. Количество живых нейронов с отростками после культивирования in vitro в течение 24 ч. Предварительная инкубация мозга моллюска в растворе с различными концентрациями пептида TSKY при 22–24°С (А) и при 4–6°С (Б) *, ** – разница между числом живых нейронов с отростками в опыте с инкубацией в растворе без пептида (контроль) и после инкубации с пептидом, p < 0.05, p < 0.001, соответственно; непарный t-критерий Стьюдента.
Таким образом, под влиянием пептида TSKY при 4–6°C происходит увеличение (в 3.5 раза) общего числа живых клеток (рис. 11Б). Наиболее выраженный эффект наблюдался при концентрации пептида 10-5M. Кроме того, под действием пептида наблюдалось снижение (рис. 12А), либо тенденция к снижению (рис. 12Б), числа нейронов с отростками вне зависимости от температуры инкубации мозга с пептидом.Имеются данные, что концентрация пептида TSKY в мозге зимоспящих животных, находящихся в состоянии спячки, приблизительно в 2 раза выше, чем в мозге активных животных, и этот пептид способен значительно снижать скорость сердечных сокращений у 36-часовых куриных эмбрионов (Kramarova et al., 1996). Показано также, что охлаждение мышей, которым предварительно ввели пептид TSKY, индуцировало у животных длительную гипотермию (Игнатьев и др., 2005).
Таким образом, полученные нами результаты, позволяют предположить, что нервные клетки моллюска при обработке пептидом TSKY могут переходить в новое физиологическое состояние (возможно, в состояние гипометаболизма/гипобиоза), в котором могут находиться в течение продолжительного промежутка времени без потери жизнеспособности, но со снижением функциональных свойств.

18
5.2. Влияние нейропептида TSKY на сохранность нейронов в процессе криоконсервации
Для изучения влияния пептида TSKY на выживаемость и регенеративную способность нейронов после криоконсервации мозга провели 3 варианта экспериментов: 1) без добавления TSKY: инкубирование ганглиев в криозащитном растворе с 2М ДМСО в течение 20 мин при 4-6°С и последующее замораживание; 2) с добавлением 10-5 М TSKY в криозащитный раствор и последующим замораживанием в этом же растворе; 3) 10-5 М TSKY добавляли только в раствор для оттаивания и отмывания от криозащитной среды.
Первый вариант эксперимента служил контролем. Во втором варианте эксперимента мы не обнаружили в клеточной культуре разницы в количестве живых нейронов, выделенных из оттаянного мозга, предварительно обработанного и замороженного в криозащитной среде с 10-5 М TSKY, по сравнению с контролем (39.5 ± 24.4%, n=12) (рис. 13).
Рис. 13. Влияние пептида TSKY (10-5М) на жизнеспособность нейронов в культуре (24 ч) после криоконсервации мозга. Количество живых нейронов и нейронов с отростками (%): 1 (контроль) – в отсутствие пептида в инкубационной среде и в криозащитном растворе; 2 - после предварительной инкубации мозга с пептидом и с добавлением пептида в криозащитный раствор; 3- добавление пептида при оттаивании и отмывании мозга от криозащитной среды. ***, * – указывают на достоверную разницу между значениями количества живых нейронов и нейронов с отростками относительно контроля, p < 0.0004 и p < 0.033, соответственно; непарный t-критерий Стьюдента.
Однако количество нейронов, формирующих отростки, в процентном отношении к числу живых нейронов было одинаковым в контроле (рис. 13,1) и опыте (рис. 13,2). Для выяснения возможных проявлений криоповреждений в период оттаивания мы провели третий вариант экспериментов. Пептид добавили в раствор, где происходило оттаивание ганглиев и отмывка от криопротектора. Результаты показали, что количество живых нейронов в опыте было в 2 раза выше относительно контроля (n = 10), однако количество нейронов с отростками относительно общего числа живых было на 10% ниже по сравнению с тем же контролем (рис. 13, 3). Предположительно, физические процессы в период оттаивания (например, перекристаллизация) вызывают такие же повреждения, как и в период замораживания, и присутствие в этот период пептида TSKY защищает нейроны от возможных негативных воздействий микрочастиц льда (МКЧЛ). Вероятно, в данных условиях пептид TSKY, может участвовать и в репарационных процессах нейронов.
На основании полученных результатов можно сделать вывод, что пептид TSKY не оказывает криопротекторного действия в процессе замораживания, но, возможно, обладает гипометаболическими свойствами, что снижает функциональную активность нейронов, уменьшая чувствительность нейронов к повреждениям, тем самым проявляя нейропротекторный эффект.

19
Для выяснения причин отсутствия положительного влияния TSKY при использовании его непосредственно при криоконсервации мы мы проанализировали его влияние на характер кристаллизации и формирование МКЧЛ при замораживании растворов до -196°С.
5.3. Изучение возможных криозащитных свойств TSKY
Была проведена видеосъемка процесса замораживания от 20°С до -196°С физиологического раствора с добавлением следующих компонентов: 20% L-15 (Leibowitz); 20% L-15 + TSKY (10-5 М); 20% L-15 + 2М ДМСО; 20% L-15 + TSKY (10-5М) + 2М ДМСО.
Анализ видеоизображений начального этапа кристаллообразования в растворах без пептида (рис. 14а,б) и с добавлением пептида TSKY (10-5 М) (рис. 14в,г) показал, что TSKY не влиял на характер кристаллизации. Кроме того, присутствие TSKY не смещало значения температурной точки кристаллизации растворов.
При температурах ниже -100°С происходило растрескивание льда (Rabin et al., 2006; Андреев и др., 2009) и формирование МКЧЛ разной формы и размеров в зависимости от состава раствора.
Рис. 14. Криомикроскопия протектирующих сред: а,д) 20% L-15 (Leibowitz); б,е) 20% L-15 + TSKY (10-5 М); в,ж) 20% L-15 + 2М ДМСО; г,з) 20% L-15 + TSKY (10-5 М) + 2М ДМСО. а-г начало кристаллизации (раскадровка с видео); д-з – сформированные микрочастицы льда при -196°. Шкала 100 мкм.
На рис. 14д показано, что при замораживании в физиологическом растворе с 20% L-15 формируются МКЧЛ с гранями в виде правильных геометрических фигур с острыми углами (ромбы, треугольники, трапеции), размером от 50 до 100 мкм. Подобную форму и размеры имеют МКЧ льда после замораживания дистиллированной воды и физиологического раствора (Андреев и др., 2009). При замораживании раствора с 20% L-15 и 2М ДМСО наблюдалось формирование округлых и шестиугольных МКЧЛ со сглаженными краями (рис. 14ж). Добавление пептида TSKY (10-5 М) в эти два раствора не изменяло характера льдообразования (рис. 14е,з).
Отсутствие эффекта пептида TSKY на температурную точку кристаллизации растворов, характер начального этапа кристаллизации и формообразование МКЧЛ подтверждает наше предположение, что TSKY не является классическим криопротектором. Мы предполагаем, что пептид TSKY проявляет нейропротекторные свойства, защищая клетки от механического и осмотического стресса, вызванных рекристаллизацией льда и выравниванием осмотических градиентов во время или после оттаивания. Кроме этого не исключено, что, благодаря этому свойству, TSKY может влиять на репарационные процессы в поврежденной ткани.

20
ЗАКЛЮЧЕНИЕ
В результате проведенных исследований мы получили данные, которые свидетельствуют, что после криоконсервации изолированного мозга моллюска Lymnaea stagnalis L. нейроны сохраняются живыми (флуоресцентный анализ Live/Dead) и восстанавливают функциональную активность. Мозг моллюска (1.5 – 2.7 мм) – это кольцо ганглиев, которые содержат ~20000 полифункциональных нейронов и включают богатые вненейрональные области, состоящие из нескольких видов глиальных клеток, длинных аксонов, синаптических структур. Криоконсервации таким образом подвергалась интегрированная структура, включающая разнородные элементы.
Эксперименты показали, что в органной культуре после криоконсервации мозга нейроны восстанавливали электрические характеристики (МП, генерация ПД и ПСП) до характерных для этих нейронов значений. Однако, синаптических связей между нейронами зарегистрировано не было. Такого рода повреждения нервной ткани могли происходить в центральных областях ганглиев, в нейропиле, и/или в комиссурах и коннективах, соединяющих ганглии. В клеточной культуре удалось показать, что нейроны, выделенные из криоконсервированного мозга, сохраняли способность к регенерации нервных отростков и формированию контактов. На этом основании можно предположить, что при определенных условиях в органной культуре поврежденные области нервной ткани могут быть восстановлены.
Первостепенное значение имеют исследования, направленные на предотвращение криоповреждений путем модификации криозащитных сред. Привлекательно использовать для этих целей природные агенты, участвующие в биохимических механизмах холодо- и морозоустойчивости. Интерес представляют, с нашей точки зрения, те, которые могут снижать уровень метаболизма ткани (или органа), т.к. в гипометаболическом состоянии клетки более устойчивы к стрессовым ситуациям. Предположительно такими свойствами может обладать пептид TSKY (Thr-Ser-Lys-Tyr), выделенный из мозга зимоспящих сусликов Spermophillus undulates (Ziganshin et al., 1994; Kramarova et al., 1996). Нам удалось впервые показать, что пептид TSKY, добавленный в криозащитный раствор на на стадии оттаивания, может проявлять нейропротекторные свойства и оказывать метаболический эффект, увеличивая жизнеспособность ткани мозга моллюска на этапе размораживания. Эти результаты расширяют возможности в усовершенствовании криозащитных сред, которые могут быть использованы для криоконсервации нервной ткани.
ВЫВОДЫ
1. Показано, что после криоконсервации изолированного мозга моллюска Lymnaea stagnalis L. нейроны сохраняют жизнеспособность и электрическую активность (МП, ПД, ПСП) при органотипическом культивировании. Зарегистрированы ответы нейронов криоконсервированного мозга моллюска на аппликацию серотонина.
2. Синаптические связи между идентифицированными нейронами не были зарегистрированы в криоконсервированном мозге, в отличие от контроля. Связи не восстанавливались на протяжении 16 суток органотипического культивирования. Предположено, что причинами отсутствия связей могут быть криоповреждения межклеточных структур в нейропиле и комиссурах.
3. В клеточной культуре показано, что нейроны после криоконсервации сохраняют функциональную активность: формируют нервные отростки и устанавливают контакты друг с другом.

21
4. Впервые показано нейропротекторное действие пептида TSKY, выделенного из мозга зимоспящих животных, на стадии оттаивания криоконсервированного мозга моллюска. При замораживании изолированного мозга моллюска пептид TSKY не проявлял криопротекторных свойств.
5. Анализ видеоизображений процесса замораживания до температуры жидкого азота показал, что пептид TSKY не влияет на характер кристаллизации и формообразование МКЧ льда, что подтверждает отсутствие у TSKY криопротективных свойств в период замораживания.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
Статьи в рецензируемых журналах: 1. Н.А. Ивличева, Е.В. Дмитриева, М.А. Костенко, Э.Н. Гахова. Криоконсервированные нейроны моллюска способны к морфологической дифференцировке в культуре in vitro// Биофизика, 49(4), 2004, 710-714. 2. Н.А. Ивличева, С.Н. Мякишева, Э.Н. Гахова. Нервная клетка – модель для изучения механизма действия криопротекторов и эффективности криоконсервации // Ветеринарная патология, 1(20), 2007, 42-43. 3. Н.А. Ивличева, Л.И. Крамарова, Р.Х. Зиганшин, В.Г. Цыганова, В.В. Рогачевский, Э.Н. Гахова. Влияние пептида Thr-Ser-Lys-Tyr на жизнеспособность изолированных нейронов Lymnaea stagnalis L. //Биологические мембраны, 29(3), 2012, 195-199. 4. Н.А. Ивличева, Э.Н. Гахова. Культура дифференцированных нейронов взрослого моллюска – альтернативная модель изучения процессов регенерации нервной системы// Биомедицина, 6, 2007, 89-96. 5. С.Н. Мякишева, Н.Ю. Сахарова, Н.А Ивличева. Криоконсервация культивируемых клеток нейробластомы мыши N1E-115 для биомедицинских исследований // Биомедицина, 6, 2007, 153-157.
Статьи в книгах: 6. L.I. Kramarova, N.A. Ivlicheva, R.H. Ziganshin, A.A. Andreev, E.N. Gakhova. The hibernation-related peptide TSKY acts as a neuroprotector in cultured pond snail neurons // Living in a Seasonal World: Thermoregulatory and Metabolic Adaptations, R. Bieber, A. Millesi (Eds.) Springer-Verlag GmbH, Heidelberg, Deutschland, chapter 18, 2012, 201-210.
Статьи в сборниках: 7. Н.А. Ивличева, И.А. Чистопольский, Э.Н.Гахова. Криоконсервация окологлоточного кольца ганглиев пресноводного моллюска Lymnaea stagnalis L. // Биофизика живой клетки, 9, 2008, 58-60. 8. Н.А. Ивличева, В.Г. Цыганова, В.В. Рогачевский, Л.И. Крамарова, Р.Х. Зиганшин, Э.Н. Гахова. Влияние регулятора естественного гипобиоза, пептида TSKY на жизнеспособность изолированных нейронов Lymnaea stagnalis в культуре in vitro // В сб. статей международной конференции «Рецепция и внутриклеточная сигнализация», г.Пущино, Т.2, 2011, 505-510.
Тезисы: 9. Н.А. Ивличева, М.А. Костенко. Способность нейронов моллюска Lymnaea stagnalis L. после криоконсервании формировать в культуре in vitro ветвистые отростки // В сб. тезисов VIII международной школы конференции молодых ученых, г. Пущино, 2004, с.83. 10. Н.А. Ивличева, Е.В. Дмитриева, М.А.Костенко, Э.Н. Гахова. Изучение жизнеспособности изолированных нейронов моллюска Lymnaea stagnalis L. после криоконсервации изолированного

22
мозга методом культуры клеток in vitro // В сб. тезисов Всероссийской конференции молодых исследователей «Физиология и медицина», г. Санкт-Петербург, 2005, с.46. 11. Н.А. Ивличева, С.Н Мякишева, М.А. Костенко. Изучение влияния рН среды на сезонную способность формировать отростки нейронами моллюска Lymnaea Stagnalis L. в культуре in vitro// В сб. тезисов IX международной школы конференции молодых ученых, г. Пущино, 2005, с.116. 12. N.A. Ivlicheva, S.N. Myakisheva, E.N. Gakhova. The neurite regeneration and outgrowth in vitro culture before and after cryopreservation of isolated brain of snail Lymnaea stagnalis L. depending on the medium pH // Simpler Nervous Systems: VIII East European Conference of the International Society for Invertebrate Neurobiology, September 13-17, thesis of lectures, Kazan, 2006, p. 34. 13. Н. А. Ивличева, И. А. Чистопольский. Восстановление функциональных свойств нейронов в культуре in vitro после криоконсервации изолированной нервной системы моллюска Lymnaea stagnalis (-196°C) // В сб. тезисов XI международной школы конференции молодых ученых, г. Пущино, 2007, с.248-249. 14. N.A. Ivlicheva, I.A. Chistopolskiy, E.N. Gakhova. Isolated neurons from cryopreserved (-196°С) snail brain retain viability. // Simpler Nervous Systems: IX East European Conference of the International Society for Invertebrate Neurobiology, September 9-13/ thesis of lectures, St. Petersburg, 2009, p. 46. 15. Н.А. Ивличева, И.А. Чистопольский, Э.Н. Гахова. Восстановление активности нейронов после криоконсервации изолированной нервной системы моллюска Lymnaea stagnalis (-196°C) // В сб. тезисов XIII международной школы конференции молодых ученых, г. Пущино, 2009, с.103. 16. Н.А. Ивличева, Чистопольский, Э.Н. Гахова. Простая нервная система: анализ жизнеспособности нейронов изолированного мозга Lymnaea stagnalis после криоконсервации (-196°C) //В сб. тезисов III Всероссийского с международным участием конгресса студентов и аспиратнов-биологов «Симбиоз-Россия 2010», г.Нижний Новгород, 2010, с.169-170. 17. Н.А. Ивличева, Э.Н. Гахова, B.Г. Цыганова, В.В. Рогачевский, Р.Х. Зиганшин, Л.И. Крамарова. Нейропротекторный эффект пептида TSKY при низких и сверхнизких температурах // В сб. тезисов 7го международного междисциплинарного конгресса «Нейронаука для медицины и психологии», Судак, Крым, Украина, 2011, с.193-194. 18. Л.И. Крамарова, Э.Н. Гахова, Р.Х. Зиганшин, Н.А. Ивличева. Эндогенные гипометаболические факторы и их возможное применение при трансплантации и криоконсервации нервной ткани // В сб. тезисов 7го международного междисциплинарного конгресса «Нейронаука для медицины и психологии», Судак, Крым, Украина, 2011, с.234-235. 19. Н.А. Ивличева, Л.И. Крамарова, Р.Х. Зиганшин, А.А. Андреев, В.Г. Цыганова, Э.Н. Гахова. Влияние пептида TSKY на жизнеспособность нейронов в культуре после криоконсервации изолированного мозга пресноводного моллюска Lymnaea stagnalis L. // В сб. тезисов 8го международного междисциплинарного конгресса «Нейронаука для медицины и психологии», Судак, Крым, Украина, 2012, с.179-180. 20. Ивличева Н.А., Крамарова Л.И., Андреев А.А., Зиганшин Р.Х., Гахова Э.Н. Нейропротекторный эффект гипометаболического фактора TSKY на нейроны моллюска при подготовке к криоконсервации (-196°С) // В сб. тезисов IV Съезда биофизиков России, г.Нижний Новгород, Т.2, 2012, с. 60.


















![Звуки [Н], [Н ], буквы Н, н](https://img.dokumen.tips/doc/110x75/5681480b550346895db53a76/-5681480b550346895db53a76.jpg)










