Embed Size (px)
Citation preview

ARTICLE
VELAFRONS COAHUILENSIS, A NEW LAMBEOSAURINE HADROSAURID (DINOSAURIA:ORNITHOPODA) FROM THE LATE CAMPANIAN CERRO DEL PUEBLO FORMATION,
COAHUILA, MEXICO
TERRY A. GATES,*,1 SCOTT D. SAMPSON,1 CARLOS R. DELGADO DE JESÚS,2 LINDSAY E. ZANNO,1
DAVID EBERTH,3 RENÉ HERNANDEZ-RIVERA,4 MARTHA C. AGUILLÓN MARTÍNEZ,2 and JAMES I. KIRKLAND5
1University of Utah, Dept of Geology and Geophysics/ Utah Museum of Natural History, 1390 E Presidents Circle, Salt LakeCity, UT, 84112, USA, [email protected], [email protected], [email protected];
2Secretaría de Educación y Cultura-Museo del Desierto, Dept. of Paleontology. Prol. Pérez Treviño 3745, Parque las Maravillas.Saltillo, Coahuila, México. 25015, [email protected], [email protected];
3Royal Tyrrell Museum of Paleontology, PO Box 7500, Drumheller, T0J 0Y0, Alberta, Canada, [email protected];4Instituto de Geología, Dept. of Paleontology, UNAM, Mexico City, M-04510, DF, Mexico, [email protected];
5Utah Geological Survey, 1594 W North Temple Ste 3110, PO Box 146100, Salt Lake City, 84114-6100, USA,[email protected]
ABSTRACT—A new lambeosaurine hadrosaurid, Velafrons coahuilensis, is described as the first lambeosaurine from theCerro del Pueblo Formation of Coahuila, Mexico, and the first lambeosaurine genus to be named from North Americain more than 70 years. Although the holotype specimen is a juvenile individual—as evidenced by its incomplete crestdevelopment and relative size compared to other North American lambeosaurines—ontogeny independent autapomor-phies have been identified including quadrate with narrow quadratojugal notch and a postorbital with well developed,dorsally positioned squamosal process. Additionally, this taxon is unique in that the prefrontal is not dorsally deflectedand anteroposteriorly expanded as in other lambeosaurine taxa of its size, but rather retains the frontal-prefrontal“clamp” present in smaller individuals of other taxa. Phylogenetic analysis places Velafrons in a polytomy with numerousother fan-crested lambeosaurines. The crest structure of Velafrons more closely resembles that of Corythosaurus andHypacrosaurus because it possesses an anteriorly projecting nasal process over the dorsal premaxilla process. Biogeo-graphically, Velafrons is one of three distinct hadrosaurids known from approximately 73.5 Ma—two lambeosaurines andone hadrosaurine—all restricted to the southern region of the Western Interior Basin of North America.
INTRODUCTION
The Cerro del Pueblo Formation (CdP Fm) in the state ofCoahuila, Mexico (Fig. 1) has been acclaimed as a productivefossiliferous unit for decades (Wolleben, 1977; Vega and Feld-mann, 1991; Kirkland et al., 2001), yet thus far it has been subjectto limited paleontological work. The majority of research in theregion to date has focused on the stratigraphy and sedimentologyof the Difunta Group (Fig. 1), a suite of formations bounded bythe Parras Shale below and the Rancho Nuevo Formation above,of which the CdP Fm forms the base (Murray et al., 1962; Mc-Bride et al., 1974; Kirkland et al., 2001; Eberth et al., 2004).Eberth and colleagues (2004) estimated the age of the CdP Fm tobe approximately 73.5 Ma based on detailed magnetostratigra-phy (32n.3r–32n.2n chron). Overall, the CdP Fm represents aregressive sequence—from the underlying marine Parras Shaleto the terrestrial overlying Cerro Huerto Formation—of coastalplain settings punctuated by small marine incursions (Eberth etal., 2004).
Throughout the formation, including the small marine beds,both vertebrate and invertebrate fossils are common. Fossils in-clude ammonites, pelecypods, gastropods, mosasaurs, turtles,crocodiles, mammals, and dinosaurs (Wolleben, 1977; Vega andFeldmann, 1991; Hernandez, 1992; Hernandez and Kirkland,
1993; Hernandez et al., 1995; Hernandez Rivera, 1996; Aguillon-Martinez et al., 1998; Aguillon et al., 1998; Rodriguez de la Rosaand Cevallos Ferriz, 1998; Kirkland et al., 2001; Brinkman et al.,2002; Garcia and Reynoso, 2002; Kirkland and Aguillon-Martinez, 2002; Eberth et al., 2003; Perrilliat et al., 2003; Rod-riguez de la Rosa, 2003; Rodriguez de La Rosa et al., 2003).Although the dinosaurian fauna of the CdP Fm has not beenexplored extensively, its diversity already surpasses that of anyother Cretaceous deposits in Mexico. Dinosaurian constituentsinclude troodontid, ornithomimid, and tyrannosaurid theropods(Rodriguez de la Rosa and Cevallos Ferriz, 1998), an unde-scribed ceratopsid, a putative protoceratopsid-grade ceratopsian,a hadrosaurine (non-crested) hadrosaurid, and a new lambeo-saurine (crested) hadrosaurid.
In 1986, the Royal Ontario Museum undertook the first largepaleontological expedition to the CdP Fm. Their efforts wereconcentrated on hadrosaurid bonebeds around the town of Presade San Antonio, with sporadic collecting from Rincon Colorado(Fig. 1). Subsequently, “Dinamation International Society” re-turned to Rincon Colorado and established several quarries inthe area, including quarry 7A, which produced the skeleton ofthis new lambeosaurine taxon. The quarry was worked season-ally from 1992 to 2001, yielding a mostly complete postcranialskeleton but no skull material. In 2002, a joint expedition involv-ing the Utah Museum of Natural History, the Royal Tyrrell Mu-seum of Paleontology, and the Museo del Desierto in Saltillo,Coahuila, reinitiated excavations at the 7A quarry, recovering*Corresponding author.
Journal of Vertebrate Paleontology 27(4):917–930, December 2007© 2007 by the Society of Vertebrate Paleontology
917

the disarticulated skull and several cervical vertebrae. The skulland postcranium of the holotype specimen were disarticulatedbut closely associated in a highly cemented calcareous grey mud-stone. The quarry is associated with a brackish water molluscanassemblage that includes Aphrodina tippana, Cerithium pueb-loensis, and Nerita, with some of the bones encrusted by Flem-ingostrea subspatula (Kirkland et al., 2001, fig 13g; Kirkland etal., 2006).
Here we describe the first lambeosaurine hadrosaurid fromthe Cerro del Pueblo Formation and the first new North Ameri-can lambeosaurine genus named in over 70 years. Further, weaddress the issue of crest development in this lambeosaurinetaxon compared to that of other lambeosaurine hadrosauridsalong with the biogeography of late Campanian hadrosaurids inthe Western Interior Basin (WIB).
Institutional Abbreviations—CMN, Canadian Museum ofNature, Ottawa, Ontario, Canada; MOR, Museum of the Rock-ies, Bozeman, Montana, USA; ROM, Royal Ontario Museum,Toronto, Ontario, Canada; CPC, Colección Paleontológica deCoahuila (Paleontological Collection of Coahuila) Saltillo, Coa-huila, Mexico; TMP, Royal Tyrrell Museum of Paleontology,Drumheller, Alberta, Canada; UMNH, Utah Museum of Natu-ral History, Salt Lake City, Utah, USA.
SYSTEMATIC PALEONTOLOGY
DINOSAURIA Owen, 1842ORNITHISCHIA Seeley, 1888ORNITHOPODA Marsh, 1882
HADROSAURIDAE Cope, 1869LAMBEOSAURINAE Parks, 1923
VELAFRONS, gen. nov.
Etymology—Vela (�sail, Spanish) + frons (�forehead,Latin); meaning “sailed forehead,” in reference to the sail-likecrest that is found on the forehead of this taxon.
Diagnosis—as for type and only species.
VELAFRONS COAHUILENSIS, sp. nov.
Etymology—coahuil (� Coahuila, the Mexican state in whichthe fossil was discovered) + ensis; meaning “from Coahuila.”
Holotype—CPC-59, partial subadult skull including left andright premaxillae and maxillae, right jugal, nasal, quadrate,mostly complete skull roof, dentary, and partial postcrania.
Type Locality—7A quarry, about 1 km northeast of the townRincon Colorado, Coahuila, Mexico (Fig. 1). The site occursapproximately 335 m from the base of the Cerro del PuebloFormation (Eberth et al., 2004).
Diagnosis—Lambeosaurine hadrosaurid characterized by thefollowing autapomorphies: quadrate with narrow quadratojugalnotch; postorbital with a dorsally positioned, high arching squa-mosal process; ceratobranchial with rounded anterior end; aswell as the unique suite of characters: a flat fan-like crest createdby the nasals and premaxillae, a jugal with poorly developedposteroventral flange positioned more posteriorly than otherlambeosaurine taxa, and an anteriorly recurved posterior pro-cess, and a well-developed raised squamosal shelf.
DESCRIPTION
The skull of CPC-59 consists of a complete dentary (lackingteeth), ceratobranchial, mostly complete left and right premax-illae, left and right maxillae, right jugal, quadrate, and nasal, anda virtually complete skull roof with fragmentary braincase (Fig.2). The entire skull was disarticulated, except for the skull roof,and covered in a thick iron crust obscuring some sutures andsubtle surface morphology, such as foraminae. In particular, thebraincase suffers from preservational issues that prevent a de-tailed reconstruction and description.
Based on a number of features (relative size, vertebral sutureclosure, conformation of skull elements; see Discussion), it isvery likely that CPC-59 represents a juvenile individual. Theskulls, and particularly the crests, of lambeosaurines change dra-matically during ontogeny (Dodson, 1975; Evans et al., 2005).
FIGURE 1. Map showing Cretaceous-aged fossiliferous basins in Coahuila, Mexico, and the stratigraphy of the Parras Basin. Modified from Eberthet al. (2004:fig. 2, 3). Quarry 7A is located approximately 1 km northeast of Rincon Colorado. Abbreviations: P de S.A., Presa de San Antonio.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 27, NO. 4, 2007918
Thus, it is almost certain that the crest of Velafrons does notrepresent the final adult form. Nonetheless, work by Evans(2005) demonstrates that juveniles possess a number of genus-level diagnostic characters. The morphological changes associ-ated with crest growth in this taxon will be discussed more thor-oughly below.
The postcranium of this specimen is not described here due tospace constraints, but is currently under study for publication ata later date.
Cranium
The skull roof is mostly complete except for portions of thefrontal and the ventral processes of the postorbital and squamo-
sal. An iron crust obscures several of the sutures and there iscrushing through the right temporal bar. Nonetheless, the skullroof provides critical information about the morphology andphylogenetic relationships of Velafrons.
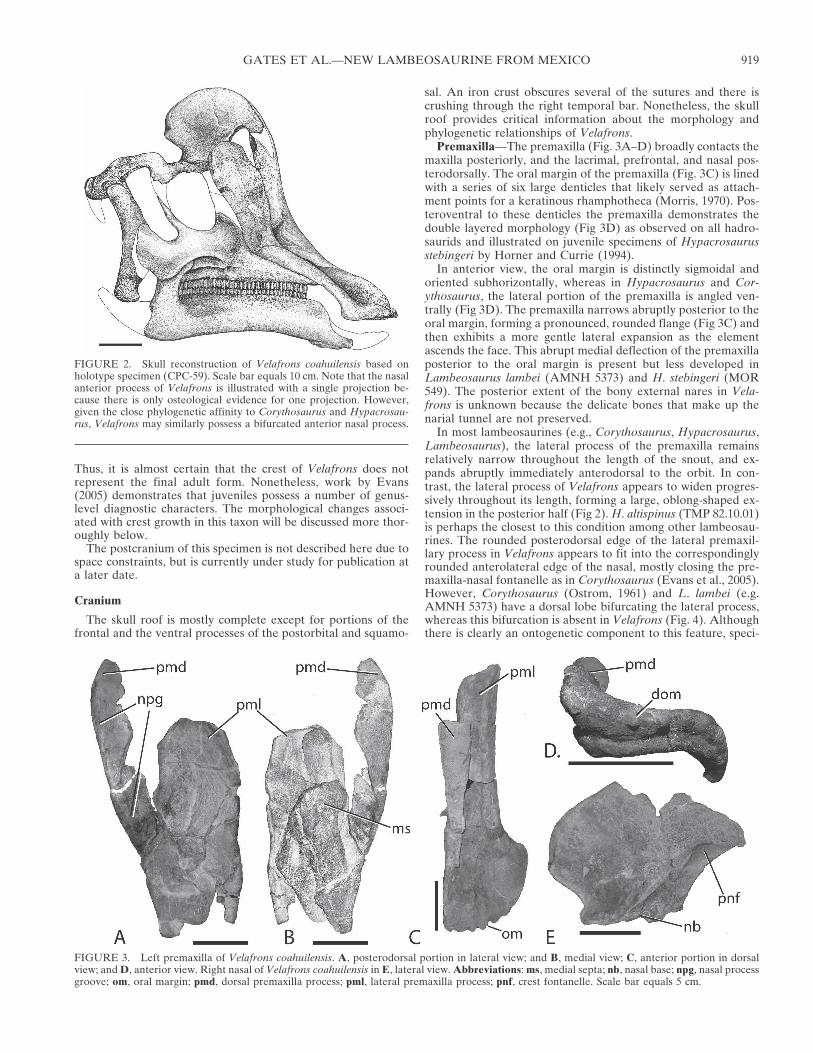
Premaxilla—The premaxilla (Fig. 3A–D) broadly contacts themaxilla posteriorly, and the lacrimal, prefrontal, and nasal pos-terodorsally. The oral margin of the premaxilla (Fig. 3C) is linedwith a series of six large denticles that likely served as attach-ment points for a keratinous rhamphotheca (Morris, 1970). Pos-teroventral to these denticles the premaxilla demonstrates thedouble layered morphology (Fig 3D) as observed on all hadro-saurids and illustrated on juvenile specimens of Hypacrosaurusstebingeri by Horner and Currie (1994).
In anterior view, the oral margin is distinctly sigmoidal andoriented subhorizontally, whereas in Hypacrosaurus and Cor-ythosaurus, the lateral portion of the premaxilla is angled ven-trally (Fig 3D). The premaxilla narrows abruptly posterior to theoral margin, forming a pronounced, rounded flange (Fig 3C) andthen exhibits a more gentle lateral expansion as the elementascends the face. This abrupt medial deflection of the premaxillaposterior to the oral margin is present but less developed inLambeosaurus lambei (AMNH 5373) and H. stebingeri (MOR549). The posterior extent of the bony external nares in Vela-frons is unknown because the delicate bones that make up thenarial tunnel are not preserved.
In most lambeosaurines (e.g., Corythosaurus, Hypacrosaurus,Lambeosaurus), the lateral process of the premaxilla remainsrelatively narrow throughout the length of the snout, and ex-pands abruptly immediately anterodorsal to the orbit. In con-trast, the lateral process of Velafrons appears to widen progres-sively throughout its length, forming a large, oblong-shaped ex-tension in the posterior half (Fig 2). H. altispinus (TMP 82.10.01)is perhaps the closest to this condition among other lambeosau-rines. The rounded posterodorsal edge of the lateral premaxil-lary process in Velafrons appears to fit into the correspondinglyrounded anterolateral edge of the nasal, mostly closing the pre-maxilla-nasal fontanelle as in Corythosaurus (Evans et al., 2005).However, Corythosaurus (Ostrom, 1961) and L. lambei (e.g.AMNH 5373) have a dorsal lobe bifurcating the lateral process,whereas this bifurcation is absent in Velafrons (Fig. 4). Althoughthere is clearly an ontogenetic component to this feature, speci-
FIGURE 2. Skull reconstruction of Velafrons coahuilensis based onholotype specimen (CPC-59). Scale bar equals 10 cm. Note that the nasalanterior process of Velafrons is illustrated with a single projection be-cause there is only osteological evidence for one projection. However,given the close phylogenetic affinity to Corythosaurus and Hypacrosau-rus, Velafrons may similarly possess a bifurcated anterior nasal process.
FIGURE 3. Left premaxilla of Velafrons coahuilensis. A, posterodorsal portion in lateral view; and B, medial view; C, anterior portion in dorsalview; and D, anterior view. Right nasal of Velafrons coahuilensis in E, lateral view. Abbreviations: ms, medial septa; nb, nasal base; npg, nasal processgroove; om, oral margin; pmd, dorsal premaxilla process; pml, lateral premaxilla process; pnf, crest fontanelle. Scale bar equals 5 cm.
GATES ET AL.—NEW LAMBEOSAURINE FROM MEXICO 919
mens of other genera of equivalent size to Velafrons (e.g., C.casuarius, ROM 1947) already possess the small dorsal exten-sion, indicating that its absence in Velafrons may be unrelated todevelopmental stage.
The dorsal process of the premaxilla in Velafrons remains deli-cate and slender throughout its length, contacting its oppositemedially and flanked laterally by the anterior process of thenasal. The small size of the Velafrons dorsal process contrastswith that seen in equivalent sized (and presumably aged) indi-viduals of other genera (Fig. 4; see below). As reconstructed atthis developmental stage, the crestal portion of the dorsal pro-cess terminates at the front of the crest, well anterior to the orbit.Overall, given the presumed subadult age of CPC-59, the condi-tion in Velafrons most closely resembles that of H. altispinus(TMP 82.10.01), which retains a relatively narrow dorsal processforming the anterior portion of the crest and contributing only athin process to the dorsum of this structure.
A large median septum extends posterodorsally on the medialside of the premaxilla (Fig 3B). When articulated, the premax-illae meet along this median septum, forming an “s-loop” likethat observed in Corythosaurus and L. lambei, but absent in H.altispinus and all species of Parasaurolophus (Weishampel,1981).
Nasal—As in Corythosaurus, Hypacrosaurus, and Olorotitan(Godefroit et al., 2003) the nasal (Fig. 3E) of Velafrons is arelatively large, flattened element that forms the bulk of thedorsal portion of the crest. It is bounded anteriorly by the pre-maxilla, ventrolaterally by the prefrontal, and posteroventrallyby the frontal. For the purposes of description, the nasal can besubdivided into a thickened, transversely expanded ventral por-tion and a relatively flattened dorsal portion that comprises themajority of the element.
The ventral portion is transversely narrow posteriorly and ex-pands anteriorly in association with the internal chamber of thenasal vestibule (Fig. 2). The nasal-frontal contact is formed bythe posterolateral corner of the nasal base slotting into a right-angled notch present on the frontal such that the frontal platformunderlaps the nasal. The frontal supports the base of the nasal inall other North American lambeosaurines, except that in thesetaxa—including juveniles—the posterolateral contact is notsquared as in Velafrons (Fig. 3E), but instead is rounded to sup-port a nasal with differing morphology. In Velafrons, the nasal-prefrontal contact is more complex, with the nasal slotting into awell-developed clamp formed by the prefrontal above and thefrontal below. The premaxillary contact is less certain, since thepremaxillary portion of this union is disarticulated.
As preserved, the flattened dorsal portion of the nasal isrounded and roughly fan-shaped. Medially, the nasal forms a flatcontact surface for its counterpart that extends throughout theposterior half and most of the height of this element. The onlyexception is a well-developed, ventrally located depression thatmay be incipient development of the narial system into the nasal.The top of the nasal in CPC-59 is not preserved from the mid-point to its anterior edge (Fig 3E). However, a distinctive contactsurface preserved on the dorsal process of the premaxilla indi-
←
FIGURE 4. Comparison of skull and crest development between threetaxa of juvenile lambeosaurines. A, Velafrons coahuilensis; B, Corytho-saurus casuarius; C, Lambeosaurus sp. Dark grey regions represent thearea of the crest occupied by the nasal whereas the light grey shows theextent of the premaxilla for each specimen. Abbreviations: na, nasal; pf,prefrontal; pmd, dorsal premaxilla process; pml, lateral premaxilla pro-cess; pnf, crest fontanelle. Skulls not to scale. Note that the nasal anteriorprocess of Velafrons is illustrated with single projection because there isonly osteological evidence for one projection. However, given the closephylogenetic affinity to Corythosaurus and Hypacrosaurus, Velafronsmay similarly possess a bifurcated anterior nasal process.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 27, NO. 4, 2007920

cates that an elongate process of the nasal extended forward toclasp the dorsal premaxillae laterally (Fig. 4). As reconstructedin Figure 2, the nasal possesses a single anteriorly projectingprocess that overlaps the dorsal premaxillary process becausethat is the only process of which there is osteological evidence.However, both Corythosaurus and Hypacrosaurus have a bifur-cated anterior process that may well have been present but notpreserved on CPC-59, especially given the overall similarity innasal morphology with the latter two taxa. The anteroventralportion of the nasal is composed of a nearly vertical, thickenedmargin that makes up the lateral edge of an anteroventrally fac-ing premaxilla-nasal fontanelle visible on the lateral side of theskull (Fig. 2) as on Corythosaurus, Lambeosaurus, and Hypac-rosaurus (Fig. 4). This latter taxon differs from other taxa in thatthe fontanelle closes quite early in development (Evans et al.,2005). Posteriorly, the crest rises vertically a short distance fromthe base of the nasal and then curves posterodorsally to form thecurved rear margin of the crest (Fig 3E). An equivalent mor-phology is present in juveniles of both C. casuarius and H. alti-spinus (Evans et al., 2005).
Maxilla—The maxilla (Fig. 5A, B) is typical of lambeosaurinehadrosaurids, being relatively gracile with a pronounced dorsalprocess near its midsection and a long ramp-like premaxillaryshelf anteriorly (Fig. 5A). A maxillary foramen occurs at thebase of the dorsal process (Fig. 2). The dorsal process is shapedlike a right-angled triangle in lateral view, with a steeply angledanterior margin and a vertical posterior margin. It rises to aheight nearly equal to the depth of the main maxilla body some-what posterior to the midpoint of the maxilla. The majority ofthe posterior side of this process articulates with the palatine,and a small portion of this element is preserved on the rightmaxilla. A large opening, the maxillopalatine foramen, occurs atthe junction of these elements near the base of the dorsal pro-cess. A third foramen occurs on the lateral side of the maxillarybody, immediately below the jugal process. Hypacrosaurus ste-bingeri (MOR 549) has two foramina at this location.
The jugal articulation is represented by a ventral ridge andlarge facet on the lateral side of the dorsal process. It appearsthat the jugal covered most of the lateral side of the dorsal pro-cess, not unlike that in other lambeosaurines.
The ectopterygoid ridge forms the prominent overhang seenon the posterior half of the maxilla in lateral view. The ectoptery-goid shelf is wider than the premaxillary shelf throughout itslength then rounds off abruptly at the posterior margin of themaxilla. However, the shelf appears to be shorter anteroposte-riorly than in Corythosaurus (Ostrom, 1961) or H. stebingeri(MOR 549), possibly caused by posterior positioning of the dor-sal process. The maxilla of Parasaurolophus cf. cyrtocristatus(UMNH VP 16666.1) is more similarly shaped, including an ap-parent posterior shift of the dorsal process. The palatine processis located on the medial side of the ectopterygoid shelf, anteriorto the pterygoid process (Fig. 5A, B).
The virtually complete left maxilla contains approximately 33tooth positions, each corresponding to a medially positioneddental foramen. The tooth row (∼248 mm in length) is distinctlysinusoidal in ventral view, with the most medial portion in theanterior region and the most lateral portion located posteriorly,a condition seen in other lambeosaurines.
Jugal—Overall, the jugal (Fig. 5C) most closely resembles thatof the Russian lambeosaurine Amurosaurus riabinini (Godefroitet al., 2004). The element contacts the maxilla anteriorly, thelacrimal and postorbital dorsally, and the quadratojugal andquadrate posteriorly. The outer margin of the anterior process isnot preserved so the morphology of this region cannot be fullyreconstructed. However, the preserved portion of the anteriorprocess suggests that this structure was dorsoventrally symmetri-cal, as in Corythosaurus (Fig. 4; Ostrom, 1961). Parasaurolophuscf. cyrtocristatus (UMNH VP 16666.1) is dissimilar to Velafrons
because its anterior process is tall, anteroposteriorly compressed,and possesses a virtually straight anterior margin. A prominentlacrimal facet is present on the dorsal margin of the anteriorprocess, closely resembling that of Corythosaurus, but differingdrastically from the large lacrimal suture interfingering of P.walkeri (ROM 768). The postorbital process is triangular at itsbase and simultaneously flattens and twists dorsally such that ittransitions from facing anteriorly to facing laterally. The postor-bital process is broken and offset 1 cm above the base, but theapproximate angle of ascent is 75º (Fig. 5C).
The jugal of Velafrons is distinctive in the position of the pos-teroventral flange and the size and shape of the posterior pro-cess. The posteroventral flange is poorly developed and moreposteriorly positioned compared to other North American lam-beosaurines, particularly Lambeosaurus lambei (e.g., AMNH5373) and both species of Hypacrosaurus (e.g. MOR 549, TMP82.10.01). Velafrons, like L. lambei and juvenile H. altispinus(CMN 2246) possesses a curved outer margin of the posteriorprocess, but unlike L. lambei the curvature extends dorsally until
FIGURE 5. Left maxilla of Velafrons coahuilensis in A, lateral view;and B, medial view. Right jugal of Velafrons coahuilensis in C, lateralview. Abbreviations: dp, dorsal process; ectp, ectopterygoid process; ja,jugal articulation; lf, lachrymal facet; nf, nutrient foramina; palp, palatineprocess; pms, premaxillary shelf; pop, postorbital process; pp, posteriorprocess; ptp, pterygoid process. Scale bar equals 5 cm.
GATES ET AL.—NEW LAMBEOSAURINE FROM MEXICO 921

the apex is directly above the posteroventral flange. This pro-nounced curvature of the posterior process results in a narrowlateral temporal fenestra (Fig. 4).
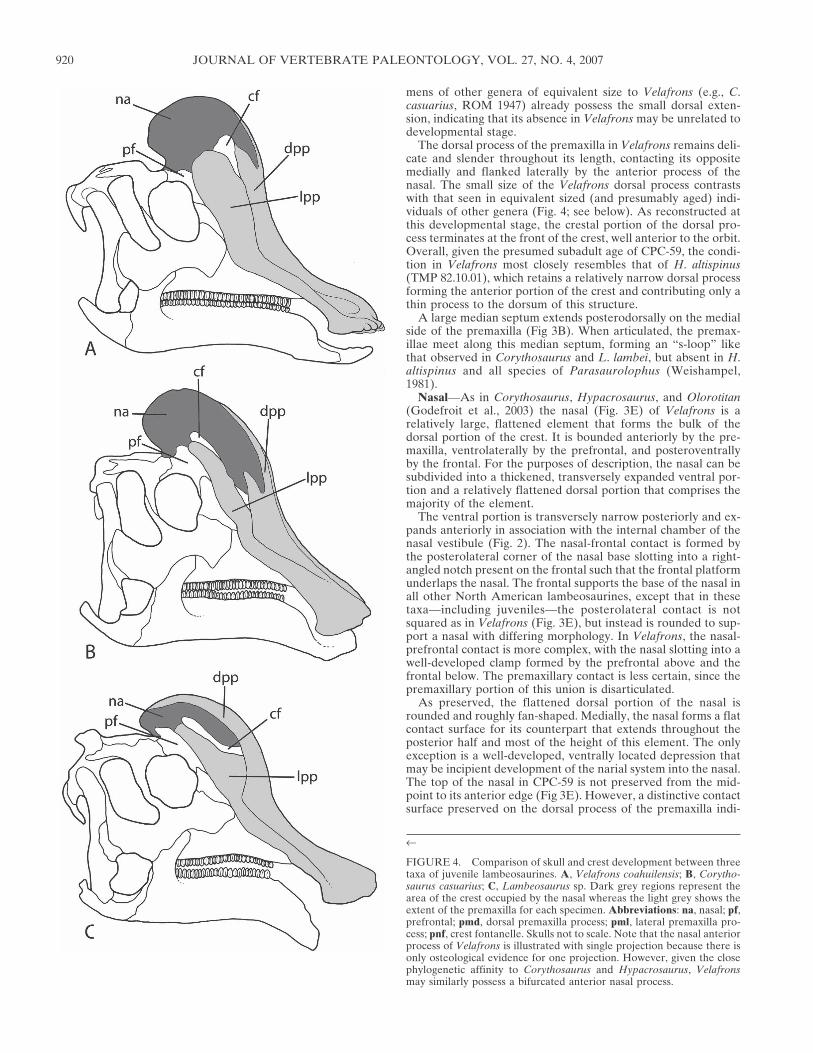
Prefrontal—The prefrontal (Figs. 6; 7A, B) is located on theanterolateral region of the skull roof. Laterally, the prefrontalmakes up the rugose anterodorsal portion of the orbital rim inassociation with the co-ossified supraorbital (Maryanska and Os-mólska, 1979). As in other lambeosaurine taxa, the prefrontalinterdigitates with the postorbital posterolaterally, forming araised bony mound. Medially, the prefrontal articulates with thefrontal, nasal, and possibly the premaxilla, although the connec-tion with the latter element is uncertain. Of these elements, thecontact with the frontal is by far the most extensive, dominatingthe posterior and ventromedial sides of the prefrontal. Contraryto the morphology seen on Amurosaurus and Jaxartosaurusaralensis (Godefroit et al., 2004), the prefrontal does not formany portion of the frontal platform, but instead overlaps thefrontal medially for a short distance before it deflects slightlydorsally, forming the roof of a low angled bony clamp for thebase of the nasal (Fig. 6). In contrast, the prefrontal in otherlambeosaurines of comparable size is already expanded and de-flected dorsally to almost a 90º angle, and developed anteropos-teriorly over the frontal to form a shallow dish shape. The com-parative ontogeny of this structure is discussed more fully below.
Frontal—The frontals (∼75 mm mediolaterally, 75 mm antero-posteriorly sensu Godefroit et al. [2004]) form the anteromedialregion of the skull roof, articulating with the prefrontals, theorbitosphenoids and laterosphenoids, the postorbitals, and pari-etals. Most of the anterior portions of the frontals are missingfrom CPC-59. A small section preserved on the right frontal (Fig.7A, D) indicates that the frontal-nasal contact is virtually square(92º angle between the lateral and posterior margins). H. stebin-geri (MOR 553S) has a slightly square frontal-nasal contact,whereas Corythosaurus (TMP 82.19.72) is well rounded posteri-orly, and the Asian lambeosaurines Amurosaurus and Jaxarto-saurus possess two posterior arches (Godefroit et al., 2004). Theprefrontals articulate with the anterolateral side of the frontals.As described previously, a slight overlap between the two ele-ments forms a clamp that grips the ventrolateral extension ofthe nasal base. As in other lambeosaurines (Horner et al., 2004),the postorbital also articulates with the lateral side of the frontal,thereby excluding the latter from contributing to the orbitalrim (Fig. 7A, B, D, E). The frontals are depressed medially atthe triradiate contact of these three elements. Medial to thisdepression is the frontal doming characteristic of juvenile lam-beosaurines (Gilmore, 1937; Godefroit et al., 2004; Fig. 7D). Theparietals contact the frontals posteriorly via a long, V-shapedsuture. It is unclear if the frontals bifurcate posteriorly to accept
an interfrontal process from the parietal as in other hadrosau-rids.
Postorbital—As is typical of hadrosaurids, the postorbital is atriradiate structure (Fig. 7D) contacting the prefrontals, frontals,laterosphenoids, parietals, and squamosals. Anteriorly, it articu-lates with the prefrontal via a robust contact that forms an ob-servable mound (Fig. 7D). The lateral side of the postorbital isrugose, marking the posterior half of the dorsal orbital rim andco-ossification with the supraorbital (Maryanska and Osmolska,1979). The frontals articulate medially in a straight, interdigitat-ing suture (Fig. 7D). A large concave depression is present onthe posteroventral surface of the postorbital, located almost di-rectly ventral to the squamosal process and anteromedial to thebase of the jugal process. Just posterior to the jugal process andventral to the squamosal process, an additional small processprojects posterolaterally on the right postorbital (the same areais not preserved on the left). This process is dorsoventrally com-pressed, but the entire morphology of this structure is unclear asit is broken near the base. This unusual projection has not beendescribed from any other lambeosaurine taxon and may repre-sent an autapomorphy of Velafrons; however, due to the frag-mentary nature of preservation, this potential character is cur-rently unverifiable.
The laterosphenoid articulates with the postorbital in a pocketat the junction of the three branches, and the parietal immedi-ately dorsal to the laterosphenoid. The shape and position of thesquamosal process is autapomorphic for Velafrons (Fig. 2; 7E).The base of the process is positioned toward the dorsal surface ofthe postorbital. The process ascends at a steep angle and termi-nates near the level of the raised squamosal shelf. This confor-mation differs from other hadrosaurids, such as Corythosaurus(Ostrom, 1961) and Parasaurolophus (Sullivan and Williamson,1999) where the squamosal process ascends posterodorsally at ashallow angle to contact a small squamosal shelf (Fig. 4). Con-sequently, the dorsal rim of the infratemporal fenestra in Vela-frons is raised above the dorsal margin of the orbit by approxi-mately 2.5 cm (Fig. 2; 7E). The condition resembles, yet is alsodistinct from, that of Amurosaurus (Godefroit et al., 2004),where it is also considered an autapomorphy, and H. stebingeri(MOR 553S). The squamosal process bifurcates posteriorly, mid-way through its length (Fig. 7E).
Parietal—In dorsal view, the parietal (Fig. 7D) is a slightlyhourglass shaped element that extends anterolaterally to form anelongate, angled contact with the frontals medially and the post-orbitals laterally. As is typical, there is an extensive ventral con-tact with the laterosphenoid. In all other hadrosaurids, an inter-frontal process typifies the medial junction of these arms. Thisstructure is not seen in the type of Velafrons, although it is mostlikely due to postmortem crushing and diagenesis. The body ofthe parietal constricts posteriorly in conjunction with the supra-temporal fenestrae, and widens at the articulation with the su-praoccipital and the squamosals. The posterodorsal region of theparietal bears a large vertical process that bisects the paired,raised squamosals (Fig. 7D). Similar to Hypacrosaurus, the pro-cess extends anteriorly over the supratempoal fenestrae to residedorsal to the rear margin of the frontals.
Squamosal—The paired squamosals (Fig. 7) form the majorityof the posterodorsal region of the skull. However, in dorsal view,the squamosals are separated by a thin, median extension of theparietal, and only seen to coalesce when one views the skull roofin posterior view (Fig. 7F). This condition differs from Amuro-saurus (Godefroit et al., 2004) and Hypacrosaurus stebingeri(MOR 553S) that fully separate the squamosals by the parietal,and is most similar to Corythosaurus (Ostrom, 1961). In dorsalview, the squamosals form a raised shelf by a forward expansionof the median rami over the parietal and supratemporal fenes-trae (Fig. 7D). This anterior projection of the squamosal is morestrongly developed in Velafrons than in Corythosaurus, although
FIGURE 6. Anterior view of articulated braincase showing detail ofprefrontal “clamp”. Abbreviations: fns, frontal-nasal suture; pfc, pre-frontal “clamp”. Scale bar equals 5 cm.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 27, NO. 4, 2007922
a large squamosal shelf is also present in H. stebingeri (MOR553S). Parasaurolophus tubicen bears a large posterodorsally di-rected process on the squamosal instead of the squamosal shelf(Sullivan and Williamson, 1999). The transversely broad postor-bital process extends anteriorly almost to the body of the post-orbital, participating in the formation of the large canopy-likestructure over the supratemporal fenestrae. The quadrate coty-lus lies at an angle of about 45º to the horizontal, and is posi-tioned on the ventrolateral border of the squamosal. Both theprecotyloid and postcotyloid processes are broken at their junc-tion with the quadrate cotylus. In posterior view, a small tongue-like supraoccipital facet overlaps the supraoccipital 1 cm (sensuGodefroit et al., 2004).
Palatoquadrate
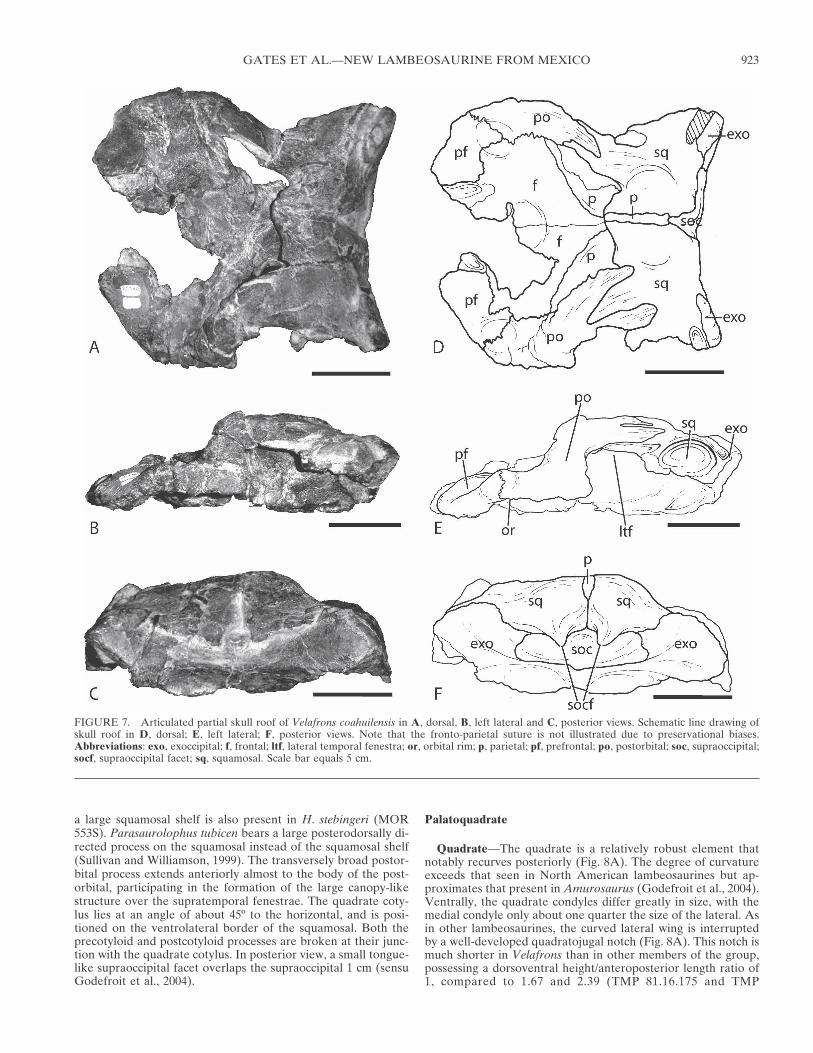
Quadrate—The quadrate is a relatively robust element thatnotably recurves posteriorly (Fig. 8A). The degree of curvatureexceeds that seen in North American lambeosaurines but ap-proximates that present in Amurosaurus (Godefroit et al., 2004).Ventrally, the quadrate condyles differ greatly in size, with themedial condyle only about one quarter the size of the lateral. Asin other lambeosaurines, the curved lateral wing is interruptedby a well-developed quadratojugal notch (Fig. 8A). This notch ismuch shorter in Velafrons than in other members of the group,possessing a dorsoventral height/anteroposterior length ratio of1, compared to 1.67 and 2.39 (TMP 81.16.175 and TMP
FIGURE 7. Articulated partial skull roof of Velafrons coahuilensis in A, dorsal, B, left lateral and C, posterior views. Schematic line drawing ofskull roof in D, dorsal; E, left lateral; F, posterior views. Note that the fronto-parietal suture is not illustrated due to preservational biases.Abbreviations: exo, exoccipital; f, frontal; ltf, lateral temporal fenestra; or, orbital rim; p, parietal; pf, prefrontal; po, postorbital; soc, supraoccipital;socf, supraoccipital facet; sq, squamosal. Scale bar equals 5 cm.
GATES ET AL.—NEW LAMBEOSAURINE FROM MEXICO 923
2002.12.74, respectively) for other lambeosaurine quadrates.Hypacrosurus stebingeri possesses a quadratojugal notch that dif-fers from that of Velafrons in that the ventral border of theformer taxon arches dorsally at the anterior opening of thenotch, whereas Velafrons is narrower and the ventral borderremains flat. A weak depression outlines the entire quadratoju-gal notch, presumably indicating that the quadratojugal com-pletely covered the paraquadratic foramen (Weishampel et al.,1993).
The well-developed pterygoid wing is roughly quadrangular inshape (Fig. 8B). It consists of a steeply angled (∼70º ) concavedorsal margin, a straight and steeply angled medial edge (60º),and finally a slightly convex ventral border. Specimens of H.stebingeri (e.g. MOR 553) have a quadrate morphology similar tothat of Velafrons, whereas other lambeosaurines such as Cor-ythosaurus, Lambeosaurus, and Amurosaurus, possess a ptery-goid wing that regularly slopes at approximately 45º, and arounded medial margin (Parks, 1923; Ostrom, 1961; Godefroit etal., 2004). A vertically elongate fossa is present posteromediallyat the junction of the pterygoid wing and the shaft. On the pos-terior side of the shaft, a small spur projects medially, level withthe ventral border of the pterygoid wing (Fig. 8B), appearingotherwise only in H. stebingeri (MOR 553). In all other observedlambeosaurine taxa, the spur is represented by only a minutebump in the same area of the quadrate.
Braincase
The braincase of CPC-59 is poorly preserved. Fragments ofthe supraoccipital, exoccipital, and basisphenoid are completeenough to describe portions of their morphology. In contrast, theorbitosphenoid and laterosphenoid, do not provide sufficientmorphology to warrant description. None of the cranial nerveforamina or braincase sutures can be observed.
Supraoccipital—The supraoccipital (Fig. 7C, F) is a stout el-ement, triangular in posterior view. It is flanked laterally andventrally by the exoccipitals, and the parietal and squamosaldorsally. Overall, the morphology and relationships of this ele-ment do not differ from that of other lambeosaurine taxa (Os-
trom, 1961; Gilmore, 1937). Poor preservation of the ventral sideof the supraoccipital prevents further description.
Exoccipital—Only the dorsomedial portions of this elementare preserved (Fig. 7C, F) and observed to articulate with thesquamosals, supraoccipital, and each other. The ventral-mostarticulation of the exoccipitals along the midline marks thedorsal margin of the foramen magnum. They form a deepershelf in Parasaurolophus cf. cyrtocristatus (UCMP 143270), thanin Velafrons, pushing the foramen magnum further into theskull. However, the depth of the foramen magnum may varythrough ontogeny, since Horner and Currie (1994:fig. 21.7f) il-lustrate a juvenile Hypacrosaurus with a relatively shallow fora-men magnum.
Basisphenoid—Although the anterior half of the basisphe-noid is severed, much of the posterior portion is preserved. Twolarge semi-ovoid sutures that articulate with the basioccipitalaccentuate the posterior aspect of the basisphenoid. The basi-pterygoid processes descend ventrolaterally, separated at anangle of 65º. The distal ends of the processes are rounded, lack-ing the lateral projections present on Hypacrosaurus stebingeri(MOR 553S) and Corythosaurus (Ostrom, 1961). A wing-likeprocess present on the right lateral side may represent the alarprocess of the basisphenoid.
Lower Jaw and Hyoid Apparatus
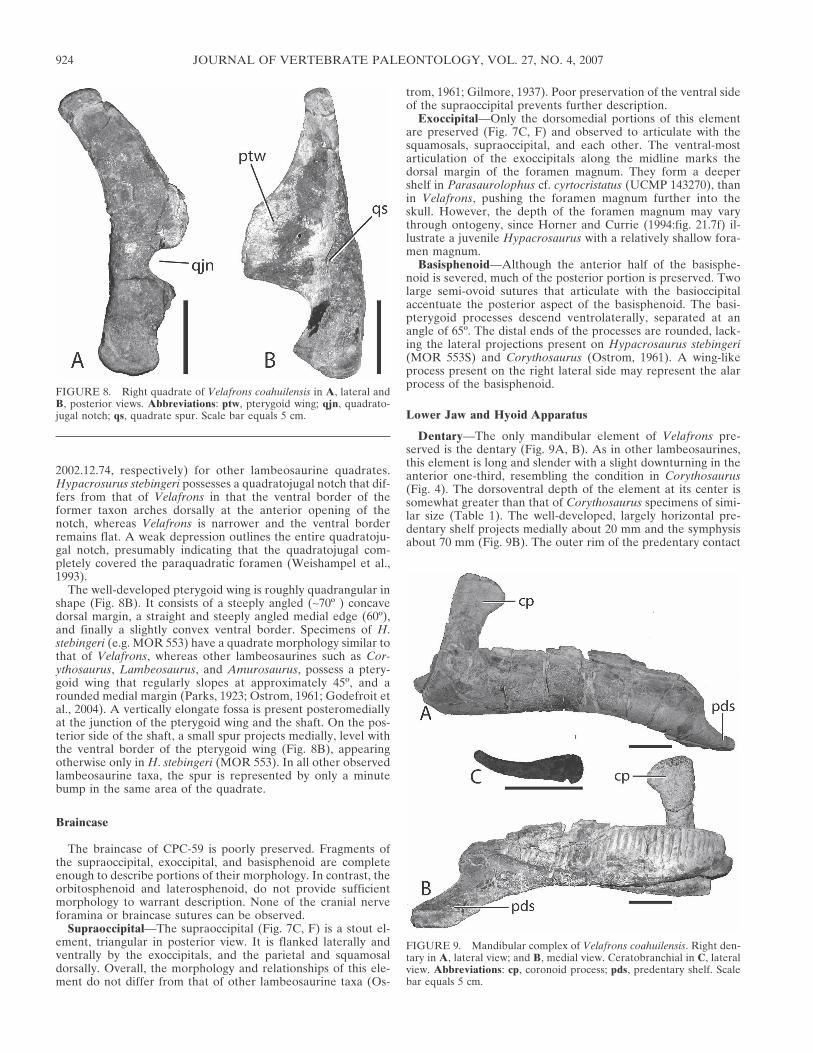
Dentary—The only mandibular element of Velafrons pre-served is the dentary (Fig. 9A, B). As in other lambeosaurines,this element is long and slender with a slight downturning in theanterior one-third, resembling the condition in Corythosaurus(Fig. 4). The dorsoventral depth of the element at its center issomewhat greater than that of Corythosaurus specimens of simi-lar size (Table 1). The well-developed, largely horizontal pre-dentary shelf projects medially about 20 mm and the symphysisabout 70 mm (Fig. 9B). The outer rim of the predentary contact
FIGURE 8. Right quadrate of Velafrons coahuilensis in A, lateral andB, posterior views. Abbreviations: ptw, pterygoid wing; qjn, quadrato-jugal notch; qs, quadrate spur. Scale bar equals 5 cm.
FIGURE 9. Mandibular complex of Velafrons coahuilensis. Right den-tary in A, lateral view; and B, medial view. Ceratobranchial in C, lateralview. Abbreviations: cp, coronoid process; pds, predentary shelf. Scalebar equals 5 cm.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 27, NO. 4, 2007924
surface curves sharply posteriorly to become confluent with den-tary body. In lateral view, the margin of this structure upturnstoward the dorsal edge of the body at almost a 90º angle, asteeper angle than observed in most other taxa.
The tooth row extends well posterior to the coronoid processand includes 34 tooth positions. No teeth are preserved in thedentary so the number of teeth per alveolus cannot be ascer-tained. The coronoid process (Fig. 9A) angles weakly anteriorlyand made up mostly of the dentary. The dorsal portion of thisprocess possesses an anterior flange while the posterior region isflush with the shaft. This condition is similar to that present inHypacrosaurus stebingeri (MOR 549), although the dorsal mar-gin in that specimen curves less than in Velafrons. Medially, alarge facet indicates that the splenial extended about half thelength of the dentary (Fig. 9B). Below this groove, a portion ofthe angular is present, demonstrating that it wrapped around themedial side of the dentary, as in other hadrosaurids (Weishampelet al., 1993).
Hyoid—The morphology of the ceratobranchial (Fig. 9C) issimilar to that of other hadrosaurids (Ostrom, 1961). The ante-rior portion is dorsoventrally expanded, but unlike other taxa,the head of the hyoid is rounded rather than square. The elementtapers posteriorly and bends dorsally to terminate in a blunt,rounded end, approximately 80 mm long.
PHYLOGENETIC ANALYSIS
A phylogenetic analysis was performed using the Evans andReisz (2007) character matrix and a combination of MacCladeversion 4.06 (Maddison and Maddison, 2000) and PAUP version4.0b10 (Swofford, 2002). In the Majority Rules consensus tree of27 most parsimonious trees produced from the branch andbound search algorithm (Fig. 10; 18 taxa [Nipponosaurus wasremoved from this analysis, see Evans and Reisz (2007)] withProbactrosaurus gobiensis, Telmatosaurus transsylvanicus, andGryposaurus incurvimanus as outgroups, 94 characters, treelength 113, CI: 0.89, RI: 0.92, RC: 0.82), Velafrons is positedinside a polytomy with Corythosaurus, Olorotitan, a clade con-taining the two species of Hypacrosaurus and a clade containingthe two species of Lambeosaurus. Within this polytomy, all ofthe taxa share fan-shaped crests, but as has been noted through-out this manuscript the development of the crest within Vela-frons is more similar to Corythosaurus and Hypacrosaurus thanit is Lambeosaurus or Olorotitan. Both Corythosaurus and Hyp-acrosaurus have long forking anterior nasal processes that sur-round the dorsal process of the premaxilla. The dorsal process, inturn, onlaps the nasal between these nasal processes (Fig. 4).This relationship between the anterior nasal process and thedorsal premaxilla process is unique to these two taxa (Evans etal., 2005). Velafrons shares the long anterior nasal process with
Corythosaurus and Hypacrosaurus, but the type specimen ofVelafrons does not preserve this region of the crest therefore, theexact morphology cannot be determined. Interestingly, eventhough Velafrons and Amurosaurus possess many similar char-acteristics including jugal morphology and a raised postorbitalposterior process, the latter taxon is posited as more primitivethan Velafrons.
DISCUSSION
The holotype specimen of Velafrons appears to represent asubadult individual. This assessment is based on relative size anddegree of crest development, as well as comparison with overallskull size with those of other juvenile lambeosaurine specimens.Yet, despite incomplete crest development, the specimen pos-sesses a number of taxonomically significant characters that canbe differentiated from ontogenetic variation. Horner (1992; andthis study) observed that, other than size, the morphology ofjugals and quadrates do not change significantly during growth,and further that these elements are morphologically virtuallyindistinguishable from adult elements at subadult growth stages.Therefore, diagnostic characters present in these elements ofVelafrons are here regarded to be valid and not the result ofontogenetic variation. Conversely, the postorbital does undergosignificant changes through ontogeny. In young individuals lack-ing full crest development, the squamosal process of the postor-bital lies nearly horizontal, but during growth the squamosalprocess angles dorsally because the caudal-most margin of thisprocess articulates with a raised squamosal shelf. The degree ofangling depends on the height of the squamosal shelf in a par-ticular genus of lambeosaurine. Velafrons, however, is outsidethe known range of variation in this feature, relocating the baseof the squamosal process to the dorsal surface of the postorbital.The squamosal process then rises near vertically and levels out tocontact the squamosal shelf.
Comparison With ?Lambeosaurus laticaudus
Morris (1981) described ?Lambeosaurus laticaudus from apartial lambeosaurine skull found in the El Gallo Formation ofBaja California. Skull elements includes a partial left premaxilla,as well as a maxilla, dentary, and jugal. He assigned this speci-men to the genus Lambeosaurus based on a single character:long open external nares of the premaxilla, which he posited tobe a generic autapomorphy of Lambeosaurus. However, numer-ous other lambeosaurines have elongate open external nares,including North American forms such as Corythosaurus andHypacrosaurus. Further, the oral margin of the premaxilla is
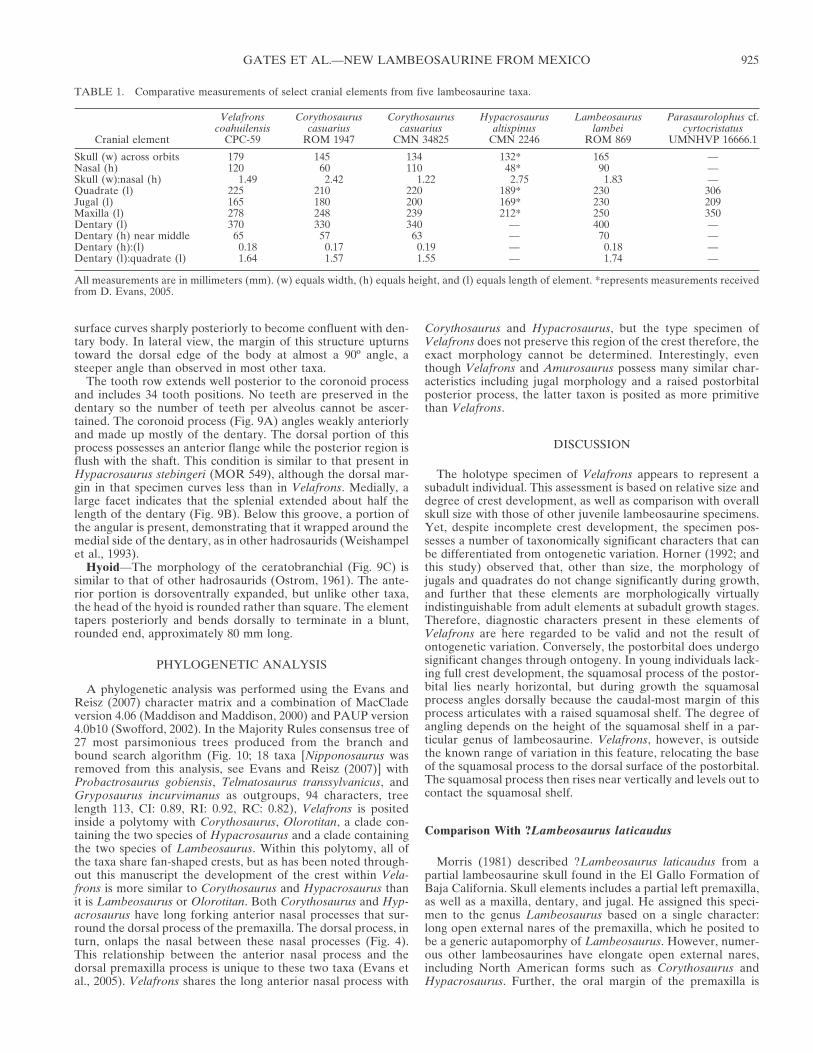
TABLE 1. Comparative measurements of select cranial elements from five lambeosaurine taxa.
Cranial element
Velafronscoahuilensis
CPC-59
Corythosauruscasuarius
ROM 1947
Corythosauruscasuarius
CMN 34825
Hypacrosaurusaltispinus
CMN 2246
Lambeosauruslambei
ROM 869
Parasaurolophus cf.cyrtocristatus
UMNHVP 16666.1
Skull (w) across orbits 179 145 134 132* 165 —Nasal (h) 120 60 110 48* 90 —Skull (w):nasal (h) 1.49 2.42 1.22 2.75 1.83 —Quadrate (l) 225 210 220 189* 230 306Jugal (l) 165 180 200 169* 230 209Maxilla (l) 278 248 239 212* 250 350Dentary (l) 370 330 340 — 400 —Dentary (h) near middle 65 57 63 — 70 —Dentary (h):(l) 0.18 0.17 0.19 — 0.18 —Dentary (l):quadrate (l) 1.64 1.57 1.55 — 1.74 —
All measurements are in millimeters (mm). (w) equals width, (h) equals height, and (l) equals length of element. *represents measurements receivedfrom D. Evans, 2005.
GATES ET AL.—NEW LAMBEOSAURINE FROM MEXICO 925

unlike other species of Lambeosaurus and more closely re-sembles that of Hypacrosaurus, exhibiting a highly denticulatedoral margin and rounded, down-turned lateral edge.
A deep tail not observed in other species of Lambeosaurusdefines the species name, laticaudus. However, a similarly deeptail is also present in Hypacrosaurus altispinus (Brown, 1913). Infact, Morris (1981:457) states that, “Were it not for the opennarial crest ?L. laticaudus might easily have been called Hypac-rosaurus sp. because this latter genus is the only hadrosauridthought to have a tall, flattened tail.” We consider the taxon?Lambeosaurus laticaudus to be a nomen dubium because thetype specimen does not have any synapomorphies that unam-biguously unite it with any lambeosaurine genus. Nor does itpossess autapomorphies that would permit erection of a newgenus. Based on the similarities of the premaxilla and the deeptail, this taxon may represent the southernmost extent of thegenus Hypacrosaurus (see also Evans and Reisz, in press), butany such referral must await discovery of additional material.From this point forward, we refer to ?L. laticaudus as the “Bajalambeosaur.”
Even though the Baja lambeosaurine specimen is fragmentary,comparison with Velafrons reveals that the two Mexican taxa are
likely not equivalent. The premaxilla of Velafrons does not sharethe rounded down-turned morphology of the Baja specimen,instead possessing a uniformly arcuate oral margin with a sharpconstriction on the lateral edge. Both taxa share a highly den-ticulated oral margin, but the amount of denticulation is variablebetween individuals and is not a diagnostic character at the genuslevel. It is not clear if Velafrons shares the elongate, open naresof the Baja lambeosaurine because they are poorly preserved.The remainder of the Baja lambeosaurine skull is too fragmen-tary for further comparison.
Comparative Ontogeny
Since Velafrons is currently known from a single specimen,there is little than can be said about the ontogenetic trajectory ofthis taxon. Nevertheless, a number of skull features are consis-tent with a juvenile stage of development that closely resemblesthat of other lambeosaurines (Table 1). A number of authorshave studied ontogenetic changes in crest growth and develop-ment within various lambeosaurine taxa (Parks, 1923; Sternberg,1935; Dodson, 1975; Evans et al., 2005). Thus, a lengthy discus-sion is not given here. However, it is important to establish key
FIGURE 10. Majority rules consensus cladogram showing phylogenetic position of Velafrons coahuilensis. The tree was produced using the Evansand Reisz (in press) character matrix, tree generation in PAUP ver. 4.0b10 (Swofford, 2002), and tree manipulation in MacClade ver. 4.06 (Maddisonand Maddison, 2000). Tree statistics: 18 taxa, 94 characters, Length: 113, CI: 0.89, RI: 0.92, RC: 0.82. Numbers on stems are boot strap values of 1000replicates.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 27, NO. 4, 2007926

similarities shared by Velafrons and juveniles of other lambeo-saurine taxa so as to highlight potentially unique developmentalcharacteristics.
Based on the absolute size of the quadrate and nasal and to alesser extent the dentary (Velafrons is larger by 30 mm), Vela-frons appears to be close to the developmental stage illustratedby the Corythosaurus specimen CMN 34825 (Fig. 4), although,there are greater differences between the size of other skull el-ements in these specimens as well as in other lambeosaurineindividuals listed in Table 1. The transverse width of the skullroof does not appear to be consistent across lambeosaurines. Theskull roof of Velafrons is wider than all other individuals of simi-lar size and developmental stage. However, it is possible thatminor crushing of the measured skulls could account for at leastsome of these deviations.
There appears to be no correlation to the height of the nasaland the width of the skull at the orbits. In general, the crestdevelopment in Velafrons parallels other lambeosaurines, in thatthe nasal is positioned dorsal to the frontals, and the premaxillapossesses lateral and dorsal processes that extend rearward tocontact the nasal. More specificly is the presence of a long an-terior process of the nasal (Evans et al., 2005) shared exclusivelywith Corythosaurus and Hypacrosaurus. The premaxilla-nasalfontanelle, located on the lateral side of the crest between thenasal and the lateral premaxillary process, is open at this stage ofdevelopment in Velafrons and Corythosaurus. Hypacrosaurus,on the other hand, closes the fontanelle early in its development.Consequently, this is one of only two characters that can be usedto distinguish immature individuals of Hypacrosaurus from Co-rythosaurus (Evans et al., 2005).
Velafrons possesses the frontal-prefrontal “clamp” supportstructure present in immature individuals of Corythosaurus,Lambeosaurus, and Hypacrosaurus (e.g., TMP 86.26.31), but ap-pears to have retained this feature longer through ontogeny. Asthe crest develops in these taxa, the prefrontal elongates antero-posteriorly, extending its medial wall dorsally to support the baseof the growing nasal bone. Simultaneously, the nasal loses theventrolateral extension and forms a bowl-like base, residing inthe foundation formed from the modified prefrontal. Yet, asmentioned above, the nasal of Velafrons maintains the ventro-lateral expansion and the frontal-prefrontal clamp, in spite of theincongruence in prefrontal development between it and otherfan-crested lambeosaurines of equivalent crest size.
Sternberg (1935) described Tetragonasaurus cranibrevis—amisidentified juvenile Lambeosaurus (Dodson, 1975; Evans etal., 2005)—that possesses a much smaller nasal, yet more ad-vanced modification of the prefrontal than Velafrons. In addi-tion, a juvenile skull roof (ROM 694) only 145 mm wide at theorbits, already possesses an upturned prefrontal. Without agrowth series of Velafrons, it is unclear when (or even if) theprefrontal would have transformed into the upturned condition.Several possible growth trajectories may be hypothesized forfuture growth of the crest, given the current state of develop-ment. One alternative is that the crest remained small and didnot develop the advanced supporting structure of other corytho-saurs. Another possibility is that the crest may undergo a periodof rapid modification late in ontogeny. A third possibility is thatVelafrons was larger at adult size than other lambeosaurines andthe apparent delay in development is an artifact of more rapidgrowth rates producing a larger skull when compared to otherNorth American lambeosaurines of equivalent age. In otherwords, if Velafrons reached larger adult body size by increasinggrowth rate rather than prolonging growth as has been demon-strated in other dinosaurs (Erickson et al., 2001), then otherequivalently sized juvenile lambeosaurine taxa would be ex-pected to be to be older, more developed individuals. Ultimately,this may explain why the holotype of Velafrons appears ontoge-netically less developed than other lambeosaurines of equivalent
size. This last alternative may be the most likely of the threesince, within the North American Western Interior Basin, south-ern hadrosaurids in general appear to be larger than their north-ern counterparts (personal observation), although no specificstudy has quantitatively verified a size difference.
All of the changes associated with crest development men-tioned above drastically alter the skull morphology between ju-venile and adult growth stages. Therefore, specific apomorphiesbased on crest characteristics in juvenile lambeosaurines are notreliable because those features alter through growth. The apo-morphies distinguishing Velafrons from all other lambeosaurinetaxa are based on skull elements that alter little with crest de-velopment of this group (see Dodson, 1975; Evans et al., 2005).As a result, we hypothesize that the diagnostic features of Vela-frons will be present on both juvenile and adult skulls, becausethese same bones undergo minimal shape change through on-togeny in other closely related taxa. Although, once adult fossilmaterial of Velafrons is discovered, more apomorphies based onadult crest morphology may further differentiate this taxon.
Biogeography
Velafrons lived near the southernmost point of the NorthAmerican continent around 73.5 Ma (Eberth et al., 2004). TheBaja lambeosaurine may have occupied a similar latitude be-cause the terrestrial deposits of the “El Gallo” Formation havedrifted northward, upwards of 2000 km, along the San AndreasFault since the Late Cretaceous (Page and Engebretson, 1984;Bartow, 1991). Within the Parras Basin, only one other hadro-saurid has been described, a hadrosaurine from the contempo-raneous Sabinas Basin referred to Kritosaurus (Kirkland et al.,2006).
The only hadrosaurid fauna known from this time period inNorth America is found in the southern region of the continentdue to the Bearpaw Transgression inundating portions of Mon-tana and Alberta, (Tsujita and Westermann, 1998), while an un-conformity developed in southern Utah (Bowers, 1990; Eaton etal., 1999). Parasaurolophus tubicen (Wiman, 1931; Sullivan andWilliamson, 1999) and Kritosaurus (Horner, 1992; consideredNaashoibitosaurus in Horner et al., 2004) are found in sedimentsof the Kirtland Formation of northern New Mexico, equivalentin age to the Parras Basin (Fig. 11; although Horner [1992] ten-tatively attributes a skull from the Bear Paw Shale to Kritosau-rus). The distribution of these hadrosaurids at 73.5 Ma appearsto follow the same pattern developed by hadrosaurids earlier inthe Campanian, with the single hadrosaurine genus (but not nec-essarily species), Kritosaurus, spanning large geographic regionsand the three lambeosaurine genera (i.e., Parasaurolophus, theBaja lambeosaurine, and Velafrons) having smaller, more iso-lated distributions (Gates and Evans, 2005). Interestingly, Para-saurolophus, the tube-crested genus, stratigraphically co-occurswith members of the Corythosaurini (sensu Evans and Reisz, inpress) clade at two different intervals of the late Campanian. P.walkeri is found coincident—stratigraphically and geographi-cally—with Corythosaurus at the base of the Dinosaur Park For-mation, Alberta, Canada, around 76 Ma. Similarly, at 73.5 Ma, P.tubicen is found in sediments in New Mexico, USA, stratigraphi-cally equivalent to Velafrons in Coahuila, Mexico.
TABLE 2. Character coding of Velafrons coahuilensis based on thecharacter matrix presented in Evans and Reisz (in press).
111?? 11?01 ???11 ???11 11111 11111 01?11 00?1? ???1? ?1111 ?11?1111?1 ????1 ????? ????? ????? ????? ????? ????
Characters 11, 12, 13, 16, 18, 33, 37, 38, and 41 were coded as questionmarks because characters rely on adult conditions not present in CPC-59,or uncertain morphology of the crest.
GATES ET AL.—NEW LAMBEOSAURINE FROM MEXICO 927

CONCLUSIONS
Velafrons is the first genus of North American lambeosaurineto be named in over 70 years, and is the first dinosaur to beidentified to the species level from the Cerro del Pueblo Forma-tion, of Coahuila, Mexico. This taxon is characterized by a fan-shaped Corythosaurus-type crest, with unique features of thejugal, postorbital, quadrate, and ceratobranchial. The crest mor-phology is more similar to Corythosaurus and Hypacrosaurusthan to Lambeosaurus because it possesses a long anterior pro-cess of the nasal that extensively overlaps the lateral surface of
the premaxilla dorsal process. Phylogenetic analysis places Vela-frons within a polytomy with other fan-crested lambeosaurinesincluding Corythosaurus, Hypacrosaurus, Lambeosaurus, andOlorotitan.
The type specimen of Velafrons is a juvenile individual, butcomparison of skull morphology and growth of other similarly-sized taxa verifies that the apomorphies seen in Velafrons aredistinct and likely not ontogenetically variable. The size propor-tions of most skull elements correspond with those seen in otherNorth American juvenile lambeosaurines. The most obvious dif-ference is the relatively large size of the nasal and relativelyunderdeveloped prefrontal. Velafrons appears to have exhibitedmore rapid growth of the nasal than in other, similarly-sized taxa.Yet at the same time, it retains the morphology of the prefrontalseen in the youngest individuals of other lambeosaurines, that ofa nasal “clamp” instead of a “bowl”.
Biogeographically, Velafrons is significant because there areonly three other hadrosaurid taxa currently known from the lat-est Campanian (approximately 73.5 Ma): the hadrosaurine Kri-tosaurus, the lambeosaurine Parasaurolophus tubicen, and anunidentifiable lambeosaurine from Baja California. The dino-saur fauna of the Cerro del Pueblo Formation, has only recentlybeen targeted for detailed study. Results to date suggest that ithas potential to become one of the most interesting and impor-tant Late Cretaceous faunas from North America. It offers aunique opportunity to increase our understanding of dinosaurdiversity from an interval when terrestrial sediments are other-wise limited. Moreover, it provides opportunities to explore pa-leoecological patterns and processes in a terrestrial ecosystem atthe southernmost extreme of the North American continent, anarea that appears to have been strongly influenced by resurgentcoastal flooding.
ACKNOWLEDGMENTS
We sincerely thank Mike Getty and the 2002 field crew fortheir assistance in excavating the holotype specimen; RosarioGomez for her invaluable assistance with logistics; “Pato” (JoséLópez Espinosa), “Nacho” (José Ignacio Vallejo González), Ru-ben A. Rodriguez-de la Rosa, and Don DeBlieux for preparationand field work; Jerry Golden for his outstanding efforts prepar-ing the skull. Thanks also to David Evans for many excellentconversations about lambeosaurine ontogeny and for access tounpublished data, and to Jim Gardner, Kevin Seymour, CarlMehling, and Jack Horner for access to collections and speci-mens. Funding was provided by National Geographic Societyand the University of Utah.
LITERATURE CITED
Aguillon-Martinez, M. C., B. Espinosa, and J. I. Kirkland. 1998. Rostrode pez sierra, Ischyrhyza mira (Familia Schlerorhinchidae) de laFormacion Lutita Parras (Campaniano Tardio, Cretacico Superior)Coahuila, Mexico. Face of a saw fish, Ischyrhyza mira (Schlerorhin-chidae) from the Lutita Parras Formation (upper Campanian, Cre-taceous); Coahuila, Mexico. Memoria del . . . Congreso Nacional dePaleontologia. Resumenes 6:2–3.
Aguillon, M. C., I. J. Vallejo, R. Hernandez, and J. I. Kirkland. 1998.Dinosaur trackway from the Cerro del Pueblo Fm., Difunta Group(latest Campanian, Cretaceous), Coahuila, Mexico. Journal of Ver-tebrate Paleontology 18:23A.
Bartow, J. A. 1991. The Cenozoic evolution of the San Joaquin Valley,California. U.S. Geological Survey Professional Paper P 1501:40.
Bowers, W. E. 1990. Geologic map of Bryce Canyon National Park andvicinity, southwestern Utah. Miscellaneous Investigations Series,U.S. Geological Survey Report I-2108:1–17.
Brinkman, D., D. Eberth, S. D. Sampson, M. C. Aquillon Martinez, C. R.Delgado de Jesus, and R. Rodriguez de la Rosa. 2002. Paleontology
FIGURE 11. Biogeographic distribution of hadrosaurid taxa from 79.5-71.5 Ma in the Western Interior Basin of North America. The rightcolumn lists geographic regions where Campanian fossiliferous forma-tions have produced significant hadrosaurid specimens. The horizontalrows list formations present in each of the geographic regions. Hadro-saurid taxa found within each formation are listed below the formationname and their time spans are illustrated with either white or black bars.The white bars represent hadrosaurine taxa while the black bars depictlambeosaurine taxa. Grey regions show marine sediments. Modifiedfrom Gates and Evans (2005).
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 27, NO. 4, 2007928

and stratigraphy of the dinosaur-bearing Cerro del Pueblo Forma-tion, southern Coahuila, Mexico. Journal of Vertebrate Paleontol-ogy 22:38A–39A.
Brown, B. 1913. A new trachodont dinosaur, Hypacrosaurus, from theEdmonton Cretaceous of Alberta. Bulletin of the American Mu-seum of Natural History 32:395–406.
Cope, E. D. 1869. Synopsis of the extinct Batrachia, Reptilia, and Avesof North America. Transactions of the American Philosophical So-ciety 14:1–252.
Dodson, P. 1975. Taxonomic implications of relative growth in lambeo-saurine hadrosaurs. Systematic Zoology 24:37–54.
Eaton, J. G., R. L. Cifelli, J. H. Hutchison, J. I. Kirkland, and J. M.Parrish. 1999. Cretaceous vertebrate faunas from the KaiparowitsPlateau, south-central Utah; pp. 345–353 in D. D. Gillette (ed.),Vertebrate Paleontology in Utah. Utah Geological Survey, SaltLake City.
Eberth, D., S. D. Sampson, R. A. Rodriguez de La Rosa, M. C. AguillonMartinez, D. B. Brinkman, and J. Lopez Espinoza. 2003. Las Agu-ilas; an unusually rich Campanian-age vertebrate locale in southernCoahuila, Mexico. Journal of Vertebrate Paleontology 23:47A.
Eberth, D. A., C. R. Delgado de Jesus, J. F. Lerbekmo, D. B. Brinkman,R. A. Rodriguez de la Rosa, and S. D. Sampson. 2004. Cerro delPueblo Fm (Difunta Group, Upper Cretaceous) Parras Basin, south-ern Coahuila, Mexico; reference sections, age, and correlation. Re-vista Mexicana de Ciencias Geologicas 21:335–352.
Erickson, G. M., K. C. Curry, and S. A. Yerby. 2001. Dinosaurian growthpatterns and rapid avian growth rates. Nature 412:429–433.
Evans, D. C., C. Forster, and R. R. Reisz. 2005. The type specimen ofTetragonosaurus erectofrons (Ornithischia: Hadrosauridae) and theidentification of juvenile lambeosaurines; pp. 349–366 in P. J. Currieand E. B. Koppelhus (eds.), Dinosaur Provincial Park: A spectacularecosystem. Indiana University Press, Indianapolis.
Evans D. C., and R. Reisz. 2007. Anatomy and relationships of Lam-beosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithis-chia) from the Dinosaur Park Formation, Alberta. Journal of Ver-tebrate Paleontology 27:373–393.
Garcia, R., and V. H. Reynoso. 2002. The southernmost record of theturtle Bothremys (Testudine: Pleurodira) in the Cerro del PuebloFormation, near Saltillo, Coahuila, Mexico. Journal of VertebratePaleontology 22:56A.
Gates, T. A., and D. C. Evans. 2005. Biogeography of Campanian had-rosaurid dinosaurs from western North America; pp. 33–39 in D. R.Braman, F. Therrien, E. B. Koppelhus, and W. Taylor (eds.), Dino-saur Park Symposium short papers, abstracts, and programs. RoyalTyrrell Museum of Paleontology, Drumheller, Alberta.
Godefroit, P., Y. L. Bolotsky, and V. R. Alifronov. 2003. A remarkablehollow-crested hadrosaur from Russia: an Asian origin for lambeo-saurines. Systematic Paleontology 2:143–151.
Godefroit, P., Y. L. Bolotsky, and J. V. Itterbeeck. 2004. The lambeo-saurine dinosaur Amurosaurus riabinini, from the Maastrichtian ofFar Eastern Russia. Acta Palaeontologica Polonica 49:585–618.
Hernandez, R. 1992. New dinosaur finds in the Cerro del Pueblo For-mation (Upper Cretaceous, Campanian) from Coahuila State,Mexico. Journal of Vertebrate Paleontology 12:32A.
Hernandez, R., and J. I. Kirkland. 1993. The rediscovery of a rich up-permost Campanian dinosaur locality in the Cerro del Pueblo Fm.,Coahuila, Mexico. Journal of Vertebrate Paleontology 13:41A.
Hernandez, R., M. C. Aguillon, C. R. Delgado, and N. R. Gomez. 1995.The Mexican Dinosaur National Monument. Journal of VertebratePaleontology 15:34A.
Hernandez Rivera, R. 1996. Dinosaurios. Secretaria de Educacion Pub-lica de Coahuila, Mexico, Saltillo, 68 pp.
Horner, J. R. 1992. Cranial morphology of Prosaurolophus (Ornithis-chia: Hadrosauridae) with descriptions of two new hadrosaurid spe-cies and an evaluation of hadrosaurid phylogenetic relationships.Museum of the Rockies Occasional Paper 2:1–119.
Horner, J. R., and P. J. Currie. 1994. Embyonic and neonatal morphologyand ontogeny of a new species of Hypacrosaurus (Ornithischia,Lambeosauridae) from Montana and Alberta; pp. 312–336 in K.Carpenter, K. F. Hirsch, and J. R. Horner (eds.), Dinosaur Eggs andBabies. Cambridge University Press, Cambridge.
Horner, J. R., D. B. Weishampel, and C. Forster. 2004. Hadrosauridae;pp. 438–463 in D. B. Weishampel, P. Dodson, and H. Osmólska(eds.), The Dinosauria. University of California Press, Berkeley.
Kirkland, J. I., and M. C. Aguillon-Martinez. 2002. Schizorhiza; a unique
sawfish paradigm from the Difunta Group, Coahuila, Mexico. Re-vista Mexicana de Ciencias Geologicas 19:16–24.
Kirkland, J. I., R. Hernandez Rivera, M. C. Aguillon Martinez, C. R.Delgado de Jesus, R. Gomez-Numez, and I. Vallejo. 2001. The LateCretaceous Difunta Group of the Parras Basin, Coahuila, Mexicoand its vertebrate fauna. Universidad Autonoma del Estado deHidalgo, Avances en Invesigacion 3:133–172.
Kirkland, J. I., R. Hernandez, T. A. Gates, G. S. Paul, S. Nesbitt, S.-B.C.I., and J. P. Garcia-de la Garza. 2006. A large hadrosaur from theSabinas Basin, Northern Mexico; pp. 299–315 in S. G. Lucas andR. M. Sullivan (eds.), Late Cretaceous vertebrates from the WesternInterior. New Mexico Museum of Natural History and ScienceBulletin, Albuquerque.
Maddison, W. P., and D. R. Maddison. 2000. MacClade 4.0: Analysis ofphylogeny and character evolution. Sinauer Associates, Sunderland,Massachusetts.
Marsh, O. C. 1882. Classification of the Dinosauria. American Journal ofScience (Third Series) 23:81–96.
Maryanska, T., and H. Osmólska. 1979. Aspects of hadrosaurian cranialanatomy. Lethaia 12:265–273.
McBride, E. F., A. E. Weidie, J. A. Wolleben, and R. C. Laudon. 1974.Stratigraphy and Structure of the Parras and La Popa basins, North-eastern Mexico. Geological Society of America Bulletin 85:1603–1622.
Morris, W. J. 1970. Hadrosaurian dinosaur bills-morphology and func-tion. Contributions in Science of the Los Angelas County Museumof Natural History 193:1–14.
Morris, W. J. 1981. A new species of hadrosaurian dinosaur from theUpper Cretaceous of Baja California-?Lambeosaurus laticaudus.Journal of Paleontology 55:453–462.
Murray, G. E., A. E. Weidie, Jr., D. R. Boyd, R. H. Forde, and P. D.Lewis, Jr. 1962. Formational divisions of Difunta Group, Parras Ba-sin, Coahuila and Nuevo Leon, Mexico. Bulletin of the AmericanAssociation of Petroleum Geologists 46:374–383.
Ostrom, J. H. 1961. Cranial morphology of the hadrosuarian dinosaurs ofNorth America. Bulletin of the American Museum of Natural His-tory 122:33–186.
Owen, R. 1842. Report on British fossil reptiles. Report of the BritishAssociation of Advanced Sciences 9:60–204.
Page, B. M., and D. C. Engebretson. 1984. Correlation between thegeologic record and computed plate motions for central California.Tectonics 3:133–155.
Parks, W. A. 1923. Corythosaurus intermedius, a new species of trach-odont dinosaur. University of Toronto Studies, Geological Series15:5–57.
Perrilliat, M. d. C., B. Espinosa, and F. J. Vega. 2003. High diversity offreshwater gastropods from the Maastrichtian Cerro del Pueblo For-mation, Coahuila; environmental and stratigraphic implications. Ab-stracts with Programs—Geological Society of America 35:31.
Rodriguez de la Rosa, R., and S. R. S. Cevallos Ferriz. 1998. Vertebratesof the El Pelillal locality (Campanian, Cerro del Pueblo Formation),southeastern Coahuila, Mexico. Journal of Vertebrate Paleontology18:751–764.
Rodriguez de la Rosa, R. A. 2003. Pterosaur tracks from the latest Cam-panian Cerro del Pueblo Formation of southeastern Coahuila,Mexico. Geological Society Special Publications 217:275–282.
Rodriguez de La Rosa, R. A., D. A. Eberth, D. B. Brinkman, S. D.Sampson, and J. Lopez Espinoza. 2003. Dinosaur tracks from thelate Campanian Las Aguilas locality, southeastern Coahuila,Mexico. Journal of Vertebrate Paleontology 23:90A.
Seeley, H. G. 1888. On Thecospondylus daviesi (Seeley), with some re-marks on the classification of the Dinosauria. Quarterly Journal ofthe Geological Society of London 44:79–86.
Sternberg, C. M. 1935. Hooded hadrosaurs of the Belly River Series ofthe Upper Cretaceous: A comparison, with descriptions of new spe-cies. Bulletin of the Geological Survey of Canada 77:1–37.
Sullivan, R. M., and T. E. Williamson. 1999. A new skull of Parasauro-lophus (Dinosauria: Hadrosauridae) from the Kirtland Formation ofNew Mexico and a revision of the genus. New Mexico Museum ofNatural History and Science Bulletin 15:1–52.
Swofford, D. L. 2002. PAUP: Phylogenetic Analysis Using Parsimony(and other methods) version 4.0 b10. Sinauer Associates, Sunder-land, Massachusetts.
Tsujita, C. J., and G. E. G. Westermann. 1998. Ammonoid habitats andhabits in the Western Interior Seaway; a case study from the Upper
GATES ET AL.—NEW LAMBEOSAURINE FROM MEXICO 929

Cretaceous Bearpaw Formation of southern Alberta, Canada. Pal-aeogeography, Palaeoclimatology, Palaeoecology 144:135–160.
Vega, F. J., and R. M. Feldmann. 1991. Fossil crabs (Crustacea, Deca-poda) from the Maastrichtian Difunta Group, northeastern Mexico.Annals of Carnegie Museum 60:163–177.
Weishampel, D. B. 1981. The nasal cavity of lambeosaurine hadrosaurs(Reptilia: Ornithischia): Comparative anatomy and homologies.Journal of Paleontology 55:1046–1057.
Weishampel, D. B., D. B. Norman, and D. Grigorescu. 1993. Telmato-
saurus transsylvanicus from the Late Cretaceous of Romania: themost basal hadrosaurid dinosaur. Palaeontology 36:361–385.
Wiman, C. 1931. Parasaurolophus tubicen n. sp. aus der Kreide in NewMexico. Nova Acta Regieae Societatis Scientiarum Upsaliensis, Ser.IV 7:1–11.
Wolleben, J. A. 1977. Paleontology of the Difunta Group (Upper Cre-taceous-Tertiary) in northern Mexico. Journal of Paleontology 51:373–398.
Submitted August 17, 2006; accepted May 7, 2007.
JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 27, NO. 4, 2007930





























