Embed Size (px)
Citation preview

MICROSCOPY RESEARCH AND TECHNIQUE
Occurrence of Bacillus amyloliquefaciens as a Systemic Endophyte of Vanilla
Orchids
JAMES F. WHITE, JR.1*, MÓNICA S. TORRES1, RAYMOND F. SULLIVAN2,
RABIH E. JABBOUR3, QIANG CHEN1, MARIUSZ TADYCH1, IVELISSE
IRIZARRY1, MARSHALL S. BERGEN1, DAPHNA HAVKIN-FRENKEL1, FAITH C.
BELANGER1
1Department of Plant Biology and Pathology, Rutgers University, New Brunswick, New
Jersey, USA, Email: [email protected]
2U.S. Army CBRNE Analytical & Remediation Activity, Aberdeen Proving Ground,
Maryland, USA
3U.S. Army Edgewood Chemical Biological Center, Aberdeen Proving Ground,
Maryland, USA
KEY WORDS lipopeptides, plant disease protection, defensive mutualism, endospores
ABSTRACT We report the occurrence of Bacillus amyloliquefaciens in vanilla orchids
(Vanilla phaeantha) and cultivated hybrid vanilla (V. planifolia x V. pompona) as a
systemic bacterial endophyte. We determined with light microscopy and isolations into
culture that tissues of V. phaeantha and the cultivated hybrid were infected by a bacterial
endophyte and that shoot meristems and stomatal areas of stems and leaves were densely
colonized. We identified the endophyte as B. amyloliquefaciens using DNA sequence
data. Since additional endophyte-free plants and seed of this orchid were not available,
additional studies were performed on surrogate hosts Amaranthus caudatus, Ipomoea
tricolor and I. purpurea. Plants of A. caudatus inoculated with B. amyloliquefaciens
demonstrated intracellular colonization of guard cells and other epidermal cells,
confirming the pattern observed in the orchids. Isolations and histological studies suggest
that the bacterium may penetrate deeply into developing plant tissues in shoot meristems,

forming endospores in maturing tissues. We demonstrated that B. amyloliquefaciens
produced fungal inhibitors in culture. Further, in controlled experiments using morning
glory seedlings, we demonstrated that the bacterium promoted seedling growth and
reduced seedling leaf necrosis due to pathogens. We detected the gene for
phosphopantetheinyl transferase (sfp), an enzyme in the pathway for production of
antifungal lipopeptides, and purified the lipopeptide ‘surfactin’ from cultures of the
bacterium. Based on our data, we hypothesize that B. amyloliquefaciens is a robust
endophyte and defensive mutualist of vanilla orchids. Whether the symbiosis between
this bacterium and its vanilla hosts can be managed to protect vanilla crops from diseases
is a question that should be evaluated in future research.
INTRODUCTION
There is growing evidence that plants in natural populations are inhabited by non-
pathogenic microbes (Stone et al., 2000; Compant et al., 2005; Johnston-Monje and
Raizada, 2011). Microbial components of the plant microbiome are both bacterial and
fungal and may exist on plant surfaces and interiors (James, 2000; Angus and Hirsch,
2013; Arnold and Lutzoni, 2007; Reinhold-Hurek and Hurek, 2011). Although we do not
thoroughly understand how most non-pathogenic microbes interact with plants or with
one another, we do have several examples. For instance, research on endophytic fungi
demonstrates that they may enhance host plant resistance to biotic and abiotic stresses,
thus protecting plants from herbivory, diseases or environmental stresses such as soil
heavy metals, drought, or extreme temperatures (Redman et al, 2002; Clarke et al., 2006;
Hamilton et al., 2012; White and Torres, 2012; Schardl et al., 2013).
Vanilla orchids are hemi-epiphytic vines that climb trees and often become fully
epiphytic through loss of soil roots (Bayman et al., 2011). Currently, vanilla bean
production worldwide is threatened by an epidemic of fungal root rot diseases that is
projected to curtail supply of vanilla in coming years (Bayman et al., 2011). Due to
economic considerations, pesticides to reduce losses are not generally used in production
of vanilla beans (Bayman et al., 2011). In an effort to identify defensive bacterial

endophytes that might eventually be used to control diseases and support growth in
cultivated vanilla plantations we initiated studies on vanilla endophytes.
MATERIALS AND METHODS
Plant Materials and Survey of Vines
A single plant of hybrid vanilla (V. planifolia x V. pompona) was obtained from
naturalized populations in Puerto Rico. A single plant of Vanilla phaeantha was obtained
from a commercial source. From these two original plants numerous individual clones
were produced through fragmentation of vines. This was done by cutting vines into
segments that were approximately 1m in length and new roots and shoots were generated
on vines. In addition, five plants of V. planifolia were obtained from a commercial source
and confirmed Bacillus-free via failure to isolate from leaves. To survey for Bacillus
endophytes in natural populations of V. phaeantha, populations of the orchid were
located in Fakahatchee Strand Preserve State Park in South Florida. Because plants of V.
phaeantha are protected, sampling for the bacterium was done non-destructively. To
sample for Bacillus in plants, heat sterilized cotton applicators were rubbed into fluid
visible in the shoot tips of V. phaeantha plants, then placed into sterile tubes for transport
to the laboratory 3 to 5 days later. Cotton applicators were pasteurized to eliminate non-
endospore formers by heating applicators to 50ºC for two hours. Pasturized cotton
applicators were rubbed onto the surface of 10% Tripticase Soy Agar (TSA; Difco)
plates, incubated at laboratory ambient temperature and examined after 1 week for the
characteristic white, dry colonies of B. amyloliquefaciens with abundant endospores
(Figs. 1 and 2).
Isolation of Bacteria from Plants
For initial isolations of B. amyloliquefaciens from plants (three plants each of V.
phaeantha and hybrid vanilla), shoot tips, leaves and roots down to five nodes from the
shoot tip and 5cm stem segments from each internode down to the fourth internode were

removed from plants. All tissues were surface disinfected in 4% sodium hypochlorite
solution for 30 min with constant agitation, followed by agitation in sterile water for 10
min. Plant tissue segments were then excised into pieces approximately 2-3 mm2 and
plated onto 10% TSA (three tissue pieces/plate; and 5 plates per tissue sample), then
incubated for seven days at laboratory ambient temperature.
To access penetration of bacteria into stem tissues a five mm diam cork borer was
heat sterilized and used to core the center from six cm-long stem segments (surface
disinfected as above) cut from the third internode from the stem tips of five plants of V.
phaeantha. Stem cores were then rinsed in sterile water and cut into pieces 2-3 mm diam;
then fifteen pieces were plated onto 10% TSA medium and incubated for seven days at
laboratory ambient temperature. After incubation plates were examined to determine the
percentage of tissue pieces showing B. amyloliquefaciens growth (Table 1).
Identification of Bacteria
For bacterial identification, ~900 base pairs of the 16S rDNA region were
obtained (Baker et al., 2003). Sequences were compared to sequences available in the
NCBI GenBank database to identify the closest matches. For amplification of the
phosphopantetheinyl transferase (sfp) gene a 594bp fragment was amplified by PCR
using two primers, sfp-f (5’-ATGAAGATTTACGGAATTTA-3’) and sfp-r (5’-
TTATAAAAGCTCTTCGTACG-3’) (Hsieh et al., 2004). The amplification was
conducted using a program of denaturing at 94oC for 1 min, annealing at 46oC for 30 s
and extension at 72oC for 1 min, for a total of 36 cycles. The PCR product was analyzed
by 1% agarose gel electrophoresis and then sent for sequencing. Both sequences were
submitted to GenBank (16S rDNA = KF765481; sfp gene = KF765482).
Histology of Infected Plants
To visualize bacteria in biofilms of vanilla shoots (Figs. 3-7) and epidermal cells
of vanilla orchid tissues were stained with the nucleic acid stain SYTO®9/Propidium
Iodide provided in the Live/Dead Bacterial Viability Kit (Life Technologies, Carlsbad,
CA) and observed using fluorescence microscopy on a Zeiss Axioskop® using the filter
system for DNA excitation (485⁄498 nm for Fig. 4). To visualize the process of tissue

entry in vanilla and Amaranthus tissues, confocal microscopy was employed using the
nucleic acid stain SYTO13® (Life Technologies, Carlsbad, CA). Tissues were examined
using a Zeiss LSM 710® confocal microscope using lasers 48 (for nucleic acids) and 633
nm (for chlorophylls) with a 100x objective (Figs. 5,6, 11-14). To make 3-D
reconstructions (Figs. 11, 12 and 14), images were created using the z-stack application
in the Zen® software package. Images were constructed using slices through the upper
epidermal cell layer of tissues (24µm @ 0.3µm section intervals; 84 sections).
To visualize bacteria on the surfaces of young rapidly growing vanilla leaves
bordering the shoot tip, thin free-hand sections of epidermal layers were made using a
razor. These were then stained using safranin-o (0.1% aqueous) at room temperature for 5
min at ambient laboratory temperature (Fig. 7). To stain endospores in older leaf tissues,
longitudinal sections were stained using the endospore specific stain malachite green
(0.5% aqueous) for 15 min at 60C. Sections were washed with several changes of water
for 10 minutes by moistening tissues with water then blotted dry until no more green
coloration emerged from tissues (Gerhardt et al., 1994). Tissues were counterstained with
safranin-o as above, then examined with light microscopy (Figs. 8-10; Gerhardt et al.,
1994). To visualize bacteria within tissues of Vanilla phaeantha, leaves and stem cores
were cut into 1 x 3mm pieces and fixed in FAA fixative for 24 hours, then dehydrated
and embedded in LR White® Acrylic resin. Sections 1 µm -thick were made using glass
knives and stained on a slide warmer for 30 seconds using safranin-o (0.1% aqueous),
rinsed, and counterstained for 30 seconds using toluidine blue (0.1% aqueous; Gerhardt et
al., 1994). Sections were visualized using light microscopy (Figs. 15-17).
Amaranthus caudatus Infection Experiment
To evaluate the colonization of plant tissues by B. amyloliquefaciens an
inoculation experiment was conducted using Amaranthus caudatus. Seeds of A. caudatus
were surface disinfected using 4% sodium hypochlorite for 10 mins with constant
agitation, then washed for 5 mins in sterile distilled water. Twenty-five seeds were then
placed on each of 10 Petri dishes containing moist filter paper. One ml of a suspension of
B. amyloliquefaciens (concentration OD 600 nm = 0.5) was then added to five of the
plates, and one ml of sterile water was added to the other five plates. Plates were sealed

in clear plastic zip-lock bags and incubated on a lab bench for 10 days, after which both
treatments were examined and seedling root lengths measured and seedlings weighed.
Evaluation of Endospore Dormancy and Isolation Frequency
To evaluate the extent to which endospore dormancy within tissues of vanilla
vines reduces isolation frequency from mature plant tissues, we conducted a bacterial
isolation experiment using media: 1.) alanine-glucose agar (1% L-alanine: 1% glucose:
1.2% Agar Noble); and 2.) 10% TSA (0.4% Tripticase Soy Agar, Difco: 1.2% Agar
Noble). In addition, to trigger endospore germination some of the tissues were heated for
two hours in an oven at 60ºC prior to isolation from plant tissues. Plant tissues were
surface disinfected as previously described and cut into 2 mm2 pieces. Five Petri plates of
each treatment were prepared with three tissue pieces on each plate. Plates were
incubated for seven days in laboratory ambient conditions prior to examination for
presence of B. amyloliquefaciens and results recorded in Table 2.
Antibiosis Experiments
To examine in vitro production of antifungal compounds B. amyloliquefaciens
was streaked onto 10% TSA plates in a line. Plugs 7 mm in diam of fungi Alternaria
alternata, Colletotrichum dematium, Fusarium oxysporum and Lasiodiplodia theobromae
were placed on either side of the bacterial line at a distance of 2 cm from the bacterial
streak. Fifteen replicates were made for each fungus and the plates were incubated at lab
ambient temperature for ten days, after which zones of inhibition between bacterial
streaks and fungal colonies were measured (Table 3).
Experiments Using Morning Glory Seedlings
In the first experiment seeds of Ipomoea tricolor (cultivar Blue star) were treated
by soaking for 24 hours in sterile water or in a suspension of B. amyloliquefaciens
(concentration OD 600 nm = 0.5). Seeds were then placed into Petri plates (ten seeds
/plate and five plates/treatment) containing sterile moist filter paper and incubated in
Ziplock® plastic bags in laboratory ambient temperature for ten days, after which plates
were examined and seedlings assessed for effects on growth (Fig. 18). In a second

experiment seeds of Ipomoea purpurea (cultivar Crimson rambler) and Ipomoea tricolor
(cultivar Blue star) were soaked in a suspension of B. amyloliquefaciens or sterile water
as above, then planted in a flat containing potting mix comprised of three parts peat moss
to two parts perlite. Twenty-five seeds were planted in rows with each treatment/variety
forming a single row. The flat was then placed under a mister to maintain high moisture
around seedlings during the germination process. After two weeks twenty seedlings of
each treatment were harvested and the cotyledon leaf area was estimated (leaf length x
width). The percentage of seedlings in each treatment showing necrotic lesions was also
determined. These data are summarized in Table 4.
Fungal Inhibitor Purification and Identification
Bacillus amyloliquifaciens lipopeptides were produced in a liter of 10% tryptic
soy broth inoculated with 5 ml of an overnight culture of the bacterium and incubated for
24 hrs at 30°C shaking at 150 rpm. The bacterium was pelleted and the pH of the
supernatant was reduced to 3 with concentrated acetic acid and further reduced to pH 2
with concentrated hydrochloric acid. The precipitate from the acidified supernatant was
collected by centrifugation and resuspended in 45 ml of butanol. Butanol insoluble
material was pelleted by centrifugation and further removed by a 0.22 µ syringe filter.
The sample was introduced into the Thermo Scientific LTQ XL Linear Ion Trap Mass
Spectrometer by autosampler injection of 10 ml of the sample into a 100 ml min-1 HPLC
flow of methanol:water:formic acid (50:49.9:0.1) buffer without chromatographic
separation. The APCI-MS was tuned in positive mode to the 1036.7 m/z analyte with a
vaporizer temperature of 450°C, a capillary temperature of 200°C and an 80/20 N2 sheath
gas/auxillary gas flow rate. Collision induced dissociation energy of 50% was used to
produce the most effective fragmentation.
Toothpick Inoculation Experiment
Wooden toothpicks were sterilized by autoclaving in a 1% sucrose solution.
Sterilized toothpicks were then placed on the surface of 10% TSA plates that were
inoculated with plugs of Alternaria alternata. After fungal mycelium colonized
toothpicks, they were removed and dipped in a suspension on B. amyloliquefaciens

(concentration 600 nm OD = 0.5) or sterile water. Twenty toothpicks for each treatment
were then used to wound leaves of greenhouse maintained plants of V. planifolia. After
ten days the diameters of necrotic zones around toothpicks were measured.
RESULTS
Preliminary Survey
A preliminary survey of plants of Vanilla planifolia (4 plants), V. pompona (2
plants), a hybrid vanilla [V. planifolia x V. pompona] (6 plants) and V. phaeantha (10
plants) demonstrated presence of a bacterial endophyte in aerial tissues of vines of the
hybrid vanilla and the North American native species V. phaeantha. A white irregular
bacterial colony that produced masses of ellipsoidal to cylindrical refractive endospores
was consistently isolated on 10% TSA from surface sterilized pieces of shoot meristems,
stems and leaves of all plants of both the hybrid vanilla and V. phaeantha (Figs. 1 and 2).
Natural populations of V. phaeantha in Fakahatchee Strand Preserve State Park near
Naples, Florida were also sampled and found to also contain B. amyloliquefaciens in 8 of
14 plants sampled.
Bacterial Identification
For bacterial identification, the 16S rDNA and the phosphopantetheinyl
transferase (sfp) genes were sequenced. The vanilla endophyte sequence for the 16S
rDNA sequence (NCBI Accession KF765481) was similar to B. amyloliquefaciens strain
FZB42 (NCBI Accession NR_075005.1), a strain commonly used in biocontrol products.
The sequence for the sfp gene (GenBank Accession KF765482) was identical to the sfp
gene in B. amyloliquefaciens strains E1PA (EMBL-EBI Accession KC711052.1).
Phylogenetic trees constructed using our sequences demonstrated placement in the
species B. amyloliquefaciens for both sequence regions (trees not shown).
Observations on Shoot Tips

Examination of shoot meristems showed presence of liquid exudate from all
plants shown to be infected by B. amyloliquefaciens (Fig. 3). Microscopic examination of
primordial leaf tissues in the shoot apex revealed presence of bacterial biofilms
containing rods and coccoid forms (Fig. 4). Bacterial biofilms were seen to form over
epidermal cell junctions in differentiating leaves of shoot tissues (Fig. 5). Bacteria were
observed to densely colonize cells associated with stomatal openings, especially guard
cells and adjacent epidermal cells (Figs. 6-10; also 11-14).
Histological Studies and Distribution of Bacillus in Plant Tissues
Using confocal microscopy bacteria were observed to accumulate at cell
junctions of primordial leaves (Figs. 5 and 6). Examination of young leaves bordering the
shoot tip showed that bacteria also accumulate around guard cells and subsidiary cells of
stomata (Figs. 5-7). An examination of older leaves shows an abundance of endospores
characteristic of B. amyloliquefaciens on the surface of and within guard cells and
subsidiary cells (Figs. 8 and 9). In addition, endospores were occasionally seen in clusters
within epidermal cells (Fig. 10). In some slide preparations endospores were seen to fill
stomatal cavities (Fig. 9). Sections through vascular bundles of leaves also show rods that
resembled those of B. amyloliquefaciens in xylem, phloem and parenchyma cells but
endospores were not evident (Figs. 15-17).
Isolations of B. amyloliquefaciens from cores from the center of stems confirm
that the bacterium penetrated deeply into stem tissues (Table 1). Mature vanilla plant
tissues that were heat pre-treated prior to isolation of bacteria showed a significant
increase in isolation frequency of B. amyloliquefaciens into alanine-glucose agar (Table
2).
Confirmation of Intracellular Penetration by B. amyloliquefaciens
In Amaranthus caudatus inoculation experiments B. amyloliquefaciens was
observed to colonize leaf tissues in a comparable way to that observed in vanilla.
Numerous cells of the bacterium were seen to enter guard cells and were often seen close
to chloroplasts (Figs. 11 and 12). In other epidermal cells bacteria were intercellular, but
remained in the periphery of the cell in cytoplasm (Fig. 14). Biofilms of bacteria formed

on the leaf surface over cell junctions (Fig. 13). Un-inoculated control plants did not
show biofilm formation on seedling leaves, and intracellular bacteria were not observed
in confocal microscopic observation of the plant tissues. Amaranthus seedlings colonized
by B. amyloliquefaciens had seedling leaves that were brighter red in color and smaller
than in the water treated controls. Bacterial colonized seedlings were on average smaller
(seedling wet wt = 5.19 ± 0.79 mg vs 7.82 ± 1.1 mg (mean ± standard deviation) for
water controls). Seedlings colonized by B. amyloliquefaciens also had notably shorter
roots than controls (10.9 ± 3.48 mm vs 25.45 ± 4.74 mm for water controls).
Fungal Inhibitors and Disease Control
Isolates of B. amyloliquefaciens from both V. phaeantha and the hybrid vanilla
were found to produce fungal inhibitors in culture (Table 3). We further amplified the
phosphopantetheinyl transferase (sfp) gene for iturin A production from the V. phaeantha
isolate. The supernatant from a liquid culture of B. amyloliquefaciens was acid
precipitated and analyzed. After Atmospheric Pressure Chemical Ionization Mass
Spectrometry (APCI-MS), a prominent 1036.7 (M+H)+ ion (Fig. 19) was further
investigated by MS/MS using resonance excitation collision induced dissociation (CID).
Although CID resulted in highly efficient precursor ion fragmentation, it was subject to a
low mass cutoff and did not allow trapping of fragment masses below 27.5% of the
precursor mass. Pulsed Q Collision Induced Dissociation (PQD) was attempted to
observe predicted low m/z fragments but very few ions of any m/z were observed and no
low m/z ions were observed (data not shown). Nevertheless, observed ions with m/z
>285 convincingly showed the 1036.7 m/z ion to be C15 surfactin, a powerful surfactant
lipopeptide compound produced by various Bacillus spp. (Fig. 20). In addition, common
morning glory seedlings derived from seeds treated with B. amyloliquefaciens showed
significantly larger seedling leaves (cotyledons) than those of water treated controls;
bacterial treated seedlings demonstrated a lower incidence of fungal lesions/necrosis than
seedlings without bacteria (Table 4). Microscopic examination of the cotyledons of
morning glory seedlings treated with the bacterium (see Fig. 18) revealed dense bacterial
biofilms on seedling leaves (Fig. 13), while water treated controls lacked any evidence of
biofilms on seedling leaves. In experiments using Bacillus-free Vanilla planifolia leaves

wounded using toothpicks bearing the fungal pathogen Alternaria alternata, wounds
created using toothpicks bearing only the fungus had a greater lesion diameters after a 4-
day incubation period than leaves wounded using toothpicks treated with the fungal
pathogen and B. amyloliquefaciens (fungus only lesion diam = 7.1538 ± 0.67791 mm
(mean ± standard deviation); fungus + bacterium lesion diam = 2.84615 ± 0.93264 mm;
N = 15; a T-test for paired samples analysis gave p < 0.01).
DISCUSSION
This study provides evidence that Bacillus amyloliquefaciens is both epiphytic
and endophytic in vanilla plants. While not exhaustively examined, the isolation of the
bacterium from tissues of the hybrid vanilla from Puerto Rico and plants of V. phaeantha
in South Florida suggest that B. amyloliquefaciens may be a widespread symbiont of
vanilla orchids. The fact that we did not isolate it from several plants of Vanilla planifolia
and V. pompona may be an indication that it is not present in all vanilla orchids.
Histological studies on V. phaeantha leaf tissues show that bacteria form biofilms
in shoot tips on leaf and stem primordia. These biofilms concentrate at epidermal cell
junctions and over stomata. This may be due to nutrient leakage from stomata and cell
junction areas. Based on observations made using vanilla plants and the surrogate host
Amaranthus caudatus, intracellular infection of plant tissues appears to occur beneath
these biofilms. In the process of infection rods appear to convert into spherical cells, as
observed in the infection of seedling leaves of Amaranthus caudatus (Figs. 11-14). These
spherical bacterial cells are likely cell wall deficient bacteria, often called L-forms.
Similar L-forms have been shown to occur in Bacillus subtilis and many other bacteria
when they enter into the cytoplasm of eukaryotic cells (Leaver et al., 2009). L-form
development is accompanied by reduced virulence and long-term persistence in the host,
thus bacterial L-forms of pathogenic bacteria have been considered to be symbiotic or
latent phases (Amijee et al., 1992; White et al., 2014). Based on the position of bacteria
in epidermal cells, the L-forms appear to be limited to the cytoplasm (Figs. 11 and 12).
Guard cells may be rich in nutrients and cytoplasmic contents due to presence of

chloroplasts there. The presence of bacteria around the margins of other epidermal cells
(Figs. 11 and 14) may be due to the large central vacuole in those cells. We did not
observe entry of B. amyloliquefaciens into the vacuole but rather it remained in the
cytoplasmic component of cells.
It seems likely that entry of the bacterium into cells of the plant is accomplished
through use of cell wall degrading enzymes. Confocal microscopy of the inner top walls
of two guard cells (Fig. 12) shows the bacteria in guard cells, and surrounding the
bacteria are darker halos that may represent softened or altered plant cell walls. Other
symbiotic bacteria are also thought to enter plant cells using cell wall degrading enzymes
(see Kovtunovych et al., 1999; Compant et al., 2005).
Entry of B. amyloliquefaciens into guard cells and subsidiary cells could affect the
functionality of stomata. Even if guard cells continued to function after entry of bacteria,
the eventual proliferation of bacteria in the stomatal chambers could prevent their closure
or reduce gas exchange with the exterior (Fig. 9). It is not clear to what extent
functionality of stomata is affected by the bacterial endophyte. However, if stomatal
functioning is reduced, B. amyloliquefaciens could be considered a weak pathogen of
vanilla orchids. Along these lines, in inoculation experiments involving Amaranthus
caudatus we recorded reductions in size and weight of seedlings as a result of infection
by B. amyloliquefaciens. This reduced growth could be a reflection of a parasitic
interaction between the bacterium and Amaranthus and could be a further indication of
the cost of the interaction to vanilla plants (Cheplick, 2007).
Two lines of evidence suggest that B. amyloliquefaciens penetrates vanilla plant
tissues but very rapidly goes dormant in most tissues. We observed abundant endospore
formation in leaf epidermal cells (Figs. 8-10). In addition, heat pre-treatments of mature
plant tissues prior to isolation attempts on alanine-glucose agar increased the rate of
isolation from tissues. Because bacterial endospores are heat activated (Keynan et al.,
1964), enhanced isolation of the bacterium from tissues after heat treatments is consistent
with a scenario where bacteria are present as endospores in plant tissues.

It is unclear how deeply B. amyloliquefaciens penetrates into vanilla orchid
tissues. We have confidence that the bacterium penetrates into epidermal layers because
we observed rods and endospores characteristic of the bacterium in these tissues, and our
infection experiments using Amaranthus caudatus confirmed at least superficial infection
into epidermal cells. However, we isolated the bacterium from deep within stem cores
where epidermal layers had been removed, suggesting that the tissues of the vanilla
orchid may be permeated in all parts by the bacterium. In sections of stem vascular
bundles we were able to visualize aggregations of bacterial rods in xylem, phloem and
parenchyma cells (Figs. 15-17). However, because we did not observe endospores
forming within tissues of vascular tissues, and our stains were non-specific, we cannot be
sure that they belong to B. amyloliquefaciens. Regardless of this uncertainty, the
consistent isolation of this bacterium from internal tissues of the stem shows that it
penetrates deeply into plant tissues. In addition, since endospores generally form as a
result of nutrient restriction (Keynan et al., 1964), it seems reasonable that the availability
of a continuous supply of sugars and other nutrients within vascular tissues of vanilla
vines reduces tendency of the bacterium to form endospores and could account for lack of
any evidence of endospores in vascular tissues (Figs. 15-17).
Vertical dissemination of the bacterium is facilitated by growth of the bacterium
within shoot tips of vanilla orchids that enables it to colonize all primary leaves and
stems as they are formed. Any new stems arising from axillary buds and aerial roots also
become colonized by the bacterium. Thus the bacterium is able to move from older stems
to newly initiated meristems. It is possible that this movement is due to bacterial
movement in vascular tissues that connect all parts of the plant. Alternatively, movement
of bacteria from older stems to newly initiated meristems may be due to surface
movement of bacteria on plants, since bacteria may move using swarming motility even
when surface plant moisture is lacking. Although, how B. amyloliquefaciens moves
within plants is unknown, it is certain that bacterial colonization of new stems enables the
bacterium to transmit to new clonally produced plants. In vanilla orchids clonal
propagation occurs frequently with most new plants arising from fragmentation of
parental vines. In agricultural production vines are broken into segments to initiate new
vines (Bayman et al., 2011). In our greenhouse culture of the hybrid vanilla and V.

phaeantha we produced dozens of new plants using fragmented vines and in every case
plants retained B. amyloliquefaciens. We were not able to evaluate whether the bacterium
would vector through seeds since our plants did not produce fruits.
Liquid accumulation in shoot tips of infected vines suggests another mechanism
for horizontal transmission of the bacterium to uninfected plants. The bacterium could
move horizontally between plants via insects that feed on the shoot meristem liquid. We
were able to repeatedly isolate B. amyloliquefaciens from shoot apex liquid. Insects, such
as flies, may vector bacteria to other plants when they feed or drink from the liquid in the
shoot tip. It is equally possible that bacteria that flow out of stomata may be dispersed by
a rain splash mechanism to adjacent plants. The high concentrations of bacteria that we
observed associated with the surfaces of vanilla plants makes this water splash
mechanism highly probable. However, additional work is needed to determine whether
either of these proposed dissemination mechanisms are important in natural populations.
Beneficial effects of infection by B. amyloliquefaciens on infected vanilla plants
is difficult to prove because all of our hybrid and V. phaeantha plants contained the
bacterium and controlled experiments using vanilla orchids were not possible. However,
we were able to demonstrate that B. amyloliquefaciens secretes surfactin, a lipopeptide
composed of a 7 amino acid cyclic ring and an alkyl tail of varied length and structure
(Raaijmakers et al., 2010). Surfactins have demonstrated fungitoxicity towards
Magnaporthe grisea, Sclerotinia sclerotiorum, and Fusarium verticillioides (Tendulkar et
al., 2007; Ongena and Jacques, 2007; Snook et al., 2009; Alvarez et al., 2012). Induction
of host plant systemic resistance by surfactins has also been demonstrated through
stimulation of the lipoxygenase pathway (Ongena and Jacques, 2007). Surfactin
lipopeptides have been shown to reduce bacterial colonization of plants by inhibition of
bacterial motility and biofilm formation (Chowdhery et al., 2013). Because of the
capacity to suppress root and foliar diseases in plants, several strains of B.
amyloliquefaciens have been employed in biocontrol formulations of commercial
products (Ongena and Jacques, 2007). In our experiments we found that the vanilla
Bacillus demonstrated inhibitory antagonism toward several fungal species when cultured
together. In another experiment using morning glory seedlings with and without the

bacterium, we found that the bacterium promoted seedling growth and reduced seedling
leaf lesions, probably due to damping off fungi (Fig. 18; Table 4). While not definitive,
we have enough evidence to hypothesize that B. amyloliquefaciens is a naturally
occurring defensive mutualist of vanilla orchids (Clay, 1988; Elliot et al., 2000).
Validation of the defensive nature of the vanilla Bacillus will require controlled
experiments with Bacillus-infected and -free vanilla plants. Whether, this bacterium has
potential to control diseases that are currently affecting vanilla orchids (Vanilla planifolia
and V. pompona) in commercial operations will require additional evaluation.
CONCLUSIONS
We present evidence that B. amyloliquefaciens is a common systemic endophyte
of some species of vanilla orchids. This is the first plant where it has been shown that
growth-promoting bacteria colonize plants through shoot tissues rather than root tissues.
We further show that the bacterium colonizes plant cells intracellularly. This has not been
shown in many other plants and where it has been shown, cells invaded have been root
cells (Paungfoo-Lonhienne et al., 2010; White et al., 2014). We demonstrated that B.
amyloliquefaciens contains genes for and produces antifungal lipopeptides; and that
application of the bacterium to morning glory seeds promoted seedling growth and
reduced disease. On the basis of the studies reported here we hypothesize that B.
amyloliquefaciens is a widespread defensive mutualist of some species of vanilla orchids.

Figs. 1‐2. Bacillus amyloliquefaciens. 1. Vanilla stem segments showing characteristic colonies of the bacterium emerging from plant tissues on 10% TSA medium. 2. Densely‐stained endospores (arrows) and longer, lightly‐stained, vegetative cells (stained with malachite green endospore stain).

Figs. 3-6. Vanilla phaeantha bearing Bacillus amyloliquefaciens. 3. Shoot tip showing
liquid exudate (arrow) at shoot opening of infected plants. 4. Biofilm on primordial leaf
in shoot showing rods (arrow) stained with SYTO9®. 5. Confocal image of surface of
leaf in shoot tip showing bacteria (arrows) in biofilms at cell junctions stained with
SYTO13®. 6. Confocal image showing bacteria (arrows) on and around guard cells
stained with SYTO13®.

Figs. 7-10. Vanilla phaeantha. 7. Bacterial accumulation (arrows) around stoma on primordial leaf bordering shoot tip, stained with safranin (0.1%, aqueous). 8. Bacterial endospores (arrows) on guard cells of an older leaf, stained with endospore stain malachite green (0.1%, aqueous) followed by safranin. 9. Bacterial endospores (arrows) in a substomatal chamber of an older leaf, stained with endospore stain malachite green (0.1%, aqueous) followed by safranin. 10. Clusters of endospores (arrows) within an epidermal cell of an older leaf, stained with malachite green.

Figs. 11, 12, 14. Confocal microscopy of Bacillus amyloliquefaciens in a seedling leaf of Amaranthus caudatus stained with SYTO13. 11. Image showing spherical bacterial L-forms (orange; arrows) within epidermal cells. White arrows indicate bacterial L-forms in guard cells (chloroplasts are blue). Black arrows indicate spherical bacterial L-forms in adjacent epidermal cells. 12. Maximum intensity projection showing spherical bacterial L-forms (arrows) within guard cells near chloroplasts (red). 13. Surface of morning glory seedling leaf showing bacterial biofilm (arrows) after seeds were treated with B. amyloliquefaciens. 14. Maximum intensity projection using a subset of z-stack showing bacterial accumulations at periphery of epidermal cells (arrows).
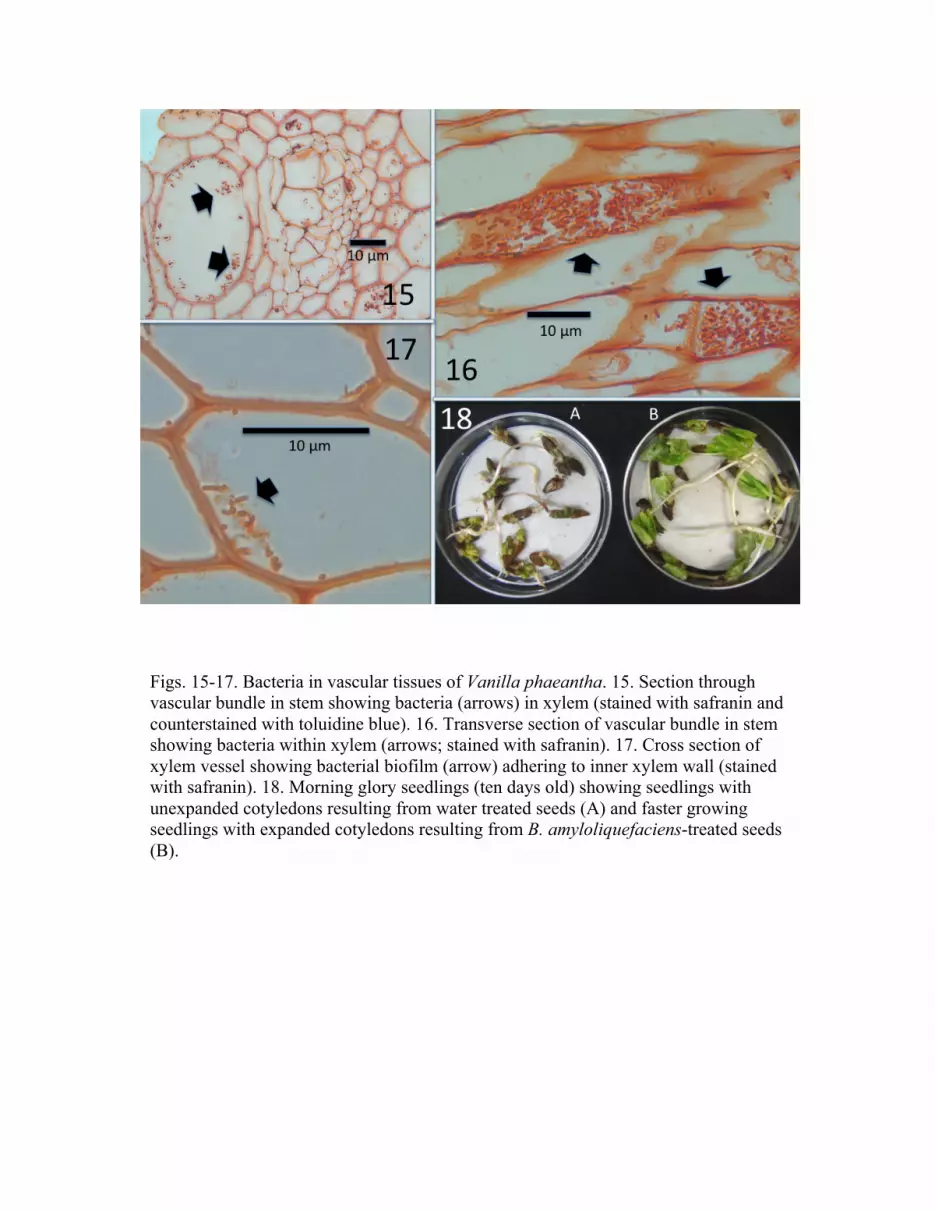
Figs. 15-17. Bacteria in vascular tissues of Vanilla phaeantha. 15. Section through vascular bundle in stem showing bacteria (arrows) in xylem (stained with safranin and counterstained with toluidine blue). 16. Transverse section of vascular bundle in stem showing bacteria within xylem (arrows; stained with safranin). 17. Cross section of xylem vessel showing bacterial biofilm (arrow) adhering to inner xylem wall (stained with safranin). 18. Morning glory seedlings (ten days old) showing seedlings with unexpanded cotyledons resulting from water treated seeds (A) and faster growing seedlings with expanded cotyledons resulting from B. amyloliquefaciens-treated seeds (B).

Fig. 19. APCI mass spectrum of the precipitated supernatant from an overnight culture of
the Bacillus amyloliquifaciens endophyte. Annotated peaks correspond to surfactins with
various fatty acid tail (C) lengths and charged ions.

Fig. 20. APCI-MS/MS spectrum (averaged from 31 spectra) of the 1036.7 m/z precursor
ion using collision induced dissociation (CID) (50% collision energy). The upper right
box lists predicted fragment ions with observed ions underlined and labeled in the
averaged spectrum. The structure of Surfactin A is shown (upper left).

TABLE 1. Isolation of B. amyloliquefaciens from epidermal tissues and stem cores of V.
phaeantha onto TSA medium1
Tissue Vine number
1 2 3 4 5
Epidermis 91.67% 100% 100% 100% 0%
Core 100% 88% 100% 100% 83.3%
1Percentage of pieces showing emergence of bacterium (N=15)
TABLE 2. Effect of medium and heat treatment on isolation frequency
Tissue Pre-treatment Alanine-glucose
agar
10% Trypticase Soy
Agar
Shoot tip No heat 100% 100%
Leaf 1 No heat 0% 100%
Leaf 2 No heat 20% 100%
Leaf 3 No heat 0% 100%
Leaf 4 2 hr @ 60C 100% 100%
Leaf 5 2 hr @ 60C 73.33% 100%
Leaf 6 2 hr @ 60C 0% 0%
Stem No heat 40% 86.67%
Stem 2 hr @ 60C 78.57% 93.33%

TABLE 3. Fungal pathogen inhibition by B. amyloliquefaciens in culture1
Fungal Pathogens
Plant Source Fusarium
oxysporum
Lasiodiplodia
theobromae
Colletotrichum
dematium
Alternaria
alternata
Hybrid vanilla 4.8 ± 1.3 5.8 ± 0.92 11.4 ± 1.26 11.4 ± 0.97
V. phaeantha 3.8 ± 0.83 7.7 ± 1.34 10.9 ± 2.85 9.2 ± 1.4
1 Data is mean ± standard deviation of inhibition zone diameters in mm (N = 20)
TABLE 4. Bacillus amyloliquefaciens effects on morning glory seedling cotyledon size
and necrosis
Cultivar Water control B. amyloliquefaciens
Leaf area
(mm2)
Percent
necrotic
Leaf area (mm2) Percent
necrotic
Crimson
rambler
251.9 ± 92.21 90% 483.3 ± 132.1 0%
Blue star 435.4 ± 240.87 100% 742.5 ± 448.4 0%
1Significant difference for both varieties when water controls are compared to bacterial
treatments (Student T test; P < 0.05; N=20).

REFERENCES
Alvarez F, Castro M, Principe A, Borioli G, Fischer S, Mori G, Jofré E. 2012. The plant-
associated Bacillus amyloliquefaciens strains MEP218 and ARP23 capable of
producing the cyclic lipopeptides iturin or surfactin and fengycin are effective in
biocontrol of sclerotinia stem rot disease. J Appl Microbiol 112: 159-174.
Amijee F, Allan EJ, Waterhouse RN, Glover LA, Paton AM. 1992. Non-pathogenic
association of L-form bacteria (Pseudomonas syringae pv. phaseolicola) with bean
plants (Phaseolus vulgaris L.) and its potential for biocontrol of halo blight disease.
Biocontrol Sci Technol 2: 203-214.
Angus AA, Hirsch AM. 2013. Biofilm formation in the rhizosphere: Multispecies
interactions and implications for plant growth. Pages 703-712, in: de Bruijn FJ (ed.)
Molecular Microbial Ecology of the Rhizosphere, Volume 2. John Wiley & Sons, Inc.
Arnold AE, Lutzoni F. 2007. Diversity and host range of foliar fungal endophytes: are
tropical leaves biodiversity hotspots? Ecology 88, 541-549.
Bacon CW, Hinton DM. 2011. In planta reduction of maize seedling stalk lesions by the
bacterial endophyte Bacillus mojavensis. Canadian Journal of Microbiology 57: 1-8.
Baker GC, Smith JJ, Cowan DA. 2003. Review and re-analysis of domain-specific 16S
primers. J. Microb. Methods 55: 541-555.
Bayman P, Mosquera-Espinosa AT, Porras-Alfaro A. 2011. Mycorrhizal relationships of
vanilla and prospects for biocontrol of root rots. Pages 266-280 in: Havkin-Frenkel D,
Belanger FC (eds.), Handbook of Vanilla Science and Technology, Blackwell
Publishing Ltd.
Cheplick GP. 2007. Costs of fungal endophyte infection in Lolium perenne genotypes
from Eurasia and North Africa under extreme resource limitation. Environmental and
Experimental Botany 60: 200–210.
Chowdhury SP, Dietel K, Rändler M, Schmid M, Junge H, Borriss R, Hartmann A,
Grosch R. 2013. Effect of Bacillus amyloliquefaciens FZB42 on lettuce growth and
health under pathogen pressure and its impact on the rhizosphere bacterial

community. PLoS ONE 8 (7), e68818.doi: 10.1371/journal.pone.0068818.
Clarke BB, White JF Jr, Hurley RH, Torres MS, Sun S, Huff DR. 2006. Endophyte-
mediated suppression of dollar spot disease in fine fescue. Plant Disease 90: 994-998.
Clay K. 1988. Fungal endophytes of grasses: a defensive mutualism between plants and
fungi. Ecology 69: 10-16.
Compant S, Reiter B, Sessitsch A, Nowak J, Clement C, Barka E. 2005. Endophytic
colonization of Vitis vinifera L. by plant growth-promoting bacterium Burkholderia
sp. strain PsJN App. Environ Microbiol 71: 1685-1693.
Elliot SL, Sabelis MW, Janssen A, van der Geest LPS, Beerling EAM, Fransen J. 2000.
Can plants use entomopathogens as bodyguards? Ecol. Letters 3: 228-235.
Hamilton CE, Gundel PE, Helander M, Saikkonen K. 2012. Endophytic mediation of
reactive oxygen species and antioxidant activity in plants: a review. Fungal Diversity
54: 1-10.
Gerhardt P, Murray RGE, Wood WA, Krieg NR. 1994. Methods for General and
Molecular Bacteriology. ASM Press, Washington, DC.
Hsieh F-C, Li M-C, Lin T-C, Kao S-S. 2004. Rapid detection and characterization of
surfactin-producing Bacillus subtilis and closely related species based on PCR.
Current Microbiology 49: 186-191; DOI: 10.1007/s00284-004-4314-7.
James EK. 2000. Nitrogen fixation in endophytic and associative symbiosis. Field Crops
Research 65: 197-209.
Johnston-Monje D, Raizada MN. 2011. Conservation and diversity of seed associated
endophytes in Zea across boundaries of evolution, ethnography and ecology. PLoS
ONE 6 (6), e20396. doi:10.1371/journal.pone.0020396.
Keynan A, Evanchik Z, Halvorson H, Hastings J. 1964. Activation of bacterial
endospores. Journal of Bacteriology 88: 313-318.
Kovtunovych, G., Lar, O., Kamalova, S., Kordyum, V., Kleiner, D. and Kozyrovska, N.
1999. Correlation between pectate lyase activity and ability of diazotrophic Klebsiella
oxytoca VN 13 to penetrate into plant tissues. Plant Soil 215: 1-6.

Leaver M, Dominguez-Cuevas P, Coxhead JM, Daniel RA, Errington J. 2009. Life
without a wall or division machine in Bacillus subtilis. Nature 457: 849-853.
Ongena M, Jacques P. 2007. Bacillus lipopeptides: versatile weapons for plant disease
biocontrol. Trends in Microbiology 16: 115-125.
Paungfoo-Lonhienne C, Rentsch D, Robatzrk S, Webb RI, Sagulenko E, Nasholm T,
Schmidt S, Lonhienne TGA. 2010. Turning the table: plants consume microbes as a
source of nutrients. PLoS ONE 5(7), e11915. doi: 10:1371/journal.pone.0011915.
Porras-Alfaro A, Bayman P. 2011. Hidden fungi, emergent properties: Endophytes and
microbiomes. Annu Rev Phytopathol 49: 291-315.
Raaijmakers JM, de Bruijn I, Nybroe O, Ongena M. 2010. Natural functions of
lipopeptides from Bacillus and Pseudomonas: more than surfactants and antibiotics.
FEMS Microbiol Rev 34: 1037–1062; DOI:10.1111/j.1574-6976.2010.00221.x.
Redman RS, Sheehan KB, Stout RG, Rodriguez RJ, Henson JM. 2002. Thermotolerance
generated by plant/fungal symbiosis. Science 298: 1581.
Reinhold-Hurek B, Hurek T. 2011. Living inside plants: bacterial endophytes. Current
Opinion in Plant Biology 14: 435-443.
Schardl CL, Florea S, Pan J, Nagabhyru P, Bec S, Calie PJ. 2013. The epichloae: alkaloid
diversity and roles in symbiosis with grasses. Current Opinion in Plant Biology 16:
480-488.
Snook ME, Mitchell T, Hinton DM, Bacon CW. 2009. Isolation and characterization of
Leu7-surfactin from endophytic bacterium Bacillus mojavensis RRC 101, a
biocontrol agent for Fusarium verticilioides. Agric Food Chem 57: 4287-4292.
Stone JK, Bacon CW, White JF Jr. 2000. An overview of endophytic microbes:
endophytism defined. In: Bacon CW, White JF Jr. (eds) Microbial Endophytes,
Marcel-Dekker, New York, USA, pp. 3-30.

Tendulkar SR, Saikumari YK, Patel V, Raghotama S, Munshi TK, Balaram P, Chattoo
BB. 2007. Isolation, purification and characterization of an antifungal molecule
produced by Bacillus licheniformis BC98 and its effect on phytopathogen
Magnaporthe grisea. J Appl Microbiol 103: 2331-2339.
White JF, Torres MS. 2010. Is endophyte-mediated defensive mutualism oxidative stress
protection? Physiologia Plantarum 138: 440-446.
White JF, Torres MS, Somu MP, Johnson H, Irizarry I, Chen Q, Zhang N, Walsh E,
Tadych M, Bergen M. 2014 In press. Hydrogen peroxide staining to visualize
intracellular bacterial infections of seedling root cells. Microscopy Research and
Technique (In press)
*Correspondence to: James F. White, Department of Plant Biology and Pathology,
Rutgers University, 59 Dudley Road, New Brunswick, NJ 08901, USA. E-mail:
[email protected]. Received ____________2014;
REVIEW EDITOR: Professor Alberto Diaspro.
Contract grant sponsor: USDA NIFA Multi-State Project 3147, the John E. and Christina
C. Craighead Foundation, and the New Jersey Agricultural Experiment Station. The
authors are grateful to Dr. Maurine Bonness and Mike Owen for assisting with sampling
plants of V. phaeantha in cypress swamps of the Fackahatchee Strand Preserve State Park
in South Florida.





























