Embed Size (px)
Citation preview

Pesticide Biochemistry and Physiology 65, 44–54 (1999)Article ID pest.1999.2426, available online at http://www.idealibrary.com on
The Toxicity and Physiological Effects of Bacillus thuringiensisToxins and Formulations on Thaumetopoea pityocampa, the
Pine Processionary Caterpillar
Carolina Rausell, Amparo Consuelo Martınez-Ramırez, Inmaculada Garcıa-Robles,and Marıa Dolores Real1
De in
atedthur
1 T96 39
0048CopyriAll rig
partamento de Genetica, Facultad de Ciencias Biologicas, Doctor Moliner 50, 46100 Burjassot, Valencia, Spa
Received January 19, 1999; accepted May 25, 1999
Toxicity of Bacillus thuringiensis Cry1B, Cry1C, and Cry1E toxins and B. thuringiensis-based bioinsectic-ides (Cordalene, Dipel, Foray 48B, and Foray 76B) was investigated in Thaumetopoea pityocampa (proces-sionary moth) larvae. Cry1B was toxic, while Cry1C and Cry1E were nontoxic. The toxicity of thebioinsecticides analyzed was of the same magnitude as that of the active purified toxins and determinedmainly by one of the toxins present in the bioformulate. We also demonstrated that only the active toxinswere able to produce cytotoxic effects on the midgut epithelial cells and specifically bound to T. pityocampa
o
brush border membrane. Analysis of the receptor mcompete for the Cry1A binding site. This finding prthuringiensis toxins against T. pityocampa. q1999 AINTRODUCTION
Thaumetopoea pityocampa, the pine proces-sionary caterpillar, is the most importantendemic pine pest in the Mediterranean area,not only because of its high defoliating power,but also due to the human health problemscaused by the urticating hairs of the larvae (1).Although there is an increasing demand for envi-ronmentally friendly alternative methods, effortsto control this lepidopteran insect involve mainlythe use of chemical insecticides, particularlyinsect growth inhibitors (2).
Insect viruses (cytoplasmic polyhedrosisvirus, nuclear polyhedrosis virus, and granulosisvirus), fungi (Beauveria bassiana), pheromones,parasites, and especially bacteria (mainly prepa-rations based on Bacillus thuringiensis Berlinervar. kurstaki) have been investigated for the bio-logical control of T. pityocampa (3). In somecases, these insect pathogens have been evalu-
as biological control agents but only B.ingiensis is already in operational use (3–5).
o whom correspondence should be addressed. Fax: 348 30 29. E-mail: [email protected].
4-3575/99 $30.00ght q 1999 by Academic Presshts of reproduction in any form reserved.
odel of the active toxins showed that Cry1B did notvides useful information for optimizing the use of B.cademic Press
B. thuringiensis is an infectious bacterium thatproduces insecticidal proteins that are proteolyti-cally activated by insect gut proteases and spe-cifically bind to receptors in the insect midgut.Once the toxins are bound, pores are formedthat alter membrane permeability and eventuallycause lysis of the epithelial cells, killing theinsect (6). Each B. thuringiensis strain exhibitsdifferent insecticidal activity spectra due primar-ily to the diversity of toxins that it can produce.Several well-characterized B. thuringiensis iso-lates have been industrially formulated and arecurrently applied against Lepidoptera (the mostdestructive group of plant pests), Diptera (vec-tors of human disease), and Coleoptera (7).
Despite the important environmental damageand economic losses caused by T. pityocampa,basic laboratory research on this pest is unfortu-nately limited. Most of the research on proces-sionary moth has been done in the field and,regarding B. thuringiensis, it has focused mainlyon determining the optimal timing of applica-tion. This is a critical aspect in the control of
T. pityocampa, because the duration of egghatching is very long and only those larvae inearly instars die with the current doses used.4

EFFECT OF B. thuringiensis ON THE
Other formulation bio-characteristics, such asactivity, stability, potency, and dose-responserelationships, are also being investigated as sig-nificant factors affecting field efficacy, whichstill remains relatively inconsistent compared touse of chemical insecticides (8, 9).
Due to its environmental safety, B. thurin-giensis has become a major tool in forest protec-tion programs for the control of defoliatingLepidoptera (7, 10). In order to demonstrate thatB. thuringiensis-based bioinsecticides representa good alternative approach for the control ofT. pityocampa, the first step to be taken is toexplore the potential use of lepidopteran-activetoxins in this insect. In a previous paper, weestablished that Cry1A-type B. thuringiensistoxins were highly toxic to first instar T. pityo-campa larvae (11) and, according to the acceptedmode of action of B. thuringiensis toxins, nowwe demonstrated that they specifically bind tothe insect midgut (11). To give a complete pic-ture of the activity spectrum of B. thuringiensisin this insect, in the present research, toxicities ofother lepidopteran-active toxins (Cry1B, Cry1C,
and Cry1E) and B. thuringiensis-based bioinsec-ticides (FORAY 48B, FORAY 76B, DIPEL, andCORDALENE) have been investigated. Wehave also evaluated the histopathologicalresponse to B. thuringiensis toxins and we haveanalyzed the receptor model of the active toxins.MATERIAL AND METHODS
Bacillus thuringiensis Toxins
The Cry1Aa, Cry1Ab, Cry1B, Cry1C, andCry1E toxins were obtained by Plant GeneticSystems (Gent, Belgium) as recombinant pro-teins expressed in Escherichia coli. The Cry1Aagene was cloned from B. thuringiensis var. aiza-wai HD-68, the Cry1Ab gene from B. thurin-giensis var. berliner 1715, and the Cry1E gene
from B. thuringiensis var. darmstadiensis HD-146. The Cry1B and Cry1C genes were clonedfrom B. thuringiensis var. entomocidus HD-110.The Cry1Ac toxin was isolated from B. thurin-giensis var. kurstaki HD73. The toxic fragmentof each d-endotoxin was obtained by trypsinPROCESSIONARY CATERPILLAR 45
digestion (12) and further purified as describedin Hofmann et al. (13).
Insect Toxicity Assays
T. pityocampa larvae were collected from nat-ural populations in Burjassot (Valencia, Spain).An artificial diet has not yet been developed forthis insect, therefore, larvae were fed on pineneedles (Pinus sylvestris) at room temperature.
Bioassays were carried out to determine thesensitivity of first instar T. pityocampa larvae toB. thuringiensis toxins. Each bioassay consistedof five doses of the corresponding trypsin-acti-vated crystal protein (Cry1B, Cry1C, andCry1E) prepared in PBS containing 0.1% BSAand a control of PBS, pH 7.4, 0.1% BSA. FreshP. sylvestris needles were dipped in a suspensionof the crystal protein and allowed to air dry.Three groups of twenty 1-day-old larvae wereplaced onto the coated pine needles for 4 days,after which the coated needles were replacedwith fresh, untreated needles. Mortality wasrecorded 4 days later. Toxicity data were evalu-ated by probit analysis using the Polo PC pro-gram (14). LC50 measurements refer to theconcentration of crystal protein which, whenapplied as a uniform covering on the needles,produces 50% mortality.
Using the previously described procedure,toxicity of the commercial formulations COR-DALENE (7.5 BIU/l, Ecogen), DIPEL LA(16.55 BIU/l, Abbot), FORAY 48B (12.7 BIU/l, Novo Nordisk), and FORAY 76B (20 BIU/l,Novo Nordisk) was also tested on T. pityocampafirst instar larvae. The relative composition ofthe Cry1A toxins in the different B. thuringiensiscommercial formulations is available only forDIPEL, which has been reported to be 28%Cry1Aa, 53% Cry1Ab, and 19% Cry1Ac (15).CORDALENE contains Cry1Aa, Cry1A(c), andCry2A protein toxins (Ecogen Europe S.r.l.,pers. comm.).
Toxin Administration and Tissue SectionStaining
Second instar larvae were fed with pine nee-dles previously dipped into solutions containing

46 RAUSEL
1 mg/ml Cry1Aa, 10 ng/ml Cry1Ab, 10 ng/mlCry1Ac, 10 ng/ml Cry1B, 1 mg/ml Cry1C, and 1mg/ml Cry1E trypsin-activated toxins. In controllarvae, pine needles soaked in bovine serumalbumin solution (1 mg/ml) were used instead.Three larvae were used in each experiment andtheir midguts were isolated 3 h 30 min afterfeeding.
Histological tissue section preparation, sec-tioning, and staining was performed accordingto Bravo et al. (16). Midguts were fixed in 10%Bouin Hollande’s sublimate fixative (17). After30-min incubation, tissues were transferred tofresh fixative solution for 24 h and washed for12 h in distilled water.
The tissue was dehydrated in 70% ethanol for1 h, 96% ethanol for 2 h, and twice 2 h in 100%ethanol. Tissue was infiltrated in xylol: 100%ethanol (50:50, v/v) for 1 h, xylol for 1 h,xylol:Paraplast (50/50, v/v) at 568C for 12 h,and finally 100% Paraplast for 48 h. Afterembedding, the Paraplast was hardened at 08C.
Transverse 5-mm-thick sections were cut witha HM-330 Microm microtome and placed ontomounting glasses coated with a 1 mg/ml poly-L-lysin solution.
Tissue sections were deparafinated andhydrated by successive immersions for 3 mintwice in xylol 100%, 3 min three times in abso-lute ethanol, and in distilled water for 1 min.Surplus Bouin Hollande’s sublimate wasremoved by incubation twice in Lugol solution(0.5% I2, 1% KI in distilled water) and sectionswere bleached with 5% sodium thiosulphate.Finally, sections were washed in distilled water,equlibrated for 5 min in TST buffer (10 mMTris–HCl, pH 7.6, 150 mM NaCl, 1 mM thimer-osal, 0.1% Triton X-100) and stained with theHeidenhain’s Azan method (18).
Binding of Toxins to Midgut Histological
SectionsMidgut histological sections of early secondinstar larvae were prepared following the proce-dure described in the previous section, andbinding of B. thuringiensis toxins was immuno-chemically detected according to Bravo et al.
L ET AL.
(19). Nonspecific binding was blocked by 1-hincubation of rehydrated sections with 0.5 ml of3% BSA in TST buffer (10 mM Tris–HCL, 100mM NaCl, 1 mM thimerosal, 0.1% (v/v) TritonX-100, pH 7.6). Tissue sections were incubatedfor 1 h with 0.3 ml of Cry1Aa, Cry1Ab, Cry1Ac,Cry1B, Cry1C, and Cry1E toxins (10 mg/ml inTST buffer). Unbound toxin was removed byrinsing in two changes of 0.5 ml of TST buffer.Bound toxin was immunochemically visualized,incubating overnight with a monoclonal primaryantibody specific against the toxins (1 mg/ml inTST buffer) and 1 h with a 1:1000 dilution ofan alkaline phosphatase rabbit anti-mouse sec-ondary antibody (Sigma A-4312). Color devel-opment was obtained by incubation of 0.5 mlof BCIP/NBT solution (1.75 mg 5-bromo-4-chloro-3-indoyl phosphate and 3.75 mg 4-nitro-tetrazolium chloride in 5 mM MgCl2, 100 mMTris, 100 mM NaCl, pH 9.5) for 10 min. Aftercolor reaction, the tissue sections were dehy-drated by successive incubations in ethanol andxylol and finally mounted in Entellan (Merk,Germany). Controls without toxin and withoutprimary antibody were also included.
Preparation of Brush Border MembraneVesicles (BBMV) and Binding ofBiotinylated Toxins
BBMV from T. pityocampa early secondinstar larvae were prepared as described byWolfersberger et al. (20). The final pellet wasresuspended in ice-cold MET buffer (0.3 M man-nitol, 5 mM EGTA, 17 mM Tris–HCI, pH 7.5)and immediately frozen and stored at 2808C.The amount of BBMV protein was measuredusing Bradford’s procedure (21) with the Bio-Rad (Richmond, CA) kit, using BSA as astandard.
Cry1Aa, Cry1Ab, and Cry1Ac were biotinyl-
ated according to the Denolf et al. procedure (22)and binding assays were performed following aPlant Genetic System modified protocol of VanRie et al. (23). Ten nanograms of biotinylatedCry1Aa or Cry1Ab toxins and 30 ng of biotinyl-ated Cry1Ac toxin were used per 100 ml of thereaction mixture. The toxin bound to BBMV was
EFFECT OF B. thuringiensis ON THE
separated from the free toxin by centrifugation at15,000g. The resulting pellet, containing BBMVproteins and the bound biotinylated toxin, wassubjected to SDS–PAGE and electroblotting.Nonspecific binding was blocked by 2-h incuba-tion of the filter with 2% Tween 20 in TBSbuffer (50 mM Tris, 150 mM NaCl, pH 7.5).Following 45-min incubation with 1:3000(v/v) dilution of Streptavidin–alkaline phospha-tase (Amersham, U.K.) in TBS buffer, the filterwas washed four times for 5 min and for 15 minwith 2% Tween 20 in TBS buffer. The CSPD
chemiluminiscence substrate (Boehringer Man-nheim, Germany) was added to the filter andexposed to an X-ray film to detect the boundtoxin. For competition experiments, binding ofthe biotinylated toxins was analyzed in the pres-ence of a 300-fold excess of unlabeled toxins(Cry1Aa, Cry1Ab, Cry1Ac, and Cry1B).RESULTS
Bioassays of Cry1 Toxins withThaumetopoea pityocampa
Concentration–mortality assays with Cry1B,Cry1C, and Cry1E were added to the previouslyreported data for the Cry1A-type proteins (11)
on Table 1. All three Cry1A-type toxins wereCry1Aaa 1.86 6 0.36Cry1Aba 3.55 6 1.11Cry1Aca 2.68 6 0.55Cry1B 16.98 6 4.65 1Cry1CCry1E
a Data from Rausell et al. (11).* 90% fiducial limits.
PROCESSIONARY CATERPILLAR 47
Histopathological Effects of Bacillusthuringiensis Toxins on the Midgut ofThaumetopoea pityocampa
Toxins that proved to be active against T. pity-ocampa were administered to first instar larvae,and cytopathological changes in midgut epithe-lial cells were investigated. Typical histopatho-logical effects, including disruption of the brushborder membrane, vacuolization of cytoplasm,and swelling and disintegration of the cells, wereobserved in midgut tissue sections (Fig. 1). Nocytological changes were detected in the midgutepithelium of larvae treated with the nontoxicCry1C and Cry1E proteins (Fig. 1).
Bioassays of Bacillus thuringiensis-BasedBioinsecticides with Thaumetopoeapityocampa
LC50 and LC90, as well as the slopes obtainedfrom toxicity assays using commercial formula-tions, were of the same magnitude for all ofthem (Table 2), and within the same value rangeobtained for the Cry1A toxins previouslyassayed.
In order to determine to what extent the toxic-ity generated by commercial formulations isinfluenced by its toxin content, probit analysis
of the combined concentration–mortality values of the toxins and the bioinsecticides previouslyhighly toxic, the order of efficiency beingCry1Ac . Cry1Ab 5 Cry1Aa (11). Cry1B assayed was carried out. From all possible com-binations, the statistical analysis using theshowed a different behavior, with a higher LC50
but a LC90 similar to those determined for Cry1A POLO-PC program (14) accepted as a homoge-neous set of data, from which a single concentra-toxins. Cry1C and Cry1E showed no activity
against T. pityocampa. tion–mortality curve could be inferred, only the
TABLE 1Slope, LC50, and LC90 Determined by the POLO-PC Program for Concentration–Mortality Assays with Bacillus
thuringiensis Toxins against First Instar Thaumetopoea pityocampa Larvae
Toxin Slope 6 SE LC50 (pg/ml) [FL95] LC90 (pg/ml) [FL95]
956 [586–1552]* 4680 [2434–35899]*895 [694–1687]* 2054 [1276–14902]*379 [279–543] 1141 [735–2859]830 [1721–1918] 2178 [2040–2635]
Not toxicNot toxic
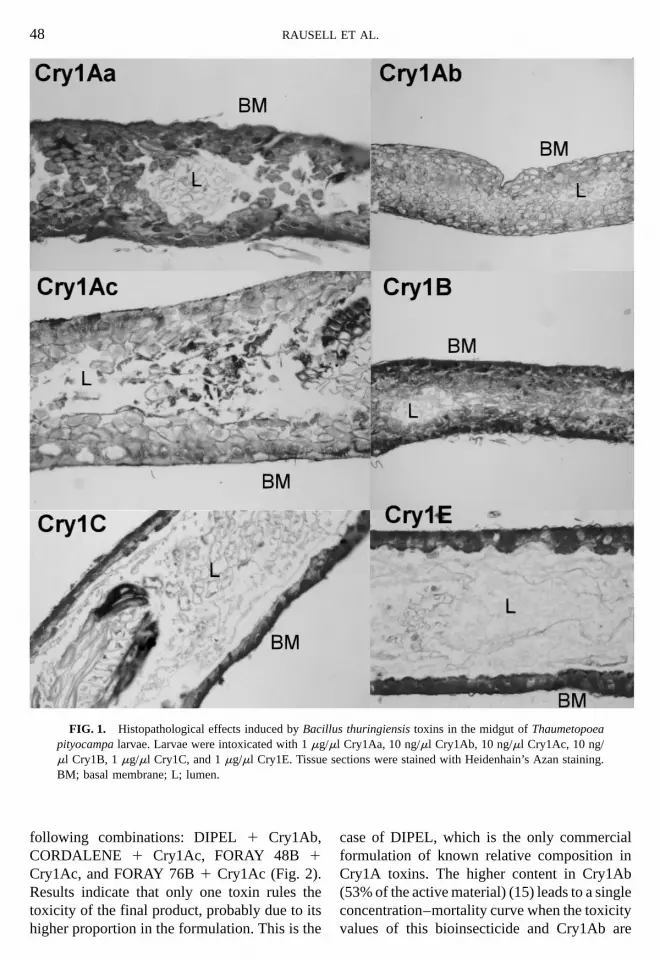
FIG. 1. Histopathological effects induced by Bacillpityocampa larvae. Larvae were intoxicated with 1 mg/mml Cry1B, 1 mg/ml Cry1C, and 1 mg/ml Cry1E. TissueBM; basal membrane; L; lumen.
following combinations: DIPEL 1 Cry1Ab,CORDALENE 1 Cry1Ac, FORAY 48B 1Cry1Ac, and FORAY 76B 1 Cry1Ac (Fig. 2).Results indicate that only one toxin rules thetoxicity of the final product, probably due to itshigher proportion in the formulation. This is the
us thuringiensis toxins in the midgut of Thaumetopoeal Cry1Aa, 10 ng/ml Cry1Ab, 10 ng/ml Cry1Ac, 10 ng/
sections were stained with Heidenhain’s Azan staining.
48 RAUSELL ET AL.
case of DIPEL, which is the only commercialformulation of known relative composition inCry1A toxins. The higher content in Cry1Ab(53% of the active material) (15) leads to a singleconcentration–mortality curve when the toxicityvalues of this bioinsecticide and Cry1Ab are

Slope, LC50, and LC90 Determined by the POLO-PC Program for Concentration–Mortality Assays with Bacillusthuringiensis-Based Bioinsecticides against First Instar Thaumetopoea pityocampa Larvae
Product Slope 6 SE
DIPEL ES 2.62 6 0.382FORAY 48B 0.990 6 0.191FORAY 76B 1.478 6 0.264CORDALENE 1.373 6 0.228
combined. However, the combination of concen-tration–mortality responses of DIPEL andCry1Aa or Cry1Ac was not accepted as a homo-geneous set of data.
Immunocytochemical Detection of theBinding of Bacillus thuringiensis Toxins toThaumetopoea pityocampa Midgut TissueSections
Specific binding of Cry1Aa, Cry1Ab,Cry1Ac, and Cry1B to the brush border mem-brane of T. pityocampa larvae was observed(Fig. 3). The nontoxic Cry1C and Cry1E proteins(Fig. 3) and controls without toxin or primaryantibody (data not presented) showed no bindingto the midgut brush border membrane.
Binding of Biotinylated Toxins toThaumetopoea pityocampa BBMV
To establish the receptor model for the toxinsactive against T. pityocampa, binding competi-tion experiments with biotinylated toxins onearly stage larvae were performed. Cry1Aa,Cry1Ab, and Cry1Ac, previously shown to beactive (11), were biotinylated and competition
EFFECT OF B. thuringiensis ON THE PROCESSIONARY CATERPILLAR 49
TABLE 2
experiments in the presence of an excess of non-labeled Cry1Aa, Cry1Ab, Cry1Ac, and Cry1Bwere carried out (Fig. 4). Results indicate thatall Cry1A-type toxins compete were able to bind
to brush border midgut membrane with eachother for the same binding sites. We also demon-strated that Cry1B did not share them.DISCUSSION
In this study, we investigated the relationshipsamong toxicity, histopathological effects, and
LC50 (pg/ml) [FL95] LC90 (pg/ml) [FL95]
667 [502–978] 2834 [1700–7806]264 [135–513] 5294 [2059–35088]304 [146–733] 2236 [868–41014]488 [369–697] 3824 [2078–11198]
binding of Cry1 B. thuringiensis toxins in T.pityocampa larvae.
In a previous paper we reported that Cry1Aa,Cry1Ab, and Cry1Ac toxins were highly toxicto T. pityocampa larvae (11). In the presentresearch we demonstrated that Cry1B was alsohighly active against this insect pest, whileCry1C and Cry1E were found to be nontoxic(Table 1).
In conclusion, toxicity assays showed thatCry1A and Cry1B B. thuringiensis toxins werehighly active against T. pityocampa and, there-fore, suitable for controlling this insect pest.
In order to compare the efficacy of the toxins,not only the LC50 value has to be taken intoaccount but also the slope of the concentration–mortality regression line because, in fact, LC90
represents the minimum level of efficacyrequired to appropriately reduce a pest in thefield (24). In the case of Cry1B, although ithas a significantly higher LC50 than that of theCry1A toxins, both types of toxins have a similarLC90 and, therefore, comparable field efficacyis expected for both of them.
Intoxication assays, carried out with firstinstar T. pityocampa larvae forced to feed onindividual toxins, showed that those concentra-tions causing 98% or higher mortality (probit.7) in bioassays were able to produce cytotoxic
effects (Fig. 1). Furthermore, such results corre-lated with the existence of in vitro binding ofthe active toxins to the brush border membraneof the larvae midgut (Fig. 3). All the facts arein accordance with the proposed mode of actionof lepidopteran-active B. thuringiensis toxins, inwhich binding to specific receptors in the insect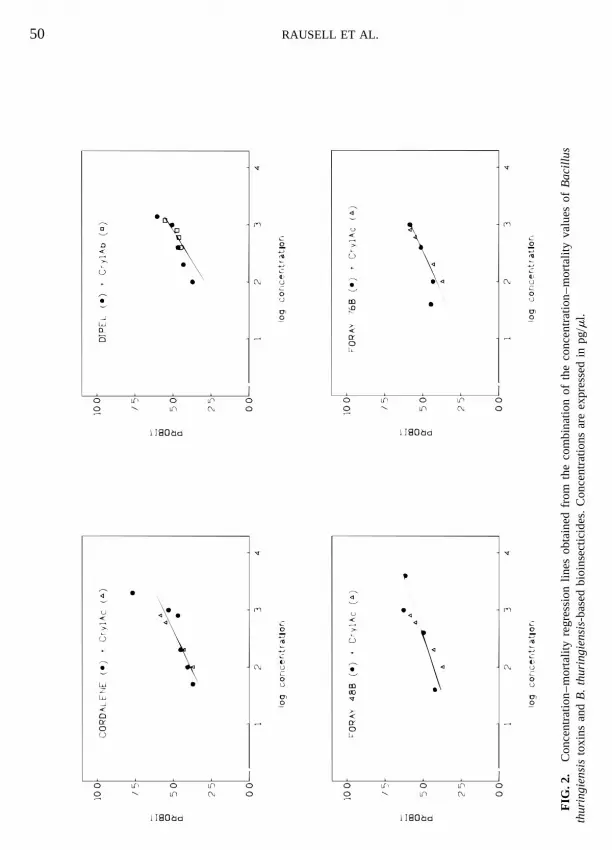
50
RAUSELL ET AL.onli
nes
obta
ined
from
the
com
bina
tion
ofth
eco
ncen
trat
ion–
mor
tali
tyva
lues
ofB
acil
lus
dbi
oins
ecti
cide
s.C
once
ntra
tion
sar
eex
pres
sed
inpg
/ ml.
FIG
.2.
Con
cent
rati
on–m
orta
lity
regr
essi
thur
ingi
ensi
sto
xins
and
B.
thur
ingi
ensi
s-ba
se

FIG. 3. Binding of Bacillus thuringiensis toxins tolarvae midguts. BM; basal membrane; MV; microvilli;
midgut epithelium determines the insecticidalactivity of toxins.
Different methods were used in order to dem-onstrate the binding of activated toxins to spe-cific receptors. Specific binding of activated
toxins was detected in insect cell cultures, histo-logical sections, and brush border membranevesicles, and the bound toxin was localized byusing specific antibodies or labeling the toxinswith fluorescein, 125I, or biotin (25, 13, 16, 23,22). Calculation of binding parameters is possi-ble only when performing in vitro experimentsbrush border membrane of Thaumetopoea pityocampaL; lumen.
with 125I-labeled toxins and brush border mem-brane vesicles (23).
An alternative method with biotinylated tox-ins, coupled to Enhanced Chemiluminescence(ECL) detection, has been recently developed
EFFECT OF B. thuringiensis ON THE PROCESSIONARY CATERPILLAR 51
by Plant Genetic Systems (Gent, Belgium). Thismethod provides qualitative information aboutbinding sites and their relationships and over-comes some of the disadvantages related to theuse of radioactive isotopes. Using this technique,we concluded that Cry1Aa, Cry1Ab, andCry1Ac recognize the same binding site (Fig.

52
RAUSELL ET AL.cede
tect
ion
ofin
vitr
obi
ndin
gof
biot
inyl
ated
toxi
nsto
earl
yse
cond
inst
arT
haum
etop
oea
pity
ocam
pabi
otin
ylat
edC
ry1A
b;(C
)bi
otin
ylat
edC
ry1A
c.F
orea
chas
say:
lane
1,bi
otin
ylat
edto
xin;
lane
2,ea
rly
Vs;
lane
3,bi
otin
ylat
edto
xin
and
proc
cess
iona
rym
oth
BB
MV
s;la
nes
4,5,
and
6,bi
otin
ylat
edto
xin
and
nce
ofan
exce
ssof
Cry
1Aty
peto
xins
(Cry
1Aa,
Cry
1Ab,
and
Cry
1Ac,
resp
ecti
vely
);an
dla
ne7,
biot
inyl
ated
nth
epr
esen
ceof
anex
cess
ofC
ry1B
toxi
n.A
rrow
spo
int
atba
nds
corr
espo
ndin
gto
the
biot
inyl
ated
toxi
ns
FIG
.4.
Enh
ance
dch
emil
umin
esce
nB
BM
Vs.
(A)
Bio
tiny
late
dC
ry1A
a;(B
)se
cond
inst
arpr
occe
ssio
nary
mot
hsB
BM
proc
cess
iona
rym
oth
BB
MV
sin
the
pres
eto
xin
and
proc
cess
iona
rym
oth
BB
MV
si
(60
KD
aap
prox
).

ACKNOWLEDGMENTS
EFFECT OF B. thuringiensis ON THE
4), confirming earlier data from binding experi-ments with 125I-radiolabeled Cry1Ab toxin (11).Moreover, we demonstrated that this binding sitewas not recognised by Cry1B (Fig. 4, lanes 7).These data indicate that in T. pityocampa at leasttwo different types of receptors can be distin-guished. One receptor binds the three Cry1A-type toxins and another one recognizes theCry1B toxin.
Results provide very useful information foroptimizing the use of B. thuringiensis toxinsagainst T. pityocampa. Mixtures or rotations oftoxins with different target sites may be a valu-able strategy in delaying the development ofresistance (26). This is an important concernbecause the use of B. thuringiensis products withimproved efficacy is expected to increase and,therefore, the level of selection for resistanceshould also be enhanced (27). In T. pityocampa,combined or rotational application of Cry1Bwith a Cry1A toxin should be an appropriatechoice.
Commercial B. thuringiensis products arebased on spore–crystal mixtures and consist ofd-endotoxins as the active principle. Therefore,we could expect efficacy of these formulationsto correlate with their toxin content. However,there is considerable confusion regarding thisissue. Van Frankenhuyzen et al. (28) reportedsynergistic interactions of HD-1 toxins (a strainthat forms the basis for most formulations usedin agriculture and forestry) in three insect species(Lymantria dispar, Choristoneura pinus, andOrygia leucostigma). Tabashnik (29) reevalu-ated these data, as well as previously publisheddata on the interaction of B. thuringiensis toxins,but failed to find any evidence of synergism.Nevertheless, Lee et al. (30) observed synergismwith a mixture of Cry1Aa and Cry1Ac andantagonism with a mixture of Cry1Aa andCry1Ab in L. dispar.
Our study, carried out with B. thuringiensis-based bioinsecticides, showed that the toxicityof bioformulates was of the same magnitudeas that of individual toxins and, therefore, nosynergism was observed. However, a synergisticeffect of a toxin combination in the mixture thatmight be masked by the antagonistic effect of
PROCESSIONARY CATERPILLAR 53
another toxin combination in the formulationcannot be ruled out.
Interestingly, statistical comparison of theconcentration–mortality values of bioinsectic-ides and individual toxins indicated that the tox-icity of bioformulates is determined mainly byone of the toxins present in the bioinsecticide(Cry1Ab or Cry1Ac). Taking into account thatCry1Ab toxicity decreases with age as a conse-quence of the loss of a binding site in T. pityo-campa (11) and the very long duration of egghatching, in order to improve their efficacy, B.thuringiensis-based commercial formulations
against this insect should be enriched in Cry1Ac,the toxin that displays the highest toxicity.By jointly considering our research and thatof Rausell (11), we can conclude that the enrich-ment of bioformulates with Cry1Ac and Cry1Bwill allow an effective and reliable use of Crytoxins in T. pityocampa management.
Research was funded by the Spanish Ministerio de Agri-cultura Pesca y Alimentacion (Project No. AGF93-1171).Carolina Rausell and Inmaculada Garcıa-Robles were sup-ported by grants from the Spanish Ministerio de Educaciony Ciencia and the Conselleria Valenciana de’Educacio iCiencia, respectively.
REFERENCES
1. G. Demolin, J. F. Abgrall, and L. Bouhot-Delduc, Evo-lution de l’aire de la processionaire du Pin en France,Les cahiers du DSF 1, 26 (1996).
2. N. Romanyc and D. Cadahia coord., “Plagas de insectosen las masas forestales espanolas” (Ministerio deAgricultura, Pesca y Alimentacion ed.) pp 47–84, ISBN84-7479-125-1 (1992).
3. A. Battisti, S. Longo, R. Tiberi, and O. Triggiany,Results and perspectives in the use of Bacillus thurin-giensis Berl. Var. kurstaki and other pathogens againstThaumetopoea pityocampa (Den. et Schiff.) in Italy
(Lep., Thaumetopoeidae), Anz. Schalingskde. Pflan-zenschutz Umweltschutz 71, 72 (1998).4. G. Demolin and A. Millet, Essais insecticides contre laProcessionaire du pin Thaumetopoea pityocampaSchiff.). Action comparative de differentes specialitescommerciales Bactospeine, Dipel, Thuricide et Dimilin,Ann. Sci. For. 38, 389 (1981).
5. G. Demolin, J. C. Martin, and P. Lavanceau, Lutte contre

54 RAUSELL
la Processionaire du Pin. L’evolution des insecticides abase de Bacillus thuringiensis, Phytoma 452, 13 (1993).
6. S. S. Gill, E. A. Cowles, and P. V. Pietrantonio, Themode of action of Bacillus thuringiensis endotoxins,Annu. Rev. Entomol. 37, 615 (1992).
7. K. Van Frankenhuyzen, The challenge of Bacillus thur-ingiensis, in “Bacillus thuringiensis, An EnvironmentalBiopesticide: Theory and Practice” (P.F. Entwistle, J.S. Cory, M. J. Bailey, and S. Higgs, Eds.), pp. 1–35,Wiley, New York, 1993.
8. D. R. Dent, The use of Bacillus thuringiensis as aninsecticide, in “Exploitation of Microorganisms” (M. G.Ford, Ed.), pp. 19–44, Chapman & Hall, London, 1993.
9. E. Schnepf, N. Crickmore, J. Van Rie, D. Lereclus, J.Baum, J. Feitelson, D.R. Zeigler, and D.H. Dean, Bacil-lus thuringiensis and its pesticidal crystal proteins,Microbiol. Mol. Biol. Rev. 62, 775 (1998).
10. K. van Frankenhuyzen, The development of Bacillusthuringiensis for control of spruce budworm, Choristo-neura fumiferana, in Canada, in “Bacillus thuringiensisBiotechnology and Environmental Benefits” (J.Y. Feng,Ed.), pp. 405–423, Hua Shiang Yuan, Taipei, Taiwan,1995.
11. C. Rausell, A. C. Martınez-Ramırez, I. Garcıa-Robles,and M.D. Real, A binding site of Bacillus thuringiensisCry1Ab toxin is lost during larval development in twoforest pests, submitted for publication.
12. H. Hofte, H. De Greve, J. Seurinck, S. Jansens, J. Mahil-lon, C. Ampe, J. Vandekerckhove, H. Vanderbruggen,M. Van Montagu, M. Zabeau, and M. Vaeck, Structuraland functional analysis of a cloned delta-endotoxin ofBacillus thuringiensis berliner 1715, Eur. J. Biochem.161, 273 (1986).
13. C. Hofmann, H. Vanderbruggen, H. Hofte, J. Van Rie,S. Jansen, and H. Van Mellaert, Specificity of Bacillusthuringiensis d-endotoxins is correlated with the pres-ence of high affinity binding sites in the brush bordermembrane of target insect midguts, Proc. Natl. Acad.Sci. USA 85, 7844 (1988).
14. LeOra Software, POLO-PC. LeOra Software, Berkeley,California, 1987.
15. Y. B. Liu, B. E. Tabashnik, and M. Pusztai-Carey, Field-
evolved resistance to Bacillus thuringiensis toxin Cry1Cin diamondback moth (Lepidoptera: Plutellidae), J.Econ. Entomol. 89, 798 (1996).16. A. Bravo, S. Jansens, and M. Peferoen, Immunocyto-chemical localization of Bacillus thuringiensis insectici-dal crystal proteins in intoxicated insects, J. Invertebr,Pathol. 60, 237 (1992)
17. G. Clark, Heidenhain’s Azan modification stainingmethod, in “Staining Procedures (G. Clark, Ed.), pp.117–118, Williams & Wilkins, London, 1981.
18. P. Brandtzaeg, Tissue preparations methods for immu-nocytochemistry, in “Techniques in Immunocytochem-
ET AL.
istry” (G. R. Bullock and P. Petrusz, Eds.), pp. 1–76,Academic Press, London, 1982.
19. A. Bravo, K. Hendrickx, S. Jansens, and M. Peferoen,Immunocytochemical analysis of specific binding ofBacillus thuringiensis insecticidal crystal proteins tolepidopteran and coleopteran midgut membranes, J.Invertebr. Pathol. 60, 247 (1992).
20. M. G. Wolfersberger, P. Luthy, A. Maure, P. Pareti, F.V. Sacchi, B. Giordana, and G. M. Hanozet, Preparationand partial characterization of aminoacid transportingbrush border membrane vesicles from the larval midgutof the cabbage butterfly (Pieris brassicae), Comp. Bio-chem. Physiol. 86, 301 (1987).
21. M. M. Bradford, A rapid and sensitive method for thequantitation of microgram quantities of protein utilizingthe principle of protein-dye binding, Anal. Biochem.72, 248 (1976).
22. P. Denolf, S. Jansens, J. Van Rie, D. Degheele, and M.Peferoen, Biotinylation of Bacillus thuringiensis insec-ticidal crystal proteins, Appl. Environ. Microbiol. 59,1821 (1993).
23. J. Van Rie, S. Jansens, H. Hofte, D. Degheele, and H.Van Mellaert, Specificity of Bacillus thuringiensis d-endotoxins: Importance of specific receptors on thebrush border membrane of the midgut of target insects,Eur. J. Biochem. 186, 239 (1989).
24. R. C. Reardon and D. L. Wagner, Impact of Bacillusthuringiensis on nontarget lepidopteran species inbroad-leaved forests, in “Biorrational Pest ControlAgents” (F. R. Hall and J. W. Barry, Eds.), pp 284–292,ACS Symp. Series, Washington DC, 1995.
25. C. Hofmann, P. Luthy, R. Hutter, and V. Pliska 1988.Binding of the delta-endotoxin from Bacillus thurin-giensis to brush-border membrane vesicles of the cab-bage butterfly (Pieris brassicae), Eur. J. Biochem. 173,85 (1988).
26. B. E. Tabashnik, Evolution of resistance to Bacillusthuringiensis, Annu. Rev. Entomol. 39, 47 (1994).
27. G. Kennedy and M. Whalon, Managing pest resistanceto Bacillus thuringiensis endotoxins: Constraints andincentives to implementation, J. Econ. Entomol. 88,454 (1995).
28. K. van Frankenhuyzen, J. L. Gringorten, R. E. Milne,D. Gauthier, M. Pusztai, R. Brousseau, and L. Masson,Specificity of activated CryIA proteins from Bacillusthuringiensis subsp. kurstaki HD-1 for defoliating forestLepidoptera, Appl. Environ. Microbiol. 57, 1650 (1991).
29. B. E. Tabashnik, Evaluation of synergism among Bacil-lus thuringiensis toxins, Appl. Entomol. Microbiol. 58,3343 (1992).
30. M. K. Lee, A. Curtiss, E. Alcantara, and D. H. Dean,Synergistic effect of the Bacillus thuringiensis toxinsCryIAa and CryIAc on the gypsy moth, Lymantria dis-par, Appl. Environ. Microbiol. 62, 583 (1996).








