Embed Size (px)
Citation preview

JOURNAL OF ULTRASTRUCTURE RESEARCH 70, 275-286 (1980)
The Structure of Sperm from the Lobster, Homarus americanus
PRUDENCE TALBOT*' t AND PRANEET CHANMANON *J
*Bodega Marine Laboratory, Bodega Bay, California 94923, and tDepartment of Biology, University of California, Riverside, California 92521
Received August 9, 1979
The structure of sperm from the lobster, Homarus americanus, was examined using light and electron microscopy. These sperm consist of an acrosome, subacrosomal region, collar, nucleus, and three spikes which are extensions of the nucleus. The aerosome is a cylinder projecting from the sperm's antierior pole. It is comprised of inner acrosomal material, outer acrosomal material, and an apical cap, and is limited by a single continuous membrane. The subacrosomal material is finely granular with most of it lying in a fossa in the base of the aerosome. The collar, a region subjacent to the subacrosomal material, contains small mitochrondria and a pair of centrioles. The subaerosomal material and collar are in direct continuity with the ehromatin of the nucleus. The nucleus itself is limited by a highly folded, thick membrane which probably represents a composite of the plasma membrane and nuclear envelope. The chromatin is granular or fibrillar and is uneondensed. The spikes are compartments of the nucleus and contain microtubules ensheathed by sheets of membrane. Ionophore A23187 which is fluorescent inserts into membranes of cells and was used to study the distribution of spike membranes in sperm with the aid of a fluorescent microscope. The mierotubule/membrane complexes of the spikes converge in the region of the collar and interconnect to form a three-sided vault the apex of which terminates immediately beneath the base of the aerosome. These lobster sperm are similar in structure to other reptantian sperm.
The unusual structure of decapod sperm has attracted much research interest. The morphology and development of such sperm have been described by both light microscopists (2-4, 14-19, 23, 25-27, 30, 34, 42, 45, 46) and electron microscopists (1, 5- 12, 20-22, 24, 28, 31-33, 35-40, 44, 48, 49). In addition, features of the fertilization se- quences have been analyzed ultrastruc- turaUy for two genera of crabs (6, 21). The structure of lobster sperm has been inves- tigated previously for three genera: Ne- phrops (9-13), Homarus vulgaris (36-40), and Panulirus (44). In brief, these studies have shown that lobster sperm are charac- terized by a highly complex acrosomal ves- icle, an uncondensed nucleus, and three (Homarus, Nephrops) or more (Panulirus) spikes which are extensions of the nuclear compartment. Although the spikes are packed with microtubules, they are station- ary and do not contribute to sperm motility.
' Present address: Department of Biology, Texas Woman's University, Denton, Texas 76201.
Reproduction in lobsters is not well un- derstood, and the sperm's unusual form suggests that fertilization may not follow the plan of more commonly studied inver- tebrates, such as the sea urchin. For this reason, we initiated studies on the repro- ductive systems of male and female lobsters with the long range goal of understanding gametogenesis, gamete morphology, ovula- tion, and fertilization. This paper reports our findings on the ultrastructure of mature sperm from Homarus americanus: a sub- sequent article describes this sperm's acro- some reaction. Our results extend the ear- lier works of Pochon-Masson (40) on Hom- arus vulgaris, although they differ some- what in interpretation.
MATERIALS AND METHODS
The testes and vas deferens from sexually mature lobsters (Homarus americanus) were dissected during June and July. These organs were rinsed in seawater (SW) to disperse the sperm which, after 20 rain, were transferred to a fresh container for further study,
The structure of lobster sperm was examined using
275 0022-5320/80/030275-12502.00/0 Copyright © 1980 by Academic Press, Inc. All rights of reproduction in any form reserved.

276 TALBOT AND CHANMANON
light and transmission electron microscopy (LM and TEM). For LM, fresh and formaldehyde-fixed sperm in SW were examined and photographed using a Reichert microscope equipped with phase contrast and dark field optics. For TEM, testes, vas deferens, or sperm pellets were fixed 1-3 hr in 3% glutaralde- hyde/SW or in 1% acrolein/3% glutaraldehyde/SW, washed several hours in SW, postfixed 0.5-1.0 hr in 1% osmium tetroxide/SW, washed in SW, dehydrated in a graded acetone series, infiltrated in acetone/plas- tic, and embedded in plastic (43). Thin sections were cut on a Porter-Blum MT-2B or LKB-I ultramicro- tome using a diamond knife. Sections were picked up on copper grids, stained with uranium and lead salts, and examined with a Hitachi H-500 electron micro- scope.
The ionophore A23187 was also used to study sperm structure. A23187 inserts into lipid bilayers and cellu- lar membranes (41) and, because it is fluorescent, its distribution in cells can be directly visualized with a fluorescent microscope. It is useful in identifying cel- lular regions which have an abundance of membrane. Sperm were fixed 20 rain in 4% formaldehyde/SW, washed in SW, incubated 3 rain in 10/~M A23187/SW, washed again in SW, and examined with a Reichert uv microscope equipped with a fluorescein filter set. Regions of the sperm with high concentration of A23187 fluoresced bright blue.
RESULTS
H. a m e r i c a n u s sperm consist of an acro- some, subacrosomal region, collar, nucleus, and three spikes which are compartments of the nucleus (Figs. 1, 2, and 17). The sperm measures 19.3 ttm from the anterior end of the acrosome to the base of the nucleus. Each spike measures 37.8 ~m from the distal tip to the point of confluence with the nucleus proper. Each region will be considered in detail.
The acrosome. The acrosome ofH. amer-
i canus sperm is an elongated vesicle which forms a prominent cylinder at the sperm's anterior end (Figs. 1 and 2). It measures approximately 14 × 3-3.5/~m and is limited by a continuous, single membrane (acroso- real membrane). The tripartite image of this membrane is asymmetric; the leaflet adjacent to the acrosomal contents is thicker and more electron dense than the one adjacent to the plasma membrane (Figs. 5 and 10). It is often difficult to resolve the acrosomal membrane at the
base of the acrosome because the electron- dense contents of the acrosome at that point obscure the inner leaflet. However, examination of favorably sectioned mate- rim shows that the acrosomal membrane is present in this region (Fig. 10).
The contents of the acrosome are heter- ogeneous. For convenience in this report, they will be referred to as the "outer acro- somal material," the "inner acrosomal ma- terial," and the "apical cap" (Figs. I and 2).
The outer acrosomal material comprises the bulk of the acrosome and extends from the lateral edges of the apical cap to the base of the vesicle (Figs. 1 and 2). This mater ia l is cont inuous laterally with the acrosomal membrane and medially with the inner acrosomal material. It gives a strong positive reaction when treated with periodic acid-Schiffs reagent (PAS) and, ultrastructurally it has moderate-to-low electron density. Small wedge-shaped pro- jections of greater electron density radiate inward from the periphery of the outer acrosomal material (Figs. 1, 2, and 4).
The inner acrosomal material which gives a weak PAS reaction forms a highly electron-dense column running the length of the acrosome's center (Fig. 1). At the acrosome's anterior and posterior margins, this material flares out to form a flange at the base of the apical cap and a cup around the subacrosomal material (Figs. 1, 2, 6, and 7). The inner acrosomal material condenses during spermiogenesis; in mature sperm of the vas deferens, it appears homogeneous and electron dense in cross section (Fig. 4). In some testicular sperm, however, conden- sation is not yet complete; when this is the case, the periphery of the inner acrosomal material has a feathery appearance (not shown) and the center of the core is only moderately electron dense (Fig. 5). At the base of the acrosome, the inner acrosomal material is continuous with the wedge- shaped projections of the outer acrosomal material (Fig. 10). We do not yet know if the wedge-shaped projections and inner ac-

LOBSTER SPERM STRUCTURE
,..PM
_AM
- O A M
277
I
Fro. 1. Schematic diagram of H. americanus sperm. Arrows indicate approximate levels of section in Figs. 3-5. The acrosome includes the inner acrosomal material, outer acrosomal material, and apical cap. The apical cap includes four zones. The entire acrosome is limited by the acrosomal membrane. This membrane lines the fossa at the base of the acrosome, but is difficult to resolve here because of the electron density of the inner acrosomal material. Portions of the spike membranes are represented by dashed lines as the spikes are not drawn to full length. AC, apical cap; IAM, inner acrosomal material; PM, plasma membrane; AM, acrosomal membrane; OAM, outer acrosomal material; NC, nuclear cuff; SM, subacrosomal material; M, mitochondrion: CE, centriole; N, nucleus; MMC, microtuble/membrane complex; S, spikes.
rosomal mater ia l are biochemical ly the same.
T h e apical cap, located a t the anterior- mos t por t ion of the acrosome, m a y be sub- divided into four regions (Figs. 1-3 and 6- 8). T h e outer three of these are organized concentr ical ly into a ring, while the four th par t ia l ly fills the center of this ring (Figs. 1-3). T h e ring's ou te rmos t region (zone 1) has a regular subs t ruc ture which appears crystall ine (Figs. 3, 6, and 8). T h e interior of zone I is lined by an electron-dense layer (zone 2) which also appears crystalline, (Figs. 2, 3, 6, and 8). Zone 3, which is modera te ly electron dense and also crystal- line, lines zone 2 and is formed by a cup- shaped extension of the inner acrosomal mater ia l (Figs. 1, 2, 7, and 8). When a hand lens is used to examine micrographs of the apical cap, the pa t t e rn of the crystal is clearly cont inuous f rom the zone to the next. For example, Fig. 8 shows a section which grazes th rough several areas of the
apical cap. In cross-section, e lectron-lucent circles are outl ined by a mater ia l of mod- erate or heavy electron density. In tangen- tial sections, these circles appear tubular; close examina t ion of these tubules reveals tha t each is cont inuous across the three zones.
T h e innermos t region of the apical cap (zone 4) is modera te ly electron dense (equivalent to zone 1) and lacks crystalline structure. Zone 4 part ial ly fills in the center of the ring formed by zones 1-3; its poster ior region is cont inuous with the inner acroso- mal material . T h e apical cap has a deep depression in its center (Figs. 1 and 2), and unlike the outer acrosomal material , gives a weak posit ive react ion with PAS reagent. T h e p lasma m e m b r a n e around the anter ior edge of the apical cap is th rown out into abou t 16 finger-like projections. These are filled with a modera te ly electron-dense ma- terial (Fig. 9) which occupies the space be tween the p lasma and acrosomal mere-

oo
t~
©
c~

LOBSTER S P E R M S T R U C T U R E 279
brane and is not part of the acrosomal vesicle, per se.
The subacrosomal material and collar. At the base of the acrosome is a fossa sufficiently large (about 5/~m deep) to be seen with the light microscope (Fig. 2, in- sets). This fossa is occupied by the subacro- somal material, which appears finely gran- ular and moderately electron dense in thin sections (Figs. 2, 9, 10, and 16). In addition, it is neither PAS nor Feulgen positive. Ex- tending out from the base of the fossa is the subacrosomal material, a thin band of which follows the contour of the acrosomal vesicle around its lateral surfaces (Figs. 9 and 10). The posterior region of the suba- crosomal material is not only sometimes more electron dense than the material in the fossa proper (Fig. 10), but also is con- tinuous with the chromatin of the nucleus (Figs. 9 and 10).
Previous workers (37, 40) have referred to the region of the sperm immediately beneath the subacrosomal zone as the "col- lar." Included in this region are centrioles (not shown) and mitochondria (Figs. 9, 10, and 16). The latter are small, have poorly defined cristae, and contain electron-dense granules. Generally, these organelles are concentrated immedia te ly beneath the subacrosomal region, but occasionally they are observed extending a short distance out into the spikes. The spikes (see below) join each other in the collar (Figs. 15 and 16). Finally, the mitochondria, centrioles, and the spikes are all continuous with the chro- matin.
Nucleus and spikes. The Feulgen-posi-
rive nucleus contains uncondensed chro- matin which is finely fibrillar or granular and moderately electron dense. The chro- matin is continuous with the membrane limiting the sperm, but does not project out into the spikes. The nucleus extends around the acrosome's lateral edges, forming a small cuff containing chromatin at the ac- rosome's base (Fig. 9). Anterior to this cuff is the plasma membrane which is thinner and less electron dense than the membrane covering the cuff, nucleus, and spikes (Fig. 9). At high magnifications, the "plasma membrane" of the cuff, nucleus, and spikes sometimes has a pentilaminar appearance, suggesting that the membranes in these regions are a composite of true plasma membrane fused with former nuclear en- velope. Apparently, the nuclear envelope has degenerated between the base of the acrosome and the nucleus. The membrane limiting the body of the nucleus is thrown into many folds which greatly increase the membrane's surface area over the nucleus (Fig. 2, inset, and Fig. 11).
H. americanus sperm have three spikes, all of which are Feulgen negative. When sperm are suspended in SW, the spikes are stationary and the sperm show no mobility except for Brownian movements. The spikes appear to be lined by nuclear enve- lope fused with the plasma membrane. Each spike's compartment is continuous with the nucleus, and all spikes are packed with microtubules surrounded by sheets of membrane which appear pentalaminar. Ionophore A23187 stains the spikes and part of the collar as indicated by bright blue
Fta. 2. Longitudinal section through a mature H. americanus sperm showing inner acrosomal material, outer acrosomal material, apical cap, subacrosomal material, nucleus, and spikes. The acrosomal cap is indented anteriorly. Small wedge-shaped projections radiate inward from the periphery of the acrosome. Insets: (a), darkfield micrograph of an unfixed sperm showing the acrosome, subacrosomal material, nucleus, and spikes; (b), dark-field micrograph of a fixed sperm. Notice that after formaldehyde fixation the inner acrosomal material is visible. Insets: (a), × 2000; (b), x 10 000. Abbreviations used in this and the following figures: a, acrosome; ac, apical cap: am, acrosomal membrane; c, collar: ce, centriole; fp, finger-like projection; Jam, inner acrosomal material: m, mitochondrion: mmc, micro tubule /membrane complex; mt, microtubules; n, nucleus; ne, nuclear cuff; (~am. outer acrosomal material; pro, plasma membrane; s, spikes; sm, subacrosomal material.

280 TALBOT AND
fluorescence in these regions (Figs. 13-15). We presumed this to be due to insertion of A23187 into membranes of the spike and collar region. Similar staining has been ob- served in hamster sperm midpieces which contain an abundance of membranes due to the presence of mitochondria (Talbot, un- published data). The ionophore "stain" was especially useful in elucidating the struc- ture of the spike microtubule/membrane complex (Fig. 15). The microtubule/mem- brane complex in each spike bifurcates forming two branches in the vicinity of the collar. Each branch then joins with those from the adjacent spikes forming a three- sided vault of microtubule/membrane com- plex in the collar (Figs. 15 and 17). These results were confirmed in thin sections (Fig. 16). The detailed structure of this vault as
CHANMANON
reconstructed from ionophore-treated and thin-sectioned sperm is shown schemat- ically in Fig. 17.
DISCUSSION
H. americanus sperm follow the basic reptantian (lobsters, crayfish, crabs) sperm pattern (see 44), and morphologically closely resemble sperm from Nephrops (13) and H. vulgaris (40), the European lobsters. In particular, the arrangement of the acro- some, collar, nucleus, and spikes are similar in H. americanus, H. vulgaris, and Ne- phrops. It is interesting to note that while sperm of Panulirus, the spiny lobster, also follow the basic reptantian pattern, their acrosomes are small and lens-shaped and more than three spikes project from the nucleus (44).
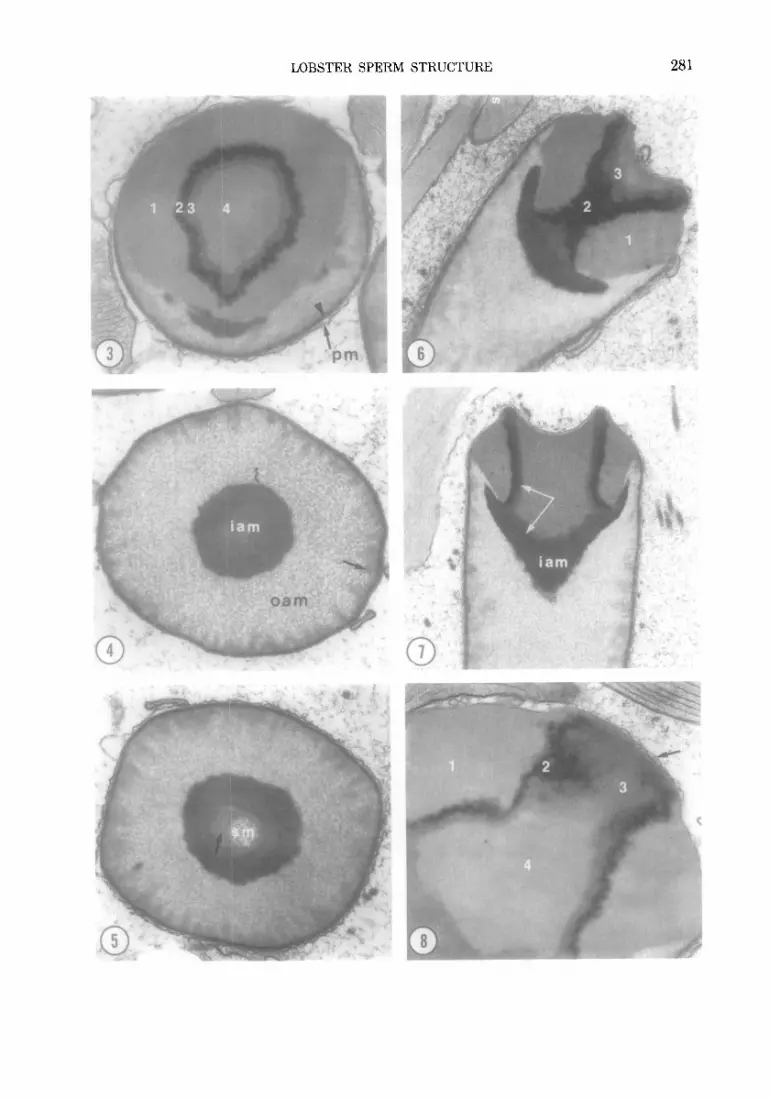
Fins. 3-5. Cross-sections through the acrosome at three levels (see Fig. 1 for approximate level of section). Figure 3 shows zones 1, 2, 3, and 4 of the apical cap. The plasma membrane surrounds the entire cell; the acrosomal membrane is indicated by an arrow. In Fig. 4 the section passes through the middle region of the acrosome. The inner acrosomal material is completely condensed and wedge-shaped projections (arrow) are evident around the periphery of the acrosomal vesicle. Figure 5 shows the base of the acrosome. In this sperm, the inner acrosomal material was not completely condensed (arrow) and is less electron dense. A portion of the subacrosomal material is shown. Fig. 3, x 23 300; Fig. 4, X 18 000; Fig. 5, × 20 000.
Fins. 6-8. Longitudinal-to-oblique sections through the acrosome's apical cap. The four zones of the apical cap are shown. Zones 1-3 appear crystalline, but zone 4 does not. The inner acrosomal material is continuous with zone 3 (see arrows in Fig. 7). Fig. 6, x 17 100; Fig. 7, x 13 800; Fig. 8, x 27 600.
Fins. 9-10. Longitudinal sections through the base of the acrosome, collar, and nucleus. The subacrosomal material and mitochondria are continuous with the chromatin. Part of the subacrosomal material extends around the basolateral surface of the acrosome (white arrows). The chromatin forms a cuff around the base of the acrosome (large arrow). The difference in thickness of the membrane limiting the nucleus and the plasma membrane around the acrosome can be compared in Fig. 9. The acrosomal membrane lining the fossa can be resolved in Figs. 8 and 9 (small black arrows). Regions of microtubule/membrane complex are evident in the collar. Figure 9 also shows the apical cap of an adjacent sperm; small finger-like projections extend between the acrosomal and plasma membrane Fig. 9, x 17 751; Fig. 10, x 29 100.
Fro. 11. Section through nucleus and microtubule/membrane complex (arrow) showing the complex, folded nature of the membrane over the nuclear region, x 12 319.
Fro. 12. Cross-section through a spike showing microtubules and sheaths of the membrane (arrow) wrapped around tubules.
FIGs. 13-15. Formaldehyde-fixed sperm treated with ionophore A23187 then examined with a fluorescent microscope. Intense fluorescence is seen in the spikes and collar. The nuclear region in Fig. 13 is also fluorescent, probably because of the large amount of folded membrane in this region. The bifurcation of the microtubule/ membrane complex of the spike can be seen in Fig. 15. x 1940.
FIG. 16. Electron micrograph through the collar showing spike-to-spike articulation. Each spike bifurcates as it approaches the collar and joins a branch coming from adjacent spikes. This results in formation of a triangle of microtubule/membrane complex in the collar. The points of this triangle are bent downward and the entire structure forms a three-sided vault, x 26 675.
a~
m~

bo
o0
b~
o H
>
>.
2~
o ~Z

©
E~
H
H
C

284 TALBOT AND CHANMANON
)[j L FIG. 17. Schematic diagram showing the three-
sided vault formed by the joining of the microtubule/ membrane complexes in the collar. Mitochrondria and centrioles are present in the central triangle of the vault apex. Portions of the subacrosomal material are continuous with the microtubule/membrane complex.
The acrosome in Homarus and Nephrops is an elongated cylindrical extension off the anterior end of the sperm; and in bo th H. americanus and Nephrops (10), the outer acrosomal material is strongly PAS posi- tive. The acrosome of H. americanus, like tha t of Panulirus (44), is limited by a single continuous membrane. In contrast, the ac- rosome of H. vulgaris is limited by two membranes (40), and a continuous tripar- t i t e membrane which is occasionally dou- bled has been repor ted in Nephrops (13). Brown has repor ted the existence of two membranes around the acrosome of the crabs, Callinectes (6). In general, sperm acrosomes are limited by a single mem- brane which is thought to be impor tant in segregating acrosomal enzymes within the sperm. The significance of the double or partial double membrane systems observed around some decapod acrosomes is not yet known, but their existence is a fur ther dis- t inction between the typical flagellated and atypical decapod sperm.
The chromat in of H. americanus sperm
is slightly more homogeneous and electron dense than tha t in other decapods (e.g., Panulirus); nevertheless it is clearly uncon- densed when compared to the chromatin of flagellated sperm, such as those of the ham- ster (47). The membrane surrounding the nuclear region of spikes of H. americanus appears to be a composite of the plasma membrane fused to elements of the nuclear envelope. A similar situation exists in other decapods (6, 9, 28, 37); however in some natant ians (29), vestiges of the nuclear en- velope are completely absent. In Homarus, as in many other decapods (e.g., 6, 28, 39), the nuclear envelope degenerates at the interface between the nucleus and subacro- somal region. While this may facilitate in- corporat ion of the sperm nucleus into the egg at fertilization (29), the absence of the envelope at this region is not a constant feature among decapods (22, 44).
Pochon-Masson (40) has stated tha t the spikes of H. vulgaris become directed to- ward the oocyte during sperm a t tachment to the chorion. Although we have in unpub- lished work observed spike flexure forward toward the acrosome during harsh treat- ments (osmotic shock), bending of the spikes was not normally observed in H. americanus except during the acrosome re- action (see following paper) when they flexed away from the acrosome and became aligned parallel to each other. Observations of numerous sperm suspended in SW have lead us to the conclusion tha t these ap- pendages are s tat ionary (except during the acrosome reaction), and tha t only Brown- ian movement is observed in a SW suspen- sion of nonreact ing H. americanus sperm.
Th e microtubules of the spikes are prob- ably cytoskeletal and are maintaining spike structure. Like the microtubules of Panu- lirus sperm (44), these are not sensitive to colchicine. The complete significance of the mic ro tubu le /membrane complex of Hom- arus and other decapod sperm is not clear from the present li terature. Quite likely an understanding of the function of these or-

LOBSTER SPERM STRUCTURE 285
ganelles will come after the details of ferti- lization mechanisms in Homarus have been clarified.
A new finding of this study is that the ionophore A23187 can be used to stain membranes for observation with a fluores- cent microscope. This permitted a useful and rapid overview of the distribution of the microtubule/membrane complex in Homarus sperm, and helped clarify the manner of spike-to-spike articulation. It should have applications in other cell sys- terns having localized regions of abundant membrane.
In 1895, Herrick (19) made the following statement regarding Homarus sperm: "nothing is known. . .about how the sper- matozoa reach the eggs and fertilize them." The mechanisms of fertilization in this un- usual group remain almost as puzzling to- day as they were 84 years ago. Our study provides the first ultrastructural analysis of sperm from H. americanus as a prelude to future investigations into the nature of lob- ster reproduction. New information on ac- rosome structure, spike articulation, and membrane organization have been pre- sented, which will provide an important basis for understanding the changes in sperm structure associated with acrosome reactions and fertilization.
We extend our thanks to Mr. Richard Sk. Chacon who assisted in obtaining Fig. 16 and to Marcia Kooda for helping prepare Fig. 17. Supported by NOAA Grants R/NP-1-7N and R/A-29 and an intercampus travel grant from the University of California.
REFERENCES
1. ANDERSON, W. A., AND ELLIS, R. A. (1967) Z. Zellforsch. Mikrosk. Anat. 77, 80.
2. ANDREWS, E. A. (1904) Anat. Anz. 25, 456. 3. BINFORD, R. (1913) J. MoTphol. 24, 147. 4. BOWEN, R. H. (1925) Anat. Rec. 31, 201. 5. BROWN, A., KLEVE, M. G., AND CLARK, W. H.
(1976) Amer. Zool. 16, 180. 6. BROWN, G. G. (1966) J. Ultrastruct. Res. 14, 425. 7. BROWN, G. G. (1967) in METZ, C. B., AND MON-
ROY, A. (Eds.), Fertilization, Academic Press, New York.
8. CHEVAILLIER, P. (1965) C.R.H. Acad. Sci. 262, 3225.
9. CHEVAILLIER, P. (1966) C.R.H. Acad. Sci. 262, 1546.
10. CHEVAILLIER, P. (1966) J. Microsc. 5, 739. 11. CHEVAILLIER, P. (1967) J. Microsc. 6, 853. 12. CHEVAILLIER, P. (1968) J. Microsc. 7, 107. 13. CHEVAILLIER, P., AND MAILLET, P. L. (1965) J.
Microsc. 4, 679. 14. FASTEN, N. (1914) J. Morphol. 25, 587. 15. FASTEN, N. (1918) Biol. Bull. 34, 277. 16. FASTEN, N. (1921) Biol. Bull. 41, 288. 17. FASTEN, N. (1924) J. Morphol. Physiol. 39, 47. 18. FASTEN, N. (1926) Biol. Bull. 50, 277. 19. HERRICK, F. H. (1895) Bull. U.S. Fish. Comm. 1-
252. 20. HINSCH, G. W. (1969) J. Ultrastruct Res. 29, 525. 21. HINSCH, G. W. (1971) J. Ultrastruct. Res. 39, 86. 22. HINSCH, G. W. (1973) Canad. J. Zool. 51, 421. 23. HUDINAGA, N. (1942) Japan. J. Zool. 10, 305. 24. KAYE, G. I., PAPPAS, G. D., YASUZUMI, G., AND
YAMAMOTO, H. (1961) Z. Zellforsch. Mikrosk. Anat. 53.
25. KING, J. E. (1948) Biol. Bull. 94, 244. 26. KOLTZOFF, N. K. (1906) Arch. Mikrosk. Anat. 67,
364. 27. LABBE, A. (1903) C.R.H. Acad. Sci. 137,272. 28. LANGRETII, S. G. (1969) J. Cell. Biol. 43, 575. 29. Lu, C. S. (1976) Dissertation, University of Hous-
ton. 30. McCROAN, J. E. (1940) Cytologia 11, 136. 31. MosEs, M. J. {1956) J. Biophys. Biochem. Cytol.
2, 215. 32. MOSES, M. J. (1961) J. Biophys. Biochem. Cytol.
9, 222. 33. MOSES, M. J. (1961) J. Biophys. Biochem. Cytol.
10, 301. 34. NATH, V. (1933) Quart. J. Microsc. Soc. 75, 543. 35. POCHON-MASSON, J. (1963) C.R.H. Acad. Sci. 256,
2226. 36. POCHON-MASSON, J. (1965) C.R.H. Acad. Sci. 260,
3762. 37. POCHON-MASSON, J. (1965) C.R.H. Acad. Sci. 260,
5093. 38. POCHON-MASSON, J. (1965) C.R.H. Acad. Sci. 260,
5352. 39. POCHON-MASSON, J. (1968) Ann. Sci. Nat. Zool.
Paris 10, 1. 40. POCttON-MASSON, J. (1968) Ann. Sci. Nat. Zool.
Paris 10, 369. 41. PRESSMAN, B. C., HARRIS, E. J., JAGGER, W. S.,
AND JOHNSON, J. H. (1967) Proc. Nat. Acad. Sci. USA 58, 1949.
42. RUTHMANN, A. (1958) J. Biophys. Biochem. Cytol. 4, 267.
43. SPURR, A. R. {1969) J. Ultrastruct. Res. 26, 31. 44. TALBOT, P., AND SUMMERS, R. G. (1978) J. Ultra-
struct. Res. 64, 341.

286 TALBOT AND CHANMANON
45. TERNI, T. (1937) Biol. Bull. 73, 394. 46. WORLEY, E. K. (1939) Cellule 48, 22. 47. YANAGIMACHI, R., AND NODA, Y. D. (1970) Amer.
J. Anat. 128, 367.
48. YAsuzumI, G. (1960) J. Biophys. Biochem. Cytol. 7, 73.
49. YASUZUMI, G., KAYE, G. I., PAPPAS, G. D., YA- MAMOTO, H., AND TSUBO, I. (1961) Z. Zellforsch. Mikrosk. Anat. 53, 141.





























