Embed Size (px)
Citation preview

The reproductive ecology of Medicago citrina (Font Quer) Greuter
(Leguminosae): a bee-pollinated plant in Mediterranean islands
where bees are absent
C. Perez-Banon1, A. Juan
1, T. Petanidou
2, M
aA. Marcos-Garcıa
1, and M. B. Crespo
1
1Centro Iberoamericano de la Biodiversidad, Universidad de Alicante, Alicante, Spain2Department of Geography, University of the Aegean, Building of Geography, University Hill, Mytilene,Greece
Received November 18, 2002; accepted January 16, 2003Published online: October 16, 2003� Springer-Verlag 2003
Abstract. The reproductive ecology of Medicagocitrina was studied in the Columbretes archipel-ago (Mediterranean Sea), where bees are absent.The flowers were self-compatible, homogamous,nectar producers, depended on large floral visi-tors for tripping and pollination. Several charac-teristics of the reproductive biology of M. citrinaindicate an adaptation to an isolated environmentcharacterised by scarcity of appropriate pollina-tors. As a result of this scarcity, fruit and seedset under natural conditions were significantlylower compared to hand-pollination treatments.The main flower visitors were mostly flies andblowflies, with Eristalis tenax (Syrphidae) andCalliphora vicina (Calliphoridae) being the mostefficient pollinators. We argue that because M.citrina faces pollen limitation in the small islets ofColumbretes, it presents an interesting case studyof pollination related to island endemism anddiversity.
Key words: Blowflies, endemism, hoverflies, iso-lation, Mediterranean islands, migratory pollin-ators, nectar secretion, reproductive ecology,tripping.
Introduction
The Mediterranean Region probably has moreisolated plant populations than most otherregions. Heterogeneous geographical barriers(e.g. high mountains, many small islands andarchipelagos), extreme climates and intensiveland use by man over millennia, have sculp-tured a diverse patchwork of ecosystems. Onsmall Mediterranean islands, pioneer popula-tions may originate from a few founderindividuals and develop under conditions ofcomplete geographical and genetic isolation(Blondel and Aronson 1999). Such islandsprovide unique natural laboratories for study-ing the process of evolution (Carlquist 1974).However, many islands are difficult to accessand, as a consequence, the reproductive biol-ogies of their plants and animals are poorlystudied (Anderson et al. 2001).
Geographical isolation of plants on islandsis often associated with pollen limitation ofseed and fruit set due to small pollinating
Plant Syst. Evol. 241: 29–46 (2003)DOI 10.1007/s00606-003-0004-3

faunas or lack of adequate pollinators (McCul-len 1990, Howarth and Mull 1992, Andersonet al. 2001), as well as with Allee-effects thatcan occur in small plant populations (Petani-dou et al. 1991, 1995a, b; Oostermeijer et al.1994, 2000). Pollinator scarcity and reducedpollinator services are often associated withhigh selfing-rates (Ramsey and Vaughton1996, Fausto et al. 200l, Anderson et al.2001, references cited therein), or a highfrequency of anemophily (Berry and Calvo1989, Anderson et al. 2001). On the otherhand, small and isolated populations of pre-dominantly outbreeding, self-compatible spe-cies may lose genetic variability by genetic driftand inbreeding (Barrett and Kohn 1991, Raij-mann et al. 1994, Fischer and Matthies 1998,Menges and Dolan 1998, Young et al. 1999).This ‘‘genetic erosion’’, when not counteractedby gene flow may result in reduced fitness,which may lead to local extinction even if theecological conditions are relatively suitable forthe species (Ouborg et al. 1991; Treuren et al.1991; Ouborg 1993; Oostermeijer et al. 1992,1994, 1998; Groom 2001).
We examine the pollination ecology ofMedicago citrina, an endangered species thatis usually dependent upon bees for pollination,but is also found in areas where bees arelacking (Lesins and Lesins 1979). One sucharea is the volcanic archipelago of Columbre-tes, located c. 55 km east of the coast ofCastellon (eastern Spain) where no permanentbee population exists (Espanol 1958, GarcıaMarı et al. 1991). Our aim is to describe thepollination ecology of M. citrina within thisinsular community, and identify possible adap-tations to the absence of bees. Specifically,we (1) examine the reproductive ecology ofM. citrina under natural conditions in Colum-bretes; (2) measure the pollination effectivenessof non-bee pollinators; and, (3) compare nectarrewards and reproductive characteristics
(pollen viability, floral fecundity, selfing rate,etc.) of these island populations with theirmainland con-specifics. We discuss our findingsin the context of in situ species management andmaintenance of biodiversity in isolated environ-ments. The results presented here are part of amajor study on the reproduction ecology ofplants in insular ecosystems, with the aim toexplore possible viability constraints associatedwith their isolation and to pinpoint the vulner-ability and risks related to their conservation(Perez-Banon 2000, Juan 2002).
Material and methods
The study species and its distribution
Medicago citrina (Font Quer) Greuter [�M. arbo-rea var. citrina Font Quer; �M. arborea subsp.citrina (Font Quer) O. Bolos & Vigo], is a perennialwoody leguminous shrub which grows up to 2 m inheight.
Flowers have a typical papilionoid leguminousstructure and are born in axillary racemes of 8–17flowers, with hundreds of racemes on each branch.Nectaries are located at the bottom of the corollatube. M. citrina, like many papilionoid leguminousflowers is dependent on a process known as‘‘tripping’’ for its pollination (Free 1993). Atanthesis, the style and the staminal column remainenclosed in the keel until pressed down by theweight of a specialised visiting insect (normally amedium- to large-sized bee) so that the style andstaminal column quickly move out of the keel, andthe flower is tripped.
The species is known only from a few Medi-terranean islands of the Spanish coast, the Colum-bretes archipelago, some islets around Ibiza (Juanet al. 1999), around Cabrera (Bibiloni et al. 1993)within the Balearic Islands, and the small isletEscull del Cap near Alicante (Serra et al. 2001)(Fig. 1).
In the Columbretes archipelago,M. citrina onlygrows on three small islets: Grossa, Ferrera andForadada. The establishment dates from after thevolcanic events that gave rise to the archipelago,
Fig. 1. Location of the four study populations (dots) of M. citrina. 1. Marine Reserve of Columbretesarchipelago. 2. Escull del Cap (Alicante). 3. Ibiza. 4. National Park of Cabrera. Subpopulations are markedwith either solid squares or arrow heads
c
30 C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands

C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands 31

between 1 and 0.3 mya (Aparicio et al. 1991). Mostlikely, these populations originated from theBalearic Islands, following bird dispersal (Juan,unpublished data). Its limited distribution (fewhundreds of plants on a few tiny islands) andvulnerability of habitats have lead to M. citrinabeing listed as an endangered plant in Spain(Spanish Royal Decree 439/1990) and assigned asendangered by IUCN (Aizpuru et al. 2000).
Nomenclature of plant taxa follows Greuteret al. (1989) and Mateo and Crespo (2001).
Study site
The fieldwork was carried out from February toJune in 1997 and 1998, in the Marine Reserve ofColumbretes archipelago which has prevailingMediterranean climate (mean annual temperature18.6 �C, mean annual rainfall 315 mm). All exper-iments and observations were carried out during theflowering peak of the species (late February–April).
Medicago citrina is the dominant woody spe-cies in Foradada and Ferrera (Fig. 1), locatedmainly on the northern slopes. On Grossa, thereare approximately 100 individuals introduced byman from the nearby Ferrera and Foradada,after the native population went extinct followingovergrazing (rabbits) and fires. The pollinationtreatments, experiments and observations wereconducted mainly on the island Grossa. Systematicfieldwork on the large native populations ofForadada and Ferrera was prevented by the strongwinds prevailing in the archipelago during mostdays of the flowering period.
The dominant vegetation of Grossa island iscomposed of Suaeda vera Forssk., Lavatera sp., andLobularia maritima (L.) Desv. subsp. columbretensisR. Fern. (endemic to the archipelago), togetherwith herb communities dominated by Euphorbiaterracina L. and Medicago littoralis Rohde.
Flowering characteristics
Flowering phenology of M. citrina was recordedin the Grossa population from February to June1997. We counted the number of inflorescencesbearing open flowers on randomly selected 4permanent branches of 15 plants throughout theseason. Counting was carried out every 3–5 daysfrom the end of February to April. At other timescounts were carried out at least once every fifteen
days, when weather and sea conditions allowedvisits to the archipelago.
Flower life span (= duration of anthesis) wasstudied on flowers selected at random and locatedat different positions on two plants (N¼ 31; N¼ 32,respectively). The flowers were marked at the budstage and observed daily until they withered.Commencement of flower anthesis was defined bythe opening of floral petals, when insect access tothe flower was made possible.
Nectar secretion
Flowers were selected at random at differentpositions of the plant/branch. They were markedat the bud stage during the blooming peak and eachbranch was covered with a metal cage (70 cmlength, 15 cm diameter) with fine gauze to preventinsect access. Nectar secretion rates were measuredby emptying the whole quantity of nectar accumu-lated at the bottom of the flower tube with 2-l1 to10-ll calibrated ‘‘Drummond’’ microcapillarytubes. Nectar concentration (expressed in % w/wsucrose) was measured immediately after nectarretrieval for each flower with a ‘‘Bellingham andStanley’’ pocket refractometer. Sugar content (inmg of equivalent sucrose) was calculated as:volume · (concentration/100) · nectar density(Dafni 1992).
Within day nectar secretion. Daily nectarsecretion rate was studied on lst-day flowers(N¼ 47) on several caged branches. Freshlyopened flowers were emptied of their nectar at10:00 h and sampled thereafter at c. 4-h intervals(14:00 and 17:00–18:00 h, hereafter called ‘‘noon’’and ‘‘evening’’ samples, respectively). Emptyingthe flowers earlier than 10:00 h was not possiblebecause dew covered the flowers every morningand diluted the nectar.
Nectar secretion in relation to flower age. Toassess the nectar secretion pattern during floweranthesis, we sampled the nectar from the first dayof opening until withering (N¼ 121, 64, 44, 35, 14,15, 20 flowers for 1st to 7th day -the last possibleday of flower anthesis-, respectively). Sampling wascarried out on the caged flowers once per day,between 10:00 and 11:00 h.
Pollen viability
The effect of flower age on pollen viability wasexamined using flowers enclosed at the bud stage in
32 C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands

metal cages covered with fine gauze (N¼ 42). Overthe following seven days, all opening flowers fromthree plants were marked each day with a differentcolour of waterproof paint. On the seventh daythree flowers of each age were collected on separatePetri dishes. Flowers were artificially tripped byexerting a slight pressure on the keel, and theexposed pollen was checked for viability. Forty-twosamples were tested (7 days · 3 flowers · 2 samplesper flower).
To assess the effect of pollen exposure on itsviability, lst-day flowers from three plants werecollected in a Petri dish (N¼ 21), and then artifi-cially tripped. Pollen was sampled from anthersfrom three flowers daily for the next seven days (atc. 10:00 h) and tested for viability.
Pollen viability was assessed using a germina-bility test. Preliminary tests with different sucrosesolutions, showed that a 30% solution was themost appropriate for the germination of M. citrinapollen; this solution was used for the preparation ofBrewbaker-Kwack medium. Under a dissectingmicroscope, each pollen sample (c. 500 grains)was mixed well with a droplet of Brewbaker-Kwackmedium (Kearns and Inouye 1993). The droplets(= samples) were left hanging from the lid of aPetri dish at room temperature and after 48 h,germination was stopped by freezing (Dafni 1992).Each sample was transferred onto a slide, perma-nently mounted and examined under a microscope.Only the pollen grains that germinated successfullyand produced tubes were considered viable. Pollenviability was expressed as the percentage of germi-nating pollen grains.
Reproductive success after different pollinationtreatments
We conducted the following pollination treatments:spontaneous self-pollination, manual self- andcross-pollination, and simple tripping withoutfurther manual pollination. Insects were excludedfrom flowers with metal cages (70 cm length, 15 cmdiameter) covered with fine gauze, which enclosedwhole branches. The cages remained closed afterthe pollination treatments were applied until thebeginning of fruit set. Two sets of treatments werecarried out, one at the beginning (late February)and the other at the end (mid April) of the 1998flowering peak.
The results from only 5 plants were used, out ofthe 15 originally enclosed, because the cages on theremaining 10 were destroyed by bad weather. Allflowers used were selected at random at differentpositions of the plants and branches.
Spontaneous self-pollination: The flowers andinflorescences at the bud stage were colour-markedand enclosed in metal cages to be left undisturbeduntil fruit harvest (N¼ 234).
Hand self-pollination: Flower buds were cagedand marked, and then tripped and self-pollinatedby hand on the first day they opened (N¼ 100).Self-pollination was carried out by removinganthers of other 1st-day flowers of the same plantwith a pair of fine forceps and rubbing them gentlyover the stigma of the 1st-day flowers to be treated.
Hand cross-pollination: A total of 107 flowerbuds were covered and marked as above. The1st-day flowers were tripped and cross-pollinatedmanually by rubbing newly opened floral anthersof 1st-day flowers of neighbouring plants, over thestigmas of the study plant.
Hand-pollination of emasculated flowers: Thesetreatments allowed us to reduce the risk ofunwanted self pollen transfer and self-pollinationduring the hand-pollinations described above, andhence check for existing differences between handself- and cross-pollinations. The treated flowerswere carefully emasculated at the bud stage, whencorolla length was equal to calyx length. All thebranches were covered as described above. Emas-culated flowers were either self-pollinated withfresh pollen from flowers of the same plant(N¼ 64), or cross-pollinated with fresh pollen fromneighbouring plants (N¼ 49).
Pollination after tripping: A total of 173 flowerbuds were bagged, marked and tripped during theirfirst day of anthesis. The tripped flowers were leftuntouched until harvest inside metal cages coveringthe whole branches.
To determine whether tripping was essentialfor pollination (and subsequent fruit set), all theflowers used to measure flower life span (N¼ 63)were followed until fruit set. We assessed thepercentage of fruit set for flowers that had beentripped naturally.
Tripping rate under natural conditions wasestimated using 351 flowers marked at the budstage and left untouched. The number of flowerstripped naturally by any means, e.g. flower visitors,
C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands 33

wind or high temperatures (Free 1993), wascounted at the stage of withering.
Natural pollination: Natural pollination successwas measured using flowers (N¼ 2735) marked atthe bud stage from 4 plants and left uncaged to befreely visited by insects.
Reproductive success and flower age
A second set of experimental treatments to inves-tigate fecundity in relation to flower age wascarried out at the beginning and the end of theflowering season, using 152 and 237 flowers,respectively. The flowers were caged at the budstage, and were marked as soon as they opened,which resulted in a continuum of flowers that hadbeen open for different lengths at time (1–7 days).On the seventh day all marked flowers wereartificially tripped and self-pollinated by hand.
In all the above treatments, fruits were har-vested when fully ripe approximately 2–3 monthsafter pollination treatments. After harvesting, seedswere counted and were of two types: badly formed(shrivelled or empty) or full, normal seeds. The firstcategory was considered unfertilised and/oraborted, and the second as viable. The reproductivesuccess of these flowers was estimated as fruit set(percentage of fruits over the total number offlowers marked), and seed set (number of seeds/fruit).
Flower visitors
Observations of flower visitors were carried outduring both years of the study. Systematic surveys(insect visitor counts) were performed in 1997 bytwo observers recording the number of insectsvisiting the flowers of three mature plants on a2 · 2 m permanent plot for 15-min periods. Thecounts were conducted twice per day, during themorning (between 9:00 and 11:00 h) and the earlyafternoon (14:30–16:00 h) for 22 days during theflowering peak (27 February–22 March). Thepollination efficiency of flower visitors was assessedby measuring both pollen transfer by differentinsect species and fruit and seed set resulting fromtheir visits.
Pollen loads on insects: Pollen (con- andheterospecific) adhering to the insect body of themain flower visiting species was collected andexamined. The loads were sampled from insects
captured after they had just visited a flower ofM. citrina in the vicinity of the study plot duringpeak flowering. Immediately after capture, theinsects were killed and the pollen from all bodyparts was removed with a sticky piece of fuchsin gel(Beattie 1972). The pollen was counted under alight microscope and identified by using referencematerial from the field. Microphotographs of nonexamined insects, taken with a scanning electronmicroscope (SEM; JEOL 840, at 20 kV), illustratedwhere M. citrina pollen adhered on the insect.
To check for the dependence of insects onM. citrina pollen, a series of gut content analyseswas carried out. After pollen removal from theexternal body, the insects were dissected under amicroscope and pollen from different parts of thegut was collected (Gilbert 1993). The samples weremounted and stained with a sticky piece of fuchsingel (Beattie 1972) and the pollen grains identifiedand counted as above. The number of individualsexamined varied among species (N¼ 15–20). Weexamined the hoverflies Eristalis tenax (Linnaeus1758) and Eupeodes corollae (Fabricius 1794), andthe blowflies Calliphora vicina Robineau-Desvoidy,1830 and Lucilia sericata (Meigen 1826). The totalnumber of pollen grains counted per sample variedbetween 200–9000.
Fruit and seed set resulting from floral visits bydifferent insects: Assuming that caged pollinatorsmay not show the same levels of efficiency as thosein natural conditions and may act atypically aseither making few visits because they are disturbedor visiting flowers by necessity, we carried out thefollowing experiment in order to assess the polli-nation efficiency of the flower visitors. Insects wereenclosed in metal cages, like the ones describedpreviously. When in anthesis, 1–2 insects ofEristalis tenax, Eupeodes corollae, Calliphora vicinaor Lucilia sericata were kept in each cage until allflowers withered (number of flowers caged in 1997:NE. tenax¼ 125, NE. corollae¼ 52, NC. vicina¼ 219,NL. sericata¼ 132; in 1998: NE. tenax¼ 165, NE. corol-
lae¼ 97, NC. vicina¼ 119, NL. sericata¼ 103). Consider-ing that the variability in flower numbers in thecages may bias the pollination efficiency measure-ments, we decided to consider not only the fruit/seed set, but also the tripping rates caused by therespective insects. These results would be interest-ing only in the case that fruit/seed set in all cageswere at the same time non proportional to flowernumbers and tripping rates. As, indeed, this
34 C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands

assumption was met (they were not proportional:see Results section and Fig. 10), the efficiencyresults were considered. When necessary, deadinsects were replaced with living insects. Pollinationeffectiveness was measured as percentage of fruitset in 1997 and 1998 and as seed set in 1998.
The transient character of some flower-visitinginsects in the Columbretes archipelago was indi-cated by their irregular presence relative to othermore constant floral visitors. This was assessed byregular counts of Eristalis tenax and Calliphoravicina visiting the flowers of the dominant plantsM. citrina and Daucus gingidium L. on Grossaisland (late February–mid April and May–midJune 1997, respectively). The counts were madeby two observers on a daily basis from 10:00 to11:00 h by observing insects visiting the flowers ofeither species in a randomised transect.
Data analysis
Prior to statistical analysis, the data were tested fornormality using Kolmogorov-Smirnov one-sampletests. In several cases the data were not normallydistributed, and therefore we applied non-paramet-ric tests (Sokal and Rohlf 1981). Variation inreproductive success with pollination treatmentswas tested by Kruskal–Wallis ANOVA (K–S) fol-lowed by separate Mann–Whitney U-tests (M–W)after application of the Bonferroni correction.Percentages obtained by pollen germination treat-ments for tripped and non-tripped flowers wereevaluated using G-tests. Throughout the paper, themeans and standard errors are given.
Results
Flowering phenology and anthesischaracteristics
The flowering period of M. citrina was long,lasting from the end of January until the end ofMay, with a peak from mid February to midApril. The flowers are homogamous and theanthers ripened about 2 days before anthesis,when the corolla was about twice as long asthe calyx. Flower life span of non-trippedflowers was significantly longer than that oftripped ones (8.0 ± 0.2 days, N¼ 27 and5.7 ± 0.4 days, N¼ 25; M–W U[27,25]¼ 114.5
P < 0.000). Post-tripped flowers remainedattractive with a brilliant lemon yellow colourand turgent corolla for 1.9 ± 0.2 days(N¼ 15), after which they showed a loss offloral colour and turgor.
Nectar secretion
Daily pattern. Nectar of lst-day flowers wasmainly secreted in the morning (Fig. 2). Vol-ume followed the same pattern as sugarcontent. Morning volume and sugar contentwere significantly higher than in the afternoon(volume: 0.0 < U < 16.0, P < 0.0002; sugarcontent: 0.0 < U < 37.0, P < 0.0051).
Nectar secretion in relation to flower
age. Nectar was secreted throughout anthesis(Fig. 3). The highest nectar secretion wasrecorded during the first three days, with a peakon the second day, although it did not differsignificantly from the nectar secreted on the firstand the third day (M–WU tests withBonferronicorrections: P > 0.05). A moderate amountwas secreted during the fourth day, thereafterfollowed by a dramatic decrease. These valueswere significantly lower than those of the firstthree days (M–W U tests, P < 0.05).
Fig. 2. Daily nectar secretion pattern of 1st-dayflowers of M. citrina. Samples were taken three timesper day over three consecutive days (N22 March¼ 14;N23 March¼ 14; N24 March¼ 19)
C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands 35

Pollen viability
The overall pollen viability differencesbetween tripped and non-tripped flowerswere not significant (G¼ 9.627, P > 0.05;Fig. 4). However, viability of trippedflowers dropped dramatically after the secondday of anthesis, whereas that of the non-tripped flowers remained high until the fifthday.
Reproductive success of M. citrina
From February to April the flowers remainedfecund from the first to the sixth day ofanthesis, whereas no seeds were producedwhen flowers were pollinated from the seventhday onwards (Fig. 5). In both periodsfecundity, calculated as seed set, steadilydecreased from the second day onwards,showing a strong dependence on flower-age(K–W H[5, 152]¼ 20.3815, P < 0.0011; K–WH[6, 237]¼ 29.3643, P < 0.0001, for Februaryand April, respectively). In both cases, flowersthat had been tripped and pollinated duringthe first day of anthesis yielded the highest seedset, compared to those tripped and pollinatedlater. However, after the Bonferroni correctionwas applied most of these differences werefound to be non-significant. There was nodifference in floral fecundity between the twoperiods studied (P > 0.05).
There were no significant differences be-tween seed sets resulting after the treatments ofFebruary and April (M–W U tests; P > 0.05).Seed set after different pollination treatmentscarried out in April varied greatly (Fig. 6;K–WH[4, 1414]¼ 488.0, P < 0.000 for the totalnumber of seeds; K–W H[4, 1414]¼ 492.4,P < 0.000 for viable number of seeds). Man-ual self- and cross-pollination yielded a greater
Fig. 3. Nectar secretion pattern of virgin, non-tripped flowers of M. citrina. We present the sugarcontent secreted daily throughout the whole floweranthesis (N¼ 121, 64, 44, 35, 14, 15, 20 flowers for 1stto 7th day, respectively)
Fig. 4. Pollen viability expressed as per-centage of germinated pollen grainsthroughout the whole life span of non-tripped and tripped flowers of M. citrina(3 flowers and 6 samples for each day andcase). Standard errors are given. Pollen intripped flowers was continuously exposed,whereas it was protected in non-trippedflowers
36 C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands

number of seeds (total or viable) than any ofthe other treatments (M–W U tests withBonferroni corrections, P < 0.000). No sig-nificant difference in the number of seeds wasfound between free-pollinated and simplytripped flowers, i.e. without intentional pollentransfer (M–W U test, P > 0.05). Spontane-
ous self-pollination resulted in a much lowersuccess than free-pollination or simple tripping(P < 0.05). Percent fruit set showed the samepattern: 13.6% for naturally-pollinated, 10.4%for simply tripped, 72.5% for hand-selfed,70.1% for hand-crossed and 4.2% for sponta-neously-selfed flowers. It is noteworthy that
Fig. 5. Reproductive success ofM. citrina after synchronous trippingand self-pollination of flowers of dif-ferent ages in February and April. Wegive the mean number of seeds perfruit (columns), the sample numberand the standard error (bars)
Fig. 6. Reproductive success of M. citrina for all pollination treatments. Columns represent the mean numberof seeds per fruit (total¼ viable + aborted, and only viable). Percent seed set is given at the top of eachcolumn. Bars are standard error
C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands 37

fruit set after natural-pollination was only13.6%, equivalent to a small fraction of theflowers that were freely tripped under naturalconditions (45.3%). No fruit was producedamong the untripped flowers (N¼ 63).
There were no differences in seed set (totalor viable) between hand self- and cross-pollinated flowers (Fig. 6; M–W U tests,P > 0.05). Similar results were obtained forflowers that had been originally emasculated inorder to control for pollen source (P > 0.05).
Flower visitors and their efficiency
Our investigation of the existing pollinatorpool in the archipelago confirmed the absenceof permanent bee-colonies from Columbretes.Medicago citrina flowers were mainly visitedby flies, mostly Calliphoridae (89%) andSyrphidae (11%). Calliphora vicina was themost dominant visitor among blowflies(86.5%), whereas the frequency of Luciliasericata was much lower (2.5%). Six hoverflyspecies visited M. citrina, but visited flowersless frequently than C. vicina. Most frequentwere Eupeodes corollae (5.6%) and Eristalistenax (4.5%), whereas Episyrphus balteatus,Meliscaeva auricollis and Scaeva pyrastri wereobserved only occasionally on the flowers(0.9%). In fact, our long-term observationscarried out on a daily basis demonstrated thatsome of the existing hoverfly populations in
the archipelago are very irregular flower visi-tors. Such insects are E. tenax and otherhoverflies that may not appear to the flowersfor a couple of consecutive days (Fig. 7).Considering their anthophilous character, thisirregularity may imply that the populations ofthese insects are so variable in the archipelagothat it can be explained only by their transienthabit. All the observed species visited flowersmainly during the morning (9:00–11:00 h)when dew covering the flowers started toevaporate, and the volume and sugar contentof nectar was high (Figs. 2, 8). The rate ofvisits decreased dramatically early in theafternoon, when the flies shifted to flowers ofother species, mainly Lobularia maritima andEuphorbia terracina.
Among the other occasional visitors, themigratory butterfly Vanessa cardui (Linnaeus1758) and the migratory hawkmoth Macro-glossum stellatarum (Linnaeus 1758) visited theflowers for nectar, but very rarely tripped them.
Analysis of gut contents showed that cal-liphorids contained almost no pollen grains intheir guts. Syrphids, on the other hand, werenot great pollen consumers of M. citrina, andthe two main syrphid visitors, Eristalis tenaxand Eupeodes corollae, had only 2.6% and1.0%, respectively, of M. citrina pollen in theirguts. The syrphids were probably visitingM. citrina flowers mainly for nectar andL. maritima flowers for pollen as indicated by
Fig. 7. Daily counts of Cal-liphora vicina and Eristalistenax visiting the flowers ofM. citrina and Daucus gingi-dium on the island Grossa inthe Columbretes archipelagothroughout the floweringseason of 1997. Values arethe numbers of insects visit-ing flowers in a transectbetween 10:00–11:00h
38 C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands

the gut content analyses (E. tenax: 97.4%;E. corollae: 99.0%).
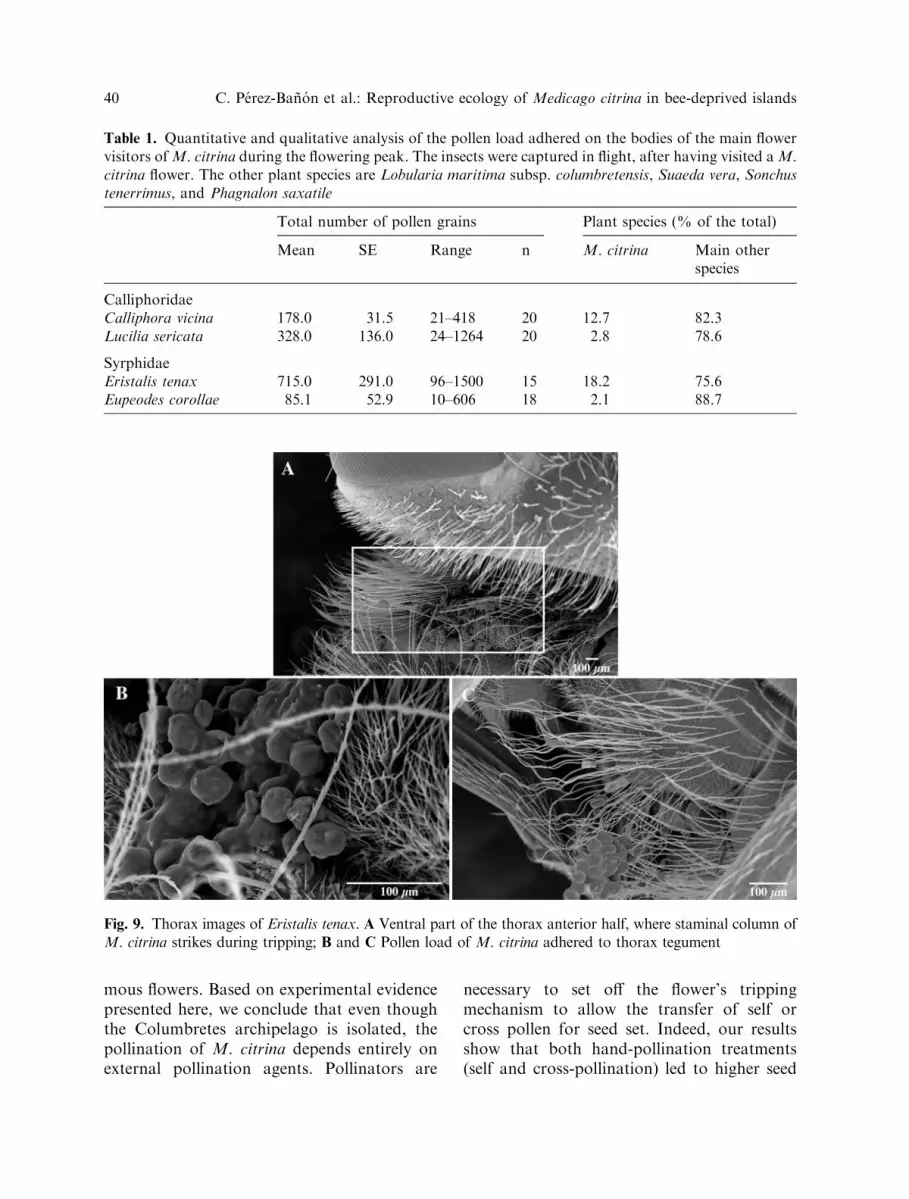
Examination of the pollen that adhered tothe bodies of the insects visiting Medicagoflowers, showed that E. tenax bore the highestabsolute number and percentage, followed bycalliphorids (Table 1). The microphotographsshow that the largest quantities of M. citrinapollen were located on the ventral part of thethorax of E. tenax (Fig. 9). This implies thatpollen transfer and deposition occurred sterno-tribically, as is the case for M. sativa (Smallet al. 1987).
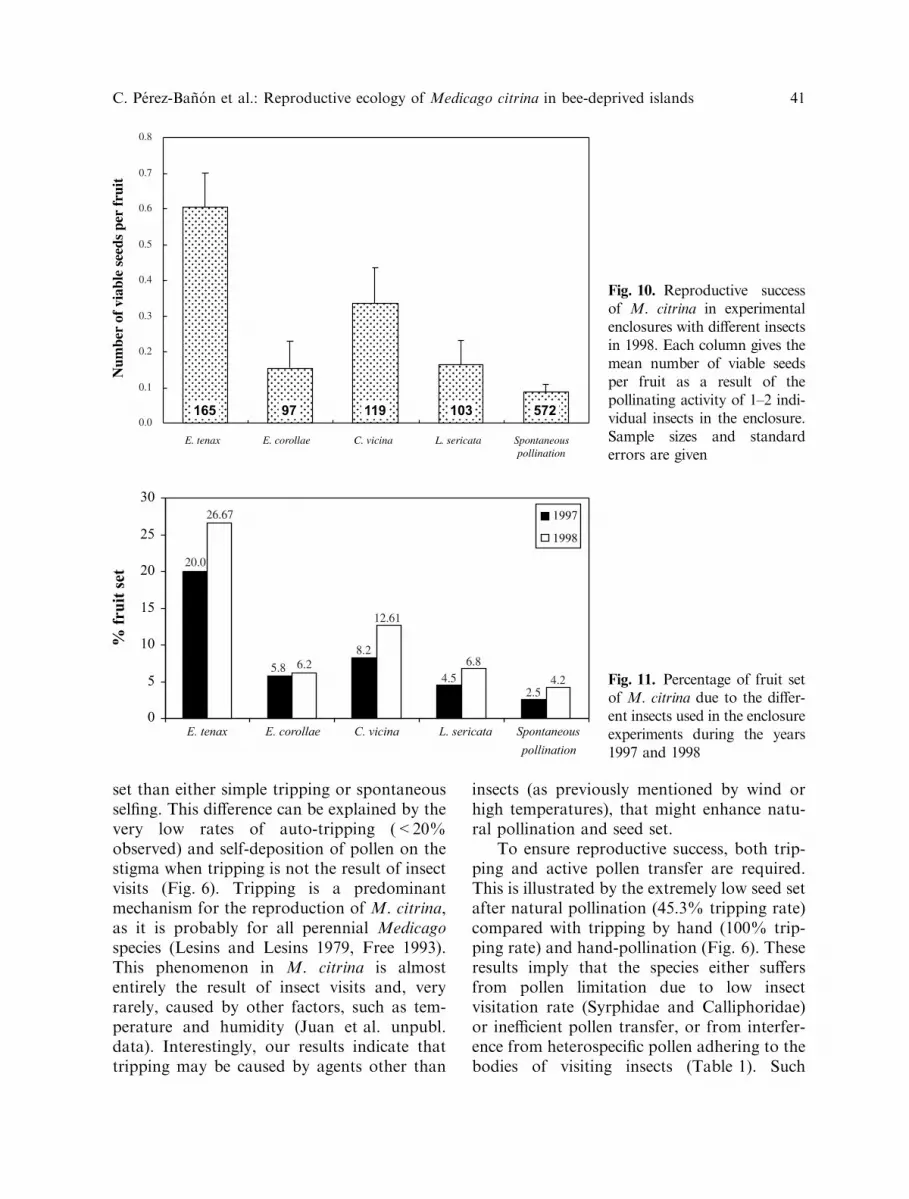
Enclosure experiments used to assess thepollination efficiency of flower visitors showedthat the highest seed sets were produced byE. tenax and C. vicina. E. tenax was moreefficient for setting seed than C. vicina (M–WU tests: P < 0.05). In all cases, seed set andseed production were significantly higher than
spontaneous pollination (Fig. 10; M-W U testsbased on the viable number of seeds:P < 0.001). Similar results were obtainedfrom fruit sets from enclosure experimentscarried out in 1997 and 1998 (Fig. 11): fruit setwas highest in the case of E. tenax, followed byC. vicina, E. corollae, and L. sericata. Thetripping rates affected by the insects inside theenclosures were as follows: E. tenax (66.4%and 55.8% for 1997 and 1998, respectively),C. vicina (50.2%; 54.6%), L. sericata (17.2%;13.6%), E. corollae (14.6%; 11.3%). Sponta-neous (automatic) tripping rate was 19.8% in1997 and 16.5% in 1998.
Discussion
Breeding system of M. citrina
Medicago citrina is a fully self-compatiblespecies bearing homogamous but non-autoga-
Fig. 8. Visitation rates of thetwo main insect groups recordedin the permanent plot during theflowering peak (27 February–22March 1997) in the morning(9:00–11:00) and early in theafternoon (14.00–16.00)
C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands 39
mous flowers. Based on experimental evidencepresented here, we conclude that even thoughthe Columbretes archipelago is isolated, thepollination of M. citrina depends entirely onexternal pollination agents. Pollinators are
necessary to set off the flower’s trippingmechanism to allow the transfer of self orcross pollen for seed set. Indeed, our resultsshow that both hand-pollination treatments(self and cross-pollination) led to higher seed
Table 1. Quantitative and qualitative analysis of the pollen load adhered on the bodies of the main flowervisitors ofM. citrina during the flowering peak. The insects were captured in flight, after having visited aM.citrina flower. The other plant species are Lobularia maritima subsp. columbretensis, Suaeda vera, Sonchustenerrimus, and Phagnalon saxatile
Total number of pollen grains Plant species (% of the total)
Mean SE Range n M. citrina Main otherspecies
CalliphoridaeCalliphora vicina 178.0 31.5 21–418 20 12.7 82.3Lucilia sericata 328.0 136.0 24–1264 20 2.8 78.6
SyrphidaeEristalis tenax 715.0 291.0 96–1500 15 18.2 75.6Eupeodes corollae 85.1 52.9 10–606 18 2.1 88.7
Fig. 9. Thorax images of Eristalis tenax. A Ventral part of the thorax anterior half, where staminal column ofM. citrina strikes during tripping; B and C Pollen load of M. citrina adhered to thorax tegument
40 C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands
set than either simple tripping or spontaneousselfing. This difference can be explained by thevery low rates of auto-tripping (<20%observed) and self-deposition of pollen on thestigma when tripping is not the result of insectvisits (Fig. 6). Tripping is a predominantmechanism for the reproduction of M. citrina,as it is probably for all perennial Medicagospecies (Lesins and Lesins 1979, Free 1993).This phenomenon in M. citrina is almostentirely the result of insect visits and, veryrarely, caused by other factors, such as tem-perature and humidity (Juan et al. unpubl.data). Interestingly, our results indicate thattripping may be caused by agents other than
insects (as previously mentioned by wind orhigh temperatures), that might enhance natu-ral pollination and seed set.
To ensure reproductive success, both trip-ping and active pollen transfer are required.This is illustrated by the extremely low seed setafter natural pollination (45.3% tripping rate)compared with tripping by hand (100% trip-ping rate) and hand-pollination (Fig. 6). Theseresults imply that the species either suffersfrom pollen limitation due to low insectvisitation rate (Syrphidae and Calliphoridae)or inefficient pollen transfer, or from interfer-ence from heterospecific pollen adhering to thebodies of visiting insects (Table 1). Such
Fig. 10. Reproductive successof M. citrina in experimentalenclosures with different insectsin 1998. Each column gives themean number of viable seedsper fruit as a result of thepollinating activity of 1–2 indi-vidual insects in the enclosure.Sample sizes and standarderrors are given
Fig. 11. Percentage of fruit setof M. citrina due to the differ-ent insects used in the enclosureexperiments during the years1997 and 1998
C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands 41

interference may come from pollen fromthe abundantly co-flowering Lobularia mariti-ma subsp. columbretensis, whose pollen wasoften found on the pollinator’s body andtherefore effectively competes M. citrina pollenon the floral stigma (Waser 1978, Petanidouet al. 1995a) or may simply clog the floralstigma.
Medicago citrina starts flowering early andextends throughout most of the season, whichis unusual under Mediterranean conditions(Petanidou et al. 1995c). The extremely longflowering period is combined with a longflower life span, even in flowers that have beentripped since their first day of anthesis (8.0days vs 5.7 in tripped flowers). This span ismuch higher than that recorded in similarspecies in other Mediterranean communities,which usually range between 2.3 and 3.2 days(Petanidou 1991). The long flower life spanmay indicate a lack of appropriate pollinatorsin the community. A further response to theabsence of its usual pollinators (bees) is thepattern of nectar secretion. Nectar is secretedmainly early in the morning, a period whenlarge-sized thermoregulating bees can forage,and less in the evening and minimally duringthe noon period (Heinrich 1993, Stone 1994).The flowers are more attractive during the firstthree days of anthesis, the period of themaximal nectar secretion, coinciding with theperiod of the highest potential floral fecundity.Indeed, our results show that floral fecundity ismaximal during the first day of anthesis,whereas it decreases steadily from the secondday onwards. The same holds true for pollenviability that is highest during the first andsecond day of anthesis. However, in non-tripped flowers pollen maintains relatively highviability for 5 days, whereas in tripped ones(i.e. exposed since the first day of anthesis) theviability drops dramatically after the secondday. All the above results indicate a likelypollination partnership in which large efficientbees may play a major role, as have been alsoobserved in all M. citrina populations of theBalearic Isles (Juan 2002). This is also indi-cated by the morphological characteristics of
M. citrina which exhibits classic characteristicsof many bee-flowers (i.e. vivid yellow, zygo-morphic-papilionoid) which require obligatorytripping for their pollination (Fig. 6).
Flower visitors and efficient pollinators
Columbretes have been reported as islandswithout bee permanent populations (Espanol1958, Garcıa Marı et al. 1991), which wasconsistent with our surveys during the flower-ing seasons of 1996–1998. The occurrence ofbees on the islands is very unusual andcorresponds to no more than a few wind-drifted individuals per year. The absence ofbees may have an enormous impact on thepollination ecology and reproductive successof the native M. citrina, which appears to be apredominantly bee-pollinated species.
The flower visitors of M. citrina in theColumbretes archipelago were almost exclu-sively flies. Blowflies visited the flowers onlyfor nectar, whereas hoverflies also visited theflowers, but for pollen. In both groups, insectvisitation coincided with the high rates ofnectar secretion in the morning, whereas fewerindividuals visited the flowers during the sec-ond phase of nectar secretion (18:00 h) whichhad a lower rate.
Based on results for the M. citrina pollentransfer on insect bodies (Table 1), the effec-tuated tripping rates compared to spontaneoustripping rates (50–66% vs. 16–20%), and thehigh seed set in the enclosure experiments(Fig. 10), it is concluded that the most efficientpollinators of M. citrina flowers in Columbre-tes were Eristalis tenax and Calliphora vicina.Eupeodes corollae and L. sericata were lessfrequent trippers and showed extremely lowpollinator efficiencies when compared to spon-taneous pollination. Based on field data (sur-veys and enclosure experiments), we suggestthat E. tenax and C. vicina are the usualtripping agents because of their size andproboscis length (5–7 and 4–5 mm, respec-tively, compared to 2–3 mm of L. sericata andE. corollae). This morphological characterallows them to reach the nectar accumulated
42 C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands

at the bottom of the corolla tube and agitatethe auricles of the wing petals, and thereforeinitiate the tripping process.
An evaluation of flower visitors, based onan analysis of pollen transfer by insects,percentage of effectuated tripping and repro-ductive success in enclosures, indicates that thepollen vector of the highest relative importancewas E. tenax. This efficiency as a pollinatormay be enhanced by the presence of manypalynophilic hairs on the insect body. Accord-ing to Holloway (1976), direct pollen feeding inE. tenax has been supplemented and probablyreplaced by a pollen-collecting system operat-ing as the fly collects nectar. According to thesame author, the effectiveness of E. tenax isincreased by the fact that, during the trippingprocess, the pollen grains are mainly wide-spread on the anterior half of the ventralsurface of the thorax, where they can remainfor several days. However, the efficiency ofE. tenax in absolute terms is still quite low tobe truly considered an efficient pollinator ofM. citrina flowers.
Absence of well adapted pollinators andconsequent low pollinator efficiency is not theonly problem M. citrina faces with respect toits reproduction in Columbretes. In addition,the major pollinator E. tenax, is a migratoryspecies and an accidental visitor to the archi-pelago (Fig. 7; Aubert and Goeldlin de Tiefe-nau 1976, 1981; Aubert and Jaccard 1981).Our 2-year data show some fluctuations ofE. tenax populations during the floweringperiod implying that the populations are notpermanent in Columbretes, and probablyoriginating from the adjacent coastal popula-tions. The presence of the necrophagousblowfly Calliphora vicina, which is of marginalimportance for the reproduction of M. citrina,depends mainly on the availability of birdcarcasses, as well as the human activities on theislands.
Our results show that the reproduction ofM. citrina in Columbretes is mainly based onautogamy. This holds true even in cases whenfloral tripping is carried out by casual pollina-tors (E. tenax and C. vicina), although it is
difficult to ascertain the extent of self- orcross-pollination. According to our experienceand the resulting data (Fig. 6), during thetripping process some self pollen is depositedon the stigma either autonomously or byinsects. The high degree of inbreeding is alsosupported by pollen loads on the flies, whichconsist of very low percentages of M. citrinapollen.
Conservation biology and management
On the Grossa islet, M. citrina has been facingserious problems of survival due to non-sustainable management in the past, butnowadays those problems have disappeared.On Foradada and Ferrera, the plants sufferfrom ‘‘outbreaks of herbivores’’. For example,during the period 1995–98 a severe attack byIcerya purchasi Maskell, 1878 (Hemiptera,Margarodidae) destroyed c. 44% of the flow-ering individuals of Foradada (SanchezCodoner, pers. comm.).
Columbretes constitute an environmentwhere bee pollinators are virtually absent,and, as a result, the plants are pollen limited.Although the lack of bees can be perceived as adeficit of the island community, when all of thepopulations within the greater archipelago areconsidered, it appears that the populations arestill viable. The natural absence of bees may bea common intrinsic phenomenon to the islandevolutionary system, which may constitute atrigger factor for local adaptations and highendemic rates (Brown and Lomolino 1998,Anderson et al. 2001).
Regarding the observed plant-pollinatorpartnership under the existing isolation con-straints, the migratory hoverfly E. tenax, andthe blowfly C. vicina are very important inproviding pollination services, hence support-ing the persistence of M. citrina in the archi-pelago. Such casual relationships of ‘‘casual’’pollinator agents with small isolated popula-tions in tiny Mediterranean islands are worthyof systematic study. For instance, it would beinteresting to know to what extent plantcharacteristics like endemism, rarity or popu-
C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands 43

lation size are related to these fragile, informalplant-pollinator partnerships like that ofM. citrina. Perhaps in the future, biogeogra-phers should draw more attention to theseinformal cases of island pollination, focusingon aspects illustrating the subtle pollinatingterritories (e.g. migratory paths and strips),which have evolved and maintained to date thesplendid Mediterranean nature garden.
The project has been financially supported bythe investigation projects GV-C-RN-12-069-96 andBSO2000-0148. We thank Conselleria de MedioAmbiente (Generalitat Valenciana) for allowing usto carry out this study in the Columbretes NaturalPark, and the staff of the park, namely JuanJimenez and Jose Vicente Escobar, that madealways our demands possible. Special thanks toall the guards of the Park for their help, patienceand understanding during the fieldwork. The paperhas benefited greatly from the criticism by GerardOostermeijer, Graham Rotheray, Dan Eisikowitchand Simon Potts that reviewed different versions ofthe manuscript.
References
Aizpuru I., Ballester G., Banares A., Bascones J. C.,Benito J. L., Blanca G., Blanche C., Blanco E.,Bramwell D., Bueno A., Cabezudo B., CarqueE., Cirujano S., Crespo M. B., Devesa J. A.,Dıaz T. E., Domınguez F., Fabregat C.,Fernandez J. A., Galicia D., Goni D., GuemesJ., Guzman D., Hernandez J. E., Herranz J. M.,Herrero-Borgonon J. J., Laguna E., Lopez G.,Lopez S., Marrero M. V., Marrero A., MateoG., Monserrat P., Moreno J. C., Naranjo J.,Nava H. S., Navarro B., Ortiz S., Rico E.,Rosello J. A., Saez L., Sainz H., Sanchez-GomezP., Serra L., Sese J. A., Ugarte R., Vaquero J.,Vargas P., Villar L. (2000) Lista Roja de FloraVascular Espanola (valoracion segun categorıasUICN). Conserv. Veg. 6: 11–38.
Anderson G. J., Bernardellom G., Stuessy T. F.,Crawford D. J. (2001) Breeding system andpollination of selected plants endemic to JuanFernandez islands. Amer. J. Bot. 88: 220–233.
Aparicio A., Mitjavila J. M., Arana V., Villa I. M.(1991) La edad del volcanismo de las islasColumbrete Grande y Alboran (Mediterraneooccidental). Bol. Geol. Miner. 102: 562–570.
Aubert J., Goeldlin de Tiefenau P. (1976) Douzeans de captures systematics de Syrphides (Dip-teres) au col de Bretolet (Alpes valaisannes).Mitt. Schweiz. Entomol. Ges. 44: 115–142.
Aubert J., Goeldlin de Tiefenau P. (1981) Obser-vations sur les migrations de Syrphides (Dipt.)dans les Alpes de Suisse occidentale. Bull. Soc.Entomol. Suisse 54: 367–370.
Aubert J., Jaccard M. (1981) La migration desSyrphides (Dipteres) dans le Jura vaudois. Bull.Soc. entomol. Suisse 54: 377–388.
Barrett S. C. H., Kohn J. R. (1991) Genetics andevolutionary consequences of small populationsize in plants: implications for conservation. In:Falk D. A., Holsinger K.E. (eds.) Genetics andconservation of rare plants. Oxford UniversityPress, Oxford, pp. 3–30.
Berry P. E., Calvo R. N. (1989) Wind pollination,self incompatibility and altitudinal shifts inpollination systems in the high Andean genusEspeletia (Asteraceae). Amer. J. Bot. 76: 1602–1614.
Beattie A. J. (1972) A technique for the study ofinsect-borne pollen. Pan-Pacific Entomologist47: 82.
Bibiloni G., Alomar G., Rita J. (1993) Floravascular dels illots i addicions a la flora deCabrera Gran. In: Alcover J. A., Ballesteros E.,Fornos J. J. (eds.) Historia natural de l’Arxipe-lag de Cabrera Consell Superior d’lnvestigacionsCientifiques. Ed. Moll. Mallorca, pp. 179–206.
Blondel J., Aronson J. (1999) Biology and wildlifeof the Mediterranean region. Oxford UP, NewYork.
Brown J. H., Lomolino M. V. (1998) Biogeogra-phy. Sinauer Associates, Sunderland, Massa-chussets.
Carlquist S. (1974) Island biology. Columbia Uni-versity Press, New York.
Dafni A. (1992) Pollination ecology – a practicalapproach. Oxford University Press, Oxford.
Espanol F. (1958) Contribucion al conocimiento delos artropodos y moluscos terrestres de las IslasColumbretes. Misc. Zool. 1: 3–37.
Fausto Jr J. A., Eckhart V. M., Geber M. A.(2001) Reproductive assurance and the evolu-tionary ecology of self-pollination in Clarkiaxanthiana (Onagraceae). Amer. J. Bot. 88: 1794–1800.
Fischer M., Matthies D. (1998) RAPD variation inrelation to population size and plant fitness in
44 C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands

the rare Gentianella germanica (Gentianaceae).Amer. J. Bot. 86: 811–819.
Free J. B. (1993) Insect pollination of crops.Academic Press, London.
Garcıa Marı F., Ferradgut F., Navarro V.,Laborda R., Costa-Comelles J. (1991) Nuevasaportaciones al conocimiento de los artropodosde las Islas Columbretes. In: Alonso L. A.,Carretero J. L., Garcıa Carrascosa M. A. (eds.)Islas Columbretes. Contribucion al estudio de sumedio natural (Coleccion Monografias, n� 5),Valencıa: Conselleria de Obras Publicas, Urban-ismo y Transportes – Agencia del Medi Ambient– Generalitat Valenciana, pp. 155–179.
Gilbert F. S. (1993) Hoverflies. Naturalist Hand-books 5. Cobret & Disney.
Greuter W., Burdet H. M., Long G. (1989) Med-Checklist. 4. Conservatoire et Jardin botaniquesde la Ville de Geneve. Italy.
Groom M. J. (2001) Consequences of subpopula-tion isolation for pollination, herbivory, andpopulation growth in Clarkia concinna. Biol.Conserv. 100: 55–63.
Heinrich B. (1993) The hot-blooded insects: strat-egies and mechanisms of thermoregulation.Harvard University Press, Cambridge.
Holloway B. A. (1976) Pollen-feeding in hoverflies(Diptera: Syrphidae). New Zeal. J. Zool. 3: 339–350.
Howarth F. G., Mull W. P. (1992) Hawaiian insectsand their kin. University of Hawaii Press,Honolulu.
Juan A. (2002) Studies on morphology, geneticvariability and reproductive biology ofMedicagocitrina (Font Quer) Greuter (Leguminosae).Bases for its conservation. Unpublished Ph.D.Thesis, University of Alicante.
Juan A., Crespo M. B., Rios S. (1999) Medicagocitrina (Font Quer) Greuter (Leguminosae):variabilidad morfologica, ecologıa y estadoactual de sus habitats. Proceedings of theXXXIX Reunion Cientıfica de la SociedadEspanola para el Estudio de los Pastos S. E. E.P. (Almerıa) 87–91.
Kearns C. A., Inouye D. W. (1993) Techniques forpollination biologists. University Press of Colo-rado, Niwot, Colorado.
Lesins K. A., Lesins I. (1979) Genus Medicago(Leguminosae). A taxogenetic study. Den Haag,Boston & London.
McCullen C. K. (1990) Reproductive biology inGalapagos Islands angiosperms. Monogr. Syst.Bot. 32: 35–45.
Mateo G., Crespo M. B. (2001) Manual para ladeterminacion de la flora valenciana. 2 edition.40 Molinier, Valencia.
Menges E. S., Dolan R. W. (1998) Demographicviability of populations of Silene regia inmidwestern prairies: relationships with firemanagement, genetic variation, geographic loca-tion, population size and isolation. J. Ecol. 86:63–78.
Oostermeijer J. G. B., Luijten S. H., Petanidou T.,Kos M., Ellis-Adam A. C., den Nijs J. C. M.(2000) Pollination in rare plants: is populationsize important? In: The Scandinavian Associa-tion for Pollination Ecology honours KnutFaegri. I. Matematisk NaturvidenskapeligeKlasse, Skifter, Ny Serie 39. Det Norske Vi-denskaps-Akademi, Oslo, pp. 201–213.
Oostermeijer J. G. B., Luijten S., Kwak M.,Boerrigter E., den Nijs H. (1998) Rare plantsin peril: on the problems of small populations.Levende Natuur 99: 134–141.
Oostermeijer J. G. B., Nijs J. C. M. den, RaijmannL. E. L., Menken S. B. J. (1992) Populationbiology and management of the marsh gentian(Gentiana pneumonanthe L.), a rare species inThe Netherlands. Bot. J. Linn. Soc. 108: 117–130.
Oostermeijer J. G. B., van Eijck M. W., den NijsJ. C. M. (1994) Offspring fitness in relation topopulation size and genetic variation in the rareperennial plant species Gentiana pneumonanthe.Oecologia 97: 289–296.
Ouborg N. J. (1993) Isolation, population size andextinction: the classical and metapopulationapproaches applied to vascular plants along thedutch Rhine-system. Oikos 66: 298–308.
Ouborg N. J., van Treuren R., van Damme J.M.M.(1991) The significance of genetic erosion in theprocess of extinction. II. Morphological varia-tion and fitness components of varying size ofSalvia pratensis L. and Scabiosa columbaria L.Oecologia 86: 359–367.
Perez-Banon C. (2000) Biology of the syrphids(Diptera: Syrphidae) of the insular ecosystems inthe Community of Valencia: aspects of theinsect-plant relationship. Unpublished Ph.D.Thesis. University of Alicante.
C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands 45

Petanidou T. (1991) Pollination ecology in aphryganic ecosystem. Unpublished Ph.D. Thesis.Aristotle University.
Petanidou T., den Nijs H., Ellis-Adam A. (1991)Comparative pollination ecology of two raredutch Gentiana species in relation to populationsize. Acta Hort. 288: 308–312.
Petanidou T., den Nijs J. C. M., OostermeijerJ. G. B. (1995a) Pollination ecology and con-straints on seed set of the rare perennial Gentianacruciata L. in The Netherlands. Acta Bot. Neerl.44: 55–74.
Petanidou T., den Nijs J. C. M., OostermeijerJ. G. B., Ellis-Adam A. C. (1995b) Pollinationecology and patch-dependent reproductive suc-cess of the rare perennial Gentiana pneumonanthein The Netherlands. New Phytologist 129: 155–163.
Petanidou T., Ellis W. N., Margaris N. S., VokouD. (1995c) Constraints on flowering phenologyin a phryganic (East Mediterranean) ecosystem.Amer. J. Bot. 82: 607–620.
Raijmann L. E. L., Leeuwen N. C. van, Kersten R.,Oostermeijer J. G. B., Nijs J. C. M. den.,Menken S. B. J. (1994) Genetic variation andoutcrossing rate in relation to population size inGentiana pneumonanthe L. Conserv. Biol. 8:1014–1026.
RamseyM., VaughtonG. (1996) Inbreeding depres-sion and pollinator availability in a partiallyself-fertile perennial herb Blandfordia grandiflora(Liliaceae). Oikos 76: 465–474.
Serra L., Perez Botella J., Izquierdo J. (2001)Medicago citrina (Font Quer) Greuter (Legumi-nosae) en la Penınsula Iberica. Anal. Jard. Bot.Madrid 59: 158–159.
Small E., Lassen P., Brookes B. S. (1987) Anexpanded circumscription of Medicago (Legumi-nosae, Trifolieae) based on explosive floweropening. Willdenowia 16: 415–437.
Sokal R. R., Rohlf F. J. (1981) Biometry: theprinciples and practice of statistics in biologicalresearch, 2nd edn. W. H. Freeman, San Fran-cisco, NY.
Stone G. N. (1994) Activity patterns of females ofthe solitary bee Anthophora plumipes in relationto temperature, nectar supplies and body size.Ecol. Entomol. 19: 177–189.
Treuren R. van., Bijlsma R., Ouborg N. J., DeldenW. Van (1991) The significance of geneticerosion in the process of extinction. I. Geneticdifferentiation in Salvia pratensis and Scabiosacolumbaria in relation to population size. Her-edity 66: 181–189.
Waser N. M. (1978) Interspecific pollen transferand competition between co-occuring plantspecies. Oecologia 36: 223–236.
Young A. G., Brown A. H. D., Zich F. A. (1999)Genetic structure of fragmented populations ofthe endangered Daisy Rutidosis leptorrhynch-oides. Conserv. Biol. 13: 256–265.
Addresses of the authors: C. Perez-Banon,A. Juan, Ma A. Marcos-Garcıa, M. B. Crespo,Centro Iberoamericano de la Biodiversidad,Universidad de Alicante, P.O. Box 99, E-03080Alicante, Spain; T. Petanidou (e-mail: [email protected]), Department of Geography, Univer-sity of the Aegean, Building of Geography, Uni-versity Hill, GR-81100 Mytilene, Greece.
46 C. Perez-Banon et al.: Reproductive ecology of Medicago citrina in bee-deprived islands





























