Embed Size (px)
Citation preview

lable at ScienceDirect
Quaternary Science Reviews 110 (2015) 131e151
Contents lists avai
Quaternary Science Reviews
journal homepage: www.elsevier .com/locate/quascirev
The remarkable Panthera pardus (Felidae, Mammalia) record from Equi(Massa, Italy): taphonomy, morphology, and paleoecology
Elena Ghezzo*, Lorenzo RookUniversity of Florence, Via La Pira 4, Florence, 50121, Italy
a r t i c l e i n f o
Article history:Received 5 October 2014Received in revised form18 December 2014Accepted 19 December 2014Available online
Keywords:Panther pardusCarnivore guildIntraspecific variabilityLate PleistoceneEqui caveItaly
* Corresponding author. Tel.: þ39 3334365026.E-mail address: [email protected] (E. Ghezzo).
http://dx.doi.org/10.1016/j.quascirev.2014.12.0200277-3791/© 2014 Elsevier Ltd. All rights reserved.
a b s t r a c t
Leopard remains are rare in the European fossil record, probably a consequence of its solitary and elusivehabits. Equi, dating back to Marine Isotope Stage 3 (MIS3), represents a rich and outstanding exception.Historical excavations (from 1911 to about 1917) at Equi allowed more than 200 remains to be recoveredof this large felid, a sample attesting the richest leopard-bearing site in the Pleistocene of Europe. TheEqui site testifies the survival of a prosperous leopard population in northwestern Italy (the Apuan Alpsarea) during the pre Last Glacial Maximum Late Pleistocene.
Almost all skeletal elements are represented in the collection (except ribs, and most of axial skeleton).At Equi, leopard is represented by both young and adult animals and, up to now, it seems the first andunique Pleistocene evidence of cave usage for cubs raised in Europe.
This study focuses on the whole record of leopard from Equi, housed in the Museum of Natural Historyof Florence, and is aimed to analyze the taphonomical assemblage, to provide an anatomical descriptionof the remains and the comparison to the European evidences. Moreover, we infer the paleoecology ofleopards, and their relationships with other large carnivores from the Equi guild (Ursus spelaeus, Canislupus, and Panthera leo spelaea). The site was probably an occasional large carnivores den, used over timeby different large carnivore taxa, perhaps in different seasons or times (mutual avoiding).
Our morphological analysis allows us to recognize sexual dimorphic characters in the cranium, and alarge morphological variability in body size.
© 2014 Elsevier Ltd. All rights reserved.
1. Foreword
Panthera pardus, the largest spotted cat in Africa and Asia, withseveral recognized subspecies, is a quite common, solitary, andopportunistic predator in the wild (Miththapala et al., 1996;Uphyrkina et al., 2001; Sunquist and Sunquist, 2002). It shows agreat size variability within its range: generally speaking, in opencountry the animals are larger than in the forest, from 20 to 30 kgup to 90 Kg, with a large variability in food habits (Hayward et al.,2006). It can live in a wide range of habitats, most commonlyassociated with cover forests and mountain terrain, from sea levelup to over than 5.000 m (Sunquist and Sunquist, 2002). At presentthe species is critically endangered in some parts of its range, listedas Near Threatened in the IUCN Red Data Book (Sunquist andSunquist, 2002; Gavashelishvili and Lukarevskiy, 2008; Henschelet al., 2008).
A general revision of European Pleistocene leopards has beenrecently provided by Fischer (2000), Sauqu�e and Cuenca-Besc�os(2013), and Diedrich (2013): P. pardus was firstly determined inEurope in late Early Pleistocene at Vallonnet Cave (Moulle et al.,2005) and at about 600 ky at Mauer (Schütt, 1969), and was quitecommon across the continent during the Middle and Late Pleisto-cene (Diedrich, 2013) with a Late Pleistocene northern limit of therange at the site of Niederlehme-Rixdorfer Horizont in Berlin(Fischer, 2000). In Europe leopards disappeared during the LatePleistocene, with a strong decrease at about the LGM, probablysurviving only in Eastern Europe until historical times (Spassov andRaychev, 1997).
Molecular phylogenies indicate that African leopards are themost primitive ones, those from western and central Asia evolvedlater fromAfrican ancestors, and those of northern and eastern Asiashows the youngest genome sequence (Miththapala et al., 1996;Uphyrkina et al., 2001). Uphyrkina et al. (2001) inferred leopards'origins of African and Asian subspecies should be dated around0.47e0.825 Ma and 0.17e0.30 Ma respectively, even though these

E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151132
data are completely in disagreement with paleontological evidence(O'Regan and Menter, 2009). On the other hand, Lei et al. (2011)estimate P. pardus origins at about 4.35 Ma ago, a time older thanthe first sure fossil evidence of the species in Africa at ca 2.0 Ma(Werdelin and Peign�e, 2010). As a matter of fact the question aboutleopard earlier appearance and differentiation is still far from beingresolved.
Even though many authors attempt to recognize fossil subspe-cies from morphological analysis (Schütt, 1969; Kotsakis andPalombo, 1979; Spassov and Raychev, 1997; Diedrich, 2013), wepoint out that, till today, leopard occurrences are too scarce to allowfor firm conclusions, and specimens are classified into differentsub-groups without a bold cladistical and/or genetic analysis. Boule(1906) recognized Panthera pardus fossilis characterized by moremassive dental features and an elongated lower carnassial incomparison to the p4 (Bonifay, 1971). Similar characters aredifferently notable both in other fossils and extant subspecies andare not so precisely identified by the author. We emphasize that ifintraspecific variability was taken into account, also the Diedrich(2013) hypothesis of successive migration waves from Africawould not be justified, and the four subspecies Panthera pardusbegoueni (Fraipont, 1923), Panthera pardus sickenbergi (Schütt,1969), Panthera pardus antiqua (Cuvier, 1835), and Panthera par-dus spelaeus (B€achler, 1936) should be better considered as a singlechronospecies P. pardus Linnæus, 1758 (instead of different sub-species characteristics of Early, Middle, and Late Pleistocene). Therole of leopards in the Pleistocene carnivore guild is related to theinteraction with other carnivores and with prey, and, according toSaoqu�e et al. (2014) and de Ruiter and Berger (2001), the leopard
Fig. 1. The location of Equi in the Tuscany region (A). Equi is located on the northern side of tper i Beni Archeologici della Toscana). The valley of Fagli has a V-shape and the cave entra
presence can influence the mammal assemblage structure, reba-lancing the species occurrences. For this reason, understanding itsbehavior in comparison to other predators (including humans) iscrucial.
The present paper is devoted to tracking the leopards' profilefrom Equi, considering taphonomy, morphology, aging and rela-tionship with other species recorded at the site, aiming for a betterviewof the Equi mammal assemblage, and to the role of this speciesin the Late Pleistocene of Europe.
2. The site of Equi
Equi is a small town on the northern side of the Apuane Alps(Fig. 1AeB). The topography of the area is characterized by driedenvironment and V-shaped fluvial valleys, even though some areaswere interested by Late Pleistocene glaciation events. Evidence ofsmall glaciers is spread most on the north-eastern side of theApuane Alps (Braschi et al., 1986; Giraudi, 2004, 2011).
The Equi paleontological site is a cave and shelter located justout and above of the town, on the left side of the fluvial valley ofFagli (coordinates WGS84 44.150, 10.167) (Fig. 1C). The cave depositrecords Holocene evidence (Medieval pottery and Neolithic re-mains), and Late Pleistocene vertebrate fossils associated withMusterian artifacts (Regalia, 1911; De Stefani, 1917; Del Campana,1923, 1954; Branchini, 1928; Battaglia, 1932; Graziosi, 1934;Rellini, 1935, 1937; Paribeni et al., 2009; Bigagli et al., 2013).
The first news about the paleontological remains from Equidates back to 1909, when G. Poderzana from the ArchaeologicalMuseum of La Spezia discovered and collected some fossils from
he Apuane Alps, above the homonymous town (B) (photo by P. Nannini, Soprintendenzance is north-east oriented (C).

E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151 133
the external shelter. Soon after, the Geology and PaleontologyMuseum of Florence University (at that time “Regio Istituto di StudiSuperiori”) continued the study and excavation of the site, foralmost a decade, under the direction of Prof. C. De Stefani. Thanks tothe excavation of the shelter deposit, the entrance of the cave wasfound in 1911. The 1909e1917 excavation allowed for the collectionof thousands of animal remains in association with human arte-facts. Also, two hearths were identified in the deposit, at differentdepths, but unfortunately not preserved today (cfr. Ghezzo et al.,2014b).
The work of De Stefani's team (described by De Stefani, 1917)and the detailed information record for almost each find, reportedon labels, allowed for a recent revaluation of the species distribu-tion within the depositional succession, and the identification of aquite homogeneous fauna assemblage along the succession(Ghezzo et al., 2014b, cfr. Fig. 11).
In 1921, according to the Italian law, the site of Equi was declareda site of paleontological importance, under the responsibility of thelocal Soprintendenza Archeologica. Again in late 1900, in accor-dance with an Italian law regarding cultural heritage, the Equi sitewas included in the areas of special planning and control forarchaeological and landscape importance (Paribeni et al., 2009;Iardella et al., 2011).
Since the De Stefani excavation times, the evidence of twodistinct phases of human presence (ceramics and Palaeolithic ar-
Beta sample Type of sample Layer Measured age 13C/12C Conventional age 2 Sigma calibration
360175 Panthera pardus ilium fragment HB? 32,910 ± 260 BP �19.6 0/00 33,000 ± 260 BP Cal BC 36,630e34,840Cal BP 38,570e36,800
360173 Rupicapra rupicapra bone fragment (diaphysys) F 26,960 ± 140 BP �20.9 0/00 27,030 ± 140 BP Cal BC 29,480e29,240Cal BP 31,430e31,190
360176 Rupicapra rupicapra femur fragment (diaphysys) B 27,560 ± 160 BP �20.5 0/00 27,630 ± 160 BP Cal BC 30,030e29,480Cal BP 31,980e31,430
tefacts) induced a lengthy harsh scientific debate between thehighest authorities in the field (Mochi, 1916, 1920, 1929; Rellini,1917, 1924, 1929) which was resolved only fifteen years later,when an excavation field season by the Italian Institute of HumanPalaeontology (Graziosi, 1934; Rellini, 1935) confirmed the absenceof ceramics within Palaeolithic levels.
After the extensive excavations by De Stefani, the deposit wasnot systematically and coherently excavated again. Although, inaddition to the work by the Italian Institute of Human Paleontologyin Rome, small excavations were undertaken by various researchersin the late 1960s to early 1970s (Ambrosi and Fabbri, 1975) andlate 1970s (Guerri, 1980, 1982).
In 2009, under an agreement among the Soprintendenza per iBeni Archeologici della Toscana, the Municipality of Fivizzanoand the Apuane Alps Regional Park, a new project began, and isstill ongoing, with the aim of validating the site with newexcavations.
Historical excavation from 1911 up to about 1917 resulted in anextremely rich faunal collection (presently housed in the NaturalHistory Museum, Geology and Paleontology Section, of the Uni-versity of Florence). It consists of small and large vertebrates, fromamphibians to mammals. Among mammals, carnivores are themost represented with an enormous record of bears remains,several felines and canids (wolves, foxes, and the rare dhole; DelCampana, 1923; Ghezzo and Rook, 2014). The determined faunaconsists of bats, birds, herpetofauna (Bartolini et al., 2014) and fish,in addition to Ursus spelaeus Rosenmüller, 1794, Canis lupusLinnæus, 1758, Vulpes vulpes (Linnæus, 1758), Cuon alpinus (Pallas,1811), Panthera leo spelaea (Goldfuss, 1810), P. pardus (Linnæus,
1758), Lynx lynx (Linnæus, 1758), Mustela nivalis Linnaeus, 1766,Mustela putorius Linnæus, 1758, Martes martes Linnæus, 1758,Martes foina (Erxleben, 1777), Cervus elaphus Linnæus, 1758, Susscrofa Linnæus, 1758, Rupicapra rupicapra (Linnæus, 1758), Capraibex Linnæus, 1758, Capra hircus (Linnæus, 1758) (only from theupper layers), Lepus sp., Marmota marmota (Linnæus, 1758), Castorfiber Linnæus, 1758 (just one tooth), Glis glis (Linnæus, 1758), Eli-omys quercinus (Linnaeus, 1766), Arvicola amphibius (Linnæus,1758), Microtus nivalis (Martins, 1842) (De Stefani, 1917; DelCampana, 1923, 1954).
The “Equi” fauna, though not re-examined in detail in recenttimes, is generally correlated (Caloi and Palombo, 1994) with themiddle of Marine Isotope Stage 3 (MIS 3; approximately in therange of 27e50 ka; cfr. Siddall et al., 2008). The recent new exca-vation survey of the cave confirmed the presence of in-situarchaeological deposits with radiometric ages on excavated mate-rial consistent with the biochronological interpretation of the his-torical collection (radiocarbon ages of 43,700 ± 1900 and44,000 ± 2200 BP; Bigagli et al., 2013).
During our revision of the De Stefani collection of historical Equiexcavations, three bone samples with registered stratigraphicprovenance were sent to the Beta Analytic Inc. (Miami, Florida, Usa)for radiometric dating (Accelerator Mass Spectrometry, AMS,analysis). All the three remains contained sufficient collagen anddata from these analyses are listed below:
These dates are also consistent with the biochronologicalinterpretation and with the recent radiometric dating of specimensrecently excavated from older layers (Bigagli et al., 2013). ThePanthera pardus specimen (sample #360175) tentatively referred tolayer HB seems to indicate an age inversion from a layer strati-graphcally superimposed to that of samples #370173 and #360176.The degree of uncertainty due to possible bias in the data availablefor the historical collections (samples were excavated in1911e1917), do not allow us however to exclude the provenance ofthe specimen from an older layer.
Despite this uncertainty, the overall information furnishedby theradiometric datingof these three samples fromhistorical collectionsallowus to chronologically place the Equi P. pardus recordwithin themiddle to latest portion of the MIS3, a time span characterized inMediterranean Europe (noticeably France, Italy, Greece and Turkey)by significant glaciation phases (Hugues andWoodward, 2008) andby climate-driven changes in the vegetation (Harrison and Prentice,2003; Fletcher et al., 2010). It is noticeable that the Equi sampletestify the survival of the European Leopard until a period of severeclimate, with mean temperatures progressively diminishing to-wards the Last Glacial Maximum (Mangerud et al., 2004).
3. Materials and methods
This study focuses on the Equi leopard fossils recovered duringthe fieldwork by De Stefani and his team (1911e1917) and housedin the Natural History Museum, Geology and Paleontology Section,of the University of Florence (IGF), firstly described by Del Campana(1954) as P. pardus antiqua.

E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151134
The Equi P. pardus collection consists in more than 200 bones(some of which are exposed in the Florence Natural HistoryMuseum permanent exhibition). Most of cranial and postcranialbones are almost completely intact and well preserved.
Morphological and dental nomenclature follows Barone (2003a;2003b) and Hillson (1996). Measurements (data available as onlinesupplementary data) follow von der Driesch (1976), while ages atdeath are here determined on the basis of epiphysis junctions anddental eruption. For comparative purposes we had access to thecollections of the Natural History Museum of Verona and Venice(Italy), the Natural History Museum of Berlin (Germany) and theHungarian Natural History Museum (Budapest), where leopardfossils are hosted, discovered in the Galerian sites of Soave Sentiero(Pasa, 1947) Cer�e Cave (Ghezzo et al., 2014a), and Upponyi (Hank�o,2007), and the Late Pleistocene leopards remains from FumaneCave (Cassoli and Tagliacozzo, 1991), Lambrecht Cave (Janossy,1986), Niederlehme (Fischer, 2000), Baumann's Cave (Diedrich,2013). Skeletons of extant leopards were also considered, hostedin the Natural History Museum, Zoology Section “La Specola” of theUniversity of Florence (MZF), the osteological collections of theArcheozoology Laboratory of the University of Siena and theVertebrate Paleontology Laboratory of the University of Ferrara. Allthese specimens are from adult animals; only one cranium recordsdata about sex (i.e. the female MZF1221-4126 from China) and onlya few records the region of capture (i.e. China, Angola and Somalia).
Additional comparative data are derived from bibliographicsources (Schütt, 1969; Kotsakis and Palombo, 1979; Spassov andRaychev, 1997; Nagel, 1999; Cardoso and Regala, 2006;Baryshnikov, 2011; Marciszak et al., 2011; Diedrich, 2013; Saoqu�eet al., 2014b).
Main abbreviations: C-M1: total length of the canine and cheekteeth; LP4: total length of the upper carnassial; p-m: total lengthof lower dental cheek series; m1L: total length of the lowercarnassial; Mc: metacarpal; Mt: metatarsal; Gl: maximum length;sd: medio-lateral minimumwidth; bd: distal width; btr: width ofthe trochlea.
4. Taphonomic remarks
With a total number of 224 determined finds, Equi currentlyrepresents the most abundant collection of leopards across Europe,far exceeding the Late Pleistocene sites of Zafarraya (Barroso et al.,2003), Wildkirchli Cave (Switzerland), Baumann's Cave (Germany),the assemblage from Cueva de Los Rincones (Spain) and Caune del’Arago and Hortus (France) (Testu, 2006, 2011; Diedrich, 2013;Saoqu�e et al., 2014a).
Adults are dominant (60% of the whole collection), followed byyoung animals (Fig. 2A). Adult age-stages are also predominantamong different anatomical regions (Fig. 2B). The occurrence ofalmost one perinatal cub is also testified by one maxillar with de-ciduous teeth in eruption (IGF6120V, Fig. 3AeC) and by few post-cranial bones, i.e. a fragile tibia diaphysis with un-ossifiedextremities (IGF5343V) (Fig. 16).
As observed by Del Campana (1954), all the anatomical regionsof leopards were sampled, with the exception of ribs, vertebrae andthird phalanxes that are very under-represented (Fig. 2D), both foryoung and adult animals.
Autopodials are the most frequent elements in the collection,with a total number of 46 determined metapodials (Fig. 2C).
The Minimum Number of Individuals (MNI), calculated consid-ering humeri and Mt within the stratigraphic distribution (Ghezzoet al., 2014b, cfr. Fig. 11), is 6 for the humeri (one for each layer a-a0,ab, hb, and i, and two right humeri in the layer l) or the Mt fre-quencies (one for each layer a-a0, ab, c, g, hb, and l), but thisapproximation is probably underestimated.
There are rare examples of gnawing and fluitation (sensu Fisher,1995; Lyman, 1994) within the entire assemblage, most on theproximal and distal ends of long bones. However, there are nodigested leopard bones. Evidences of peridepositional larval dam-age of periosteum are on IGF6141V (Fig. 3D), characterized bytwelve pinholes on the diaphysis (about 2.5 mm of diameter eachone) never reaching the spongy lumen of the bone. Similar occur-rences were observed in several fossils but have just recently beenanalyzed by Fejfar and Kaiser (2005), Harris and Timm (2013), andHolden et al. (2013).
5. Systematic paleontology
Order Carnivora Bowdich, 1821.Family Felidae Fischer Von Waldheim, 1817.Subfamily Felinae Fischer Von Waldheim, 1817.Genus Panthera Oken, 1816P. pardus Linnæus, 1758
5.1. Referred specimens
See Appendix.
5.2. Description
5.2.1. CraniaThe site of Equi returned five well preserved leopard crania
(Figs. 4 and 5). Three of them (IGF10036V, IGF15110V, andIGF15111V/1) are almost intact. The neurocranial and splancnoc-ranial sides are preserved as well as the zygomatic arcs, muzzlesand teeth.
IGF10036V (Figs. 4C, F, 5AeE) is the largest and most massive,compared to the other samples from the same site. On the contrary,IGF15111V/1 (Figs. 4A, 5FeG) is the smallest and most slightly builtcranium, and also the most damaged one (probably ruined duringthe excavation). IGF15111V/1 slightness is a feature not related to ayoung individual because all sutures are almost ossified and thetooth wear is at an advanced stage.
In lateral view, the crania have a straight sagittal crest.Compared with the cranium of extant leopards it is not projectingso far upward. With the exception of IGF185V/1, this crest sticks outslightly over the occipital surface.
Dorsal outline of the muzzle is convex, shorter than the neu-rocranial portion, with the nasal bones frontally sloped. The fronto-nasal suture is slightly sunken-in, and produces a deep depression(especially evident in IGF10036V and IGF15110V). The canine rootsproduce a swelling of the maxillar bones, ending above theinfraorbital foramen. IGF10036V is the only specimen showing themaxillary suture. This line follows the orbital edge and ends close tothe fronto-nasal suture. In ventral view, the three complete crania(IGF10036V, IGF15110V, and IGF15111V/1) show awide palate, withthe choanae opened between the canines.
The post orbital constriction is not marked in IGF185V/1,IGF10036V and IGF15110V, while is well pronounced in IGF184Vand IGF15111V/1. The braincases are elongated, elliptical (barrel-like) or spherical (globe-like) (Table 1).
5.2.1.1. Upper teeth. Incisives series is preserved only in IGF185V/2(Fig. 6). The I1 and I2 are frontally flat and their lingual outlinecurves forming an enlarged crown surface. There is a short dia-stema between incisives and canines. The presence of P2 is variable.IGF15111V/1 bears it only on the left side. IGF10036V has P2 on bothsides, while IGF15110V has no P2s at all (even the alveoli). DelCampana (1954) reported that the first premolars of IGF15110V

Fig. 2. Pie chart of leopard bones' age-stage (A), and age-stage frequencies for anatomical regions (B). Pie chart describing metapodial leopard remains from Equi (C). Generalrepresentation of leopard specimens from Equi (D).
Fig. 3. Fossil of perinatal cub of leopard (IGF6120V) in lingual (A), labial (B) and ventral (C) views. D: putative larval holes on diaphysis surface of a young humerus (IGF6141V).
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151 135
were lost during the lifetime and the alveolus was obliterated fromthe re-growth of the maxillar. In our opinion, in this skull the P2never erupted because there is no sign of bone remodeling on themaxillary bone. The third premolars are worn in all specimens.IGF15111V/1 shows an unusual wear of distal side of paracone,probably due to awrong occlusion of the maxillary and mandibulardental margins.
The carnassials are massive, deeply worn and morphologicallyvariable among the specimens. IGF10036V do not bear the leftcarnassial (probably lost for post-depositional events). The bestpreserved is the left P4 of IGF15111V/1, which has a slightly bi-lobedmetacone (similar to IGF185V/1). The latter is smaller than the
paracone tip (a characteristic that does not seem related to thewearstage). The distal lobe is stretched posteriorly. Protocone is trans-versally developed on the palatal surface and do not create aconstriction between paracone and metacone.
The M1 alveolus is preserved in all the crania, and is still presentonly on the left side of IGF15111V/1.
5.2.1.2. Mandibles. All the specimens have a rectilinear horizontalbranch (Fig. 7). The synphysis is long and inclined. No lower in-cisives and second premolars (or their alveoli) are found in ourspecimens.

Fig. 4. Cranial leopard remains from Equi in top view: IGF15111V/1 (A); IGF15110V (B); IGF10036V (C); IGF185V/1 (D); IGF184V (E). IGF10036V, IGF185V/1, and IGF184V are alsorepresented in lateral views (F, G, H, respectively).
Fig. 5. IGF10036V and IGF15111V/1 in detail. IGF10036V in dorsal (A), ventral (B), caudal (C), left (D) and right (E) views. Cranium IGF15111V/1 in dorsal view (G) and with mandibleIGF15111V/2 in left side view (F).
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151136
5.2.1.3. Lower teeth. Incisive alveoli are small, showing only a slightdimensional increase from i1 to i3. Cheek teeth are short andembricated. Canines are elongated, with the typical leopard middlegrooves on the labial side of the crown that exclude a determina-tion to other similar-sized species, such as the genus Puma orPanthera onca (Diedrich, 2013; Saoqu�e et al., 2014b). There is nodiastema between incisives and canines, and a short diastemaseparates the canine from the third premolar. The p3 is a smalltooth; mesial and distal ends of paraconid bear one small tip.
The p4 is larger: the protoconid mesial edge is inclined, long andstraight, while the distal one is shorter and curved. As a result, thecuspid ends before the midpoint of the tooth.
The m1 is similar in length to the p4. A short valley separatesprotoconid and paraconid. The latter is ever higher than the formerone, and occupies more than half of the tooth-blade length. Distally,a small bulge, when present, never creates a real metaconid.
5.2.2. Forelimb5.2.2.1. Humerus. The sample includes eleven humeri. These arelong and don't show a marked rotation of the diaphysis. IGF5342Vis complete and its diaphysis shows evidence of pectoralis musclestress (Fig. 8). IGF5352V and IGF5351V are significantly smaller, dueto the ontogenetic growth. The distal sovracondilar foramen iselongated to the distal end and its process is massive. The

Table 1Morphological characters of the specimens of P. pardus from Equi compared to other Pleistocene and Holocene skulls. OC-MZF: osteological collection of the Natural HistoryMuseum of the University of Florence.
Morphological characters of leopard crania
Fronto-nasal suture is slightly sunken-in Post-orbital constrinction Upper diastema Neurocranium feature P2 r/l
IGF15110V Yes Large Short Barrel 0/0IGF15111V/1 e Narrow Short Globe 0/1IGF10036V Yes Narrow No Barrel 1/1IGF185V/1 e Narrow Short Barrel 0/-IGF184V e Narrow e Barrel �/�Manga Larga No Large No Barrel 1/1Monte Sacro No Large No Barrel 1/1 (?)8748 (OC-MZF) No Large No Globe 0/18439 (OC-MZF) Yes (small) Large e Barrel �/�
Fig. 6. Premaxillar IGF185V/2 with incisive serie in frontal (A) ventral (B) and dorsal(C) views.
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151 137
trochanter ventral outline is rounded and curves from the lateral tothe medial side. IGF5341V (Fig. 8B) is from a very young animal anddoes not preserve the proximal epiphysis (un-ossified); the samespecimen instead preserves the distal epiphysis showing the ossi-fication line at the contact with the diaphysis.
5.2.2.2. Ulna. Ulnae in the sample are represented by six speci-mens. Only two are complete (Fig. 9), with the both the extremitiesossified. Ulnae show a great size variability. The articular concavityof IGF5402V (Fig. 9C) is similar to the homologous in IGF5354V(Fig. 9B), but the former is larger and has a greater outline.
5.2.2.3. Radius. The sample includes eight radii. The diaphysis ofthe radius is not compressed and is transversally elliptical. Thetuberosity is large and produces a hump just under the proximalepiphysis on the latero-distal surface. IGF5337V is the slenderradius specimen in the sample belonging to an adult animal(Fig. 10).
5.2.2.4. Metacarpals. The McII (Fig. 11) has a sub-rectilineardiaphysis. Frontally, there is a small bulge at the proximalmedial margin. The articular surface with the trapezoid is uniqueand has a triangular shape. There is a concavity on the middle sideof the surface. It ends before the distal convex elongation, which iscurved behind and medially directed. On the lateral view, there isan elongated frontal socket just under the epiphysis, where theMcIII is joined; above this concavity, on the antero-lateral cornerthere is a horizontal convexity. The proximal-lateral edge of McIIIis sub-rectangular and flat (Fig. 12): it curves and forms a sizeableconvexity directed to the medial surface. Medially, the articularsurface is divided into two sides: frontally, the first one is hori-zontally elongated, stretching out and sloping to the medial plane.Behind, there is a flat and short limit of the surface. Laterally andfrontally, there is only one concavity for the location of the McIV,similar to that described on the previous bone. The diaphysiswidth slightly increases from the proximal to the distal end.Comparing to the other metacarpals, the McIV is slender (Fig. 13),with the exception of IGF5297V, which has a massive andenlarged diaphysis. There is a tuberosity above the distal side ofthe diaphysis, at the radial flexor muscle insertion. The lateral sideof the proximal end is rectilinear or slightly convex. Behind, thereis a deep convexity. The articular surface is sloping and convex,rectangular in shape, ending frontally with a curved large edge.Medially, there is an articular protrusion, with a sinuous trend(concave and convex). On the plantar side, the proximal articu-lation ends whit horizontal steps. For the junctionwithMcIII thereis a sloping surface.
We note the differences in dimensions and robustness for thethree McV (Fig. 14). IGF5309V is longer and larger than the other
two McV. The proximal epiphysis is deeply convex and the anteriorlimit ends with an acute angle. Laterally, it develops a bulge andforms a semilunar convex surface. The diaphysis has a rectangulartransversal section.

Fig. 7. Leopard mandibles from Equi: IGF6122V, IGF6120V, and IGF6103V in labial (A, B, and C) and lingual (A0 , B0 , and C0 , respectively) views; IGF10037V/1 in lingual (D) and labial(D0) views; IGF10037V/2 in labial (E) and lingual (E0) views; IGF6102V in lingual (F) and labial (F0) views; IGF185V/4 in labial (G) and lingual (G0) views; IGF6104V in lingual (H) andlabial (H0) views.
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151138
5.2.3. Hindlimb
5.2.3.1. Femur. Only one femur from an adult animal is keptwithin the historical excavations collections (IGF5347V, found in1918). This bone is broken along the diaphysis and the brokenedge was smoothed after the deposition. Nine more femurs in the
collection from Equi belong to young or sub-young individuals(Fig. 15).
5.2.3.2. Tibia/Fibula. No found of tibias (nor fibulae) from adultanimals are kept within the Equi historical collection with the

Fig. 8. A) right leopard humerus (IGF5342V) with evidence of pectoralis muscle stresson the proximal diaphysis; B) young right humerus of leopard (IGF5341V).
Fig. 9. Left ulna of leopard in lateral view: from an adult animal (IGF5355V) (A) and acub (IGF5354V) without the proximal and distal epiphysis (B); specimen IGF5402V (C)is massive and probably from a male or old animal.
Fig. 10. Radious of leopards: IGF5337V is from a left paw of an adult animal (A) andIGF5340V is a left radious from a younger one (B).
Fig. 11. Left McII (IGF5303V) in dorsal (A), proximal (C e not in scale), and medial (B)views.
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151 139

Fig. 12. Right McIII (IGF5302V) in dorsal (A), proximal (Ce not in scale), and lateral (B)views.
Fig. 13. Right McIV (IGF5293V) in dorsal (A), proximal (Ce not in scale), and lateral (B)views.
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151140
exception of the specimen in exposition (numbered as nn46 inAppendix). IGF5346V (Fig. 16) has a sub-rectangular distaldiaphysis in transversal section and a developed medialmalleolus.
5.2.3.3. Metatarsal. IGF5311V is a MtII (Fig. 17). The proximal end isthinner than the distal one, and its massive diaphysis forms amedial concavity. The proximal articulation is formed by a medialconcavity that is frontally bulged and posteriorly directed to themedial side. Laterally, there are two well marked surfaces, bothconcave and joined to the MtIII.
The MtIII (Fig. 18) has a massive and curved diaphysis. It isfronto-distally flattened and rectilinear. The proximal articulation isjoined to the middle cuneiform bone. Frontally, it is enlarged andcurved. Medially, it forms a brief constriction and a distal bulge.
The MtIV (Fig. 19) has a less curved diaphysis. The proximal endis clearly directed toward the caudal side. The articular area for thelateral cuboid is convex, slightly higher on the distal edge, andfrontally tilted. Like for the McIV, it has a marked convexity for thejunction with the last metatarsus. MtV is well represented byIGF5318V (Fig. 20). Two are probably from the same very younganimal (IGF5325V and IGF5324V): proximal and distal epiphysisare unfused (and consequently smaller size and a different kind offossilization and color). The MtV diaphysis has a triangular sectionand the proximal epiphysis show a lateral bi-lobed edge. Fromthese two lobes, the articular surface is medially inclined. Thelateral junction to MtIV is formed by an enlarged frontal structureand a distal plane.
6. Discussion
6.1. General overview
The site of Equi is a very rich deposit. The site has been known inliterature for almost one century, but fossils and the available in-formation about the original excavations were disconnected andnever considered together. Until a recent re-evaluation analysis ofthe historical collection (Ghezzo et al., 2014b), the context wasvirtually ignored and a comparison with the new research (startedsince 2009) could not have been possible. On the basis of theavailable information from original labels in the Florence museumcollection Ghezzo et al. (2014b) proved that there are not veryremarkable differences in the distribution of large carnivores'anatomical elements through the stratigraphic record. Wolves andleopards were the most numerous animals in the site (notconsidering bear remains) and their distribution increases in theuppermost layers. The occurrence of the same species throughoutthe sequence is probably related to climatic conditions within themiddle to later part of the MIS3.
The lack of axial skeleton and autopodial ends in the collectionseems not to be due to scavenging, but, most likely, it could berelated to the effects of excavators' selection rather than to taph-onomic agents (even though such consideration is partially indisagreement with the presence of well preserved small bones in

Fig. 14. Left McV (IGF5309V) in dorsal (A), proximal (Ce not in scale), and medial (B)views.
Fig. 15. Right femurs of leopards in frontal view: adult (IGF5347V) (A) and juvenile (B:IGF5350V; C: IGF5349V) specimens.
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151 141
the Equi collection, implying a favorable depositional environmentand careful recovery work during the field work in early times).
Few peculiar features of the leopard bones from Equi testify theoccurrence of disease and consumption, not related at the momentto a precise type of carnivores. Usually, bone consumption is relatedto hyena activity, but there is no evidence, at Equi, of hyenafrequentation. Examples of different predator carcasses being sor-ted by hyenas when scavenging is recorded at several open air sites(Bottrop, Perick and Keppler Cave; Diedrich, 2009, 2012), and at thesite of Gran Dolina (level TD10-1) where lion remains were sorteddue to human action (Blasco et al., 2010). These sites were char-acterized by a common preservation pattern for bone accumula-tions, with large predators carcasses damaged most at fore andhind limbs (as a consequence, vertebrae are not destroyed and areusually preserved). The preservation pattern is the opposite at Equi,where entire skull, and fore and hind limbs are well preserved,meanwhile, the axial skeleton is almost completely missing.
Not so far from the Equi deposit, faunal guild from Buca dellaIena was described and dated back to the MIS3 (about 41 ky) byPitti and Tozzi (1971). Layer D of the cave represents a typical hyenaden site with the high frequency of the predator in association toseveral large preys (horses, deers, and aurochs) and few othercarnivores, such as U. spelaeus and Meles meles. As mentionedabove, in the Equi assemblage the differences are notable, both forspecific occurrences and frequencies; few herbivorous are repre-sented and several species of carnivores are recorded, such as lions,wolves, foxes, dholes, and mustelids.
Recently, many studies about leopards as bone collector weremade in comparison to hyenas, and their role as scavengers seem to
have been underestimated (de Ruiter and Berger, 2001; Carlson andPickering, 2003; Pickering and Carlson, 2004; O'Regan and Menter,2009; Pickering et al., 2011; Saoqu�e et al., 2014b), as for dholes andwolves (Fosse et al., 2012; Mallye et al., 2012; Fourvel et al., 2014),even though extant leopards use caves where trees are rare, justavoiding food-competitors and never as a durable den (de Ruiterand Berger, 2001 and references therein). In this case, the pres-ence of many fossils of leopards in opposition to the rare toppredator (lion) could imply a borderline environment. According toseveral authors (Balme et al., 2007; Bhattarai and Kindlmann, 2012;Steinmetz et al., 2013), the largest predators (i.e. lions in Africa andtigers in Asia) occupy the best areas to maximize their hunting andfitness success, where the prey availability represents the mainstayfor the range selection (Lovari et al., 2014). The subordinate pred-ators' ranges (leopards, wolves, and the smaller dholes and foxes)are the results of avoiding the largest carnivorans, and balancingthe disadvantages of prey frequency with less selective habits, theiragility and (not for the former) pack organization. Also, the steepvalley of Fagli was probably considered by leopards in order toavoid other predators (such as hyenas), and bears for hibernation. Alast hypothetical scenario for explain a so high frequency of leopardin the site of Equi was already depicted by Diedrich (2012, 2013) forother European Pleistocene sites, with the opportunity of scavengebears remains.
The sites with several carnivorans species are not infrequentduring the Pleistocene. Argant et al. (2007) found P. onca gombas-zoegensis and Panthera spelaea fossilis in association to Ursusdeningeri and Canis mosbachensis in the site of Chateau Breccia(France). Diedrich (2013) described the younger association in theWildkirchli Cave (Switzerland) and Baumann's Cave (Germany),where Panthera leo spelaea, U. spelaeus, Panthera pardus spelaea and
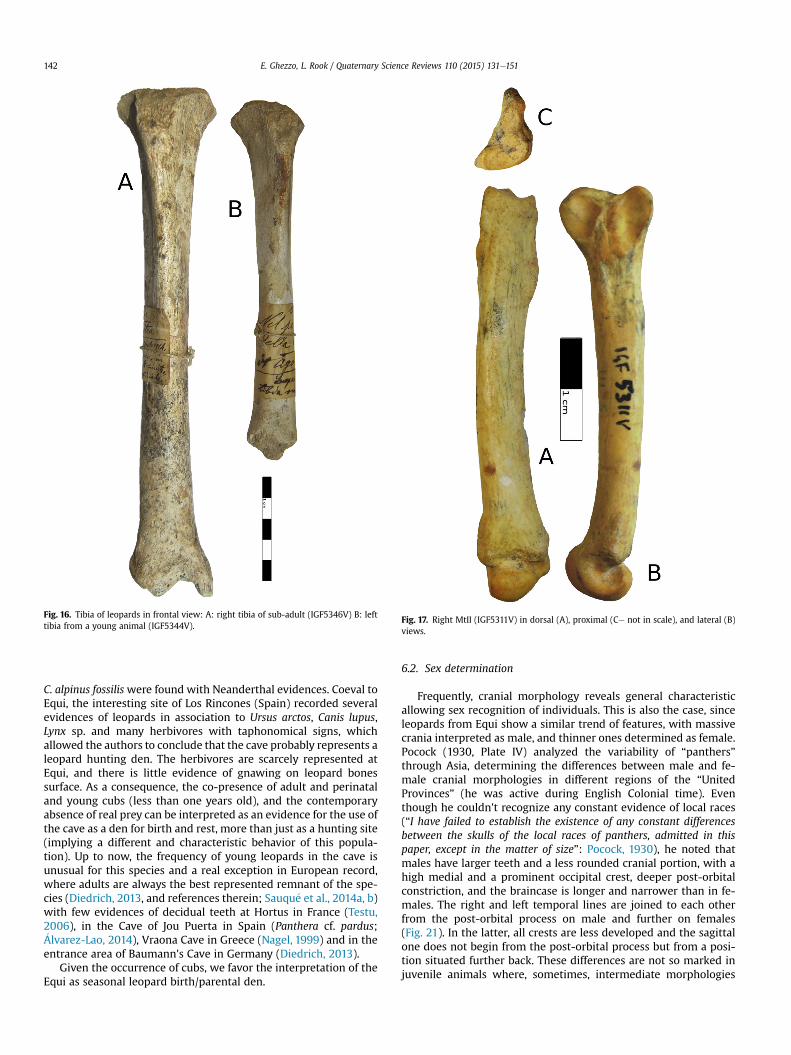
Fig. 16. Tibia of leopards in frontal view: A: right tibia of sub-adult (IGF5346V) B: lefttibia from a young animal (IGF5344V).
Fig. 17. Right MtII (IGF5311V) in dorsal (A), proximal (Ce not in scale), and lateral (B)views.
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151142
C. alpinus fossiliswere found with Neanderthal evidences. Coeval toEqui, the interesting site of Los Rincones (Spain) recorded severalevidences of leopards in association to Ursus arctos, Canis lupus,Lynx sp. and many herbivores with taphonomical signs, whichallowed the authors to conclude that the cave probably represents aleopard hunting den. The herbivores are scarcely represented atEqui, and there is little evidence of gnawing on leopard bonessurface. As a consequence, the co-presence of adult and perinataland young cubs (less than one years old), and the contemporaryabsence of real prey can be interpreted as an evidence for the use ofthe cave as a den for birth and rest, more than just as a hunting site(implying a different and characteristic behavior of this popula-tion). Up to now, the frequency of young leopards in the cave isunusual for this species and a real exception in European record,where adults are always the best represented remnant of the spe-cies (Diedrich, 2013, and references therein; Sauqu�e et al., 2014a, b)with few evidences of decidual teeth at Hortus in France (Testu,2006), in the Cave of Jou Puerta in Spain (Panthera cf. pardus;�Alvarez-Lao, 2014), Vraona Cave in Greece (Nagel, 1999) and in theentrance area of Baumann's Cave in Germany (Diedrich, 2013).
Given the occurrence of cubs, we favor the interpretation of theEqui as seasonal leopard birth/parental den.
6.2. Sex determination
Frequently, cranial morphology reveals general characteristicallowing sex recognition of individuals. This is also the case, sinceleopards from Equi show a similar trend of features, with massivecrania interpreted as male, and thinner ones determined as female.Pocock (1930, Plate IV) analyzed the variability of “panthers”through Asia, determining the differences between male and fe-male cranial morphologies in different regions of the “UnitedProvinces” (he was active during English Colonial time). Eventhough he couldn't recognize any constant evidence of local races(“I have failed to establish the existence of any constant differencesbetween the skulls of the local races of panthers, admitted in thispaper, except in the matter of size”: Pocock, 1930), he noted thatmales have larger teeth and a less rounded cranial portion, with ahigh medial and a prominent occipital crest, deeper post-orbitalconstriction, and the braincase is longer and narrower than in fe-males. The right and left temporal lines are joined to each otherfrom the post-orbital process on male and further on females(Fig. 21). In the latter, all crests are less developed and the sagittalone does not begin from the post-orbital process but from a posi-tion situated further back. These differences are not so marked injuvenile animals where, sometimes, intermediate morphologies

Fig. 18. Right MtIII (IGF5313V) in dorsal (A), proximal (Ce not in scale), and lateral (B)views.
Fig. 19. Right MtIV (IGF5292V) in dorsal (A), proximal (Ce not in scale), and lateral (B)views.
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151 143
are shown. Finally, Pocock supposes that the skulls of both sexestend to flatten along the top as age advances, becoming lower andlonger than in youngeadult individuals. Sunquist and Sunquist(2002) add that, in general, males' are larger than females' of thesame regions, often reaching a 30e50% heavier body mass.
On the basis of Pocock's (1930) analysis, a sex determination ispossible for the Equi fossil material. Crania IGF10036V and IGF184Vhave masculine features, while the frailer (with under developedsagittal crests) IGF15111V/1 and IGF185V/1 seem to be betterattributable to female individuals. IGF15110V cranium has sub-spherical outline of the neurocranium, undeveloped sagittal crestand long temporal lines, but the bone appears massive as a male, sowe cannot rule out that this specimen represents a very old female.According, the skulls from Allekoaitze and Vjeternica Cave areconsidered a female and a male, respectively (Corral, 2012;Miculini�c, 2012).
Diedrich (2013) tried to infer the two genders from the size oftheir crania. A similar conclusion was possible for lions (Turner,1984) and could be considered true within a well-defined popula-tion, but it is not possible to compare cranial remains from farterritories and long geological times, without considering climaticoscillations and the typical morphological and size flexibility of thisspecies. In the previous study of carnivorans from Equi, Del
Campana (1954) reported that total lengths of males and femalesof modern leopards from different regions (South Africa, Congo,eastern Africa, and central India), respectively 178e247 and205e218 mm, showing a quite extensive dimensional overlapping.The shorter skulls from Equi, IGF15111V/1 and IGF15110V arewithinthe smallest male size range, and IGF185V/1 is the most massiveand largest skull, besides the upper limit of length for modernleopards: a result exactly opposite to the morphological conclusion.
Anyway, the general rule that males are larger than femalescould be considered valid, but the range and size variability of theseanimals don't allow per se inferring the genders, and theirmorphology should be preferable to size analysis for sex determi-nation (Fig. 22).
6.3. Body size
Dimensionally speaking, the crania from Equi fall within therange of extant leopards. They are generally massive, and all largerthan extant subspecies for the palatal and neurocranium width(Fig. 23). Comparative material of jaguars (P. onca) and cougars(Puma concolor) are intermediate in size among P. pardus and lions,and within the leopard-lynx lower limit respectively. Leopardsfrom Equi are quite larger than the conspecific extant animals. It isalso notable that all leopard crania from Equi had a shorter muzzle

Fig. 20. Right MtV (IGF5318V) in medial (A), proximal (Ce not in scale), and lateral (B)views. Distal end is broken by gnawing.
Fig. 21. Female and male comparative morphologies of leopard crania (from Pocock,1930; modified): major differences are on temporal lines junctions, the developmentof the sagittal crest, the feature of the braincase, more or less evident postorbital
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151144
in comparison to extant relatives, even though the check teeth arealways 2.8 times longer (in average) than the carnassial for both thespecies.
As mentioned above for leopards, a strongest overlapping ofcranial condilobasal lengths in the two genders is also found in thesimilar middle-sized felids P. concolor, varying between about 110and less than 200 (Giannini et al., 2010). The skulls of American andEuropean cougars are shorter than leopards, being rounded both inthe braincase and splancnocranial regions.
Upper teeth reveal a clear separation of leopards from lynx inthe LP4/C-M1 ratio (Fig. 24A). The specimens from Equi are set onthe lower limit of variability and match with the leopard fromPortugal from the Late Pleistocene and with P. pardus from Angola(specimen 8748 from Natural History Museum of Florence). Asconsequence, and again, the muzzles of the leopard from Equi canbe considered quite short. The split is so evident also for lower teethfollowing the same pattern: the extant leopard set is partiallyoverlying the lynx's, while specimens from Equi are larger and closeto the coeval leopard tooth fromGrotta del Capriolo (Pitti and Tozzi,1971) (Fig. 24B).
The sizes of upper carnassials from Equi are within the leopard'srange. The Villafranchian European cougar shows smaller uppercarnassial, recording dimensions between 20 and 23.2 mm (Cherinet al., 2013), and shorter lower check teeth (Fig. 24B), meanwhilejaguar carnassials are larger, falling closer to lion size.
Unfortunately, information about the original position of eachcranium in the stratigraphy of Equi is completely missing. Instead,the lower m1 distribution seems to have a distributional pattern.IGF10037V/1, IGF6104V, and IGF10037V/2 are larger thanIGF6102V, IGF6103V and IGF15111V/2 and both are well grouped intwo sets. Anyway, this distribution is not justified by the strati-graphic provenance: the information were reported only for
IGF6104V, IGF6102V, which were found just below the surface inthe shelter and cave respectively, and for IGF6103V, which is older(layer hb in Ghezzo et al., 2014b).
The m1L/m1B ratio diagram (Fig. 25) show that the three mostrepresentative sites of Los Rincones, Vraona and Equi, have com-parable trendlines but the latter has clearly smaller teeth. Thisevidence must be considered with the shorter muzzles and largepalate of Equi leopards, concluding a local variability of the popu-lation. At the same time, P. concolor is well defined by its shorterand slender lower carnassial and P. onca gombaszoegensis is quitelarger than Late Pleistocene leopards.
Postcranial bones are more fragmented and there are only oneentire humerus and radius (Figs. 8 and 10). The bones are welldiscerned from lions and the humerus is included into the extantleopard variability for the trochlear width (Fig. 26A). The largesthumerus, IGF5342V, is also the older one (layer b, just above thesecond hearth; cfr. Ghezzo et al., 2014b) and its size is betterexplainable with sex dimorphism or wide intraspecific variability.As we can see in Fig. 10, the size of radius varies in relation toontogeny and intraspecific variability: it is an individual-relatedcharacter. The length patterns of metacarpals and metatarsals forthe species of big cats allow us to well-define the limit betweenlions and leopards, for extant and for the fossil materials too. Lynxesmetapodials tend to be longer than leopards ones, but theweight ofdiaphysis is larger for the latter in comparison to the slenderer lynxand consequently they cannot be confused (Fig. 27). There are not atrend among leopards from Middle to Late Pleistocene and extantanimals, and an overlap between this species and cougar (bothP. concolor and Puma pardoides) can be observed at least for theMcIV.
Nagel (1999) noted a particularly robustness of feet bones ofleopard from Vraona Cave. Compared to Equi, the specimens seemeffectively larger, but the absolute ranges have not a statisticalsignificance (the variance between the two collections is about1e2 mm in metapodials both for minimum and maximum SD).
6.4. Specific determination and morphological comparison
Morphology is quite variable in leopards (Table 1) and thedimensional analysis must also be taken into account for a bettergrounded determination.
constriction.

Fig. 22. Total length vs. condilus width scatter diagram of fossils and extant species: black dots are from extant animals from different regions (a single specimen has a declaredgender); the gray dot is the Monte Sacro Middle Pleistocene cranium (M.P.U.R./V.1191: Kotsakis and Palombo, 1979); the gray-black dot is the cranium from Algar do Manga Larga(Cardoso and Regala, 2006); white dots are fossils crania described by Diedrich (2013); square, triangle and diamond represent the crania from Equi. Genders were defined on thebasis of morphological characters.
Fig. 23. Basion-prostion vs. palatal weight (A) and euryoneeuryon (B). In comparison to extant leopards, the fossils from Equi are larger, stoutly built and massive, and clearlydistinct from other species of felids.
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151 145

Fig. 24. Dental comparison among upper (A) and lower (B) teeth of leopards from Equi (black squares), extant (white circles) and extinct leopards (Arago e white squares, un-published data; Grotta del Capriolo e gray square, Pitti and Tozzi, 1971; Manga Larga e gray-black circle, Cardoso and Regala, 2006), extant and extinct lions (white triangles) andlynxes (white rhombus), and cougar (gray diamonds).
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151146
Leopards are smaller than lions and the teeth are slender.Compared to P. pardus, themandibles of P. onca gombaszoegensis arehigher and the canine is larger. The lower carnassial is also massiveand the two tips are closer to each other (Spassov and Raychev,1997; Hemmer et al., 2001). Leopards are quite larger than Lynxlynx, and in this case both the morphology and dimension allowingcorrect determination.
Spassov and Raychev (1997), Nagel (1999), Diedrich (2013) andSaoqu�e et al. (2014b) described the morphological affinities of theLate Pleistocene leopards to Panthera uncia. We excluded the snowleopard for determining the specimens from Equi because thecrania clearly diverge both for size and morphology from P. uncia.The most visible difference between the extant snow leopard andAfrican leopard is the very concave dorsal edge of the shortsplancnocranium in the former. In the mandible, the symphysis isquite vertical and the diastema shorter. The shortness of snowleopard diastema doesn't fit with modern leopard and the speci-mens from Equi are within the variability of the latter (Fig. 28). Thesame trend is shown for almost all the Middle and Late Pleistocenespecimens include in this analysis. On the contrary, the diastema ofthe mandible from Str�ask�a Sk�al�a is very short and was firstlydetermined as snow leopard by Thenius (1969) and included toP. pardus by Hemmer (1971).
Diedrich (2013) resumed the evolutive trends of Europeanleopards identifying the most ancient animal with elongated skulls,medium-wide frontals and low-crowned P4. The youngest animalsassumed and a progressive change of these characters, with a shortand enlarged skull and developed cusps of P4. The features of in-dividual variability offered by the leopard cranial record from Equipoint out a different scenario, given that the variability observed inliterature can be easily explain with intraspecific (intrapopula-tional) variability. The cranial length is variable as much as thefrontal widths, and P4 crown height varies in relation to thewearing stage. IGF15110V is very similar to the cranium from Algarda Manga Larga Cave (Portugal) (Cardoso and Regala, 2006) with aquite short muzzle, a barrel-like braincase and a large postorbitalconstriction. They differ for the sunken-in feature of the dorsal endof the nasals but this particular features is also notable on the skullfrom Allekoaitze Cave (Spain) (Corral, 2012). This last characterrepresents a plesiomorphic feature, being visible also on skulls oflions, tigers and leopards (Ghezzo and Bergamo, submitted forpublication). Otherwise, the skull from Monte Sacro (Rome) haslong splancno and neurocranium and a developed sagittal crest(Kotsakis and Palombo, 1979). IGF10036V is similar to such skull,showing a comparable development of neurocranial side andsagittal crest, but the splancnocranium is verticalized and the

Fig. 25. Length and breadth of lower carnassials and respective trendlines of the main leopard collections, namely Equi, Los Rincones and Vraona Cave. The lines are quite parallel toeach other and precisely specify for the shape of leopards m1s.
Fig. 26. Postcranial comparison among different species of Felidae. A: humerus(IGF5342V) B: radius (IGF5339V); total length (Gl) and distal breadth (Bd) ratios.
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151 147
muzzle results shorter (Fig. 5). In this specimen, the postorbitalconstriction and the frontals are quite narrowed, showing moreaffinities to the modern specimen from the Horn of Africa (from theVenice Museum of Natural History) but unlike the presumed trendfor Late Pleistocene European leopards.
As mentioned above, the presence of P2 is quite variable in theskulls from Equi, while they are present at the cave of L'Observ-atoire and Monte Sacro, lacking instead in the specimens fromLunel-Viel (Bonifay, 1971; Kotsakis and Palombo, 1979) (Table 1).
Two dental morphotypes were recognized by Spassov andRaychev (1997) for the lower p4-m1 and we noted that the leop-ards from Equi are ascribable to the “gracile type” (Fig. 29) with aconcave lingual edge of the carnassial and especially for the mesio-distal symmetry of p4. The mesial side of these teeth are equallydeveloped than the distal one, giving a rectangular shape to thetooth. The modern comparative specimen from Angola (MZF8748)shows a triangular p3 and a narrow m1, implying a not definitivedifferentiation between the two groups in modern animals. Theauthors identified the robust type as belonging to archaic animalsand the gracile one to the Late Pleistocene leopards. In fact, themandible from Mauer, identified as P. pardus sickenbergi by Schütt(1969) has a clear triangular p3 and a shorter and rectilinearlingual edge of the m1 (Fig. 7 in Schütt, 1969).
Mandible IGF10037V/2 and IGF6103V show a little obliquesymphysis. The p4/m1 ratio of the specimens fromEqui (85e93%) isout of the variability of Triagalnata (98%) (Spassov and Raychev,1997); in modern leopards it ranges between 84 up to 101%,including both the studied collections. Spassov and Raychev (1997)stressed the shortness feature of the Triagalnata and Vraona dia-stema and effectively they are shorter than those from Equi(13e16 mm, n.7), and even of modern leopards (13-20.4, n.12).Again, diastema is 12e12.7 mm in P. concolor and 22 mm (n ¼ 3) inPanthera onca gombaszoegenesis from V�ertessz}ol}os II and extantjaguar (considered together), so the two species results discerniblefrom leopards for this measurement.
Lower teeth are embricated, the horizontal ramus is high, bothunder the third premolar as well as the last molar, and the lower

Fig. 27. Metacarpals (A) and metatarsal (B). Comparative dimensional range of Greatest length (Gl) in Holocene and Late Pleistocene big cats.
Fig. 28. Diastema length of different species of Felidae. Modern leopards are partiallyincluded to lion's variability but are well-divided to P. uncia. The same is generally forthe leopard fossils with the exception of the short mandible of Triagalnata and theproblematic Str�ask�a Sk�ala specimen.
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151148
edge is linear and straight. Madurell-Malapeira et al. (2010) andTestu et al. (2011) stated that usually there are two mental fossae inleopards and three in P. uncia and P. concolor. In the specimens fromEqui, IGF10037V/1 (and perhaps the broken IGF6104V) has twoforamina. All the other ones have three nervous insertions, like themandible of P. pardus from Arag�o (Testu et al., 2011) and P. pardusfrom Los Rincones (Ri10/C1/2010) (Sauqu�e and Cuenca-Besc�os,2013).
The m1B/m1L ratio for the specimens from Equi is between 28and 40, really under the ratio expressed for the other Late Pleis-tocene European leopards (Vraona and Los Rincones), and inter-preted as a local variation of the species.
Postcranial bones of leopards are rare in the fossil record. Thespecimens from Equi are quite well preserved but most are fromjuvenile individuals, with the diaphysis unfused. Nagel (1999)noted the robustness of leopard feet bone from Vraona Cave anddetermined the whole collection as P. pardus vraonensis, aiming toemphasize the differences of this particular population, waiting formore exhaustive comparisons. As mentioned above, the compari-son between these specimens and Equi reveals a massive featurefor the former, but the Gl/SD ratio for postcranial is quite similar inboth the collections. In this case, it should be useful a large andmore complete comparison among fossils and modern speciesbefore to infer conclusions about a real differentiation.
To conclude, as proven for other large felidae taxa, such asHomotherium latidens (Galobart et al., 2003) and cave lion (Burgeret al., 2004; Sotnikova and Nickolskiy, 2006), specific and subspe-cific determination are often a consequence of the rarity of finds,the distance among sites and chronology, or are due to differentauthors' points of view, more than being a real and realisticdivergence among two or more evolutive branches.

Fig. 29. Comparison between the types recognized by Spassov and Raychev (1997) and the p4-m1 from Equi. 1) robust type; 2) gracile type. Equi teeth: IGF185V/3 (A); mirroredIGF6102V (B); IGF6103V (C); mirrored IGF10037V/1 (D); IGF10037V/2 (E). Black lines show the differences between the two types.
E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151 149
7. Conclusions
Leopards combine the power and strength of the larger catswith the grace and versatility of the smaller felinae.
For the first time from its discovery at the beginning of 19thcentury, the study of leopard collection from Equi allows for thedefinition of the morphological characterization, of a quitecontemporaneous population (mid to late MIS3) within a singularcontext. Surprisingly, leopards from Equi were characterized by agreat size variability both for cranial and limbs, maintaining just afew constant characters, such as the lower edge of mandibles, thelack of a real metaconide on m1, and the furrow at the nasaljunction.
We revealed the frequency of different age stages and both sexesand proved that morphological variability is more closely related togender differences than to subspecific determination.
The presence of cubs is a very interesting occurrence, never seenin other European Late Pleistocene sites with this high frequency,and with this study we proved, also for European leopards, thatcaves were being used for births in addition to rest and possiblybear hunting or scavenging, and not a fortuitous frequentation nora stable hunting station.
Sex dimorphism was proven by cranial features more thandental morphology or size, namely: the development of the sagittalcrest and its junction with temporal lines, the features of thebraincase, and the width of the postorbital constriction.
Leopards and smaller carnivores probably engaged in a mutualavoidance, while the presence of several bear remains implies thatthe cave was likely used for different reasons in different seasonsand times.
Direct radiocarbon dating confirms the previous chronologicalattribution to the MIS3 and the previous study about leopard bonedistribution through the stratigraphical record, and determined theEqui collection to be one of the latest examples of a leopard largepopulation in Europe just before the climate decreasing trend of theLGM.
Finally, thanks to the Equi collection, we can affirm that the sizeand morphologies of rare European leopards can be attributed tointraspecific variability and that the subspecific determinations arenot provable for the European paleontological record. Even thoughwe don't know the precise position of cranial finds into the strati-graphic record of Equi, they are undeniably younger than 44 ky(probably at about 33e27 ky ago according to the recent datings).The cranial and postcranial features include almost all the
variability of European specimens, almost for the Late Pleistocene,and we can admit that the p4-m1 morphology could be preferredfor the subspecific determination.
Accordingly, we identify the Equi leopards belonging to P. pardusspelaea, considering the peculiar morphological features as localvariability within the larger geographical range of the Late Pleis-tocene species.
Acknowledgments
For the access to paleontological and osteological collections weare very grateful to the curators of the Natural History Museum ofthe University of Florence, Dr. Elisabetta Cioppi (Geological andPaleontological section) and Dr. Paolo Agnelli (Zoology section “LaSpecola”), the Natural History Museum of Berlin, Dr. Oliver Hampe(Vertebrate Paleontology section) and the Hungarian Natural His-tory Museum, Prof. Mih�aly Gasparik (Department of Palaeontologyand Geology), as well as to to Dr. P. Boscato (Siena) and Prof. B. Sala(Ferrara). The Vertebrate Palaeontology Laboratory of the Univer-sity of Florence is acknowledged for specimens preparation andrestoration.
The revision of the paleontological collection from the Equi cave,is framed within a wider project aimed to the re-evaluation andvalorisation of the historical Paleontological collections of theMuseum of Natural History, University of Florence, partially sup-ported by the University of Florence grants (Fondi di Ateneo, to LR).EG received support from a DAAD grant (The German AcademicExchange Service, award codenumber: A/13/71401), and from theSYNTHESYS Project http://www.synthesys.info/(project numberHU-TAF-4000), which is financed by European CommunityResearch Infrastructure Action under the FP7 “Capacities” Program.
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.quascirev.2014.12.020.
References
�Alvarez-Lao, D.J., 2014. The Jou Puerta cave (Asturias, NW Spain): a MIS3 largemammal assemblage with mixture of cold and temperate elements. Palae-ogeogr. Palaeoclimatol. Palaeoecol. 393, 1e19.
Ambrosi, A.C., Fabbri, M., 1975. I santuari “d’Abri” nelle Apuane e i livelli medievalidella Tecchia di Equi (Prov. di Massa Carrara). Archeol. Mediev. 2, 367e377.
Argant, A., Argant, J., Jeannet, M., Erbajeva, M., 2007. The big cats of the fossil siteChateau Breccia Northern Section (Saone-et-Loire, Burgundy, France):

E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151150
stratigraphy, palaeoenvironment, ethology and biochronological dating. CourierForschungsinstitut Senckenberg 259, 121e140.
B€achler, E., 1936. Das Wildkirchli. Eine Monographie. Buchdruckerei H. Tschudy,p. 254.
Balme, G., Hunter, L., Slotow, R., 2007. Feeding habitat selection by hunting leopardsPanthera pardus in a woodland savanna: prey catchability versus abundance.Anim. Behav. 74, 589e598.
Barone, R., 2003a. In: Anatomia comparata dei mammiferi domestici. osteologia,vol. 1. Edagricole (ed.).
Barone, R., 2003b. In: Anatomia comparata dei mammiferi domestici. splancnologia.Apparecchio digerente e respiratorio, vol. 3. Edagricole (ed.).
Barroso, C., Riquelme, J., Moigne, A.M., Banes, L., 2003. Las faunas de grandesmamíferos del Pleistoceno Superior de la cueva del Boquete de Zafarraya.Estudio paleontol�ogico y paleoecol�ogico. El pleistoceno Superior de la cueva delBoquete da Zafarraya. Arqueol. Monogr. 15, 169e222.
Bartolini, S., Cioppi, E., Rook, L., Delfino, M., 2014. Late Pleistocene fossils and thefuture distribution of Rana temporaria along the Apennine Peninsula (Italy).Zool. Stud. 53, 76. http://dx.doi.org/10.1186/s40555-014-0076-5.
Baryshnikov, G.F., 2011. Pleistocene Felidae (Mammalia, Carnivora) from the KudaroPaleolithic cave sites in the Caucasus. Proc. Zool. Inst. RAS 315 (3), 197e226.
Battaglia, R., 1932. Note su alcune industrie paleolitiche della cerchia alpina e dellealpi apuane. Atti Accad. Veneto-Trentino-Istriana 22, 93e105.
Bhattarai, B.P., Kindlmann, P., 2012. Interaction between Bengal tiger (Pantheratigris) and leopard (Panthera pardus): implication for their conservation. Bio-divers. Conserv. 21, 2075e2094.
Bigagli, C., Iardella, R., Palchetti, A., Paribeni, E., 2013. Fivizzano (Ms). Equi Terme, saggionella Grotta della Tecchia. Not. Soprintend. Beni Archeol. Toscana 8, 187e193.
Blasco, R., Rosell, J., Arsuaga, J.L., Bermúdes de Castro, J.M., Carbonell, E., 2010. Thehunted hunter: the capture of a lion (Panthera leo fossilis) at the Gran Dolinasite, Sierra de Atapuerca, Spain. J. Archaeol. Sci. 37, 2051e2060.
Bonifay, M.-F., 1971. Carnivores quaternaires du sud-est de la France. In: M�emoiresdu Mus�eum national d’Histoire Naturelle, S�eries C, p. 339.
Branchini, I., 1928. Esame tipologico dei manufatti della Grotta d'Equi (Lunigiana).Arch. Antropol. Etnol. 58, 48e63.
Braschi, S., Del Freo, P., Trevisan, L., 1986. Ricostruzione degli antichi ghiacciai sulleAlpi Apuane. Atti Soc. Tosc. Sci. Nat. Mem. Serie A 93, 203e219.
Burger, J., Rosendahl, W., Loreille, O., Hemmer, H., Eriksson, T., G€otherstr€om, A.,Hiller, J., Collins, M.J., Wess, T., Alt, K.W., 2004. Molecular phylogeny of theextinct cave lion Panthera leo spelaea. Mol. Phylogenet. Evol. 30, 841e849.
Caloi, L., Palombo, M.R., 1994. Le faune a grandi mammiferi del Pleistocene supe-riore dell’Italia centrale: biostratigrafia e paleoambiente. Boll. Serv. Geol. Ital.111, 77e104.
Cardoso, J.L., Regala, F.T., 2006. O Leopardo, Panthera pardus (L., 1758), do Algar daManga Larga (Planalto de Santo Ant�onio, Porto de M�os). Comun. Geol. 119e144.
Carlson, K.J., Pickering, T.R., 2003. Intrinsic qualities of primate bones as predictorsof skeletal element representation in modern and fossil carnivore feeding as-semblages. J. Hum. Evol. 44, 431e450.
Cassoli, P.F., Tagliacozzo, A., 1991. Considerazioni paleontologiche, paleoecologiche earcheozoologiche sui macromammiferi e gli uccelli dei livelli del PleistoceneSuperiore del Riparo di Fumane (VR) (Scavi 1988-1991). Boll. del Mus. Civ. StoriaNat. Verona 18, 349e446.
Cherin, M., Iurino, D.A., Sardella, R., 2013. Earliest occurrence of Puma pardoides(Owen, 1846) (Carnivora, Felidae) at the Plio/Pleistocene transition in westernEurope: new evidence from the Middle Villafranchian assemblage of Montopoli,Italy. C. R. Pal�evol 12, 165e171.
Corral, J.C., 2012. T�ecnicas aplicadas en la preparaci�on de un cr�aneo cuaternario dePanthera pardus (Linneo, 1758) de Ataun (cueva Allekoaitze, Guipúzcoa,Espa~na). Bol. Geol. Miniero 123 (2), 127e138.
Cuvier, G., 1835. Recherches sur les ossemens fossiles ou l'on r�etablit les caract�eresde plusieurs animaux dont les r�evolutions du globe ont d�etruit les esp�eces, vol.4. G. Dufour et E. d'Ocagne, Paris.
de Ruiter, D.J., Berger, L.R., 2001. Leopards as taphonomic agents in dolomitic caves-implications for bone accumulations in the hominid-bearing deposits of southAfrica. J. Archaeol. Sci. 27, 665e684.
De Stefani, C., 1917. La Grotta preistorica di Equi nelle Alpi Apuane. Archi. Antropol.Etnol. 46, 41e82.
Del Campana, D., 1923. Sopra un Cuon e una Mustela del Quaternario di Equi (ApliApuane). Regia Accademia Nazionale dei Lincei. Classe di Scienze Fisiche,Matematiche e Naturali. Rendiconti Ser. 5 (32), 170e172.
Del Campana, D., 1954. Carnivori quaternari della Tecchia e della Caverna di Equinelle Alpi Apuane (Mustelidi, Canidi, Felidi). Palaeontogr. Ital. 44, 1e42.
Diedrich, C.G., 2009. Steppe lion remains imported by Ice Age spotted hyenas intothe Late Pleistocene Perick Caves hyena den in northern Germany. Quat. Res. 71,361e374.
Diedrich, C.G., 2012. Cave bear killers and scavengers from the last ice age of centralEurope: feeding specializations in response to the absence of mammoth steppefauna from mountainous regions. Quat. Int. 255, 59e78.
Diedrich, C.G., 2013. Late Pleistocene leopards across Europe e northernmost Eu-ropean German population, highest elevated records in the Swiss Alps, com-plete skeletons in the Bosnia Herzegowina Dinarids and comparison to the IceAge cave art. Quat. Sci. Rev. 76, 167e193. http://dx.doi.org/10.1016/j.quascirev.2013.05.009.
Fejfar, O., Kaiser, T.M., 2005. Insect bone-modification and paleoecology of Oligo-cene mammal-bearing sites in the Doupov Mountains, Northwestern Bohemia.
Palaeontol. Electron. 8 (1), 8A:11p, 1MB. http://palaeo-electronica.org/paleo/2005_1/fejfar8/issue1_05.htm.
Fischer, K., 2000. Ein Leoparden-Fund, Panthera pardus (L., 1758), aus dem jung-pleistoz€anen Rixdorfer Horizont von Berlin und die Verbreitung del Leopardenim Pleistoz€an Europas. Mitt. Mus. Nat.kd. Berl. Geowiss. Reihe 3, 221e227.
Fisher Jr., J.W., 1995. Bone surface modifications in zooarchaeology. J. Archaeol.Method Theory 2 (1), 7e68.
Fletcher, W., S�anchez-Go~ni, M.F., Allen, J., Cheddadi, R., Combourieu-Nebout, N.,Huntley, B., Lawson, I., Londiex, L., Magri, D., Margari, V., Müller, U.,Maughton, F., Novenko, E., Roucroux, K., Tzedakis, P., 2010. Millennial-scalevariability during the last glacial in vegetation records from Europe. Quat. Sci.Rev. 29, 2839e2864.
Fosse, P., Nuria, S., Smietana, W., Okarma, H., Wajrak, A., Fourvel, J.B., Madelaine, S.,Montserrat, E.-N., C�aceres, I., Yravedra, J., Brugal, J.P., Prucca, A., Haynes, G.,2012. Bone modification by modern wolf (Canis lupus): a taphonomic studyfrom their natural feeding places. J. Taphon. 10 (3e4), 197e217.
Fourvel, J.-B., Fosse, P., Brugal, J.-B., Cregut-Bonnoure, E., Slimak, L., Tournepiche, J.-F., 2014. Characterization of bear remains consumption by Pleistocene largecarnivores (Felidae, Hyaenidae, Canidae). Quat. Int. 339e340, 232e244.
Fraipont, C., 1923. Crane de Panth�ere ou de Lynx g�eant provenent de la caverne deTrois-Fr�eres (Ari�ege). Rev. Anthropol. 33, 42.
Galobart, �A., Pons-Moy�a, J., Ant�on, M., Maroto, J., 2003. Descripci�on del material deHomotherium latidens (Owen) de los yacimientos del Pleistoceno inferior deIncarcal (Girona, NE de la Península Ib�erica). Paleontol. i Evol. 99e141.
Gavashelishvili, A., Lukarevskiy, V., 2008. Modelling the habitat requirements ofleopard Panthera pardus in west and central Asia. J. Appl. Ecol. 45, 579e588.
Ghezzo, E., Bergamo, A., Critical overview about cranial variability of European andAfrican lions (Panthera leo ssp.). Ann. Paleontol. submitted for publication.
Ghezzo, E., Rook, L., 2014. Cuon alpinus (Pallas, 1811) (Mammalia, Carnivora) fromEqui (Late Pleistocene, Massa-Carrara, Italy): anatomical analysis and paleo-ethological contextualisation. Rendiconti Lincei 25, 491e504. http://dx.doi.org/10.1007/s12210-014-0345-6.
Ghezzo, E., Bert�e, D.F., Sala, B., 2014a. The revaluation of Galerian Canidae, Felidaeand Mustelidae of the Cer�e Cave (Verona, Northeastern Italy). Quat. Int.239e240, 76e89. http://dx.doi.org/10.1016/j.quaint.2012.12.031.
Ghezzo, E., Palchetti, A., Rook, L., 2014b. Recovering data from historical collections:stratigraphic and spatial reconstruction of the outstanding carnivoran recordfrom the Late Pleistocene Equi cave (Apuane Alps, Italy). Quat. Sci. Rev. 96,168e179. http://dx.doi.org/10.1016/j.quascirev.2014.03.012.
Giannini, N.P., Segura, V., Giannini, M.I., Flores, D., 2010. A quantitative approach tothe cranial ontogeny of the puma. Mamm. Biol. 75, 547e554.
Giraudi, C., 2004. The Appennine glaciations in Italy. In: Ehlers, J., Gibbard, P.L.(Eds.), Quaternary Glaciations-extent and Chronology. Part I: Europe. Elsevier,Amsterdam, pp. 215e224.
Giraudi, C., 2011. Middle Pleistocene to Holocene glaciations in the Italian Apen-nines. In: Ehlers, J., Gibbard, P.I., Hughes, P.D. (Eds.), AA: VV: Quaternary Gla-ciations e Extent and Chronoly. A Closer Look, Developments in QuaternarySciences Series Editor: Van der Meer J. J.M, vol. 15, pp. 211e219 (chapter 17).
Graziosi, P., 1934. Esplorazione paletnologica del territorio etrusco. Studi Etruschi 8,307e313.
Guerri, M., 1980. Equi Terme. Riv. Sci. Preist. 35, 360e362.Guerri, M., 1982. Grotta di Equi Terme (Prov. Massa Carrara). Riv. Sci. Preist. 37,
305e306.Hank�o, E.P., 2007. A revision of three Pleistocene subspecies of Panthera, based on
mandible and teeth remains, stored in Hungarian collections. Fragm. Palae-ontol. Hung. 24e25, 25e43.
Harris, J.M., Timm, R.M., 2013. Paleoecological and taphonomic implications ofinsect-damaged Pleistocene vertebrate remains from Rancho La Brea, SouthernCalifornia. PLoS ONE 8 (7), e67119. http://dx.doi.org/10.1371/journal.pone.0067119.
Harrison, S.P., Prentice, I.C., 2003. Climate and CO2 controls on global vegetationdistribution at the last glacial maximum: analysis based on palaeovegetationdata, biome modelling and palaeoclimate simulations. Glob. Change Biol. 9,983e1004.
Hayward, M.W., Henschel, P., O'Brien, J., Hofmeyr, M., Balme, G., Herley, G.I.H., 2006.Prey preferences of the leopard (Panthera pardus). J. Zool. 270, 298e313. http://dx.doi.org/10.1111/j.1469-7998.2006.00139.x.
Hemmer, H., 1971. Zur kenntnis pleistozaner mitteleuropaischer Leoparden(Panthera pardus). Neues Jahrb. Geol. Pal€aontol. Abh. 138 (1), 15e36.
Hemmer, H., Kahlke, R.-D., Vekua, A.K., 2001. The jaguar, Panthera oncagombaszoegensis (Kretzoi, 1938) (Carnivora: Felidae) in the late LowerPleistocene of Akhalkalaki (South Georgia; Transcaucasia) and its evolutionaryand geological significance. Geobios 34 (4), 475e786.
Henschel, P., Hunter, L., Breitenmoser, U., Purchase, N., Packer, C., Khorozyan, I.,Bauer, H., Marker, L., Sogbohossou, E., Breitenmoser-Wursten, C., 2008. Pantherapardus. The IUCN Red List of Threatened Species. Version 2014.3. www.iucnredlist.org. Downloaded on 08 December 2014.
Hillson, S., 1996. Teeth. The University of Cambridge Press.Holden, A.R., Harris, J.M., Timm, R.M., 2013. Paleoecological and Taphonomic Im-
plications of Insect-damaged Pleistocene Vertebrate Remains from Rancho LaBrea, Southern California. PLoS ONE 8 (7), e67119. http://dx.doi.org/10.1371/journal.pone.0067119.
Hugues, P.P., Woodward, J.C., 2008. Timing of glaciation in the Mediterraneanmountains during the last cold stage. J. Quat. Sci. 23, 575e588.

E. Ghezzo, L. Rook / Quaternary Science Reviews 110 (2015) 131e151 151
Iardella, R., Palchetti, A., Paribeni, E., 2011. 1909-2009: 100 anni di ricerche allaTecchia di Equi Terme (MS). In: 150 anni di Preistoria e Protostoria in Italia.Volume dei Riassunti della XLVI Riunione Scientifica dell’Istituto Italiano diPreistoria e Protostoria, pp. 51e52.
Janossy, D., 1986. Pleistocene Vertebrate Faunas of Hungary. In: Developments inPalaeontology and Stratigraphy, vol. 8. Elsevier, Amsterdam, p. 208.
Kotsakis, T., Palombo, M.R., 1979. Un cranio di Panthera pardus (L.) del Pleistocenemedio-superiore di monte sacro (Roma). Geol. Romana 18, 137e155.
Lei, W., XiaoBing, W., LiXin, Z., ZhiGang, J., 2011. Mitogenomic analysis of the genusPanthera. Sci. China 54 (10), 917e930.
Linnaeus, C., 1766. Systema naturae sive regna tria naturae, secundum classes,ordines, genera, species, cum characteribus, differentiis, synonymis, locis, XIIed. Laurentii Salvii, Holmiae.
Linnæus, C., 1758. Systema naturæ per regna tria naturæ, secundum classes,ordines, genera, species, cum characteribus, differentiis, synonymis, locis.Tomus I (in Latin). Laurentius Salvius, Holmiæ (Stockholm).
Lovari, S., Pokheral, C.P., Jnawali, S.R., Fusani, L., Ferretti, F., 2014. Coexistence of thetiger and the common leopard in a prey-rich area: the role of prey partitioning.J. Zool. http://dx.doi.org/10.1111/jzo.12192.
Lyman, R.L., 1994. Vertebrate Taphonomy. Cambridge University Press.Madurell-Malapeira, J., Alba, D.M., Moy�a-Sol�a, S., Aurell-Garrido, J., 2010. The Ibe-
rian record of the puma-like cat Puma pardoides (Owen, 1846) (Carnivora,Felidae). C. R. Palevol 9, 55e62.
Mallye, J.-B., Costamagno, S., Boudadi-Maligne, M., Prucca, A., Lauroulandie, V.,Thi�ebaut, C., Mourre, V., 2012. Dhole (Cuon alpinus) as a bone accumulator andnew taphonomic agent? the case of Noisetier Cave (French Pyrenees). J. Taphon.10 (3e4), 318e346.
Mangerud, J., Ehlers, J., Gibbard, P. (Eds.), 2004. Quaternary Glaciations: Extent andChronology 1: Part I Europe. Elsevier, Amsterdam.
Marciszak, A., Krajcarz, M.T., Krajcarz, M., Stefaniak, K., 2011. The first record ofleopard Panthera pardus Linnaeus, 1758 from the Pleistocene of Poland. Actazool. crac. 54A (1e2), 39e46.
Miculini�c, K., 2012. Fossil remains of leopard (Panthera pardus) from VjetrenicaCave, Popovo polje, BiH (PhD thesis). Zagreb University.
Miththapala, S., Seifensticker, J., O'Brien, S.J., 1996. Phylogeographic subspeciesrecognition in leopards (Panthera pardus): Molecular genetic variation. Conserv.Biol. 10 (4), 1115e1132.
Mochi, A., 1916. Sull'industria preistorica della Grotta di Equi. Arch. Antropol. Etnol.46, 210e212.
Mochi, A., 1920. Contro la contemporaneit�a dell’Orso speleo con l'Uomo neolitico.Replica al Prof. De. Stefani. Arch. Antropol. Etnol. 49, 143e153.
Mochi, A., 1929. Osservazioni alla memoria del Prof. R. Battaglia intitolata “Il Pale-olitico Superiore in Italia. Bull. Paletnol. Ital. 49, 1e11.
Moulle, P.-E., Echassoux, A., Lacobat, F., Desclaux, E., Bailon, S., 2005. L'environne-ment animal des premiers habitants de l'Europe m�editerrann�eenne: les grandsmammif�eres contemporains de l'homme du Vallonnet, donn�ees taxonomiqueset biostratigraphiques pour la deuxi�eme moiti�e du Pl�eostoc�ene inf�erieur. In:Molines, N., Moncel, M.-H., Monnier, J.L. (Eds.), Les premiers peuplements enEurope. Colloque international: donn�ees r�ecentes sur les modalit�es depeuplement et sur le cadre chronostratigraphique, g�eologique etpal�eog�eographique des industries du Pal�eolithique ancien et moyen en Europe(Rennes, 22-25 sept. 2003), John and Hedge, Oxford, BAR Intern. Series S1364,pp. 105e113.
Nagel, D., 1999. Panthera pardus vraonensis n.ssp a new leopard from the Pleistoceneof Vraona. Greece. Neues Jahrb. Geol. Pal€aontol. Monatsh. 3, 119e150.
O'Regan, H.J., Menter, C.G., 2009. Carnivora from the Plio-Pleistocene hominin siteof Drimolen, Gauteng, South Africa. G�eobios 42, 329e350.
Paribeni, E., Iardella, R., Palchetti, A., 2009. Fivizzano (MS). Ricerche alla Tecchia diEqui Terme. Not. Soprintend. Beni Archeol. Toscana 5, 138e140.
Pasa, A., 1947. I mammiferi di alcune antiche brecce veronesi. In: Memorie delMuseo Civico di Storia Naturale di Verona, 1, pp. 1e111.
Pickering, T.R., Carlson, K.J., 2004. Baboon taphonomy and its relevance to theinvestigation of large felid involvement in human forensic cases. Forensic Sci.Int. 144, 37e44.
Pickering, T.R., Heaton, J.L., Zwodeski, S.E., Kuman, K., 2011. Taphonomy of bonesfrom Baboons killed and eaten by wild leopards in Mapungubwe National Park,South Africa. J. Taphon. 9 (2), 117e159.
Pitti, C., Tozzi, C., 1971. La Grotta del Capriolo e la Buca della Iena presso Mommio(Camaiore, Lucca). Riv. Sci. Preist. 26, 213e258.
Pocock, R.I., 1930. The panthers and ounces of Asia. Volume II. J. Bombay Nat. Hist.Soc. 34, 63e82, 307e336.
Regalia, O.E., 1911. Fauna glaciale in due grotte di Equi (Alpi Apuane). Riv. Ital.Paleontol. 17, 24e28.
Rellini, U., 1917. La Grotta preistorica di Equi nelle Alpi Apuane. Bull. Paletnol. Ital.42, 67e74.
Rellini, U., 1924. Appunti sul Paleolitico Italiano-Oggetti delle caverne di Equi e diTenerano nelle Alpi Apuane. Bull. Paletnol. Ital. 44, 1e34.
Rellini, U., 1929. Nota sulle “Osservazioni alla Memoria del Prof. R. Battaglia...” diAldobr. Mochi. Bull. Paletnol. Ital. 49, 98e101.
Rellini, U., 1935. La ceramica della caverna di Equi. Bull. Paletnol. Ital. 55, 67e69.Rellini, U., 1937. La stirpe di Neanderthal nel Lazio. Bull. Paletnol. Ital. 1, 5e56.Rosenmüller, J.C., 1794. Quedam de ossibus fossilibus animalis cuiusdam, historiam
eius et cognitionem accuratiorem illustrantia, dissertatio, quam d. 22. Octob.1794 ad disputandum proposuit Ioannes Christ. Rosenmuller Hesberga-Francus,LL.AA.M. In: Theatro anatomico Lipsiensi Prosector assumto socio Io. Chr. Aug.Heinroth Lips. Med. Stud. Cum tabula aenea. Leipzig, pp. 1e34.
Saoqu�e, V., RabalGarc�es, R., Sola-Almagro, C., Cuenca-Besc�os, G., 2014a. Boneaccumulation by leopards in the Late Pleistocene in the Moncayo Massif (Zar-agoza, NE Spain). PlosOne 9 (3), e92144. http://dx.doi.org/10.1371/journal.pone.0092144.
Saoqu�e, V., Rabal-Garc�es, R., Cuenca-Besc�os, G., 2014b. Carnivores from Los Rin-cones, a leopard den in the highest mountain of the Iberian range (Moncayo,Zaragoza, Spain). Hist. Biol. Int. J. Paleobiol. 1e28. http://dx.doi.org/10.1080/08912963.2014.977882.
Sauqu�e, V., Cuenca-Besc�os, G., 2013. The Iberian Peninsula, the last Europeanrefugium of Panthera pardus Linnaeus 1758 during the Upper Pleistocene.Quaternaire 24 (1), 35e48.
Schütt, Von G., 1969. Panthera pardus sickenbergi n. subsp. Aus den Mauerer Sanden.Neues Jahrbuch für Geologie und Pal€aontologie. Monatshefte Begründet 1807,pp. 299e310.
Siddall, M., Rohling, E.J., Thompson, W.G., Waelbroeck, C., 2008. MIS 3 sea-levelfluctuations: data synthesis and new out look. Rev. Geophys. 46, RG4003.http://dx.doi.org/10.1029/2007RG000226.
Sotnikova, M., Nickolskiy, P., 2006. Systematic position of the cave lion Pantheraspelaea (Goldfuss) basedon cranial andd ental characters. Quat. Int. 142e143,218e228. http://dx.doi.org/10.1016/j.quaint.2005.03.019.
Spassov, N., Raychev, D., 1997. Late Wurm Panthera pardus remains from Bulgaria:the European fossil leopards and the question of the probable species survivaluntil the Holocene on the Balkans. Hist. Nat. Bulg. 7, 71e96.
Steinmetz, R., Seuaturien, N., Chutipong, W., 2013. Tigers, leopards, and dholes in ahalf-empty forest: assessing species interactions in a guild of threatened car-nivores. Biol. Conserv. 163, 68e78. http://dx.doi.org/10.1016/j.biocon.2012.12.016.
Sunquist, M., Sunquist, F., 2002. Wild Cats of the World. The University of Chicagopress e Chicago and London.
Testu, A., 2006. Etude pal�eontologique et biostratigraphique des Felisae et Hyaenidaepl�eistoc�enes de l’Europem�editerran�eenne (PhD thesys). Universite de Perpignan.
Testu, A., Moigne, A.-M., de Lumley, H., 2011. La panth�ere Panthera pardus desniveaux inf�erieurs de la Caune de l’Arago �a Tautavel (Pyr�en�ees-orientales,France) dans le contexte des Felidae (Felidae, Pantherinae) de taille moyenne duPl�eistoc�ene Europ�een. Quaternaire 4, 271e281.
Thenius, E., 1969. Uber das Vorkomen fossiler Schneeleoparden (Subgenus Uncia,Carnivora, Mammalia). Saugetierkundl. Mitt. 17, 234e242.
Turner, A., 1984. Dental sex dimorphism in European lions (Panthera leo L.) of theUpper Pleistocene: palaeoecological and palaeoethological implications. Ann.Zool. Fenn. 21, 1e8.
Uphyrkina, O., Johnson, W., Quigley, H., Miquelle, D., Marker, L., Bush, M.,O'Brien, S.J., 2001. Phylogenetics, genome diversity and origin of modernleopard, Panthera pardus. Mol. Ecol. 10, 2617e2633.
von der Driesch, A., 1976. A Guide to the Measurement of Animal Bones fromAcrchaeological Sites. Peabody Museum of Archaeology and Ethmology, Har-vard University, p. 358.
Werdelin, L., Peign�e, S., 2010. Carnivora. In: Werdelin, L., Sanders, W.J. (Eds.),Cenozoic Mammals of Africa. University of California Press, pp. 603e657.





























