Embed Size (px)
Citation preview

Plant and Soil 254: 155–165, 2003.© 2003 Kluwer Academic Publishers. Printed in the Netherlands.
155
The regulation of nodulation, nitrogen fixation and ammoniumassimilation under a carbohydrate shortage stress in the Discariatrinervis-Frankia symbiosis
Claudio Valverde & Luis Gabriel Wall1Programa de Investigacion sobre Interacciones Biologicas, Departamento de Ciencia y Tecnologia, UniversidadNacional de Quilmes, Roque Saenz Peña 180, Bernal, B1876BXD, Argentina. 1Corresponding author∗
Received 19 July 2002. Accepted in revised form 20 August 2002
Key words: dark stress, Discaria trinervis, Frankia, nitrogenase, nitrogen fixation, nodules
Abstract
N2-fixation is sensitive to limitation in the availability of newly synthesised carbohydrates for the nodules. We de-cided to explore the response of the D. trinervis - Frankia symbiosis to a transient decrease in carbohydrate supplyto nodules. Feedback inhibition of nodulation as well as nodule growth was not released by a 6-day dark stressin D. trinervis nodulated plants. However, nitrogen fixation and assimilation were affected by the imposed stress.Nitrogenase activity was totally inhibited after 4 days of darkness although high levels of nitrogenase componentswere still detected at this time. Degradation of FeMo and Fe nitrogenase subunits – both at similar rates – wasobserved after 6 days of dark stress, revealing the need for inactivation to precede enhancement of protein turnover.Glutamine synthetase (GS), malate dehydrogenase (MDH) and asparagine synthetase (AS) polypeptides were alsodegraded during the dark stress, although at a lower rate than nitrogenase. ARA and nitrogenase were totallyrecovered 8 days after resuming normal illumination. It seems that current nitrogenase activity and ammoniumassimilation are not, or are only weakly linked with the feedback control of nodulation in D. trinervis. Theseobservations give support to the persistence of an autoregulatory signal in mature nodules that is not sensitive totransient shortages of carbon supply and sustains the inhibition of nodulation in the transient absence of N2 fixation.
Abbreviations: AS – asparagine synthetase; GS – glutamine synthetase MDH – malate dehydrogenase
Introduction
Actinorhizal plants make use of ammonium that issymbiotically fixed within root nodules that havebeen induced and colonised by filamentous bacteriafrom the genus Frankia (Wall, 2000). The processof nitrogen fixation is highly dependent on an ad-equate energy supply and low O2 pressure. Theformer is thought to be accomplished by shuttlingof dicarboxylic acids from the host cell cytoplasmto Frankia cells (Huss-Danell, 1997). The latter taskinvolves several strategies that include a variableFrankia lipidic cell envelope, expression of haemo-
∗ FAX No: +54-11-4365-7182. e-mail: [email protected]
globin like protein, variable respiration rates and reg-ulation of O2 diffusion through nodule intercellularspaces (Huss-Danell, 1997). Nodules receive carbo-hydrates in the form of phloem delivered sucrose.Sucrose is split into monomers by sucrose synthaseor invertase activities, thus providing substrates fordicarboxylic acid synthesis and assimilation of fixedammonia into amides and/or ureides (Vance, 2000). Inthis way, the metabolisms of carbon and nitrogen areintertwined.
The formation of root nodules is regulated by hostmechanisms that control the location and growth ofnodules. Rapid autoregulation of nodulation has beendescribed in Alnus incana and Discaria trinervis, amechanism that systemically down-regulates infection

156
a few days after nodulation has been initiated in theinoculated root zone, before the onset of nitrogen fix-ation (Wall, 2000). An additional mechanism operateswhen nodules are mature and functional; root infec-tion as well as growth of nodules are regulated bythe N content of the shoot (Valverde et al., 2000).However, some subtle differences in regulation of nod-ulation have been observed between A. incana and D.trinervis. While the excision of nodules results in newnodule formation in both plants, the elimination of N2fixing activity by incubation of nodulated roots in anAr:O2 atmosphere resulted in new nodule formationonly in A. incana (Valverde, 2000; Wall, 2000). Theseparate evolutionary origin of these two symbiosesmay account for the different regulatory responses. A.incana (root hair-infected host) belongs to the cladeI of actinorhizal plants, as proposed by Swensenand Mullin (1997), while D. trinervis (intercellularinfected host) belongs to clade Ill.
Nitrogenase activity is sensitive to a wide rangeof perturbations (including cold stress, drought stress,defoliation or dettoping, a high supply of ammoniumand flooding), most probably as a consequence of re-duced availability of newly synthesised carbohydratesin the nodules (Huss-Danell, 1997). In Alnus glu-tinosa, a primary effect of a 1-hour darkening was a60% reduction of sucrose supply to nodules (Wheeler,1971). Storage carbohydrates in the root nodules aremainly unavailable for nitrogen fixation (Wheeler,1971). In A. incana, a darkening treatment of 4 daysinduced degradation of nitrogenase proteins and anirreversible loss of structure of Frankia vesicles (Lun-dquist and Huss-Danell, 1991a; Vikman et al., 1990).The reduction of photosynthesis during such stresstreatment was associated with the recovery of rootsusceptibility for infection by Frankia and nodule de-velopment (Wall and Huss-Danell, 1996). Altogether,these observations suggested that current nitrogenaseactivity would be involved in the feedback regulationof nodulation in mature nodules. In this paper, we ex-plored for the Discaria trinervis – Frankia symbiosis,whether treatments that cause a temporal decrease ofcarbohydrate supply result in perturbation of feedbackregulation of nodulation. We also analysed the timecourse of activity and protein levels of enzymes in-volved in N2 fixation and assimilation (nitrogenasecomponents, glutamine synthetase (GS), malate de-hydrogenase (MDH) and asparagine synthetase (AS))during and after a transient dark stress.
Materials and methods
Plant growth, Frankia growth and inoculation
Seeds of Discaria trinervis (Hooker et Arnot) Reichewere surface-sterilised and germinated as previouslydescribed (Valverde and Wall, 1999a). Seedlings at thecotyledonary stage (12–14 d after the start of germin-ation), were aseptically transferred to growth pouches(Mega International, Minneapolis, USA). Each pouchwith four seedlings contained 12.5 mL of nutrientsolution diluted to 1/10 of full strength without min-eral nitrogen (Huss-Danell, 1978). Pouches were keptin the greenhouse (Universidad Nacional de Quilmes,Bernal, Argentina; 34◦ 7′ S, 58◦ 3′ W) where meanmaximum temperature was 24 ◦C, mean minimumtemperature was 20 ◦C and relative humidity ranged50 – 95%. Incandescent lamps (400 W, Osram, Brazil)supplemented natural light such that the photoperiodwas of 16 h.
Frankia strain BCU110501 (Chaia, 1998) wasused as inoculum. Bacteria were grown in static BAPminimal medium with 55 mM of glucose as C source(Chaia, 1998), at 28 ◦C for 4 weeks. Cells were har-vested by centrifugation (1000 g, 5 min), washed withnutrient solution without N, and resuspended in asmall volume (3 – 4 mL) of the same solution. Thecell suspension was homogenised by repeated passagethrough needles, first with 0.8 mm gauge and then with0.5 mm gauge (three times each). Frankia biomassin the homogenate was estimated by determination ofpacked cell volume (Nittayajarn and Baker, 1989).
Each seedling was inoculated by dripping 200µL of inoculum containing 5 µL of packed Frankiacells, from the root tip to the uppermost zone of theroot. Pouches were subsequently watered with nutrientsolution without N. About 2–3 weeks following inocu-lation the bottom edge of the pouches was cut off andpouches were placed all together into a separate biggerplastic bag. Nutrient solution was added to a level ofabout 1-cm and was refilled as necessary or renewedat least once a week. The pouches took solution by ca-pillarity through the supporting paper. Non-inoculatedplants never formed nodules.
Treatments to reduce carbohydrate supply to nodules.Recording of new nodules
In a first experiment, the effect of reduction of leaf areaor light intensity on nodulation was assessed on 18weeks-old plants that were growing with N2 fixation

157
as the only source of N. The nodulation status of theseseedlings at the time the treatments started was 16.47nodules per plant (SE = 0.48, n =211). Pre-existingnodules were marked with a water-proof marker penon the plastic surface of the pouch in order to be able toidentify new nodules at the end of the experiment. Thewhole nodulated root system was re-inoculated with2.5 µL of Frankia BCU110501 cells. Groups of rein-oculated plants (n = 20) were treated as follows: (A)control plants remained untreated; (B) the lower halfof leaves was removed; (C) the upper half of leaveswas removed; (D) all leaves were removed; (E) shootswere partially shaded with a mesh that reduced lightintensity to ca. 20% during 6 days; (F) shoots weretotally shaded with a black cloth during 6 days. Nineweeks after treatments were applied, nodulation (ex-amination of the root system in the pouch with nakedeye) and growth (dry matter) were measured.
In a second experiment, the effect of different de-grees of light intensity reduction on nodulation wasassessed on 13 weeks-old nodulated plants with anodulation status similar as the previous experiment.Pre-existing nodules were marked with a water-proofmarker pen on the plastic surface of the pouch in or-der to be able to identify new nodules at the end ofthe experiment. Groups of plants (n = 12) were re-inoculated with 10 µL of Frankia BCU110501 cells,and immediately placed under shading cages that re-duce the intensity of light to 100% (control), 54%.26%, 12%, 8% and 0.3% of normal light conditions,for a period of 20 days. Temperature was not af-fected with this treatment. Control plants were leftuncovered. New nodulation status of the plants wasexamined 6 weeks after restoration to normal light.
In order to analyse the response of nitrogenasecomponents to dark stress, 14 weeks-old nodulatedplants were placed under a black cloth that reduced thelight intensity to less than 0.5% of normal light con-ditions. Then, nitrogenase activity was estimated asacetylene reducing activity periodically after the startof the dark treatment. After measuring nitrogenaseactivity, nodules were harvested and total soluble pro-teins were extracted. The activities of GS and MDHwere measured in the crude nodule extracts, and the re-lative amounts of GS, MDH and AS polypeptides werestudied by Western blotting. Nitrogenase componentswere immunodetected in Frankia extracts from thesedimented fraction of crushed nodules
Acethylene reducing activity
Nitrogenase activity was estimated as acetylene re-duction activity (ARA) of intact plants in pouches(Valverde et al. 2000). Each pouch was incubated atroom temperature (24 ◦C) in a gas-tight cuvette withan available volume of ca. 800 mL. The plants werekept in light (controls) or dark (treatments) during in-cubations. Pouches were cut open along their sides tofacilitate gas diffusion, and shoots were gently bent tofit into the cuvette. C2H2 was injected to a final con-centration of 5% (v/v). A gas sample of 0.5 mL wastaken 30–45 min after C2H2 injection, and analysedfor C2H4 production in a Konik KNK-3000-HRGCgas chromatograph with a Porapak-Q80/100 column(2 m long, 1/8′′ diameter) run at 35 ◦C. Injection cham-ber temperature was 110 ◦C. Carrier gas was N2 at aflow rate of 38 mL per minute. The flame ionisationdetector was operated with air at a flow rate of 300 mLper minute and H2 at a flow rate of 45 mL per minute,at a temperature of 170 ◦C. Acetylene reducing activ-ity is expressed as nmoles of ethylene produced perhour and per mg of nodule fresh weight.
Protein extraction from nodule host cells
Root nodules (45 – 130 mg fresh weight) were separ-ated from plants used for ARA estimation and kepton ice in polypropylene microtubes until extractionof proteins. Nodules were ground on ice manually in700 µL of extraction buffer (100 mM N-morpholinomethane sulphonate (MES), 15% v/v ethyleneglycol,2% v/v 2-mercaptoethanol, 100 mM sucrose, 1 mMphenylmethylsulfonylfluoride (PMSF), pH 6.8) with aTeflon pestle (Egli et al. 1989). Nodule homogenateswere centrifuged at 25 000 g for 20 min. The super-natant was transferred to new microtubes and used forassaying GS and MDH activities, and protein content.The rest of the nodule cell cytoplasm extract was im-mediately frozen at 86 ◦C. The pellet containing hostcell debris, Frankia filaments and vesicle clusters weresaved for extraction of Frankia proteins.
Protein extraction from cultured Frankia cells andvesicle clusters
Frankia BCU110501 cells grown in liquid me-dium were collected by centrifugation in microtubes(25 000 g for 10 min). Cell fresh weight was estimatedby weighing pellets. Cells were washed with homo-genization buffer (50 mM Tris-HCI, 1 mM EDTA,

158
2.5% w/v SDS, pH 8.0) and resuspended in an-other 500 µL of homogenization buffer (Lundquistand Huss-Danell, 1991 a). A preparation of FrankiaBCU110501 vesicle clusters was obtained from ef-fective D. trinervis nodules by differential filtration ofnodule homogenates (Akkermans et al. 1981). Plants(20 weeks old) were growing with N2 as the onlysource of N. A nodule mass of 1.8 g fresh weightwas placed in a Petri dish on ice and chopped with ascalpel blade. Five ml of cold distilled water were ad-ded to resuspend cell debris, and immediately passedfirst through a 100-µM nylon mesh, then through a 20-µM nylon mesh. The material remaining in the 20-µMmesh was washed with cold distilled water, resuspen-ded in 1 mL of distilled water and transferred to apolypropylene microtube. The quality of the vesiclecluster suspension was checked by light microscopy.Vesicle clusters were sedimented (500 g for 5 min),washed with homogenization buffer (50 mM TrisHCl,1 mM EDTA, 2.5% w/v SDS, pH 8.0) and resuspendedin 500 µL of homogenization buffer.
Frankia cell suspension, vesicle clusters suspen-sion and pellets from nodule homogenates were keptat 100 ◦C for 5 min with occasional vortexing, andthen sonicated (Fisher Scientific F60 sonifier) with atip for 30 s at an output of 3–5 watts (three pulses witha 30 s interval in between). The sonicated suspensionswere centrifuged for 10 min at 25 000 g, and the super-natant (protein extract) stored at −86 ◦C (Lundquistand Huss-Danell, 1991 a).
Protein determination
Protein content in nodule host cell extracts was es-timated by the Bradford protein assay adapted forELISA microplates using BioRad� Protein reagentas indicated by the manufacturer. Bovine serum albu-min (BSA, Sigma) was used as standard. The contentof protein in Frankia culture extracts, vesicle clusterextracts and nodule Frankia extracts were determinedwith the bicinchoninic acid protein assay (Smith et al.1985). Standard BSA (Sigma) solutions were preparedin homogenization buffer.
Enzyme assays
Glutamine synthetase (GS) activity was determinedby the ADP-dependent transferase assay, which wasscaled down and adapted to a microplate (Romanovet al. 1998). Five µL of nodule extract was added to100 µL of reaction mixture (20 mM Tris.acetate, 8.75
mM hydroxylamine, 1 mM EDTA, 2.25 mM MnCI2,17.5 mM NaH2AsO4, 2.75 mM ADP, 35 mM glutam-ine, pH 6.4) and incubated at 37 ◦C for 3–5 h. Reactionceased after adding 100 µL of ferric reagent (3.2%w/v FeCl3, 4% w/v TCA, 0.5 N HCl). γ -glutamylhydroxamate production was measured in an ELISAmicroplate reader at 530 nm. The GS unit was definedas the amount of enzyme that catalyses the productionof 1µmol γ -glutamyl hydroxamate per min.
Malate dehydrogenase (MDH) activity was meas-ured in nodule extracts by monitoring disappearanceof NADH at 340 nm (Egli et al. 1989). The assaybuffer contained 100 mM bicine (pH 8.5), 3 mM ox-alacetate and 0.16 mM NADH. Ten µL of sample wereadded to initiate reaction in 1 mL of reaction mixture,at room temperature (24 ◦C). One MDH unit was con-sidered as the amount of enzyme that catalyses thereduction of 1 µmol of oxalacetate per min.
Western blot analysis of PAGE separated proteins
SDS-PAGE was performed according to the proced-ure of Laemmli (1970). For immunodetection of ni-trogenase components, samples mixed with loadingbuffer (20 µL) containing 50 µg (dinitrogenase) or20 µg (dinitrogenase reductase) were loaded onto anEC12O Minivertical Gel System (F-C Apparatus Co.,USA). The gels were 0.75 mm thick and had a totalacrylamide concentration of 4% in the stacking zoneand 10% in the separating zone. Electrophoresis wasperformed at 20 mA, constant current, at room tem-perature until the tracking dye reached the bottomof the gels. Gels, Whatman filter papers and PVDFmembrane (Immobilon P, Millipore) were incubated inblotting buffer (25 mM Tris, 192 mM glycine, 20% v/vmethanol, pH 8.3) for 15 min. Polypeptides were elec-troblotted (Burnette, 1981) onto PVDF membranes inan EC14O Mini Blot Module (E-C Apparatus Co.,USA) for 90 min at 4 ◦C and 250 mA constant current.The voltage was about 40–45 V. Electrophoretic sep-aration of polypeptides from nodules host cell extractsfor immunodetection of GS, MDH and asparaginesynthetase (AS) proteins was performed as above de-scribed, except that the amount of protein loaded was20 µg for GS, 10 µg for MDH and 50 µg for AS.
Immunochemical detection of proteins was per-formed essentially as described by Blake et al. (1984).PVDF membranes were incubated for 60 min at 37 ◦Cin a blocking solution containing 5% w/v skim milk(Molico) in TBS (50 mM Tris. HCl, 150 mM NaCl,pH 8.0). The membranes were incubated overnight

159
Table 1. Defoliation or transient shading did not affect the feedback regulation over nodulation in Discariatrinervis. Values are means ± SE. Different superscripts denote significant statistical difference at the 5%level (ANOVA)
Treatment Control 50% upper 50% lower 100% 80% 99.7%
leaves leaves leaves shading shading
removed removed removed (6 days) (6 days)
Plant
biomass (mg 224 ± 17a 218 ± 23a 240 ± 22a 153 ± 15b 214 ± 18a 250 ± 21a
DM)
New nodules 0.2 ± 0.1a 0.0 ± 0.0a 0.1 ± 0.1a 0.0 ± 0.0a 0.0 ± 0.0a 0.2 ± 0.2a
Number of
plants with 3 0 1 0 0 1
new nodules
Proportion of
nodule tissue 5.0 ± 0.4ac 4.4 ± 0.3ab 4.5 ± 0.2a 3.6 ± 0.2b 4.6 ± 0.2a 5.5 ± 0.5c
(% DM)
Number of 18 20 20 20 19 20
plants
at 4 ◦C with primary antisera diluted 1:2000 (anti-dinitrogenase), 1:5000 (anti-dinitrogenase reductase),1:5000 (anti-GS), 1:1000 (anti-MDH) and 1:4000(anti-AS), in 5% w/v skim milk-0.05% v/v Tween 20in TBS. After first incubation with antibodies, mem-branes were rinsed 3 × 2 min with 5% w/v skim milk-0.05% v/v Tween 20 in TBS. The secondary antibody,goat anti-rabbit immunoglobulin G (IgG) conjugatedwith alkaline phosphatase (Dako NS, Denmark) wasdiluted 1:1000 with 5% w/v skim milk-0.05% v/vTween 20 in TBS and incubated with membranes atroom temperature for 2.5 h. Membranes were rinsed3 × 2 minutes with 0.05% v/v Tween 20 in TBSand once for 2 min with alkaline phosphatase buffer(100 mM Tris, 100 mM NaCI, 5 mM MgCl2 pH 9.5).Blots were stained with a BCIP and NBT phosphatasesubstrates (Sigma).
The primary antisera against Azotobacter vine-landii dinitrogenase and the ‘universal antiserum mix-ture’ against Rhodospirillum rubrum and Azotobactervinelandii dinitrogenase reductases were a kind giftfrom Dr. P.W. Ludden (University of Winsconsin,USA). The primary antisera against cytosolic MDHisoform and AS were a kind gift of Dr. C.P. Vance(University of Minnesota, USA). The primary anti-serum against GS was a kind gift of Dr. T. Gordon(Institute of Grassland and Environmental Research,UK).
For molecular mass determinations, low molecularmass markers (14.3–66 kDa; Sigma) or high molecu-lar weight markers (30–200 kDa) were included in thegels. After electroblotting, the PVDF membrane stripcontaining the markers was stained with CoomasieBlue R. Dry and stained blots were digitalised in a HP-Scanjet 6200C scanner. The relative intensity of bandswas quantified through Image Pro-Plus v4.0 software.
Results
A transient reduction in carbohydrate supply tonodules does not release the feedback regulation ofnodulation in Discaria trinervis
Treatments were applied to nodulated D. trinervisplants to reduce the supply of photosynthates to nod-ules. Photosynthetic leaf area was reduced by partialor total defoliation, which also reduced the N demandof the shoot. Alternatively, partial or total shadingreduced incident radiation for 6 days. Changes in nod-ulation status was analysed 9 weeks after treatments.None of these treatments resulted in new nodulationat all (Table 1), although root systems had been re-inoculated just before treatments were applied. Theinoculum was infective on control seedlings, whichdeveloped an average of 22.9 nodules (SE ± 2.0).
160
Table 2. Long term reduction of incident radiation (20 days) did not infIuence the feedback regulation ofnodulation in Discaria trinervis. Values are means ± SE. Different superscripts denote significant statisticaldifference at the 5% level (ANOVA)
Residual 100% 54% 26% 12% 8% 0.3%
light
intensity
Plant 204 ± 24ac 233 ± 16c 184 ± 17abc 179 ± 27abc 142 ± 16b 162 ± 20ab
biomass
(mg DM)
New 4.3 ± 1.8a 0.8 ± 0.5ab 2.3 ± 1.2ab 0.5 ± 0.3b 7.9 ± 1.8c 2.0 ± 1.3ab
nodules
Number 15 12 12 13 13 11
of plants
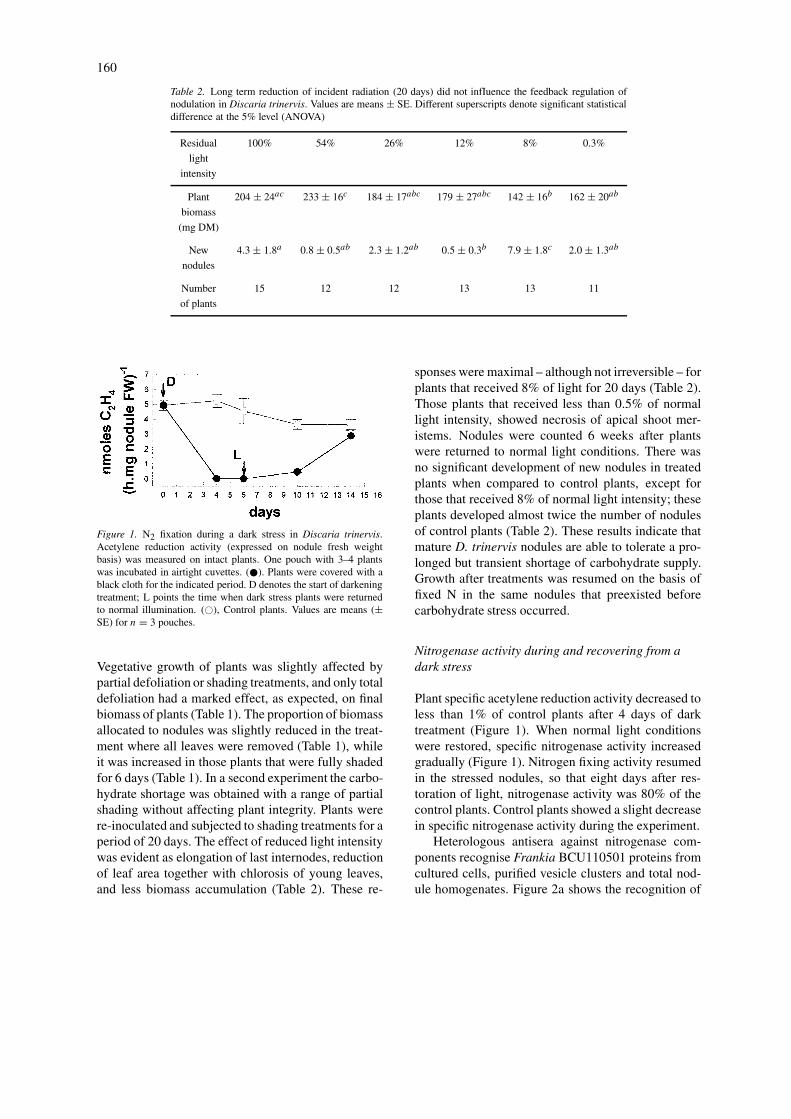
Figure 1. N2 fixation during a dark stress in Discaria trinervis.Acetylene reduction activity (expressed on nodule fresh weightbasis) was measured on intact plants. One pouch with 3–4 plantswas incubated in airtight cuvettes. (�). Plants were covered with ablack cloth for the indicated period. D denotes the start of darkeningtreatment; L points the time when dark stress plants were returnedto normal illumination. (�), Control plants. Values are means (±SE) for n = 3 pouches.
Vegetative growth of plants was slightly affected bypartial defoliation or shading treatments, and only totaldefoliation had a marked effect, as expected, on finalbiomass of plants (Table 1). The proportion of biomassallocated to nodules was slightly reduced in the treat-ment where all leaves were removed (Table 1), whileit was increased in those plants that were fully shadedfor 6 days (Table 1). In a second experiment the carbo-hydrate shortage was obtained with a range of partialshading without affecting plant integrity. Plants werere-inoculated and subjected to shading treatments for aperiod of 20 days. The effect of reduced light intensitywas evident as elongation of last internodes, reductionof leaf area together with chlorosis of young leaves,and less biomass accumulation (Table 2). These re-
sponses were maximal – although not irreversible – forplants that received 8% of light for 20 days (Table 2).Those plants that received less than 0.5% of normallight intensity, showed necrosis of apical shoot mer-istems. Nodules were counted 6 weeks after plantswere returned to normal light conditions. There wasno significant development of new nodules in treatedplants when compared to control plants, except forthose that received 8% of normal light intensity; theseplants developed almost twice the number of nodulesof control plants (Table 2). These results indicate thatmature D. trinervis nodules are able to tolerate a pro-longed but transient shortage of carbohydrate supply.Growth after treatments was resumed on the basis offixed N in the same nodules that preexisted beforecarbohydrate stress occurred.
Nitrogenase activity during and recovering from adark stress
Plant specific acetylene reduction activity decreased toless than 1% of control plants after 4 days of darktreatment (Figure 1). When normal light conditionswere restored, specific nitrogenase activity increasedgradually (Figure 1). Nitrogen fixing activity resumedin the stressed nodules, so that eight days after res-toration of light, nitrogenase activity was 80% of thecontrol plants. Control plants showed a slight decreasein specific nitrogenase activity during the experiment.
Heterologous antisera against nitrogenase com-ponents recognise Frankia BCU110501 proteins fromcultured cells, purified vesicle clusters and total nod-ule homogenates. Figure 2a shows the recognition of

161
Figure 2. Recognition of Frankia BCU11050I nitrogenase compon-ents by heterologous antisera. Western blots probed with a primaryantiserum against Azotobacter vinelandii dinitrogenase (Panel A)and a mixture of primary antisera against Rhodospirillum rub-rum and Azotobacter vinelandii dinitrogenase reductases. Proteinsamples (20 µg): (1) cultured Frankia BCU110501; (2) a sonicatedhomogenate of Discaria trinervis nodules infected with FrankiaBCU110501; (3) Frankia BCU110501 purified vesicle clusters fromD. trinervis nodules; (4) nodule host cell cytoplasm extract; (5) D.trinervis root extract. Arrowheads point the recognised polypeptidesin Frankia BCU110501 protein samples (A, 53 kDa; B, 43 kDa).
Frankia BCU110501 polypeptides after probing anelectroblotted membrane with an antiserum againstAzotobacter vinelandii dinitrogenase protein. A com-mon band of 53 kDa was detected in N2 fixing Frankiacell extracts, in vesicle clusters from N2 fixing nodulesand also in sonicated N2 fixing nodules. This apparentmolecular weight of a dinitrogenase like polypeptideis in accordance with reported masses of the FeMocomponent of dinitrogenases from other diazotrophs(55 – 60 kDa; BRENDA, 2001). No band was ob-served in supernatant of nodule host cell extracts orroot extracts. A second band of 36 kDa was recognisedby this antiserum only in Frankia cell extracts. Therecognition of Frankia BCU110501 proteins by anti-dinitrogenase reductase serum is shown in Figure 2b.The antiserum (a mixture of antibodies directed againAzotobacter vinelandii and Rhodospirillum rubrumdinitrogenase reductase proteins) cross-reacted withpolypeptides of apparent molecular masses of 43 kDain Frankia cell extracts (lane 1), 40 kDa in total nodulesonicates (lane 2), 44 kDa in vesicle clusters extract(lane 3), and with a 35 kDa polypeptide in roots ex-tract (lane 5). Since dinitrogenase polypeptide was notdetected in the same sample, we conclude that the 35kDa band detected by the anti-dinitrogenase reductase
Figure 3. Frankia BCU110501 dinitrogenase protein in darkstressed Discaria trinervis nodules. A black bar indicates the dur-ation of dark stress. Nodule homogenates were centrifuged and thesediment containing host cell debris and Frankia vesicle clusterswas sonicated to solubilize Frankia proteins. 50 µg of protein wereloaded.
Figure 4. Frankia BCU110501 dinitrogenase-reductase protein indark stressed Discaria trinervis nodules. A black bar indicates theduration of dark stress. Nodule homogenates were centrifuged andthe sediment containing host cell debris and Frankia vesicle clusterswas sonicated to solubilize Frankia proteins. 20 µg of protein wereloaded.
serum was an artefact. The molecular mass of detec-ted polypeptides is slightly higher than the Fe-proteinfrom other diazotrophs (27.5 – 36 kDa; BRENDA,2001). Nevertheless, both antisera cross-reacted withproteins from nitrogen fixing tissues. We thus considerthese antisera as a useful tool for the analysis of thestability of nitrogenase proteins in this experiment.
The nitrogenase components in dark stressed D.trinervis plants
Western blot analysis of protein extracts from symbi-otic Frankia clearly showed that the FeMo protein wasstill present at high level in stressed nodules after 4days of darkness, while acetylene reduction was neg-ligible (Figures 3 and 5a). FeMo protein was reducedto a minimum by day 6 of dark stress, and started torecover in normal light conditions (Figures 3 and 5a).By day 8 after restoring normal illumination, the level

162
Figure 5. The levels of Frankia BCU110501 nitrogenase proteinsin dark stressed Discaria trinervis nodules. The intensity of bandsdetected in Western blots shown in Figures 3 and 4 was obtained byimage digitalisation. Results are presented in relative arbitrary units.D denotes the start of darkening treatment; L points the time whendark stress plants were placed under normal illumination. (�), Darkstressed plants. (�), Control plants.
of expression of FeMo protein was fully recovered. Asimilar pattern was observed for the Fe protein dur-ing the experiment (Figures 4 and 5b), although about15% of protein remained by day 6 of darkness. Thecalculated ratio of FeMo protein to Fe protein wassimilar for control and dark treated plants by the timenitrogenase activity was lost (i.e. after 4 days of darkstress), but was 50% lower for dark treated plants thanfor control plants 2 days later (Figure 5c).
The activity and protein level of GS, MDH and ASduring and recovering from dark stress
Polyclonal antibodies against alfalfa cytosolic MDH(cMDH), alfalfa AS and common bean GS recognisedpolypeptides of similar molecular weights that are ex-pressed in D. trinervis nodule extracts (Valverde andWall, 2001). We used these antibodies to monitor theamount of GS, MDH and AS nodule proteins in thisexperiment, together with their enzymatic activities.
Nodule GS specific activity dropped to a minimumafter 4 days of darkness but recovered soon to thepre-stress level by 6 days of darkness (Figure 6). Theamount of GS subunit was reduced to ca. 50% byday 6 of darkness (Figure 6) and returned to normallevels with normal illumination. In control plants, spe-cific GS activity dropped steadily over the 14-days
Figure 6. Activity and levels of GS in dark stressed Discariatrinervis nodules. GS activity (panel A) and immunoblotting (panelB) were assayed on supernatant of nodule homogenates. Panel A:D denotes the start of darkening treatment; L points the time whendark stress plants were placed under normal illumination. (�), Darkstressed plants. (�), Control plants. Values are means (± SE)for n = 3 pouches. Panel B: a black bar indicates the durationof dark stress. Twenty µg of protein were loaded. The antiserumagainst bean GS cross reacts with a 52 kDa D. trinervis polypeptide(Valverde and Wall, 2003).
period (Figure 6), although the amount of GS proteinwas rather constant (Figure 6). The specific activityof MDH in dark stressed plants fell progressively dur-ing the 6 days of darkening, and could not recover tothe pre-stress level. Darkening resulted in a minimalamount of MDH polypeptides by day 6 (Figure 7).After returning to normal illumination, the majorcMDH cross-reactive band was rapidly recovered buttwo smaller anti cMDH-reactive polypeptides showeda slower recovery (Figure 7). In control plants, specificMDH activity remained higher than stressed plants,and the amount of MDH-like polypeptides increasedmainly for the fast migrating bands (Figure 7). In thecase of AS, the dark stress caused a partial reduction(50%) of the protein level in nodules, that was totallyrecovered when light was restored (Figure 8). No sig-nificant change occurred in the amount of AS subunitin control plants during the experiment (Figure 8).
163
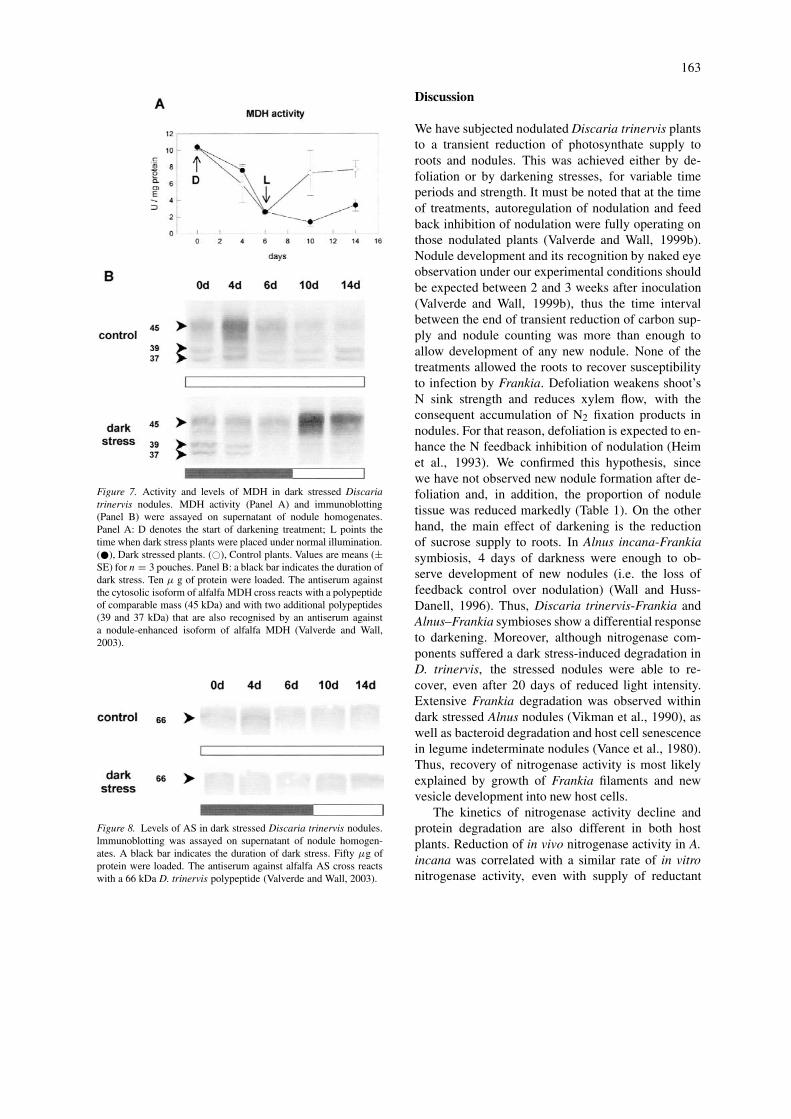
Figure 7. Activity and levels of MDH in dark stressed Discariatrinervis nodules. MDH activity (Panel A) and immunoblotting(Panel B) were assayed on supernatant of nodule homogenates.Panel A: D denotes the start of darkening treatment; L points thetime when dark stress plants were placed under normal illumination.(�), Dark stressed plants. (�), Control plants. Values are means (±SE) for n = 3 pouches. Panel B: a black bar indicates the duration ofdark stress. Ten µ g of protein were loaded. The antiserum againstthe cytosolic isoform of alfalfa MDH cross reacts with a polypeptideof comparable mass (45 kDa) and with two additional polypeptides(39 and 37 kDa) that are also recognised by an antiserum againsta nodule-enhanced isoform of alfalfa MDH (Valverde and Wall,2003).
Figure 8. Levels of AS in dark stressed Discaria trinervis nodules.lmmunoblotting was assayed on supernatant of nodule homogen-ates. A black bar indicates the duration of dark stress. Fifty µg ofprotein were loaded. The antiserum against alfalfa AS cross reactswith a 66 kDa D. trinervis polypeptide (Valverde and Wall, 2003).
Discussion
We have subjected nodulated Discaria trinervis plantsto a transient reduction of photosynthate supply toroots and nodules. This was achieved either by de-foliation or by darkening stresses, for variable timeperiods and strength. It must be noted that at the timeof treatments, autoregulation of nodulation and feedback inhibition of nodulation were fully operating onthose nodulated plants (Valverde and Wall, 1999b).Nodule development and its recognition by naked eyeobservation under our experimental conditions shouldbe expected between 2 and 3 weeks after inoculation(Valverde and Wall, 1999b), thus the time intervalbetween the end of transient reduction of carbon sup-ply and nodule counting was more than enough toallow development of any new nodule. None of thetreatments allowed the roots to recover susceptibilityto infection by Frankia. Defoliation weakens shoot’sN sink strength and reduces xylem flow, with theconsequent accumulation of N2 fixation products innodules. For that reason, defoliation is expected to en-hance the N feedback inhibition of nodulation (Heimet al., 1993). We confirmed this hypothesis, sincewe have not observed new nodule formation after de-foliation and, in addition, the proportion of noduletissue was reduced markedly (Table 1). On the otherhand, the main effect of darkening is the reductionof sucrose supply to roots. In Alnus incana-Frankiasymbiosis, 4 days of darkness were enough to ob-serve development of new nodules (i.e. the loss offeedback control over nodulation) (Wall and Huss-Danell, 1996). Thus, Discaria trinervis-Frankia andAlnus–Frankia symbioses show a differential responseto darkening. Moreover, although nitrogenase com-ponents suffered a dark stress-induced degradation inD. trinervis, the stressed nodules were able to re-cover, even after 20 days of reduced light intensity.Extensive Frankia degradation was observed withindark stressed Alnus nodules (Vikman et al., 1990), aswell as bacteroid degradation and host cell senescencein legume indeterminate nodules (Vance et al., 1980).Thus, recovery of nitrogenase activity is most likelyexplained by growth of Frankia filaments and newvesicle development into new host cells.
The kinetics of nitrogenase activity decline andprotein degradation are also different in both hostplants. Reduction of in vivo nitrogenase activity in A.incana was correlated with a similar rate of in vitronitrogenase activity, even with supply of reductant

164
and ATP (Lundquist and Huss-Danell, 1991b). Thisargues in favour of a dark stress-induced degrada-tion of nitrogenase protein (observed mainly for theFeMo component), with a concomitant loss of activ-ity. In D. trinervis, complete loss of activity was alsoobserved after 4 days of darkness (Figure 1), but nitro-genase polypeptides were not yet degraded (Figures3–5). The minimal amount of Fe and FeMo poly-peptides occurred after 6 days of dark stress (Figures3–5). Degradation kinetics were comparable for bothnitrogenase components. In A. incana nodules, dark-ening had a more profound effect on FeMo proteinrather than Fe protein (Lundquist and Huss-Danell,1991a,b). When dark stress was alleviated, the re-covery of protein level was similar for both subunits(Figures 3–5). These results clearly indicate that ametabolic down regulation of nitrogenase activity pre-cedes the degradation of its components. The cascadeof biochemical events leading to nitrogenase degrada-tion during a dark stress is, however, not clarified yet.It was proposed (Lundquist and Huss-Danell, 1991a,b)that the elevation of pO2 due to a reduced respira-tion rate in the presence of low amounts of C sourcesmight trigger nitrogenase inactivation and subsequentdegradation, as occurs in Anabaena where exposure ofcell extracts to O2 was necessary before degradation ofFeMo protein (Murry et al., 1983).
The regulatory pattern of some enzymes involvedin ammonia assimilation (such as GS, MDH and ASproteins) during the dark stress was more complexthan observed for nitrogenase, but in general they wereless sensitive to the dark stress. There was a partialreduction of polypeptide levels by day 6 of darkening,with a subsequent rapid recovery after returning tonormal light conditions (Figures 6–8). This negativeregulation of protein level seemed to follow in time,the inhibition of nitrogen fixation. An apparent activ-ation of GS was observed at the time its protein levelwas the lowest (Figure 6). The low concentration ofammonium (due to nitrogenase inactivation) could bethe reason for activation of GS. On the other hand, thesustained reduced activity of MDH after alleviation ofdark stress was not correlated with the total level ofMDH polypeptides, but it rather correlated with theamount of smaller MDH-like polypeptides (Figure 7).
A temporal shortage of carbon supply to nodulesindeed prevented nitrogen fixation (Figure 1). Devel-opment of new nodules however, was not activated.Thus, it seems that current nitrogenase activity andammonia assimilation are weakly (or even not) linked
with the feedback control of nodulation in D. trinervis.It may be that the limitation of C supply to nodules thatwere imposed to D. trinervis nodules was not enoughto cause a significant reduction of the N content ofthe shoot; thus the feedback control of nodulationmediated by N-status signals (Parsons et al., 1993;Valverde et al., 2000) was still operative during andafter C stresses. An alternative explanation could bethat an autoregulation-like signal (Valverde and Wall,1999b) originating in nodules may not be sensitiveto transient shortages of carbon supply. Thus, theseobservations give support to the existence of othersources of negative regulatory signals over nodulation(Wall, 2000).
Acknowledgements
We thank P.W. Ludden for kindly providing nitro-genase antibodies, C.P. Vance for MDH and AS an-tibodies, and T. Gordon for GS antibodies. We aregreatly indebted to Dr. Pedro Balatti for the GC fa-cility. This work was funded by Universidad Nacionalde Quilmes (Argentina, grant 827-0201/99) and by theInternational Foundation for Science (IFS, Sweden,grant D2952-1). L.G.W. is member of the ResearchCareer of CONICET. CV. holds a fellowship fromCONICET.
References
Akkermans A D L, Huss-Danell K and Roloefsen W 1981 Enzymesof the tricarboxilic acid cycle and the malate-aspartate shuttle inthe N2-fixing endophyte of Alnus glutinosa. Physiol. Plant. 53,289–294.
Blake M S, Johnston K H, Russell-Jones G J and Gotschlich EC 1984 A rapid, sensitive method for detection of alkalinephosphatase conjugated anti-antibody on western blots. Anal.Biochem. 136, 175–79.
BRENDA. The comprehensive enzyme information system. www.brenda.uni-koeln.de
Burnette W N 1981 ‘Western blotting’: Electrophoretic transfer ofproteins from sodium dodecyl sulfate-polyacrylamide gels to un-modified nitrocellulose and radiographic detection with antibodyand radioidinated Protein A. Anal. Biochem. 112, 195–203.
Chaia E 1998 Isolation of an effective strain of Frankia fromnodules of Discaria trinervis (Rhamanceae). Plant Soil 205,99–102.
Egli M A, Griffith S M, Miller S S, Anderson M P and Vance C P1989 Nitrogen assimilating enzyme activities and enzyme pro-tein during development and senescence of effective and plantgene-controlled ineffective alfalfa nodules. Plant. Physiol. 91,898–904.

165
Heim I, Hartwig U A and Nösberger J 1993 Current nitrogen fixa-tion is involved in the regulation of nitrogenase activity in whiteclover (Trifolium repens L.). Plant Physiol. 103, 1009–1014.
Huss-Danell K 1978 Nitrogenase activity measurements in intactplants of Alnus incana. Physiol. Plant., 43, 342–348.
Huss-Danell K 1997 Actinorhizal symbioses and their N2 fixationNew Phytol. 136, 375–405.
Laemmli U K 1970 Cleavage of structural proteins during theassembly of the head of the bacteriophage T4. Nature 227,680–685.
Lundquist PO and Huss-Danell K 1991a Nitrogenase activity andamounts of nitrogenase proteins in a Frankia-Alnus incanasymbiosis subjected to darkness. Plant Physiol. 95, 808–813.
Lundquist PO and Huss-Danell K 1991b Response of nitrogenaseto altered carbon supply in a Frankia-Alnus incana symbiosis.Physiol. Plant. 83, 331–338.
Murry M A, Hallenbeck P C, Esteva D and Benemann J R 1983Nitrogenase inactivation by oxygen and enzyme turnover inAnabaena cylindrica. Can. J. Microbiol. 29, 1286–1294.
Nittayajarn A and Baker DD 1989 Methods for the quantification ofFrankia cell biomass. Plant Soil 118, 199–204.
Parsons R, Stanforth A, Raven J A and Sprent J 1993 Nodulegrowth and activity may be regulated by a feedback mechanisminvolving phloem nitrogen. Plant Cell Environ. 16, 125–136.
Romanov V I, Gordon A J, Minchin F R, Witty J F, Skot L, James CL and Tikhonovich I A 1998 Physiological and biochemical char-acteristics of FN1, a ‘fixation impaired’ mutant of pea (Pisumsativum L.). J. Exp. Bot. 49, 1789–1796.
Smith P K, Krohn R I, Hermanson G T, Mallia A K, Gartner F H,Provenzano M D, Fujimoto E K, Goeke NM, Olson B J andKienk D C 1985 Measurement of protein using bicinchoninicacid. Anal. Biochem. 150, 76–85.
Swensen S M and Mullin B C 1997 Phylogenetic relationshipsamong actinorhizal plants. The impact of molecular systemat-ics and implications for the evolution of actinorhizal symbiosis.Physiol. Plant. 99, 565–573.
Valverde C and Wall L G 1999a Time course of nodule devel-opment in Discaria trinervis (Rhamnaceae)-Frankia symbiosis.New Phytol. 141, 345–354.
Valverde C and Wall L G 1999b Regulation of nodulation in Dis-caria trinervis (Rhamnaceae)-Frankia symbiosis. Can. J. Bot.77, 1302–1310.
Valverde C 2000 ‘Regulación de la nodulación radicular en Ia sim-biosis Discaria trinervis – Frankia’. Ph. D. Thesis, UniversidadNacional de La Plata, Argentina.
Valverde C, Wall L G and Huss-Danell K 2000 Regulation of nod-ulation and nodule mass relative to nitrogenase activity and ni-trogen demand in seedlings of Discaria trinervis (Rhamnaceae).Symbiosis 28, 49–62.
Valverde C and Wall L G 2003 Ammonium assimilation in Discariatrinervis root nodules. Regulation of enzyme activities and pro-tein levels by the availability of macronutrients (N, P and C).Plant Soil 254, 139–153.
Vance C P, Johnson L E B, Halvorsen A M, Heichel G H andBarnes D K 1980 Histological and ultrastructural observationsof Medicago sativa root nodule senescence after foliage removal.Can. J. Bot. 58, 295–309.
Vance C P 2000 Amide biosynthesis in root nodules of temperatelegumes. In Prokaryotic Nitrogen Fixation: A Model System forAnalysis of a Biological Process. Ed. E W Triplett. pp. 589–607.Horizon Scientific Press, Wymondham, UK.
Vikman P -A, Lundquist P -O and Huss-Danell K 1990 Respiratorycapacity, nitrogenase activity and structural changes of Frankiain symbiosis with Alnus incana, in response to prolonged dark-ness. Planta 182, 617–625.
Wall L G 2000 The actinorhizal symbiosis. J. Plant Growth Reg. 19,167–182.
Wall L G and Huss-Danell K 1996 La actividad fijadora de nitrógenocomo factor de regulación de la nodulación en la simbiosis Alnusincana – Frankia. In Proceedings of the XXI Meeting of theArgentinean Society of Plant Physiology (Mendoza, Argentina,20–22 March 1996). pp. 450–451.
Wheeler C T 1971 The causation of the diurnal changes in nitrogenfixation in the nodules of Alnus glutinosa New Phytol. 70, 487–495.





























