Embed Size (px)
Citation preview

The growth inhibitory effect of conjugated linoleic acid on MCF-7cells is related to estrogen response system
Vijayender Rao Durgam, Gabriel Fernandes*
Department of Medicine/Clinical Immunology, The University of Texas Health Science Center at San Antonio,7703 Floyd Curl Drive, San Antonio, TX 78284-7874, USA
Received 28 January 1997; revision received 21 February 1997; accepted 21 February 1997
Abstract
Conjugated linoleic acid (CLA) has been shown to have a direct oncostatic action on MCF-7 human breast cancer cells inculture. However, the mechanism involved is not fully elucidated. In this study we have examined whether the inhibitor actionis related to the estrogen responsiveness of MCF-7 cells. Our results demonstrate that CLA selectively inhibits proliferation ofER positive MCF-7 cells as compared with ER negative MDA-MB-231 cells. Cell cycle studies indicated that a higherpercentage of CLA treated MCF-7 cells remained in the G0/G1 phase as compared to control and those treated with linoleicacid (LA). CLA also inhibited expression of c-myc in MCF-7 cells. These results demonstrate that CLA may inhibit MCF-7cell growth by interfering with the hormone regulated mitogenic pathway. We are reporting for the first time the involvementof CLA, a dietary factor, in the regulation of hormone mediated mitogenic pathways in ER positive breast cancer cellproliferation in vitro. Published by Elsevier Science Ireland Ltd.
Keywords:MCF-7 cell growth; CLA; Cell cycle; c-myc expression
1. Introduction
The role of bioactive lipids in pathophysiologicalprocesses such as cancer, inflammation and relateddiseases is well recognized. The importance of dietaryfatty acids and their metabolites, in the modulation ofmolecular events in a proliferating tumor cell, hasbeen recently investigated [1–4]. The role of linoleicacid in promoting [5–8], andq-3 fatty acids in inhi-biting [9–11] tumor growth is well established.Although the exact mechanism involved for promot-
ing these effects is not yet known, arachidonic acidderived eicosanoids appear to play an important rolein increasing malignancy [12,13].
Regulation of gene expression by hormones is wellestablished. MCF-7 estrogen responsive and MDA-MB-231 unresponsive human breast cancer celllines were extensively investigated in defining themechanism by which hormones affect breast tumorcell proliferation. MCF-7 contains both estrogenreceptor (ER) [14] and progesterone receptor (PR)[15]. MDA-MB-231 do not respond to estrogen andthe ER gene is controlled transcriptionally in the ERnegative breast cancer cell line [16]. Reports indicatethat the estrogen induced MCF-7 cell growth is
CancerLetters116 (1997)121–130
0304-3835/97/$17.00 Publishedby ElsevierScienceIrelandLtd.PI I S0304-3835(97)00192-4
* Corresponding author. Tel.: +1 210 5674679; fax: +1 2105674592.

accompaniedby changes in the expressionof earlyprotooncogenessuchas c-myc and c-fos [17,18]. Anegative modulation of theconcentrationof receptorsfor estrogenandprogestinsin varioustissuesby ara-chidonic acid andothernon-esterified fatty acidshasbeenreported [19].
Recently the anti-carcinogenicpropertyof conju-gated linoleic acid,an isomeric derivative of linoleicacid containing conjugated double bonds, wasreported[20,21]. Conjugatedlinoleic acid is a naturalconstituent of manyfoods. It is higher in foodsfromruminants,suchas meatand dairy products (valuesranged from 0.6 to 5.6 mg CLA/g fat) as comparedto foodsfrom non-ruminants.It wasinitially identifiedas an antimutagenic agentfrom grilled groundbeef[20]. Inhibition of benzopyreneinducedfore-stomachneoplasiain miceby CLA wasfirst reported by Ha etal. [22]. It wasfurthershown thatdietaryCLA is alsoeffective in the preventionof DMBA inducedmam-mary tumors[21]. Otherstudies indicatedthata diet-ary level of 0.1% CLA is sufficient to produceasignificant inhibition of carcinogeninducedrat mam-mary tumors[23].
CLA has beenshown to have a direct inhibitoryaction on MCF-7 human breastcancer cells in culture[24]. However, the mechanism involved in causingthe inhibitory effect is not yet understood.In thepre-sentstudywe examinedwhethertheinhibitory actionis relatedto the estrogen responsesystemof MCF-7cells. In order to test this hypothesis,we determinedtherateof proliferation of estrogen responsiveMCF-7cells and unresponsive MDA-MB-231 cells cocul-tured with CLA and LA and compared them withthe cells grown on normal media without addingany fatty acids.We alsoexamined whether the CLAtreated MCF-7 cells would reversebackto prolifera-tion stateupontransfer to normalmedia.Furthermore,wehavealso examinedcell cyclephasedistribution ofproliferatingMCF-7cellsandmeasuredthelevelof c-myc expression. Our results indicated that CLA isquite effective in inhibiting hormone responsiveMCF-7 cell growth as compared to MDA-MB-231cells.This inhibition maybedueto adelay in progres-sion throughtheG1 checkpoint duringthecell cycleby down regulation of c-myc expression. Further-more, these results also indicate a possible role forCLA in down regulating c-myc expression throughinterferencewith the estrogenresponsesystem. This
may be oneof the possible mechanismsfor reducedMCF-7 cell growth by CLA. This is the first directevidenceof theinvolvementof CLA, a dietaryfactor,in thenegativeregulation of hormonemediatedbreastcancercell growth.
2. Materials and methods
2.1. Determination of rate of cell proliferation
Human breast cancer cells, estrogenresponsiveMCF-7 and unresponsive MDA-MB-231, wereobtained from Dr Kent C. Osborne, Division ofOncology, The University of TexasHealth ScienceCenter at San Antonio. The MCF-7 cell line wasmaintained in IMEM media (Biofluids, Rockville,MD) with 5% calf serum (JRH Biosciences,Lenexa,KS) and10−8 M insulin. MDA-MB-231 cell line wasmaintained on IMEM media with 10% calf serum.The linoleic acid was obtained from Sigma (St.Louis, MO) and conjugatedlinoleic acid was a giftfrom Dr MichaelW. Pariza(FoodResearchInstitute,University of Wisconsin, Madison,WI).
The cells were cultured in 12-well costar plateswith 1.5–2.0 × 106 cells/ml in IMEM complete med-ium with added fatty acids at a concentration of3.5 × 10−5 M in ethanol (ethanol not exceeding0.1% in medium).The cells were grown at 37°C inan incubator with 95% air and 5% CO2 for up to 6days, with every day counting of viable/deadcellsafter treating with trypsin EDTA (Cellgro, Herdon,VA) and the viability was determined by stainingwith 0.4% trypan blue (Sigma). Simultaneously, thecellswereplatedin a 96-well plateandincubatedforupto 96h. Fromzerohourafterevery 24h, 0.2mCi of(3H)-thymidine (specificactivity, 25 Ci/mmol; Amer-sham,Arlington Heights,IL) wasaddedto eachwellandafter1 h cellswerewashedwith salinephosphatebuffer, trypsinized and harvested with Titertek cellharvester (Flow Laboratories, Rockville, MD) andincorporated (3H)-thymidine was countedfor 1 minusing Packard Tri-carb 460 (Packard Instruments,Downers groove, IL). For studiesto determine theeffect of different concentrations of fatty acids onthe growth of MCF-7 cells, three concentrations(1.7,3.5 and7.1 × 10−5 M) of fatty acidsweretestedundersimilar conditionsdescribed above.For media
122 V.R. Durgam, G. Fernandes/ CancerLetters116 (1997)121–130
replacement studies, the cells were incubated eitherwith media aloneor media wasaddedwith fatty acidsfor 4 daysin duplicates.In onesetof wells themediawith added fatty acids was replaced with normalmedia. The other set of wells was continued inmedia with addedfatty acids. Then the incubationwascontinuedfurther for up to 6 days.
2.2. Isolation of RNA and Northern Blot analysis
TheMCF-7cells grownin normal mediaandmediaaddedwith LA andCLA (3.5 × 10−5 M) weregrownin T-75cell cultureflasks.After 72h of incubationthecells weretreated with trypsin EDTA andcell viabi-lity was determined. The total RNA was isolatedusing guanidium thiocynate hydrochloride method[25]. Total RNA (20 mg) was electrophoresedandelectroblotted on to nitrocellulose (Schleicher andSchuell) and fixed to membraneby UV irradiation(Stratalinker 1800,Statagene).The blots werehybri-dized in a buffer with 50% formamide at 42°C. ThecDNA probes usedin this studywerec-myc (1.4 kbEcoRI/ClaI fragment, Oncor,Gaithersberg, MD) andGAPDH (1.2kb,Pst.1 fragment,ATCC).Af terwash-ing the blots with high stringency(0.1 × SSC,0.5%SDS at 65°C for 2 × 15 min) they were exposed at−80°C to Kodak XAR-5 film with DuPont intensify-ing screens.The intensified auto radiograph bandswere measured semiquantitatively by video imageanalysis using the NIH image1.4 program. The dataareexpressed asthe ratio of the intensity of the spe-cific RNA bandafter normalization to that of consti-tutively expressedGAPDH mRNA.
2.3. Cell cycleanalysis
The MCF-7 cells grown in normal media (control)andmediaaddedwith CLA andLA (3.5 × 10−5 M) for72 h under the aboveconditions were usedfor cellcycleanalysis.Thecellswerestained with amodifiedKrishantechnique[26] (50mg/ml propidium iodideina hypotonic sodiumcitrate solutionwith 0.3%NP-40and 0.1 mg/ml RNase-A) at 1.0 × 106 cells/ml, vor-texed andstained for 30 min at room temperature inthedark.Prior to flow cytometricmeasurements,sam-ples were filtered througha 37 mm nylon meshinto12 × 75mmtubesandstoredat4°C until analysiswascompleted within 24 h. Al l samples were analyzed
with anEPICS ELITE flow cytometer (CoulterCyto-metry, Miami, FL) using a 15 mW argon ion laseroperated at 6 A of power at 488nm. Photomultipliertubevoltagewasadjustedfor eachcontrol sample topositiontheG0/G1 to channel 240on a 1024channelpresentation.Histogramswereanalyzedfor cell cyclecompartments using multi Cycle-Plus version 3.0(Phoenix Flow Systems,SanDiego,CA). Histogramsinclude50K collectedeventsin orderto maximizethestatistical validity of thecompartmental analysis.Thecell cycle analysis experimentwascarried out at theflow cytometerlaboratoryof theCancerTherapy andResearchCenter in SanAntonio, TX.
2.4. Statisticalanalysis
Cell culture experiments with a minimum of sixvalueswereobtainedand the valueswereexpressedas mean± SEM. The proliferative responseof cellswith andwithout fatty acidswasanalyzedstatisticallyusingStudent’s t-test, andP , 0.05 wasconsideredsignificant.
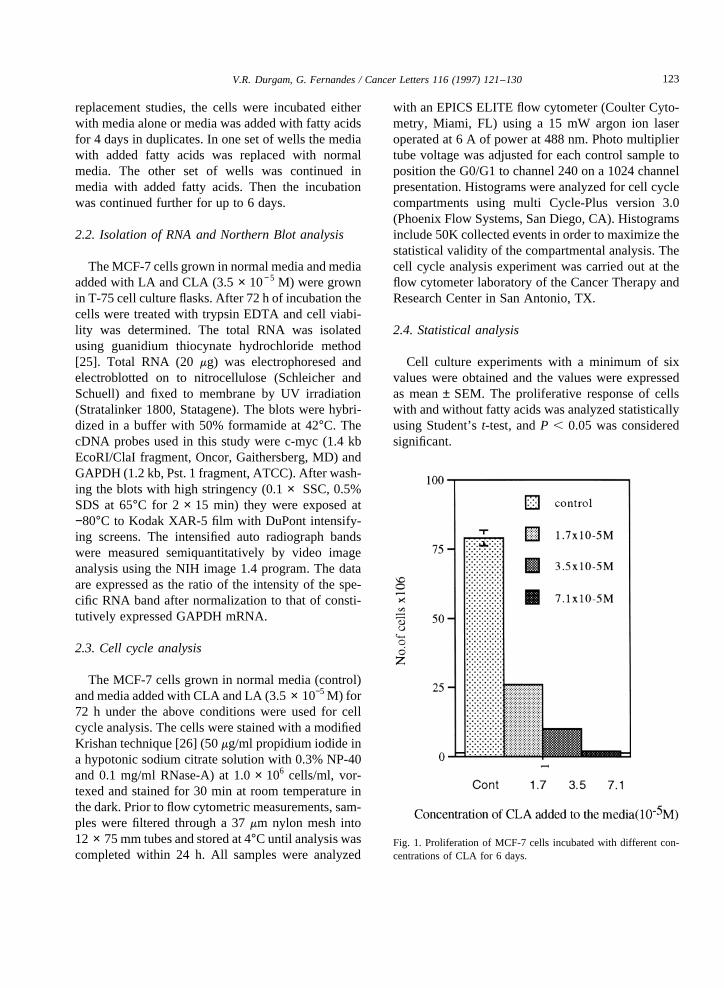
Fig. 1. Proliferationof MCF-7 cells incubatedwith different con-centrationsof CLA for 6 days.
123V.R. Durgam, G. Fernandes/ CancerLetters116 (1997)121–130
3. Results
The proliferative response of MCF-7 and MDA-MB-231 cells co-cultured with CLA and LA(3.5 × 10−5 M) and media alone were comparedbyusing both the viable cell countbasedon the trypanblue exclusionmethodand by thymidine incorpora-tion assay. Among threeconcentrationsof fatty acidstested(1.7,3.5,7.1 × 10−5 M), the3.5 × 10−5 M con-centrationwasfoundto beoptimumwithout anycyto-toxicity to the MCF-7 cells (Fig. 1).
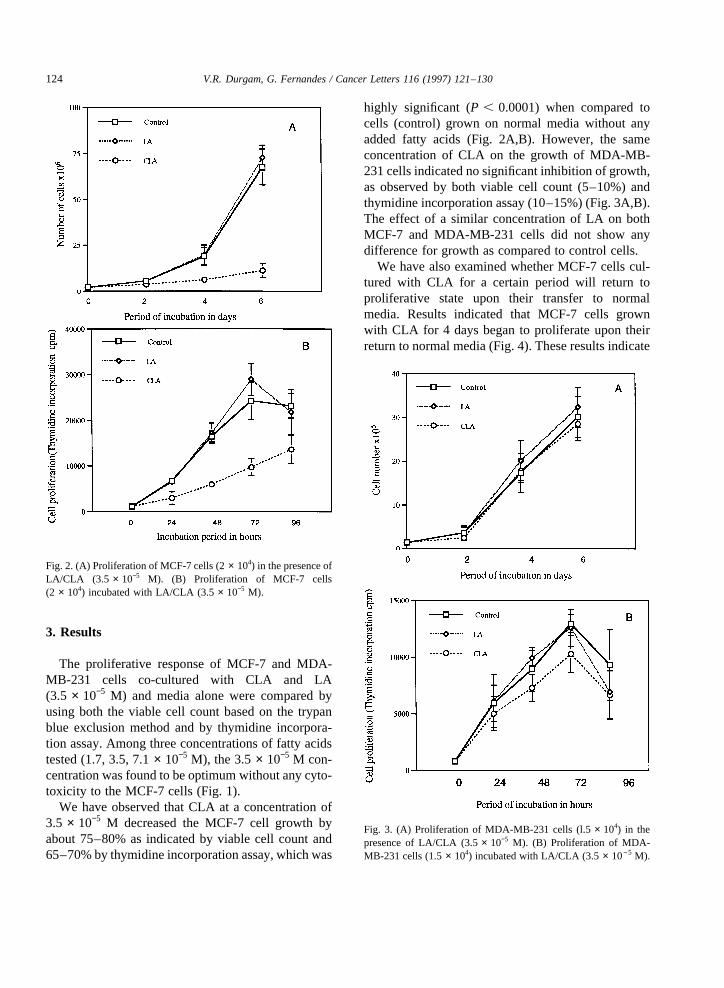
We haveobserved that CLA at a concentrationof3.5 × 10−5 M decreasedthe MCF-7 cell growth byabout75–80% as indicatedby viable cell countand65–70%by thymidineincorporationassay,whichwas
highly significant (P , 0.0001) when compared tocells (control) grown on normal media without anyaddedfatty acids (Fig. 2A,B). However, the sameconcentration of CLA on the growth of MDA-MB-231cells indicatednosignificant inhibition of growth,as observed by both viable cell count (5–10%) andthymidineincorporation assay(10–15%)(Fig. 3A,B).The effect of a similar concentration of LA on bothMCF-7 and MDA-MB -231 cells did not show anydifferencefor growthascompared to control cells.
We havealsoexamined whether MCF-7 cells cul-tured with CLA for a certain period will return toproliferative state upon their transfer to normalmedia. Results indicated that MCF-7 cells grownwith CLA for 4 daysbeganto proliferateupontheirreturnto normalmedia (Fig. 4). Theseresultsindicate
Fig. 2. (A) Proliferation of MCF-7cells(2 × 104) in thepresenceofLA/CLA (3.5 × 10−5 M). (B) Proliferation of MCF-7 cells(2 × 104) incubatedwith LA/CLA (3.5 × 10−5 M).
Fig. 3. (A) Proliferation of MDA-MB-231 cells (l.5 × 104) in thepresenceof LA/CLA (3.5 × 10−5 M). (B) Proliferation of MDA-MB-231 cells (1.5 × 104) incubatedwith LA/CLA (3.5 × 10−5 M).
124 V.R. Durgam, G. Fernandes/ CancerLetters116 (1997)121–130
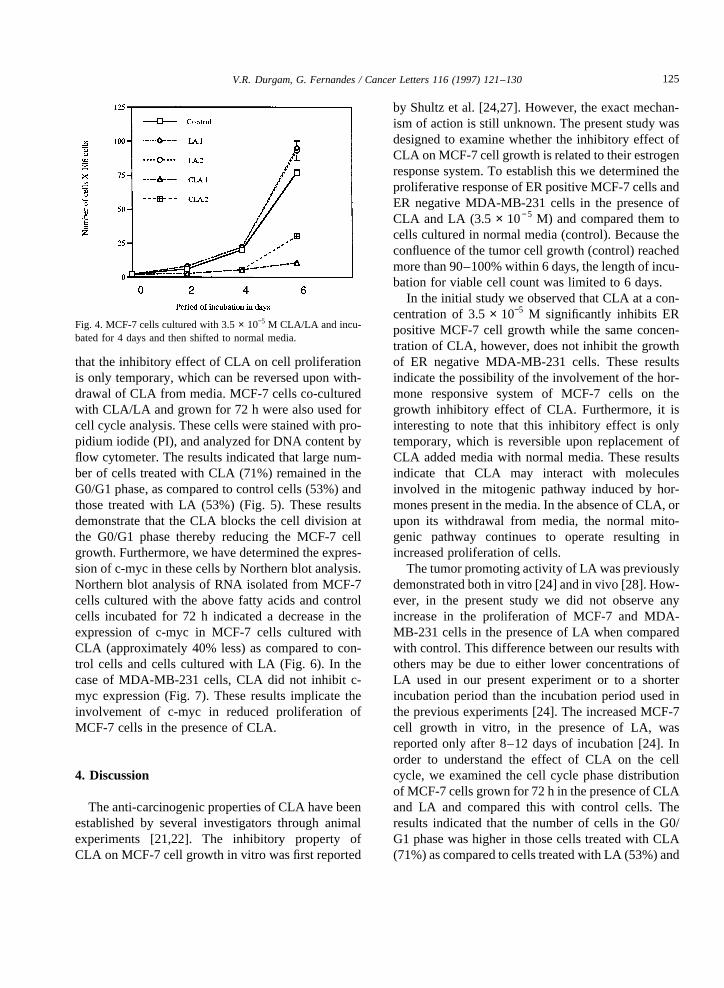
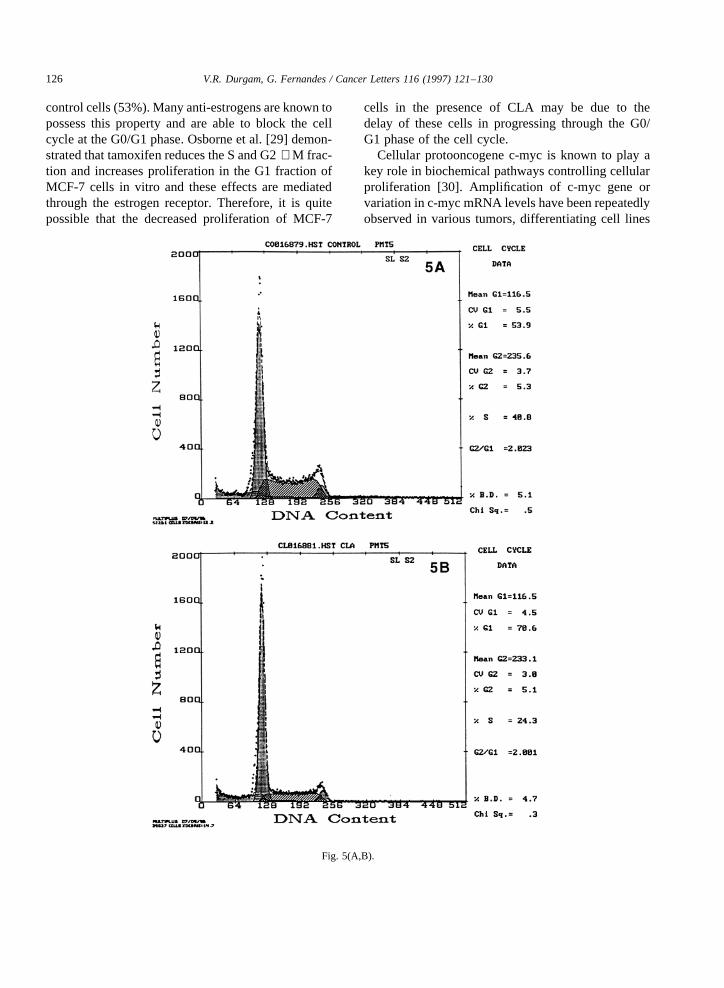
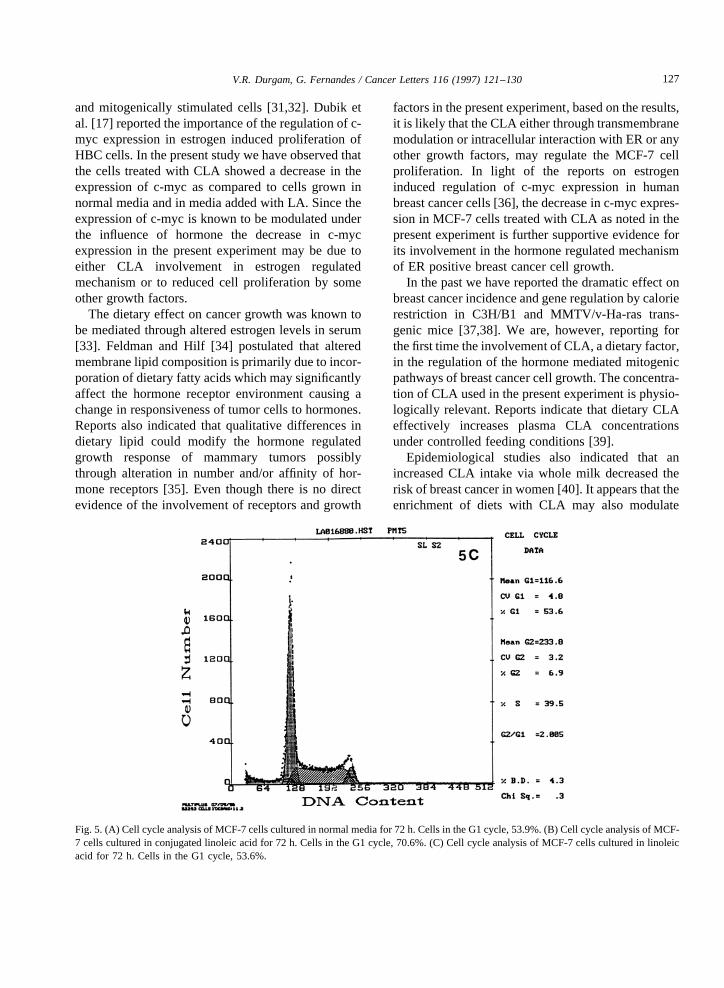
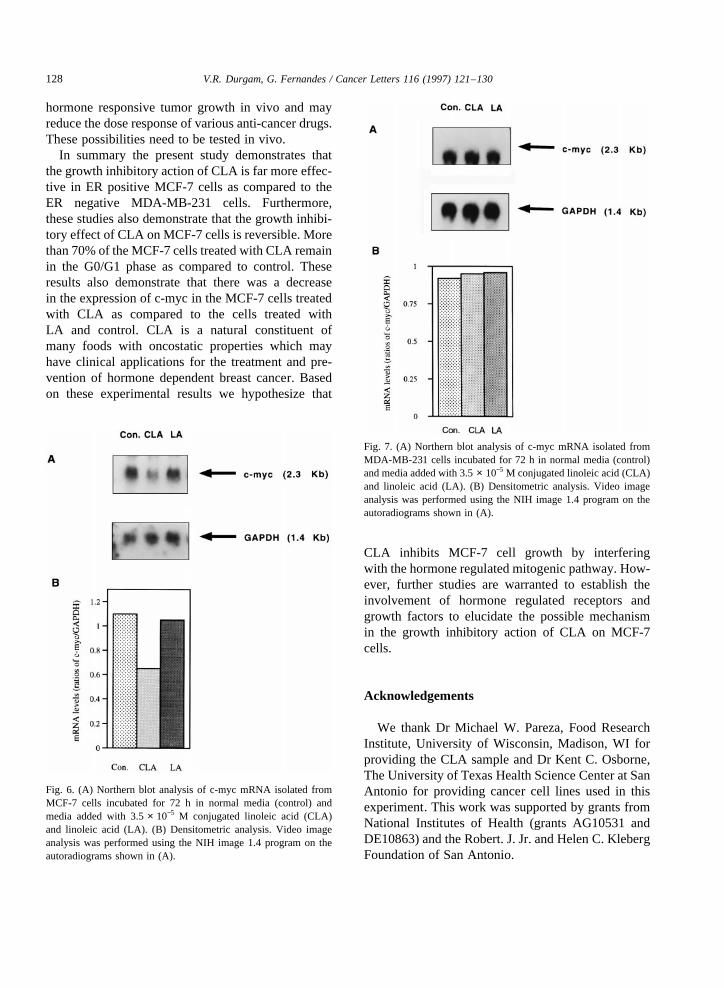
that the inhibitory effectof CLA on cell proliferationis only temporary, which canbe reversed uponwith-drawal of CLA from media. MCF-7 cells co-culturedwith CLA/LA andgrown for 72 h werealso usedforcell cycleanalysis.Thesecells werestainedwith pro-pidium iodide(PI), andanalyzedfor DNA contentbyflow cytometer. Theresults indicatedthat largenum-ber of cells treated with CLA (71%) remainedin theG0/G1 phase, ascomparedto control cells (53%)andthose treatedwith LA (53%) (Fig. 5). Theseresultsdemonstratethat the CLA blocks the cell division atthe G0/G1 phasethereby reducing the MCF-7 cellgrowth. Furthermore,wehavedeterminedtheexpres-sionof c-mycin thesecellsby Northernblot analysis.Northernblot analysisof RNA isolatedfrom MCF-7cells cultured with the abovefatty acids andcontrolcells incubatedfor 72 h indicated a decreasein theexpression of c-myc in MCF-7 cells cultured withCLA (approximately 40% less)ascomparedto con-trol cells and cells cultured with LA (Fig. 6). In thecaseof MDA-MB-231 cells, CLA did not inhibit c-myc expression(Fig. 7). Theseresults implicate theinvolvement of c-myc in reduced proliferation ofMCF-7 cells in the presenceof CLA.
4. Discussion
Theanti-carcinogenic propertiesof CLA havebeenestablishedby severalinvestigators through animalexperiments [21,22]. The inhibitory property ofCLA onMCF-7cell growthin vitro wasfirst reported
by Shultzet al. [24,27]. However, the exactmechan-ism of actionis still unknown. The presentstudy wasdesignedto examine whether the inhibitory effect ofCLA onMCF-7cell growthis relatedto theirestrogenresponsesystem.To establish this we determined theproliferative responseof ERpositive MCF-7cellsandER negative MDA-MB -231 cells in the presenceofCLA andLA (3.5 × 10−5 M) andcompared themtocellscultured in normalmedia (control). Because theconfluenceof thetumorcell growth (control) reachedmorethan90–100%within 6 days,thelength of incu-bationfor viable cell countwaslimit ed to 6 days.
In the initial studywe observedthatCLA at a con-centration of 3.5 × 10−5 M significantly inhibits ERpositive MCF-7 cell growth while the same concen-tration of CLA, however, doesnot inhibit thegrowthof ER negative MDA-MB-231 cells. Theseresultsindicatethepossibility of the involvementof thehor-mone responsive system of MCF-7 cells on thegrowth inhibitory effect of CLA. Furthermore, it isinteresting to note that this inhibitory effect is onlytemporary, which is reversible upon replacement ofCLA addedmediawith normalmedia.Theseresultsindicate that CLA may interact with moleculesinvolved in the mitogenicpathway induced by hor-monespresentin themedia. In theabsenceof CLA, orupon its withdrawal from media, the normal mito-genic pathway continues to operate resulting inincreasedproliferationof cells.
Thetumorpromoting activity of LA waspreviouslydemonstratedbothin vitro [24] andin vivo [28]. How-ever, in the presentstudy we did not observeanyincrease in the proliferation of MCF-7 and MDA-MB-231 cells in the presenceof LA whencomparedwith control.This differencebetweenour resultswithothersmay be due to either lower concentrationsofLA used in our presentexperiment or to a shorterincubation period than the incubation periodusedinthepreviousexperiments[24]. The increasedMCF-7cell growth in vitro, in the presenceof LA, wasreported only after 8–12 daysof incubation [24]. Inorder to understand the effect of CLA on the cellcycle, we examined the cell cycle phasedistributionof MCF-7cellsgrown for 72h in thepresenceof CLAand LA and compared this with control cells. Theresultsindicated that the numberof cells in the G0/G1 phasewashigherin thosecells treatedwith CLA(71%)ascompared to cells treatedwith LA (53%)and
Fig. 4. MCF-7 cellsculturedwith 3.5 × 10−5 M CLA/LA andincu-batedfor 4 daysandthenshiftedto normalmedia.
125V.R. Durgam, G. Fernandes/ CancerLetters116 (1997)121–130
control cells(53%).Manyanti-estrogensareknown topossess this property and are able to block the cellcycle at theG0/G1 phase. Osborneet al. [29] demon-stratedthattamoxifen reducestheSandG2 + M frac-tion and increasesproliferation in the G1 fraction ofMCF-7 cells in vitro and theseeffects are mediatedthrough the estrogenreceptor. Therefore, it is quitepossible that the decreased proliferation of MCF-7
cells in the presence of CLA may be due to thedelay of these cells in progressing through the G0/G1 phaseof the cell cycle.
Cellular protooncogene c-myc is known to play akey role in biochemicalpathways controlling cellularproliferation [30]. Amplification of c-myc gene orvariationin c-mycmRNA levelshavebeenrepeatedlyobservedin varioustumors, differentiating cell lines
Fig. 5(A,B).
126 V.R. Durgam, G. Fernandes/ CancerLetters116 (1997)121–130
and mitogenicallystimulatedcells [31,32]. Dubik etal. [17] reported theimportanceof theregulation of c-myc expression in estrogen inducedproliferation ofHBC cells.In thepresentstudywehaveobservedthatthe cells treatedwith CLA showeda decreasein theexpressionof c-myc as compared to cells grown innormal media andin media addedwith LA. Sincetheexpressionof c-myc is knownto bemodulatedunderthe influence of hormone the decrease in c-mycexpressionin the presentexperiment may be due toeither CLA involvement in estrogen regulatedmechanismor to reducedcell proliferation by someother growth factors.
The dietary effect on cancer growth wasknown tobe mediatedthrough altered estrogenlevels in serum[33]. Feldmanand Hilf [34] postulated that alteredmembranelipid compositionis primarily dueto incor-poration of dietaryfatty acidswhichmaysignificantlyaffect the hormonereceptor environmentcausingachangein responsiveness of tumorcells to hormones.Reportsalso indicatedthat qualitativedifferences indietary lipid could modify the hormone regulatedgrowth response of mammary tumors possiblythrough alteration in number and/oraffinity of hor-monereceptors [35]. Even though thereis no directevidenceof the involvement of receptors andgrowth
factors in thepresentexperiment, basedontheresults,it is likely thattheCLA eitherthroughtransmembranemodulationor intracellular interactionwith ERor anyother growth factors, may regulatethe MCF-7 cellproliferation. In light of the reports on estrogeninduced regulation of c-myc expression in humanbreastcancer cells [36], thedecreasein c-mycexpres-sion in MCF-7 cells treatedwith CLA asnotedin thepresentexperimentis furthersupportive evidenceforits involvement in thehormoneregulatedmechanismof ER positivebreastcancer cell growth.
In thepastwe havereportedthedramatic effectonbreastcancer incidenceandgeneregulation by calorierestriction in C3H/B1 and MMTV /v-Ha-ras trans-genic mice [37,38]. We are, however, reporting forthefirst timetheinvolvement of CLA, adietaryfactor,in the regulation of the hormonemediatedmitogenicpathwaysof breastcancer cell growth. Theconcentra-tion of CLA usedin thepresentexperimentis physio-logically relevant. Reports indicatethat dietaryCLAeffectively increases plasma CLA concentrationsundercontrolled feeding conditions[39].
Epidemiological studies also indicated that anincreased CLA intake via whole milk decreased therisk of breastcancerin women[40]. It appearsthattheenrichmentof diets with CLA may also modulate
Fig. 5. (A) Cell cycleanalysisof MCF-7cellsculturedin normalmediafor 72h. Cellsin theG1cycle,53.9%.(B) Cell cycleanalysisof MCF-7 cellsculturedin conjugatedlinoleic acid for 72 h. Cells in theG1 cycle,70.6%.(C) Cell cycleanalysisof MCF-7 cellsculturedin linoleicacid for 72 h. Cells in the G1 cycle, 53.6%.
127V.R. Durgam, G. Fernandes/ CancerLetters116 (1997)121–130
hormone responsive tumor growth in vivo and mayreducethedoseresponseof variousanti-cancerdrugs.Thesepossibilities needto be testedin vivo.
In summary the presentstudy demonstratesthatthegrowthinhibitory action of CLA is far moreeffec-tive in ER positive MCF-7 cells ascomparedto theER negative MDA-MB -231 cells. Furthermore,these studiesalsodemonstratethat thegrowth inhibi-tory effectof CLA onMCF-7cellsis reversible.Morethan70%of theMCF-7cellstreatedwith CLA remainin the G0/G1 phaseas compared to control. Theseresults also demonstratethat there was a decreasein theexpression of c-myc in theMCF-7 cells treatedwith CLA as compared to the cells treated withLA and control. CLA is a natural constituent ofmany foods with oncostatic properties which mayhaveclinical applicationsfor the treatment and pre-vention of hormonedependentbreastcancer. Basedon these experimental results we hypothesizethat
CLA inhibits MCF-7 cell growth by interferingwith thehormoneregulatedmitogenicpathway.How-ever, further studies are warranted to establishtheinvolvement of hormone regulated receptors andgrowth factors to elucidatethe possiblemechanismin the growth inhibitory action of CLA on MCF-7cells.
Acknowledgements
We thank Dr Michael W. Pareza,Food ResearchInstitute, University of Wisconsin,Madison,WI forproviding the CLA sample andDr Kent C. Osborne,TheUniversity of TexasHealth ScienceCenteratSanAntonio for providing cancer cell lines usedin thisexperiment. This work wassupportedby grantsfromNational Institutes of Health (grantsAG10531 andDE10863)andtheRobert.J.Jr.andHelenC. KlebergFoundation of SanAntonio.
Fig. 6. (A) Northernblot analysisof c-myc mRNA isolatedfromMCF-7 cells incubatedfor 72 h in normal media (control) andmedia addedwith 3.5 × 10−5 M conjugatedlinoleic acid (CLA)and linoleic acid (LA). (B) Densitometric analysis.Video imageanalysiswasperformedusing the NIH image1.4 programon theautoradiograms shownin (A).
Fig. 7. (A) Northernblot analysisof c-myc mRNA isolatedfromMDA-MB-231 cells incubatedfor 72 h in normalmedia(control)andmediaaddedwith 3.5 × 10−5 M conjugatedlinoleic acid(CLA)and linoleic acid (LA). (B) Densitometric analysis.Video imageanalysiswasperformedusing the NIH image1.4 programon theautoradiogramsshownin (A).
128 V.R. Durgam, G. Fernandes/ CancerLetters116 (1997)121–130

References
[1] Tillotson, J., Darzynkiewicz, Z., Cohen, L. and Ronai, J.(1994)Effectsof linoleic acid on breasttumor cell prolifera-tion areassociatedwith changesin p53proteinexpression.In:Proceedings of the Fourth Annual Symposiumon Diet andBreastCancer,Vol. 364,p. 175.Editor: K. Weisburger. Ple-num Press,New York (abstract).
[2] Lu, J.,Jiang,C., Fountaine,S. andThompson,H. (1995)Rasmay mediatemammarycancerpromotionby high fat, Nutr.Cancer,23, 283–290.
[3] Sumida,C., Graber,R. and Nunez,E. (1988) Role of fattyacidsin signaltransductionmodulatorsandmessengers,Pros-taglandinsLeukotrienes EssentialFatty Acids, 8, 612–616.
[4] Durgam,V.R. andFernandes,G. (1997)Roleof dietaryfattyacids on breast cancer: molecular approaches. Immunol.Infect. Dis., in press.
[5] Rao,G.A. andAbraham,S. (1976)Enhancedgrowth rateoftransplanted mammaryadenocarcinomainducedin C3Hmiceby dietary linoleate,J. Natl. CancerInst., 56, 431–432.
[6] Ip, C., Carter,C.A. andIp, M. (1985)Requirement of essen-tial fatty acids for mammarytumorigenesis in rats, CancerRes.,45, 1997–2001.
[7] Fernandes, G. and Venkatraman, J.T. (1991) Modulation ofbreastcancergrowth in nudemice by n-3 lipids. In: WorldReview of Nutrition and Dietetics, Vol. 66, pp. 488–503.Editors:A.P. Simopoulos, R.R. Kifer, R.E. Martin andS.M.Barlow. Karger,Basel.
[8] Fernandes, G. and Venkatraman, J.T. (1990) Micronutrientsand lipid interactions in cancer. In: Micro Nutrients andImmuneFunctions. Editors: A. Bendichand R.K. Chendra.Ann. N. Y. Acad.Sci., 587,78-91.
[9] Charles,W.H., Michael,R.B.K. andJohn,P.N. (1995)Diet-ary q-3 fats reducesproliferation and tumor yield at color-ectal anastomosis in rats, Gastroenterology, 109, 431–439.
[10] Jurkowski, J.J. and Cave, W.T. (1995) Dietary effects ofmenhadenoil on thegrowthandmembranelipid compositionof rat mammarytumors,J.Natl. CancerInst.,74,1145–1150.
[11] Fernandes, G. and Venkatraman, J.T. (1992) Possiblemechanisms throughwhich dietarylipids andcalorierestric-tion modulatethe growthof breastcancer.In: Exercise,Cal-ories,Fat andCancer,Vol. 322, pp. 185–201. Editor: M.M.Jacobs.PlenumPress,Washington,DC.
[12] Rose, D.P. and Conally, J.M. (1990) Effect of fatty acidinhibitors of eiconsanoid synthesis on the growth of ahumanbreastcancercell line in culture, CancerRes.,50,7139–7144.
[13] Croft, K.D., Bellin, L.J., Legge, F.M. and Vandongen,R.(1987) Effect of diet enriched in eicosapentanoic acid ordocosohexanoic acid on prostanoid metabolism in rats,Lipids, 22, 647–651.
[14] Brooks,S.C.,Locke, E.R. and Soule,H.D. (1993) Estrogenreceptorin humanbreasttumorcell line (MCF-7) from breastcarcinoma,J. Biol. Chem.,248,6251–6256.
[15] Horwitz, K.B. and McGurie, W.L. (1978) Estrogencontrolprogesteronereceptorin humanbreastcancercorrelationwith
nuclearprocessingof estrogenreceptor,J. Biol. Chem.,253,2228–2233.
[16] Weigel,R.J.anddeConinck,E.C.(1993)Transcriptionalcon-trol of estrogenreceptorin estrogenreceptornegativebreastcarcinoma,CancerRes.,53, 3472–3474.
[17] Dubik, D., Dembiniski, T.C. andShiu,R. (1987)Stimulationof c-myc oncogeneexpressionassociatedwith estrogeninducedproliferation of humanbreastcancercells, CancerRes.,47, 6517–6521.
[18] Greenberg,M.E. and Ziff, E.B. (1984) Stimulation of 3T3cells induces transcription of the c-fos protooncogene,Nature,311,433–438.
[19] Mitsuhashi,N., Takano,A. andKato, J. (1986)Inhibition ofthe binding of R-5020andrat uterineprogesteronereceptorsby long chain fatty acids,Endocrinol.Jpn.,33, 93–96.
[20] Pariza,M.W., Loretz,L., Storkson, J.andHolland,N. (1983)Mutagensand modulatorsof mutagenesisin fried groundbeef,CancerRes.,43, 244.
[21] Ip, C., Chin, S., Scimeca,J. andPariza,M.W. (1991)Mam-mary cancerpreventionby conjugated linoleic acid, CancerRes.,51, 6118–6124.
[22] Ha, Y.L., Storkson,J. andPariza,M.W. (1990)Inhibition ofBenzo(o)pyreneinduced mouse forestomach neoplasia byconjugateddienoicderivativesof linoleic acid, CancerRes.,50, 1097–1101.
[23] Ip, C., Singh, M., Thompson,H. and Scimeca,J. (1994)Effect of CLA on carcinogenesis. In: Diet andBreastCancer,Vol. 364,pp. 59–65. Editor: K.E. Weisburge.PlenumPress,Washington,DC.
[24] Schultz,T., Chew, B. and Seaman,W. (1992) Differentialstimulatoryandinhibitory responsesof humanMCF-7 breastcancercells to linoleic acid and conjugatedlinoleic acid inculture,AnticancerRes.,12, 21–43.
[25] Chomczynski, P.andSacchi,N. (1987)SinglestepmethodofRNA isolation by acid guanidiumthiocynate-phenol-chloro-form extraction,Anal. Chem,162,156–159.
[26] Krishan,A. (1975)Rapidflow cytometricanalysisof mam-maliancellscycleby propidiumiodidestaining,J.Cell Biol.,66, 188–195.
[27] Shultz,T.D., Chew,B.P.,Seaman,W.R. andLuedecke,L.O.(1992)Inhibitory effectof linoleic acidandb-caroteneon thein vitro growthof humancancercells,CancerLett., 63,125–133.
[28] Rose,D.P. and Conally, J.M. (1993) Effects of dietary q-3fatty acidson humanbreastcancergrowth andmetastasis innudemice, J. Natl. CancerInst., 85, 1743–1747.
[29] Osborne,K.C., Hobbs,K. and Gary, M.C. (1985) Effect ofestrogenandanti estrogenon growthof humanbreastcancercells in athymicnudemice, CancerRes.,45, 584–590.
[30] Stiles, C.D. (1983) The molecularbiology of cell cycle ofPGDF,Cell, 33, 653–655.
[31] Blanchard,J.M.,Piechaczyk,K.M., Dani,C., Chanbard,J.C.,Franchhi,A., Pouyssegur, J. and Jeantur,P. (1985) c-mycgene is transcribed at high rate in G0 arrestedfibroblastsand is post transcriptionally regulatedin responseto growthfactors,Nature,317,443–445.
[32] Escot,C., Theilet,C., Lidereau,R., Spyratos, F., Chaampene,
129V.R. Durgam, G. Fernandes/ CancerLetters116 (1997)121–130

M., Gest,J.andCallahan,R. (1986)Geneticalterationsof thec-myc protooncogene (MYC) in human primary breastcarcinoma,Proc. Natl. Acad. Sci. USA, 83, 4834–4838.
[33] Chan,P.C.,Head,J.F.,Cohen,L.A. andWynder,E.L. (1977)Influenceof dietaryfat on the inductionof mammarytumorsby N-nitrosomethylurea: associated hormone changesanddifferencesbetweenSpragueDawley andF344rats,J. Natl.CancerInst., 59, 1279–1283.
[34] Feldman,J.M. and Hilf, R. (1985) A role of estrogensandinsulin binding in the dietary lipid alterationof R3230ACmammary carcinoma growth in rats, Cancer Res., 45,1964–1972.
[35] Feldman,J.M. andHilf, R. (1985)Effect of diabeteson thedietary lipid alteration of R3230AC mammary carcinomagrowth in rats, Proc. Soc. Exp. Biol. Med., 179, 105–112.
[36] Santos,G.F.,Scott,G.K., Lee,W.M.F., Liu, E. andBenz,C.(1988) Estrogeninducedpost transcriptional modulationof
c-myc protooncogene expressionin human breast cancercells, J. Biol. Chem.,263,95–99.
[37] Fernandes,G., Yunis, E.J. and Good,R.A. (1976) Suppres-sion of adenocarcinoma by the immunological consequencesof calorie restriction,Nature,263,504–507.
[38] Fernandes,G.,Chandrasekar,B., Troyer,D.A., Venkatraman,J.T. andGood,R.A. (1995)Dietary lipids andenergyintakeaffects mammarytumor incidenceand gene expressioninMMTV/v-H a-ras, Proc. Natl. Acad. Sci. USA, 92, 6494–6498.
[39] Huang, Y.C., Lloyd, M.S., Leudecke, O. and Shultz, D.P.(1994)Effect of cheddarcheeseconsumptionon plasmacon-jugatedlinoleic acid concentrations in men, Nutr. Res.,14,373–386.
[40] Knekt,P.,Jarvinon,R.S.,Pukkala,E. andAromea, A. (1996)Intake of dairy products and risk of breastcancer,Br. J.Cancer,73, 687–691.
130 V.R. Durgam, G. Fernandes/ CancerLetters116 (1997)121–130





























