Embed Size (px)
Citation preview

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Eugeniya]On: 13 October 2010Access details: Access Details: [subscription number 927989511]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of Natural HistoryPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713192031
The genus Eurycercus Baird, 1843 (Cladocera: Eurycercidae) in theNeotropicsEugeniya I. Bekkera; Alexey A. Kotova; Lourdes M. A. Elmoor-Loureirob
a A.N. Severtsov Institute of Ecology and Evolution, Moscow, Russia b Laboratyrio de Zoologia,Universidade Católica de Brasília, Taguatinga, DF, Brazil
Online publication date: 12 October 2010
To cite this Article Bekker, Eugeniya I. , Kotov, Alexey A. and Elmoor-Loureiro, Lourdes M. A.(2010) 'The genusEurycercus Baird, 1843 (Cladocera: Eurycercidae) in the Neotropics', Journal of Natural History, 44: 41, 2481 — 2508To link to this Article: DOI: 10.1080/00222933.2010.488752URL: http://dx.doi.org/10.1080/00222933.2010.488752
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Journal of Natural HistoryVol. 44, Nos. 41–42, November 2010, 2481–2508
ISSN 0022-2933 print/ISSN 1464-5262 online© 2010 Taylor & FrancisDOI: 10.1080/00222933.2010.488752http://www.informaworld.com
TNAH0022-29331464-5262Journal of Natural History, Vol. 1, No. 1, Sep 2010: pp. 0–0Journal of Natural HistoryThe genus Eurycercus Baird, 1843 (Cladocera: Eurycercidae) in the NeotropicsJournal of Natural HistoryE.I. BekkerEugeniya I. Bekkera*, Alexey A. Kotova and Lourdes M.A. Elmoor-Loureirob
aA.N. Severtsov Institute of Ecology and Evolution, Leninsky Prospect 33, Moscow 119071, Russia; bLaboratyrio de Zoologia, Universidade Católica de Brasília, QS 7 lote 1, Bloco M Sala 331, 71966–700, Taguatinga, DF, Brazil
(Received 7 December 2009; final version received 22 April 2010)
In Brazil and Columbia, we found three different species of Eurycercus Baird, 1843(Cladocera: Eurycercidae), belonging to two different subgenera: (1) E. (Eurycercus)cf. lamellatus (O. F. Müller, 1776), (2) E. (Bullatifrons) meridionalis sp. nov. and(3) E. (Bullatifrons) dumonti sp. nov. Appearance of the first species in Brazilcould be a result of introduction due to human activities. The two other speciesare apparently Neotropical endemics. There is no opportunity to associate thepreviously described E. lamellatus var. minuta Birabén, 1939 with any taxa foundby us, because the type material is lost and no eurycercids are present in its typelocality now; therefore this taxon is species incorrigenda with minimal chanceof being re-evaluated.
Keywords: Branchiopoda; Anomopoda; taxonomy; new species; South America
Introduction
Having analysed the previous literature, Forró et al. (2008) found that the number ofrecently known cladoceran species and endemics is significantly smaller in the Neo-tropics than in the Palaearctic (189 vs 245 and 73 vs 119, respectively). Mostly this isa consequence of an apparently insufficient study of South American fauna insteadof its “poorness”. A series of very detailed descriptions and re-descriptions of theCladocera was made by Paggi (1992, 1996, 1999). Recent investigations revealedadditional new, mostly endemic, taxa (Kotov and Elmoor-Loureiro 2008; Kotov et al.2010; Sinev and Elmoor-Loureiro 2010); some “old” species were also re-evaluatedand re-described (Van Damme et al. 2005; Kotov 2009), but there are many revisionsstill required before study of the Neotropical fauna can be considered adequate. Atpresent, the best studied region of the Neotropics is its northern border because ofthe efforts of Mexican researchers (Elías-Gutiérrez et al. 2006, 2008). However, manyregions of South America are problematic for freshwater biodiversity assessment.
A peculiarity of recent studies of the Neotropical cladocerans is the great attentionpaid to the family Chydoridae Dybowski et Grochowski, 1894. Recent “morphological”taxonomists have concentrated on this group, which is the most diverse among theAnomopoda and Cladocera in toto (Forró et al. 2008). Another related family,Eurycercidae Kurz, 1875 sensu Dumont and Silva-Briano, 1998 with a single genusEurycercus Baird, 1843, has not attracted the attention of researchers in the Neotropics
*Corresponding author. Email: [email protected]
Downloaded By: [Eugeniya] At: 10:42 13 October 2010

2482 E.I. Bekker et al.
since Birabén (1939). Paggi (1995) pointed out that Eurycercus lamellatus var. minutaBirabén, 1939 is most probably a separate taxon. But Eurycercus, not common in theNeotropics, has not previously been studied in detail. Before this study, there wereonly a few records of Eurycercus in temperate South America, all of them as Palae-arctic Eurycercus (Eurycercus) lamellatus (O.F. Müller, 1785). In Brazil, E. lamellatuswas recorded twice from the Southern Region (Montú and Gloeden 1986; Serafim-Junior et al. 2003).
In contrast, since the pioneer Frey (1973) publication, knowledge on the taxon-omy and distribution of the genus Eurycercus Baird, 1843 in the Holarctic has greatlyadvanced. The genus was subdivided into three subgenera (Frey 1975), a series ofnew species was found (Frey 1973, 1975, 1978; Hann 1982, 1990), and some “old”taxa were re-described (Hann 1990; Kotov 2000b). Frey (1995) declared that SouthAfrican populations belong to a separate species, but this taxon was not formallydescribed. Also Eurycercus has played an important role in studies of functional mor-phology (Fryer 1963; Kotov 1998) and life history (Hann 1985), and is the onlycladoceran genus for which the differentiation and subsequent development of thehead and each thoracic appendage has been studied in embryogenesis (Kotov 1995).Dumont and Silva-Briano (1998) suggested that the subfamily rank of EurycercinaeKurz, 1875 should be raised to family Eurycercidae.
Our study of a limited number of populations from Brazil and Columbia (weunderline once more that Eurycercus is a rare genus in the Neotropics), revealedthree different species, described in the present paper, belonging to two differentsubgenera, Eurycercus (Eurycercus) Baird, 1843 and Eurycercus (Bullatifrons) Frey,1975.
Materials and methods
The sample containing the type series of E. meridionalis sp. nov. was collected with aplankton net (130-µm mesh) attached to a handle and with a wide-mesh (1 cm) coverover its opening. The net was carried through the macrophytes. The sample was fixedwith 70% ethanol. Other samples were obtained from our colleagues.
Specimens were selected from all samples under a binocular stereoscopic micro-scope, placed on slides and studied whole under a high-power optical microscope. Atleast five parthenogenetic females from each locality (where possible) were dissectedfor analysis of appendages. Drawings were prepared using a camera lucida attachedto an Alphaphot compound microscope.
A system of enumeration for different setae on thoracic limbs proposed by Kotov(2000a) for alonine chydorids and then tested for other anomopods (Kotov et al.2010), including Eurycercus (Kotov 2000b), was used.
In general, we followed the morphometric measurements suggested by Hann(1982). At the same time, the terminology used here is different from that of Frey(1975, 1978) and Hann (1982, 1990), but follows Kotov (2000a, b) which co-ordinatesbetter with that used for other groups of the Anomopoda; abbreviations for morpho-logical structures are taken predominantly from Kotov et al. (2002). Also, regressionstatistics for species’ discrimination (Hann 1982), requiring dissections of many spec-imens, was not possible given our relatively limited and unique material, sometimesobtained from remote localities. For our analysis of instar variability of some charac-ters, we dissected only the five smallest and five largest females of each species, and
Downloaded By: [Eugeniya] At: 10:42 13 October 2010

Journal of Natural History 2483
compared ranges of obtained measurements. If a parameter was not measured for aspecimen (i.e. IP or MHP because of unsuccessful dissection), we did not try to dis-sect more specimens to obtain additional information, preferring to save as manyundissected animals as possible as paratypes. Therefore, we can only comment oninstar variability of some characters when there is a very obvious difference betweenthe smallest and largest females. We preferred to search for better characters forspecies discrimination than the morphometric regressions suggested by Hann (1982),and we believe that we have found them.
Abbreviations
Collections: AAK, personal collection of A. A. Kotov, Moscow, Russia; EL, personalcollection of L. M. A. Elmoor-Loureiro, Taguatinga, Brazil; MCN, Museu de CiênciasNaturais, Fundação Zoobotânica do Rio Grande do Sul, Porto Alegre, Brazil;MGU, Zoological Museum of M. V. Lomonosov Moscow State University, Moscow,Russia; MNRJ, Museu Nacional/UFRJ, Rio de Janeiro, Brazil.
In the text and illustrations: IDL, inner distal lobe of limb I; IP, interpore distance;ODL, outer distal lobe of limb I.
For measurements: BL, body length; BH, body height; HL, head length; HW,head width; MHP, major head pore diameter; IP, interpore distance; VL, valvelength; PL, postabdomen length; PH, postabdomen height; NT, number of preanalteeth; CL, claw length; DS, length of more distal spine on base of postabdominalclaw; BS, length of more basal spine on base of postabdominal claw; AL, antenna Ilength; DA, diameter of antenna I.
Systematics
Eurycercus (Eurycercus) cf. lamellatus (O. F. Müller, 1776)(Figure 1)
Material studied
One juvenile female from lower reaches of Rio Nhamundá, Central Amazonia, Pará,Brazil, AAK 2003-030.
Short description of single juvenile female
In lateral view body subrectangular (Figure 1A). Dorsal margin interrupted by arelatively shallow depression behind major head pore. In anterior view, body moder-ately compressed laterally, median dorsal keel well developed, although not sharp(Figure 1B). Head with relatively long rostrum. Compound eye relatively small,ocellus was not found, but the pigment could have been dissolved by irregularities inthe preservation technique. A single major “head pore” as a ringed, suboval field ofspecial cuticle; a minute, elongated ovoid lateral pore located at either side of majorpore, closer to it (Figure 1C). Labrum with a medium-sized median keel, terminatingin a broadly rounded apex. Postabdomen with subparallel dorsal and ventral margins.Distal anal embayment shallow. Armature of the preanal margin as a series of 101
Downloaded By: [Eugeniya] At: 10:42 13 October 2010
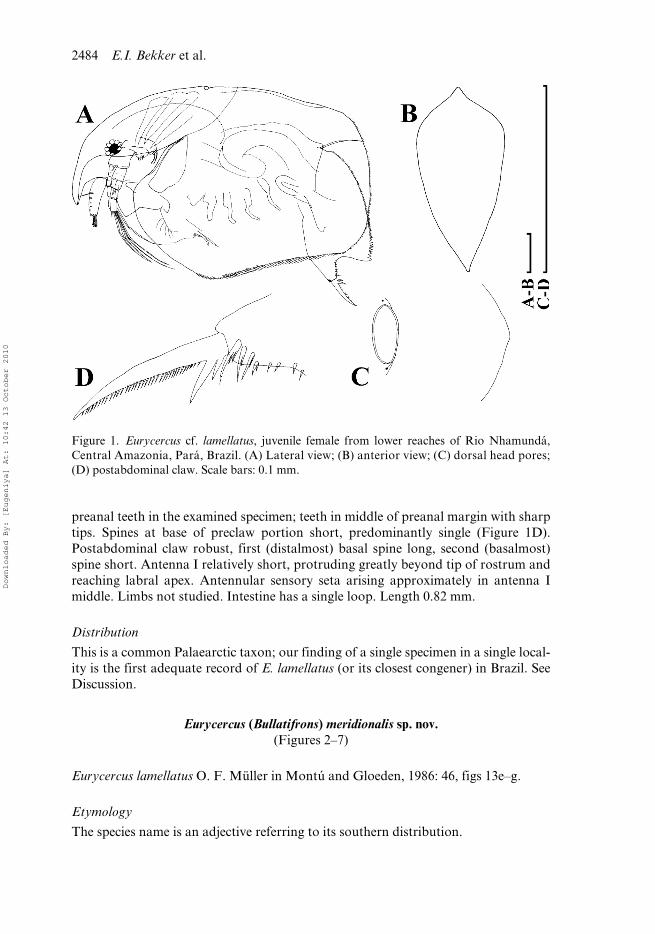
2484 E.I. Bekker et al.
preanal teeth in the examined specimen; teeth in middle of preanal margin with sharptips. Spines at base of preclaw portion short, predominantly single (Figure 1D).Postabdominal claw robust, first (distalmost) basal spine long, second (basalmost)spine short. Antenna I relatively short, protruding greatly beyond tip of rostrum andreaching labral apex. Antennular sensory seta arising approximately in antenna Imiddle. Limbs not studied. Intestine has a single loop. Length 0.82 mm.
Distribution
This is a common Palaearctic taxon; our finding of a single specimen in a single local-ity is the first adequate record of E. lamellatus (or its closest congener) in Brazil. SeeDiscussion.
Eurycercus (Bullatifrons) meridionalis sp. nov.(Figures 2–7)
Eurycercus lamellatus O. F. Müller in Montú and Gloeden, 1986: 46, figs 13e–g.
Etymology
The species name is an adjective referring to its southern distribution.
Figure 1. Eurycercus cf. lamellatus, juvenile female from lower reaches of Rio Nhamundá,Central Amazonia, Pará, Brazil. (A) Lateral view; (B) anterior view; (C) dorsal head pores;(D) postabdominal claw. Scale bars: 0.1 mm.
Downloaded By: [Eugeniya] At: 10:42 13 October 2010

Journal of Natural History 2485
Type locality
Canal de Sarita, Estação Ecológica do Taim, Rio Grande do Sul, Brazil, coordinates:32.6408° S, 52.62333° W. The type series was collected on 29 December 2001 byL.M.A. Elmoor-Loureiro.
The Estação Ecológica do Taim is located within a larger area called the TaimHydrological System, at the extreme south of Brazil (32°30′ to 32°40′S, 52°20′ to52°40′ W). The reserve encompasses a variety of habitats such as beaches, dunes, for-ests, grasslands and, especially, lakes and wetlands. The complex of lakes andwetlands is locally known as “banhado”, where the emergent macrophytes couldcover more than 30% of the area, with predominance of Scirpus californicus, Scirpusgiganteus and Zizaniopsis bonariensis; among the fluctuant macrophytes, Salviniaherzogii, Azolla caroliniana and Lemma valdiviana are the most frequent (Paz 2003).The climate of the region is subtropical humid or type Cfa, according to the Köppenclassification (Waechter and Jarenkow 1998).
Type material
Holotype. Adult parthenogenetic female, mounted on a slide in PVL, MNRJ20411.
Paratypes. Three females in alcohol, MNRJ20410; three females in alcohol,MCN2554; two adult parthenogenetic females, AAK M-0904; 11 dissected partheno-genetic females on slides EL00890-00900 and EL00904.
Other material studied
Ten females from an artificial pond caused by sand mining at Parque Municipal doIguaçu, Curitiba, Estado do Paraná, Brazil, collected on 12 May 2006 by G. P. Neves,AAK M-0905. Coordinates: 25°31′48″ S, 49°13′22″ W.
The Parque Municipal do Iguaçu, located at the Metropolitan Region of Curit-iba, has a complex of natural and artificial ponds, marginal to the upper Iguaçu River;large macrophyte stands, particularly of the genus Eichhornia and Pistia, occur at theedge of the ponds (Abilhoa and Agostinho 2007). The specimens of E. meridionaliswere sampled from an artificial pond, formed in a depression in the ground, causedby sand mining. For ponds of Parque Municipal do Iguaçu, Abilhoa and Agostinho(2007) reported pH between 6.8 and 7.8, and concentration of dissolved oxygenbetween 4.35 and 6.73 mg/l. The climate in both regions is also subtropical humid ortype Cfa, according to the Köppen classification, with mean temperature between10°C and 22°C (Abilhoa and Agostinho 2007).
Diagnosis
Adult parthenogenetic female. Dorsal margin with a dorsal bubble bearing headpores. In anterior view, body wide, median dorsal keel absent. Rostrum short; poste-rior portion of head shield with a distinct straight section of the margin immediatelyanterior to the point of mandibular articulation; lateral pores elongated. Labral keelmedium-sized, terminating in a broadly rounded apex. Postabdomen with a shallowdistal anal embayment, with 102–122 preanal teeth. Teeth at base of preclaw portionshort, predominantly clustered (doubled or tripled). Antenna I relatively short, butprotruding greatly beyond tip of rostrum; antennular sensory seta arising somewhat
Downloaded By: [Eugeniya] At: 10:42 13 October 2010

2486 E.I. Bekker et al.
basally to antenna I middle; denticles in rows encircling antennular surface speciallysmall. IDL of limb I with three bisegmented setae, among them, a remarkable stronghook-like seta, but not as large as in E. macracanthus. IDL supplied with five to ninelong distal spinules, six to ten long proximal spinules, four to seven very short mar-ginal spinules and seven to nine short basal spinules. Limb II scraper 5 with 20–25denticles, scraper 6 with 18–23 denticles, scraper 7 with 19–23 denticles; filter platewith eight setae. Nine setae in filter plate III; eight setae in filter plate IV, eight setaein filter plate V. Intestine with a single loop. Length up to 2.02 mm.
Description
Parthenogenetic female. In lateral view body subovoid in larger females (Figures 2A,E,3A) and subrectangular in small females (Figures 2F, 3B), maximum height of thebody in its middle portion (BH/BL = 0.53–0.68 in juveniles, 0.54–0.69 in largestadults). Dorsal margin evenly convex, interrupted only by a bubble-like head pore(Figures 2B, 3C). Posterodorsal angle expressed, more or less rounded in both adultsand juveniles. Posterior margin slightly convex, smoothly rounded, posteroventralangle broadly rounded. In larger adults ventral margin with a slight prominenceimmediately anterior to the margin middle. In anterior view, body wide, not compressedlaterally, maximum width of body at level of mandibular articulation. Median dorsalkeel is absent. Parthenogenetic females carry few eggs in the brood pouch.
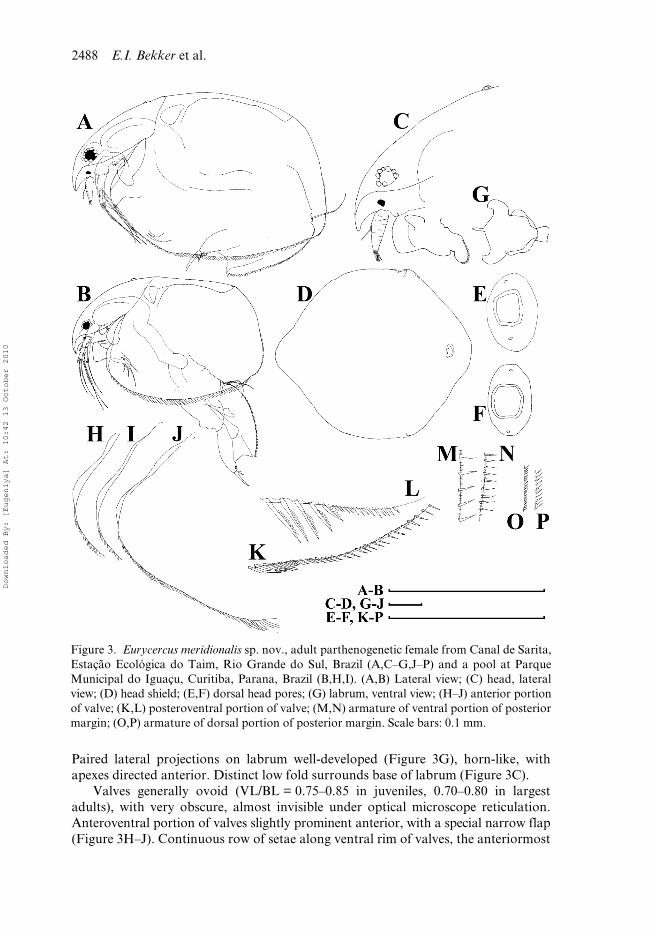
Head (Figure 3C) large, with dorsal margin regularly arched from rostrum toregion of dorsal head pores. Border line between head shield and valves obscure inpreserved animals. Rostrum well expressed, short. Frontal head pore as transversesplit, located somewhat anteriad to antennule bases on ventral surface of head(Figure 2H,I). Compound eye rather large, located near dorsal margin of head markedlycloser to rostral extremity than to head pores. Ocellus small, located at antennulebase, closer to eye than to tip of rostrum. Head shield little longer than wide (HW/HL = 0.82–0.83 in juveniles, 0.76–0.91 in largest adults), with maximum height atlevel of mandibular articulation. Its anterior portion (= portion anterior to level ofmandibular articulation) more than twice as large as posterior one (Figure 3D),broadly rounded with slightly projected rostral region; posterior portion with adistinct straight section of the margin immediately anterior to the point of mandibulararticulation (Figure 3D). A single major “head pore” (dorsal organ) as a ringed,suboval field of special cuticle located on a dorsal bubble (Figures 2B,C),MHP = 0.022 mm in juveniles, 0.022–0.024 mm in largest adults. A minute, elongatedovoid lateral pore located at either side of major pore, closer to it (Figures 2C, 3E,F),distance between lateral head pores growing proportionally with growth of headshield (IP/HW = 0.06–0.08 in juveniles, 0.05–0.07 in largest adults). Head pores“projecting from the rounded head like a bubble with the median pore at the top” is adiagnostic trait of the subgenus Bullatifrons according to Frey (1975: 293).
Labrum as a fleshy body, with a medium-sized median keel, terminating in abroadly rounded apex, keel anterior margin slightly convex, without setulation,posterior margin almost straight. Distal labral plate with rich setulation (Figure 3C).
margin of head, head pores and postabdomen; (E) adult; (F) juvenile; (G) instar variability ofpostabdomen general shape; (H) head, ventral view; (I) frontal head pore. Scale bars: 1 mm(A,E–G); 0.1 mm (B–D,H); 0.01 mm (I).
Downloaded By: [Eugeniya] At: 10:42 13 October 2010
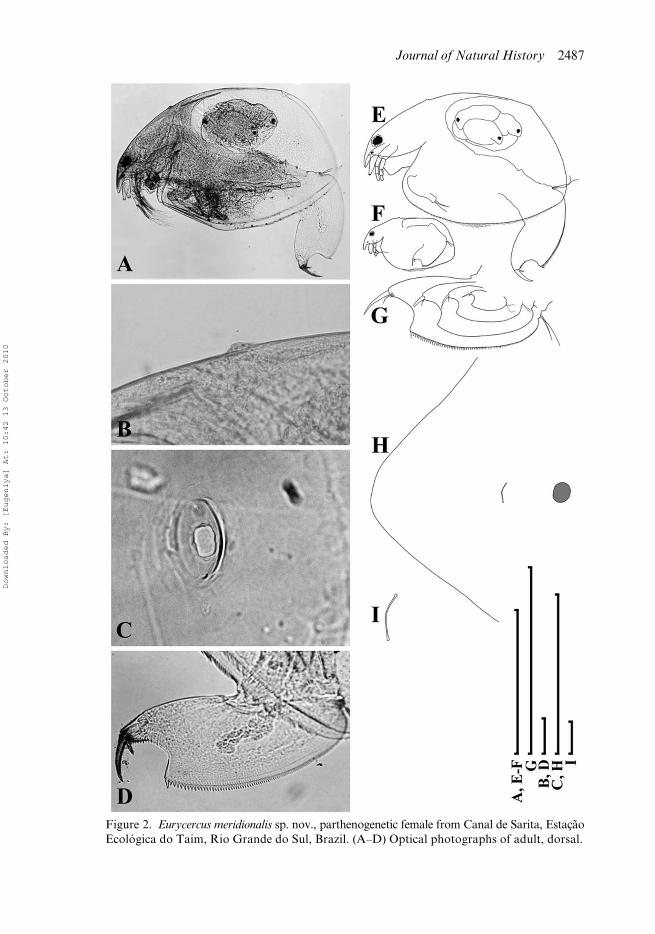
Journal of Natural History 2487
Figure 2. Eurycercus meridionalis sp. nov., parthenogenetic female from Canal de Sarita, EstaçãoEcológica do Taim, Rio Grande do Sul, Brazil. (A–D) Optical photographs of adult, dorsal.
Downloaded By: [Eugeniya] At: 10:42 13 October 2010
2488 E.I. Bekker et al.
Paired lateral projections on labrum well-developed (Figure 3G), horn-like, withapexes directed anterior. Distinct low fold surrounds base of labrum (Figure 3C).
Valves generally ovoid (VL/BL = 0.75–0.85 in juveniles, 0.70–0.80 in largestadults), with very obscure, almost invisible under optical microscope reticulation.Anteroventral portion of valves slightly prominent anterior, with a special narrow flap(Figure 3H–J). Continuous row of setae along ventral rim of valves, the anteriormost
Figure 3. Eurycercus meridionalis sp. nov., adult parthenogenetic female from Canal de Sarita,Estação Ecológica do Taim, Rio Grande do Sul, Brazil (A,C–G,J–P) and a pool at ParqueMunicipal do Iguaçu, Curitiba, Parana, Brazil (B,H,I). (A,B) Lateral view; (C) head, lateralview; (D) head shield; (E,F) dorsal head pores; (G) labrum, ventral view; (H–J) anterior portionof valve; (K,L) posteroventral portion of valve; (M,N) armature of ventral portion of posteriormargin; (O,P) armature of dorsal portion of posterior margin. Scale bars: 0.1 mm.
Downloaded By: [Eugeniya] At: 10:42 13 October 2010

Journal of Natural History 2489
members short (Figure 3A), setae then sharply increasing in size posterior to theprominence on ventral margin, and finally gradually decreasing in size to posteroventralvalve portion. Posteroventral angle with a row of thin spinules, small denticlesbetween them (Figure 3K,L). This row continues to ventral portion of posteriormargin (Figure 3M,N), whereas dorsal portion of posterior margin armed with small,curved, densely located spinules lacking setules between them (Figure 3O,P).
Thorax without external traces of segmentation, with six limb pairs. Abdomenthick, no abdominal projections on dorsal part of all segments (Figures 2A, 3B).
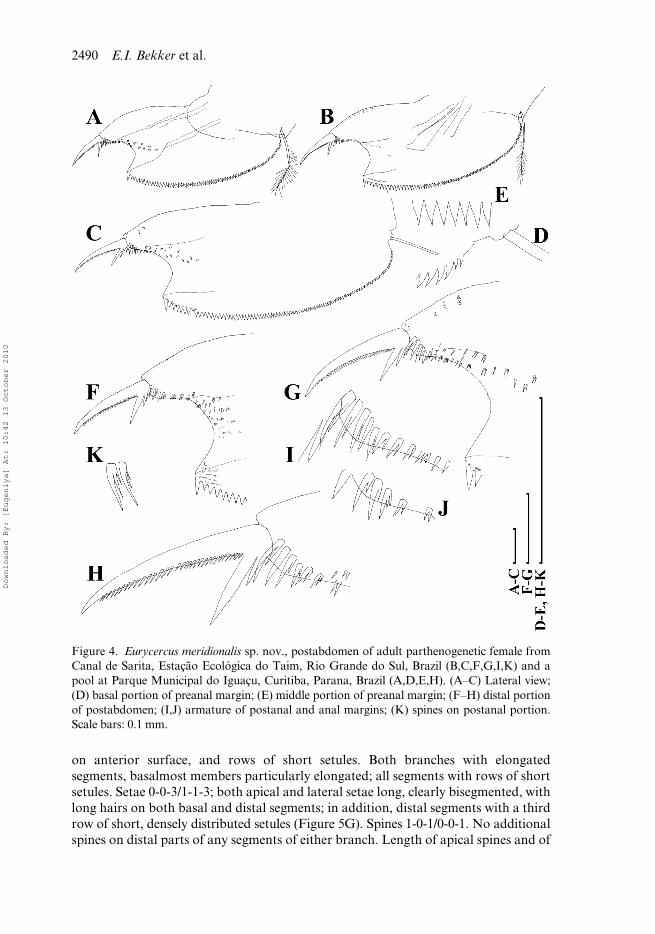
Postabdomen as a large (PL/TL = 0.33–0.40 in juveniles and the same in largestadults), broad (PH/PL = 0.55–0.65 in juveniles, 0.51–0.57 in largest adults), flattenedplate with subparallel dorsal and ventral margins (Figures 2D, 4A–C). Anus opensdistally; hence the whole slightly convex dorsal margin represents the preanal marginof postabdomen. Dorsodistal (preanal) angle well expressed, distal anal embaymentshallow, dorsal portion of distal (= anal) margin straight to slightly concave. Postanalangle obtuse, rounded (Figures 4F–J). Preclaw portion of postabdomen, named as“distal expansion” by Hann (1990), as a conical prominence. Ventral margin ofpostabdomen slightly convex, a conspicuous, chitinized thickening of integument dis-tally to claw base (Figure 4G). Armature of the preanal margin as a series of preanalteeth (NT = 78–96 in juveniles, 102–122 in largest adults), slightly and graduallyincreasing in size in distal direction; a small gap lacking any teeth (Figure 4D) at baseof postabdominal setae; teeth in middle of preanal margin with sharp tips (Figure 4E);distalmost tooth considerably larger than the others, located just on dorsodistal angleof postabdomen. On preclaw portion of postabdomen there are crescentic clusters ofspines (homologues of lateral setae or postanal teeth of chydorids), distalmost mem-bers particularly large, predominantly clustered (Figure 4I–K), teeth at base of pre-claw portion short, also predominantly clustered (doubled or triplet). Subparallelrows of minute setules on whole lateral surface of postabdomen.
Postabdominal setae short (about half of preanal margin length), bisegmented,distal segment shorter than basal one and bilaterally setulated. Setae are located on adistinct, nut-like base.
Postabdominal claw relatively robust (CL/PL = 0.29–0.33 in juveniles, 0.26–0.29in largest adults); with massive base; weakly and evenly tapered in distal direction,and slightly curved (Figure 4F–H). Two pectens of denticles on outer surface of dor-sal side of claws (Figure 4H). Two basal spines, first (distal) long (DS/CL = 0.31–0.45in juveniles, 0.31–0.37 in largest adults), second (basal) short (BS/CL = 0.15–0.21 injuveniles, 0.12–0.18 in largest adults; BS/DS = 0.32–0.52 in juveniles, 0.36–0.48 inlargest adults), located dorsally immediately at base of claw.
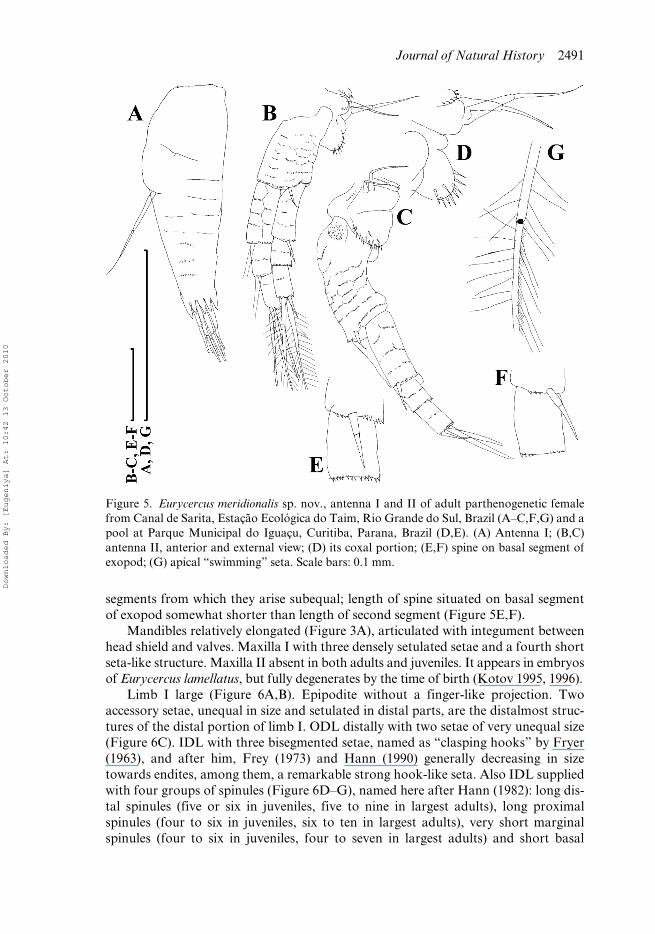
Antenna I (antennule) relatively short (AL/BL = 0.10–0.11 in juveniles, 0.09–0.12in largest adults; AL/DA = 2.7–3.0 in adults); with maximal width in basal half, withits distal two-thirds evenly tapering distally (Figure 5A); protruding greatly beyond tipof rostrum (Figure 3C). Slender antennular sensory seta long (about half antenna Ilength), arising somewhat basally to antenna I middle (Figure 5A). Nine bisegmentedaesthetascs, with pointed teeth around them. No setules at anterior margin ofantenna I. Numerous short rows of minute denticles encircling antennular surface.
Antenna II relatively short. Several projections on its external surface in coxalregion (Figure 5B–D). On one of them with two bisegmented setae, unequal in length,next projection with semi-circular row of setules; distalmost projection inflated, withnumerous strong spinules. Massive basal segment with a relatively long seta distally
Downloaded By: [Eugeniya] At: 10:42 13 October 2010
2490 E.I. Bekker et al.
on anterior surface, and rows of short setules. Both branches with elongatedsegments, basalmost members particularly elongated; all segments with rows of shortsetules. Setae 0-0-3/1-1-3; both apical and lateral setae long, clearly bisegmented, withlong hairs on both basal and distal segments; in addition, distal segments with a thirdrow of short, densely distributed setules (Figure 5G). Spines 1-0-1/0-0-1. No additionalspines on distal parts of any segments of either branch. Length of apical spines and of
Figure 4. Eurycercus meridionalis sp. nov., postabdomen of adult parthenogenetic female fromCanal de Sarita, Estação Ecológica do Taim, Rio Grande do Sul, Brazil (B,C,F,G,I,K) and apool at Parque Municipal do Iguaçu, Curitiba, Parana, Brazil (A,D,E,H). (A–C) Lateral view;(D) basal portion of preanal margin; (E) middle portion of preanal margin; (F–H) distal portionof postabdomen; (I,J) armature of postanal and anal margins; (K) spines on postanal portion.Scale bars: 0.1 mm.
Downloaded By: [Eugeniya] At: 10:42 13 October 2010
Journal of Natural History 2491
segments from which they arise subequal; length of spine situated on basal segmentof exopod somewhat shorter than length of second segment (Figure 5E,F).
Mandibles relatively elongated (Figure 3A), articulated with integument betweenhead shield and valves. Maxilla I with three densely setulated setae and a fourth shortseta-like structure. Maxilla II absent in both adults and juveniles. It appears in embryosof Eurycercus lamellatus, but fully degenerates by the time of birth (Kotov 1995, 1996).
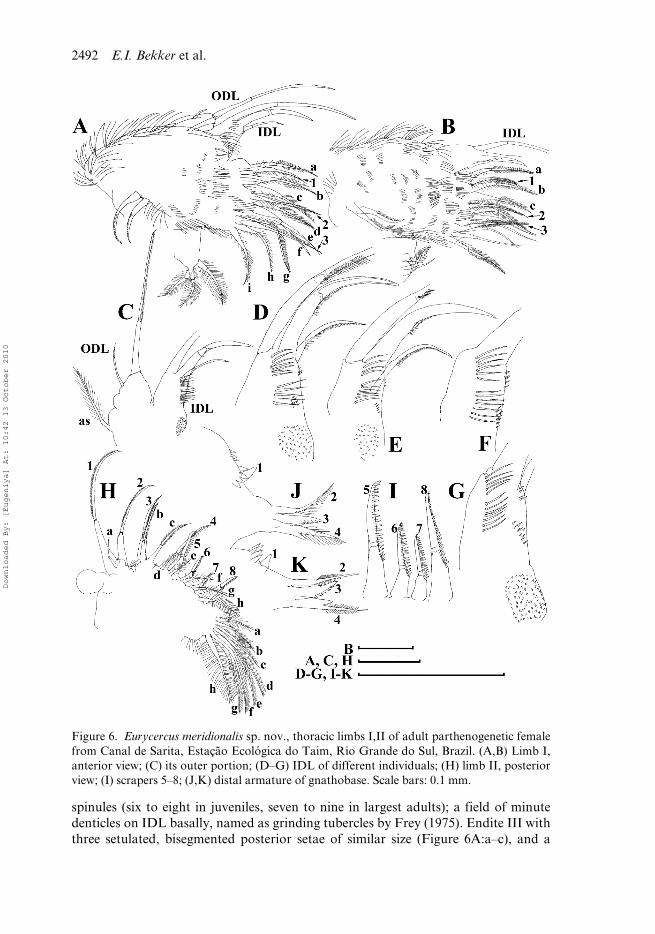
Limb I large (Figure 6A,B). Epipodite without a finger-like projection. Twoaccessory setae, unequal in size and setulated in distal parts, are the distalmost struc-tures of the distal portion of limb I. ODL distally with two setae of very unequal size(Figure 6C). IDL with three bisegmented setae, named as “clasping hooks” by Fryer(1963), and after him, Frey (1973) and Hann (1990) generally decreasing in sizetowards endites, among them, a remarkable strong hook-like seta. Also IDL suppliedwith four groups of spinules (Figure 6D–G), named here after Hann (1982): long dis-tal spinules (five or six in juveniles, five to nine in largest adults), long proximalspinules (four to six in juveniles, six to ten in largest adults), very short marginalspinules (four to six in juveniles, four to seven in largest adults) and short basal
Figure 5. Eurycercus meridionalis sp. nov., antenna I and II of adult parthenogenetic femalefrom Canal de Sarita, Estação Ecológica do Taim, Rio Grande do Sul, Brazil (A–C,F,G) and apool at Parque Municipal do Iguaçu, Curitiba, Parana, Brazil (D,E). (A) Antenna I; (B,C)antenna II, anterior and external view; (D) its coxal portion; (E,F) spine on basal segment ofexopod; (G) apical “swimming” seta. Scale bars: 0.1 mm.
Downloaded By: [Eugeniya] At: 10:42 13 October 2010
2492 E.I. Bekker et al.
spinules (six to eight in juveniles, seven to nine in largest adults); a field of minutedenticles on IDL basally, named as grinding tubercles by Frey (1975). Endite III withthree setulated, bisegmented posterior setae of similar size (Figure 6A:a–c), and a
Figure 6. Eurycercus meridionalis sp. nov., thoracic limbs I,II of adult parthenogenetic femalefrom Canal de Sarita, Estação Ecológica do Taim, Rio Grande do Sul, Brazil. (A,B) Limb I,anterior view; (C) its outer portion; (D–G) IDL of different individuals; (H) limb II, posteriorview; (I) scrapers 5–8; (J,K) distal armature of gnathobase. Scale bars: 0.1 mm.
Downloaded By: [Eugeniya] At: 10:42 13 October 2010

Journal of Natural History 2493
setulated, stiff anterior seta 1 with a small, slender sensillum near its base. Endite IIwith three posterior setae (Figure 6A:d–f) analogous to those on eIII, and a stiff,setulated anterior seta 2, a very small sensillum near its base. Endite I with threeposterior setae (g–i), and a stiff anterior seta 3. Two ejector hooks anteriorly on outerportion of limb corm (Figure 6A,B). Well-developed maxillar process, earlier shownin embryos to be a remainder of gnathobase I (Kotov 1995), bearing three slender,fully and densely setulated setae on inner side of limb base.
Limb II with ovoid epipodite lacking a finger-like projection; exopodite as a smalllobe (Figure 6H). At inner side of limb, a row of eight stiff scrapers; setae 1–3 withmore delicate feathering, setae 4–8 with relatively robust denticles (Figure 6I). Scraper5 with 15–19 denticles in juveniles and 20–25 denticles in largest adult; scraper 6 with15–17 denticles in juveniles and 18–23 denticles in largest adult; scraper 7 with 13–19denticles in juveniles and 19–23 denticles in largest adult. Posteriorly on limb corm 8soft setae: distalmost one (Figure 6H:a) short; next two (Figure 6H:b,c) longest,relatively stout, bilaterally armed in distal portions by short setules; basalmost ones(Figure 6H:d–h) similarly feathered by long hairs. Distal armature of gnathobasewith four setae (Figure 6J–K), one of them a minute sensillum (1), located far fromthe others, a row of denticles (four or five in juveniles, four to eight in largest adults)crossing near it. Filter plate with eight long, densely setulated setae; distalmost seta offilter plate clearly smaller than the others, second and third are slightly shorter thanothers. Additional bunch of setules basal to filter plate.
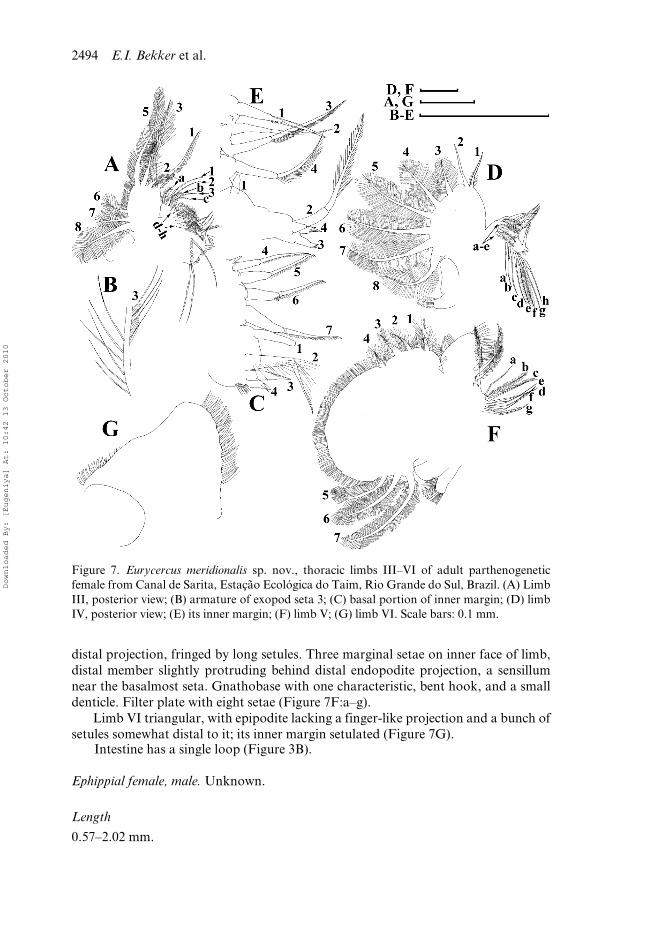
Limb III with relatively large epipodite lacking a finger-like projection (Figure 7A).Exopodite flat, smaller than those of limbs IV and V. Distally, five setae of unequalsize (Figure 7A:1–5), among them seta 3 with a special three-dimensional armature(supplied with two rows of strong setules and a row of delicate setules, Figure 7B);distal group consists of three setae (6–8), seta 8 especially long. Distal portion of innerlimb part forms a partly isolated lobe, distal endite sensu Kotov (2000a, b), externalendite sensu Dumont and Silva-Briano (1998) with three bisegmented anterior setae(1–3). Basal endite, internal endite sensu Dumont and Silva-Briano (1998) somewhatlarger than distal endite. Marginally, a row of four stiff setae (Figure 7C:4–7), a smallsensillum near seta 4. Seven long soft setae of subequal size (Figure 7C:a–h) on limbcorm posteriorly. Gnathobase weakly demarcated from basal endite, distal armaturewith four members, one of them (Figure 7C:1) a large, bottle-shaped sensillum locatedfar from the others. Nine setae in filter plate.
Limb IV with pre-epipodite as a setulated hillock; epipodite large, ovoid, without afinger-like projection (Figure 7D). Exopodite oval, with two distal, relatively stout,bisegmented setae of unequal size, armed by short setules (Figure 7D:1,2), other six setaefeathered bilaterally by very long hairs (Figure 7D:3–8). Marginally on inner limb face, arow of four stiff anterior setae (Figure 7E:1–4). Seta 1 shorter, setae 2–4 of equal size,feathered by long setules in distal part, small receptors near bases of setae 2 and 3. Poste-riorly, five soft setae (Figure 7D:a–e) with subequal length. Distal armature of gnatho-base with four members (Figure 7E:1–4). One of them a long, bisegmented seta, denselyfeathered in distal part (Figure 7E:2), two others small (Figure 7E:3,4), a large, bottle-shaped sensillum (Figure 7E:1) is a fourth member of gnathobasic armature (similar tolimb III). Filter plate IV with eight setae, middle ones longer than marginal ones.
Limb V with pre-epipodite as a setulated hillock; epipodite without a finger-likeprojection. Exopodite very large, with four short distal (Figure 7F:1–4) and threelarge lateral (Figure 7F:5–7) setae. Inner portion of limb with a protruding flap-like
Downloaded By: [Eugeniya] At: 10:42 13 October 2010
2494 E.I. Bekker et al.
distal projection, fringed by long setules. Three marginal setae on inner face of limb,distal member slightly protruding behind distal endopodite projection, a sensillumnear the basalmost seta. Gnathobase with one characteristic, bent hook, and a smalldenticle. Filter plate with eight setae (Figure 7F:a–g).
Limb VI triangular, with epipodite lacking a finger-like projection and a bunch ofsetules somewhat distal to it; its inner margin setulated (Figure 7G).
Intestine has a single loop (Figure 3B).
Ephippial female, male. Unknown.
Length
0.57–2.02 mm.
Figure 7. Eurycercus meridionalis sp. nov., thoracic limbs III–VI of adult parthenogeneticfemale from Canal de Sarita, Estação Ecológica do Taim, Rio Grande do Sul, Brazil. (A) LimbIII, posterior view; (B) armature of exopod seta 3; (C) basal portion of inner margin; (D) limbIV, posterior view; (E) its inner margin; (F) limb V; (G) limb VI. Scale bars: 0.1 mm.
Downloaded By: [Eugeniya] At: 10:42 13 October 2010

Journal of Natural History 2495
Differential diagnosis
See Table 1 and Discussion for differentiation of American species of Eurycercus(Bullatifrons).
Distribution
The species is distributed in the South Atlantic and the Upper Río Paraná basins ofBrazil (Estado do Paraná and Estado do Rio Grande do Sul).
Eurycercus (Bullatifrons) dumonti sp. nov.(Figures 8–11)
Etymology
This species is dedicated to Professor Henri J. Dumont, renowned Belgian investiga-tor of the Cladocera and other freshwater animals, who collected the type series.
Type locality
Lake Regadera near Bogotá, Columbia. Co-ordinates: 4.4000° S, 74.1500° W.
Type material
Holotype. Adult parthenogenetic female in 96% alcohol, MGU ML 97. Label of theholotype: “Eurycercus (Bullatifrons) dumonti sp.nov., 1 parth. � from Lake Regaderanear Bogota, Columbia, coll. in 16.09.1992 by H.J. Dumont, HOLOTYPE”.
Paratypes. Fifteen females, MGU ML 98; 10 females, AAK 2009-075.No hydrochemical information was taken during sampling. Lake Regadera
(Ebmolsa la Regadera) is a large, shallow, artificial water reservoir in the watersystem serving Bogota town, suffering from high anthropogenic pressure, i.e. organicpollution.
Other material studied
Eleven parthenogenetic females from Lagoa dos Patos, connected to Rio Invinhema,Paraná floodplain, Mato Grosso do Sul, Brazil, coll. in September 1992 by F. A.Lansac-Tôha. Coordinates: 22°49′33.66″ S, 53°33′09.9″ W.
Lagoa dos Patos is a small lagoon (c. 1.2 km2), with depths ranging between 2.8and 4.8 m, and connected by a permanent channel with Invinhema River, at theupper Paraná River floodplain; dense stands of aquatic macrophytes are present,particularly of the genera Eichhornia, Salvinia and Polygonum (Serafim-Junior et al.2003). Rodrigues et al. (2002) have investigated the limnological parameters ofPatos Lagoon, such as: dissolved oxygen (1.0–9.0 mg/l), pH (c. 5.8–7.5), conductivity(c. 20–50 μS/cm), total nitrogen (c. 250–1300 mg/l), total phosphorus (c. 10–90 μg/l).The climate of the region is subtropical humid or type Cfa, according to the Köppenclassification; the mean annual rainfall is 1500 mm, and the mean temperature isbelow 18°C in winter, with rare frosts, and above 22°C during warmer months(Souza et al. 2009).
Downloaded By: [Eugeniya] At: 10:42 13 October 2010

2496 E.I. Bekker et al.
Diagnosis
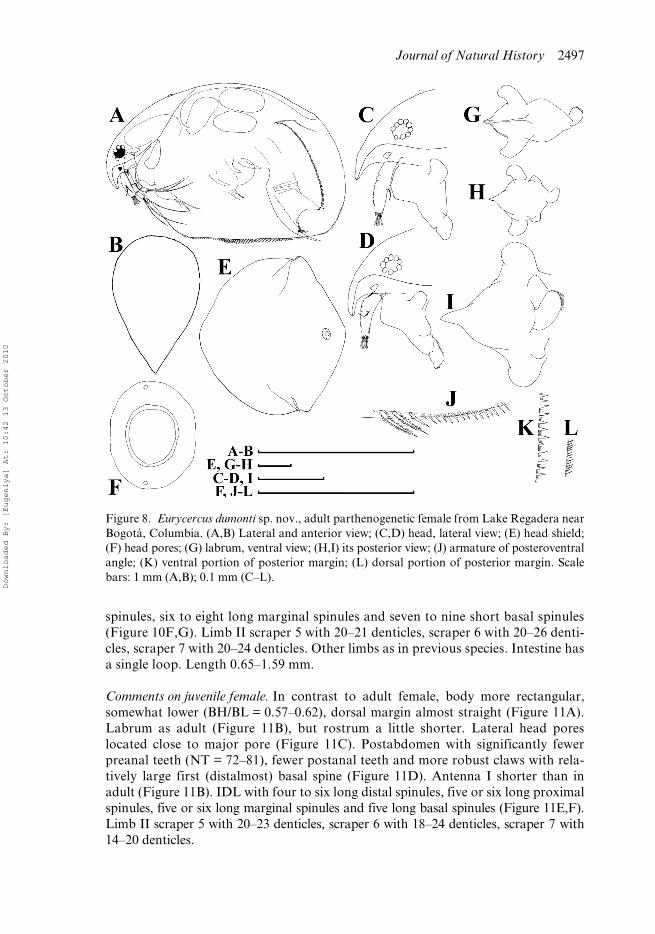
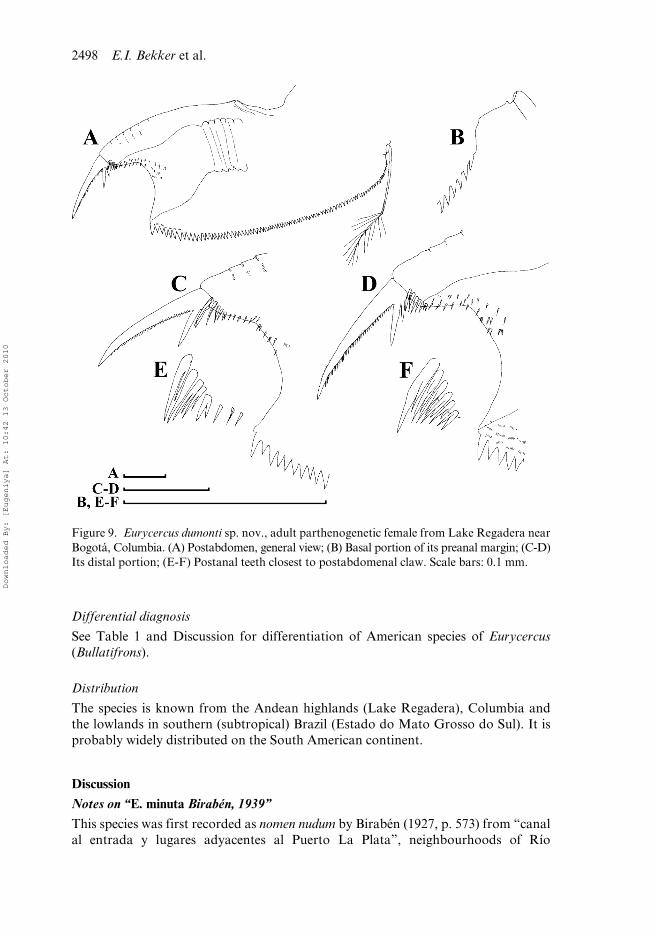
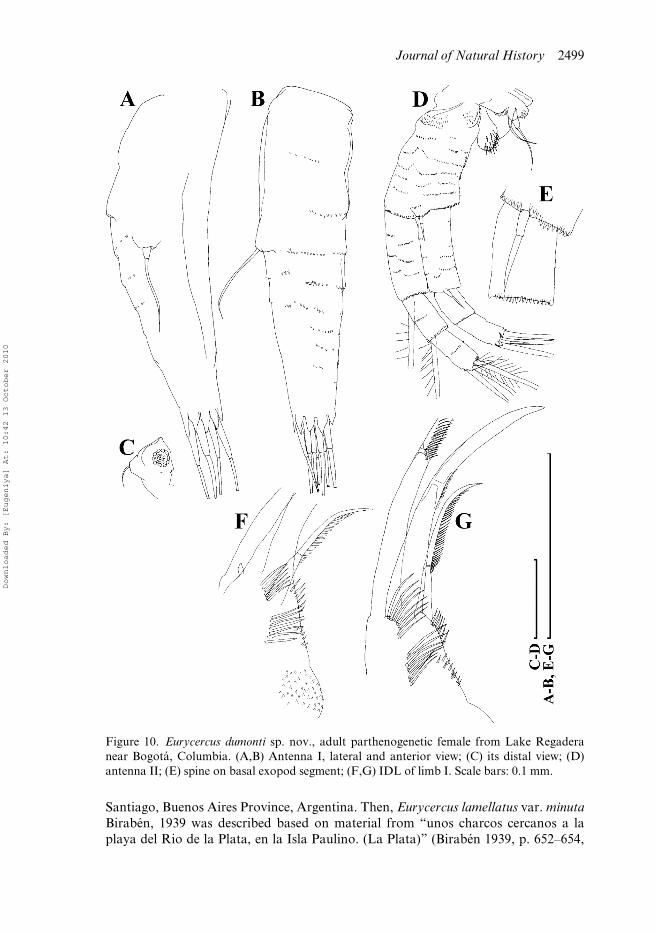
Dorsal margin with a dorsal bubble bearing head pores. In anterior view, body wide,median dorsal keel absent. Rostrum short; posterior portion of head shield lacking astraight section of the margin immediately anterior to the point of mandibular articu-lation; lateral head pores ovoid. Labrum with a small, rounded median keel. Postab-domen with a shallow distal anal embayment and 80–93 preanal teeth. Teeth at baseof preclaw portion long, thin, predominantly single. Antenna I protruding greatlybeyond labral keel and tip of rostrum; elongated; antennular sensory seta arisingapproximately in its middle; denticles in rows encircling antennular surface speciallysmall, located on clearly visible ridges. IDL of limb I with three bisegmented setae,among them, a remarkable strong hook-like seta, but not as large as in E. macracanthus.IDL supplied with seven or eight long distal spinules, seven to nine long proximalspinules, six to eight long marginal spinules and seven to nine short basal spinules.Limb II scraper 5 with 20–21 denticles, scraper 6 with 20–26 denticles, scraper 7 with20–24 denticles. Nine setae in filter plate III; eight setae in filter plate IV, eight setaein filter plate V. Intestine has a single loop. Length 0.65–1.59 mm.
Short description
Adult parthenogenetic female. In lateral view body subovoid in larger females (BH/BL = 0.61–0.64) (Figure 8A). Dorsal margin interrupted by a dorsal bubble bearinghead pores. In anterior view, body wide, not compressed laterally, median dorsal keel isabsent (Figure 8B). Parthenogenetic females carry few eggs in the brood pouch. Headwith short rostrum (Figure 8C,D). Head shield little wider than long (HW/HL about1.1) with maximum height posterior to level of mandibular articulation, posteriorportion lacking a straight section of the margin immediately anterior to the point ofmandibular articulation (Figure 8E). A single major “head pore” as a ringed, subovalfield of special cuticle; a minute, circular lateral pore located at either side of major pore,closer to it (Figure 8F). Labrum with a small, rounded median keel (Figure 8C–D,G–I).Armature of posterior valve margin as in previous species (Figure 8J–L). Postabdo-men broad (PH/PL about 0.50–0.55), with subparallel dorsal and ventral margins(Figure 9A). Distal anal embayment shallow (Figure 9C,D). Armature of the preanalmargin as a series of preanal teeth (NT = 80–93); small gap lacking any teeth at baseof postabdominal setae (Figure 9B); teeth in middle of preanal margin with sharptips. Teeth at base of preclaw portion long, thin, predominantly single, while longerdistalmost teeth doubled or tripled (Figure 9E,F). Postabdominal claw robust (CL/PLabout 0.3); first (distalmost) basal spine long (DS/CL about 0.3), second (basalmost)spine short (BS/CL about 0.16; BS/DS about 0.4–0.5). Antenna I short relative tobody length (AL/BL about 0.12), but protruding greatly beyond labral keel and tipof rostrum; elongated (AL/DA = 3.1–3.5) (Figure 10A,B), triangular in section(Figure 10C). Antennular sensory seta arising approximately in its middle. Denticlesin rows encircling antennular surface specially small, located on clearly visible ridges.On antenna II, spine situated on basal segment of exopod somewhat shorter thanlength of second segment (Figure 10D,E). In general, limbs are very similar to those inthe previous species. Limb I with IDL supplied with three bisegmented setae generallydecreasing in size towards endites, among them, a remarkable strong hook-likeseta, but not as large as in E. macracanthus. IDL supplied with four groups ofspinules, namely: seven or eight long distal spinules, seven to nine long proximal
Downloaded By: [Eugeniya] At: 10:42 13 October 2010
Journal of Natural History 2497
spinules, six to eight long marginal spinules and seven to nine short basal spinules(Figure 10F,G). Limb II scraper 5 with 20–21 denticles, scraper 6 with 20–26 denti-cles, scraper 7 with 20–24 denticles. Other limbs as in previous species. Intestine hasa single loop. Length 0.65–1.59 mm.
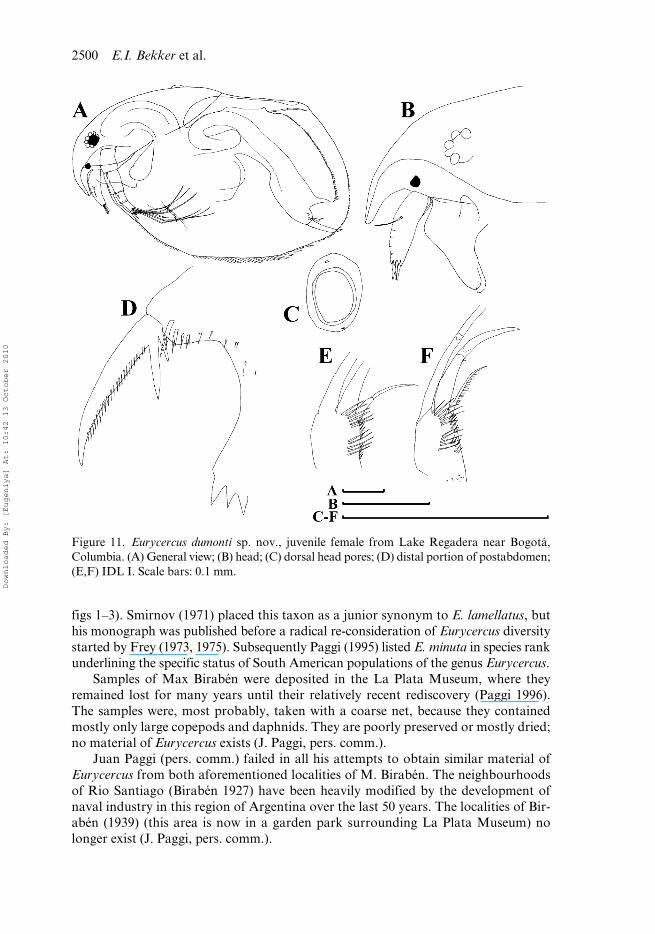
Comments on juvenile female. In contrast to adult female, body more rectangular,somewhat lower (BH/BL = 0.57–0.62), dorsal margin almost straight (Figure 11A).Labrum as adult (Figure 11B), but rostrum a little shorter. Lateral head poreslocated close to major pore (Figure 11C). Postabdomen with significantly fewerpreanal teeth (NT = 72–81), fewer postanal teeth and more robust claws with rela-tively large first (distalmost) basal spine (Figure 11D). Antenna I shorter than inadult (Figure 11B). IDL with four to six long distal spinules, five or six long proximalspinules, five or six long marginal spinules and five long basal spinules (Figure 11E,F).Limb II scraper 5 with 20–23 denticles, scraper 6 with 18–24 denticles, scraper 7 with14–20 denticles.
Figure 8. Eurycercus dumonti sp. nov., adult parthenogenetic female from Lake Regadera nearBogotá, Columbia. (A,B) Lateral and anterior view; (C,D) head, lateral view; (E) head shield;(F) head pores; (G) labrum, ventral view; (H,I) its posterior view; (J) armature of posteroventralangle; (K) ventral portion of posterior margin; (L) dorsal portion of posterior margin. Scalebars: 1 mm (A,B); 0.1 mm (C–L).
Downloaded By: [Eugeniya] At: 10:42 13 October 2010
2498 E.I. Bekker et al.
Differential diagnosis
See Table 1 and Discussion for differentiation of American species of Eurycercus(Bullatifrons).
Distribution
The species is known from the Andean highlands (Lake Regadera), Columbia andthe lowlands in southern (subtropical) Brazil (Estado do Mato Grosso do Sul). It isprobably widely distributed on the South American continent.
Discussion
Notes on “E. minuta Birabén, 1939”
This species was first recorded as nomen nudum by Birabén (1927, p. 573) from “canalal entrada y lugares adyacentes al Puerto La Plata”, neighbourhoods of Río
Figure 9. Eurycercus dumonti sp. nov., adult parthenogenetic female from Lake Regadera nearBogotá, Columbia. (A) Postabdomen, general view; (B) Basal portion of its preanal margin; (C-D)Its distal portion; (E-F) Postanal teeth closest to postabdomenal claw. Scale bars: 0.1 mm.
Downloaded By: [Eugeniya] At: 10:42 13 October 2010
Journal of Natural History 2499
Santiago, Buenos Aires Province, Argentina. Then, Eurycercus lamellatus var. minutaBirabén, 1939 was described based on material from “unos charcos cercanos a laplaya del Rio de la Plata, en la Isla Paulino. (La Plata)” (Birabén 1939, p. 652–654,
Figure 10. Eurycercus dumonti sp. nov., adult parthenogenetic female from Lake Regaderanear Bogotá, Columbia. (A,B) Antenna I, lateral and anterior view; (C) its distal view; (D)antenna II; (E) spine on basal exopod segment; (F,G) IDL of limb I. Scale bars: 0.1 mm.
Downloaded By: [Eugeniya] At: 10:42 13 October 2010
2500 E.I. Bekker et al.
figs 1–3). Smirnov (1971) placed this taxon as a junior synonym to E. lamellatus, buthis monograph was published before a radical re-consideration of Eurycercus diversitystarted by Frey (1973, 1975). Subsequently Paggi (1995) listed E. minuta in species rankunderlining the specific status of South American populations of the genus Eurycercus.
Samples of Max Birabén were deposited in the La Plata Museum, where theyremained lost for many years until their relatively recent rediscovery (Paggi 1996).The samples were, most probably, taken with a coarse net, because they containedmostly only large copepods and daphnids. They are poorly preserved or mostly dried;no material of Eurycercus exists (J. Paggi, pers. comm.).
Juan Paggi (pers. comm.) failed in all his attempts to obtain similar material ofEurycercus from both aforementioned localities of M. Birabén. The neighbourhoodsof Rio Santiago (Birabén 1927) have been heavily modified by the development ofnaval industry in this region of Argentina over the last 50 years. The localities of Bir-abén (1939) (this area is now in a garden park surrounding La Plata Museum) nolonger exist (J. Paggi, pers. comm.).
Figure 11. Eurycercus dumonti sp. nov., juvenile female from Lake Regadera near Bogotá,Columbia. (A) General view; (B) head; (C) dorsal head pores; (D) distal portion of postabdomen;(E,F) IDL I. Scale bars: 0.1 mm.
Downloaded By: [Eugeniya] At: 10:42 13 October 2010

Journal of Natural History 2501
As a consequence we have no chance to find any type or topotype material. Butthe description and pictures of Birabén (1939) are not detailed enough to determineeven a subgenus of his species. The small size (0.58–0.65 mm) of his specimens couldbe a reflection of their juvenile age (note that the female from the author’s fig. 1 doesnot contain eggs). So we have no chance to associate E. minutus with any taxa foundby us (especially keeping in mind that two of them are found in southern Brazil, quiteclose to La Plata). So Birabén’s taxon is species incorrigenda with limited chance thatit can be re-evaluated.
Comments on E. (E.) cf. lamellatus in Brazil
Our single specimen was found in the material of Brandorff et al. (1982), but theauthors did not record this species; probably this single specimen was missed. Eurycercus(Eurycercus) lamellatus s. str. is a very common Palaearctic taxon (Smirnov 1971;Alonso 1996). During most of the twentieth century, only two species were recog-nized in the world fauna: E. lamellatus and E. glacialis. As a result, many specimensfrom non-Palaearctic localities were determined under the former name (Frey 1971).Now it is obvious that they belong to other species, described subsequently, such asE. (E.) microdontus Frey, 1978 from North America, or awaiting description, such asSouth African E. (E.) sp. nov. (Frey 1993). Many populations earlier determined as“E. lamellatus”, especially from South America, in reality belong to the subgenusE. (Bullatifrons) (Frey 1975).
Here we found a single specimen which belongs to E. (E.) lamellatus or its closestcongener. Our specimen could not belong to “North American” E. (E.) microdontusbecause of its moderately compressed body and relatively low dorsal keel. There is areal chance that we have here E. (E.) lamellatus s. str., which was introduced to theAmazon basin from the Palaearctic by human activities. The final conclusion can onlybe drawn after obtaining new, better material on this taxon from South America.
Diagnostic traits of Eurycercus (Bullatifrons) species and analogies with other anomopod groups
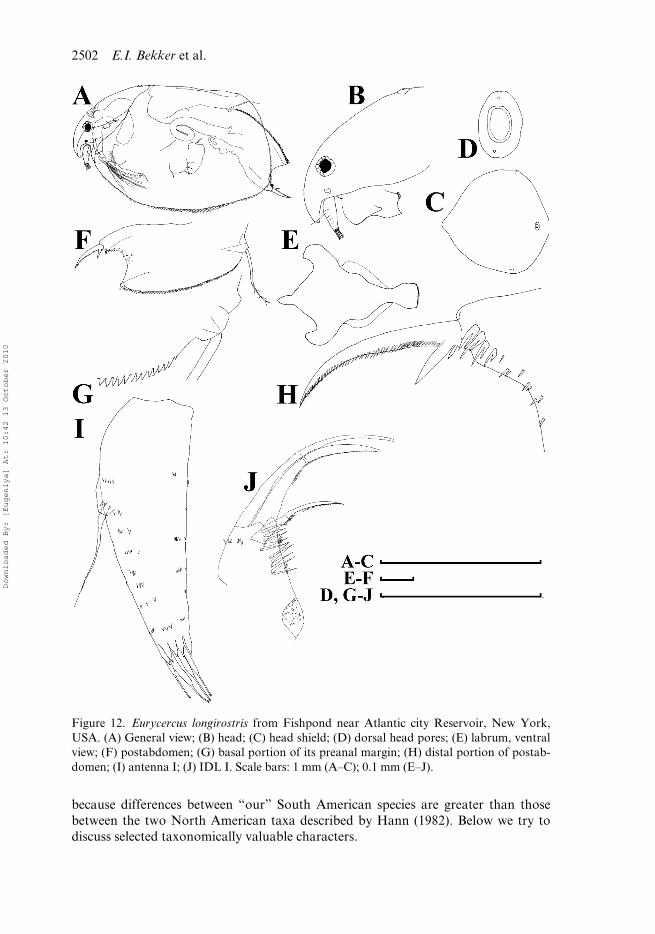
Until now, four species of the subgenus Bullatifrons were known: Eurycercus macra-canthus Frey, 1973 from the Russian Far East and China (Frey 1973; Chen Shou-Zhonget al. 1995; Minakawa et al. 2006), E. pompholygodes Frey, 1975 from Swedenand Norway (Frey 1975; Hessen and Walseng 2008), E. vernalis Hann, 1982 andE. longirostris Hann, 1982 from North America (Hann 1982; Elías-Gutiérrez et al.1997). The latter two species are indistinguishable morphologically except for somedifferences in the ontogenetic changes of some morphometric characters. Here were-studied several populations of longirostris and vernalis (Figure 12A) for their accu-rate comparison with South American taxa. We could confirm that there are no mor-phological differences between the adult females of the former and the latter,furthermore, preliminary phylogenetic data based on the CO1 gene sequences suggestthat they are conspecific (Kotov, unpublished results). The revealed differencesbetween American taxa of E. (Bullatifrons) are reflected in Table 1.
In contrast to the system of species determination based on comparison ofontogenetic changes of some morphometric characters (Hann 1982), we used hereanother approach, namely differentiation based on largest adult females. It was possible
Downloaded By: [Eugeniya] At: 10:42 13 October 2010
2502 E.I. Bekker et al.
because differences between “our” South American species are greater than thosebetween the two North American taxa described by Hann (1982). Below we try todiscuss selected taxonomically valuable characters.
Figure 12. Eurycercus longirostris from Fishpond near Atlantic city Reservoir, New York,USA. (A) General view; (B) head; (C) head shield; (D) dorsal head pores; (E) labrum, ventralview; (F) postabdomen; (G) basal portion of its preanal margin; (H) distal portion of postab-domen; (I) antenna I; (J) IDL I. Scale bars: 1 mm (A–C); 0.1 mm (E–J).
Downloaded By: [Eugeniya] At: 10:42 13 October 2010
Journal of Natural History 2503
Both South American taxa are relatively small, and E. dumonti sp. nov. is apparentlyamong the smallest eurycercids in the world (Table 1). The body size is traditionallyregarded as an important diagnostic character of the anomopod species (Smirnov 1971;Kotov and Elías-Gutiérrez 2009). However, unfortunately, having limited material of fewspecimens from a population, it is very difficult to use this character because we can con-clude that the range of size is underestimated, but have no idea of the maximum size.
The relative length of the labral keel is a ‘traditional’ diagnostic character forEurycercus (Frey 1973). It works for the closest relatives of our two new species, namelyfor differentiation of a Palaearctic pair of species, E. (B.) macracanthus andE. (B.) pompholygodes (Frey 1975). In most eurycercids the keel is large (approximatelyas long as antenna I, sometimes even longer), and only in some species of the subgenusEurycercus (Bullatifrons) (E. pompholygodes and E. dumonti sp. nov.), is it small.
Some anomopod genera have well-developed teeth or hillocks on the preanalmargin. The function of these structures is not always obvious, but they are charac-teristic just for benthic animals (Kotov 2006). Investigators of such groups try to usetheir number and external appearance in systematics. Characters of teeth or hillocksare found to be very important diagnostic features in Ilyocryptus (Kotov and Elías-Gutiérrez 2009) and Leydigia (Kotov 2009), but in these groups the number of thepostanal structures is many times less as compared with Eurycercus. Very importantfor taxonomy of the latter group is the conclusion of Hann (1990) about the differ-ent shape of postanal teeth in E. (Teretifrons) glacialis Lilljeborg, 1887 andE. (T.) nigracanthus Hann, 1990. But we think that counting the number of preanalteeth in Eurycercus (following Frey 1971 and subsequent papers) is significantly lesshelpful for taxonomy compared with the aforementioned groups because of their
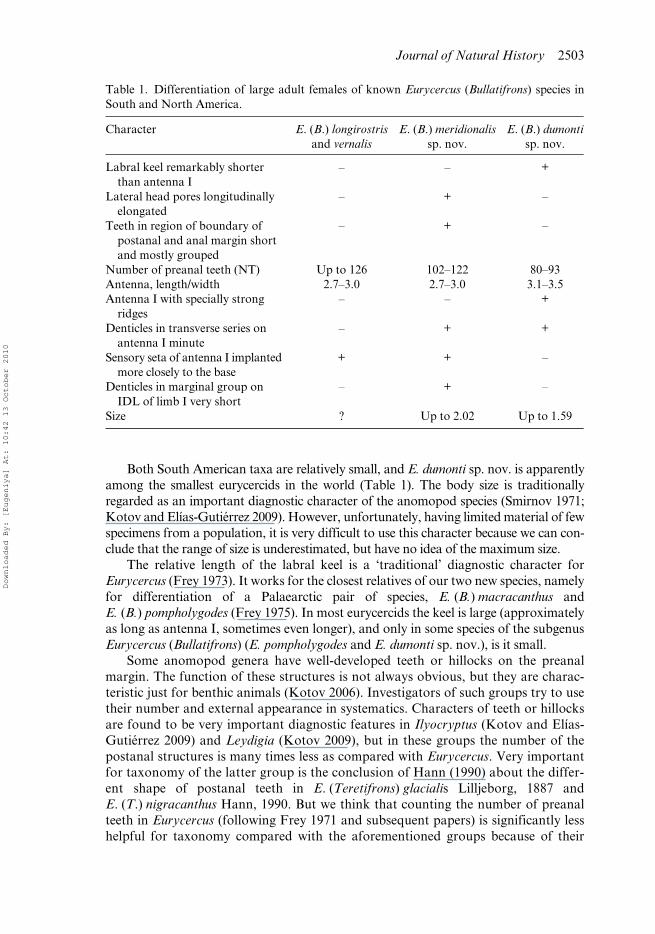
Table 1. Differentiation of large adult females of known Eurycercus (Bullatifrons) species inSouth and North America.
Character E. (B.) longirostrisand vernalis
E. (B.) meridionalissp. nov.
E. (B.) dumontisp. nov.
Labral keel remarkably shorter than antenna I
– – +
Lateral head pores longitudinally elongated
– + –
Teeth in region of boundary of postanal and anal margin short and mostly grouped
– + –
Number of preanal teeth (NT) Up to 126 102–122 80–93Antenna, length/width 2.7–3.0 2.7–3.0 3.1–3.5Antenna I with specially strong
ridges– – +
Denticles in transverse series on antenna I minute
– + +
Sensory seta of antenna I implanted more closely to the base
+ + –
Denticles in marginal group on IDL of limb I very short
– + –
Size ? Up to 2.02 Up to 1.59
Downloaded By: [Eugeniya] At: 10:42 13 October 2010

2504 E.I. Bekker et al.
large number. The number of any elements in any nonoligomerized rows is normallyvery variable among specimens (Smirmov 1971). We believe that a search fortaxonomically helpful characters in more oligomerized structures is better for Euryc-ercus taxonomy than the ‘traditional’ counting of numerous postabdominal teeththat vary greatly in number.
Peculiarities of teeth at the anal–postanal boundary are found by us to be valua-ble for discrimination of the American eurycercid species. Other characters of thepostabdomen do not work. Discrimination of close species based on mostly double/mostly single postanal denticles was introduced to the taxonomy of Chydorinae byFrey (1988) and subsequently confirmed to be useful (Smirnov et al. 2006). It worksfor the discrimination of these two South American taxa.
It is well-known that the length of antenna I is an important diagnostic characterfor subgeneric and even species differentiation in Eurycercus (Frey 1971, 1975; Hann1990). In the case of American Eurycercus (Bullatifrons) this character also works. InE. longirostris, E. vernalis and E. meridionalis sp. nov., length of antenna I is aboutthe length of the labral keel, while in E. dumonti sp. nov., the labral keel reaches onlyabout 0.5–0.7 of antenna I length. The question is whether antenna I is elongated, oris the labral keel shortened? Such questions could be resolved only by a phylogenyreconstruction, which for Eurycercus is a task for the future. At this stage, E. dumontisp. nov. seems to be a taxon retaining more plesiomorphies (a relatively short labralkeel and a relatively long antenna I).
In the past, the fine characters of antenna I attracted little attention of investigators(see papers by Frey 1971, 1973, 1975); this appendage must be re-examined in allpreviously described species of the subgenus Eurycercus (Bullatifrons). It is a quiteremarkable fact that taxonomically valuable characters for the eurycercids (i.e. rela-tive length of antenna I, expression of ridges and denticles on its body) are the sameas we found previously for the ilyocryptids (Kotov and Elías-Gutiérrez 2009),another family containing a single genus and regarded as “primitive”, having a basalposition on the Anomopoda phylogenetic tree.
Groups of setules on IDL are more highly valuable for Eurycercus taxonomybecause some of them are reduced in some species (Hann 1982, 1990). In the case ofE. meridionalis sp. nov., the marginal group is not fully absent, but contains shortdenticles in contrast to other species with long denticles. So, importance of IDLspinulation is confirmed by our study.
In reality, some groups of spinules on IDL are characteristic not only of eurycerc-ids, but of at least some Chydoridae, both alonines (Alonso 1996; Kotov 2000a, 2009;Sinev and Kotov 2000; Van Damme et al. 2005) and chydorines (Alonso 1996; VanDamme and Dumont 2007; Kotov 2008), and at least some Macrothricidae (Alonso1996; Silva-Briano and Dumont 2001; Kotov 2007). Spinulation of IDL is not norm-ally used for species discrimination in the aforementioned anomopod groups becausethe small size of the animals makes the IDL armature indistinguishable under anoptical microscope, even using maximum magnification. But the value of the setuleson IDL for species and subspecies discrimination is confirmed for a large-sizedchydorid Leydigia (Kotov 2009), phylogenetically distant from the eurycercids. Wethink that spinulation of IDL is a good source of additional characters for thesystematics of relatively large chydorids and macrothricids.
In reality, all characters found to be valuable for the eurycercid taxonomy in thepresent paper, were already used previously for species discrimination in other
Downloaded By: [Eugeniya] At: 10:42 13 October 2010

Journal of Natural History 2505
anomopod groups – chydorids and ilyocryptids. Our study underlined again theurgency for a taxonomist in a particular group of the Anomopoda to look at thesystematics of other groups with the aim of finding analogies in taxonomically valuablecharacters, reflecting convergences in evolutionary history.
General comments on Eurycercus in South America
Few previous authors reported the occurrence of E. lamellatus in South America(Montú and Gloeden 1986; Serafim-Junior et al. 2003). We found that ‘Eurycercuslamellatus’ illustrated by Montú and Gloeden (1986) from Lagoa dos Patos in RioGrande do Sul (note that that ‘our’ locality with the same name for E. dumonti sp.nov. is from another state, Mato Grosso do Sul), belongs to another subgenus, that isE. (B.) meridionalis sp. nov. It is a remarkable fact that the authors incorrectly deter-mined the species, but correctly illustrated a little protuberance at the head poreregion, suggesting that the specimens belong to the E. (Bullatifrons) subgenus, and alarge labral keel. Any other previous records of E. lamellatus from the Neotropicsshould be considered with caution. Although a single specimen of E. (E.) cf. lamellatuswas really found by us in a single locality, most specimens belonged toE. (B.) meridionalis sp. nov. and E. (B.) dumonti sp. nov., which are apparently morecommon in South America.
Our revision has changed ideas on the worldwide distribution of Eurycercus.Now it is clear that the subgenus E. (Bullatifrons), most diverse within the genus, isdistributed in the Neotropics, and that the Neotropics are as rich in Eurycercus spe-cies as the Nearctic zone. We demonstrate here (1) the necessity of an accurate studyof the cladoceran specimens for their adequate determination and (2) the necessity ofinformative descriptions and illustrations in faunistic publications. Unfortunately,previous authors published for some regions (including the Neotropics) lists of taxa,many of which could not be associated with any real species. This information couldnot be used for any kind of subsequent analysis, which in such cases ends up dealingwith artificial groups instead of real taxa (see discussion in Kotov 2008; Kotov et al.2010). Only re-examination of the cladoceran species from different regions couldprovide adequate information for recent attempts to analyse the biogeography, quan-titative biodiversity and other aspects of the cladoceran distribution.
Acknowledgements
We are very grateful to Professor N.N. Smirnov for help during different phases of our workand supplying us with some samples, Dr J.C. Paggi for valuable consultations, Professor H.J.Dumont, Dr F.A. Lansac-Tôha and M.Sci. Gilmar Perbiche Neves for the samples with Euryc-ercus from South America. This work is supported by the Russian Foundation for BasicResearch (grant 09-04-00201-a for E.I.B. and A.A.K.), the ‘Biodiversity’ Programme of thePresidium of Russian Academy of Sciences (for E.I.B. and A.A.K.) and Russian Science Sup-port Foundation (for A.A.K.), and the Conselho Nacional de Desenvolvimento Científico eTecnológico (grant 555288/2006-4 for L.M.A.E.L.).
References
Abilhoa V, Agostinho AA. 2007. Aspectos biológicos de duas espécies de Astyanax (Teleostei,Characidae) em lagoas marginais do alto Rio Iguaçu, Paraná, Brasil. Estud Biol. 29(66):23–32.
Downloaded By: [Eugeniya] At: 10:42 13 October 2010

2506 E.I. Bekker et al.
Alonso M. 1996. Crustacea, Branchiopoda. In: Ramos MA, editor. Fauna Iberica. Volume 7.Madrid: Museo Nacional de Ciencias Naturales; 486 p.
Birabén M. 1927. Observaciones sobre la microfauna de Rio Santiago. Physis 8:572–574.Birabén M. 1939. Los Cladóceros de la familia Chydoridae. Physis 17:651–671.Brandorff G.-O, Koste W, Smirnov NN. 1982. Structure of rotiferan and crustacean communu-
ties of the lower Rio Nhamundá, Amazonas, Brazil. Stud Neotr Fauna Envir. 17:69–121.Chen Shou Zhong, Tang Wen Qiao, Zhang Peng. 1995. A new record of Cladocera (Eurycercus
macrocanthus Frey, 1973) in China. Sichuan J Zool. 14:68–69.Dumont HJ, Silva-Briano M. 1998. A reclassification of the anomopod families Macrothricidae
and Chydoridae, with the creation of a new suborder, the Radopoda (Crustacea: Branchi-opoda). Hydrobiologia 384:119–149.
Elías-Gutiérrez ME; Ciros-Pérez J; Gutiérrez-Aguirre M, Cervantes-Martínez A. 1997. Achecklist of the littoral cladocerans from Mexico, with descriptions of five taxa recentlyrecorded from the Neovolcanic Province. Hydrobiologia 360:63–73.
Elías-Gutiérrez M, Kotov AA, Garfias-Espejo T. 2006. Cladocera (Crustacea: Ctenopoda,Anomopoda) from southern Mexico, Belize and northern Guatemala, with some biogeo-graphical notes. Zootaxa. 1119:1–27.
Elías-Gutiérrrez M, Suárez-Morales E, Gutiérrez-Aguirre M, Silva-Briano M, Granados-Ramírez JG, Garfias-Espejo T. 2008. Cladocera y Copepoda de las aguas continentales deMéxico. Guía ilustrada. México, D.F.: UNAM, CONABIO, ECOSUR, SEMARNAT-CONACYT; 322 pp.
Forró L, Korovchinsky NM, Kotov AA, Petrusek A. 2008. Global diversity of cladocerans(Cladocera; Crustacea) in freshwater. Hydrobiologia 595:177–184.
Frey DG. 1971. Worldwide distribution and ecology of Eurycercus and Saycia (Cladocera).Limnol Oceanogr. 16:254–308.
Frey DG. 1973. Comparative morphology and biology of three species of Eurycercus(Chydoridae, Cladocera) with description of Eurycercus macrocanthus sp. n. Int Rev gesHydrobiol. 58:221–267.
Frey DG. 1975. Subgeneric differentiation within Eurycercus (Cladocera, Chydoridae) and anew species from Northern Sweden. Hydrobiologia 46:263–300.
Frey DG. 1978. A new species of Eurycercus (Cladocera, Chydoridae) from Southern UnitedStates. Tulane Stud Zool Bot. 20:1–25.
Frey DG. 1988. Separation of Pleuroxus laevis Sars, 1861 from two species in North America:Pleuroxus straminius Birge, 1879 and Pleuroxus chiangi n. sp. (Cladocera, Chydoridae).Can J Zool. 66:2534–2563.
Frey DG. 1993. The penetration of cladocerans into saline waters. Hydrobiologia 267:233–248.Fryer G. 1963. The functional morphology and the feeding mechanism of the chydorid
cladoceran Eurycercus lamellatus (O. F. Müller). Trans R Soc Edinb. 65:335–381.Hann BJ. 1982. Two new species of Eurycercus (Bullatifrons) from Eastern North America
(Chydoridae, Cladocera). Taxonomy, ontogeny, and biology. Int Rev ges Hydrobiol.67:585–610.
Hann BJ. 1985. Influence of temperature on life history characteristics of two sibling species ofEurycercus (Cladocera, Chydoridae). Can J Zool. 63:891–898.
Hann BJ. 1990. Redescription of Eurycercus (Teretifrons) glacialis (Cladocera, Chydoridae),and description of a new species, E. (T.) nigracanthus, from Newfoundland, Canada. CanJ Zool. 68:2146–2157.
Hessen DO, Walseng B. 2008. The rarity concept and the commonness of rarity in freshwaterzooplankton. Freshwat Biol. 53:2026–2035.
Kotov AA. 1995. The development of head and thoracic appendages of Eurycercus lamellatus(Crustacea, Daphniiformes) in later embryogenesis. Zool Zh. 74:39–50 (in Russian).
Kotov AA. 1996. Fate of the second maxilla during embryogenesis in some AnomopodaCrustacea (Branchiopoda). Zool J Linn Soc. 116:393–405.
Downloaded By: [Eugeniya] At: 10:42 13 October 2010

Journal of Natural History 2507
Kotov AA. 1998. Molar surface structure and work of mandibles of Eurycercus lamellatus(Anomopoda, Crustacea). Zool Zh. 77:548–557 (in Russian).
Kotov AA. 2000a. Analysis of Kozhowia Vasiljeva & Smirnov, 1969 (Chydoridae, Anomop-oda, Branchiopoda), and a description of Parakozhowia n. gen. Hydrobiologia 437:17–56.
Kotov AA. 2000b. Morphology and variability of Eurycercus lamellatus (O.F. Müller, 1776)(Branchiopoda: Anomopoda: Eurycercidae) from Lake Glubokoe, Moskow Area, centralRussia. Arthropoda Selecta. 9(3):159–173.
Kotov AA. 2006. Adaptations of the Anomopoda (Cladocera) for benthic mode of life. Zool.Zh. 85:1043–1059.
Kotov AA. 2007. Revision of the hirsuticornis-like species of Macrothrix Baird, 1843(Cladocera: Anomopoda: Macrothricidae) from Subantarctic and temperate regions ofthe southern hemisphere. J Nat Hist. 41:2569–2620.
Kotov AA. 2008. Finding of Pleuroxus smirnovi sp. nov. from the Pamir region revealedtoday’s imperfect state of systematics and biogeography of the Chydorinae (Cladocera:Chydoridae). Internat Rev Hydrobiol. 93:200–209.
Kotov AA. 2009. A revision of Leydigia Kurz, 1875 (Anomopoda, Cladocera, Branchiopoda),and subgeneric differentiation within the genus. Zootaxa 2082:1–68.
Kotov AA, Elías-Gutiérrez M. 2009. A phylogenetic analysis of Ilyocryptus Sars, 1862(Cladocera: Ilyocryptidae). Internat Rev Hydrobiol. 94:208–225.
Kotov AA, Elías-Gutiérrez M, Williams JL. 2002. A preliminary revision of sordidus-like spe-cies of Ilyocryptus Sars, 1862 (Anomopoda, Branchiopoda) in North America, withdescription of I. bernerae n. sp. Hydrobiologia 472:141–176.
Kotov AA, Elmoor-Loureiro LMA. 2008. Revision of Ilyocryprus Sars, 1862 (Cladocera:Ilyocryptidae) of Brazil with description of two new subspecies. Zootaxa 1962:49–64.
Kotov AA, Sinev AY, Berrios VL. 2010. The Cladocera (Crustacea: Branchiopoda) of six highaltitude water bodies in the North Chilean Andes, with discussion of Andean endemism.Zootaxa 2430:1–66.
Minakawa N, Tanaka S, Bogatov V. 2006. Occurrence of Eurycercus (Teretifrons) glacialisLilljeborg, 1887 (Cladocera, Chydoridae) on Sakhalin Island. Biodiversity and Biogeographyof the Kuril Islands and Sakhalin. 2:111–114.
Montú M, Gloeden IM. 1986. Atlas dos Cladocera and Copepoda (Crustacea) do estuário daLagoa dos Patos (Rio Grande, Brasil). Neritica 1(2):1–134.
Paggi JC. 1992. A new species of Ilyocryptus (Anomopoda: Macrothricidae) from Argentina.Hydrobiologia 231:141–151.
Paggi JC. 1995. Crustacea, Cladocera. In Lopretto EC, Tell G. editors. Ecossistema de Aguas Con-tinentales: Metodologías para su estudio. La Plata: Ediciones Sur. ; Tomo III; p. 909–951.
Paggi JC. 1996. Daphnia (Ctenodaphnia) menucoensis (Anomapoda; Daphniidae): a new spe-cies from athalassic saline waters in Argentina. Hydrobiologia 319:137–147.
Paggi JC. 1999. Status and phylogenetic relationships of Daphnia sarsi Daday, 1902 (Crustacea:Anomopoda). Hydrobiologia 403:27–37.
Paz AR. 2003. Análise dos principais fatores intervenientes no comportamento hidrodinâmicodo banhado do Taim (RS) [Master thesis]. [Porto Alegre (Rio Grande do Sul)]: Universi-dade Federal do Rio Grande do Sul.
Rodrigues LC, Train S, Roberto MC, Pagioro TA. 2002. Seasonal fluctuation of some limno-logical variables on a floodplain lake (Patos lagoon) of the Upper Paraná River, MatoGrosso do Sul State, Brazil. Braz. Arch Biol Technol. 45:499–513.
Serafim-Junior M, Lansac-Tôha FA, Paggi JC, Velho LFM, Robertson B. 2003. Cladocerafauna composition in a river–lagoon system of the upper Paraná river floodplain, with anew record for Brazil. Brazil J Biol. 63:349–356.
Silva-Briano M, Dumont HJ. 2001. Wlassicsia, Bunops and Onchobunops (Anomopoda), threerelated genera. Hydrobiologia 442:1–28.
Sinev AY, Elmoor-Loureiro LMA. 2010. Three new species of chydorid cladocerans of sub-family Aloninae (Branchipoda: Anomopoda: Chydoridae) from Brazil. Zootaxa 2390:1–25.
Downloaded By: [Eugeniya] At: 10:42 13 October 2010

2508 E.I. Bekker et al.
Sinev AY, Kotov AA. 2000. Redescription of Alona labrosa Vasiljeva & Smirnov, 1969(Chydoridae, Anomopoda, Branchiopoda). Hydrobiologia 439:125–139.
Smirnov NN. 1971. Chydoridae of the world fauna. Fauna SSSR. Rakoobraznie. 1(2):1–531(in Russian).
Smirnov NN, Kotov AA, Coronel J. 2006. Partial revision of the aduncus-like species of Pleu-roxus Baird, 1843 (Chydoridae, Cladocera) from the southern hemisphere with commentsof subgeneric differentiation within the genus. J Nat Hist. 40:1617–1639.
Souza MC, Kawakita K, Slusarski SR, Pereira GF. 2009. Vascular flora of the Upper ParanáRiver floodplain. Braz J Biol. 69:735–745.
Van Damme K, Dumont HJ. 2007. Limb morphology of the carnivorous anomopods Anchis-tropus emarginatus Sars, 1862 and Pseudochydorus globosus (Baird, 1843) (Crustacea:Branchiopoda: Anomopoda). Ann Limnol. 43:271–284.
Van Damme K, Kotov AA, Dumont HJ. 2005. Redescription of Leydigia parva Daday, 1905and assignment to Parvalona gen. nov. (Cladocera: Anomopoda: Chydoridae). J NatHist. 39:2125–2136.
Waechter JL, Jarenkow JA. 1998. Composição e estrutura do componente arbóreo das matasturfosas do Taim, Rio Grande do Sul. Biotemas. 11:45–69.
Downloaded By: [Eugeniya] At: 10:42 13 October 2010





























