Embed Size (px)
Citation preview

RESEARCH PAPER
The distribution of TPX2 in dividing leaf cells of the fernAsplenium nidusE. Panteris, I.-D. S. Adamakis & K. ChanoumidouDepartment of Botany, School of Biology, Aristotle University of Thessaloniki, Thessaloniki, Greece
INTRODUCTION
Cell division in higher plants requires the organisation of sev-eral successive microtubule arrays, some of which are plant-specific. The preprophase microtubule band, the perinuclearprophase spindle, the acentrosomal metaphase and anaphasespindle, the phragmoplast, the perinuclear early interphasemicrotubules and the cortical interphase array follow eachother as mitosis and cytokinesis proceed in plant cells (seeamong others Hasezawa & Kumagai 2002). The assembly andorganisation of the above arrays depend on several factors,such as the c-tubulin complexes (Binarova et al. 2006; Pastu-glia et al. 2006; see also Kong et al. 2010), the Ran GTPase(Pay et al. 2002; Jeong et al. 2005; Zhao et al. 2008; Meier &Brkljacic 2009) and the Aurora kinases (Van Damme et al.2004; Demidov et al. 2005; Kawabe et al. 2005). Apart fromthe above, a homologue of the Targeting Protein for Xklp2(TPX2) has recently been identified in Arabidopsis thaliana,Nicotiana tabacum and Trandescantia virginiana, involved inspindle assembly during prophase (Vos et al. 2008). In theabove plants, TPX2 is intranuclear during interphase and isexported from the nucleus during prophase, interacts withAurora kinases and possibly also participates in microtubulenucleation (Vos et al. 2008; see also Petrovska et al. 2012).After nuclear envelope breakdown, TPX2 in A. thaliana andN. tabacum is localised at the kinetochore microtubule bun-dles during metaphase and anaphase and is degraded duringtelophase (see above). Its importance for cell division hasbeen further proven as nuclear envelope breakdown andmitosis were inhibited by microinjection of anti-TPX2 (see
above). Apart from A. thaliana and N. tabacum, the presenceof TPX2 or TPX2-related protein homologues in other plantspecies has also been reported (Vos et al. 2008; Evrard et al.2009).
Microtubule organisation in ferns is characterised by somediscernible peculiarities in comparison to angiosperms. Whilein the latter plants preprophase band narrowing is F-actinmediated (Mineyuki & Palevitz 1990; Eleftheriou & Palevitz1992), in ferns this process seems to be F-actin-independent(Panteris et al. 1992; further unpublished observations). Inangiosperms the metaphase spindle may not emanate directlyfrom the prophase spindle (Zachariadis et al. 2000, 2004;Lloyd & Chan 2006; see also Panteris et al. 2011; Panteris &Adamakis 2012), while in ferns the former seems to be adirect outcome of the latter (Zachariadis et al. 2004). In an-giosperms, the reinstatement of cortical microtubules atinterphase is preceded by an array of radially arranged peri-nuclear microtubules (see among others Wick 1985; Flanderset al. 1990; Hasezawa & Nagata 1991), while in ferns the cor-tical array is usually organised without the mediation of peri-nuclear microtubules (Panteris et al. 1991, 1995).Furthermore, a distinct population of microtubules lines thenew cell wall in post-cytokinetic cells of ferns (Jenni et al.1990; Panteris et al. 1991; Zachariadis et al. 2003) but not ofangiosperms.
Taking into account the above features of microtubuleorganisation in ferns, we studied the possible presence anddistribution of TPX2 in dividing leaf cells of the fern Aspleni-um nidus. A polyclonal anti-AtTPX2 antibody, raised againstthe conserved TPX2 signature motif (Vos et al. 2008), identi-
Keywords
Asplenium nidus; cell division; cytokinesis;
microtubules; mitosis; TPX2.
Correspondence
E. Panteris, Department of Botany, School of
Biology, Aristotle University of Thessaloniki,
Thessaloniki GR-541 24, Macedonia, Greece.
E-mail: [email protected]
Editor
S. Wick
Received: 2 February 2012; Accepted: 19
March 2012
doi:10.1111/j.1438-8677.2012.00615.x
ABSTRACT
Plant cell division requires the dynamic organisation of several microtubule arrays.The mechanisms of regulation of the above arrays are under rigorous research.Among several factors that are involved in plant microtubule dynamics, the Target-ing Protein for Xklp2 (TPX2) has been found to play a role in spindle organisation,in combination with Aurora kinases, in dividing cells of angiosperms. Microtubuleorganisation in dividing cells of ferns exhibits certain peculiarities. Accordingly, thepresence and distribution of a TPX2 homologue might be helpful in understandingthe patterns and regulatory mechanisms of microtubule arrays in this plant group.In this study, a putative TPX2 homologue was identified using Western blotting inthe fern Asplenium nidus. It was found, using immunostaining and CLSM, that it isco-localised with perinuclear preprophase microtubules and the prophase spindle,and follows the microtubule pattern during metaphase ⁄ anaphase and telophase.During cytokinesis, while in angiosperms TPX2 is degraded, in A. nidus the TPX2signal persists, co-localising with the phragmoplast. In early post-cytokinetic cells, aTPX2 signal is present on the nuclear surface facing the daughter cell wall and,thereafter it is co-localised with the fern-specific microtubule aggregation that linesthe new wall, which is possibly involved in cortical microtubule assembly.
Plant Biology ISSN 1435-8603
Plant Biology 15 (2013) 203–209 ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands 203

fied a putative TPX2 homologue, the distribution of whichexhibits important differences from that observed in A. thali-ana and N. tabacum cells. Leptomycin-B treatment (Kudoet al. 1998) was also applied to confirm the intranuclearlocalisation of TPX2 during interphase.
MATERIAL AND METHODS
Plant material and treatments
Asplenium nidus plants were cultivated in the lab. The apicalareas of coiled young leaves were used for all the procedures,as they comprise a large population of dividing cells. All thechemicals and reagents were purchased from Sigma (Tauf-hirchen, Germany), Merck (Darmstadt, Germany) and Appli-chem (Darmstadt, Germany), unless otherwise stated.Leptomycin-B, a potent inhibitor of nuclear export (Kudo etal. 1998; see also Vos et al. 2008), was initially dissolved as a100 lM stock in absolute methanol and kept at )20 �C; work-ing solution for treatment was prepared by diluting to500 nm. The lamina of the coiled leaf apical area was cut intopieces ca. 4 · 4 mm. The pieces were put in small dishes with500 nm leptomycin-B solution and treated for 4 h in the darkat room temperature on a rocking platform (Biorad, Hercules,CA, USA) at 35 rpm. Similar treatment with 0.5% methanolwas applied as a control. Oryzalin (a kind gift of Dr. HartmutQuader, Biozentrum, Hamburg, Germany) was dissolved as a10 mm stock in absolute acetone and stored at 4 �C; workingsolution was 10 lm, and for control 0.1% acetone was applied.The treatment was similar to that with leptomycin-B.
Immunostaining and fluorescence microscopy
Control as well as leptomycin-B- and oryzalin-treated leafmaterial was prepared as described in Panteris & Karali(2008) with some modifications. First, the leaves or leaf pieceswere cut into ca. 2 · 2 mm samples, immediately pre-fixedfor 20 min at room temperature in 1% (w ⁄ v) paraformalde-hyde (PFA) in PEM (50 mm PIPES, 5 mm EGTA, 5 mm
MgSO4, pH 6.8) with addition of 5% (v ⁄ v) DMSO. All thefollowing steps were also performed at room temperature,unless otherwise stated. After pre-fixation, the samples werefixed for 1 h in 4% (w ⁄ v) PFA in PEM with 5% (v ⁄ v) DMSO.After three 10-min washes in PEM, the samples were treatedwith 0.1% (w ⁄ v) NaBH4 in PEM twice for 30 min, then over-night. After washing, the cell walls were partially digested with2% (w ⁄ v) cellulase (Onozuka R10; Yakult Honsha, Tokyo,Japan) and 2% (w ⁄ v) macerozyme R10 (Serva, Heidelberg,Germany) in PEM for 1 h. After removing the enzymes, thesamples were treated with absolute methanol at )20 �C for20 min. Then, extraction and blocking with 5% (v ⁄ v) DMSOand 1% (v ⁄ v) Triton X-100 + 3% (w ⁄ v) BSA was performedfor 1 h. The samples were incubated overnight with monoclo-nal rat anti-a-tubulin (YOL 1 ⁄ 34, AbD Serotec, Kidlingto,UK) and polyclonal rabbit anti-AtTPX2 (Vos et al. 2008),diluted 1:80 and 1:3000 in PEM, respectively. Some sampleswere incubated only with anti-AtTPX2. Following three 30-min rinses with PEM, the samples were incubated overnightwith FITC-anti-rat and TRITC-anti-rabbit 1:80 in PEM. Afterfurther washing, the samples were mounted in a drop of anti-fade solution (glycerol:PEM 2:1 (v ⁄ v) + 0.5% (w ⁄ v) p-phe-nylendiamine). The preparations were examined under a Ni-
kon D-Eclipse C1 confocal laser scanning microscope (CLSM)with the appropriate wavelengths and filters, and images wererecorded according to the manufacturer’s instructions. Insome specimens, the DNA was counterstained with10 lgÆml)1 Hoechst 33258 and they were observed under aNikon Eclipse i80 microscope (Nikon Instruments EuropeBV, Amstelvee, the Netherlands) equipped with epifluores-cence. Digital images were processed with Adobe PhotoshopCS2, with linear settings only.
Western blotting of leaf extracts
For Western immunoblotting of putative TPX2 isoforms,extracts of untreated coiled leaf areas of A. nidus were ho-mogenised at 4 �C in a buffer containing 20 mm b-glycero-phosphate, 20 mm HEPES, pH 7.5, 20 mm NaF, 2 mm
EDTA, 0.2 mm Na3VO4, 10 mm benzamidine, 5 mm DTT,1% (v ⁄ v) Triton X-100 and a commercial protease inhibitorcocktail. Lysates were incubated on ice for 10 min and thencentrifuged at 4 �C for 20 min at 16,000 g. Clarified superna-tants were collected and total protein was measured using theBradford assay.
Total protein extracts were boiled with 0.33 (v ⁄ v) of SDS-polyacrylamide gel electrophoresis sample buffer (10% w ⁄ vSDS, 13% v ⁄ v glycerol, 300 mm Tris–HCl, pH 6.8, 130 mm
dithiothreitol) and 0.2% (w ⁄ v) bromophenol blue. Proteinswere separated by SDS-PAGE on 12% (w ⁄ v) SDS-polyacryl-amide gels and transferred electrophoretically onto nitrocellu-lose membranes (0.45 lm). Membranes were blocked in 5%(w ⁄ v) BSA powder in TBST buffer (20 mm Tris–HCl, pH7.5, 137 mm NaCl, 0.1% (v ⁄ v) Tween 20) overnight at 4 �C,then washed in TBST (three times for 5 min each) and thenincubated for 2 h at room temperature with polyclonal anti-
Fig. 1. Western blot of Asplenium nidus leaf proteins with polyclonal
antibody against the conserved TPX2 signature motif of Arabidopsis thali-
ana. A single band ca. 100 kDa is identified.
TPX2 in Asplenium nidus Panteris, Adamakis & Chanoumidou
204 Plant Biology 15 (2013) 203–209 ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands
AtTPX2 diluted 1:10,000 in TBST buffer containing 5%(w ⁄ v) BSA powder. Following two 5-min washes in TBSTXS(1x TBS, 1% (w ⁄ v) BSA, 1% (w ⁄ v) Triton X and 1% (w ⁄ v)SDS) one wash with a 0.8% (w ⁄ v) NaCl solution and finallyan incubation of 5 min with a 5% (w ⁄ v) non-fat milk solu-tion in TBST, the membranes were incubated for 2 h atroom temperature with horseradish peroxidase-conjugatedsecondary antibodies (1:2500) diluted in TBST buffer con-taining 5% (w ⁄ v) non-fat milk powder. After washing inTBSTXS buffer (three times for 5 min) and one wash with a0.8% (w ⁄ v) NaCl solution, proteins were detected usingenhanced chemiluminescence (Cell Signalling, Beverly, MA,USA).
RESULTS AND DISCUSSION
A putative TPX2 homologue in A. nidus
A single band at ca. 100 kDa was identified by Western blot-ting of the leaf extract (Fig. 1), suggesting that the putative A.nidus TPX2 homologue may be rather smaller than those ofA. thaliana, N. tabacum and T. virginiana, which have a massof 120 kDa (Vos et al. 2008). Also, A. nidus TPX2 seems tobe more stable than those of the above angiosperms, whichby Western blotting were labelled as bands mainly at 65 kDa,probably due to their high lability (see above). Interestingly,
the molecular mass of the putative A. nidus TPX2 homologueis similar to those of Xenopus laevis at ca. 100 kDa (Witt-mann et al. 2000) and human at ca. 98 kDa (Garrett et al.2002) cells.
The distribution of TPX2 signal in dividing leaf cells
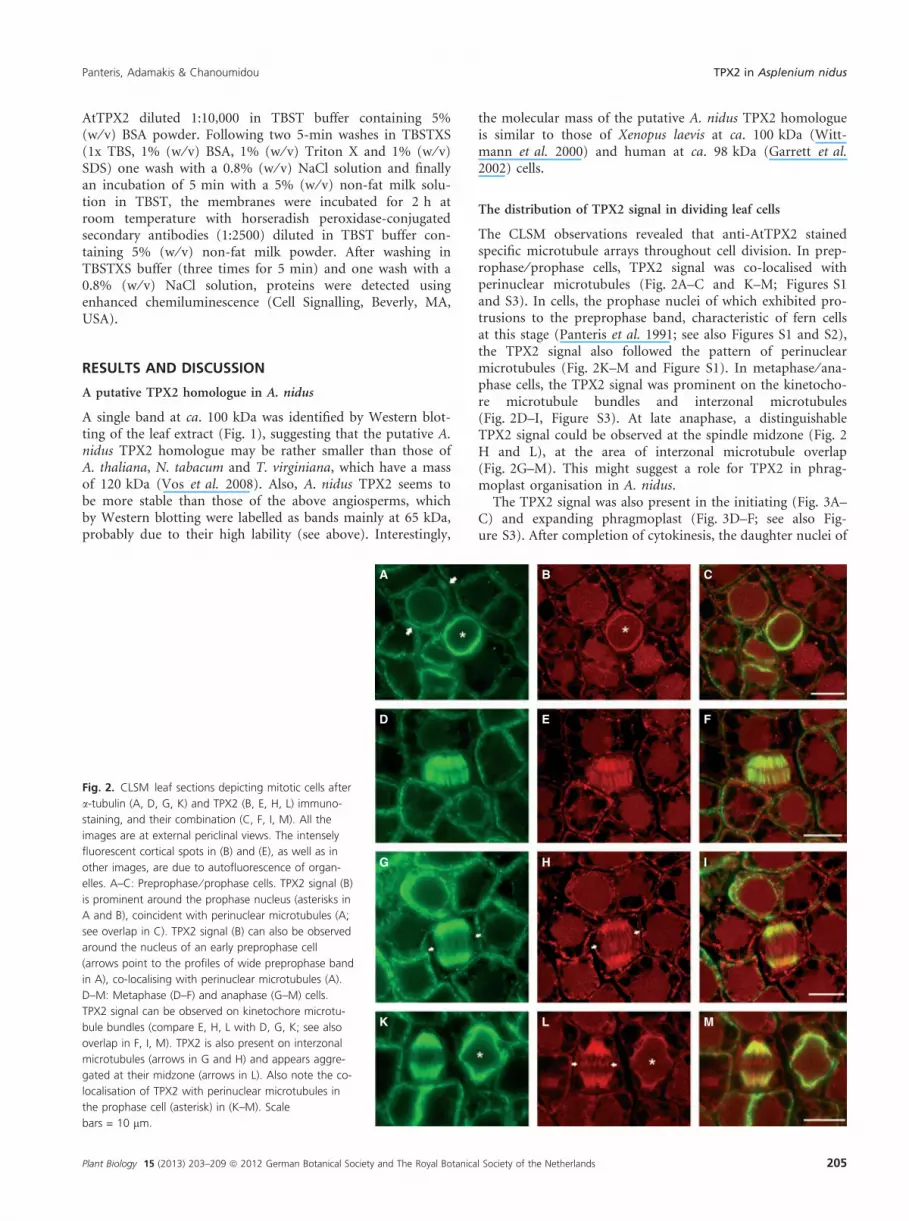
The CLSM observations revealed that anti-AtTPX2 stainedspecific microtubule arrays throughout cell division. In prep-rophase ⁄ prophase cells, TPX2 signal was co-localised withperinuclear microtubules (Fig. 2A–C and K–M; Figures S1and S3). In cells, the prophase nuclei of which exhibited pro-trusions to the preprophase band, characteristic of fern cellsat this stage (Panteris et al. 1991; see also Figures S1 and S2),the TPX2 signal also followed the pattern of perinuclearmicrotubules (Fig. 2K–M and Figure S1). In metaphase ⁄ ana-phase cells, the TPX2 signal was prominent on the kinetocho-re microtubule bundles and interzonal microtubules(Fig. 2D–I, Figure S3). At late anaphase, a distinguishableTPX2 signal could be observed at the spindle midzone (Fig. 2H and L), at the area of interzonal microtubule overlap(Fig. 2G–M). This might suggest a role for TPX2 in phrag-moplast organisation in A. nidus.
The TPX2 signal was also present in the initiating (Fig. 3A–C) and expanding phragmoplast (Fig. 3D–F; see also Fig-ure S3). After completion of cytokinesis, the daughter nuclei of
A B C
D E F
G H I
K L M
Fig. 2. CLSM leaf sections depicting mitotic cells after
a-tubulin (A, D, G, K) and TPX2 (B, E, H, L) immuno-
staining, and their combination (C, F, I, M). All the
images are at external periclinal views. The intensely
fluorescent cortical spots in (B) and (E), as well as in
other images, are due to autofluorescence of organ-
elles. A–C: Preprophase ⁄ prophase cells. TPX2 signal (B)
is prominent around the prophase nucleus (asterisks in
A and B), coincident with perinuclear microtubules (A;
see overlap in C). TPX2 signal (B) can also be observed
around the nucleus of an early preprophase cell
(arrows point to the profiles of wide preprophase band
in A), co-localising with perinuclear microtubules (A).
D–M: Metaphase (D–F) and anaphase (G–M) cells.
TPX2 signal can be observed on kinetochore microtu-
bule bundles (compare E, H, L with D, G, K; see also
overlap in F, I, M). TPX2 is also present on interzonal
microtubules (arrows in G and H) and appears aggre-
gated at their midzone (arrows in L). Also note the co-
localisation of TPX2 with perinuclear microtubules in
the prophase cell (asterisk) in (K–M). Scale
bars = 10 lm.
Panteris, Adamakis & Chanoumidou TPX2 in Asplenium nidus
Plant Biology 15 (2013) 203–209 ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands 205
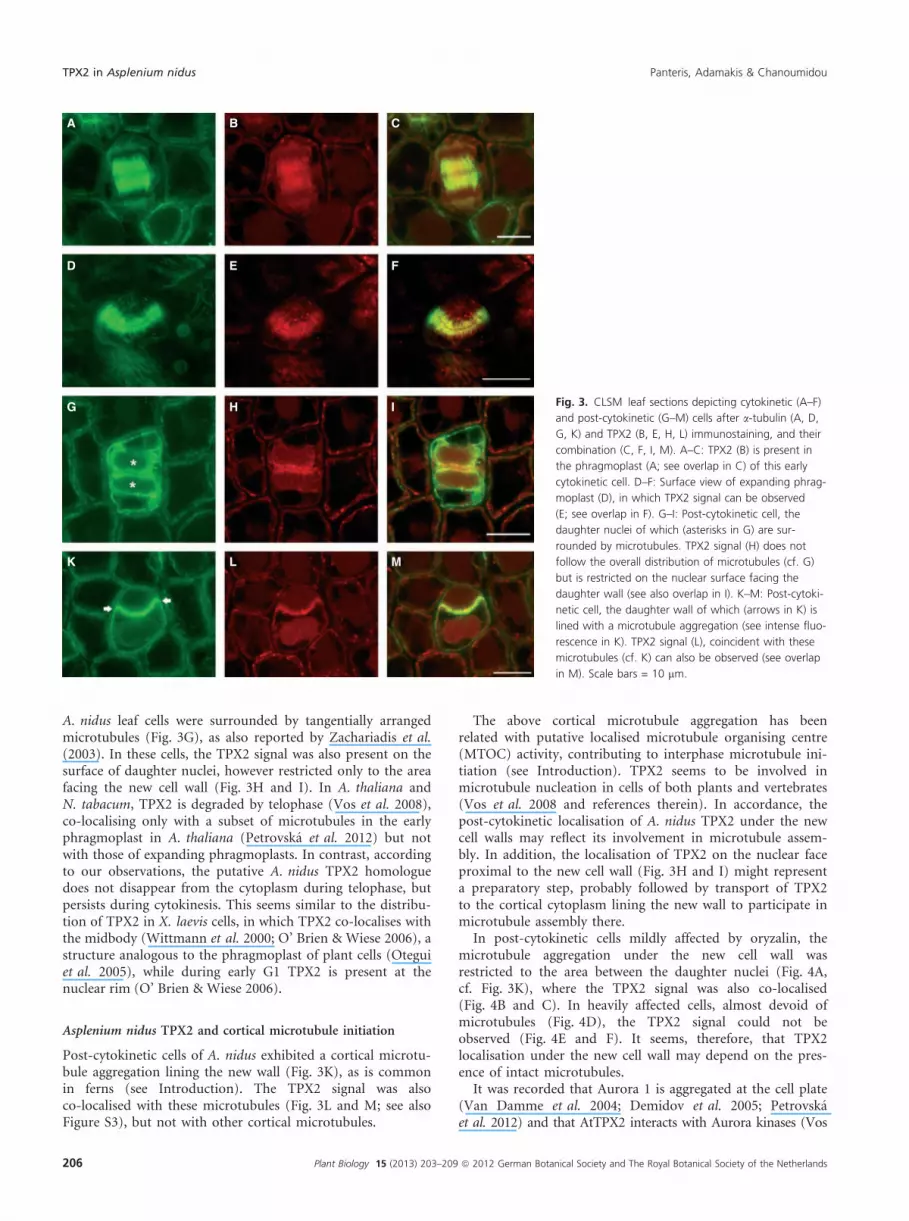
A. nidus leaf cells were surrounded by tangentially arrangedmicrotubules (Fig. 3G), as also reported by Zachariadis et al.(2003). In these cells, the TPX2 signal was also present on thesurface of daughter nuclei, however restricted only to the areafacing the new cell wall (Fig. 3H and I). In A. thaliana andN. tabacum, TPX2 is degraded by telophase (Vos et al. 2008),co-localising only with a subset of microtubules in the earlyphragmoplast in A. thaliana (Petrovska et al. 2012) but notwith those of expanding phragmoplasts. In contrast, accordingto our observations, the putative A. nidus TPX2 homologuedoes not disappear from the cytoplasm during telophase, butpersists during cytokinesis. This seems similar to the distribu-tion of TPX2 in X. laevis cells, in which TPX2 co-localises withthe midbody (Wittmann et al. 2000; O’ Brien & Wiese 2006), astructure analogous to the phragmoplast of plant cells (Oteguiet al. 2005), while during early G1 TPX2 is present at thenuclear rim (O’ Brien & Wiese 2006).
Asplenium nidus TPX2 and cortical microtubule initiation
Post-cytokinetic cells of A. nidus exhibited a cortical microtu-bule aggregation lining the new wall (Fig. 3K), as is commonin ferns (see Introduction). The TPX2 signal was alsoco-localised with these microtubules (Fig. 3L and M; see alsoFigure S3), but not with other cortical microtubules.
The above cortical microtubule aggregation has beenrelated with putative localised microtubule organising centre(MTOC) activity, contributing to interphase microtubule ini-tiation (see Introduction). TPX2 seems to be involved inmicrotubule nucleation in cells of both plants and vertebrates(Vos et al. 2008 and references therein). In accordance, thepost-cytokinetic localisation of A. nidus TPX2 under the newcell walls may reflect its involvement in microtubule assem-bly. In addition, the localisation of TPX2 on the nuclear faceproximal to the new cell wall (Fig. 3H and I) might representa preparatory step, probably followed by transport of TPX2to the cortical cytoplasm lining the new wall to participate inmicrotubule assembly there.
In post-cytokinetic cells mildly affected by oryzalin, themicrotubule aggregation under the new cell wall wasrestricted to the area between the daughter nuclei (Fig. 4A,cf. Fig. 3K), where the TPX2 signal was also co-localised(Fig. 4B and C). In heavily affected cells, almost devoid ofmicrotubules (Fig. 4D), the TPX2 signal could not beobserved (Fig. 4E and F). It seems, therefore, that TPX2localisation under the new cell wall may depend on the pres-ence of intact microtubules.
It was recorded that Aurora 1 is aggregated at the cell plate(Van Damme et al. 2004; Demidov et al. 2005; Petrovskaet al. 2012) and that AtTPX2 interacts with Aurora kinases (Vos
A B C
D E F
G H I
K L M
Fig. 3. CLSM leaf sections depicting cytokinetic (A–F)
and post-cytokinetic (G–M) cells after a-tubulin (A, D,
G, K) and TPX2 (B, E, H, L) immunostaining, and their
combination (C, F, I, M). A–C: TPX2 (B) is present in
the phragmoplast (A; see overlap in C) of this early
cytokinetic cell. D–F: Surface view of expanding phrag-
moplast (D), in which TPX2 signal can be observed
(E; see overlap in F). G–I: Post-cytokinetic cell, the
daughter nuclei of which (asterisks in G) are sur-
rounded by microtubules. TPX2 signal (H) does not
follow the overall distribution of microtubules (cf. G)
but is restricted on the nuclear surface facing the
daughter wall (see also overlap in I). K–M: Post-cytoki-
netic cell, the daughter wall of which (arrows in K) is
lined with a microtubule aggregation (see intense fluo-
rescence in K). TPX2 signal (L), coincident with these
microtubules (cf. K) can also be observed (see overlap
in M). Scale bars = 10 lm.
TPX2 in Asplenium nidus Panteris, Adamakis & Chanoumidou
206 Plant Biology 15 (2013) 203–209 ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands
et al. 2008; Petrovska et al. 2012). Especially, Petrovska et al.(2012) discussed the combined function of At Aurora 1, AtT-PX2 and c-tubulin in regulating microtubule assembly. It istempting, therefore, to assume that the presence of TPX2under the daughter cell wall in A. nidus post-cytokinetic cellsmight reflect such an interaction between Aurora kinases,TPX2 and c-tubulin, which is also localised there (see Panter-is et al. 2000), probably triggering transient MTOC activity.As a result, cortical microtubules are initiated on the daugh-ter cell wall surface of A. nidus, as is also the case in otherferns (see Introduction). In contrast, in A. thaliana andN. tabacum, in which TPX2 is degraded by telophase, suchan interaction does not occur, and interphase microtubulesoriginate from the nuclear surface (see Introduction).
Intranuclear origin of TPX2 in A. nidus cells
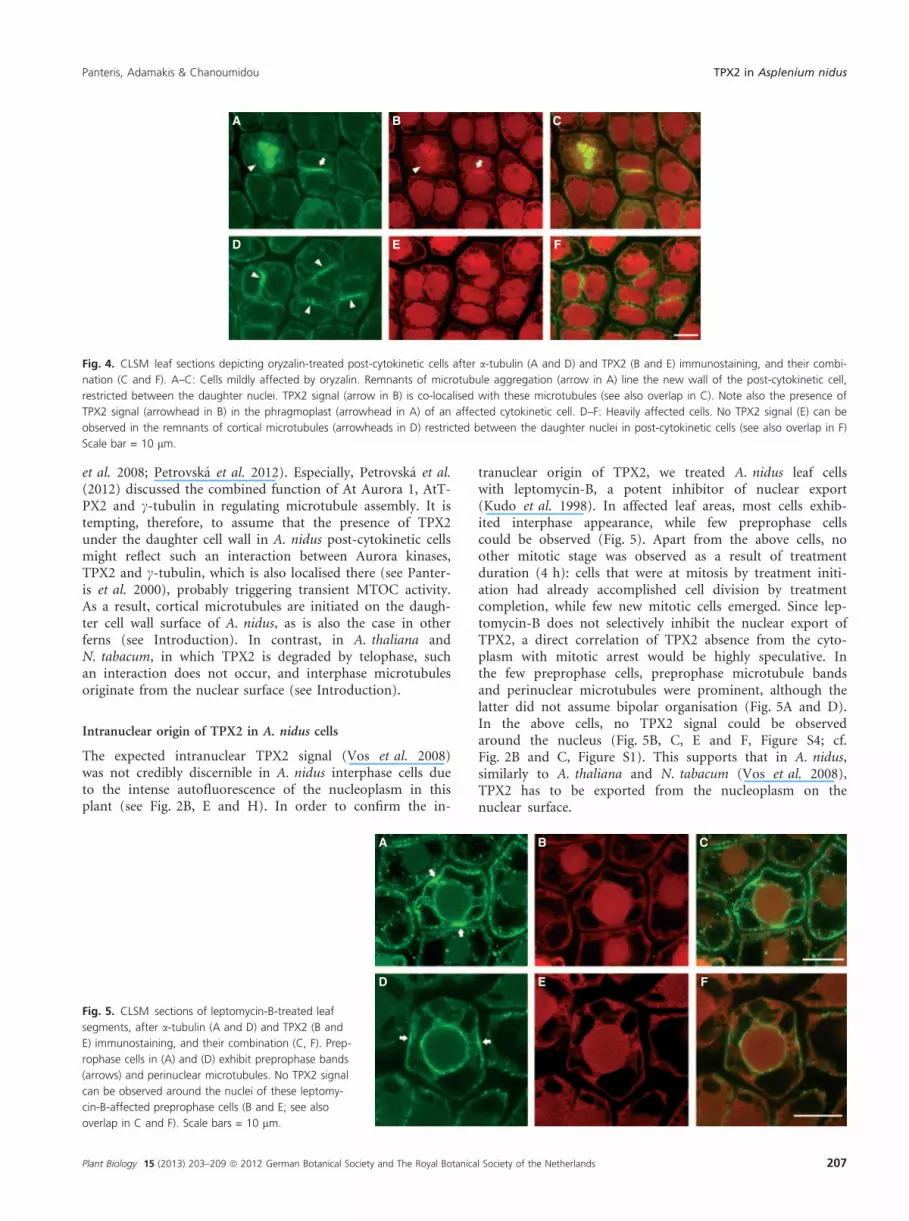
The expected intranuclear TPX2 signal (Vos et al. 2008)was not credibly discernible in A. nidus interphase cells dueto the intense autofluorescence of the nucleoplasm in thisplant (see Fig. 2B, E and H). In order to confirm the in-
tranuclear origin of TPX2, we treated A. nidus leaf cellswith leptomycin-B, a potent inhibitor of nuclear export(Kudo et al. 1998). In affected leaf areas, most cells exhib-ited interphase appearance, while few preprophase cellscould be observed (Fig. 5). Apart from the above cells, noother mitotic stage was observed as a result of treatmentduration (4 h): cells that were at mitosis by treatment initi-ation had already accomplished cell division by treatmentcompletion, while few new mitotic cells emerged. Since lep-tomycin-B does not selectively inhibit the nuclear export ofTPX2, a direct correlation of TPX2 absence from the cyto-plasm with mitotic arrest would be highly speculative. Inthe few preprophase cells, preprophase microtubule bandsand perinuclear microtubules were prominent, although thelatter did not assume bipolar organisation (Fig. 5A and D).In the above cells, no TPX2 signal could be observedaround the nucleus (Fig. 5B, C, E and F, Figure S4; cf.Fig. 2B and C, Figure S1). This supports that in A. nidus,similarly to A. thaliana and N. tabacum (Vos et al. 2008),TPX2 has to be exported from the nucleoplasm on thenuclear surface.
A B C
D E F
Fig. 4. CLSM leaf sections depicting oryzalin-treated post-cytokinetic cells after a-tubulin (A and D) and TPX2 (B and E) immunostaining, and their combi-
nation (C and F). A–C: Cells mildly affected by oryzalin. Remnants of microtubule aggregation (arrow in A) line the new wall of the post-cytokinetic cell,
restricted between the daughter nuclei. TPX2 signal (arrow in B) is co-localised with these microtubules (see also overlap in C). Note also the presence of
TPX2 signal (arrowhead in B) in the phragmoplast (arrowhead in A) of an affected cytokinetic cell. D–F: Heavily affected cells. No TPX2 signal (E) can be
observed in the remnants of cortical microtubules (arrowheads in D) restricted between the daughter nuclei in post-cytokinetic cells (see also overlap in F)
Scale bar = 10 lm.
A B C
D E F
Fig. 5. CLSM sections of leptomycin-B-treated leaf
segments, after a-tubulin (A and D) and TPX2 (B and
E) immunostaining, and their combination (C, F). Prep-
rophase cells in (A) and (D) exhibit preprophase bands
(arrows) and perinuclear microtubules. No TPX2 signal
can be observed around the nuclei of these leptomy-
cin-B-affected preprophase cells (B and E; see also
overlap in C and F). Scale bars = 10 lm.
Panteris, Adamakis & Chanoumidou TPX2 in Asplenium nidus
Plant Biology 15 (2013) 203–209 ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands 207

CONCLUSIONS
We identified a putative TPX2 homologue in the fern A.nidus. Although not yet molecularly characterised, itseems similar in size to TPX2 homologues of vertebrates(ca. 100 kDa; Fig. 1). In addition, similar to its vertebrate ho-mologues, it persists during the telophase ⁄ interphase transi-tion. In particular, its co-localisation with corticalmicrotubules under the daughter cell wall in post-cytokineticcells implies its involvement in localised MTOC activity. Sincea microtubule aggregation under the daughter cell wall hasalso been reported in post-cytokinetic cells of bryophytes (Ap-ostolakos & Galatis 1992), it might be interesting to investi-gate whether TPX2 exhibits a similar localisation, establishinga common feature among land cryptogamic plants.
ACKNOWLEDGEMENTS
We are grateful to Professor Anne-Catherine Schmit (Institutde Biologie Moleculaire des Plants, CNRS, Strasbourg,France) for the anti-AtTPX2 antibody. Dr. Penelope Mavra-gani (School of Biology, Aristotle University) and Dr. Anasta-sia Tsingotjidou (Faculty of Veterinary Medicine, Aristotle
University) kindly provided access to the microscopes usedin this study.
SUPPORTING INFORMATION
Additional Supporting Information may be found in theonline version of this article:
Figure S1. CLSM sections through the nuclei (A–C) andprojections of 20 sections (D–F) of A. nidus leaf protodermafter a-tubulin (A and D) and TPX2 (B and E) immuno-staining, and their combination (C and F).
Figure S2. CLSM sections of prophase cells after a-tubulinimmunostaining.
Figure S3. Fluorescence microscopy images of A. nidusdividing cells after a-tubulin (A and D) and TPX2 (B and E)immunostaining, and DNA counterstaining with Hoechst33,258 (C and F).
Figure S4. Projection of 20 CLSM sections of the celldepicted in Fig. 5D–F.
Please note: Wiley-Blackwell is not responsible for the con-tent or functionality of any supporting materials supplied bythe authors. Any queries (other than missing material) shouldbe directed to the corresponding author for the article.
REFERENCES
Apostolakos P., Galatis B. (1992) Patterns of microtu-
bule organization in two polyhedral cell types in
the gametophyte of the liverwort Marchantia palea-
cea Bert. New Phytologist, 122, 165–178.
Binarova P., Cenklova V., Prochazkova J.,
Doskocilova A., Volc J., Vrlık M., Bogre L. (2006)
c-Tubulin is essential for acentrosomal microtubule
nucleation and coordination of late mitotic events
in Arabidopsis. The Plant Cell, 18, 1199–1212.
Demidov D., Van Damme D., Geelen D., Blattner
F.R., Houben A. (2005) Identification and dynam-
ics of two classes of Aurora-like kinases in Arabid-
opsis and other plants. The Plant Cell, 17, 836–848.
Eleftheriou E.P., Palevitz B.A. (1992) The effect of
cytochalasin D on preprophase band organization
in root tip cells of Allium. Journal of Cell Science,
103, 989–998.
Evrard J.-L., Pieuchot L., Vos J.W., Vernos I., Schmit
A.-C. (2009) Plant TPX2 and related proteins. Plant
Signaling and Behavior, 4, 69–72.
Flanders D.J., Rawlings D.J., Shaw P.J., Lloyd C.W.
(1990) Re-establishment of the interphase microtu-
bule array in vacuolated plant cells, studied by con-
focal microscopy and 3-D imaging. Development,
110, 897–904.
Garrett S., Auer K., Compton D.A., Kapoor T.M.
(2002) hTPX2 is required for normal spindle mor-
phology and centrosome integrity during vertebrate
cell division. Current Biology, 12, 2055–2059.
Hasezawa S., Kumagai F. (2002) Dynamic changes
and the role of the cytoskeleton during the cell
cycle in higher plant cells. International Review of
Cytology, 214, 161–191.
Hasezawa S., Nagata T. (1991) Dynamic organization
of plant microtubules at the three distinct transi-
tion points during the cell cycle progression of syn-
chronized tobacco BY-2 cells. Botanica Acta, 104,
206–211.
Jenni V., Cattelan H., Roos U.-P. (1990) Immunoflu-
orescence and ultrastructure of mitosis and cell
division in the fern Athyrium filix-femina. Botanica
Helvetica, 100, 101–119.
Jeong S.Y., Rose A., Joseph J., Dasso M., Meier I.
(2005) Plant-specific mitotic targeting of RanGAP
requires a functional WPP domain. The Plant Jour-
nal, 42, 270–282.
Kawabe A., Matsunaga S., Nakagawa K., Kurihara D.,
Yoneda A., Hasezawa S., Uchiyama S., Fukui K.
(2005) Characterization of plant Aurora kinases
during mitosis. Plant Molecular Biology, 58, 1–13.
Kong Z., Hotta T., Lee Y.-R.J., Horio T., Liu B. (2010)
The c-tubulin complex protein GCP4 is required for
organizing functional microtubule arrays in Arabid-
opsis thaliana. The Plant Cell, 22, 191–204.
Kudo N., Wolff B., Sekimoto T., Schreiner E., Yoneda
Y., Yanagida M., Horinouchi S., Yoshida M. (1998)
Leptomycin B inhibition of signal-mediated nuclear
export by direct binding to CRM1. Experimental
Cell Research, 242, 540–547.
Lloyd C.W., Chan J. (2006) Not so divided: the com-
mon basis of plant and animal cell division. Nature
Reviews Molecular Cell Biology, 7, 147–152.
Meier I., Brkljacic J. (2009) Adding pieces to the puz-
zling nuclear envelope. Current Opinion in Plant
Biology, 12, 752–759.
Mineyuki Y., Palevitz B.A. (1990) Relationship
between preprophase band organization, F-actin
and the division site in Allium. Fluorescence and
morphometric studies on cytochalasin treated cells.
Journal of Cell Science, 97, 283–295.
O’ Brien L.L., Wiese C. (2006) TPX2 is required for
postmitotic nuclear assembly in cell-free Xenopus
laevis egg extracts. Journal of Cell Biology, 173, 685–
694.
Otegui M.S., Verbrugghe K.J., Skop A.R. (2005) Mid-
bodies and phragmoplasts: analogous structures
involved in cytokinesis. Trends in Cell Biology, 15,
404–413.
Panteris E., Adamakis I.-D.S. (2012) Aberrant micro-
tubule organization in dividing root cells of p60-
katanin mutants. Plant Signaling and Behavior, 7,
16–18.
Panteris E., Karali D.S. (2008) The role of new micro-
tubule assembly and MAP65 in microtubule bundle
formation in pavement cells of Asplenium nidus.
Journal of Biological Research, 10, 139–147.
Panteris E., Galatis B., Apostolakos P. (1991) Patterns
of cortical and perinuclear microtubule organiza-
tion in meristematic root cells of Adiantum capillus
veneris. Protoplasma, 165, 173–188.
Panteris E., Apostolakos P., Galatis B. (1992) The
organization of F-actin in root tip cells of Adian-
tum capillus veneris throughout the cell cycle. A
double label fluorescence microscopy study. Pro-
toplasma, 170, 128–137.
Panteris E., Apostolakos P., Galatis B. (1995) The
effect of taxol on Triticum preprophase root cells:
preprophase microtubule band organization seems
to depend on new microtubule assembly. Protoplas-
ma, 186, 72–78.
Panteris E., Apostolakos P., Graf R., Galatis B. (2000)
Gamma-tubulin colocalizes with microtubule arrays
and tubulin paracrystals in dividing vegetative cells
of higher plants. Protoplasma, 210, 179–187.
Panteris E., Adamakis I.-D.S., Voulgari G., Papado-
poulou G. (2011) A role for katanin in plant cell
division: microtubule organization in dividing root
cells of fra2 and lue1 Arabidopsis thaliana mutants.
Cytoskeleton, 68, 401–413.
Pastuglia M., Azimzadeh J., Goussot M., Camilleri C.,
Belcram K., Evrard J.-L., Schmit A.-C., Guerche P.,
Bouchez D. (2006) c-Tubulin is essential for micro-
tubule organization and development in Arabidop-
sis. The Plant Cell, 18, 1412–1425.
Pay A., Resch K., Frohnmeyer H., Fejes E., Nagy F.,
Nick P. (2002) Plant RanGAPs are localized at the
nuclear envelope in interphase and associated with
microtubules in mitotic cells. The Plant Journal, 30,
699–709.
Petrovska B., Cenklova V., Pochylova Z., Kourova H.,
Doskocilova A., Plıhal O., Binarova L., Binarova P.
(2012) Plant Aurora kinases play a role in mainte-
nance of primary meristems and control of endore-
duplication. New Phytologist, 193, 590–604.
TPX2 in Asplenium nidus Panteris, Adamakis & Chanoumidou
208 Plant Biology 15 (2013) 203–209 ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands

Van Damme D., Bouget F.-Y., Van Poucke K., Inze D.,
Geelen D. (2004) Molecular dissection of plant cyto-
kinesis and phragmoplast structure: a survey of GFP-
tagged proteins. The Plant Journal, 40, 386–398.
Vos J.W., Pieuchot L., Evrard J.-L., Janski N., Bergdoll
M., de Ronde D., Perez L.H., Sardon T., Vernos I.,
Schmit A.-C. (2008) The plant TPX2 protein regu-
lates prospindle assembly before nuclear envelope
breakdown. The Plant Cell, 20, 2783–2797.
Wick S.M. (1985) Immunofluorescence microscopy of
tubulin and microtubule arrays in plant cells. III.
Transition between mitotic ⁄ cytokinetic and inter-
phase microtubule arrays. Cell Biology International
Reports, 9, 357–371.
Wittmann T., Wilm M., Karsenti E., Vernos I. (2000)
TPX2, a novel Xenopus MAP involved in spindle
pole organization. Journal of Cell Biology, 149,
1405–1418.
Zachariadis M., Galatis B., Apostolakos P. (2000)
Study of mitosis in root-tip cells of Triticum turgi-
dum treated with the DNA-intercalating agent ethi-
dium bromide. Protoplasma, 211, 151–164.
Zachariadis M., Quader H., Galatis B., Apostolakos
P. (2003) Organization of the endoplasmic reticu-
lum in dividing cells of the gymnosperms Pinus
brutia and Pinus nigra, and of the pterophyte
Asplenium nidus. Cell Biology International, 27,
31–40.
Zachariadis M., Quader H., Galatis B., Apostolakos P.
(2004) An inhibitor of the ATP-dependent endo-
plasmic reticulum Ca2+-pump affects spindle orga-
nization in dividing cells of the angiosperm
Triticum turgidum but not in species of gymno-
sperms and pteridophytes. Journal of Biological
Research, 2, 3–19.
Zhao Q., Brkljacic J., Meier I. (2008) Two distinct,
interacting classes of nuclear envelope-associated
coiled-coil proteins are required for the tissue-spe-
cific nuclear envelope targeting of Arabidopsis Ran-
GAP. The Plant Cell, 20, 1639–1651.
Panteris, Adamakis & Chanoumidou TPX2 in Asplenium nidus
Plant Biology 15 (2013) 203–209 ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands 209









![Age, size and regeneration of old-growth white pine at Dividing Lake Nature Reserve, Algonquin Park, Ontario, [Ontario For. Res. Rep. 131]](https://img.dokumen.tips/doc/110x75/63603ed98838ef044c06c75e/age-size-and-regeneration-of-old-growth-white-pine-at-dividing-lake-nature-reserve.jpg)



















