Embed Size (px)
Citation preview

SP
PRMa
CSb
n
Acsqttrbm6i(Btsb
ePmvaBbch
cw“l
KB
Ar1
*EAztbkr
Neuroscience 151 (2008) 361–373
0d
YNAPTIC TERMINALS FROM MICE MIDBRAIN EXHIBIT FUNCTIONAL
2X7 RECEPTORCpd(omi1dfaosaats
p(2h(tarP
p(caIwopt
AMLos
rd(Tt
. MARÍN-GARCÍA,a1 J. SÁNCHEZ-NOGUEIRO,a1
. GÓMEZ-VILLAFUERTES,a,b D. LEÓNa AND. T. MIRAS-PORTUGALa*
Departamento de Bioquímica, Facultad de Veterinaria, Universidadomplutense de Madrid, Av. Puerta de Hierro s/n, 28040 Madrid,pain
Centro Nacional de Biotecnología, Consejo Superior de Investigacio-es Científicas, 28049 Madrid, Spain
bstract—P2X7 receptor has been recently localized in miceerebellar granule neuron fibers. Here, the expression of thisubunit has been detected in wild type mice midbrain, byuantitative real time-polymerase chain reaction, immunocy-ochemistry and Western blot assays. The functionality ofhis P2X7 subunit has been confirmed using microfluorimet-ic experiments in isolated synaptic terminals from mice mid-rain. 2=-3=-O-(4-benzoylbenzoyl)-ATP (BzATP) was 30-foldore potent than ATP and EC50 values were 20 �M and
30 �M respectively. Brilliant Blue G (BBG) and 1-[N,O-bis(5-soquinolinesulfonyl)-N-methyl-L-tyrosyl]-4-p h enylpiperazineKN-62) produced an inhibition in the responses induced byzATP, with IC50 values of 0.027 nM and 2.23 nM, respec-
ively. In addition, P2X7 inhibitors as ZnSO4, BBG anduramin abolished partially or totally the responses inducedy the physiological agonist ATP.
According to immunochemical and PCR assays the pres-nce of a “P2X7-like” protein in synaptosomes from validated2X7 knockout (KO) model have been detected. In KO ani-als, BzATP was sixfold more potent than ATP and the EC50
alues were 87 �M and 590 �M respectively. BBG and KN-62lso produced an inhibition in the responses induced byzATP, with IC50 value of 0.61 nM and 118 nM respectively,oth of them higher than in wild type mice. Moreover, thealcium mobilization ability of native P2X7 receptors wasigher in control compared with KO mice.
These biochemical and pharmacological experiments areonsistent with the presence of a functional P2X7 receptor inild type mice midbrain, and the existence of a less efficientP2X7-like” receptor in the KO model. © 2008 IBRO. Pub-ished by Elsevier Ltd. All rights reserved.
ey words: synaptosomes, P2X7 receptor, ATP, BzATP,BG, KN-62.
TP is an important signaling molecule, acting as a neu-otransmitter in both the peripheral nervous system and
Both authors have contributed equally.Corresponding author. Tel: �34-91-3943894; fax: �34-91-3943909.-mail address: [email protected] (M. T. Miras-Portugal).bbreviations: BBG, Brilliant Blue G; BzATP, 2=-3=-O-(4-benzoylben-oyl)-ATP; EGTA, ethylene glycol-bis(2-amino-ethylether)-N,N,N=,N=-etraacetic acid; HBM, hypertonic buffered medium; KN-62, 1-[N,O-is(5-isoquinolinesulfonyl)-N-methyl-L-tyrosyl]-4-phenylpiperazine; KO,
anockout; MOPS, 3-(N-morpholino)propanesulfonic acid; RT-PCR,eal time-polymerase chain reaction; WT, wild type.
306-4522/08$32.00�0.00 © 2008 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2007.10.038
361
NS. Moreover, there is increasing interest in the thera-eutic potential of purinergic compounds in a wide range ofisease conditions in relation to both P1 and P2 receptorsBurnstock, 2005). P2 receptors, which mediate the actionf nucleotides such as ATP, are divided in two groups:etabotropic P2Y receptors coupled to G proteins and
onotropic P2X receptors (Abbracchio and Burnstock,994; Ralevic and Burnstock, 1998; North, 2002). Sevenifferent P2X subunits (P2X1-7) have been cloned to date
rom various mammalian species. P2X subunits may occurs homo- or hetero-oligomeric assemblies of more thanne subunit, except P2X7 receptor, which does not coas-emble with any other subunit (Torres et al., 1999; Northnd Surprenant, 2002; Roberts et al., 2006). Distributionnd relative abundance of P2X receptors differ accordingo brain areas and neuronal topology (Xiang and Burn-tock, 2005; Franke and Illes, 2006; Roberts et al., 2006).
In the last years, different studies have shown theresence of P2X7 receptor in different regions of CNSSperlágh et al., 2006), such as cerebellum (Hervás et al.,003; Sánchez-Nogueiro et al., 2005; León et al., 2006),ippocampus (Armstrong et al., 2002) and cerebral cortexWirkner et al., 2005). Moreover, recent evidence of func-ional P2X7 receptors expressed in midbrain, cerebellarnd cortical synaptic terminals from rat has also beeneported (Miras-Portugal et al., 2003; Hervás et al., 2005;atti et al., 2006).
In this paper, using immunostaining procedures, sim-le and quantitative real time-polymerase chain reactionRT-PCR) and microfluorometric techniques, we haveharacterized the responses mediated by P2X7 receptorctivation in single synaptic terminals from mice midbrain.n addition, P2X7 knockout (KO) mice (Solle et al., 2001)ere used in order to thoroughly compare the functionalityf P2X7 receptor. In these KO mice, the presence of a newrotein was detected, with similar but not identical proper-ies to the genuine P2X7 receptor.
EXPERIMENTAL PROCEDURES
ll experiments carried out at the Universidad Complutense deadrid followed the guidelines of the International Council foraboratory Animal Science (ICLAS). All assays were designed inrder to minimize the number of mice necessary for statisticaltudies and their suffering.
Animals used were 60–90 days old male C57B1/6J mice,eferred from now as wild type (WT), and male P2X7 KO miceerived from Pfizer (Groton, CT, USA), generated by Solle et al.2001). Briefly, the KO mice has a deletion in the region from1527 to T1605, located at exon 13, resulting in the disruption of
he C terminus of the P2X receptor. These KO mice are viable
7nd fertile (Solle et al., 2001).
ved.

S
TRCcae2
TUea(a3Rtr
(PwS
Rp�ptUpecRsifeqcpmovfTp
I
TlmmsswAdeA(a(oa1adr(fbsleo
W
PamaLP
Tq
G
PPPPP
T
m
m
m
a
ats
P. Marín-García et al. / Neuroscience 151 (2008) 361–373362
imple and quantitative RT-PCR experiments
otal RNA was extracted from WT and KO mice midbrain using Trieagent-RNA, DNA, protein isolation reagent (Molecular Researchenter, Inc. Cincinnati, OH, USA), as previously described (Chom-zynski, 1993). RNA was quantified and RT reactions were mades previously described (Sánchez-Nogueiro et al., 2005; Leónt al., 2006), using a final volume of 50 �l of RT buffer containing�g RNA.
Simple PCR reactions were carried out using 0.625 U Ampliaq Gold DNA polymerase (Applied Biosystems, Foster City, CA,SA), in the presence of 3 mM MgCl2, dNTP mixture (0.2 mMach), specific P2X7 oligonucleotide primers (300–600 nM each),nd 3–5 �l of the RT product. After a 10 min hold at 95 °Crequired for Ampli Taq Gold DNA polymerase activation), themplification was run for 40 cycles (95 °C, 30–45 s; 55–60 °C,0–110 s; 72 °C, 45–120 s). Control reactions, H2O control andT without reverse transcriptase, were done to test possible con-
amination. Specific primers for P2X7 were designed in our labo-atory and are summarized in Table 1.
PCR amplification products were electrophoresed on a 1.5%w/v) agarose gel and stained with SYBR Gold probe (Molecularrobes, Eugene, OR, USA). Relative densities of PCR bandsere obtained by a gel imaging system Gel Logic 200 Imagingystem (Kodak, Rochester, NY, USA).
P2X7 mRNA levels were determined through quantitativeT-PCR. In the PCR reactions a commercial mixture of specificrimers and probes for P2X1, P2X2, P2X3, P2X4, P2X7 and-actin (Table 2) (Assays-on-DemandTM Gene Expressionroducts, TaqMan MGB probes, FAMTM dye-labeled), both of
hem prepared in a TaqMan PCR master Mix, No AmperaseNG were used (all from Applied Biosystems). The commercialrobes were labeled with FAM and TAMRA at the 5= and 3=nds, respectively. �-Actin served as endogenous control. Toheck the light variability in the initial concentration, the totalNA quality and the conversion efficiency of the reverse tran-cription reaction, PCR reactions were carried out with an initial
ncubation at 95 °C during 10 min for polymerase activation,ollowed by 40 cycles (melting 95 °C for 15 s, annealing andxtension 60 °C for 1 min), using an ABI PRISM 7700 Se-uence Detection System (Applied Biosystems). The thresholdycle or cT value occurs when an exponential growth PCRroduct is detected. Data analysis was carried out using 2���CT
ethod (Applied Biosystems) (Livak and Schmittgen, 2001),nce efficiency validation experiments were performed. Thesealidation experiments were made constructing standard plotsor P2X7 and �-actin amplification using the real-time PCR.hese experiments showed similar efficiencies between both
able 1. Four specific primer pairsa
Forward primer
p2x7Disrupted TGCATCACCACCTCCAAGCTCTTCCATp2x7Exon 13 GGTGCCAGTGTGGAAATTGmp2x7 GCACGAATTATGGCACCGTCp2x7Intra AGGATCCGGAAGGAGTT
mp2x7 disrupted, which amplifies a region that includes the absent relmost totally exon 13 sequence (G1438-A1795); mp2x7, which ampli
he region that codifies the intracellular epitope (A1737-A1795). Theequence. Tm, temperature of melting.
airs of primers and very close to 2.�
mmunocytochemical studies
he synaptosomal isolation from mouse midbrain was made fol-owing the procedure described by Dunkley et al. (1986) and
odified by Pintor and Miras-Portugal (1995). We consideredidbrain the brain after cortex and cerebellum removal. Synapto-
omal pellets obtained by means of a Percoll gradient, after re-uspension in 1 ml of PBS, were glued onto coverslips pretreatedith poly-L-lysine and fixed with PFA 4% in PBS (w/v) for 15 min.s primary antibodies (incubated 1 h at 37 °C) we used threeifferent anti-P2X7 antibodies that recognize an intracellularpitope (from Arg576 to Tyr595) of the P2X7 receptor, fromlomone Laboratories (Jerusalem, Israel), BD PharMingen
Franklin Lakes, NJ, USA) and Chemicon (Temecula, CA, USA),nd a primary antibody that recognizes an extracellular epitopefrom Lys136 to Cys152) of the P2X7 receptor, from Alomone Lab-ratories, all of them at 1:200 dilution. We also used as primaryntibodies anti-P2X1 (Chemicon) 1:100, anti-P2X2 (Chemicon):100, anti-P2X3 (Chemicon) 1:200, anti-P2X4 (Alomone) 1:100nd anti-synaptophysin from Sigma (St. Louis, MO, USA) at 1:500ilution. As secondary antibodies, TRITC-conjugated goat anti-abbit IgG (Sigma) and FITC-coupled goat anti-mouse IgGSigma), both at 1:500 dilution, were used. Controls were per-ormed following the same procedure but replacing primary anti-odies by the same volume of PBS-BSA solution. Data are pre-ented as mean�S.E.M. of roughly 2000 synaptosomes from ateast three different synaptosomal preparations. Significant differ-nces were determined by two-tailed Student’s t-test, a probabilityf P�0.05 was considered significant.
estern blot experiments
roteins from mice midbrain synaptic terminals were extracted,s previously described (Sánchez-Nogueiro et al., 2005). Theembranes were incubated overnight at 4 °C with the differentnti-P2X7 primary antibodies described above: from Alomoneaboratories (intracellular epitope) at 1:5000 dilution, from BDharMingen (intracellular epitope) at 1:5000 dilution, from Chemicon
able 2. Exon boundary summary of P2X primers/probes used inuantitative RT-PCR reactions and endogenous control, �-actin
ene Exon boundary
2rx1 1–22rx2 7–82rx3 1–22rx4 1–22rx7 5–6
Reverse primer Tm (°C)
CACCAGCAAGGGATCCTGGTAAAGC 58
TAGGGATACTTGAAGCCACTG 60TAGGGATACTTGAAGCCACTGT 60
TAGGGATACTTGAAGCCACT 55
e KO mice (T1517-G1616); mp2x7 exon 13, for a region that includesegion from amino- to carboxy-terminal (G79-A1795); mp2x7 intra, forrs were designed in our laboratory based on specific mouse P2X7
gion in thfies the rse prime
-Actin 6–6

(tpmoGadw
M
SmbNllpesismno2rs
stomtmi6sPsi9eleeMMf
tosfOCI
Foil
P. Marín-García et al. / Neuroscience 151 (2008) 361–373 363
intracellular epitope) at 1:1000 dilution and from Alomone Labora-ories (extracellular epitope) at 1:1000 dilution. We also used asrimary antibody anti-�-tubulin (Sigma) at 1:20,000 dilution. Next,embranes were incubated 1 h at room temperature with the sec-ndary antibody anti-rabbit IgG peroxidase linked (DakoCytomation,lostrup, Denmark), at 1:1000 in all cases for anti-P2X7 detectionnd anti-mouse IgG peroxidase linked (DakoCytomation), 1:5000ilution, for �-tubulin detection. Incubation with the control peptideas made following the manufacturer instructions.
icrofluorometric assays
ynaptosomal pellets containing 0.5 mg of protein, obtained byeans of a Percoll gradient, were resuspended in 1 ml hypertonicuffered medium (HBM) (composition in mM: NaCl 140, KCl 5,aHCO3 5, NaH2PO4 1.2, glucose 13, Hepes 10, pH 7.4) and
oaded with Fura-2 acetoxymethyl ester (Fura-2 AM) (5 �M) (Mo-ecular Probes; Leiden, The Netherlands) for 45 min at 37 °C. Afterelleting and washing, synaptosomes were deposited onto cov-rslips pretreated with poly-L-lysine for 45 min at 37 °C, to allowetting and sticking to the substrate. This time is also sufficient forntrasynaptosomal hydrolysis of the Fura-2 AM. Next, the cover-lips were washed with fresh HBM containing 1.33 mM CaCl2 andounted in a small superfusion chamber. Firstly synaptic termi-als were stimulated with increasing concentrations of the physi-logic P2X7 receptor agonist ATP or the pharmacological one=-3=-O-(4-benzoylbenzoyl)-ATP (BzATP) for 30 s to make theespective dose–response curves. In addition synaptosomes wereuperfused for 2 min with increasing concentrations of BBG (a
ig. 1. P2X7 mRNA sequence analysis from WT and KO mice midbraligonucleotides mp2x7 disrupted (A1, in green), mp2x7 exon 13 (A2,
mage obtained in qualitative analysis using the former primers. mp2x7 Disruptedadder. H2O, control carried out in absence of template. (C) Table: The amplifi
elective P2X7 receptor antagonist) (Sperlágh et al., 2006) andhen were stimulated for 30 s with 100 �M BzATP in the presencef this antagonist. Next, synaptosomes were washed with HBMedium for 90 s and stimulated again for 30 s with 100 �M BzATP
o make the inhibition curve for this antagonist. Similar experi-ents were developed using another antagonist, 1-[N,O-bis(5-
soquinolinesulfonyl)-N-methyl-L-tyrosyl]-4-phenylpiperazine (KN-2) (Bianco et al., 2006). In other set of experiments synapto-omes were then superfused for 2 min with ZnSO4 100 �M (a2X7 receptor inhibitor) (Virginio et al., 1997) and then weretimulated for 30 s with 600 �M ATP in the presence of the
nhibitor. Next, synaptosomes were washed with HBM medium for0 s and stimulated again for 30 s with 600 �M ATP. Similarxperiments were developed, but replacing ZnSO4 by 100 nM Bril-
iant Blue G (BBG) (a selective P2X7 receptor antagonist) (Sperláght al., 2006) or suramin (P2X7 receptor antagonist) (Donnelly-Rob-rts and Jarvis, 2007). All superfusions were made in the absence ofgCl2 in HBM medium. A pulse of 30 mM KCl (in the presence ofgCl2) was applied at the end of each experiment to confirm the
unctionality and viability of the synaptosomes under study.Synaptosomes were excited alternately at 340 and 380 nm
hrough a Nikon TE-200 microscope 100� lens (S Fluor 0.5–1.3il iris). The incoming light was set at 340 and 380 nm, corre-ponding to the fluorescence peaks of Ca2�-saturated and Ca2�-ree Fura-2 AM solutions. Images were obtained using anRCA-ER C 47 42–80 camera from Hamamatsu (Hamamatsuity, Japan) controlled by MetaFluor 6.2r6 PC software (Universal
maging Corporation, Cambridge, UK). Background and autofluo-
epresentative schemes of the different amplified fragments using the), mp2x7 (A3, in blue) and mp2x7 intra (A4, in red). (B) Agarose gel
in. (A) Rin purple
(B1), mp2x7 exon 13 (B2), mp2x7 (B3) and mp2x7 intra (B4). M, DNAed region and its size in every pair of primers used.

rtnGRsmasEst
S
Stcrre(t
wrKePoercf
wuaf(m(
I
Wp
Fmfn
P. Marín-García et al. / Neuroscience 151 (2008) 361–373364
escence components were subtracted at each wavelength andhe ratio 340/380 was calculated by 32-bit float arithmetic (realumbers). Ratios were calibrated into [Ca2�]i values using therynkiewicz’s equation (Grynkiewicz et al., 1985). The variablesmax, Rmin and � were calculated in vitro from the spectra ofmall Fura-2 AM droplets in Ca2�-saturated solution [compositionM: KCl 100, NaCl 10, MgCl2 1, 3-(N-morpholino)propanesulfoniccid (MOPS) 10, CaCl2 2.5 and Fura-2 AM 10 �M] and Ca2�-freeolution [composition mM: KCl 100, NaCl 10, MgCl2 1, MOPS 10,GTA 5 and Fura-2 AM 10 �M], both determined empirically in ourystem. Data are presented as the mean�S.E.M. correspondingo at least three different synaptosomal preparations.
RESULTS
imple and quantitative RT-PCR
imple PCR experiments were carried out using primershat amplified distinct regions in P2X7 cDNA (Table 1). Asan be observed in Fig. 1B1, the primers [(mp2x7 dis-upted, in green) (Fig. 1A1)] which recognized the dis-upted sequence in P2X7 gene KO amplified a 90 pb bandxclusively in the WT midbrain, as previously reportedSánchez-Nogueiro et al., 2005). In addition, when we usedhe specific primers [(mp2x7 exon 13, in purple) (Fig. 1A2)]
ig. 2. Analysis of P2X1,2,3,4,7 expression by quantitative RT-PCR inidbrain mRNA were used in the RT reactions followed by quantitativ
or P2X1,2,3,4 were found between both groups of animals. (A2) In P2X7 KO micormalized using the values obtained for �-actin mRNA and expressed as the m
hich amplified a part of exon 13 that includes the dis-upted region, a 357 pb fragment was detected in WT vsO samples (Fig. 1B2). Furthermore, another pair of prim-rs [(mp2x7, in blue) (Fig. 1A3)], directed to amplify the2X7 cDNA almost completely, showed a 1716 pb productnly in WT midbrain, as it occurred using the former prim-rs (Fig. 1B3). However, using the primers that codified theegion that includes the epitope recognized by the intra-ellular anti-P2X7 antibodies (in red) (Fig. 1A4), a 58 pbragment was detected in WT both KO midbrain (Fig. 1B4).
The expression of P2X1,2,3,4,7 receptors in midbrainas analyzed by quantitative RT-PCR. In P2X1,2,3,4 sub-nits no differences were found between WT and P2X7 KOnimals (Fig. 2A1). On the contrary, P2X7 receptor mRNA
rom KO mice was increased five times with respect to WTFig. 2A2). This increase can be produced by truncatedRNA originated by P2X7 disruption in the KO model
Solle et al., 2001).
mmunochemical analysis of P2X expression
e developed immunocytochemical analysis of P2X ex-ression in synaptosomes from WT and KO mice, to check
parations from WT and KO midbrain. Equal amounts of WT and KOing specific primers for P2X subunits. (A1) No significant differences
DNA pree PCR us
e a five times increase was observed respect to WT samples. Resultsean�S.E.M. of six different experiments. Unpaired t-test, *** P�0.001.

Fiba
P. Marín-García et al. / Neuroscience 151 (2008) 361–373 365
ig. 3. (A) P2X2 and P2X3 immunostaining in synaptosomes from WT and KO midbrain. The immunostaining with anti-P2X receptors is presentedn red. The immunostaining with anti-synaptophysin receptors is presented in green. The co-localization of both antibodies is shown in yellow. Scalear�10 �m. (B) Average of midbrain synaptic terminals expressing P2X and P2X receptors. Data are means�S.E.M. of three different experiments
2 3nd roughly 2000 synaptic terminals.

icaaPmv
eleadfssPm
bcp(ep
ws
Pm
Owttc5assvhC8mE
pwt
FIAmab 7
2
P. Marín-García et al. / Neuroscience 151 (2008) 361–373366
f, at the protein level, some other P2X subunit could beompensating for the lack of P2X7 receptor in the KOnimals. No P2X1 or P2X4 expression was found in syn-ptic terminals from both animals (not shown). P2X2 and2X3 were present in synaptosomes from WT and KOice (Fig. 3A), and the immunostaining percentage was
ery close when comparing both animal models (Fig. 3B).In a next step, the expression of the P2X7 in protein
xtracted from mice midbrain synaptic terminals was ana-yzed. As can be observed in Fig. 4A we found the pres-nce of two bands in WT and in KO and these bands werebolished when the incubation of P2X7 antibodies waseveloped in the presence of the control peptide. No dif-erent P2X7 receptor expression in proteins from midbrainynaptic terminals was found. The 65 kDa band corre-ponds to the predicted molecular weight of the mouse2X7 receptor, while the 77 kDa one may correspond to aodified state of this subunit.
Immunocytochemical studies were carried out in mid-rain synaptosomes to confirm the presence of P2X7 re-eptor in single synaptic terminals. The presence of therotein was detected in synaptosomes from both animalsFig. 4B). In WT 31.5�3% of the total synaptic terminalsxpress the P2X7 receptor. However, in KO animals, the
A
65 kDa
77 kDa
P2X7 Synaptophysin
WT
KO
B
ig. 4. P2X7 expression in midbrain synaptosomes from WT and Kdentical results were found using the other anti-P2X7 primary antibodb, anti-P2X7 primary antibody; control, control peptide incubation.idbrain. The immunostaining with anti-P2X7 receptor (from Chemicontibodies tested. The immunostaining with anti-synaptophysin is presar�10 �m. (C) Average of midbrain synaptic terminals expressing P2X000 synaptic terminals.
ercentage of synaptosomes expressing P2X7 subunit f
as lower (26.8�2%), but this difference is not statisticallyignificant (Fig. 4C).
harmacological studies of P2X7 receptor in miceidbrain synaptic terminals
nce the presence of the P2X7 messenger and proteinas detected in mice midbrain, the functional properties of
he P2X7 receptor in WT and KO animals were under-aken. Synaptic terminals were challenged with increasingoncentrations of ATP and BzATP between 100 nM andmM in the perfusion medium in absence of magnesium,
nd the dose–response curves obtained have been repre-ented in Fig. 5. The ATP dose–response curve showed aimilar affinity in WT and KO synaptic terminals, EC50
alues were 630 �M and 590 �M respectively, and slightigher maximum effect in WT synaptosomes (Fig. 5A).oncerning BzATP (Fig. 5B), EC50 values were 20 �M and7 �M for WT and KO mice, respectively. Again, theaximum effect was slightly higher in the WT animals. TheC50 values and maximal effect are summarized in Table 3.
Next, inhibition curves for BBG and KN-62 were madere-incubating for 2 min the isolated synaptic terminalsith increasing concentrations of these antagonists be-
ween 0.1 pM to 100 �M (depending on the compound),
KO
Abntrol Control
C
WT KO
0
10
20
30
40
% c
o-lo
calis
atio
n
rge
(A) Immunoblots for anti-P2X7 from Chemicon as primary antibody.g of proteins isolated from midbrain synaptic terminals were loaded.nolocalization of P2X7 receptor in synaptosomes from WT and KOented in red. Similar results were obtained with the others anti-P2X7
reen. The co-localization of both antibodies is shown in yellow. Scaleor. Data are means�S.E.M. of three different experiments and roughly
WT
Ab Co
Me
O mice.ies; 30 �
(B) Immun) is presented in g
recept
ollowed by 100 �M BzATP stimulations in presence or
asKseo0aW
rKs(
r
iaAmatsAt
Fsw5liolK
Tct
A
W
K
pirr
Fbsb(atfasb*
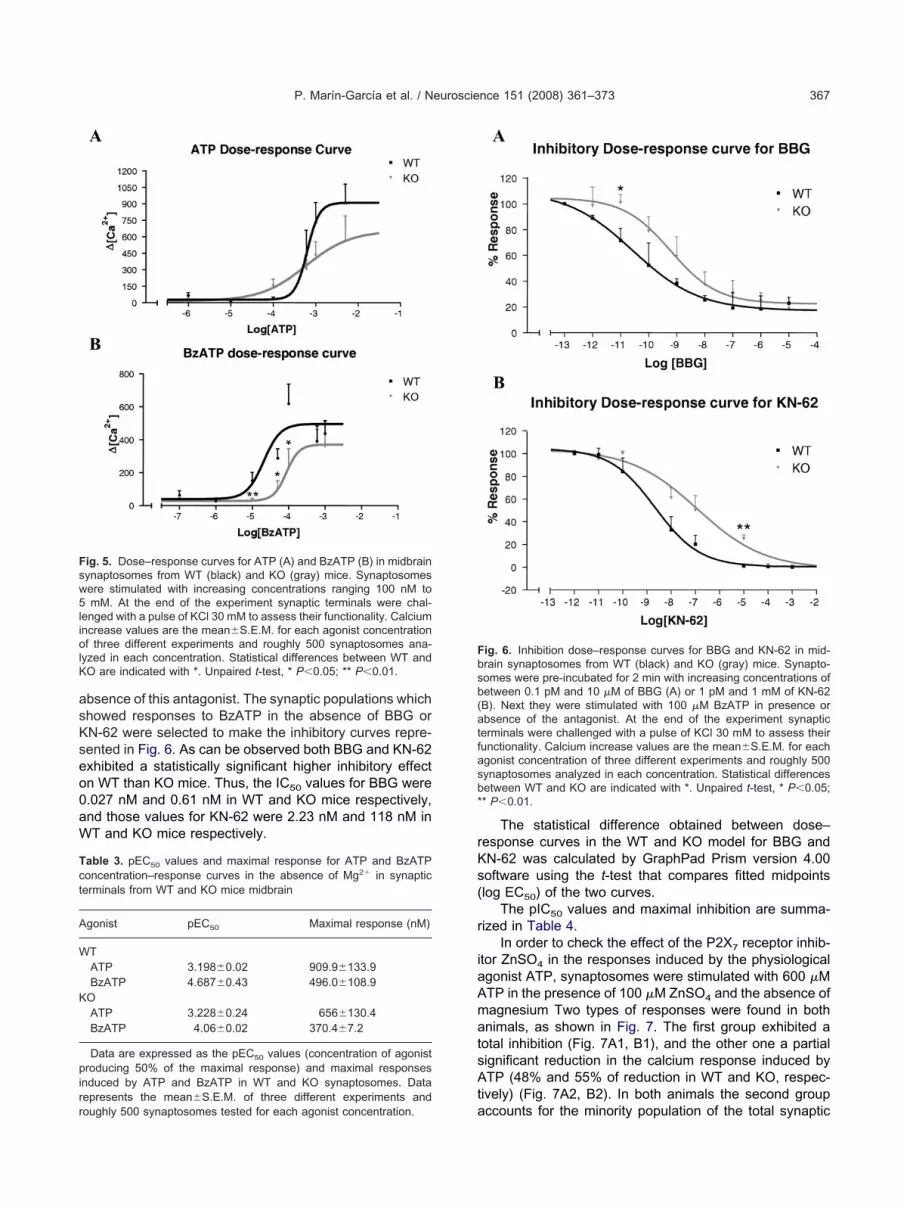
P. Marín-García et al. / Neuroscience 151 (2008) 361–373 367
bsence of this antagonist. The synaptic populations whichhowed responses to BzATP in the absence of BBG orN-62 were selected to make the inhibitory curves repre-ented in Fig. 6. As can be observed both BBG and KN-62xhibited a statistically significant higher inhibitory effectn WT than KO mice. Thus, the IC50 values for BBG were.027 nM and 0.61 nM in WT and KO mice respectively,nd those values for KN-62 were 2.23 nM and 118 nM inT and KO mice respectively.
ig. 5. Dose–response curves for ATP (A) and BzATP (B) in midbrainynaptosomes from WT (black) and KO (gray) mice. Synaptosomesere stimulated with increasing concentrations ranging 100 nM tomM. At the end of the experiment synaptic terminals were chal-
enged with a pulse of KCl 30 mM to assess their functionality. Calciumncrease values are the mean�S.E.M. for each agonist concentrationf three different experiments and roughly 500 synaptosomes ana-
yzed in each concentration. Statistical differences between WT andO are indicated with *. Unpaired t-test, * P�0.05; ** P�0.01.
able 3. pEC50 values and maximal response for ATP and BzATPoncentration–response curves in the absence of Mg2� in synapticerminals from WT and KO mice midbrain
gonist pEC50 Maximal response (nM)
TATP 3.198�0.02 909.9�133.9BzATP 4.687�0.43 496.0�108.9
OATP 3.228�0.24 656�130.4BzATP 4.06�0.02 370.4�7.2
Data are expressed as the pEC50 values (concentration of agonistroducing 50% of the maximal response) and maximal responses
nduced by ATP and BzATP in WT and KO synaptosomes. Dataepresents the mean�S.E.M. of three different experiments and
aoughly 500 synaptosomes tested for each agonist concentration.
The statistical difference obtained between dose–esponse curves in the WT and KO model for BBG andN-62 was calculated by GraphPad Prism version 4.00oftware using the t-test that compares fitted midpointslog EC50) of the two curves.
The pIC50 values and maximal inhibition are summa-ized in Table 4.
In order to check the effect of the P2X7 receptor inhib-tor ZnSO4 in the responses induced by the physiologicalgonist ATP, synaptosomes were stimulated with 600 �MTP in the presence of 100 �M ZnSO4 and the absence ofagnesium Two types of responses were found in bothnimals, as shown in Fig. 7. The first group exhibited a
otal inhibition (Fig. 7A1, B1), and the other one a partialignificant reduction in the calcium response induced byTP (48% and 55% of reduction in WT and KO, respec-
ively) (Fig. 7A2, B2). In both animals the second group
ig. 6. Inhibition dose–response curves for BBG and KN-62 in mid-rain synaptosomes from WT (black) and KO (gray) mice. Synapto-omes were pre-incubated for 2 min with increasing concentrations ofetween 0.1 pM and 10 �M of BBG (A) or 1 pM and 1 mM of KN-62B). Next they were stimulated with 100 �M BzATP in presence orbsence of the antagonist. At the end of the experiment synaptic
erminals were challenged with a pulse of KCl 30 mM to assess theirunctionality. Calcium increase values are the mean�S.E.M. for eachgonist concentration of three different experiments and roughly 500ynaptosomes analyzed in each concentration. Statistical differencesetween WT and KO are indicated with *. Unpaired t-test, * P�0.05;* P�0.01.
ccounts for the minority population of the total synaptic

tBltSat
rre9ai4tipsct
Iry
Tib
A
W
K
Fcttatr
P. Marín-García et al. / Neuroscience 151 (2008) 361–373368
erminals responding as a typical P2X7 receptor (Fig. 7A3,3). In all the cases, the synaptic terminals were stimu-
ated with a second pulse of 600 �M ATP, to verify thathese synaptosomes expressed functional P2X7 subunits.imilar results were obtained when the P2X7 receptorntagonists 100 nM BBG or 100 �M suramin were usedogether with ATP (Figs. 8 and 9). Again, two different
able 4. pIC50 values and maximal inhibition of BBG and KN-62nhibition–response curves in the absence of Mg2� for responses inducedy BzATP in synaptic terminals from WT and KO mice midbrain
ntagonist pIC50 Maximal inhibition (%)
TBBG 10.56�0.21 82.82�3.73KN-62 8.65�0.12 97.81�2.19
OBBG 9.22�0.19 77.56�3.64KN-62 6.93�0.48 98.05�1.95
ig. 7. Responses to ATP 600 �M in the presence of ZnSO4 100 �M ian be observed in WT (A1, A2) and in KO synaptosomes (B1, B2). Aerminals responding to the physiologic agonist. Synaptosomes werehe functionality of the synaptosomes. Each response represents thenalyzed. Unpaired t-test, * P�0.05. Percentage of the two response
o those synaptosomes that do not respond to 600 �M ATP in the presence oeduced response to the physiological agonist in the presence of the inhibitor.
esponses can be observed. On the one hand, some ofesponses to ATP were completely abolished in the pres-nce of the antagonist BBG (Fig. 8A1, B1) or suramin (Fig.A1, B1). On the other hand, partial inhibited responses werelso found for both antagonists. Thus, the percentage of
nhibition in ATP responses induced by BBG was 57% and5% in WT and KO, respectively (Fig. 8A2, B2), whereas in
he presence of suramin this percentage was 45% and 25%n WT and KO, respectively (Fig. 9A2, B2). This secondopulation was the minority of the typical P2X7 respondingynaptosomes (Figs. 8A3, B3, 9A3, 9B3). In all cases thealcium increase induced by ATP was bigger in WT synapticerminals than in KO (Figs. 7, 8, 9).
DISCUSSION
n the present work the presence of a functional P2X7
eceptor in mice midbrain has been reported. In the lastears several studies have indicated the presence of this
n synaptic terminals from WT (A) and KO (B). Two types of responsesulse of ATP 600 �M was applied to confirm the presence of synapticd with a pulse of KCl 30 mM at the end of the experiment to assess.E.M. of three different experiments and roughly 3000 synaptosomesWT (A3) and KO (B3). The shaded area of the pie chart corresponds
n midbraisecond pstimulatemean�Stypes for
f 100 �M ZnSO4. The dark area corresponds to those that showed a

s2W2sp(P(f(Famcfi2M
dP(
2wrsca(ipIcupnmoftmos
Fc npaireda
P. Marín-García et al. / Neuroscience 151 (2008) 361–373 369
ubunit in CNS (Armstrong et al., 2002; Hervás et al.,003; Franke et al., 2005; Sánchez-Nogueiro et al., 2005;irkner et al., 2005; León et al., 2006; Sperlágh et al.,
006). In this sense and using specific primers for P2X7
ubunit designed in our laboratory, we have confirmed theresence of the mRNA for this receptor in mice midbrainFigs. 1 and 2). We have also detected the presence of the2X7 protein in mice midbrain single synaptic terminals
Fig. 4). The presence of P2X7 subunit in synaptosomesrom different CNS regions has been previously describedLundy et al., 2002; Miras-Portugal et al., 2003; Deng andyffe, 2004; Hervás et al., 2005; Patti et al., 2006), as wells in neuromuscular junction in a wide variety of skeletaluscles (Moores et al., 2005). The fact that this receptor
o-localizes with synaptophysin in synaptic terminals con-rms the presynaptic role of P2X7 subunit (Deuchars et al.,001; Miras-Portugal et al., 2003; Hervás et al., 2005;oores et al., 2005).
The available anti-P2X7 antibodies have been used toemonstrate, as one additional proof, the presence of2X receptors at the CNS, as reported by several authors
ig. 8. Responses to ATP 600 �M in the presence of BBG 100 nM inan be observed in WT (A1, A2) and in KO synaptosomes (B1, B2). Und dark areas as in Fig. 7.
7
Deuchars et al., 2001; Sperlágh et al., 2002; Franke et al., r
005; Wirkner et al., 2005; Suadicani et al., 2006). In ourork the immunochemical characterization of the P2X7
eceptor in our animal model is a first step. Regarding thepecificity of the antibodies that recognize anti-P2X7 re-eptor, some authors have suggested that the commercialntibodies are not recognizing neuronal P2X7 proteinAnderson and Nedergaard, 2006). Microfluorometric stud-es developed on single synaptic terminals confirmed theresence of this receptor in mice midbrain synaptosomes.n this way, functional experiments have demonstrated alear calcium increase when synaptic terminals were stim-lated with the physiological P2X7 agonist ATP and theharmacological one BzATP, both in the absence of mag-esium (Fig. 5). Although in midbrain tissue we have foundRNA of all different P2X subunits assayed (Fig. 2A1),nly the presence of P2X2, P2X3 and P2X7 protein wasound in synaptic terminals (Figs. 3A and 4A). However,he responses in the absence of magnesium are mediatedainly by P2X7 subunits. So, the absence of this cation inur experimental conditions assures that most of the re-ponses to ATP or BzATP found are mediated by P2X
synaptic terminals from WT (A) and KO (B). Two types of responsest-test, * P�0.05. Assay conditions as in Fig. 7. In A3 and B3, shaded
midbrain
7
eceptors.

Bf2rt(r0roeacpP1ea
varmtss
wsBdopr
Mt
Fc npaireda
P. Marín-García et al. / Neuroscience 151 (2008) 361–373370
In synaptic terminals from WT midbrain, the fact thatzATP is much more potent than ATP is a distinctive
eature of P2X7 receptors (Anderson and Nedergaard,006). Although BzATP is an effective agonist at other P2Xeceptors, in particular, P2X1, P2X2 and P2X3, it is only athe P2X7 receptor that BzATP is more potent than ATPNorth, 2002). In midbrain mice terminals BBG inhibit theesponses induced by BzATP with an IC50 value of.027 nM. Studies with BBG at the rat and human P2X7
eceptor had demonstrated its selective antagonistic effectver this subunit (Jiang et al., 2000; North, 2002; Hervást al., 2005; Anderson and Nedergaard, 2006; Honore etl., 2006). It is to take into account that at the higher BBGoncentration, most of the responses to BzATP were com-letely abolished by BBG, as it could be expected from2X7 properties reported in the literature (Virginio et al.,997; Deuchars et al., 2001; Ireland et al., 2004; Hervást al., 2005; Moores et al., 2005). In a similar way, KN-62
ig. 9. Responses to ATP 600 �M in the presence of suramin 100 �Man be observed in WT (A1, A2) and in KO synaptosomes (B1, B2). Und dark areas as in Fig. 7.
lso inhibits the responses induced by BzATP, with an IC50 a
alue of 2.23 nM. It is well characterized that KN-62 acts asP2X7 receptor antagonist in human, but has no effect in
at P2X7 subunit (Donnelly-Roberts and Jarvis, 2007). Inouse, this compound is able to behave as a P2X7 recep-
or antagonist in microglial cells (Bianco et al., 2006). Ourtudy shows that KN-62 is also an antagonist of the P2X7
ubunit present in synaptic terminals from mouse midbrain.KO synaptosomes exhibited similar behavior, although
ith several differences. Thus, whereas WT synapto-omes showed a 30-fold increase in EC50 values of thezATP dose–response curve with respect to ATP, thisifference was reduced to sixfold in the KO model. More-ver, the BBG and KN-62 potency was also lower, com-ared with WT, with an IC50 value of 0.61 and 118 nM,espectively.
The inhibitory effect of Zn2� ions in the absence ofg2� ions agrees with the presence of P2X7 receptors at
he synaptic terminals, as has been described by other
in synaptic terminals from WT (A) and KO (B). Two types of responsest-test, * P�0.05. Assay conditions as in Fig. 7. In A3 and B3, shaded
in midbra
uthors (Virginio et al., 1997; Hervás et al., 2005) (Fig. 7).

IAPnATl
tscfwc
aeN2Pdy
ff2ieocvpm12tAPtPsPpra1dre
s6KTtsKe
td
tmpfimw2gLbitPmsMdiiaobip
tifAlaPolsa
TvNKmpps“tec“
AG
P. Marín-García et al. / Neuroscience 151 (2008) 361–373 371
n addition, when synaptosomes were superfused withTP, but in the presence of the P2X7 inhibitor ZnSO4,2X7 selective antagonist BBG or P2X7 receptor antago-ist suramin, the response to the physiological agonistTP was completely or partially inhibited (Figs. 7, 8 and 9).his reveals the heterogeneity in the P2X7 receptor popu-
ation present in mice midbrain.In addition, although both groups of animals responded
o ATP and BzATP, the response amplitude induced in KOynaptosomes was lower than in WT. Moreover, the re-overy of the basal levels of the intracellular calcium wasaster in KO synaptosomes than in WT. A similar behavioras found in cultured granule neurons from P2X7 KO miceerebella (Sánchez-Nogueiro et al., 2005).
This pharmacological characterization is in completegreement with the presence of a P2X7 receptor (Virginiot al., 1997; Lundy et al., 2002; North, 2002; Anderson andedergaard, 2006; Honore et al., 2006; Sperlágh et al.,002, 2006). Recently a novel antagonist of human and rat2X7 receptor, A-740003 (Honore et al., 2006), has beenescribed. This compound is not commercially availableet and its effect in mice has to be checked.
In a previous work, we suggested the presence of aunctional “P2X7-like” receptor in cultured granule cellsrom P2X7 KO mice cerebellum (Sánchez-Nogueiro et al.,005). The efficiency of this KO model has been confirmed
n different cells such as peritoneal macrophages (Sollet al., 2001), leukocytes (Labasi et al., 2002), osteoclast,steoblasts (Ke et al., 2003) and submandibular ductalells (Pochet et al., 2003). In CNS this model seems to bealuable because, in RT-PCR experiments using a pair ofrimers directed to amplify the disrupted region in the KOodel, this sequence was not detected in KO animals (Fig.B1), as previously reported (Sánchez-Nogueiro et al.,005). However, we have not detected a difference be-ween both mice models in the DNA regions that codify forrg576 to Tyr595 (intracellular epitope recognized by anti-2X7 antibodies used in the present work) (Fig. 1B4). Due
o these results we tried to characterize the difference in2X7 mRNA sequence between WT and KO mice throughimple RT-PCR assays, using a specific pair of primers for2X7. First, we employed specific oligonucleotides to am-lify the Trp476 to Tyr595, because in this region the alteredegion in the KO mice is included. The results have shown
357 pb product exclusively in WT but not in KO (Fig.B2). Furthermore, when the primers mp2x7 (which areirected to amplify from Ser23 to Tyr595) were used, similaresults were observed (Fig. 1B3). These results cannot bexplained by polymorphisms or gene duplication.
Moreover, quantitative RT-PCR experiments, usingpecific primers/probes that amplify between exons 5 and, have shown an increase in mRNA expression levels inO animals in respect to WT ones (Fig. 2A2). Due to theaqMan primers/probes specificity for P2X7 mRNA and
he presence of a single amplicon with exactly the sameize in both animals, the increase in P2X7 mRNA levels inO in respect to WT mice could be justified by the pres-
nce of diverse P2X7 mRNA truncated forms that include this sequence. Authors who generated the P2X7 KO modelid not exclude this possibility (Solle et al., 2001).
In addition, the immunochemical experiments showedhe presence of a “P2X7-like” receptor in the KO miceidbrain synaptic terminals with an identical expressionattern in immunostaining and in Western blot experimentsrom KO and WT midbrain mice synaptosomes (Fig. 4). Asndicated previously, some authors consider that the com-
ercial anti-P2X7 antibodies could exhibit cross-reactivityith other protein sequences (Anderson and Nedergaard,006). In this sense, Denlinger et al. (2001) have sug-ested that P2X7 C-terminal domain contains a conservedPS-binding domain. This could imply that anti-P2X7 anti-odies that recognize intracellular epitopes could be bind-
ng to this domain. However, in a previous work we showedhat P2X7 receptor is not present in macrophages from2X7 KO animals (Sánchez-Nogueiro et al., 2005). If com-ercial antibodies were recognizing the LPS protein this
hould be true in macrophages and not only in brain.oreover, we developed a BLAST against Gene BankTM
ata base to study how similar the P2X7 carboxy-terminals in respect to other proteins. We have not found a 100%dentical protein. Moreover, we carried out the sequencenalysis of this region (P2X7 C-terminal domain) and webserved that the 19 residues were exactly the same inoth animal models used in our study (data not shown). So
t is impossible that the region belongs to a LPS bindingrotein or even others known up to now.
Altogether, these results confirm the presence of func-ional P2X7 receptor in mice midbrain synaptosomes, show-ng the existence in P2X7 KO CNS mice of a subunit, withunctional properties similar but not identical to “native” P2X7.nother observation that points to the existence of a “P2X7-
ike” receptor in KO CNS mice is the fact that in thesenimals no other P2X subunits compensate the lack of the2X7 receptor (Fig. 2A1). This lack of compensation byther P2X subunits can be explained by the existence, at
east in KO CNS mice, of a “P2X7-like” receptor that as-umes the role of the “native” P2X7 subunit when this isbsent.
CONCLUSION
o summarize, these observations, together with the pre-ious results obtained in our laboratory (Sánchez-ogueiro et al., 2005) point to the existence, at least in theO mice brain, of a functional “P2X7-like” receptor whichust proceed from an alternative gene expression orseudogene, because other explanations, as polymor-hism or gene duplication, are unlikely. This “P2X7-like”hould exhibit molecular modification in respect to thenative” P2X7 receptor affecting the agonist binding site orhe ionic permeability, which confers a characteristic andxclusive kinetic behavior and implies a reduction in thehannel permeability. These results confirm the role ofnative” P2X7 receptor at the synaptic terminals in CNS.
cknowledgments—The authors are very grateful to Dr. C. A.abel (from Pfizer Inc., Groton, CT, USA) for KO generation and
o Dr. J. P. Dehaye (Laboratoire de Biochimie et de Biologie

CBwoCid
A
A
A
B
B
C
D
D
D
D
D
F
F
G
H
H
H
I
J
K
L
L
L
L
M
M
N
N
P
P
P
R
R
S
S
S
P. Marín-García et al. / Neuroscience 151 (2008) 361–373372
ellulaire, Institut de Pharmacie, Université Libre de Bruxelles,russels, Belgium) for his generous gift of the P2X7 KO mice. Thisork was supported by research grants from the Spanish Ministryf Education and Science BFU2005-02079, the Fundación Laaixa n° BM05-114-0, the Fundación Marcelino Botín, the Span-
sh Ministry of Education and Science AP2002-0059 and the Juntae Comunidades de Castilla La Mancha.
REFERENCES
bbracchio M, Burnstock G (1994) Purinoceptors: are there familiesof P2X and P2Y purinoceptors? Pharmacol Ther 64:445–475.
nderson CM, Nedergaard M (2006) Emerging challenges of assign-ing P2X7 receptor function and immunoreactivity in neurons.Trends Neurosci 29:257–262.
rmstrong JN, Brust TB, Lewis RG, MacVicar BA (2002) Activation ofpresynaptic P2X7-like receptors depresses mossy fiber-CA3 syn-aptic transmission through p38 mitogen-activated protein kinase.J Neurosci 22:5938–5945.
ianco F, Ceruti S, Colombo A, Fumagalli M, Ferrar D, Pizzirani C,Matteoli M, Di Virgilio F, Abbracchio MP, Verderio C (2006) Arole for P2X7 in microglial proliferation. J Neurochem 99:745–758.
urnstock G (2005) Purinergic signalling: therapeutic potential. An RAcad Nac Farm 71:283–319.
homczynski P (1993) A reagent for the single-step simultaneousisolation of RNA, DNA and proteins from cell and tissue samples.Biotechniques 15:532–537.
eng Z, Fyffe EW (2004) Expression of P2X7 receptor immunoreac-tivity in distinct subsets of synaptic terminals in the ventral horn ofrat lumbar spinal cord. Brain Res 1020:53–61.
enlinger LC, Fisette PL, Sommer JA, Watters JJ, Prabhu U, DubyakGR, Proctor RA, Bertics PJ (2001) Cutting edge: the nucleotidereceptor P2X7 contains multiple protein- and lipid-interaction motifsincluding a potential binding site for bacterial lipopolysaccharide.J Immunol 167:1871–1876.
euchars SA, Atkinson L, Brooke RE, Musa H, Miligan CJ, BattenTFC, Buckley NJ, Parson SH, Deuchars J (2001) Neuronal P2X7
receptors are targeted to presynaptic terminals in the central andperipheral nervous system. J Neurosci 21:7143–7152.
onnelly-Roberts DL, Jarvis MF (2007) Discovery of P2X7 receptor-selective antagonists offers new insights into P2X7 receptor func-tion and indicates a role in chronic pain states. Br J Pharmacol151:571–579.
unkley PR, Jarvie PE, Heath JW, Kidd GJ, Rostas JA (1986) A rapidmethod for isolation of synaptosomes on Percoll gradients. BrainRes 372:115–129.
ranke H, Klimke K, Brinckmann U, Grosche J, Francke M, SperlaghB, Reinchenbach A, Liebert UG, Illes P (2005) P2X7 receptor-mRNA and -protein in the mouse retina: changes during retinaldegeneration in BALBCrds mice. Neurochem Int 47:235–242.
ranke H, Illes P (2006) Involvement of P2 receptors in the growthand survival of neurons in the CNS. Pharmacol Ther 109:197–324.
rynkiewicz G, Poeni M, Tsien RY (1985) A new generation of fluo-rescent calcium indicators with greatly improved fluorescenceproperties. J Biol Chem 260:3440–3450.
ervás C, Pérez-Sen R, Miras-Portugal MT (2003) Coexpression offunctional P2X and P2Y nucleotide receptors in single cerebellargranule cells. J Neurosci Res 73:384–399.
ervás C, Pérez-Sen R, Miras-Portugal MT (2005) Presence of di-verse functional P2X receptors in rat cerebellar synaptic terminals.Biochem Pharmacol 70:770–785.
onore P, Donnelly-Roberts D, Namovic MT, Hsieh G, Zhu CZ,Mikusa JP, Hernandez G, Zhong C, Gauvin DM, Chandran P,
Harris R, Perez Medrano A, Carroll W, Marsh K, Sullivan JP,Faltynek CR, Jarvis M (2006) A-740003 [N-(1-{[(cyanoimino)(5-quinolinylamino)methyl]amino}-2,2-dimethylpropyl)-2-(3,4-dimeth-oxyphenyl)acetamide], a novel and selective P2X7 receptors an-tagonist, dose-dependently reduces neuropathic pain in the rat.J Pharmacol Exp Ther 319:1376–1385.
reland MF, Noakes PG, Bellingham MC (2004) P2X7-like receptorsubunits enhance excitatory synaptic transmission at centralsynapses by presynaptic mechanisms. Neuroscience 128:269 –280.
iang LH, Mackenzie AB, North RA, Surprenant A (2000) Brilliant BlueG selectively blocks ATP-gated rat P2X7 receptors. Mol Pharmacol58:82–88.
e HZ, Qi H, Weidema AF, Zhang Q, Panupinthu N, Crawford DT,Grasser WA, Paralkar VM, Li M, Audoly LP, Gabel CA, Jee WS,Dixon SJ, Sims SM, Thompson DD (2003) Deletion of the P2X7
nucleotide receptor reveals its regulatory roles in bone formationand resorption. Mol Endocrinol 17:1356–1367.
abasi JM, Petrushova N, Donova C, McCurdy S, Lira P, Payette MM,Brissette W, Wicks JR, Audoly LP, Gabel CA (2002) Absence ofthe P2X7 receptor alters leukocyte function and attenuates aninflammatory response. J Immunol 168:6436–6445.
eón D, Hervás C, Miras-Portugal MT (2006) P2Y1 and P2X7 recep-tors induce calcium/calmodulin-dependent protein kinase II phos-phorylation in cerebellar granule neurons. Eur J Neurosci 23:2999–3013.
ivak KJ, Schmittgen TD (2001) Analysis of relative gene expressiondata using real-time quantitative PCR and the 2-��CT method.Methods 25:402–408.
undy PM, Hamilton MG, Mi L, Gong W, Vair C, Sawyer TW, Frew R(2002) Stimulation of Ca2� influx through ATP receptors on ratbrain synaptosomes: identification of functional P2X7 receptor sub-types. Br J Pharmacol 135:1616–1626.
iras-Portugal MT, Díaz-Hernández M, Giráldez L, Hervás C, Gómez-Villafuertes R, Sen RP, Gualix J, Pintor J (2003) P2X7 receptors inrat brain: presence in synaptic terminals and granule cells. Neuro-chem Res 28:1597–1605.
oores TS, Hasdemir B, Vega-Riveroll L, Deuchars J, Parson SH(2005) Properties of presynaptic P2X7-like receptors at the neuro-muscular junction. Brain Res 1034:40–50.
orth RA (2002) Molecular physiology of P2X receptors. Physiol Rev82:1013–1067.
orth RA, Surprenant A (2002) Pharmacology of cloned P2X recep-tors. Annu Rev Pharmacol Toxicol 40:563–580.
atti L, Raiteri L, Grilli M, Parodi M, Raiteri M, Marchi M (2006) P2X7
receptors exert a permissive role on the activation of release-enhancing presynaptic �7 nicotinic receptors co-existing on ratneocortex glutamatergic terminals. Neuropharmacology 50:705–713.
intor J, Miras-Portugal MT (1995) A novel receptor for diadenosinepolyphosphates coupled to calcium increase in rat midbrain syn-aptosomes. Br J Pharmacol 115:895–902.
ochet S, Gómez-Muñoz A, Marino A, Dehaye JP (2003) Regulationof phospholipase D by P2X7 receptors in submandibular ductalcells. Cell Signal 15:927–935.
alevic V, Burnstock G (1998) Receptors for purines and pyrimidines.Pharmacol Rev 50:413–492.
oberts JA, Vial C, Digby HR, Agboh KC, Wen H, Atterbury-ThomasA, Evans RJ (2006) Molecular properties of P2X receptors. PlugersArch Eur J Physiol 452:486–500.
ánchez-Nogueiro J, Marín-García P, Miras-Portugal MT (2005)Characterization of a functional P2X7-like receptor in cerebellargranule neurons from P2X7 knockout mice. FEBS Lett 579:3783–3788.
olle M, Labasi J, Perregaux DG, Stam E, Petrushova N, Koller BH,Griffiths RJ, Gabel CA (2001) Altered cytokine production in micelacking P2X7 receptors. J Biol Chem 276:125–132.
perlágh B, Köfalvi A, Deuchars J, Atkinson L, Milligan CJ, Buckley
NJ, Vizi ES (2002) Involvement of P2X7 receptors in the regulation
S
S
T
V
W
X
P. Marín-García et al. / Neuroscience 151 (2008) 361–373 373
of neurotransmitter release in the rat hippocampus. J Neurochem81:1196–1211.
perlágh B, Vizi ES, Wirkner K, Illes P (2006) P2X7 receptors in thenervous system. Prog Neurobiol 78:327–346.
uadicani OS, Brosnan CF, Scemes E (2006) P2X7 receptors mediateATP release and amplification of astrocytic intracellular Ca2� sig-naling. J Neurosci 26:1378–1385.
orres GE, Egan TM, Voigt MM (1999) Hetero-oligomeric assembly ofP2X receptor subunits. Specificities exist with regard to possible
partners. J Biol Chem 274:6653–6659.irginio C, Church D, North RA, Surprenant A (1997) Effects of diva-lent cations, protons and calmidazolium at the rat P2X7 receptor.Neuropharmacology 36:1285–1294.
irkner K, Köfalvi A, Fischer W, Günther A, Franke H, Gröger-Arndt H,Nörenberg W, Madarász E, Vizi ES, Schneider E, Sperlágh B, IllesP (2005) Supersensitivity of P2X7 receptors in cerebrocortical cellcultures after in vitro ischemia. J Neurochem 95:1421–1437.
iang Z, Burnstock G (2005) Changes in expression of P2X purino-ceptors in rat cerebellum during postnatal development. Brain Res
Dev Brain Res 156:147–157.(Accepted 12 November 2007)(Available online 12 November 2007)





























