Embed Size (px)
Citation preview

www.elsevier.com/locate/yabbi
ABBArchives of Biochemistry and Biophysics 457 (2007) 187–196
Substrate specificity of recombinant dengue 2 virus NS2B-NS3protease: Influence of natural and unnatural basic amino acids
on hydrolysis of synthetic fluorescent substrates
I.E. Gouvea a, M.A. Izidoro a, W.A.S. Judice a, M.H.S. Cezari a, G. Caliendo b,V. Santagada b, C.N.D. dos Santos c, M.H. Queiroz c, M.A. Juliano a,
P.R. Young d, D.P. Fairlie e, L. Juliano a,*
a Departmento de Biofısica, Escola Paulista de Medicina, Universidade Federal de Sao Paulo, Rua Tres de Maio, 100 - Sao Paulo 04044-020, Brazilb Dipartimento di Chimica Farmaceutica e Tossicologica, Universita di Napoli ‘‘Federico II’’, Via D. Montesano, 49-80131 Naples, Italy
c Instituto de Biologia Molecular do Parana, Rua Prof. Algacyr Munhoz Mader 3775, CIC Curitiba 81350-010, Brazild School of Molecular & Microbial Sciences University of Queensland, St. Lucia, 4072 Brisbane, Australia
e Institute for Molecular Bioscience, University of Queensland, St. Lucia, 4072 Brisbane, Australia
Received 22 August 2006, and in revised form 30 October 2006Available online 16 November 2006
Abstract
A recombinant dengue 2 virus NS2B-NS3 protease (NS means non-structural virus protein) was compared with human furin for thecapacity to process short peptide substrates corresponding to seven native substrate cleavage sites in the dengue viral polyprotein. Usingfluorescence resonance energy transfer peptides to measure kinetics, the processing of these substrates was found to be selective for theDengue protease. Substrates containing two or three basic amino acids (Arg or Lys) in tandem were found to be the best, with Abz–AKRRSQ–EDDnp being the most efficiently cleaved. The hydrolysis of dipeptide substrates Bz–X–Arg–MCA where X is a non-naturalbasic amino acid were also kinetically examined, the best substrates containing aliphatic basic amino acids. Our results indicated thatproteolytic processing by dengue NS3 protease, tethered to its activating NS2B co-factor, was strongly inhibited by Ca2+ and kosmo-tropic salts of the Hofmeister’s series, and significantly influenced by substrate modifications between S4 and S06. Incorporation of basicnon-natural amino acids in short peptide substrates had significant but differential effects on Km and kcat, suggesting that further dissec-tion of their influences on substrate affinity might enable the development of effective dengue protease inhibitors.� 2006 Elsevier Inc. All rights reserved.
Keywords: Protease; Peptides; Dengue
The flaviviruses comprise a group of about 70 positive-stranded RNA viruses including the causative agents ofDengue, West Nile, Yellow fever, and Japanese Encephali-tis infections. Dengue fever and dengue hemorrhagic fever(DHF)1/dengue shock syndrome (DSS) are caused by oneof four serotypes of dengue virus (serotypes 1–4), with upto 100 million infections per year. Approximately 2.5
0003-9861/$ - see front matter � 2006 Elsevier Inc. All rights reserved.
doi:10.1016/j.abb.2006.11.005
* Corresponding author. Fax: +55 11 5575 90 40.E-mail address: [email protected] (L. Juliano).
1 Abbreviations used: DHF, dengue hemorrhagic fever; DSS, dengueshock syndrome; NS, non-structural.
billion people are at risk globally, mostly in tropical andsub-tropical regions, reflecting the distribution of the insectvector. Classical dengue fever has been recorded for manycenturies; however a marked increase in the more seriouscomplication, DHF/DSS has occurred only over the lastfew decades. DHF/DSS is associated with significant mor-tality, particularly in the pediatric population [1–3] and novaccine or therapeutic treatment is currently available.
The dengue virus single positive sense RNA genome isapproximately 11 kb and codes for a single polyproteinprecursor. This precursor traverses the endoplasmic reticu-lum (ER) membrane multiple times, and is processed

188 I.E. Gouvea et al. / Archives of Biochemistry and Biophysics 457 (2007) 187–196
co- and post-translationally into three structural proteins,namely, C (core), prM (precursor to membrane), and E(envelope) and seven non-structural (NS) proteins NS1,NS2A, NS2B, NS3, NS4A, NS4B, and NS5 [4]. These pro-cessing events are essential for virus replication and requirethe host proteases called signalase and furin [5,6] as well asa two-component viral protease, NS2B/NS3 [4]. The ami-no-terminal 180 amino acids of the multifunctional proteinNS3 complexed with NS2B is responsible for cleavages onthe cytoplasmic side of the ER membrane at the junctionsbetween NS2A-NS2B, NS2B-NS3, NS3-NS4A, andNS4B-NS5 as well as at internal sites within C, NS2A,NS3, and NS4A. The host cell signalase mediates cleavageson the other face of the membrane, in the ER lumen. Laterin virion maturation, prM is processed to M protein by ahost protease called furin in a post-Golgi compartment togenerate M [5,6].
Dengue NS3 protease activity does not depend on thepresence of the complete sequence of NS2B co-factor(CF), but does need a central 40 residue relatively hydro-philic domain of NS2B (CF40) to be catalytically active[6–8]. The remaining hydrophobic residues of NS2B arethought to anchor the complex to cellular membranes.Although human hepatitis C virus (HCV) NS3 proteaseis activated by a shorter synthetic peptide co-factor [9],dengue NS3 protease is only active when co-expressed withthe NS2B hydrophilic domain probably due to comple-mentary folding of enzyme with co-factor [8,10].
Dengue 2 virus NS2B/NS3 is a trypsin-like serine prote-ase with a catalytic triad composed of His51, Asp75, andSer135 folded with two anti-parallel, six-stranded b-barrelsymmetry [11]. The dengue NS2B/NS3 protease hydrolyzesits natural polyprotein substrate on the C-terminal sideafter a pair of basic residues (Lys–Arg, Arg–Arg, or Arg–Lys) or, occasionally, after Gln–Arg, interacting in the sub-sites S2 and S1 followed by small side chain amino acids(Gly, Ser, or Ala) in the subsite S01 [4]. The best recombinantconstruct so far described for expression of a soluble, activedengue protease that is resistant to autolysis was based onmodeling of the dengue virus NS3 protease complexed withits co-factor [12]. The complex contains the catalyticallyactive NS3 protease (NS3pro) connected to CF40 via anon-cleavable and flexible non-apeptide, Gly4-Ser-Gly4
linker. The fusion protein was designated CF40glyNS3pro,and this chimeric dengue 2 virus protease hydrolyzed syn-thetic chromogenic peptides containing the non-prime sideresidues of the cleavage site of dengue virus polyprotein [8].
In the present paper, we report the proteolytic activity ofa recombinant dengue 2 virus (strain NGC) chimeric prote-ase (CF40glyNS3pro) on fluorescence resonance energytransfer (FRET) peptides based on native viral polyproteinprocessing sites. In order to check whether amino acid sub-stitutions between viral strains can affect differences in pro-tease activity, we included the protease CF40glyNS3proderived from a recent low passage DEN2 isolate from aBrazilian DF case (strain BR/01-EN). The same peptidesubstrates were also assayed against recombinant human
furin to check for specificity of the dengue polyproteincleavage sites. As P3 to P1 positions of the cleavage sitesin dengue polyproteins have basic amino acids, we exploredin more detail the susceptibility to hydrolysis of FRET pep-tides containing two or three basic amino acids having mostof the combinations of Arg and Lys in tandem sequence.
Finally, in order to obtain further information for designof NS2-NS3 enzyme inhibitors, we explored the susceptibil-ity to hydrolysis of even shorter fluorescent substrates ofgeneral structure Bz–X–Arg–MCA, where X was a non-natural basic amino acid that combined a positively chargedgroup with an aromatic or aliphatic substituent at the sameside chain. Examples of X were 4-aminomethyl-phenylala-nine (Amf), 4-guanidine phenylalanine (Gnf), 4-amino-methyl-N-isopropyl-phenylalanine (Iaf), 3-pyridyl-alanine(Pya), 4-piperidinyl-alanine (Ppa), 4-aminomethyl-cyclo-hexyl-alanine (Ama), and 4-aminocyclohexyl-alanine(Aca) (see structures in Fig. 1). This P2 position seemed suit-able for the introduction of non-natural basic amino acidsbecause of the observed restricted specificity of S1 subsiteof NS2B-NS3 enzyme to Arg or Lys.
Recently, after completing this work, the functional pro-filing of the proteases of the four dengue serotypes wasreported using fluorescent peptide substrate libraries [13],and Niyomrattanakit et al. [14] reported an analysis ofthe substrate specificity of the NS3 of serotype 2 usinginternally quenched fluorescent peptides. However, ourresults using FRET peptide substrates comprising naturaldengue virus polyprotein sequences as well as unnaturalbasic amino acids provide additional information aboutthe specificity of NS2B-NS3 enzyme and further definethe best composition of natural and non-natural basicamino acids at the non-prime side substrate residues.
Materials and methods
Enzymes
Recombinant NS3 protease dengue 2 virus, strain NGC (CF40glyNS3pro), was obtained and purified as described previously [8]. Briefly thepQE9.CF40.gly.NS3pro (CF40 fused to NS3pro via a Gly4SerGly4 linker)vector was used for high level, inducible expression of amino-terminalhexahistidine-tagged recombinant enzyme. Cultures of Escherichia coli
strain SG13009, transformed with the expression plasmid, were grown in2 l of LB medium containing 100 lg ml�1 ampicillin and 25 lg ml�1
kanamycin at 37 �C until the A600 nm reached 0.6. The cells were inducedfor expression by the addition of isopropyl-b-D-thiogalactopyranose to afinal concentration of 1 mM and incubated for an additional 3 h at 30 �C.The cells were harvested by centrifugation and resuspended in 1 ml of lysisbuffer (50 mM Hepes, pH 7.5, 300 mM NaCl, 5% glycerol)/10 ml of ori-ginal culture and subjected to probe sonication (five 30-s pulses) on ice andthen centrifuged at 27,000g for 30 min at 4 �C. The supernatant waspurified by passage through a 2-ml column of Ni2+ nitrilotriacetic acid–agarose (Qiagen) pre-equilibrated with 50 mM Hepes, pH 7.5, containing300 mM NaCl. The column was extensively washed with buffer containing20 mM imidazole, and protein was then eluted from the column in buffercontaining 100 mM imidazole. Elution fractions were analyzed by 15%SDS–PAGE. Samples of pre- and post-induced cells as well as soluble andinsoluble fractions following lysis were collected and also analyzed by 15%SDS–PAGE. The hydrolytic activities of all samples were also checked onZ-RR-MCA in order to discriminate contamination activity of bacterial
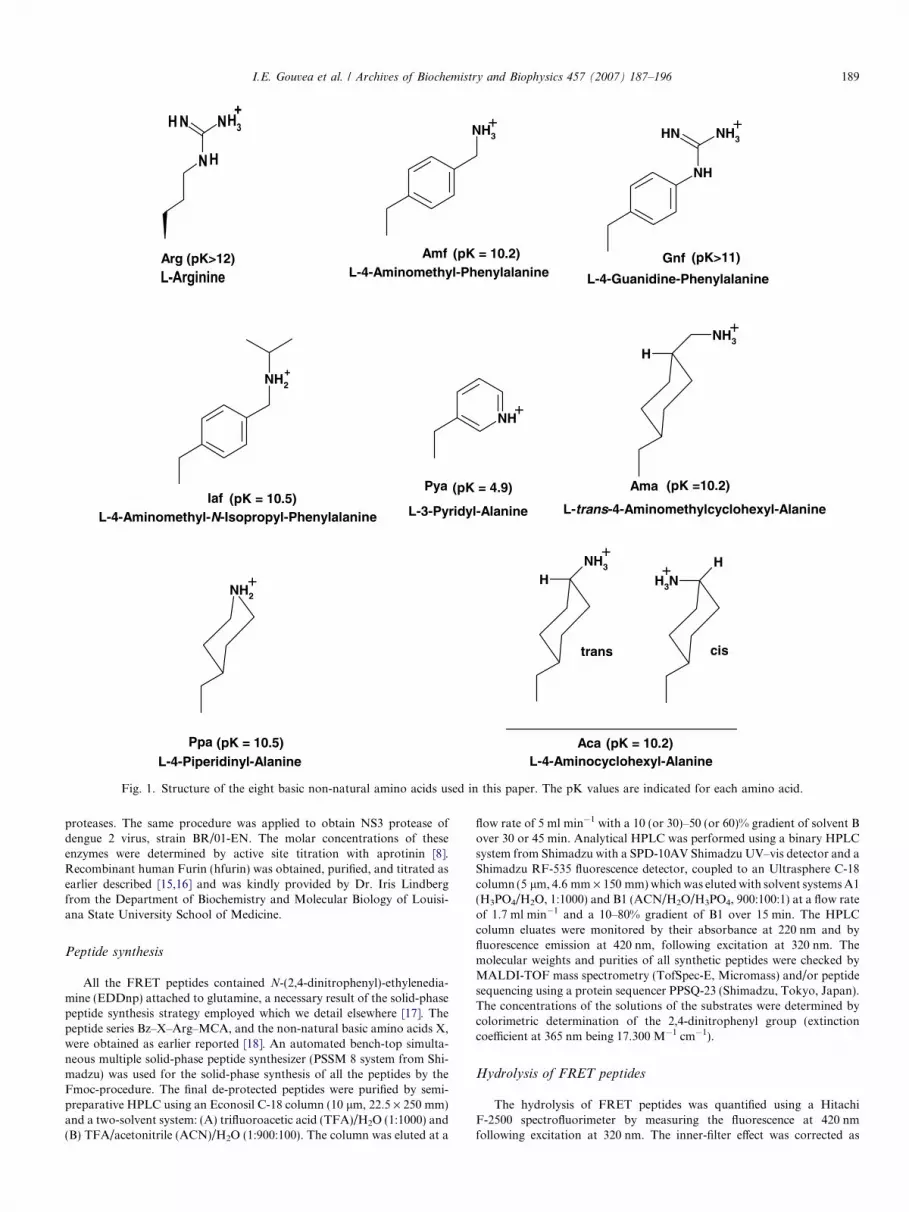
Fig. 1. Structure of the eight basic non-natural amino acids used in this paper. The pK values are indicated for each amino acid.
I.E. Gouvea et al. / Archives of Biochemistry and Biophysics 457 (2007) 187–196 189
proteases. The same procedure was applied to obtain NS3 protease ofdengue 2 virus, strain BR/01-EN. The molar concentrations of theseenzymes were determined by active site titration with aprotinin [8].Recombinant human Furin (hfurin) was obtained, purified, and titrated asearlier described [15,16] and was kindly provided by Dr. Iris Lindbergfrom the Department of Biochemistry and Molecular Biology of Louisi-ana State University School of Medicine.
Peptide synthesis
All the FRET peptides contained N-(2,4-dinitrophenyl)-ethylenedia-mine (EDDnp) attached to glutamine, a necessary result of the solid-phasepeptide synthesis strategy employed which we detail elsewhere [17]. Thepeptide series Bz–X–Arg–MCA, and the non-natural basic amino acids X,were obtained as earlier reported [18]. An automated bench-top simulta-neous multiple solid-phase peptide synthesizer (PSSM 8 system from Shi-madzu) was used for the solid-phase synthesis of all the peptides by theFmoc-procedure. The final de-protected peptides were purified by semi-preparative HPLC using an Econosil C-18 column (10 lm, 22.5 · 250 mm)and a two-solvent system: (A) trifluoroacetic acid (TFA)/H2O (1:1000) and(B) TFA/acetonitrile (ACN)/H2O (1:900:100). The column was eluted at a
flow rate of 5 ml min�1 with a 10 (or 30)–50 (or 60)% gradient of solvent Bover 30 or 45 min. Analytical HPLC was performed using a binary HPLCsystem from Shimadzu with a SPD-10AV Shimadzu UV–vis detector and aShimadzu RF-535 fluorescence detector, coupled to an Ultrasphere C-18column (5 lm, 4.6 mm · 150 mm) which was eluted with solvent systems A1(H3PO4/H2O, 1:1000) and B1 (ACN/H2O/H3PO4, 900:100:1) at a flow rateof 1.7 ml min�1 and a 10–80% gradient of B1 over 15 min. The HPLCcolumn eluates were monitored by their absorbance at 220 nm and byfluorescence emission at 420 nm, following excitation at 320 nm. Themolecular weights and purities of all synthetic peptides were checked byMALDI-TOF mass spectrometry (TofSpec-E, Micromass) and/or peptidesequencing using a protein sequencer PPSQ-23 (Shimadzu, Tokyo, Japan).The concentrations of the solutions of the substrates were determined bycolorimetric determination of the 2,4-dinitrophenyl group (extinctioncoefficient at 365 nm being 17.300 M�1 cm�1).
Hydrolysis of FRET peptides
The hydrolysis of FRET peptides was quantified using a HitachiF-2500 spectrofluorimeter by measuring the fluorescence at 420 nmfollowing excitation at 320 nm. The inner-filter effect was corrected as

190 I.E. Gouvea et al. / Archives of Biochemistry and Biophysics 457 (2007) 187–196
previously described [19]. When Bz-RR-MCA peptide was used, thecondition was changed to 460 and 380 nm, respectively.
The recombinant dengue 2 proteases were incubated in 50 mM Trisbuffer (pH 9.0, 10 mM NaCl, 20% glycerol) for 2 min at 37 �C and thereaction started by addition of the substrate. Furin assays were performedin 100 mM MES buffer, pH 7.0 containing 1 mM CaCl2. The kineticparameters kcat and Km were calculated by non-linear regression dataanalysis using the GraFit version 5.0 program (Erithacus Software, Horley,Surrey, UK). The peptide bonds cleaved in each substrate were identified byMALDI-TOF mass spectrometry analysis of the fragments after isolationby HPLC. Ki values were calculated by monitoring the hydrolysis of Z-RR-MCA in the presence of inhibitory FRET peptides as described at [20].
Enzyme assays
Dengue NS3 pH-profiles were obtained by measuring the kineticparameters of hydrolysis, kcat, Km and kcat/Km over a pH range of 7.0–11.0adjusted with 2 M NaOH and HCl. These determinations were carried outin a four-component buffer comprised of 25 mM acetic acid, 25 mM Mes,75 mM Tris and 25 mM glycine. Enzymatic activity was measured at37 �C, using the fluorimetric assay described above and the data werefitted with the GraFit software to the follow equation:
kcat=Km ¼kcat=KmðLimitÞ10pH�pKa1
102pH�pKa1�pKa2
where kcat/Km (limit) is the highest value of this kinetic parameter.The influences of salts (NaCl, Na2SO4, Na2CO3, NaOAc, Na2PO4 and
CaCl2), glycerol and detergents were investigated in 50 mM Tris, pH 9.0,using Bz–RR–MCA as substrate. The initial velocity was measured at37 �C with an enzyme NS3NGC concentration of between 22.6 and116 nM. All the kinetic data are results of at least three determinationusing different substrate concentrations and the standard errors were lessthan 5% for all determinations.
HPLC stopped kinetics
Peptide concentrations ranging from 10 to 150 lM were treated withdengue NS3 protease at 37 �C to achieve 10–15% cleavage. Reactions werestopped by the addition of trifluoroacetic acid to 0.33% and percentcleavage was measured by reverse phase-HPLC.
Docking of the best substrate to active site
Docking of the optimized substrate Abz–AKRRSQ–EDDnp wascarried out using GOLD v.2.1.2. The crystal structure of the NS3 proteasewas extracted from the PDB database (code 1DF9). The macromoleculardocking of extremely flexible molecules such as peptides into protein activesites is problematic due to the inherent difficulty of sufficiently samplingthe conformation space available to the ligands and also the comparativelyshort energy minimization routines commonly used in virtual screening/docking packages. In order to overcome the first of these limitations weutilized our knowledge of protease-substrate molecular recognition [21] toloosely constrain the substrates into an approximate extended b-strand.This enabled the side-chains to move and find their optimum location,whilst keeping the backbone in a biologically relevant conformation.
Results
Hydrolysis of peptide substrates corresponding to native
cleavage sites by dengue 2 virus proteases
CF40glyNS3pro chimeric proteases from dengue 2 virusstrains, NGC and BR/01-EN, tagged with six consecutivehistidines were purified and characterized [8]. There are10 amino acid differences in the protease encoding a region
of the genome between the two strains, all except one beingconservative. They are in the following positions; (BR/01-EN versus NGC) NS3proE20D, R28K, R61K, V77I,I114L, T119A, V139I, R141K, D168E, and Y179D. Nodifferences were found in the 40 amino acid CF40 activat-ing segment.
Seven FRET peptides containing the amino acidsequences that span the reported cleavage sites of polypro-tein processing by dengue 2 virus NS2B/NS3 of NGCstrain were synthesized with Abz (ortho-aminobenzoicacid) and EDDnp (N-[2,4-dinitrophenyl)-ethylenediamine)at N- and C-terminal end of each peptide, respectively.The FRET peptides were based on the polyprotein cleav-age sites: NS2A/NS2B, NS2B/NS3, NS3/NS4A, NS4B/NS5, Cint, NS3int, and NS4Aint. Table 1 shows the sequenc-es of these FRET peptides and the kinetic parameters fortheir hydrolysis by CF40glyNS3pro from NGC and BR/01-EN dengue 2 virus strains.
Optimal conditions of buffer composition and pH wereused for enzyme activity, as described for the hydrolysisof chromogenic peptidyl-pNa substrates [8] that character-istically has low salt concentration (see below the effects ofsalts in enzyme activity). All assayed peptides were cleavedonly at one peptide bond always hydrolyzed after the lastC-terminal basic amino acid even for peptides with fourconsecutive Arg residues. The sites of cleavage in theFRET peptides were the same as those in the polyprotein,indicating that the synthetic peptides containing Abz andEDDnp did not introduce restrictions or different interac-tions with the viral proteases. The kcat/Km values obtainedfor the hydrolysis of the seven FRET peptides byCF40glyNS3pro from NGC were approximately two timeshigher than those obtained from the BR/01-EN dengue 2virus enzyme and the best and the worst substrates werethe same for both enzymes. These results indicate thatthe structural differences between them do not affect theirpeptidase specificity but have some impact on catalyticefficiency.
Peptides 5 and 6 were hydrolyzed with the highest kcat/Km values. However, for the hydrolysis of peptide 6 the kcat
parameter was the highest among the assayed peptideswhile for the hydrolysis of peptide 5 the Km value wasthe lowest. Interestingly, peptide 5 had four consecutiveArg residues, whereas peptide 6 is the only substrate inTable 1 with two Arg residues at prime sites. Althoughthe recognition of basic residues at P2 and P1 positionsby the dengue protease is considered the key specificity fea-ture of such flavivirus enzymes [13,14,22–24], it is possiblethat an interdependence of the protease subsites whenoccupied by the substrate could justify the hydrolysis ofpeptide 2 and the resistance of peptide 7. In order to inves-tigate if the Km values obtained were not limited by innerfilter effects, HPLC stopped kinetic were performed withpeptide 1, the highest Km measurement in fluorescenceassays among natural cleaved sites. The value obtained(Km = 39.1 ± 5.7) is in agreement with the fluorescencevalue (Km = 30.0 ± 1.9)

Tab
le1
Kin
etic
par
amet
ers
for
the
hyd
roly
sis
by
CF
40gl
yNS
3pro
and
hfu
rin
of
syn
thet
icF
RE
Tp
epti
des
sub
stra
tes
bas
edo
np
oly
pro
tein
clea
vage
site
sb
yd
engu
evi
rus
NS
2B/N
S3
com
ple
xan
dh
ost
furi
n
Pep
tid
eN
o.
Seq
uen
ceA
bz–
pep
tid
yl–
ED
Dn
pC
leav
age
inp
oly
pro
tein
NS
3 NG
CN
S3 B
R/0
1h
Fu
rin
kca
t(s�
1)
Km
(lM
)k
cat/
Km
(mM
s)�
1k
cat/
Km
(mM
s)�
1k
cat
(s�
1)
Km
(lM
)k
cat/
Km
(mM
s)�
1
1R
TS
KK
RflS
WP
LN
EQ
2A/2
B0.
1730
5.7
2.9
0.02
0.44
382
EV
KK
QR
flA
GV
LW
DQ
2B/3
0.03
8.8
3.4
1.6
Res
ista
nt
3F
AA
GR
KflS
LT
LN
LQ
3/4A
0.02
17.7
1.2
0.6
Res
ista
nt
4T
TS
TR
RflG
TG
NIG
Q4B
/50.
0427
1.5
0.7
Res
ista
nt
5L
NR
RR
RflT
AG
MII
QC
int
0.04
3.9
10.2
5.1
0.23
0.33
697
6S
AA
QR
RflG
RIG
RN
QN
S3 i
nt
0.29
13.8
21.0
15.8
0.17
1.32
129
7E
PE
KQ
RT
PQ
DN
NS
4Ain
tR
esis
tan
tR
esis
tan
tR
esis
tan
tH
RR
EK
RflS
VA
LQ
prM
/M*
0.04
261.
50.
80.
810.
1845
00
Hyd
roly
sis
con
dit
ion
s:th
ep
rote
ases
wer
ein
cub
ated
in50
mM
Tri
sb
uff
er(p
H9.
0,10
mM
NaC
l,20
%gl
ycer
ol)
for
2m
inat
37�C
and
the
reac
tio
nst
arte
db
yad
dit
ion
of
the
sub
stra
te.
All
the
kin
etic
dat
aar
ere
sult
so
fat
leas
tth
ree
det
erm
inat
ion
usi
ng
diff
eren
tsu
bst
rate
con
cen
trat
ion
san
dth
est
and
ard
erro
rsw
ere
less
than
5%fo
ral
ld
eter
min
atio
ns.
I.E. Gouvea et al. / Archives of Biochemistry and Biophysics 457 (2007) 187–196 191
We also include in Table 1 the kinetic parameters forthe hydrolysis of Abz-HRREKRSVALQ-EDDnp, corre-sponding to the recognized cleavage site of furin withinthe dengue prM protein. This FRET peptide was poorlyhydrolyzed by CF40glyNS3pro, but it was very suscepti-ble to recombinant human furin (hfurin). All the otherpeptides in this series (Table 1) were also assayed againsthfurin, but only peptide 5 corresponding to the internalsite of the core protein (Cint) was significantly hydrolyzed.The other FRET peptides assayed against hfurin werepoorly hydrolyzed or resistant to hydrolysis.
In order to investigate the contribution of each aminoacid of the best substrate in its hydrolytic reaction withCF40glyNS3pro, we synthesized a series of peptidesderived from the best substrate in Table 1, namely peptide6, in which the amino acids were exchanged with those inthe corresponding position of peptide 3 (the worstsubstrate in Table 1). The kinetic parameters for theirhydrolysis are shown in Table 2. The biggest decrease inthe kcat/Km value was obtained with peptide 10, resultingfrom a single change in peptide 6 at P1 of an Arg for aLys. A significant increase in the kcat/Km value wasobserved with the hydrolysis of peptide 16 which corre-sponded to a substitution in peptide 6 at P06 of Asn byLeu. The biggest decreases in Km values were observed inpeptides 9, 15, and 16, which resulted from the replace-ments of Gln/Gly at P3, Arg/Asn at P05 and Asn/Leu atP06, respectively. The variations in the Km values are com-pensated by the kcat and in most of the cases the kcat/Km
values presented in Table 2 do not show large variations,except for peptide 10. Confirming the preference of theS1 subsite of dengue virus protease for Arg instead ofLys, we observed a significant increase in the efficiency ofhydrolysis of the peptide 3 homologue in which Lys wassubstituted by Arg (Abz–FAAGRRSLTLNLQ–EDDnp).The kinetic parameters for hydrolysis of this peptide were:kcat = 0.62 s�1, Km = 12.5 lM and kcat/Km = 5.0 s�1
mM�1.
Hydrolysis of FRET hexapeptide substrates containing two
or three natural basic amino acids
Table 3 shows the kinetic parameters for the hydrolysisof FRET hexapeptides that were designed to contain com-binations of two or three basics amino acids in tandemarrangement flanked by small side chain amino acids(peptides 17–28 in Table 3). All the susceptible peptideswere hydrolyzed at the carboxyl side of the last basic ami-no acid of the peptide sequence. The Km values of hydro-lysis of this series of FRET peptides are in generalsignificantly higher than those for hydrolysis of the 12 res-idue FRET peptides derived from the dengue virus poly-protein (Tables 1 and 2). The highest Km values wereobtained with substrates containing a pair of Lys at posi-tions P1 and P2 (see the peptides 18, 25, and 26) and thelowest Km values were observed with peptides containingthe pair Arg–Lys (see the peptides 19 and 24). In contrast,

Table 2Kinetic parameters for the hydrolysis of FRET peptides derived from Abz–SAAQRRGRIGRNQ–EDDnp (peptide 6) by CF40glyNS3pro
Peptide No. Positions in the substrates (P6–P01) kcat (s�1) Km (lM) kcat/Km (mM s)�1
P6 P5 P4 P3 P2 P1 P01 P02 P03 P04 P05 P06 P07
3 F A A G R K S L T L N L Q 0.02 17.7 1.1
Modifications of peptide 6 with the amino acids of peptide 3
6 S A A Q R R G R I G R N Q 0.29 13.8 21.18 F 0.14 7.4 19.49 G 0.09 4.0 22.0
10 K 0.04 9.8 4.611 S 0.14 5.9 23.512 L 0.11 7.2 15.513 T 0.34 16.5 20.914 L 0.18 6.9 26.515 N 0.11 4.6 23.116 L 0.16 4.5 35.9
Hydrolysis conditions and the errors of the kinetic parameter determinations were similar as described in Table 1. The standard errors were less than 5%for all determinations.
Table 3Kinetic parameters for hydrolysis by CF40glyNS3pro of model FRET peptides containing two or three basic amino acids
Peptide No. Sequence Abz–peptidyl–EDDnp kcat (s�1) Km (lM) kcat/Km (mM s)�1
17 AGRRflSAQ 0.21 20.0 10.518 AGKKflSAQ 0.02 58.8 0.319 AGRKflSAQ 0.13 21.3 6.120 AGKRflSAQ 0.20 38.0 5.321 ARRRflSQ 0.38 25.6 14.822 AKRRflSQ 1.0 31.4 31.823 ARKRflSQ 0.07 58.0 1.224 ARRKflSQ 0.09 18.3 4.925 ARKKflSQ 0.05 67.3 0.726 AKKKflSQ 0.04 69.2 0.627 AKRKflSQ 0.04 23.7 1.728 AKKRflSQ 0.07 53.2 1.329 AGRRPAQ Resistant (Ki = 11.8 lM)30 AKRRRPQ Resistant (Ki = 15.5 lM)
Z–RRfl–MCA 0.11 247 0.5
The arrows indicate the cleavage sites, which were determined by MALDI-TOF mass spectrometry of all reaction mixture after previous desalting and byisolation of the product of hydrolysis and the composition determined by amino acid analysis. Hydrolysis conditions of the kinetic parameter determi-nations were similar as described in Table 1. The standard errors were less than 5% for all determinations.
192 I.E. Gouvea et al. / Archives of Biochemistry and Biophysics 457 (2007) 187–196
the highest kcat values were obtained for the substrateswith the pair Arg–Arg (see the peptides 21 and 22). Thepresence of a third basic amino acid at the P3 position sig-nificantly increased only the kcat value of hydrolysis for thepeptides containing the Arg–Arg pair at P1 and P2. Thebest substrate derived from the combinations of the basicamino acids Lys and Arg was Abz-AKRRSQ-EDDnp(peptide 22), which was hydrolyzed with the highestkcat/Km value.
In accordance with these results, a similar fluorescent sub-strate, Bz-nKRR-ACMC (n = norleucine and ACMC =7-amino-3-carbamoylmethyl-4-methyl coumarin), wasdescribed as the best substrate for all four dengue serotypeNS2B/NS3 proteases based on the analysis of tetrapeptidepositional scanning synthetic combinatorial libraries [13].It is noteworthy that CF40glyNS3pro does not cleavebetween two basic residues in all the peptides of Table 3,indicating that the enzyme does not accept basic amino acids
at the P01 position. In order to verify if basic residues in P01impeded hydrolysis of the substrates, two peptides were syn-thesized containing Pro after the basic sequence of two andfour basic residues (peptides 29 and 30). Both peptides wereresistant to hydrolysis but they inhibited the enzyme with Ki
values in the lM range (Table 3). Under the same conditions,the commonly used commercially available peptide, Z–RR–MCA, was hydrolyzed with a high Km value (Table 3).
Hydrolysis of fluorescent peptide substrates containing basic
non-natural amino acids
The series of fluorescent peptides Bz–XR–MCA, whereX is a basic non-natural amino acid, is shown in Fig. 1.They were assayed as substrates against CF40glyNS3proand the kinetic parameters for their hydrolysis are shownin Table 4. The peptides containing 4-aminomethyl-phenyl-alanine (Amf—peptide 32) and 4-aminocyclohexyl-alanine

pH6 7 8 9 10 11 12
Rel
ativ
e A
ctiv
ity(%
)
0
20
40
60
80
100
Fig. 2. pH dependency profile for Dengue NS3 protease activity. Param-eters of hydrolysis, kcat, Km and the relationship kcat/Km were obtained foreach pH in conditions described in Materials and methods. kcat/Km wereplotted as percentage relative activities assigning the highest catalyticefficiency as 100%. Representative peptide substrates Abz-SAAQRR-GRIGRNQ-EDDnp (peptide 6—d) and Abz-FAAGRKSLTLNLQ-EDDnp (peptide 3—s) are shown.
Table 5pK values obtained from the pH-profile hydrolysis curves of syntheticFRET peptides substrates based on polyprotein cleavage sites by dengueNS2B/NS3 protease complex
Peptide No. Sequence Abz–peptidyl–EDDnp pK1 pK2
1 RTSKKRflSWPLNEQ 8.6 ± 0.2 8.9 ± 0.22 EVKKQRflAGVLWDQ 8.2 ± 0.2 9.4 ± 0.23 FAAGRKflSLTLNLQ 8.3 ± 0.2 9.2 ± 0.24 TTSTRRflGTGNIGQ 8.1 ± 0.1 9.9 ± 0.15 LNRRRRflTAGMIIQ 8.3 ± 0.1 10.2 ± 0.16 SAAQRRflGRIGRNQ 8.5 ± 0.2 10.4 ± 0.2
Z-RR-MCA 8.5 ± 0.1 10.0 ± 0.1
Salt (mM)0 100 200 300 400 500
Rel
ativ
e A
ctiv
ity(%
)
0
20
40
60
80
100
Fig. 3. Dengue NS3 salt inhibition profile. Relative activities determinedby measuring the initial velocity of hydrolysis of the peptide Z–RR–MCAby NS3 in the presence of NaOAc (h), NaCl (d), Na2SO4 (s), sodiumcitrate (,) and CaCl2 (j). The assay conditions were as described inMaterials and methods.
Table 4Kinetic parameters for hydrolysis of series of peptides derived fromBz–XR–MCA, in which X are Arg or non-natural basic amino acids(see the amino acid structures in Fig. 1) and Bz is benzoyl group
Substrates X CF40glyNS3pro
No. kcat (s�1) Km (lM) kcatt/Km (mM s)�1
31 Arg 0.109 247 0.4432 Amf 0.013 26 0.5033 Gnf 0.035 490 0.0734 Iaf 0.022 354 0.0635 Pya 0.002 43 0.0536 Ama 0.172 298 0.5837 Ppa 0.014 216 0.0738 Aca 0.013 27 0.48
Hydrolysis conditions of the kinetic parameter determinations were sim-ilar as described in Table 1. The standard errors were less than 5% for alldeterminations.
I.E. Gouvea et al. / Archives of Biochemistry and Biophysics 457 (2007) 187–196 193
(Aca—peptide 38) were hydrolyzed with the highest kcat/Km
values in this series, primarily as a result of a low Km. Thepeptide 36 that has trans-4-aminomethylcyclohexyl-alanine(Ama) is hydrolyzed with the highest kcat value in the series.It is noteworthy that the peptide containing 3-pyridyl-alanine (Pya—peptide 35) was poorly hydrolyzed but theobserved Kmvalue was similar to those obtained for pep-tides 32 and 38. Peptide 33 containing 4-guanidine-phenyl-alanine (Gnf ) is poorly hydrolyzed with a high Km value incomparison to the other peptides in the series although theside chain of Gnf has the guanidine function. Peptides 34(Iaf-4-aminomethyl-N-isopropyl–phenylalanine) and 37(Ppa—4-piperidinyl-alanine) were poorly hydrolyzed alsowith high Km values.
Effects of pH and salts on the protease activity
We determined the pH-profile for the enzyme catalyzedhydrolysis of peptides 1–7 (Table 1) and of Z-RR-MCA.All presented a similar shape profile to those obtained forthe hydrolysis of the substrates Abz-SAAQRR-GRIGRNQ-EDDnp (peptide 6—NS3int) or Abz-FAAGRKSLTLNLQ-EDDnp (peptide 3—NS3/NS4A), asshown in Fig. 2. The pK1and pK2 for all of the obtainedprofiles are presented in Table 5. No difference wasobserved for the pK1 values, whereas the pK2 values ofpeptides 1–3 were approximately one pK unit lower thatthose of peptides 4–6. These two sets of peptides differin their basic amino acid composition; peptides 1–3 haveLys while peptides 4–6 have only Arg. This difference inthe pK2 values could be related to deprotonation of thee-NH2-group of the Lys side chain. The pKa of the freeamino acid is 10, but in peptides 1–3 Lys is flanked byother basic residues with electrostatic repulsion potentiallyreducing its pK [24]. A pK2 value of 10 for the short pep-tide Z–RR–MCA pH-profile hydrolysis gives support tothis interpretation.
The anion-dependent activity of CF40glyNS3pro in thepresence of citrate, sulfate, phosphate, acetate, and chlo-ride in their sodium form are shown in Fig. 3. All the
anions inhibited the enzyme and, interestingly, the decreasein activity by salts followed the Hofmeister series: cit-rate > SO2�
4 > acetate� Cl�. CF40glyNS3pro was alsostrongly inhibited by CaCl2 to an extent similar to citrate.

Fig. 4. Colour representations of the active site region of Dengue NS3protease with the catalytic triad shown in pink. Enzyme subsites arelabelled S5–S02.
194 I.E. Gouvea et al. / Archives of Biochemistry and Biophysics 457 (2007) 187–196
Modeling of Abz–AKRRSQ–EDDnp interaction with NS3
subsites
The substrate Abz–AKRRSQ–EDDnp (peptide 22)which presented the highest kcat/Km value was modeledinto the Dengue 2 NS3 binding site occupying the S5–S02subsites (Fig. 4).The substrate occupies a shallow solventexposed active site, with the exception of the S1 pocketwhich is sufficiently deep to almost completely concealthe P1 arginine side-chain from solvent. The P2 arginineside-chain docks in an orientation directed towardsAsn152 in the shallow S2 pocket. The P3 lysine side chainmakes hydrophobic contacts through the b- and c-methyl-enes to Val155 before terminating in solvent. The S4–S5
region is hydrophobic. The P01 serine side-chain docks wellinto the small S1 0 pocket. The subsite pockets in theS3 0–S6 0 region are not well defined, and because of thisuncertainty, no constraints could be used in this region.Consequently, the substrate docked in multiple orienta-tions with one example shown in Fig. 4 where the dinitro-phenyl group is occupying a small pocket bounded byAla99, Glu103, Phe130, Pro132, and Thr134.
Discussion
The hydrolytic activities of the proteases from denguestrains NGC and BR/01-EN on FRET peptides corre-sponding to endogenous cleavage sites of the viral polypro-tein have similar specificity, which is expected due to theirhigh degree of similarity, although NGC enzyme is a moreefficient peptidase. The seven synthetic peptides (Table 1)had different distributions of basic residues, yet were selec-tive for the dengue protease over hfurin, except for thesequence derived from the internal core segment (Cint).Although cleavages of the dengue polyprotein by NS2B/NS3 and furin occur in different host cell compartments,the amino acid sequences of the cleavage sites are alsoselective for each of these enzymes. This result contraststo the high hydrolytic activity of West Nile virus NS3peptidase on furin substrates [10].
The kcat/Kmvalues associated with peptides 1–4 (Table1) are 2–10 times higher than the corresponding reported
chromogenic peptidyl-pNa substrates containing the samepeptide sequences [8]. These differences can be attributedessentially to lower Km values for the hydrolysis of FRETpeptide substrates, and this is clearly related to the occu-pancy of both prime and non-prime enzyme subsites [8]by these substrates. The earlier reported dengue 2 proteaseconstruct NS2B–NS3pro [25] hydrolyzed dansyl-labeledfluorescent peptides corresponding to polyprotein cleavagesites [26] but with significantly lower kcat/Kmvalues than weobtained with the CF40glyNS3pro enzyme construct. Themain differences are in Km values that are likely the resultof differences in the structure of the dengue 2 proteaserecombinants, since the peptide sequences of the assayedsubstrates are very similar.
The S1 subsite of CF40glyNS3pro has a restricted pref-erence for basic residues as shown by the cleavage sites inthe FRET peptides in Tables 1 and 2, which is in accordwith the reported hydrolysis of peptides derived from den-gue 2 polyprotein sequences [4,8,25] and also with the anal-ysis of positional scanning synthetic combinatorial libraries[13].
The S2 subsite of the enzyme is less selective, but alsohas a marked preference for basic amino acids, althoughdengue NS2B/NS3 protease hydrolyses peptides with Glnin this position (peptide 2 of this study) or peptides con-taining Thr, as shown by synthetic combinatorial libraries[12,13]. However, the efficiency of hydrolysis of peptidescontaining basic or non-basic residues at the P2 positionseems to depend on the nature of the amino acids in otherpositions, as demonstrated by the resistance to hydrolysisof Abz–EPEKQRTPQDNQQ–EDDnp (peptide 7 in Table1) and the susceptibility to hydrolysis of Abz–EVKKQ-RAGVLWDQ–EDDnp (peptide 2 in Table 1).
Peptide 5 was hydrolyzed with the lowest Km value andthis can be related to the presence of four Arg in positionsP1 to P4. This is in accordance with the low Ki values report-ed for inhibition of dengue virus NS2B/NS3 protease bysynthetic peptides containing three consecutive basic resi-dues, that were designed based on the products of hydroly-sis of the dengue polyprotein substrate [27]. It is noteworthythat peptide 1 containing three basic residues at positionsP1–P3 was hydrolyzed with the highest Km value in the ser-ies. This may simply be a result of the basic amino acid com-position of substrates in their P1–P3positions, or could bedue to the differences in prime site residues that influencethe enzyme-substrate interaction. The role of amino acidsin substrate prime site positions is observed by comparingpeptides 2 and 7. These substrates have the same aminoacids at positions P3–P2–P1, but peptide 2 is significantlyhydrolyzed, whereas peptide 7 is resistant.
We also explored the extended binding site ofCF40glyNS3pro from P6 to P06 by substituting amino acidsof the worst substrate (peptide 3) into corresponding posi-tions in the best substrate found in Table 1 (peptide 6). Sig-nificant variations in the Km and kcat values were observedfor the hydrolysis of the resulting peptides, but for themajority they compensated each other resulting in only

I.E. Gouvea et al. / Archives of Biochemistry and Biophysics 457 (2007) 187–196 195
small variations in kcat/Km. The only exception was thesubstitution of Arg by Lys at P1 which resulted in a 7-folddecrease in the kcat value, confirming the high preference ofthe enzyme for Arg in this position. Molecular models ofsubstrate binding with the NS3 dengue protease, in theabsence of the NS2B activating co-factor, suggested thatthe critical enzyme–substrate interactions were restrictedto the four residues spanning P2 to P02 [11]. However, thedata presented in Table 2 show that the substrate bindingsite in CF40glyNS3pro might also involve residues P6
and P06. In accordance with this view, CF40glyNS3prowas reported to be more effective on the hydrolysis of thesubstrate Boc–GRR–MCA than on the hydrolysis of N-a-benzoyl-L-Arg-p-nitroanilide (BAPNA) [7] that containsonly a P1 side chain. The crystal structure of dengue virusNS3pro without the NS2B activator in complex with aBowman–Birk inhibitor was reported [11] and a uniquebifurcated S1 pocket detected. Several non-substrate inhib-itors were synthesized containing biguanidine structuresthat potentially mimic this interaction [28], but they arepoor inhibitors with Ki values higher than 40 lM. Thesubstrate Abz-SAAQRRGRIGRNQ-EDDnp (peptide 6)and some of its analogues (peptides 9, 11, and 13–16) werehydrolyzed with kcat/Km 2–3 times higher that the bestinternally quenched peptides recently described [14]. Thissignificant difference between the susceptibility of our sub-strates could be related to the nature of the quencher groupused. Shiryaev et al. [14] used nitro-tyrosine, with is depro-tonated at pH 9 and the reported specificity of dengue NS3protease could have significant interference of the negative-ly charged side chain of nitro-tyrosine that was notobserved with our EDDnp that has no charge at any pH.
We explored in more detail the susceptibility to hydroly-sis of FRET peptides containing two or three basic aminoacids having most of the combinations of Arg and Lys intandem sequence (Table 3). All the combinations of Argand Lys in the substrates presented in Table 3 resulted insubstrates hydrolyzed with Km values systematically higherthan those observed with the hydrolysis of the peptidesderived from the polyprotein. This further indicates that,in addition to interactions between basic residues of thesubstrates with enzyme subsites S1–S3, the affinities of sub-strates and possibly inhibitors require other interactionsthan in these positions, as already suggested above.
All the peptides containing Lys in P1 resulted in sub-strates that hydrolyzed with lower kcat, however if the pep-tides with Lys in P1 had Arg at P2 they were hydrolyzedwith the lowest Km (peptides 19, 24, and 27 in Table 3).The best combination of basic amino acid was obtainedwith Abz–AKRRSQ–EDDnp (peptide 22) that was hydro-lyzed with the highest kcat value in this series.
It is noteworthy that all of the substrates containingthree basic residues were always hydrolyzed after the lastC-terminal basic amino acid, indicating that Arg or Lys isnot accepted at the S01 subsite. This view is supported bythe resistance to hydrolysis of CF40glyNS3pro of the pep-tides Abz–AGRRPAQ–EDDnp and Abz–AKRRRPQ–
EDDnp, and this occurs even with the latter peptide thatcontains the sequence KRR also present in the best sub-strate (peptide 22). The peptides Abz–AGRRPAQ–EDDnpand Abz–AKRRRPQ–EDDnp significantly inhibited theenzyme (Kivalues are shown in Table 3), therefore it is pos-sible that the last Arg of these peptides fits into the S1 sub-site of the enzyme but the Arg–Pro bond is resistant tohydrolysis. The reported cleavages of peptides containingArg at the P01 site in the positional scanning synthetic com-binatorial libraries [13] contrast with this observation. Asthe verification of the cleavage sites in this kind of libraryis not done, the cleavages in these two libraries probablyoccurred after the Arg–Arg and Arg–Lys pair of basic ami-no acids and not between them. The libraries designed toput Arg or Lys in the P01 position in fact put them at P1 posi-tion, and this occurs due to a limitation in the analysis of thehydrolysis of these libraries since the cleavage sites of theirpeptides cannot be checked.
The peptide series Bz–X–Arg–MCA, in which X is a non-natural basic amino acid with an amine or guanidine sub-stituents bound to an aliphatic or aromatic group, werehydrolyzed by CF40glyNS3pro with kcat/Km values lowerthan for Bz–Arg–Arg–MCA (peptide 32). The exceptionswere peptides containing a primary amine substituent inposition 4 of the phenyl group (Amf in peptide 32) or ali-phatic cyclohexyl group (Ama, peptide 36 and Aca, peptide38). It is noteworthy from the results presented in Tables 1–3 that the guanidine substituent of Arg fits better into the S2
subsite of CF40glyNS3pro than the primary amine of Lys.However, the inverse seems to occur when the basic substi-tuent is attached to a aromatic or aliphatic ring. Anotherinteresting result is the relatively low Km value for thehydrolysis of the peptide containing 3-pyridyl-alanine(Pya, peptide 35), in which the amino function is deproto-nated at pH 9. All these observations are likely to be impor-tant for future design of dengue protease inhibitors.
The pH-profiles of hydrolysis obtained with all FRETpeptides of Table 1 present their optimum value aroundpH 9 which did not change even after modifying the saltcomposition of the buffers and by addition of differentdetergents (data not shown). Abz and EDDnp groupsseem also not be involved in the determination of theoptimum pH since Z–RR–MCA hydrolysis pH-profilewas similar to those of the FRET peptides. The peptideacetyl–SAAQRRGRIGRN–amide was synthesized with-out the fluorescent donor–acceptor groups and its pH-profile hydrolysis was similar to all the assayedsubstrates (data not shown). The high activity ofCF40glyNS3pro at alkaline pH can not be the physiolog-ical environment of the enzyme inside the host cell andthe observed behavior of CF40glyNS3pro could not rep-resent the real function of the native protease. It is pos-sible that NS3 requires further interactions with anunknown activator. A similar situation was describedfor human kallikrein 6 that presented optimum activityat pH 9.0 but in the presence of glycosaminoglycan orkosmotropic salts, the pH shifted to 7.5 [29].

196 I.E. Gouvea et al. / Archives of Biochemistry and Biophysics 457 (2007) 187–196
The large inhibitory effect of Ca2+ on protease activitycompared with the other salts is noteworthy. This resultsuggests that Ca2+ may interact with CF40glyNS3promodifying its structure and impairing its peptidase activity.Ca2+ may, therefore, play a significant role in the control ofvirus replication in the host cell. The susceptibility ofCF40glyNS3pro activity to kosmotropic salts such as cit-rate and sulfate is a further indication that the active con-formation of this enzyme is likely to be sensitive to itsphysiological environment. The model of Abz-AKRRSQ-EDDnp docked with the NS3 protease is consistent withthe kinetic parameters of hydrolysis of this peptide, whichis the best substrate among all the examined peptides,although the enzyme we used contained the activating cen-tral 40 residue, relatively hydrophilic domain of NS2B(CF40) that is necessary for a high catalytic activity.
In conclusion, CF40glyNS3pro may have an extendedbinding site and that the affinity of substrates and possiblyinhibitors depends on a number of interactions that extendat least from positions P6 to P06. One interpretation of thisinformation is that the substrate extends outside of the cat-alytic site and may fold back onto, and interact with, resi-dues on the exterior surface of the enzyme-co-factorconstruct. Finally, our results have also indicated that theincorporation of basic non-natural amino acids in shortpeptide sequences affected the affinity and catalytic efficien-cy. Further dissection of such effects may enable use ofthese and other unnatural amino acids to develop efficientinhibitors for this viral enzyme.
Acknowledgment
This work was supported by the Brazillian researchagencies Fundacao de Amparo a Pesquisa do Estado deSao Paulo (FAPESP) and Conselho Nacional de Desen-volvimento Cientıfico e Tecnologico (CNPq) and theNational Health & Medical Research Council (NHMRC)and Australian Research Council (ARC) of Australia.We thank Martin Stoermer for performing the peptidedocking study.
References
[1] W.J. McBride, H. Bielefeldt-Ohmann, Microbes Infect. 2 (2000)1041–1050.
[2] T. Srichaikul, S. Nimmannitya, Baillieres Best Pract. Res. Clin.Haematol. 13 (2000) 261–276.
[3] J.R. Stephenson, Bull. World Health Organ. 83 (2005) 308–314.[4] T.J. Chambers, R.C. Weir, A. Grakoui, D.W. McCourt, J.F. Bazan,
R.J. Fletterick, C.M. Rice, Proc. Natl. Acad. Sci. USA 87 (1990)8898–8902.
[5] P. Keelapang, R. Sriburi, S. Supasa, N. Panyadee, A. Songjaeng, A.Jairungsri, C. Puttikhunt, W. Kasinrerk, P. Malasit, N. Sittisombut,J. Virol. 78 (2004) 2367–2381.
[6] S. Clum, K.E. Ebner, R. Padmanabhan, J. Biol. Chem. 272 (1997)30715–30723.
[7] R. Yusof, S. Clum, M. Wetzel, H.M. Murthy, R. Padmanabhan, J.Biol. Chem. 275 (2000) 9963–9969.
[8] D. Leung, K. Schroder, H. White, N.X. Fang, M.J. Stoermer, G.Abbenante, J.L. Martin, P.R. Young, D.P. Fairlie, J. Biol. Chem. 276(2001) 45762–45771.
[9] N.J. Butkiewicz, M. Wendel, R. Zhang, R. Jubin, J. Pichardo, E.B.Smith, A.M. Hart, R. Ingram, J. Durkin, P.W. Mui, M.G. Murray,L. Ramanathan, B. Dasmahapatra, Virology 225 (1996) 328–338.
[10] S.A. Shiryaev, B.I. Ratnikov, A.V. Chekanov, S. Sikora, D.V.Rozanov, A. Godzik, J. Wang, J.W. Smith, Z. Huang, I. Lindberg,M.A. Samuel, M.S. Diamond, A.Y. Strongin, Biochemical J. 393(2006) 503–511.
[11] H.M. Murthy, K. Judge, L. DeLucas, R. Padmanabhan, J. Mol. Biol.301 (2000) 759–767.
[12] R.I. Brinkworth, D.P. Fairlie, D. Leung, P.R. Young, J. Gen. Virol.80 (1999) 1167–1177.
[13] J. Li, S.P. Lim, D. Beer, V. Patel, D. Wen, C. Tumanut, D.C. Tully,J. A Williams, J. Jiricek, J.P. Priestle, J.L. Harris, S.G. Vasudevan,J. Biol. Chem. 280 (2005) 28766–28774.
[14] P. Niyomrattanakit, S. Yahorava, I. Mutule, F. Mutulis, R. Petro-vska, P. Prusis, G. Katzenmeier, J.E. Wikberg, Biochem. J. 397 (2006)203–211.
[15] A. Cameron, J. Appel, R.A. Houghten, I. Lindberg, J. Biol. Chem.275 (2000) 36741–36749.
[16] F. Jean, K. Stella, L. Thomas, G. Liu, Y. Xiang, A.J. Reason, G.Thomas, Proc. Natl. Acad. Sci. USA 95 (1998) 7293–7298.
[17] I.Y. Hirata, M.H.S. Cezari, C. Nakaie, P. Boschcov, A.S. Ito, M.A.Juliano, L. Juliano, Lett. Peptide Sci. 1 (1994) 299–308.
[18] R.L. Melo, R.C. Barbosa Pozzo, L.C. Alves, E. Perissutti, G.Caliendo, V. Santagada, L. Juliano, M.A. Juliano, Biochim. Biophys.Acta 1547 (2001) 82–94.
[19] Y. Liu, W. Kati, C.M. Chen, R. Tripathi, A. Molla, W. Kohlbrenner,Anal. Biochem. 267 (1999) 331–335.
[20] G. Salvessen, H. Nagase, R.J. Beynon, J.S. Bond (Eds.), Oxford,England:IRL Press 1989 p. 83-104.
[21] J.D.A. Tyndall, T. Nall, D.P. Fairlie, Chem. Rev. 105 (2005)973–999.
[22] L. Zhang, P.M. Mohan, R. Padmanabhan, J. Virol. 66 (1992)7549–7554.
[23] A. Cahour, B. Falgout, C.J. Lai, J. Virol. 66 (1992) 1535–1542.[24] L. Juliano, A.M.C. Paiva, Biochemistry 13 (1974) 2445–2450.[25] R. Khumthong, C. Angsuthanasombat, S. Panyim, G. Katzenmeier,
J. Biochem. Mol. Biol. 35 (2002) 206–212.[26] R. Khumthong, P. Niyomrattanakit, S. Chanprapaph, C. Angsu-
thanasombat, S. Panyim, G. Katzenmeier, Protein Pept. Lett. 10(2003) 19–26.
[27] S. Chanprapaph, P. Saparpakorn, C. Sangma, P. Niyomrattanakit, S.Hannongbua, C. Angsuthanasombat, G. Katzenmeier, Biochem.Biophys. Res. Commun. 330 (2005) 1237–1246.
[28] V.K. Ganesh, N. Muller, K. Judge, C.H. Luan, R. Padmanabhan,K.H. Murthy, Bioorg. Med. Chem. 13 (2005) 257–264.
[29] P.F. Angelo, A.R. Lima, F.M. Alves, S.I. Blaber, I.A. Scarisbrick,M. Blaber, L. Juliano, M.A. Juliano, J. Biol. Chem. 281 (2006)3116–3126.





























