Embed Size (px)
Citation preview

Neuron, Vol. 42, 163–172, April 8, 2004, Copyright 2004 by Cell Press
Subcortical Discrimination of Unperceived Objectsduring Binocular Rivalry
When the perceived object is behaviorally relevant andrequires a rapid adaptive response, this extended pro-cessing time may put the animal at a survival disadvan-
Brian N. Pasley,1,2 Linda C. Mayes,1
and Robert T. Schultz1,*1Developmental Neuroimaging Laboratory
tage. However, amygdala neurons selective for complexChild Study Centerobjects have been found in monkeys with response la-Yale Universitytencies as short as 60 ms, which is roughly twice as fastNew Haven, Connecticut 06520as IT response latencies, indicating the presence of a2 Helen Wills Neuroscience Institutemore rapid alternate visual route in primates (NakamuraUniversity of California, Berkeleyet al., 1992). Several sources of data, including anatomi-Berkeley, California 94720cal (Linke et al., 1999; Romanski et al., 1997), lesion(Miller et al., 1980), and functional brain imaging (Morriset al., 1999, 2001; Vuilleumier et al., 2003) studies, haveSummarybeen used to identify a series of subcortical visual struc-tures that plausibly comprise a subcortical pathway ter-Rapid identification of behaviorally relevant objects isminating in the amygdala. This pathway, proceedingimportant for survival. In humans, the neural com-from the retina to the superior colliculus (SC) to posteriorputations for visually discriminating complex objectsnuclei of the thalamus and onto the amygdala, bypassesinvolve inferior temporal cortex (IT). However, lessdetailed cortical processing and is thought to providedetailed but faster form processing may also occur inthe amygdala with lower-resolution but more rapidlya phylogenetically older subcortical visual systemprocessed visual input (LeDoux, 1996). Some accountsthat terminates in the amygdala. We used binocularemphasize the bias of this system for stimuli that arerivalry to present stimuli without conscious aware-informative about potential dangers (e.g., fear-relevantness, thereby eliminating the IT object representationstimuli) (LeDoux, 1996; Morris et al., 1999, 2001).and isolating subcortical visual input to the amygdala.
Although the anatomical connections underlying thisFunctional magnetic resonance imaging revealedsubcortical visual system clearly exist in nonhuman pri-significant brain activation in the left amygdala butmates (Benevento and Fallon, 1975; Jones and Burton,not in object-selective IT in response to unperceived1976; Schiller and Malpeli, 1977; Stepniewska et al.,fearful faces compared to unperceived nonface ob-2000), there is not yet completely convincing empiricaljects. These findings indicate that, for certain behav-data to argue for the functional significance of this path-iorally relevant stimuli, a high-level cortical represen-way. One piece of evidence for its functional significancetation in IT is not required for object discrimination incomes from the study of “blindsight.” In blindsight, athe amygdala.lesion in primary visual cortex (V1) prevents consciousvision in the corresponding portion of the visual fieldIntroductionand is believed to disrupt the ventral object recognitionpathway extending from V1 through IT. Nevertheless,When presented with a variety of visual objects in theblindsight patients exhibit residual abilities to detect andenvironment, the brain must be able to quickly and reli-localize visual stimuli (Weiskrantz, 1997), suggesting theably detect those that are behaviorally relevant (e.g.,activation of an alternative visual pathway. In a func-those that signal potential dangers). The amygdala istional magnetic resonance imaging (fMRI) study, Morrisbelieved to be a critical component of this perceptualand colleagues (2001) demonstrated amygdala activa-
process and has been implicated in many forms oftion to emotionally expressive faces in a blindsight pa-
affective processing, especially fear-related (LeDoux,tient who did not consciously perceive the stimuli. How-
1996). There are both cortical and subcortical amygdalar ever, because the subcortical visual stream is believedinputs (LeDoux, 1996), but the relative and perhaps to possess relatively primitive pattern vision mecha-unique contribution of each to visually driven, affective nisms, several investigators have argued that highly pro-responses in the amygdala is not yet well understood. cessed input from IT may be required to support visual
In nonhuman primates, the amygdala receives input discriminatory responses in the amygdala (Pessoa etfrom IT (Stefanacci and Amaral, 2002), the last purely al., 2002a, 2002b; Rolls, 1999, 2000). In fact, in a studyvisual area in the ventral object recognition pathway by Goebel and colleagues (2001), this same blindsight(Gallant, 2000), but not from earlier cortical visual areas patient exhibited increased activity in IT when presented(Iwai and Yukie, 1987; Webster et al., 1991). Although with images of complex objects in his blind visual field,input from IT may provide the amygdala with a hierarchi- suggesting that IT inputs to the amygdala could havecally processed and detailed object representation, this driven the observed amygdala response.processing strategy necessarily sacrifices speed for en- Visual backward masking is another phenomenon thathanced detail. Visual responses in monkey IT typically has been used to disrupt processing in IT (Morris etoccur between 100 and 140 milliseconds (ms) after stim- al., 1998; Whalen et al., 1998) in order to isolate theulus presentation (Rolls, 1999) (in humans, IT latencies subcortical pathway. These studies briefly presentedare a bit longer, i.e., 150–200 ms; Allison et al., 1999). target images of faces with emotional or neutral expres-
sions (�30 ms) followed immediately by a sustainedmasking stimulus that rendered the target image invisi-*Correspondence: [email protected]

Neuron164
ble. In macaques, the mask is known to reduce the (Blake and Logothetis, 2002). Some evidence in humans,amount of information regarding the target image by however, suggests that rivalry may be mediated by ainterrupting or overriding the firing of neurons in IT process in V1 referred to as interocular competition(Kovacs et al., 1995; Rolls et al., 1994). Importantly, the (Tong and Engel, 2001). Under this particular model,amygdala receives its major visual input from IT, but not lateral inhibition among monocular neurons in V1 isfrom earlier visual areas (Iwai and Yukie, 1987; Ste- believed to suppress input from one eye, effectivelyfanacci and Amaral, 2000; Webster et al., 1991). Because blocking further visual processing for the suppressedmasking degrades the information available in this late stimulus while the dominant monocular stimulus con-stage of cortical visual processing, it has been argued tinues on to higher-level visual areas and eventuallythat the differential amygdala response is derived exclu- reaches visual awareness (Blake, 1989; Tong, 2001). Al-sively from subcortical visual input (Morris et al., 1999, though there is some evidence that neurons in V1 and2001). Interpretation of the origin of this visual input to V2 still respond to suppressed stimuli (Leopold and Lo-the amygdala is difficult, however, since neurons in IT gothetis, 1996), data from both monkey neurophysiologyretain substantial information about the target image (Sheinberg and Logothetis, 1997) and human fMRI (Tongdespite the masking procedure (Rolls et al., 1999). In et al., 1998) converge to suggest that suppressed visualfact, under experimental conditions where the object is information is blocked from reaching IT where neuralnot “seen,” increased target-related neural activity in responses reflect the perceptually dominant and not theIT has been observed consistently across a variety of suppressed stimulus. These physiological data agreefunctional imaging (Bar and Biederman, 1999; Dehaene well with behavioral studies exploring the effects of ri-et al., 2001; Grill-Spector et al., 1999) and single-unit valry suppression on behavioral priming. These studies(Kovacs et al., 1995; Rolls and Tovee, 1994) experiments demonstrate that rivalry suppression, unlike backwardthat have employed backward masking. masking, abolishes behavioral priming for suppressed
Furthermore, IT activity related to the masked target words (Zimba and Blake, 1983) and pictorial stimuliis subject to repetition suppression, a phenomenon ob- (Cave et al., 1998), providing additional support for theserved in positron emission tomography (PET) and fMRI notion that suppression blocks visual information fromstudies in which the hemodynamic response is reduced reaching the late stages of the ventral visual pathway.for repeated compared to novel stimuli (Dehaene et al., Perceptual suppression during binocular rivalry there-2001; Henson and Rugg, 2003). Such selective modifica- fore appears well suited to test the subcortical visualtion is evidence for the sustained presence of a neural pathway to the amygdala independently of input from IT.representation that is sufficiently intact to differentiate Based on this evidence, we used a binocular rivalrythe repeated from the unrepeated stimulus. A related paradigm in which the image of either a face with aand consistent finding concerns behavioral priming of fearful expression or a chair with no apparent emotionalthe masked stimulus (Bar and Biederman, 1998; Buck- value was presented under complete perceptual sup-ner et al., 1998; Koutstaal et al., 2001). Priming is a pression in order to observe differential neural responsesphenomenon that occurs when a prior presentation of that occur in isolation from cortical input. While chairsa stimulus improves or “primes” performance during are neutral stimuli with no obvious emotional signifi-subsequent presentations. Like repetition suppression, cance, fMRI studies have found with notable consis-behavioral priming appears to depend on the presence tency that fearful faces activate the amygdala, particu-of a functional neural representation formed late in the larly on the left (Calder et al., 2001). Thus, the currentventral visual stream (Bar and Biederman, 1998; Buckner study was designed to interrogate subcortical visualet al., 1998; Koutstaal et al., 2001), suggesting that the regions for increased neural activity to suppressed fear-high-level representation of the masked stimulus per- ful faces as compared to suppressed neutral chairs. Wesists despite the masking procedure. Thus, it remains reasoned that a significant differential response inunclear whether or not the discrimination observed in
the amygdala, but not in object-selective IT, would pro-the amygdala during masking and blindsight occurs as
vide strong evidence that rudimentary discriminationa consequence of a degraded but intact neural repre-
of certain complex visual patterns does not require asentation in IT or, alternatively, originates exclusivelyhigh-level cortical representation but instead may befrom a subcortical source of input. Consequently, themediated by a subcortical visual pathway that canability of the subcortical visual pathway to discriminateoperate independently and in parallel to cortical visualcertain behaviorally relevant stimuli in isolation from theprocessing.IT object representation remains controversial (Pessoa
et al., 2002a, 2002b; Rolls, 1999, 2000).ResultsTo address this controversy, we used fMRI to measure
brain activity during a binocular rivalry paradigm as aIn binocular rivalry, perceptual dominance is promotedway to probe the ability of the amygdala to discriminateby “stimulus strength,” as defined by characteristicscomplex objects in isolation from the IT neural represen-such as high luminance and high contrast, transienttation. During binocular rivalry, each eye is presentedmotion, and complexity (Blake, 2001). We therefore pre-with a different, incompatible image. Rather than experi-sented to one eye a complex image of a house thatence fusion of these two distinct images, the observermoved back and forth sharply in order to constrain con-experiences alternating perceptual dominance of onescious perception to this image. During the movementimage or the other. Thus, while one image is perceptuallyof the house, a target image of a fearful face or a neutraldominant, the other is completely suppressed, or unper-chair gradually faded into view of the other eye, re-ceived. The neural mechanisms underlying rivalry are
poorly understood and are a subject of current debate maining in view for approximately 1.5 s before fading

Subcortical Processing of Unperceived Stimuli165
Figure 1. Binocular Rivalry Stimulus Presen-tation Procedure for a Suppressed Face Trial
Participants viewed blocks of four consecu-tive trials lasting 10 s and were instructed toindicate by button press if at any point theyperceived anything besides a house, no mat-ter how fleeting, in order to verify the success-ful suppression of the target image. Acrossall participants, unsuccessful rivalry suppres-sion trials occurred in 2% of these blocks,which were subsequently excluded from fur-ther analysis.
out again. This presentation scheme (Figure 1) main- not reach these higher-level areas of visual cortex. How-ever, because the exact location of IT object-selectivetained the perceptual suppression of the target image
during the entire trial. In addition to presentations of areas can vary somewhat between individuals and maynot be accurately reflected in a group average, we alsosuppressed chairs and suppressed fearful faces, we
also included a third condition in which a moving house functionally defined face and chair ROIs on each individ-ual’s data. This provided a more rigorous test to deter-was presented to both eyes, in order to compare re-
sponses to this nonrivalry condition. These three condi- mine if the discrimination in the amygdala was paralleledby IT activation that may have in turn driven the observedtions were indistinguishable to the participant, appearing
as a moving house in every case. amygdala activation. Mean signal changes within theseROIs during the rivalry experiment were used to test forTo assess the presence of discrimination of suppressed
objects, we compared activity during suppressed face condition-specific activations of face- and chair-selec-tive IT. An ANOVA contrasting the peak mean signalpresentations to activity during suppressed chair pre-
sentations within several a priori regions of interest change in face- and chair-selective IT cortex revealedno significant main effects for suppressed image type(ROIs). These ROIs included three nodes of the subcorti-
cal visual pathway—the SC, the thalamus, and the [suppressed face/chair, F(1,44) � 0.70, p � .41] or ROI[F(1,44) � 0.85, p � .36], nor was there a significantamygdala—as well as face- and chair-selective IT cor-
tex. To identify these category-selective areas of IT image type by ROI interaction [F(1,44) � 0.71, p � .41].Thus, as shown in Figures 3C and 3D, during rivalrous(Epstein and Kanwisher, 1998; Ishai et al., 1999; Kan-
wisher et al., 1997), we conducted an independent ob- presentations, the average response function in IT fol-lows the dominant house stimulus, with no significantject localizer experiment during the same scan session,
in which participants viewed images of fearful and neu- differences between the three conditions [repeatedmeasures ANOVA, face-selective IT: F(2,20) � 0.759,tral faces or chairs. This allowed us to functionally define
the areas of IT involved in encoding high-level face and p � .48; chair-selective IT: F(2,22) � 1.48, p � .25].However, when the amygdala ROI is included in thischair neural representations. These face and chair ROIs
overlapped with those reported by other groups (Ishai analysis, there are significant effects of image type[F(1,66) � 10.49, p � .002], ROI [F(2,66) � 3.85, p � .03],et al., 1999) and were localized in the lateral and medial
fusiform gyrus for faces and chairs, respectively. and the image type by ROI interaction [F(2,66) � 5.32,p � .008]. This result is accounted for by the significantlyAfter transformation of all 12 individual data sets into
standard stereotaxic space (Talairach and Tournoux, greater suppressed face activation of the left amygdala,compared to suppressed chair, without any significant1988), a voxel-wise random effects analysis of the group
data yielded significant left amygdala activation for a effect of image type on the cortical ROIs.Although we did not observe significant activation insuppressed face versus suppressed chair contrast
[t(11) � 4.063, p � 0.005, corrected for 5 multiple ROI any areas outside of our a priori ROIs (tested at p � .05,corrected), our slice selection primarily covered only thecomparisons; Figure 2]. In addition, inspection of the
individual data sets showed that this effect was present visual cortex and midbrain. We were therefore unableto assess the possibility that other nonvisual corticalin 9 of the 12 participants at a threshold of p � 0.05,
uncorrected. Left-lateralized activation is consistent regions may have exhibited differential responses tosuppressed objects, conceivably providing the amyg-with previous studies finding a bias toward the left amyg-
dala for fear-related stimuli (Calder et al., 2001). dala with visual input through an indirect cortical route.However, prior studies of rivalry have found that activityImportantly, in the group data, the same suppressed
face versus suppressed chair contrast produced no sig- in nonvisual areas (e.g., parietal and prefrontal cortex)is associated with conscious perception rather than thenificant differences in face and chair ROIs within IT cor-
tex (tested at a less stringent threshold of p � 0.01 to unperceived stimulus (Lumer et al., 1998; Lumer andRees, 1999), suggesting that suppressed visual informa-strengthen the claim of a null result), suggesting that
visual information regarding the suppressed objects did tion fails to reach these later processing stages. Simi-
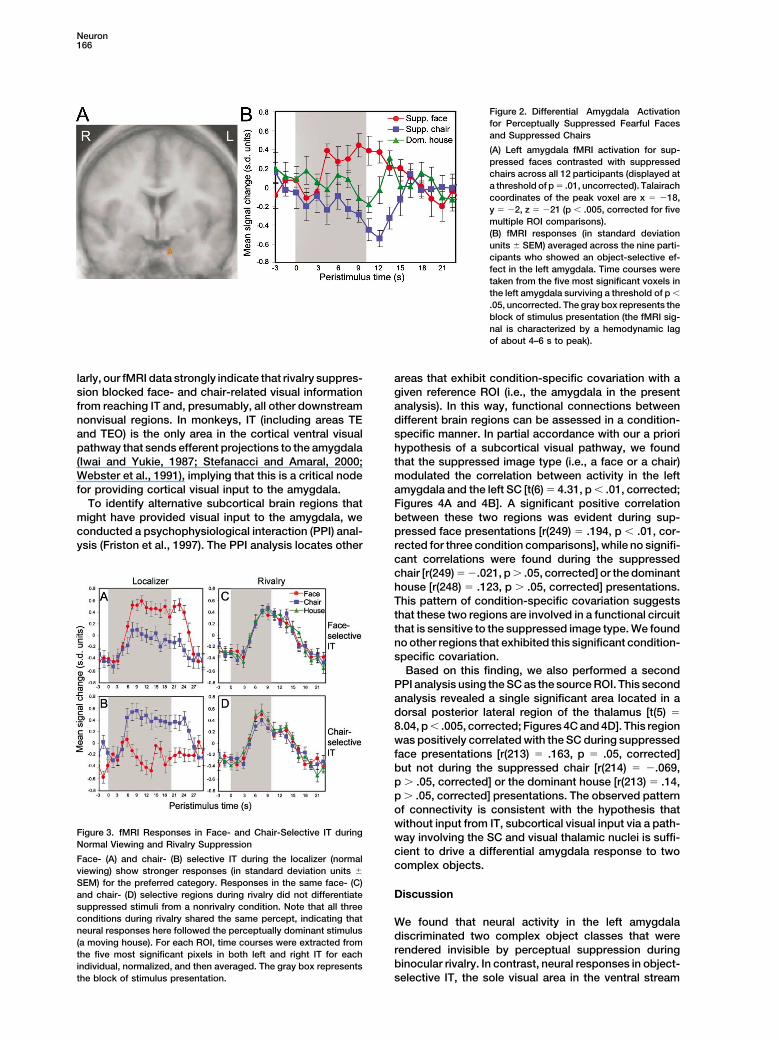
Neuron166
Figure 2. Differential Amygdala Activationfor Perceptually Suppressed Fearful Facesand Suppressed Chairs
(A) Left amygdala fMRI activation for sup-pressed faces contrasted with suppressedchairs across all 12 participants (displayed ata threshold of p � .01, uncorrected). Talairachcoordinates of the peak voxel are x � �18,y � �2, z � �21 (p � .005, corrected for fivemultiple ROI comparisons).(B) fMRI responses (in standard deviationunits � SEM) averaged across the nine parti-cipants who showed an object-selective ef-fect in the left amygdala. Time courses weretaken from the five most significant voxels inthe left amygdala surviving a threshold of p �
.05, uncorrected. The gray box represents theblock of stimulus presentation (the fMRI sig-nal is characterized by a hemodynamic lagof about 4–6 s to peak).
larly, our fMRI data strongly indicate that rivalry suppres- areas that exhibit condition-specific covariation with agiven reference ROI (i.e., the amygdala in the presentsion blocked face- and chair-related visual information
from reaching IT and, presumably, all other downstream analysis). In this way, functional connections betweendifferent brain regions can be assessed in a condition-nonvisual regions. In monkeys, IT (including areas TE
and TEO) is the only area in the cortical ventral visual specific manner. In partial accordance with our a priorihypothesis of a subcortical visual pathway, we foundpathway that sends efferent projections to the amygdala
(Iwai and Yukie, 1987; Stefanacci and Amaral, 2000; that the suppressed image type (i.e., a face or a chair)modulated the correlation between activity in the leftWebster et al., 1991), implying that this is a critical node
for providing cortical visual input to the amygdala. amygdala and the left SC [t(6) � 4.31, p � .01, corrected;Figures 4A and 4B]. A significant positive correlationTo identify alternative subcortical brain regions that
might have provided visual input to the amygdala, we between these two regions was evident during sup-pressed face presentations [r(249) � .194, p � .01, cor-conducted a psychophysiological interaction (PPI) anal-
ysis (Friston et al., 1997). The PPI analysis locates other rected for three condition comparisons], while no signifi-cant correlations were found during the suppressedchair [r(249) � �.021, p � .05, corrected] or the dominanthouse [r(248) � .123, p � .05, corrected] presentations.This pattern of condition-specific covariation suggeststhat these two regions are involved in a functional circuitthat is sensitive to the suppressed image type. We foundno other regions that exhibited this significant condition-specific covariation.
Based on this finding, we also performed a secondPPI analysis using the SC as the source ROI. This secondanalysis revealed a single significant area located in adorsal posterior lateral region of the thalamus [t(5) �8.04, p � .005, corrected; Figures 4C and 4D]. This regionwas positively correlated with the SC during suppressedface presentations [r(213) � .163, p � .05, corrected]but not during the suppressed chair [r(214) � �.069,p � .05, corrected] or the dominant house [r(213) � .14,p � .05, corrected] presentations. The observed patternof connectivity is consistent with the hypothesis thatwithout input from IT, subcortical visual input via a path-
Figure 3. fMRI Responses in Face- and Chair-Selective IT during way involving the SC and visual thalamic nuclei is suffi-Normal Viewing and Rivalry Suppression
cient to drive a differential amygdala response to twoFace- (A) and chair- (B) selective IT during the localizer (normal
complex objects.viewing) show stronger responses (in standard deviation units �
SEM) for the preferred category. Responses in the same face- (C)and chair- (D) selective regions during rivalry did not differentiate Discussionsuppressed stimuli from a nonrivalry condition. Note that all threeconditions during rivalry shared the same percept, indicating that We found that neural activity in the left amygdalaneural responses here followed the perceptually dominant stimulus discriminated two complex object classes that were(a moving house). For each ROI, time courses were extracted from
rendered invisible by perceptual suppression duringthe five most significant pixels in both left and right IT for eachbinocular rivalry. In contrast, neural responses in object-individual, normalized, and then averaged. The gray box represents
the block of stimulus presentation. selective IT, the sole visual area in the ventral stream

Subcortical Processing of Unperceived Stimuli167
Figure 4. ROI Connectivity
(A) A region of the left SC (�4, �29, �3)showed a significant condition-specific co-variation with the left amygdala (p � .01, cor-rected).(B) The scatterplot displays the correlationbetween this left SC ROI and the left amyg-dala ROI, coded by condition. The significantPPI analyses described in the text are a func-tion of a significant difference between thesuppressed face [r(249) � .194, p � .01, cor-rected for three condition comparisons] andthe suppressed chair [r(249) � �.021, p �
.05, corrected] correlations . The correlationduring house presentations was also nonsig-nificant [not displayed; r(248) � .123, p �
.05, corrected].(C) A region of the left dorsal posterior lateralthalamus (�10, �14, 12) showed a significantcondition-specific covariation with the SC(p � .005, corrected).(D) The SC and thalamic region correlatedpositively during suppressed face presenta-tions [r(213) � .163, p � .05, corrected] butwere not significantly correlated during sup-pressed chair [r(214) � �.069, p � .05, cor-rected] or house [not displayed; r(213) � .14,p � .05, corrected] presentations. The fMRIimages are displayed at a threshold of p �
.01, uncorrected.
that projects to the amygdala, were identical between viewed monocularly, is disrupted by a new “flashed”stimulus to the other eye. Kreiman and colleagues, re-rivalrous conditions (suppressed face/chair) and a nonri-
valrous control condition (dominant house). This sug- cording from single cells in the amygdalae of patientsundergoing brain surgery, identified several amygdalagests that IT cortex did not receive suppressed visual
information and thus could not contribute input to the neurons that responded selectively to complex objectsat latencies of around 200 ms, suggesting transmissionamygdala to trigger its differential response to sup-
pressed objects. In addition, we found significant corre- of visual input from IT (where latencies are typically�170 ms; Allison et al., 1999). However, when theselations between different subcortical visual structures
during suppressed face presentations (but not during preferred images were perceptually suppressed by aflashed stimulus (that was ineffective in stimulatingsuppressed chair presentations): (1) the left amygdala
correlated positively with the SC and (2) the SC corre- amygdala activity), the increased firing rate of the amyg-dala neurons returned immediately to the lower baselinelated positively with a dorsal posterior lateral region of
the thalamus. As shown in Figures 4A and 4C, these rate. This finding indicates that perceptual suppressionof the preferred image blocked the transmission of sup-correlations were highly specific to these structures.
Our results support prior studies suggesting that ri- pressed visual input to the amygdala via the corticalIT pathway.valry suppression blocks suppressed visual signals
sometime prior to IT in the ventral processing stream While Kreiman et al.’s study clearly demonstrates theblocking effect of suppression on cortical visual input,(Tong and Engel, 2001; Tong et al., 1998). On the other
hand, our data indicate that information regarding the it is unclear why the suppressed stimuli did not continueto engage the amygdala through the subcortical path-suppressed image successfully reaches the amygdala,
apparently through a subcortical pathway. One possible way described in our current results. There are severalpossibilities that may explain this apparent discrepancy.caveat to this conclusion is that fMRI may lack the sensi-
tivity to detect weak signals in IT during rivalry suppres- First, the single neurons in this study were chosen foranalysis based on their strong selectivity for differentsion and that these weak signals may yet provide the
amygdala with cortical visual input. However, this possi- complex images or object categories. However, thissubsample of amygdala neurons may not overlap with ability seems unlikely in view of past studies demonstra-
ting that fMRI has sufficient sensitivity to detect weak less selective population of neurons driven by relativelylow-resolution subcortical visual inputs. It is thereforeextrastriate signals related to unperceived stimuli during
backward masking (Bar and Biederman, 1998; Dehaene possible that amygdala neurons responsive to subcorti-cal inputs may be a distinct population and, conse-et al., 2001). Moreover, a related study that employed
“flash suppression” (Kreiman et al., 2002) provides more quently, were excluded from Kreiman et al.’s analysisbecause they did not meet selectivity requirements. Indirect evidence that perceptual suppression blocks the
transmission of cortical visual input to the amygdala. addition, this study investigated only a very small sam-ple of amygdala neurons (25 selective neurons wereFlash suppression is closely related to binocular rivalry
and occurs when the perception of a baseline image, analyzed, many of which were in the right hemisphere,

Neuron168
not the left as highlighted by the current study). The dominance in SC neurons may preserve monocularvisual input, permitting subsequent subcortical pro-fMRI blood oxygenation level-dependent (BOLD) signal,
on the other hand, reflects the simultaneous neural cessing of suppressed stimuli. Based on this physiologi-cal evidence and our connectivity analysis, it thereforeactivity of large, distributed populations of neurons
(typically around 107 per voxel). If amygdala neurons seems reasonable that the SC played at least some rolein the form processing that sustained the differentialresponsive to subcortical inputs are fewer than those
responsive to cortical inputs, detection of these rela- amygdala response to suppressed complex objects.In primates, the SC is usually implicated in visual ori-tively uncommon sets of neurons may be easier with
fMRI than with single unit recording. enting responses and directional encoding. Althoughthe visual cortex has superseded it as the primary formAnother possible difference between the current
study and that of Kreiman et al. concerns the neurophys- processing center in the primate brain, the SC still pos-sesses the ability to process simple forms, but onlyiological basis of the BOLD signal commonly measured
in fMRI. Currently under debate is whether the BOLD within a narrow range of spatial frequencies (Lomber,2002; Miller et al., 1980). A critical problem for the hy-signal is more closely linked to the spiking output of
neurons or to energy requirements associated with cu- pothesis of a subcortical pathway is to determine howprimitive pattern vision in the SC might successfullymulative activity at the synapses (Logothetis, 2003). The
amygdala activation that we observed may therefore code and propagate complex form information to othersubcortical structures. Several aspects of our resultshave been related more to increased input to the amyg-
dala rather than to actual neuronal spikes. This interpre- provide indications of what visual information is passedto the amygdala through this putative subcortical sys-tation would be consistent with the absence of single
neuron responses observed by Kreiman et al. How- tem. One potentially important observation is that thedifferential amygdala activity for suppressed objectsever, even if these fMRI results reflect an input rather
than an output process, either possibility entails the was due to both increased signal for faces and de-creased signal for chairs. This bidirectional differentialexistence of an intact visual pathway that successfully
transmits information to the amygdala during perceptual response to suppressed objects would seem to indicatethat information about both the face and the chair suc-suppression. Given the convergent evidence for cortical
blocking during suppression, the increased amygdalar ceeded in reaching the amygdala and consequentlymodulated activity there. In view of this apparent nonse-response to the suppressed face evidently resulted from
visual input whose origin was other than IT. lectivity, the SC is probably not involved in any type of“gating” mechanism (i.e., all or none output) wherebyThis conclusion is also supported by our functional
connectivity analyses. The observed pattern of connec- irrelevant stimuli are completely filtered out from furtherprocessing. Because we did not observe a differentialtivity coincides with the hypothesis of a feedforward
subcortical visual pathway to the amygdala involving fMRI response in the SC to suppressed objects, wepostulate that the processing of each suppressed objectthe SC and visual thalamus (LeDoux, 1996; Morris et al.,
1999, 2001). However, because this connectivity analy- type generated similar neuroenergetic requirementswithin this structure. In this sense, the SC may act assis is based on correlation, we cannot conclusively de-
termine the directionality or the full extent of this path- a general filter for form information, processing eachpattern without regard to behavioral relevance andway. For example, it is possible that the correlation
between the amygdala and SC is representative of a passing on a similar range of visual information for bothsuppressed object types.shared source of cortical input rather than a feedforward
subcortical pathway. In this sense, responses in these If in fact the SC is engaged in such a filtering process,then it is crucial to consider the precise nature andtwo subcortical regions may be modulated by a third
region (possibly V1 or V2, which have been found to level of detail of information that ultimately reaches theamygdala through this filter. In the absence of visualprocess aspects of suppressed stimuli; Leopold and
Logothetis, 1996). However, these cortical areas appear cortex, the SC is sensitive to the low but not high spatialfrequency components of patterns (Lomber, 2002; Millerto have no direct projections to the amygdala (Iwai and
Yukie, 1987; Stefanacci and Amaral, 2000; Webster et et al., 1980; Vuilleumier et al., 2003). While high spatialfrequencies encode individual facial features and fineal., 1991) and are not known to support the discrimina-
tion of complex objects. The SC, on the other hand, is details, low spatial frequencies encode the global con-figural structure of the face (Schyns and Oliva, 1999).an ancient midbrain structure that serves as the primary
object processing structure in avians and amphibians Low spatial frequency information about the coarsestructure of a face is particularly important for face per-and receives direct visual input from the retina in pri-
mates (Sewards and Sewards, 2002). It is therefore well ception, and it is thought that an efficient face detectionmechanism would rely more strongly upon low ratherpositioned to receive direct visual information before
incongruent signals during rivalry have been altered by than high spatial frequencies (Dailey and Cottrell, 1999).Thus, the SC appears to encode a range of spatialbinocular convergence. In addition, there is evidence
that the ocular dominance of neurons in the visually information that is particularly valuable for detecting acomplex face stimulus. This view is consistent withresponsive superficial layer of the SC is stronger for the
contralateral eye and that this dominance becomes even the observation that newborn infants exhibit longer look-ing times for face-like patterns than for other patterns,stronger after a visual cortex lesion (Rosa and Schmid,
1994). Compared to the robustness of suppression in even in the first minutes of life (Goren et al., 1975;Simion et al., 1998). At birth, the visual cortex is function-binocular extrastriate cortex, suppressive mechanisms
in the SC may be weaker for neurons that receive input ally immature (Kraemer and Sjostrom, 1998), and it isthought that this visual preference is instead supportedpredominantly from only one eye. Consequently, ocular

Subcortical Processing of Unperceived Stimuli169
by a subcortical mechanism involving the SC (Morton nized for fast detection of salient stimuli, this hierarchicaland Johnson, 1991). Thus, the positive correlation we strategy would presumably be slower than a template-observed between the amygdala and the SC during pre- matching mechanism but also considerably faster thansentations of suppressed faces may relate to a subcorti- the multistage cortical processing stream. Determiningcal pattern vision mechanism that has been configured, if either of these two mechanisms is employed by thepossibly via evolutionary pressure, to preserve the most subcortical visual pathway is an important line of fu-essential spatial information needed to support rudi- ture work.mentary detection of face-like patterns, although the Our results have demonstrated that differential activ-range of preferred patterns is likely to be quite crude. ity for a behaviorally relevant and a neutral object in the
While the SC appears to encode a range of visual amygdala can occur without input from IT and that thisinformation valuable for face detection, another impor- discrimination appears to be mediated by a subcorticaltant question concerns how the amygdala actually pro- pathway involving the SC and a dorsal posterior lateralcesses this input to achieve the pattern discrimination region within the thalamus. Our method should also pro-that underlies its role in forming responses to salient vide a means to further characterize the effects of rivalrystimuli. Amygdala neurons involved in a system for de- suppression within subcortical visual structures and totecting behaviorally relevant stimuli are believed to rep- identify the range of visual patterns to which this path-resent the conjunction of a stimulus representation with way is responsive in order to understand the analysisits behavioral relevance. This system may perform a principles that support subcortical form discrimination.type of “lookup” operation, searching available memory The computational mechanisms that underlie subcorti-for the incoming stimulus representation and then de- cal pattern vision may then yield insights into both theciding if this stimulus has a previously associated behav- ontogenetic and evolutionary development of vision.ioral value (Rolls, 1999). The incoming, high-level stimu-
Experimental Procedureslus representation provided by IT is unlikely to resemblethe relatively crude representation provided by a sub-
Participantscortical pathway. It is therefore unclear if this system canTwelve healthy participants (4 men, 4 left-handed) were paid foruse two different source representations (i.e., a detailedtheir participation and ranged in age from 18 to 34. All had normal
cortical versus a crude subcortical representation) to or corrected-to-normal vision and gave written, informed consentidentify the same value association (i.e., the behavioral in accordance with procedures and protocols approved by the Insti-value associated with the stimulus representation). tutional Review Board of the Yale University School of Medicine.
There are at least two possible models to explain howfMRI Data Acquisitionvastly dissimilar subcortical and cortical representa-In the scanner, participants viewed stimuli (subtending �5 visualtions could both trigger the same value association.degrees) through red/blue anaglyph glasses. Images were back-First, it may be that when the amygdala receives rela-projected onto a translucent screen mounted near the end of the
tively unprocessed subcortical input, it performs some- MRI gantry and were viewed through a periscopic prism system onthing analogous to simple template matching using the head coil. Behavioral response data were collected with a fiber-
optic button box, and the participant’s head was immobilized usingcertain representations that are tuned to specific viewsfoam wedges and tape across the forehead. T2*-weighted imagesof behaviorally relevant stimuli. Evidence for template-sensitive to BOLD contrast were acquired on a GE Sigma 1.5 Teslabased pattern vision has been found in other animalsscanner (LX operating system) with a standard quadrature headthat lack a visual cortex and whose pattern vision iscoil, using a gradient echo, single-shot echo planar sequence and
mediated by the thalamus and the optic tectum, the an oblique axial orientation determined for each participant (TR �homolog to the primate SC. In avians, these structures 1500 ms, TE � 40, flip angle � 60, NEX � 1, in-plane voxel size �appear to accomplish pattern recognition through “ac- 3.125 mm � 3.125 mm, 18 oblique slices, 3.3 mm thick with no
gap). In all participants, these slices covered extrastriate cortex,tive vision,” a template-matching mechanism whereinthe superior colliculus, the amygdala, and inferior aspects of thean animal actively changes its position in order to matchthalamus, and in six participants these slices also covered the entireits view with one of a limited number of stored patternthalamus. T1-weighted structural images of the same thicknesstemplates (Dawkins and Woodington, 2000). Second, it(TR � 500, TE � 14, field of view � 200 mm, 256 mm � 192 mm
might be that subcortical visual structures preceding matrix, 2 NEX) and a high-resolution 3D structural data set (3Dthe amygdala perform a hierarchical construction of an SPGR, contiguous, sagittal acquisition, 124 images with 1.2 mmobject representation, akin to processes within cortical isotropic voxels, TR � 24, TE � 5, flip angle � 45, matrix � 192 �
256, NEX � 2, FOV � 30 cm) were collected in the same session.object recognition pathway. Our results are consistentwith the results of blindsight and backward masking
Rivalry and Localizer Experimentsstudies, suggesting that the SC and a visual posteriorThe rivalry run consisted of six repetitions of three block typesthalamic nucleus may be involved in such perceptualcorresponding to one nonrivalry condition (a moving house) and
processes. Although more rudimentary and less de- two rivalrous conditions (suppressed fearful face and suppressedtailed, this subcortical representation might sufficiently chair). Stimuli were 10 s long movies created using Adobe Premiereapproximate the IT representation such that the same (Adobe Systems, Inc., San Jose, CA, http://www.adobe.com). Toamygdala neurons can effectively use either representa- allow the anaglyph glasses to filter different images to the two eyes
for the rivalrous stimuli, images of houses defined by red luminancetion to perform the necessary lookup. The pulvinar,were overlaid on the image of a simple disc defined by blue lumi-which has direct connections to both the SC (Beneventonance. The position of the red and blue lens was randomly alternatedand Fallon, 1975; Stepniewska et al., 2000) and theacross participants between the right and left eye. Each trial began
amygdala (Jones and Burton, 1976), is a likely candidate with the disc projected to one eye for 0.5 s. At this point, the housefor an intermediate stage in a subcortical visual hierar- image appeared in the other eye and began to move sharply backchy, given its involvement in basic pattern discrimination and forth while the blue disc gradually faded into a target image of
a fearful face or neutral chair also defined by blue luminance. The(Chalupa et al., 1976). For a subcortical pathway orga-

Neuron170
target image then faded back into the blue disc after approximately tical activation clusters, while at the same time eliminating singlevoxel activations that may be due to noise. Significant voxels outside1.5 s. In the nonrivalrous moving house condition, a house was
presented binocularly with movement identical to the monocular of these ROIs for which we had no a priori hypotheses were cor-rected for multiple comparisons across the whole brain.house in the rivalrous stimuli. Four trials (2.5 s each) presented
consecutively constituted one 10 s stimulus block with each block We also inspected the individual activation maps at a less strin-gent threshold of p � .05, uncorrected, in order to determine thefollowed by a 10 s rest period. Participants performed a 1-back
behavioral task indicating by button press if the current image was proportion of participants who exhibited activations for our mainsuppressed face versus suppressed chair contrast. Object-selectivethe same as or different than the previous image. We instructed
participants to press a separate button if at any point they perceived cortex was functionally defined on each individual’s 2D anatomyusing a fearful face versus chair contrast derived from each individu-anything besides a house, even small portions of a face or chair, in
order to identify and to exclude from our analysis the blocks where al’s localizer data. Time courses from the rivalry run were extractedfrom the five most significantly activated pixels in the left and rightperceptual suppression failed. Participants performed at greater
than 90% accuracy with no difference between suppressed image hemispheres in both face- and chair-selective IT. After normaliza-tion, the time courses were averaged by condition and the peaktrials (p � .5). All trials for which the participant did not respond
were treated as unsuccessful suppression trials (since they could signal amplitudes (positive or negative) for each individual wereentered into a two-way ANOVA (cortical ROI by suppressed imagenot be used to verify successful suppression). Across all partici-
pants, unsuccessful rivalry suppression trials occurred in 2% of type) to test for possible differential effects of the suppressed imageon the cortical ROIs. A second two-way ANOVA was performed thatthese blocks, which were subsequently excluded from further
analysis. included the amygdala as an additional ROI factor in order to testfor a significant differential effect of suppressed image type on theThe object localizer protocol was administered after the rivalry
protocol. It consisted of three runs with four blocks each of fearful amygdala and cortical ROIs. Finally, a repeated measures two-wayANOVA was performed on the condition-specific response functionsfaces, neutral faces, and chairs, randomly intermixed and separated
by a 10 s block of rest. Stimuli were presented as movies in a manner within each cortical ROI to test for significant differences betweenthe two rivalrous and the nonrivalrous conditions (image type bysimilar to the rivalry experiment except that each object type was
presented binocularly with eight consecutive trials constituting a time; the eight time points corresponding to an increased hemody-namic response were included, which took into account a 4.5 s lag20 s long block.for the peak hemodynamic response).
We used a PPI analysis (Friston et al., 1997) to evaluate condition-Data Analysis specific interactions between the left amygdala and our other a prioriImage analyses and tests of statistical significance were done using ROIs. In this separate analysis, we included the seven individuals forBrainvoyager 2000 (Brain Innovation, Maastricht, The Netherlands, whom our slice selection covered the dorsal posterior thalamushttp://www.BrainVoyager.com), and locally developed software was and for whom we observed individually significant left amygdalaused to inspect time courses and correlations. Motion-corrected activation (p � .05, uncorrected) resulting from a suppressed faceimages were high pass filtered at two cycles per time course and versus suppressed chair contrast. The time courses from the maxi-spatially smoothed using a Gaussian filter with a full-width half- mally activated voxel in the left amygdala in each individual weremaximum value of 7 mm. The individual data were coregistered with extracted, normalized, and entered into a separate random effectshigh-resolution 2D anatomical images for display and localization. analysis as a covariate of interest to identify significant, condition-
For group analysis, these data sets were then transformed into a specific correlations within each ROI taken from the group averagedproportional three-dimensional grid defined by Talairach and Tour- data. This analysis technique assesses whether correlations be-noux (1988) and coregistered with the high-resolution 3D data set tween regions during one condition differ significantly from thosethat was resampled to give 1 mm3 voxels. In the averaged group correlations during another condition. We also performed the samedata set, ROIs corresponding to face- and chair-selective IT areas PPI analysis using the superior colliculus as the source data. Forwere functionally defined based on a fixed effects general linear this we used ROIs that included the left and right superior colliculimodel (GLM) analysis contrasting fearful faces with chairs which, defined on each individual’s 2D anatomical images of the six individ-after convolution with a standard hemodynamic response function uals for whom our slice selection covered the entire thalamus. The(Boynton et al., 1996), were included as predictors in the GLM. same thresholds used in the main effects analysis were also usedEach ROI included contiguous voxels within ventral temporal cortex in these PPI analyses, both within and outside the ROIs.surviving a statistical threshold of p � .01, uncorrected (center Ta-lairach coordinates for each cluster were: face-selective, [right] x �
Acknowledgments47,y � �43, z � �18; [left] x � �38, y � �53, z � �17; chair-selective, [right] x � 28, y � �35, z � �10; [left] x � �27, y � �34,
This work was supported by grants from the National Institute ofz � �13). The subcortical ROIs were defined anatomically ratherChild Health and Human Development (grants PO1 HD 03008 andthan functionally because (1) we did not have an independent func-PO1 HD/DC35482). We wish to thank Hedy Sarofin, Terry Hickey,tional task that could reliably activate the SC or the visual thalamus,and Larry Win for technical assistance and members of the Percep-and (2) we did not want to assume that the localizer task (i.e., domi-tual Expertise Network for helpful discussion. Portions of this worknant face/chair presentations) would activate the same subregionswere previously presented at Cognitive Neuroscience Society, 2003of the amygdala as the rivalry task (i.e., suppressed face/chair pre-(B.N.P. and R.T.S.; fMRI evidence for emotional processing of per-sentations), based on the possibility that subregions receiving corti-ceptually suppressed faces in the amygdala during binocular rivalry)cal or subcortical input do not overlap.and at the 9th annual Human Brain Mapping conference, New York,In the rivalry task, specific effects of each condition in our a prioriNew York, June 18–22, 2003 (B.N.P. and R.T.S.; Differential amyg-group ROIs were evaluated by contrasts defined using a randomdala but not extrastriate activity is evidence for categorization ofeffects GLM with three predictors corresponding to the suppressedobjects not seen during binocular rivalry).face, suppressed chair, and dominant house conditions (separate
predictors were defined for those blocks in which rivalry suppressionfailed). Predictors were convolved with a standard hemodynamic Received: June 30, 2003response function (Boynton et al., 1996). Statistical maps of specific Revised: December 8, 2003contrasts were generated with a resampled voxel resolution of 1 Accepted: February 13, 2004mm3 corresponding to the resolution of the coregistered 3D anatomi- Published: April 7, 2004cal. Because of strong a priori hypotheses regarding our ROIs, sig-nificant voxels within these regions were identified using a statistical
Referencesthreshold of p � .005, uncorrected with a spatial extent greater thanfive contiguous 1 mm3 voxels (reported p values are corrected forfive multiple ROI comparisons). This cluster threshold was chosen Allison, T., Puce, A., Spencer, D.D., and McCarthy, G. (1999). Electro-
physiological studies of human face perception. I: Potentials gener-in an effort to minimize the possibility of excluding any small subcor-

Subcortical Processing of Unperceived Stimuli171
ated in occipitotemporal cortex by face and non-face stimuli. Cereb. Jones, E.G., and Burton, H. (1976). A projection from the medialpulvinar to the amygdala in primates. Brain Res. 104, 142–147.Cortex 9, 415–430.
Bar, M., and Biederman, I. (1998). Subliminal visual priming. Psychol. Kanwisher, N., McDermott, J., and Chun, M.M. (1997). The fusiformSci. 9, 464–469. face area: a module in human extrastriate cortex specialized for
face perception. J. Neurosci. 17, 4302–4311.Bar, M., and Biederman, I. (1999). Localizing the cortical regionmediating visual awareness of object identity. Proc. Natl. Acad. Sci. Koutstaal, W., Wagner, A.D., Rotte, M., Maril, A., Buckner, R.L., andUSA 96, 1790–1793. Schacter, D.L. (2001). Perceptual specificity in visual object priming:
functional magnetic resonance imaging evidence for a laterality dif-Benevento, L.A., and Fallon, J.H. (1975). The ascending projectionsference in fusiform cortex. Neuropsychologia 39, 184–199.of the superior colliculus in the rhesus monkey (Macaca mulatta).
J. Comp. Neurol. 160, 339–361. Kovacs, G., Vogels, R., and Orban, G.A. (1995). Cortical correlate ofpattern backward masking. Proc. Natl. Acad. Sci. USA 92, 5587–Blake, R. (1989). A neural theory of binocular rivalry. Psychol. Rev.5591.96, 145–167.Kraemer, M., and Sjostrom, A. (1998). Lack of short-latency-poten-Blake, R. (2001). Primer on binocular rivalry, including controversialtials in the VEP reflects immature extra geniculate visual functionissues. Brain Mind 2, 5–38.in delayed visual maturation (DVM). Doc. Ophthalmol. 97, 189–201.Blake, R., and Logothetis, N.K. (2002). Visual competition. Nat. Rev.Kreiman, G., Fried, I., and Koch, C. (2002). Single-neuron correlatesNeurosci. 3, 13–21.of subjective vision in the human medial temporal lobe. Proc. Natl.Boynton, G.M., Engel, S.A., Glover, G.H., and Heeger, D.J. (1996).Acad. Sci. USA 99, 8378–8383.Linear systems analysis of functional magnetic resonance imagingLeDoux, J.E. (1996). The Emotional Brain: The Mysterious Underpin-in human V1. J. Neurosci. 16, 4207–4221.nings of Emotional Life (New York: Simon & Schuster).Buckner, R.L., Goodman, J., Burock, M., Rotte, M., Koutstaal, W.,Leopold, D.A., and Logothetis, N.K. (1996). Activity changes in earlySchacter, D., Rosen, B., and Dale, A.M. (1998). Functional-anatomicvisual cortex reflect monkeys’ percepts during binocular rivalry. Na-correlates of object priming in humans revealed by rapid presenta-ture 379, 549–553.tion event-related fMRI. Neuron 20, 285–296.Linke, R., De Lima, A.D., Schwegler, H., and Pape, H.C. (1999).Calder, A.J., Lawrence, A.D., and Young, A.W. (2001). Neuropsychol-Direct synaptic connections of axons from superior colliculus withogy of fear and loathing. Nat. Rev. Neurosci. 2, 352–363.identified thalamo-amygdaloid projection neurons in the rat: possi-Cave, C.B., Blake, R., and McNamara, T.P. (1998). Binocular rivalryble substrates of a subcortical visual pathway to the amygdala. J.disrupts visual priming. Psychol. Sci. 9, 299–302.Comp. Neurol. 403, 158–170.
Chalupa, L.M., Coyle, R.S., and Lindsley, D.B. (1976). Effect of pulvi-Logothetis, N.K. (2003). The underpinnings of the BOLD functionalnar lesions on visual pattern discrimination in monkeys. J. Neuro-magnetic resonance imaging signal. J. Neurosci. 23, 3963–3971.physiol. 39, 354–369.Lomber, S.G. (2002). Learning to see the trees before the forest:Dailey, M.N., and Cottrell, G.W. (1999). Organization of face andreversible deactivation of the superior colliculus during learning ofobject recognition in modular neural network models. Neural Netw.local and global visual features. Proc. Natl. Acad. Sci. USA 99, 4049–12, 1053–1073.4054.
Dawkins, M.S., and Woodington, A. (2000). Pattern recognition andLumer, E.D., and Rees, G. (1999). Covariation of activity in visualactive vision in chickens. Nature 403, 652–655.and prefrontal cortex associated with subjective visual perception.
Dehaene, S., Naccache, L., Cohen, L., Bihan, D.L., Mangin, J.F., Proc. Natl. Acad. Sci. USA 96, 1669–1673.Poline, J.B., and Riviere, D. (2001). Cerebral mechanisms of word
Lumer, E.D., Friston, K.J., and Rees, G. (1998). Neural correlates ofmasking and unconscious repetition priming. Nat. Neurosci. 4,perceptual rivalry in the human brain. Science 280, 1930–1934.752–758.Miller, M., Pasik, P., and Pasik, T. (1980). Extrageniculostriate visionEpstein, R., and Kanwisher, N. (1998). A cortical representation ofin the monkey. VII. Contrast sensitivity functions. J. Neurophysiol.the local visual environment. Nature 392, 598–601.43, 1510–1526.
Friston, K.J., Buechel, C., Fink, G.R., Morris, J., Rolls, E., and Dolan,Morris, J.S., Ohman, A., and Dolan, R.J. (1998). Conscious and un-R.J. (1997). Psychophysiological and modulatory interactions in neu-conscious emotional learning in the human amygdala. Nature 393,roimaging. Neuroimage 6, 218–229.467–470.
Gallant, J. (2000). The neural representation of shape. In Seeing,Morris, J.S., Ohman, A., and Dolan, R.J. (1999). A subcortical path-K.K. DeValois and R.L. DeValois, eds. (San Diego, CA: Academicway to the right amygdala mediating “unseen” fear. Proc. Natl. Acad.Press), pp. 311–333.Sci. USA 96, 1680–1685.
Goebel, R., Muckli, L., Zanella, F.E., Singer, W., and Stoerig, P.Morris, J.S., DeGelder, B., Weiskrantz, L., and Dolan, R.J. (2001).(2001). Sustained extrastriate cortical activation without visualDifferential extrageniculostriate and amygdala responses to presen-awareness revealed by fMRI studies of hemianopic patients. Visiontation of emotional faces in a cortically blind field. Brain 124, 1241–Res. 41, 1459–1474.1252.
Goren, C.C., Sarty, M., and Wu, P.Y. (1975). Visual following andMorton, J., and Johnson, M.H. (1991). CONSPEC and CONLERN: apattern discrimination of face-like stimuli by newborn infants. Pedi-two-process theory of infant face recognition. Psychol. Rev. 98,atrics 56, 544–549.164–181.
Grill-Spector, K., Kushnir, T., Edelman, S., Avidan, G., Itzchak, Y.,Nakamura, K., Mikami, A., and Kubota, K. (1992). Activity of singleand Malach, R. (1999). Differential processing of objects under vari-neurons in the monkey amygdala during performance of a visualous viewing conditions in the human lateral occipital complex. Neu-discrimination task. J. Neurophysiol. 67, 1447–1463.ron 24, 187–203.Pessoa, L., Kastner, S., and Ungerleider, L.G. (2002a). AttentionalHenson, R.N., and Rugg, M.D. (2003). Neural response suppression,control of the processing of neutral and emotional stimuli. Cogn.haemodynamic repetition effects, and behavioural priming. Neuro-Brain Res. 15, 31–45.psychologia 41, 263–270.Pessoa, L., McKenna, M., Gutierrez, E., and Ungerleider, L.G.Ishai, A., Ungerleider, L.G., Martin, A., Schouten, J.L., and Haxby,(2002b). Neural processing of emotional faces requires attention.J.V. (1999). Distributed representation of objects in the human ven-Proc. Natl. Acad. Sci. USA 99, 11458–11463.tral visual pathway. Proc. Natl. Acad. Sci. USA 96, 9379–9384.Rolls, E.T. (1999). Brain and Emotion (Oxford: Oxford UniversityIwai, E., and Yukie, M. (1987). Amygdalofugal and amygdalopetalPress).connections with modality-specific visual cortical areas in ma-
caques (Macaca fuscata, M. mulatta, and M. fascicularis). J. Comp. Rolls, E.T. (2000). Neurophysiology and functions of the primateamygdala, and the neural basis of emotion. In The Amygdala: ANeurol. 261, 362–387.

Neuron172
Functional Analysis, J.P. Aggleton, ed. (New York: Oxford UniversityPress), pp. 447–478.
Rolls, E.T., and Tovee, M.J. (1994). Processing speed in the cerebralcortex and the neurophysiology of visual masking. Proc. R. Soc.Lond. B Biol. Sci. 257, 9–15.
Rolls, E.T., Tovee, M.J., Purcell, D.G., Stewart, A.L., and Azzopardi,P. (1994). The responses of neurons in the temporal cortex of pri-mates, and face identification and detection. Exp. Brain Res. 101,473–484.
Rolls, E.T., Tovee, M.J., and Panzeri, S. (1999). The neurophysiologyof backward visual masking: information analysis. J. Cogn. Neurosci.11, 300–311.
Romanski, L.M., Giguere, M., Bates, J.F., and Goldman-Rakic, P.S.(1997). Topographic organization of medial pulvinar connectionswith the prefrontal cortex in the rhesus monkey. J. Comp. Neurol.379, 313–332.
Rosa, M.G., and Schmid, L.M. (1994). Topography and extent ofvisual-field representation in the superior colliculus of the megachi-ropteran Pteropus. Vis. Neurosci. 11, 1037–1057.
Schiller, P.H., and Malpeli, J.G. (1977). Properties and tectal projec-tions of monkey retinal ganglion cells. J. Neurophysiol. 40, 428–445.
Schyns, P.G., and Oliva, A. (1999). Dr. Angry and Mr. Smile: whencategorization flexibly modifies the perception of faces in rapid vi-sual presentations. Cognition 69, 243–265.
Sewards, T.V., and Sewards, M.A. (2002). Innate visual object recog-nition in vertebrates: some proposed pathways and mechanisms.Comp. Biochem. Physiol. A Mol. Integr. Physiol. 132, 861–891.
Sheinberg, D.L., and Logothetis, N.K. (1997). The role of temporalcortical areas in perceptual organization. Proc. Natl. Acad. Sci. USA94, 3408–3413.
Simion, F., Valenza, E., Umilta, C., and Dalla Barba, B. (1998). Prefer-ential orienting to faces in newborns: a temporal-nasal asymmetry.J. Exp. Psychol. Hum. Percept. Perform. 24, 1399–1405.
Stefanacci, L., and Amaral, D.G. (2000). Topographic organizationof cortical inputs to the lateral nucleus of the macaque monkeyamygdala: a retrograde tracing study. J. Comp. Neurol. 421, 52–79.
Stefanacci, L., and Amaral, D.G. (2002). Some observations on corti-cal inputs to the macaque monkey amygdala: an anterograde tracingstudy. J. Comp. Neurol. 451, 301–323.
Stepniewska, I., Ql, H.X., and Kaas, J.H. (2000). Projections of thesuperior colliculus to subdivisions of the inferior pulvinar in NewWorld and Old World monkeys. Vis. Neurosci. 17, 529–549.
Talairach, J., and Tournoux, P. (1988). Co-planar Stereotaxic Atlasof the Human Brain (New York: Thieme).
Tong, F. (2001). Competing theories of binocular rivalry: a possibleresolution. Brain Mind 2, 55–83.
Tong, F., and Engel, S.A. (2001). Interocular rivalry revealed in thehuman cortical blind-spot representation. Nature 411, 195–199.
Tong, F., Nakayama, K., Vaughan, J.T., and Kanwisher, N. (1998).Binocular rivalry and visual awareness in human extrastriate cortex.Neuron 21, 753–759.
Vuilleumier, P., Armony, J.L., Driver, J., and Dolan, R.J. (2003). Dis-tinct spatial frequency sensitivities for processing faces and emo-tional expressions. Nat. Neurosci. 6, 624–631.
Webster, M.J., Ungerleider, L.G., and Bachevalier, J. (1991). Connec-tions of inferior temporal areas TE and TEO with medial temporal-lobe structures in infant and adult monkeys. J. Neurosci. 11, 1095–1116.
Weiskrantz, L. (1997). Consciousness Lost and Found (Oxford: Ox-ford University Press).
Whalen, P.J., Rauch, S.L., Etcoff, N.L., McInerney, S.C., Lee, M.B.,and Jenike, M.A. (1998). Masked presentations of emotional facialexpressions modulate amygdala activity without explicit knowledge.J. Neurosci. 18, 411–418.
Zimba, L.D., and Blake, R. (1983). Binocular rivalry and semanticprocessing: out of sight, out of mind. J. Exp. Psychol. Hum. Percept.Perform. 9, 807–815.




























