Embed Size (px)
Citation preview

DEVELOPMENTAL BIOLOGY 18, 6&-631 ( 1968 )
Studies on Intercellular Adhesive Selectivity’
STEPHEN ROTH”.”
Department of Biology, Case Western Resert;e Universit!y, Ckdand, Ohio
Accepted September 1.5, I’IGS
INTRODI!CTION
Adhesive selectivity between cells may play a role in morphogenetic phenomena. This property must have components at various sub- cellular levels of organization. To analyze the role of selectivity in morphogenesis at the cellular level, however, a technique is needed with which intercellular adhesive selectivities may be measured and compared. The following will attempt to satisfy this need and to begin such an analysis.
Selectivity has been defined (Steinberg, 1964) as “the faculty of a thing to interact with other things to different degrees.” In terms of intercellular adhesion, selectivity implies that a cell of a particular type can form more stable associations with one cell type than with another.
Adhesive selectivity, as defined above, has been employed to account for a large number of morphogenetic phenomena (Holtfreter, 1939; Townes and Holtfreter, 1955; Gustafson and Wolpert, 1963; Steinberg, 1964; Weston, 1963; Coman, 1944; Carter, 1965; Abercrombie and Heaysman, 1953; Abercrombie et al., 1957).
In all these cases, selective adhesion or, what amounts to the same thing, differences in adhesive stability between different cell types, have been supposed to exist and, furthermore, to exist in specific de-
1 Submitted in partial fulfillment of the requirements for the Degree of Doctor of Philosophy in the Department of Biology, Case Western Reserve \‘nivcrsity, Cleveland, Ohio.
’ Present address: McCollum-Pratt Institute, The Johns Hopkins I’nivcrsit!, Baltimore, Maryland.
’ This investigation was carried out under the tenure of developmental biolog> predoctoral training grant No. 2TlAD20 f’ IOU, the U. S. Public Health Srr~ice, arrtl NIH Grant No. 8-ROl-HD-03477 to Dr. James Weston.
GO2

INTERCELLULAR ADHESIVE SELECTIVITY 693
grees. The fact remains, however, that differences in intercellular ad- hesive stability between metazoan cells have never been demonstrated independently of the phenomena they are used to explain or from which they are predicted. To accomplish this, three requirements for measurement must be met. First, an operational criterion for inter- cellular adhesion must be selected since the mechanisms for adhesive interactions are unknown and direct measurement is impossible. The criterion selected should be a meaningful indicator or consequence of the adhesive interaction. Second, the operation by which the criterion is defined should be open to reliable and unequivocal measurement. Third, the measured criterion of one interaction should be suitable for comparison with those of other interactions. A measurement technique satisfying each of these requirements could be used to test many hypotheses which account for specific morphogenetic phenomena by assuming or predicting adhesive selectivity. Unfortunately, none of the methods previously used to measure intercellular adhesive stabili- ties satisfies all the above requirements. These techniques can be di- vided into two basic groups: physical separation (Coman, 1944; Herr- mann and Hickman, 1948; Easty et al., 1966; Weiss, 1961; Taylor, 1961) and reaggregation kinetics (Moscona, 1962; Curtis and Greaves, 1965; Steinberg and Granger, 1966; Lilien and Moscona, 1967).
One important criticism applies to all experiments attempting to measure cell adhesion by physical separation. The criterion chosen for measurement, namely ease of separation of cell from cell or cell from substrate, may not be a good indication of the actual interaction about which information is desired, namely cell adhesion to cell or cell ad- hesion to substrate. Weiss (1961) has pointed out that when two ad- herends are separated, the line of separation might be within one of the adherends rather than along the adhesive bonds holding the ad- herends together. Indeed, it has been shown (Weiss and Coombs, 1963) that cells disrupted from a glass substrate leave parts of their membranes or surfaces behind. Brooks et aL (1967) have also con- cluded from their studies of shear forces on tissue fragments that the intercellular adhesions, once formed, may be more resistant to sepa- ration than the cell membranes themselves.
Measurements using reaggregation kinetics are also inadequate since they fulfill only two of the three previously set requirements for the measurement of adhesive stability and the subsequent demonstra- tion of selectivity. The measurable criterion, probability of adhesion

604 STEPHEN ROTH
after collision, is clearly defined and does seem to give a meaningful indication of intercellular adhesion, although this remains to be demonstrated. Also, there are several methods available for measuring this criterion reliably. Its failure is with the third requirement that measurements of adhesive interactions between cell types be suitable for comparison. Adhesive stabilities between different cell types cannot easily be compared with this technique for two reasons.
First, similar dissociating agents may affect different cell types dif- ferently. That is, the same dissociating agent may injure one cell type more than another, or alternatively, two cell types may require differ- ent lengths of time to recover from similar levels of clamagt (Weiss and Kapes, 1966; Curtis, 1961) .
Second, in different cell suspensions the reaggregation technique cannot control for the effects of differences in cell size or concentration on the measured parameter. Both these extraneous variables can in- fluence the rate of collision within a cell suspension (Steinberg and Roth, 1964) and this, in turn, affects the rate of adhesion. Therefore. the reaggregation technique cannot differentiate between characteris- tic differences in the probability of adhesion among cells after collision and non-characteristic differences in the rates of collision.
Characteristic cellular differences in the probability function, if present, can be detected only if the extraneous factors affecting colli- sion can be eliminated as variables in the comparison. This has been accomplished in the following studies with the collecting-aggregate method of measuring relative adhesive stabilities (Roth and Vieston, 1967). Unlabeled aggregates serve as collecting particles and can be made of either the same cell type as a labeled suspension (an isotypic aggregate) or of a different cell type (a heterotypic aggregate). The results are presented in terms of the number of labeled cells collected by these aggregates. Adhesive interactions between different aggregate types and an identical suspension can be compared in this way under conditions where all extraneous factors influencing the number of ad- hesions, including the adhesive state and the degree of clustering of the cells of the suspension, are the same for the cell types being compared.
It should be emphasized that the measured parameter is not “ad- hesive stability,” or “cellular adhesion,” but simply the number of labeled cells collected by an aggregate during a given time period. It is assumed that this is a reflection of the probabilitv of adhesion be-

INTERCELLULAR ADHESIVE SELECTIVITY 605
tween the cell and aggregate in question. It is further assumed that this probability is one reasonable indicator of the undefined phenome- non, “intercellular adhesion.” Evidence for this assumption will be presented in the discussion.
A previous paper (Roth and Weston, 1967) has evaluated the col- lecting-aggregate system as a general method for the measurement of cell-to-cell adhesive stability. It determined the effects of parameters such as aggregate size, cell suspension density, and duration of collec- tion on the number of labeled cells collected and showed the statistical reliability of the system. That work also demonstrated that for chick embryo liver and neural retina cells, isotypic aggregates always col- lected more labeled cells than heterotypic aggregates in the same sus- pension. This paper will examine some aspects of cell selectivity by asking such questions as: Does selectivity exist between other cell types; do metabolic inhibitors and “conditioned” medium affect this system as they do others; do killed cells retain adhesive selectivity; how similar is the probability of adhesion between a cell and an aggregate to the probability of adhesion between a cell and a cell; and, finally, when does this selectivity appear and how do experimental procedures affect its manifestation?
MATERIALS AND METHODS
Dissociation and aggregation. Cells for both collecting aggregates and labeled suspensions were removed aseptically from White Leg- horn chicken embryos or from embryos of a substrain of DBA mice. Ages of the embryos from which tissues were taken will be given for each experiment. Development was measured for chicken embryos, from the onset of incubation and, for mouse embryos, from the time of appearance of a vaginal plug. Tissues were dissociated by pipetting them through a small-bore Pasteur pipette in a dissociating medium containing 0.25% crude collagenase, 0.25% crude trypsin, and 10% heated chicken serum in calcium- and magnesium-free saline. The method has been described in detail previously (Roth and Weston, 1967).
Unlabeled aggregates to be used as collecting particles were made from cell suspensions prepared 24 hours previously and containing about 2.5 x lo5 cells/ml for chicken embryo neural retina and about 6 x 105 cells/ml for all other cell types used. Three milliliters of the cell suspensions were allowed to aggregate in lo-ml DeLong culture

606 STEPHEN ROTH
flasks on a gyratory shaker bath (New Brunswick Scientific Company, New Brunswick, New Jersey, Model G76). All cultures were main- tained within a pH range of 7.2-7.6. The shaker bath was kept at 37°C and the platform was rotated at 70 rpm (diameter of rotation = % inch).
The medium used for preparing aggregates and for the collection of labeled cells consisted of 60% TC 199, 25% Hanks’ salt solution, and 15% heated chicken serum (56°C for 30 minutes; Grand Island Biologicals ) .
Labeling and radioautograph!y. Windows were cut in the shells of eggs to be labeled before 4 days of incubation. The windows were sealed with Cellophane tape until 24 hours before the labeled tissues were to be removed (usually 6 or 7 days of incubation). At this time the windows were opened and the first dose of 15 PC of tritiated thymidine (6 Clmmole; 100 &/ml) was dropped directly onto the embryo. The windows were closed again with tape and the eggs were reincubated until 4 hours before dissection when a second dose of 3 !.tC of the isotope was administered. When ll-12-dav embryos wcrc used, 20 and 10 PC were administered in the first and second doses.
Twelve-day mouse embryos were labeled by injecting pregnant mice with a single 100 &’ dose of tritiated thvmidine through a tail \rcin. hlothers were sacrificed 24 hours after the isotope dose, and the cm- bryos were immediately taken. Labeled tissues were collect~tl. dis- sociated, washed, and diluted as described above.
Unlabeled aggregates with approximately equal diamrtcrs \\‘er(-‘ selected under a dissecting microscope and were deposited in the DeLong flasks containing the labeled suspensions. Concentrations of labeled cells will be given for each experiment. Collection of labeled cells was carried out on the gyratory shaker under the same conditions described for the production of unlabeled aggregates.
After circulating in the labeled suspensions, collecting aggregates were removed and fixed in Bouin’s fluid, unless indicated otherwise. dehydrated, cleared, embedded in paraffin, and sectioned at 5 /I,. The slides were coated with Kodak NTB-2 Nuclear Track Emulsion, dried at room temperature, and exposed for 3 weeks in lightproof desiccating cans at 4%.
In each experiment, at least one flask of labeled cells of each tissue type was replaced on the shaker for an additional 12-15 hours after the collecting aggregates had been removed. The labeled aggregates thus formed were removed, treated exactly as \vere the unlabeled ag-

INTERCELLULAR ADHESIVE SELECTIVITY 607
gregates, and used as controls for labeling efficiency. These labeled aggregates indicated the percentage of labeled cells incorporated into aggregates. Labeling of cells in these aggregates always exceeded 90%.
Counting procedure. When radioautographic sections of collecting aggregates were examined, the third or quarter of the ribbon contain- ing the largest sections was estimated under a dissecting microscope. Labeled nuclei were counted on every third section in this zone to avoid recounting any single nucleus which might appear in adjacent sections. Data will be presented in terms of the mean number of la- beled nuclei per section in this zone + the standard error about the mean. The number of sections counted per aggregate will be given for each mean and standard error. When a grand mean is used, the num- ber of aggregate means contributing to the grand mean will be given.
Labeled cells on aggregate surfaces were counted without regard to their presence as single cells or as parts of cell clusters. In general, about one-half the cells counted were singles and about one-half were in clusters of at least two.
The aggregate diameters were determined by measuring the largest section with an optical micrometer. With irregularly shaped aggre- gates, the largest diameter was used.
Critique of the counting procedure. One implication of the de- scribed counting procedure is that the mean number of labeled nuclei per section should be lower than the actual number of labeled cells collected by the aggregate. For example, assume an aggregate is 0.3 mm in diameter, a cell nucleus is 5 ,J in diameter, and the mean path length of a beta particle from a tritium atom is 1 I*. Assume further that an aggregate has collected a single labeled cell. If every third section in the central fourth of the ribbon is examined, the probability that the single-labeled nucleus will be seen is about l/6 (l/4 surface area examined x about 213 chance of scoring the nucleus if it is in the examined area). If five sections are counted and the nucleus is scored, the mean number of labeled cells per third section + SE, will be 0.20 k 0.20. In other words, if six aggregates each contained one labeled cell, the counting procedure used would, on the average, yield five aggregates means of 0.0 and one aggregate mean of 0.2. It follows that an aggregate would have to collect about 30 labeled cells in order for the aggregate mean to be about one. Since about 10% of the cells are not labeled, an aggregate with a mean number of cells of one would have collected, on the average, a total of 33 cells. In short, an aggregate

608 STEPHEN ROT11
mean of zero does not necessarily mean that an aggregate has not col- lected any cells. An aggregate mean of one, moreover, indicates that an aggregate has probably collected many cells. Aggregate means will, therefore, underestimate by a constant factor the actual number of ~11s collected by an aggregate.
EXPERIMENTS AND RESCI,TS
Adhesizje Relationships betzceen Different Tbme.s
-4ggregates and suspensions were obtained from 12-day chick sternal cartilage and pectoral muscle as well as from ‘I-day, chick livers. All nine combinations of the aggregates and suspensions from these tissues were made and placed on the shaker for 6 hours. The results are pre- sented in Table 1. In each of the three suspension types, the means of similar-sized isotypic aggregates were pooled. The grand means were then set at 100% for comparison with the grand means of the hetero- typic aggregates within each suspension. The numbers of cells collected
NUMBER 0~ CELLS COLLECTED BY IIETER~TYP~(~ .\(X~RI:GATES l:.ELATI\.E TO
THAT COLLECTED BY IS~TYPIC AGGRE(:ATES IN TIIE SAME R~~si~~ss~o~.‘~
(‘art ilage Liver Peclor:~l muscle
u Time = 6 hours. ZJ Values are expressed as the percents of the mew tlrunl)er of cells collectcvl 1)~ the
isc~t ypic aggregates in each of the labeled srwpel~siorrs. i-\ctrul wlrles of the mean rltunl)er of labeled cells collected 1)~ isot yic aggregnles in cart ilagr, liver, :mtl pectoral muscle suspensions are 4.2, 60.0, and W.i, re*pect,ively.
c ?;tunl)ers in parent,heses are the numlwrs of aggreg:~tct meant: in the gr:tn~l me:1n. \Ilwre IIO~ indicated, datum is from a single aggregate.
by heterotypic aggregates are expressed as percentages of the isotypic grand mean. The numbers in parentheses following each datum are the number of aggregate means contributing to the grand mean. The results of this experiment show that for these three tissues, isotypic ag- gregates collect more labeled cells than do heterotypic aggregates in the same suspension for the same time.
In another experiment aggregates and labeled suspensions were pre-

INTERCELLULAR ADHESIVE SELECTIVITY 609
pared from livers and neural retinae of 7-day chicken embryos while aggregates alone were prepared from 13-day mouse embryo livers and neural retinae. Heterologous aggregates were placed in both labeled suspensions in iso- and heterotypic combinations. Isotypic, homologous aggregates were used as controls in each case. The results are shown in Table 2. Within both labeled suspensions-chick liver and chick neural retina-isotypic mouse aggregates collected more labeled cells than heterotypic mouse aggregates. During equivalent time periods, however, isotypic mouse aggregates collected fewer labeled cells than isotypic chicken aggregates in the same suspension.
TABLE 2 NUMBER OF CELLS COLLECTED BY Iso- AND HETEROTYPIC, HETEROLOGOUS
AGGREGATES RELATIVE TO THAT COLLECTED BY ISOTIPIC, HOMOLOGOUS AGGREGATES IN THE SAME SUSPENSIONSQ
Aggregate types, diameters =
0.42 f 0.06 mm
Chick liver Chick neural retina Mouse liver Mouse neural retina
a Time = 4% hours.
Labeled suspension type (density, in cells/ml)
Chick liver (106) Chick neural retina (IW)
loo” (2)C - - 100 (3)
65 3 10 61
b Values are expressed as the percents of the mean number of cells collected by the isotypic, homologous aggregates in each of the labeled suspensions. Actual values of the mean number of cells collected by these aggregates in chick liver and neural retina suspensions are 21.1 and 21.8, respectively.
c Numbers in parentheses are the numbers of aggregate means in the grand mean. Where not indicated, datum is from a single aggregate.
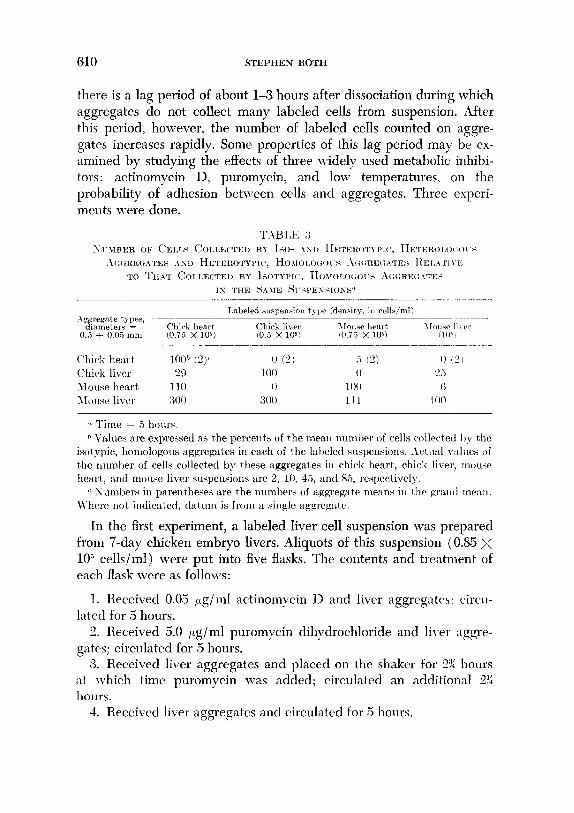
A third experiment was carried out to compare the results from all combinations of aggregates and suspensions of liver and heart ventricle cells of 7-day chick embryos and B-day mouse embryos. The results of this experiment are shown in Table 3. These data demonstrate that mouse liver aggregates from 1Sday embryos consistently collect more labeled cells than any of the other aggregates in any of the tested sus- pensions. Aside from this, however, between heart and liver cells from chick and mouse embryos, tissue specificity plays a larger role in the collection of cells by aggregates than does species specificity.
Effects of Metabolic Inhibitors
The results shown previously (Roth and Weston, 1967) indicate that
610 STEPHEN ROTH
there is a lag period of about l-3 hours after dissociation during which aggregates do not collect many labeled cells from suspension. After this period, however, the number of labeled cells counted on aggre- gates increases rapidly. Some properties of this lag period may be ex- amined by studying the effects of three widely used metabolic inhibi- tors: actinomycin D, puromycin, and low temperatures, on the probability of adhesion between cells and aggregates. Three experi- ments were done.
‘k Time = 5 hours. h Values are expressed as the percents of the mew ~luml~r of cells collectetl I)?; the
isot ypic, homologous aggregates in each of the laljeled slwpensions. Act \I:L~ v:tllles of the nrrmher of cells collected by these aggregates in chicak heart,, chick liver, T~O~ISP
heart, and mouse liver swpensions are 2, 10, 45, and ST,, respertively. c Xllmbers iu parentheses are the numbers of aggregate means in the yrn1111 me:tu.
Where llot indicated, datum is from a single aggregate.
In the first experiment, a labeled liver cell suspension was prepared from ‘i-day chicken embryo livers. Aliquots of this suspension (0.85 x 10” cells/ml) were put into five flasks. The contents and treatment of each flask were as follows:
1. Received 0.05 ,.tg/ml actinomycin D and liver aggregatc,s: eircu- lated for 5 hours.
2. Received 5.0 ;Lg/ml puromycin dihpdrochloride and liver aggre- gates; circulated for 5 hours.
3. Received liver aggregates and placed on the shaker for 2% hours at which time puromycin was added; circulated an additional 2?: hours.
4. Received liver aggregates and circulated for 5 hours.

INTERCELLULAR ADHESIVE SELECTIVITY 611
5. Received liver aggregates and placed on the shaker for 2% hours.
At the end of each treatment, the aggregates were collected and fixed. At one-fifth the concentration of the same sample of actinomycin D (0.01 pg/ml), the uptake of tritiated uridine by explants of chick embryo spinal ganglia in monolayer culture was decreased by 80% (Cowell, 1967). The results of this experiment are presented in Table 4. They show that actinomycin D had no detectable effect on the num- ber of labeled cells collected by isotypic liver aggregates, whereas puromycin decreased this number strikingly. These results indicate that some metabolic events which may be puromycin sensitive occur during the lag period which are necessary for the acquisition of ad- hesiveness between cells and aggregates. They also show that some puromycin-sensitive events occur after the lag period which are nec- essary for cell-to-aggregate adhesion.
TABLE 4 EFFECTS OF ACTINOMYCIN II AND PUROMYCIN ox THE NUMBER OF LABELED LIVER CELLS COLI,ECTED BY LIVER AGGREGATES"
Time (hours) Control Actinomycin D Puromycin
5 30.6 + 3.2 36.4 + 4.6 2.0 + 0.6 36 4 f 1.R 34.4 + 3.1 33.0 k 3.8
2p 9.8 IL 2.9 4.0 * 1.2 6.4 + 1.7
a Aggregate diameters = 0.29 * 0.05 mm. Values are expressed as mean IL SE. n = 5.
* After 24 hours on the shaker, the controls were removed and fixed and puromycin was added to the experimental flask. This flask was removed and its aggregates were fixed 2f hours later for total of 5 hours on the shaker.
The second experiment tested the effect of a temperature decrease on the probability of adhesion between isotypic combinations of ag- gregates and cells of ‘I-day chick heart ventricles and livers. The den- sities of the labeled heart and liver suspensions were 3 x lo4 and 6 X 10” cells/ml, respectively. Each suspension was aliquoted into four flasks, Two flasks of each suspension type on each shaker were in the water bath at 37°C. The other flask of each suspension type was on a platform raised above the water level on both shakers. The tempera-

612 STEPHEN ROTH
ture of the medium in the flasks on the raised platforms was the same as the room temperature throughout the experiment (2223’C). Iso- typic aggregates circulated in the suspensions for 5% hours. The results (Table 5) show that, in all cases, aggregates collected fewer cells at room temperature than did similar-sized aggregates in the same sus- pensions at 37°C. The results also show that aggregates circulated in labeled suspensions on a gyrator-y shaker collect more cells than do aggregates in the same suspension on an oscillatory shaker.
EFFECT OF DECREASED TEMPERATIJRE OS THE XURIBER OF LABELED
HEART AND LIVER CELLS COLI.EWED BY ISOTYPIC AGGREGATES"
The final experiment in this section was designed to determine when the temperature-sensitive events, in terms of labeled cell collection, were occurring. A labeled liver suspension (lo” cells/ml) was made from ‘I-day chicken embryos. The suspension was aliquoted into five flasks and after receiving isotypic aggregates, treated as follows:
1. Placed in the 37°C bath for 4 hours. 2. Placed in an ice bath (O’C) attached to the shaker for 4 hours. 3. Placed in the 37°C bath for 2 hours. 4. Placed at 0°C for 2 hours and then at 37°C for 2 additional hours. 5. Placed at 37°C for 2 hours and then at 0°C for 2 additional hours.
At the end of each treatment, the aggregates were collected and fixed. The results (Table 6) show again that the number of labeled cells collected by aggregates in 4 hours at 0°C is much less than that

INTERCELLULAR ADHESIVE SELECTIVITY 613
at 37°C. Also, the number of labeled cells collected by aggregates in flasks 4 and 5 are greater than those collected by aggregates in flask 3. There are, therefore, temperature-sensitive and insensitive cellular events which affect the formation of stable, cell-to-aggregate adhesions occurring both during and after a lag period of approximately 2 hours.
TABLE 6 EFFECTS OF Low TEMPERATURE AT VARIOUS TIMES ON THE NUMBER
OF LABELED LIVER CELLS COLLECTED BY LIVER AGGREGATES”
4 hr at 37°C 2 hr at 0°C 4 hr at 0°C 2 hr at 37°C 2 hr at 37°C ;trattt3072 0
109.2 f 5.7 3.0 * 1.1 8.2 f 1.4 27.8 k 4.4 63.2 5 9.1 95.6 _+ 4.0 1.2 f 0.2 17.8 I!z 1.5 44.9 * 4.9 89.6 f 7.8
117.4 k 8.6 1.6 k 1.4 8.4 k 2.5 39.4 f 4.4 44.2 f 1.6 73.4 k 4.1 86.2 * 5.0
86.0 + 7.0
Grand means : 98.9 1.9 11.5 56.9 65.7
a Aggregate diameters = 0.3 f 0.04 mm. Values are expressed as mean f SE. n = 5.
Effects of “Conditioned” Media
“Conditioned” medium (Moscona, 1962) has been reported to en- hance the rate of aggregation of dissociated, chick neural retina cells. A series of experiments was designed, therefore, to elucidate the ef- fects of used media on the number of labeled cells collected by un- labeled aggregates. For the purposes of these experiments, conditioned medium (CM) means nutrient medium (NM) which has supported cells of a particular type in suspension for 1 day. The densities of the conditioning cells were 2 X lo5 cells/ml for 7-day chick embryo neural retina cells, and 6 x lo5 cells/ml for 7-day chick embryo heart ven- tricle and liver cells.
Effect of homologous CM. In this experiment, a suspension of labeled liver cells from 7-day embryos was distributed into two centrifuge tubes, The cells were pelleted by centrifugation and the supernatants were discarded. The cells of one tube were then resuspended with NM, while the cells of the other tube were resuspended with an equal amount of liver-CM. Both isotypic (liver) aggregates and neural retina aggregates were added to each suspension after they had been

614 STEPHEN ROTH
aliquoted into DeLong flasks. A suspension of neural retina cells was treated identically except that the CM used for resuspension was con-
ditioned by neural retina cells. These suspensions also received isotypic as well as liver aggregates. Table 7 compares the difference in number of labeled cells collected by aggregates in NM and CM for 4 hours. In isotypic combinations of liver cells and aggregates, liver-CM increased the number of collected cells by a factor of about 25. In neural retina isotypic combinations, the number was increased by a factor more than three. The numbers of heterotypic adhesions are also increased by CSI but to a lesser degree since previous experiments havre shov\,u near]!; comparable collection by heterotypic aggregates under similar con- ditions without CM (see Roth and Weston, 1967).
Aggregate. .Suspension diameters =
(IO” cells ‘ml) 0.34 * 0.02 mm N3I
Specificity of the C&l. Two basic questions must be asked about the properties of CM. First, will any Ch4 increase the number of any iso- typic adhesions or will CM have a positive effect only on the cells and aggregates of the tissue type which conditioned it? Second, is this in- crease in collection of labeled cells due to an increase in the probability or rate of adhesion after the lag period, or do the cells acquire adhesive properties earlier while the probability of adhesion remains about the same? The following experiment was designed to answer these two questions.
Labeled suspensions of 7-day chick embryo heart ventricle (8 x lo&), liver (lOj), and neural retina (lo” cells/ml) were prepared. From each of these suspensions, 9 ml was aliquoted into each of four centri- fuge tubes. After the cells were pelleted and the NM removed, each
INTERCELLULAR ADHESIVE SELECTIVITY 615
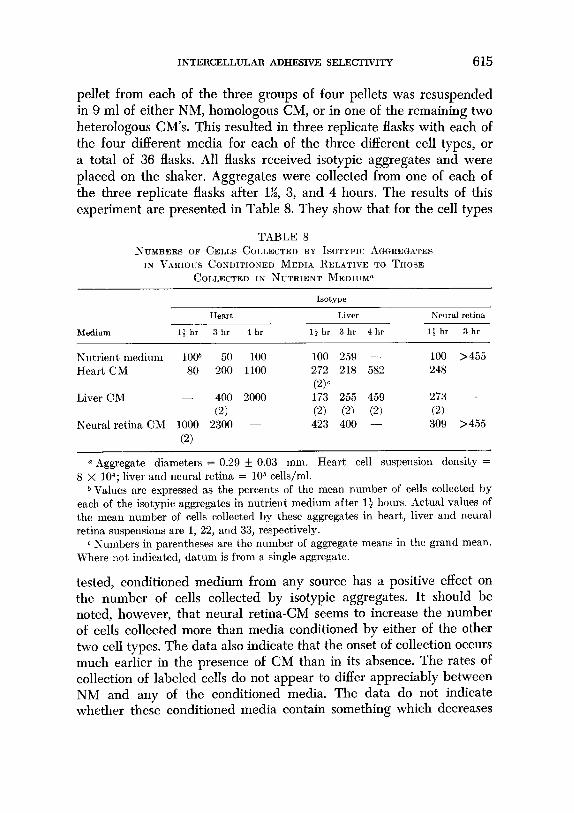
pellet from each of the three groups of four pellets was resuspended in 9 ml of either NM, homologous CM, or in one of the remaining two heterologous CM’s. This resulted in three replicate flasks with each of the four different media for each of the three different cell types, or a total of 36 flasks. All flasks received isotypic aggregates and were placed on the shaker. Aggregates were collected from one of each of the three replicate flasks after l%, 3, and 4 hours. The results of this experiment are presented in Table 8. They show that for the cell types
TABLE 8 NCYBERS OF CELLS COLLECTED BY ISOTYPIC .~GREGATES
Is VARIOUS CONDITIONED MEDIA IIELATIVE TO THOSE
COLLECTED IN NUTRIENT MEDIUIP
1sotype
Medium
Heart Liver Neural retina
13 hr 3 hr 4 hr ljhr 3hr 4hr 1: hr 3 hr
Nutrient medium lOoh 50 100 Heart CM 80 200 1100
Liver CM - 400 2000
(2) Neural retina CM 1000 2300 -
(2)
100 259 - 100 >455 272 218 582 248 -
cw 173 255 459 273 - (2) (21 @‘i 0) 423 400 - 309 >455
a Aggregate diameters = 0.29 + 0.03 mm. Heart cell suspension density = 8 X 104; liver and neural retina = lo5 cells/ml.
b Values are expressed as the percents of the mea.n number of cells collected by each of the isotypic aggregates in nutrient medium after 13 hours. Actual values of the mean number of cells collect,ed by these aggregates in heart, liver and neural retina suspensions are 1, 22, and 33, respectively.
c Numbers in parentheses are the number of aggregate means in the grand mean. Where not, indicated, datum is from a single aggregate.
tested, conditioned medium from any source has a positive effect on the number of cells collected by isotypic aggregates. It should be noted, however, that neural retina-CM seems to increase the number of cells collected more than media conditioned by either of the other two cell types. The data also indicate that the onset of collection occurs much earlier in the presence of CM than in its absence. The rates of collection of labeled cells do not appear to differ appreciably between NM and any of the conditioned media. The data do not indicate whether these conditioned media contain something which decreases

616 STEPHEN ROTH
the lag period or whether they lack something which otherwise would cause the lag period.
Low temperatures and CM. Aliquots of a labeled liver suspension (lo” cells/ml) were centrifuged and resuspended as before in NJ4 or CM (pooled from liver and neural retina suspensions). All the flasks
contained liver aggregates and were placed either at 37°C or at 22°C as previously described. The results (Table 9) show unequivocally that the negative effect of low temperatures on the collection of la- beled liver cells by liver aggregates is eliminated by CM.
Puromycin and Ch1. A labeled liver suspension (10” cells/ml) was distributed, centrifuged, and resuspended either in NM, with and with-
out puromycin (5 pg/ml), or pooled liver and neural retina CM, with and without puromycin. Isotypic aggregates circulated in these sus- pensions for 6 hours. The results (Table 10) reveal that the number of labeled cells collected in CM plus puromycin is greater than that in NM alone. The negative effect of puromycin, therefore, is overcome by the CM.

mmm~~~mm ADHESIVE SI~LJ~CTIVITY 617
Fragments versus aggregates
This experiment was performed to determine whether or not prior dissociation and reaggregation of collecting aggregates affects the se- lectivity exhibited by them. Specifically, the experiment was designed to determine the similarity between surfaces of aggregates made from cells of dissociated tissues and undissociated fragments of the tissues themselves.
Labeled suspensions were made from 7-day chick livers and from neural retinae (both at 105 cells/ml), Tissue fragments were prepared by mincing 7-day chick livers and neural retinae in watch glasses with curved scissors until most of the pieces were about the size of the most often used aggregates (about 0.3 mm). These fragments were de- posited in NM in DeLong flasks and placed on the shaker exactly as were the unlabeled suspensions for the production of collecting aggre- gates. After 1 day, the initially irregular and jagged fragments of both tissue types had rounded up and become indistinguishable from the aggregates of the same type. Each labeled suspension received iso- and heterotypic aggregates and fragments. The suspensions and col- lecting particles were placed on the shaker for 6 hours. The results (Table 11) demonstrate that the selectivity previously shown for liver and neural retina aggregates is also characteristic of fragments of these tissues. There are, as well, no important differences in the degrees of these selectivities.
TABLE 11 COMPARISON BETWEEN THE MEAN NUMBER OF LABELED CELLS COLLECTED
BY AGGREGATES AND FRAGMENTS IN IDENTICAL SUSPENSIONP
Collecting particle, Suspension, diameter = density =
0.32 f 0.02 mm 106 cells/ml Aggregate Fragment
Liver Liver 13.8 + 2.7 (5) 9.6 k 1.5 (5) 11.8 5 4.1 (5) 27.6 k 4.0 (5)
Neural retina Liver <l O-(5) <l.O-(5) Neural retina Neural retina 53.8 + 4.4 (4) 48.0 f 3.9 (4) Liver Neural retina <l.O-(5) < l .O-(5)
a Time = 6 hours. Values are expressed as mean f SE. (n)
Selectivity between Dead Cells and Aggregates
Is the demonstrated selectivity a product of the living state or can it be exhibited by dead cells or aggregates? The following experiment

618 STEPHEN ROTH
was performed to answer this question. Labeled suspensions (2 x 10” cells/ml) of 7-day chick liver and neural retina cells were prepared, and two aliquots from each suspension were centrifuged. One pellet from each suspension type was resuspended in glutaraldehyde (8.1% in 0.1 M sodium cacodylate, pH = 7.8) at 4°C while the control pellets
Smpension type
Liver (SM)
Killed liver (YM) Liver, 0.5 Selmtl rctilw 0 :;
2.0 * 0 -I(51 0 0 i 0.0 (5) 0 0 * 0.0 I.51
Killed liver, 0.X Killed Ileum1 ret itia, 0 L’
0.0 * 0.0 (a) 0.0 + 0 0 I.51
Neural retina (NM) Liver, 0.4T Sewal re+:r, 2-4
0 0 + 0.0 (-51 > IO0 (31 > too (.i!
Killed liver, 0.4 Killed twruxl retilta. 0 :I
0.0 + 0.0 (5) :I ‘L & 0.6 (31
Killed neural retina (CM) Liver, 0.21 Selllxl retilm, 0 45 t 0 I.?
1 s f 12(5l 0 6 k 0 4 (51 I 2 * 0 ti I*2
.5 ci * l.‘I . Ii1 .
I<illetl liver, 0.4 Killrcl ~wr~r:rl ret illa 0 1
2 x f 1 2 15, S (i + 2 ti I .j)
were resuspended in Hanks’ saline at room temperature. After 30 min- utes, both suspensions were washed three times by centrifugation and resuspension. The fixed suspensions were washed with sucrose (0.2 31 sucrose in 0.1 M sodium cacodylate, pH = 7.1-7.5) at 4°C while the

INTERCELLULAR ADHESIVE SELEcTnTITY 619
control suspensions were washed with Hanks’ saline at room tempera- ture. After the final centrifugation, living neural retina and liver pellets and fixed liver pellets were resuspended in NM. Fixed neural retina pellets were resuspended in pooled liver and neural retina CM.
Aggregates to be fixed were first washed in Hanks’ saline, They were then transferred to cacodylate-buffered glutaraldehyde at 4°C. After 30 minutes, they were washed by three transferrals to cacodylate- buffered sucrose also at 4°C. After one further wash in Hanks’, they were placed in the desired suspensions.
Killed and living suspensions received iso- and heterotypic, killed and living aggregates in all combinations. These cultures circulated on the shaker for 7 hours. The results are illustrated in Table 12. It is clear that the number of liver or neural retina cells collected is greatly reduced when either the cells in the suspension, aggregates, or both are fixed. However, in all combinations where either the suspension or the aggregates in it have been killed, there are more isotypic than heterotypic adhesions. This is especially apparent in cases where the aggregates are fixed but the suspensions are living.
Relatively large numbers of killed neural retina cells were collected by heterotypic as well as by isotypic, killed aggregates in the presence of CM. There was no equivalent collection of killed liver cells by killed aggregates in the absence of CM. Whether this is due to a property of neural retina cells or to the presence of CM, remains to be determined.
When cells or aggregates were killed by heat (60°C for 10 minutes) or 95% alcohol, there was no detectable collection of labeled cells by iso- or heterotypic aggregates.
Initial Adhesions
Previous experiments reported in this paper have shown that aggre- gates will selectively collect cells of their own kind as soon as the la- beled cells of the suspension become adhesive to the aggregates. That is, between the cells and aggregates tested, and under the given con- ditions, there is no period of nonselective adhesion when the number of heterotypic adhesions equals the number of isotypic adhesions. If there is similarly no period of nonselective adhesion between two types of freshly dissociated cells then one would predict that the addition of unlabeled neural retina cells to a suspension of labeled liver cells should have no effect on the number of labeled cells collected in this

620 STEPHENROTH
mixed suspension by either liver or neural retina aggregates. In order to test this prediction, the following three experiments were carried out. In all three experiments, suspensions ( lo5 cells/ml) and aggre- gates came from 7-day chicken embryos.
In the first experiment, a labeled liver suspension was divided into two aliquots. Each was centrifuged and the supernatants discarded. One aliquot was resuspended with NM, and the other was resuspended with a second suspension of unlabeled neural retina cells. As a result, the total cell concentration of the second suspension was 2 >< lo’ cells/ml, half of these being labeled liver cells and the other half being unlabeled neural retina cells. Each of the two resulting suspensions received liver and neural retina aggregates and the flasks were cir- culated on the shaker for 3 hours and 50 minutes. Table 13 summarizes the results of this experiment. In the mixed suspension there is a de- crease in the number of labeled liver cells collected by liver aggregates but an increase in the number of labeled liver cells collected by neural retina aggregates, relative to the numbers collected by similar aggre- gates in the labeled liver suspension alone. It is obvious that adhesions are occurring between the labeled liver cells and the unlabeled neural retina cells in suspension.
TABLE: 13 EFFECT OF ADDED, USLABELED NECRAL RE~lr4.4 ChxLs oN TIT~
NUMBER OF LABELED LIVER CELM COI.I,ECTED BY NEURAL RETINA AND LIVER AGGREGaTESa
Aggregates (diameters!
I>* suspension. 10s cells/ml J.* + N suspension, 2 X (mean l SE) 105 cells!ml (meall t SIC!
Liver (0.34 f 0.04 mm) 45.4 * 2 .i Ill.0 * 1.0 28.0 + x.7 11.6 !c 2 s 28.2 * 2.4
Neural retina (0.5 +I 0.05 mm) 3.x + 1.0 31.0 + 1 :; 6.S + 0.6 14.8 * l..i 2.5 + 0.9 11.8 * 1.7
n Time = 4 hours. n = 5. * Labeled.
The second experiment contains a duplicate of the first as well as its reciprocal. Labeled neural retina and mixed labeled neural retina plus unlabeled liver suspensions were prepared exactly as described above as were labeled liver and mixed labeled liver and unlabeled neural
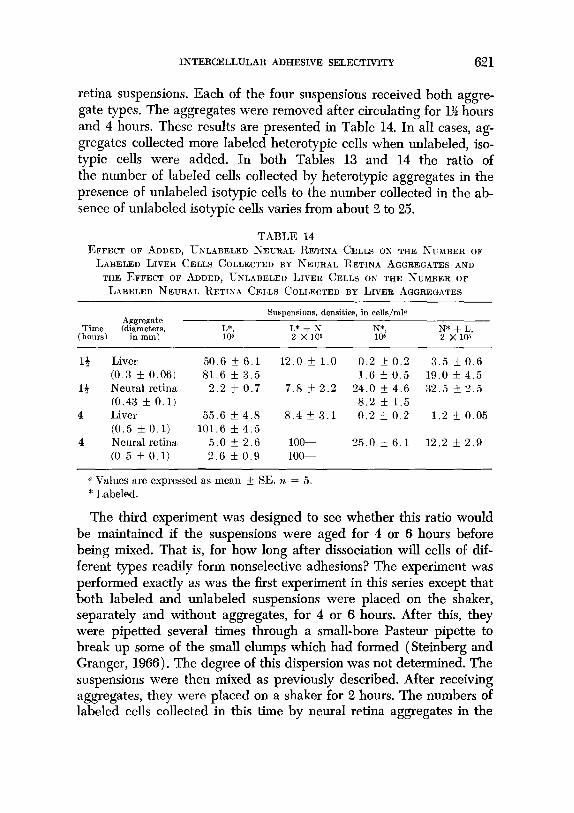
INTERCELLULAR ADHESIVE SELECTIVITY 621
retina suspensions. Each of the four suspensions received both aggre- gate types. The aggregates were removed after circulating for 1% hours and 4 hours. These results are presented in Table 14. In all cases, ag- gregates collected more labeled heterotypic cells when unlabeled, iso- typic cells were added. In both Tables 13 and 14 the ratio of the number of labeled cells collected by heterotypic aggregates in the presence of unlabeled isotypic cells to the number collected in the ab- sence of unlabeled isotypic cells varies from about 2 to 25.
TABLE 14 EFFECT OF ADDED, UNLABELED NEURAL RETINA CELLS ON THE NTXBER OF
LABELED LIVER CELLS COLLECTED BY NEURAL RETINA AGGREGATES AND
THE EFFECT OF ADDED, UNLABELED LIVER CELLS ON THE NUMBER OF
LABELED NEURAL RETINA CELLS COLI,ECTED BY LIVER AGGREGATES
Aggregate Suspensions, densities, in cells/ml~
Time ‘di”,“&s, I,*, L* + N, N*, (hours)
N* + L, 105 2 x 105 105 2 Xlos
1+ Liver 50.6 k 6.1 12.0 * 1.0 0.2 57 0.2 3.5 f 0.6 (0.3 5 0.06) 81.6 f 3.5 1.6 k 0.5 19.0 * 4.5
13 Neural retina 2.2 f 0.7 7.8 f 2.2 24.0 f 4.6 32.5 f 2.5 (0.43 It 0.1) 8.2 + 1.5
4 Liver 55.6 rk 4.8 8.4 + 3.1 0.2 5 0.2 1.2 * 0.05 (0.5 f 0.1) 101.6 k 4.5
4 Neural retina 5.0 + 2.6 loo- 25.0 f 6.1 12.2 f 2.9 (0.5 f 0.1) 2.6 IL 0.9 loo-
o Values are expressed as mean + SE. n = 5. * Labeled.
The third experiment was designed to see whether this ratio would be maintained if the suspensions were aged for 4 or 6 hours before being mixed. That is, for how long after dissociation will cells of dif- ferent types readily form nonselective adhesions? The experiment was performed exactly as was the first experiment in this series except that both labeled and unlabeled suspensions were placed on the shaker, separately and without aggregates, for 4 or 6 hours. After this, they were pipetted several times through a small-bore Pasteur pipette to break up some of the small clumps which had formed (Steinberg and Granger, 1966). The degree of this dispersion was not determined. The suspensions were then mixed as previously described. After receiving aggregates, they were placed on a shaker for 2 hours. The numbers of labeled cells collected in this time by neural retina aggregates in the

622 STEPHEN ROTH
TABLE 15 EFFECT OF ADDED, UNLABELED iXEuR.4L IIETrs.4 CELLS os THE XI-miE:R
OF L.ABELED LIVER CELLS CoLLEcTELI BY ~IKR.41, ItETIU.4 AGGREo.4W!3
DURING A ~-HOUR PERIOD AFTER TRE ~‘~I-SPEX?IOXS H.~vE BEES CIRCULATISG UNMXED FOR 4 OR 6 HOVRV
Time of incubatim ,Sllslwmaior,s, dmaities. in cells,‘ml prior to mixing __-- -_
Chows) I.*, 105 I.” + N (2 x IO”
4 1.x k 0.0X I) (i * O.(i 0.0~- 0.6 + 0 L’
6 0 o- I) 4 f 0 2 0 s + 0 4
CL Aggregate diameters = 0.9 mm. Time of :t,,, ~~~reg~at~ circrdatiotl = 2 hoiw. \.:L~II(+ are expressed as meRn f SE. rr = 5.
mixed suspensions are shown in Table 15. In both cases, the ratios do not vary significantly from one.
DISCUSSION
Properties of the Collecting-Aggregate Method
Use of the collecting-aggregate system described in this study as- sumes that the probability of forming a stable adhesion between a cell and an aggregate is a unique cellular property. The system can mani- fest and compare the probabilities of adhesion between any number of aggregates and the cells of a single suspension independent of es- traneous variables affecting the rate of initial contacts between the cells and the aggregates. The number of cells adhering to an aggregate under these conditions reflects this probability. In addition to revealing specific differences in adhesive stability, this procedure may be used to examine the effects on net adhesion of the extraneous factors them- selves. Comparisons between numbers of cells collected by aggregates in suspensions of different cell types are not valid since the extraneous variables discussed above cannot be considered equivalent. As a re- sult, adhesive stabilities between isotypes cannot be compared directly.
Probabilit!/ of Adhesion as a Measure of Adhesive Stabihty.
Because this system yields consistent, quantitative data, it is well suited for measuring the probabilities of adhesion between different cells and aggregates. It is important, however, that these probabilities be established as meaningful indicators of intercellular adhesions. -4

INTERCELLULAR ADHESIVE SELECTIVITY 623
direct relationship between these two is, of course, impossible to prove and even difficult to infer since one phenomenon has a precise, oper- ational definition while the other has usually been described in non- operational terms. The following discussion will list the evidence con- sistent with the idea that the probability of adhesion between a cell and an aggregate is a good indicator of the adhesive stability between that cell type and the cell type of the aggregate.
Tissue-specific sorting out in mixed aggregates is usually (Moscona, 1962; Steinberg, 1964; Trinkaus and Lentz, 1964), but not always ( Curtis, 1961)) attributed to tissue-specific differences in adhesive strengths. This sorting out has been described for a number of tissues. Chick limb premuscle (Moscona, 1952), chick liver (Moscona, 1957), and chick heart ventricle (Steinberg, 1963) each sort out from chick limb precartilage cells. Also, neural retina (Steinberg, 1962b) and liver cells (Steinberg, 1963) each sort out from heart ventricle cells. In ad- dition, Moscona (1957) has shown that cartilage cells, neural retina cells, and kidney cells from mouse and chick embryos sort out primarily according to tissue as if there were no species difference.
The results obtained with the collecting-aggregate technique are consistent with all these observations. Tissue-specific differences in the probabilities of adhesion have been shown between chick liver and neural retina cells, between chick cartilage, liver, and pectoral muscle cells, and between chick heart ventricle and liver cells. Tissue-specific differences have also been shown between chick and mouse embryonic liver, neural retina, and heart ventricle cells. Furthermore, this tech- nique has revealed that the probability of adhesion is slightly greater between homologous, isotypic cells and aggregates than between heterologous, isotypic cells and aggregates. This species-specific dif- ference was not detected by simple observations of aggregates after sorting out of cells had occurred. If sorting out is due to tissue-specific differences in adhesive stability, then it is clear that the data presented here are consistent with the hypothesis that the parameter being measured is a good indicator of these adhesive stabilities.
The reaggregation of previously dissociated cells is also a process involving the net formation of intercellular adhesions. It has been ob- served that low temperatures (Moscona, 1962; Steinberg and Granger, 1966) as well as puromycin (Moscona and Moscona, 1963) decrease the rate of formation of these adhesions. Actinomycin D is reported to have only a slight inhibitory effect on reaggregation during the first

624 STEPHEN ROTH
&5 hours after dissociation (Moscona and Moscona, 1963). These general results are also obtained with the collecting-aggregate tech- nique. Low temperatures (both 22’C and 0°C) greatly reduce the number of labeled liver cells collected by unlabeled liver aggregates. Puromycin has a similar effect at the same concentration used in the Moscona and Moscona (1963) system. Actinomycin D, however, has no detectable effect on labeled cell collection during the first 5 hours after cell dissociation. The results presented here also indicate that there are temperature- and puromycin-sensitive events necessarv for cell collection occurring during and after the lag period.
Moscona (1962) has also reported the existence of a cell-free super- natant with specific, cohesion-promoting abilities. This supernatant came from lo-day neural retina cell suspensions (at about 10” cells/ ml) which were rotated on a gyratory shaker for 2 hours at 100 rpm at 38°C. This used medium was also able to negate the inhibitorv ef- fect of low temperature on aggregate formation in neural retina. sus- pensions. Although this supernatant was considered to be tissue- specific, the results of the critical reciprocal experiments were not reported. Lilien and Moscona (1967) have reported similar results with a dialyzed, serum-free supernatant from monolayered neural retina cells, but again no results from reciprocal experiments went re- ported. With the method used in this study, analogous results arc ob- tained although the conditioned medium contains serum and is exposed to cells for 24 hours. Conditioned medium was found to increase th(, number of labeled cells collected largely by erasing the lag period. The present system also confirmed the finding that conditioned medium negated the inhibitory effects not only of low temperatures but of puromycin as well. Especially interesting in the light of the results TC- ported by Moscona (1962) and Lilien and Moscona (1967) is the fact that neural retina conditioned medium is more effective, in terms of labeled cells collected, than liver-conditioned medium or hcart- conditioned medium. Reciprocal experiments with liver-, heart-, and neural retina-conditioned media, however, indicate no significant tis- sue specificity.
The final correlation between probability of adhesion, as measrlrecl with the collecting-aggregate technique, and the general concept of intercellular adhesion concerns the sorting out which occurs between fused fragments ( Steinberg, 1962c). Steinberg successfully predicted that if subsequent adhesive properties were independent of the cells’

INTERCELLULAR ADHESIVE SELECTIVITY 625
starting condition, then two fused fragments of different tissues would be expected to yield the same final pattern as a mixed aggregate con- taining those same cell types. The tissues arranged themselves inter- nally or externally in the fused fragments as in the mixed aggregates, A similar prediction can be made for testing with the collecting-aggre- gate method. If aggregate surfaces after 24 hours are similar to undis- sociated fragment surfaces of the same tissue type, then these frag- Ments should collect the same numbers of labeled cells as similar sized aggregates in the same suspension. This prediction also is verified with chick embryo liver and neural retina aggregates and fragments.
Measurements of Selectivity
The results of experiments with the collecting-aggregate system are consistent with many observations on what has generally been called selective cell adhesion. Some of these results are, however, not consistent with two hypotheses which seek to account for the degree and the time of onset of this selectivity.
Degree of selectivity. SteinLerg (1963, 1964) postulates that in mixed aggregates at equilibrium where one cell type, e.g., liver, sorts out completely externally from another cell type, e.g., heart, the work. or strength, of cohesion between the heart cells should be greater than that between liver cells. ile further predicts that the work of adhesion between the two cell t);pes should be of intermediate value. If the probability of adhesion may be considered a good indicator of strength of adhesion then one would predict that, in the present experiments. heart aggregates should collect more IabeIed heart cells than would liver aggregates in the same suspension. This does happen. The hy- pothesis also predicts that heart aggregates should collect more labeled liver cells than liver aggregates in the same suspension. This never happens, IsotypJic associations are almost always more probable than heterotypic association,. c In this context then, either the probability of adhesion beu-Ig measured is not a good indicator of work of adhesion, or work of ,dhesion is not playing its postulated role in sorting out.
Onset oj Jelectivity. The present results snow that as soon as cells adhere to aggregates, they adhere selectively. For a11 the tissues ex- amined, except 13-day mouse liver, isotypic aggregates collect more labeled cells than heterotypic aggregates. These data do not agree with the hypothesis that for about 12-24 hours after dissociation cells are nonselectively adhesive (for discussion see Steinberg, 1963; Moscona,

626 STEPHEN ROTH
1965; Steinberg and Roth, 1964). After this period, the hypothesis states that the cells acquire their tissue-specific characteristics an d sorting out begins.
This hypothesis might explain why aggregates from mixed suspen- sions contain both cell types in proportions roughly similar to those of the two original suspensions (Steinberg, 196%). In order to test this hypothesis, an experiment was carried out to test the assumptions for
freshly dissociated liver and neural retina cells. This was accomplished by adding unlabeled cells to flasks containing aggregates and labeled cell suspensions. It was found that the presence of unlabeied neural retina cells decreased the number of labeled liver cells collected h\ liver aggregates as compared to the number collected in the absencr of any unlabeled neural retina cells. Some interaction seems to occur. therefore, between the freshly dissociated, labeled liver cells and un- labeled neural retina which decreases the probability that labeled cells will be collected. In addition, the presence of unlabeled neural retina cells significantly increases the number of labeled liver cells collectt~d bv unlabeled neural retina aggregates. Reciprocal experiments sub stantiate these findings. This suggests that some adhesions occur br- tween dissociated cells of the two types. It is also shown that \vhrn labeled and unlabeled cell suspensions are circulated on the gyrator\ shaker for 4-6 hours and then mixed and seeded with collecting aggrci- gates, no such effects are seen.
The simplest explanation for these results is that prior to 1 hours after dissociation, liver and neural retina cells readily adhertt to oncl another. These adhesions would result in cell clusters composed of labeled and unlabeled cells. Although a substantial number of the labeled cells in these clusters may still be able to form and maintain adhesions with isotypic aggregates and bring unlabeled cells along
with them, there may or may not be a decrease in labeled cell collcc- tion. On the other hand, if the unlabeled cells in these mixed clusters can similarly form and maintain adhesions with their isotypic aggre- gates (these aggregates are, of course, heterotypic with respect to the labeled cell type), they will take the hetcrotypic, labeled cells \\.ith them. This would result in a definite increase in the number of labeled cells collected by heterotypic aggregates when compared to thr same collection in the absence of any isotypic, unlabeled cells. By about .-I hours, the probability of adhesions between heterotypic ~11 types in suspension must decrease since the addition of unlahcled ~~~11s no

INTERCELLULAR ADHESIVE SELECTIVITY 627
longer alters the collection of labeled cells by aggregates. Therefore, if dissociated cells will adhere to heterotypic cells but not to hetero- typic aggregates, then the adhesive properties of the cell surfaces must be different from those of the aggregate surfaces. By 4 hours in sus- pension, dissociated cell surfaces no longer may be distinguished in this way from the surfaces of collecting aggregates.
The results of the experiments just described are consistent with the hypothesized existence of initial nonselectivity. The period o’f this non- selectivity, however, is no more than 4 hours long for liver and neural retina cells instead of the previously postulated 12-24 hours. Variability of this no8nselective period as a function of different tissues has not been investigated but could provide the opportunity to test Curtis’ ( 1961) timing hypothesis definitively.
A working hypothesis can be constructed at this point which will ac- count for the experimental observations reported in this paper. For this model, some assumptions must be made. The evidence that these assumptions are valid varies considerably.
First, it must be assumed that there are molecular groups on cell surfaces which are responsible for selective adhesion. Evidence for a tissue-specific molecular moiety of this type is circumstantial, at best. The fact that there is some residual adhesive selectivity between cells and aggregates that have been killed with glutaraldehyde suggests two relevant conclusions. First, selectivity cannot be explained entirely in terms of filopodial activity on a cell’s surface. Second, there is a struc- tural component in or on the cell surface which is, somehow, respon- sible for this adhesive selectivity.
A second assumption is that the selective surface component(s) would be removed or altered by dissociation with crude trypsin and collagenase. When this occurs, nonselective adhesions would be made more likely.
The final assumption is that two cells can easily adhere to one another only when both cells have lost their adhesively selective com- ponents or when they have both regained them. In the first case, the cells may be of different types. In the second case, however, adhesion will be favored only if both cells are from the same tissue. Adhesion between the two cells would be unlikely if the second cell has either different selective components or none at all. In other words, the ex- istence of specific molecular groups on one cell surface necessitates the existence of the same groups on the second cell surface. There is

628 STEPHEN ROTH
no evidence for this assumption but, as with the other three assump- tions, the methods with which it can be tested are now becoming available.
Given these assumptions, it is expected that isotypic aggregates in labeled suspensions will selectively collect labeled cells onlv after the specific binding sites have been regenerated, and then onlv if the ag- gregates and the cells possess similar tissue-specific sites. Since the aggregates are composed of cells dissociated 24 hours previouslv. they, of course, would already possess the molecular groups responsible tor selective adhesions. Because of this, adhesions between cells and heter- otypic aggregates are not favored at any time after the cells have been dissociated. When two different suspensions are mixed, one Iabcletl and the other not labeled, the cells can adhere to one another vcr\. quickly. When the specific sites are regenerated, the cells will bc al,&, to adhere to isotypic aggregates but now would pull along attachcld. labeled, heterotypic cells. In this way, heterotypic collection can 1~ increased by the addition of isotypic cells. This hypothesis fnrthel predicts that after the cells in the different suspensions havr regcner- ated their selective sites, nonselective adhesions between the cells could not easily occur. This is consistent with the observation that the addition of isotypic but unlabeled cells no longer enhances the col- lection of heterotypic, labeled cells after 4 hours.
The hypothesis described above fulfills three essential criteria for a useful working hypothesis: ( 1) it explains the observations, (2 1 its as- sumptions can each be tested, and, (3) it suggests further testable prc- dictions based on important properties of the system.
Some of these assumptions and predictions concern the existence and characterization of the molecular moieties responsible for selcc- tive and non-selective intercellular adhesion. These predictions ma\’ be tested with methods such as specific enzymatic degradation df postulated surface substances, appearance of surface label when these substances are regenerated from isotopicallv labeled precursors, and the inhibition of adhesive selectivity by specific antibodies or hapten- like compounds. The collecting-aggregate system could serve as an assav for changes in selectivity effected by the procedures listed above.
Recent modifications in the system using highly labeled cells anti scintillation counting of the collecting aggregates will allow more rapid experimentation as well as the determination of true rates of collection.

INTERCELLULAR ADHESIVE SELECTIVITY 629
SUMMARY
The probability that two cells will form and maintain an adhesion after collision occurs in suspension is assumed to indicate the adhesive stability between the cells in question. Unlabeled cell aggregates are circulated in tritiated thymidine-labeled cell suspensions where all extraneous factors which might have affected collision are held con- stant. After circulating, the aggregates are collected, sectioned and examined radioautographically for labeled cells.
Experiments are carried out with cells and aggregates from chicken embryo neural retinae, livers, sterna, heart ventricles and pectoral muscles and with mouse embryo neural retinae and heart ventricles. Isotypic aggregates (same tissue type as the cells in the suspension) always collect more labeled cells than similar-sized heterotypic aggre- gates (different tissue type).
Collections of labeled chicken embryo liver cells by unlabeled liver aggregates is decreased by low temperatures (0’ and WC) and by puromycin but not by actinomycin D. “Conditioned” medium increases the collection and counteracts the inhibitory effects of low tempera- tures and puromycin.
Some degree of adhesive selectivity is shown by neural retina and liver cells and aggregates which were killed with glutaraldehyde.
Because the presence of unlabeled cells alters the collection of la- beled cells of another type by iso- and heterotypic aggregates, it is con- cluded that nonselective adhesions occurred between freshIy dis- sociated cells of different types more readily than between cells and heterotypic aggregates.
A hypothesis is described which assumes that there are at least two different molecular moieties on the cell surface which are responsible for intercelluIar adhesion, One of these causes seIective and the other nonselective adhesions. Experiments to test predictions of the hy- pothesis are discussed.
During the course of this work, I received invaluable advice from Dr. James A. Weston and excellent technical assistance from Mrs. Bonnie Edwards.
REFERENCES
ABERCROMBIE, M., and HEAYSMAN, J. E. M. (1953). Observations on the social behavior of cells in tissue culture. I. Speed of movement of chick heart fibro- blasts in relation to their mutual contacts. E~ptl. Cell Res. 5, 111-131.
ABERCROMBIE, M., HEAYSMAN, J. E. M., and KARTHAUSER, H. M. (1957). Social behavior of cells in tissue culture. III. Mutual iniluence of sarcoma cells and fibroblasts. ExptZ. Cell Res. 13, 276-291.

630 STEPHEN ROTH
BHOOKS, D. E., MILLAH, J. S., SEAMAN, G. V. F., and VASSAR, I?. S. ( 1967). Some physicochemical factors relevant to cellular interactions. J. Cell. Physiol. 69, 155-168.
CARTER, S. B. (1965). Principles of cell motility: The direction of cell movrrnent and cancer invasion. Nature 208, 1183-1187.
COIIIAF, D. R. (1944). Decreased mutual adhesiveness, a property of cells front squamous cell carcinomas. Cancer Res. 4, 625-629.
Co\?‘ELL, L. C. (1967). An analysis of melanogenesis in cultnred chick rmhryo spinal ganglia. Thesis, Department of Biology, Case Western Reserve University.
CURTIS, A. S. G. (1961). Timing mechanisms in the specific adhesions of cells. Exptl. Cell Res., Suppl. 8, 197-122.
CURTIS, A. S. G., and GREAVES, hl. F. (1965). The inhibition of cell aggregation lry a pure serum protein. J. Emhryol. Exptl. Morph& 13, 309-326.
EASTI-, G. C., EASTY, D. M., and AhlHHOSE, E. J. (1960). Studies of cellrilar ad- hesiveness. Exptl. Cell Res. 19, 539-547.
GUSTAFSON, T., and WOLPERT, L. (1963). The cellular basis of nmrphogewsis and sea urchin development. Intern. Rec. Cytol. 15, 139-214.
HERRhlANN, H., and HICKMAN, F. (1948). VII. The adhesion of epithclimn to stroma in the cornea. Bull. Johns Hopkins Hosp. 82, 182-224.
HOLTFRETER, J. (1939). Gewebeaffinitat, ein Mittel der embryonalen Formbildung. Arch. Exptl. ZelZforsch. 23, 169-209, as translated in “Foundations in Exprri- mental Embryology” (B. Willier and J. Oppenheimer, Eds. ). Prentice-IIall, New Jersey, 1964.
LILIEN, J. E., and MOSCOXA, A. A. (1967). Cell aggregation: Its cnhancenrent In a supernatant from homologous cells. Science 157, 76-72.
MOSCONA, A. A. ( 1952). Cell suspensions from organ rucliments of chick embry-os. Exptl. Cell Res. 3, 536-539.
MOSCONA, A. A. (1957). The development in vitro of chimaeric aggregates of dis- sociated embryonic chick and mouse cells. Proc. Natl. Acad. Sci. U. S. 43, 1X4- 194.
MOSCONA, A. A. (1962). Analysis of cell recombinations in experimental synthesis of tissues in uitro. J. Cellular Comp. Physiol. 60, Suppl. 1, 65-80.
MOSCONA, A. A. ( 1965). In ‘Cells and Tissues in Culture” (E. N. Wilmer. Ed. ). Vol. I. Academic Press, New York.
MOSCONA, A. A., and MOSCONA, H. (1963). Inhibition of adhesiveness and aggre- gation of dissociated cells by inhibitors of protein and RNA synthesis. Science 142, 1076-1071.
ROTH, S. A., and WESTON, J. A. (1967). The measurement of interccllrrlar nd- hesion. PTOC. Natl. Acad. Sci. U. S. 58, 974-980.
STEIKBERG, M. S. (1962a). On th e mechanism of tissue reconstruction 1~ dis- sociated cells. I. Population kinetics, differential adhesiveness, ancl the absenct of directed migration. Proc. N&Z. Acad. Sci. U. S. 48, 1577-1582.
STEINBERG, M. S. (1962b). On the mechanism of tissue reconstruction hv disso- ciated cells. II. Time course of events. Science 137, 762-763.
STEIKBERG, M. S. (1962c). On the mechanism of tissue reconstruction 1~~ disso- ciated cells. III. Free energy relations and the reorganization of fused, hetero- nomic tissue fragments. Proc. Natl. Acud. Sci. V. S. 48, 1769-1776.

INTERCELLULAR ADHESIVE SELECTIVITY 631
STEINBERG, M. S. ( 1963). Tissue reconstruction by dissociated cells. Science 141, 401-408.
STEINBERG, M. S. ( 1964). In “Cell Membranes in Development” (M. Locke, Ed.) Academic Press, New York.
STEINBERG, M. S., and GRANGER, R. E. (1966). The re-acquisition of adhesiveness by trypsinized chick embryo cells in uitro. Am. Zoologist 6, 579. (Abstract.)
STEINBERG, M. S., and ROTH, S. A. (1964). Phases in cell aggregation and tissue reconstruction. An approach to the kinetics of cell aggregation. 1. Exptl. Zool. 157, 327-338.
TAYLOR, A. C. ( 1961). Attachment and spreading of cells in culture. Exptl. Cell Res., Suppl. 8, 154-173.
TOWNES, P. L., and HOLTFRETER, J. (1955). D irected movements and selective ad- hesions of embryonic amphibian cells. J. Exptl. Zool. 128, 53-120.
TRINKAUS, J. P., and LENTZ, J. P. (1964). Direct observation of type specific segregation in mixed cell aggregates. Deuelop. BioZ. 9, 115-136.
WEISS, L. ( 1961). Measurement of cell adhesion. ExptZ. Cell Res., Suppl. 8, 141- 153.
WEISS, L., and COOMBS, R. R. ( 1963). The demonstration of rupture of cell sur- faces by an immunological technique. Exptl. CeZZ Res. 30, 331-338.
WEISS, L., and KAPES, (1966). Observations on cell adhesion and separation fol- lowing enzyme treatment. EzytZ. Cell Res. 41, 601-608.
WESTON, J. A. ( 1963). A radioautographic analysis of the migration and localiza- tion of trunk neural crest cells in the chick. Develop. BioZ. 6, 279-310.





























