Embed Size (px)
Citation preview

ARTHRITIS & RHEUMATISMVol. 46, No. 3, March 2002, pp 824–836DOI 10.1002/art.10102© 2002, American College of Rheumatology
Stromal Cell–Derived Factor 1 (CXCL12) Induces MonocyteMigration Into Human Synovium Transplanted Onto
SCID Mice
M. C. Blades,1 F. Ingegnoli,1 S. K. Wheller,2 A. Manzo,1 S. Wahid,1 G. S. Panayi,1
M. Perretti,2 and C. Pitzalis1
Objective. The mechanisms by which monocyte/macrophage cells migrate to the joint involve a series ofintegrated adhesion and signaling events in which che-mokines and their receptors are strongly implicated.This study was undertaken to investigate the hypothesisthat stromal cell–derived factor 1 (SDF-1), a CXCchemokine (CXCL12), plays a critical role in monocyte/macrophage localization to synovium.
Methods. SDF-1 and CXC receptor 4 (CXCR4)expression in rheumatoid arthritis (RA) and osteoar-thritis synovium and graft SDF-1, tumor necrosis factor� (TNF�), and human and murine vascular markerswere examined by immunohistochemistry and double-immunofluorescence. The functional capacity of SDF-1to modulate monocyte migration into joints was inves-tigated by examining the localization of pro-myelomonocytic U937 cells into synovial tissue trans-planted into SCID mice. SDF-1, TNF�, or saline wasinjected into graft sites and response determined by thenumber of fluorescently labeled U937 cells (injectedintravenously) detected in grafts by ultraviolet micros-copy.
Results. SDF-1 and CXCR4 were highly expressedin CD68� cells in the RA synovium. SDF-1 inducedU937 cell migration in vitro and in vivo in a dose-dependent manner and, in vivo, SDF-1 was more effec-tive than TNF�. In contrast to TNF�, SDF-1 did not
induce intracellular adhesion molecule 1 in transplantmicrovasculature. Furthermore, intragraft injection ofSDF-1 did not up-regulate TNF�, or vice versa.
Conclusion. This study demonstrates, for the firsttime, that SDF-1 is functional in vivo when injected intosynovial grafts. In addition, SDF-1 is more potent thanTNF�, and its mechanisms of action appear to beautonomous. Therefore, SDF-1 may be an importantTNF-independent molecule involved in the migration toand retention of inflammatory effector cells in the joint.
The chronic synovitis of rheumatoid arthritis(RA) is characterized by an inflammatory infiltrateconsisting of HLA–DR–positive antigen-presenting cellsin close contact with activated T lymphocytes, the ma-jority of which belong to the helper/memory(CD4�,CD45RO�) Th1 phenotype (1–5). Other char-acteristic features of rheumatoid synovitis include newblood vessel formation and thickening of the lining orintimal layer (5,6). The normal intima is only 1 or 2 cellsthick, while in RA this may increase to as many as 30cells, chiefly comprising macrophage- and fibroblast-likecells (5,7). Cells of the monocyte/macrophage lineage, inparticular, are thought to be important in mediating theeffector/destructive phase of the disease, via the secre-tion of pro-erosive cytokines and other inflammationmediators (5,8). This concept is further supported by theresults of association studies demonstrating a directcorrelation between the number of infiltrating macro-phages and joint erosions (9). The origin of intimalmacrophages is uncertain, but there is evidence suggest-ing that they derive from bone marrow precursors andarrive in the joint as monocytes (10). Although othermechanisms, including local proliferation and decreasedapoptosis, can contribute to the progressive accumula-tion of intimal macrophages in the synovium, it isbelieved that this phenomenon is largely due to sus-tained monocyte recruitment from the circulation (5,11).
Supported by the Welcome Trust and the Arthritis ResearchCampaign.
1M. C. Blades, BSc, F. Ingegnoli, MD, A. Manzo, MD, S.Wahid, BSc, G. S. Panayi, MD, DSc, C. Pitzalis, MD, PhD, FRCP:Guy’s, St Thomas and King’s College School of Medicine, London,UK; 2S. K. Wheller, BSc, M. Perretti, MD: The William HarveyResearch Institute, London, UK.
Address correspondence and reprint requests to C. Pitzalis,MD, PhD, FRCP, Rheumatology Unit, 5th Floor, Thomas Guy House,Guy’s Hospital, London SE1 9RT, UK. E-mail:[email protected].
Submitted for publication July 9, 2001; accepted in revisedform October 9, 2001.
824

Leukocyte migration from the bloodstream intoperipheral tissues is a complex multistep process regu-lated by a series of integrated adhesion and signalingevents (12–15). The signaling events necessary to acti-vate surface integrins, with subsequent firm adhesionand transendothelial migration, are principally mediatedby a family of chemoattractant cytokines, or chemokines(16,17). Chemokines share structural similarities, includ-ing 4 conserved cysteines that form disulfide bonds inthe tertiary structure of the proteins (18). More than 40chemokines have been described, the majority of whichcan be incorporated into 2 large subfamilies, CXC andCC chemokines, according to whether an interveningamino acid residue spaces the first 2 cysteines in themotif or they remain adjacent (18).
Chemokines signal by binding to specific Gprotein–linked receptors that span the cell membrane 7times (19–21). Engagement of these receptors results inactivation of small GTP binding proteins, such as theRas, Rac, and Rho families, and phospholipases, withdownstream formation of inositol triphosphate and intra-cellular Ca2� mobilization, eventually leading to integrinactivation (22,23). The importance of chemokines insynovial inflammation is indicated by the fact that vari-ous chemokines are found in abundance in the RAsynovium and are biologically active. These includemacrophage inflammatory protein 1� (MIP-1�), MIP-1�, RANTES, monocyte chemotactic protein 1,interleukin-8, and epithelial neutrophil–activating pep-tide 78 (24–29). In addition, at least part of synovial fluidchemotactic activity has been shown to be chemokinerelated (26). Furthermore, their functional importancehas been confirmed by studies of treatments in animalmodels of inflammatory arthropathies. Blocking anti-bodies against these mediators significantly reduced thenumbers of leukocytes recruited to the joint and theseverity of the arthritis. Conversely, administration ofchemokines to the joints of experimental animals re-sulted in an increase in infiltrating leukocytes and sig-nificantly worse disease (24,27,30–34).
Recently, another chemokine, stromal cell–derived factor 1 (SDF-1), has been implicated in therecruitment of bloodborne leukocytes to the RA joint.SDF-1 belongs to the CXC chemokine family and hasbeen recently renamed CXCL12 (35). In contrast to themajority of chemokines, SDF-1 interacts monogamouslywith a specific receptor, LESTR/FUSIN, subsequentlytermed CXCR4 (36). Both SDF-1 expression andCXCR4 expression have been found to be essential fornormal myelo- and lymphopoiesis, both in the embryoand in the adult (37). In addition, SDF-1 has been shownto be instrumental in mobilizing hematopoetic precur-
sors from the bone marrow into the circulation andsubsequently into peripheral tissues (38). Finally, SDF-1is expressed on the luminal surface of high endothelialvenules (HEVs) in peripheral lymph nodes, and initiallyit was thought to play a central role in lymphocytemigration mainly to secondary lymphoid organs (39).However, subsequent work has shown that SDF-1 is alsoexpressed in dermal microvascular endothelium ofchronically inflamed skin, indicating a more general rolefor this chemokine in the recruitment of leukocytes tosites of inflammation (40).
The importance of SDF-1 in the recruitment ofinflammatory cells to the joint is suggested by the largeamounts of SDF-1 messenger RNA found in RA syno-vial fibroblasts (41,42). In addition, the SDF-1 receptor(CXCR4) has been shown to be expressed in high levelsby synovial CD45RO� synovial T cells (43,44). Further-more, Buckley and colleagues have shown that trans-forming growth factor �, contained in synovial fluid,could induce sustained expression of CXCR4 on T cellsand that, upon stimulation, these CXCR4-expressing Tcells exhibited integrin-dependent adhesion in vitro (44).
To date, there are no published data on theexpression of SDF-1 or CXCR4 on the monocyte/macrophage population in the synovium. Therefore, webegan this study with a systematic analysis, by immuno-histochemistry, of the tissue and cellular distribution ofSDF-1 and CXCR4 in RA and osteoarthritis (OA)synovial membrane (SM). We confirmed that proteinexpression of SDF-1 and CXCR4 in the cellular infil-trate in RA was abundant and was much higher than inOA. In addition, SDF-1 was found prominently in thelining layer, in association with CD68� as well asfibroblast-like cells. To test directly in vivo the conceptthat SDF-1 may be instrumental in the recruitment ofmonocytes to the RA SM, we used a model in whichhuman synovial membrane (HuSM) is transplanted sub-cutaneously into SCID mice (45). We demonstrated, forthe first time, that SDF-1 injected intragraft is not onlyactive in vivo, but is more potent than tumor necrosisfactor � (TNF�) in up-regulating the migration of thepro-myelomonocytic U937 cell line to synovial trans-plants. In addition, we showed that SDF-1 and TNF� areindependently regulated and act via autonomous mech-anisms. Although TNF� remains a key cytokine in thepathogenesis of RA, a significant implication of ourfindings is that SDF-1 is an important TNF-independentmolecule that likely modulates the migration of inflam-matory cells to the joint. This may have importantimplications regarding the treatment of RA that isresistant to TNF-blocking agents.
SDF-1 AND MONOCYTE LOCALIZATION TO SYNOVIUM 825

PATIENTS AND METHODS
Patients. Ten patients with RA (mean age 64.8 years,range 54–72) and 10 with OA (mean age 68.5 years, range66–76) were included in the study. All patients had RA or OAthat fulfilled the American College of Rheumatology (former-ly, the American Rheumatism Association) criteria (46,47),and all were treated with nonsteroidal antiinflammatory drugs.Additionally, all patients with RA had been or currently werereceiving 1 or more disease-modifying antirheumatic drugsand/or steroids. All patients provided informed consent asapproved by the hospital ethics committee (LREC 98/11/27).
Tissue collection, preparation, and storage. Synovialtissue was obtained during joint replacement surgery, pro-cessed for routine cryohistologic study, and stored at �70°C.Samples for transplantation were cut into 0.5-cm3 pieces,frozen in 20% DMSO (Sigma, Poole, UK) in heat-inactivatedfetal calf serum (FCS; PAA Systems, Linz, Austria), and storedin liquid nitrogen (45).
Immunohistologic analysis of SM in RA and OApatients. Serial cryosections (10 �m) were mounted on poly-L-lysine (Sigma)–coated slides and dried overnight at roomtemperature. Sections for immunohistochemical analysis werefixed in acetone at 4°C for 10 minutes, wrapped in aluminumfoil, and stored at �70°C. Sections were analyzed by variousstandard immunohistologic techniques as described below.
Indirect immuno–alkaline phosphatase staining wasperformed using the avidin–biotin–alkaline phosphatase sys-tem (Dako, Cambridge, UK) and red substrate kit (Vector,Peterborough, UK), with 50 mM levamisole added to inhibitendogenous enzyme activity, as previously described (48).Sections were dehydrated, cleared in xylene, and mountedusing DePEX (BDH Chemical, Poole, UK) for examination bylight microscopy.
Double immunofluorescence staining was carried outusing direct and indirect techniques. When the antibodiesbeing tested were raised in different species (e.g., rabbit andmouse), indirect immunofluorescence staining was performed.Sections were washed in 50 mM Tris buffered saline (pH 7.6)and incubated in nonimmune serum for 30 minutes and in thefirst primary antibody for 1 hour. After washing, the appropri-ate fluorescein isothiocyanate (FITC)– or tetramethylrhodam-ine isothiocyanate (TRITC)–conjugated secondary antibodywas applied for 1 hour. Free secondary antibody binding siteswere then blocked with nonimmune serum, followed by incu-bation in the second primary antibody, washing, and incuba-tion in conjugated secondary antibody. When directly conju-gated primary antibodies were used or if the 2 primaryantibodies being tested were raised in the same host, theappropriate directly conjugated primary antibody was applied.Finally, sections were washed and mounted in Immunofluor(ICN Flow, Basingstoke, UK) before examination using ultra-violet (UV)–fluorescence microscopy and FITC- and TRITC-specific excitation filters. Control sections with 1 or bothprimary antibodies omitted and, in the case of directly conju-gated primary antibodies, their replacement with appropriatelyconjugated irrelevant control antibodies, were included foreach sample. Sections of normal lymph node and tonsil wereused as positive controls. A list of the antibodies used ispresented in Table 1.
SM from RA and OA patients was analyzed systemat-ically, with 4 different compartments considered (lining layer,sublining, vessels, and perivascular inflammatory aggregates).Immunostaining was assessed semiquantitatively. The propor-tion of the tissue compartment staining positively was scoredfrom 0 to 4 (0 � no staining, 1 � 1–25% stained; 2 � 26–50%;3 � 51–75%; 4 � 76–100%). The staining intensity for each
Table 1. Primary and secondary antibodies used for immunohistochemistry studies*
Clone Specificity Host Source
Primary antibodies (unconjugated)F8/86 vWF Mouse Dako
CD68 Mouse Dako6.5B5 ICAM-1 Mouse †B-B15 CD11a Mouse SerotecP-87a SDF-1� Rabbit PeproTechmAb 172 CXCR4 Mouse R&D Systems9G11 Human CD31 Mouse R&D SystemsMEC 13.3 Murine CD31 Rat PharMingenmAb 610 Human TNF� Mouse R&D Systems
Primary antibodies (fluorochrome labeled)AHP062F vWF-FITC Mouse SerotecKP-1 (F7135) CD68-FITC Mouse Dako
Secondary antibodiesRabit anti-mouse–biotin Mouse Ig Rabbit (biotinylated) DakoSwine anti-rabbit–biotin Rabbit Ig Swine (biotinylated) DakoRabbit anti-rat–biotin Rat Ig Rabbit (biotinylated) DakoGoat anti-mouse Mouse Ig Goat (FITC) Becton DickinsonGoat anti-rabbit Rabbit Ig Goat (FITC and TRITC) Southern BiotechnologyRabbit anti-mouse Mouse Ig Rabbit (FITC and TRITC) Southern BiotechnologyStreptavidin–alkaline phosphatase kit NA NA Dako
* vWF � von Willebrand factor; ICAM-1 � intercellular adhesion molecule 1; SDF-1� � stromal cell–derived factor 1�; mAb � monoclonalantibody; CXCR4 � CXC receptor 4; TNF� � tumor necrosis factor �; FITC � fluorescein isothiocyanate; TRITC � tetramethylrhodamineisothiocyanate; NA � not applicable.† Kind gift from Dr. D. O. Haskard (London, UK).
826 BLADES ET AL

compartment was scored from 0 to 3 (0 � no stain; 1 � faint;2 � moderate; 3 � strong). For each antibody, 3 sections persample were assessed under blinded conditions by 2 indepen-dent observers. The interobserver variation was �5%.
In vitro studies. U937 cell culture and analysis ofCXCR4 and lymphocyte function–associated antigen 1 (LFA-1)expression by flow cytometry. The U937 human myelomonocyticcell line was cultured in RPM 1640 medium � 10% FCS. Cellswere subcultured and maintained at 0.5–1.0 � 106 cells/ml. Forfluorescence-activated cell sorter (FACS) analysis, U937 cells(0.5 � 106/ml) were resuspended in phosphate buffered saline(PBS). Aliquots (100 �l) were added to a 96-well plate intriplicate for each treatment and incubated on ice. CXCR4 andLFA-1 expression was assessed by addition of specific mono-clonal antibodies (mAb; see Table 1) at their previouslydetermined saturating concentrations. Nonspecific binding wasminimized by the addition of human IgG (16 mg/ml). Plateswere incubated at 4°C for 1 hour. Cells were washed andincubated with FITC-conjugated F(ab�)2 fragments of goatanti-mouse IgG prior to further washing and were analyzed by
flow cytometry (FACScan II analyzer; Becton Dickinson,Oxford, UK).
U937 cell chemotaxis assay. The commercially availableNeuroprobe ChemoTxplate 96-well plate (Receptor Technol-ogies, Adderbury, UK) with polycarbonate membrane filtersand 5-�m membrane pores was utilized for the U937 cellchemotaxis assay. U937 cells were washed and resuspended(4 � 106 cells/ml) in RPMI 1640 � 0.1% bovine serumalbumin. Recombinant human SDF-1 (1 ng/ml or 10 ng/ml;PeproTech, London, UK) or medium was added to the bottomwells (27 �l); the filter was placed on top and 25 �l of the U937cell suspension placed above the membrane. Plates wereincubated for 2 hours in a humidified incubator at 37°C with5% CO2. Cells remaining on top of the filter were absorbedoff, and the surface was washed. The plate was centrifuged(312g, 5 minutes), the filter removed, pelleted cells resus-pended, and 20 �l removed and counted by light microscopy.In some experiments, anti-CXCR4 mAb was preincubated atthe saturating concentration of 20 �g/ml with U937 cells for 30minutes at 37°C, prior to addition of cells to the plate. Data are
Figure 1. Expression of stromal cell–derived factor 1 (SDF-1), CXC receptor 4 (CXCR4), and intercellular adhesion molecule 1 (ICAM-1) inosteoarthritis (OA) and rheumatoid arthritis (RA) synovium. A, Immunohistochemical staining for SDF-1, CXCR4, and ICAM-1. Positive cells areseen in the vasculature (�), lining layer (arrows), sublining cells (S), and perivascular inflammatory infiltrates (L). Increased SDF-1 staining is seenin the RA sublining and perivascular inflammatory aggregates. A similar pattern is seen for CXCR4. ICAM-1 immunoreactivity was higher in RAversus OA synovium in all compartments except the vasculature (original magnification � 200). B, Semiquantitative analysis of SDF-1, CXCR4, andICAM-1 immunoreactivity. The proportion of each tissue constituent positively stained for each antibody was estimated and the sections scored from0 to 4 (see Patients and Methods for details). SDF-1 expression was significantly increased in the RA sublining and perivascular aggregates. Similarresults were obtained for CXCR4. ICAM-1 expression was significantly increased in all RA compartments except the vasculature. � � P � 0.05; ��
� P � 0.01, by Mann-Whitney nonparametric U test. Values are the mean and SEM (n � 10 for each patient group). Color figure can be viewedin the online issue, which is available at http://www.arthritisrheum.org.
SDF-1 AND MONOCYTE LOCALIZATION TO SYNOVIUM 827

reported as the mean � SEM number of cells migrated perwell.
In vivo studies. Tissue transplantation into SCID ani-mals. Beige SCID C.B-17 mice were maintained underpathogen-free conditions in the biology facilities of KingsCollege. Synovial samples were thawed, washed in saline, andkept in saline-moistened sterile gauze over ice until trans-planted. Mice were anesthetized by intraperitoneal injection of0.2 ml Dormitor (0.1 mg/ml; Smith Kline Beecham, Brentford,UK) and 0.1 ml ketamine (0.1 mg/ml, Smith Kline Beecham),an incision made in the dorsal skin behind the ear, and thetissue inserted subcutaneously. The wound was closed withsoluble suture material (Ethicon, Edinburgh, UK). Success ofthe transplantation was assessed 4 weeks postoperatively byimmunohistologic analysis.
Preparation and labeling of U937 cells. U937 cells werewashed and resuspended in serum-free PBS in preparation forlabeling. Cells were incubated with the PKH26 dye (Sigma) ata concentration of 100 �l dye/20 � 10 6 cells for 2 minutes,after which the reaction was stopped with the addition of 10 mlFCS. Cells were then washed twice to remove unbound dye andresuspended in sterile PBS. Viability, determined by trypan bluedye exclusion, was �95%. PKH26 labeling efficiency was con-firmed by UV microscopy before transplantation.
In vivo cell migration assay. PKH26-labeled U937 cells(5 � 106 cells/animal) were injected intravenously in a 100-�lvolume into transplanted SCID mice. SDF-1,TNF� (Genzyme,Haverhill, UK), or saline was injected intragraft, simulta-neously. Forty-eight hours later all animals were killed, and the
grafts were extracted and processed for cryohistologic study.U937 cell migration was assessed histologically by UV micros-copy. Results were expressed as the mean number of cells/high-power field. Approximately 100 high-power fields werecounted per transplant, from 3 cutting levels. Graft viabilitywas assessed macroscopically and by microscopy of toluidineblue–stained sections. Necrotic grafts or those comprisingmurine tissues were excluded from the study.
Assessment and quantification of human and murinevasculature within the grafts. To confirm the conservation ofhuman vasculature following transplantation and to assess celladhesion molecule expression following cytokine/chemokinestimulation of the grafts, we assessed human intercellularadhesion molecule 1 (ICAM-1), von Willebrand factor (vWF),and murine CD31 pre- and posttransplantation, by immuno-histochemistry analysis. The volume fraction (Vv) of immuno-stained human and murine vessels was determined microscop-ically, using a point counting method as previously described(45). Briefly, 2 immunostained sections per transplant wereexamined exhaustively using a �25 objective and rectangular5 � 5 eyepiece graticule.
The fraction of intersections overlying immunostainedvessels was determined for each microscope field, and themean Vv of vessels within the transplant was calculated formurine, human, and combined vascularity.
Statistical analysis. Results are expressed as the mean� SEM unless otherwise indicated. Nonparametric statisticalanalyses were performed using the PC analysis package Sig-maStat 2.0 (Jandel, SPSS UK, Woking, UK). Initially, either
Figure 2. Distribution of SDF-1 and CXCR4 in RA synovial vasculature. SDF-1 and CXCR4 (a and d), analyzed by doubleimmunofluorescence, are strongly expressed in the lining layer (�). Factor VIII–positive vessels (b and e) are also positive forSDF-1 and CXCR4 (c and f). See Figure 1 for definitions. (Original magnification � 200.) Color figure can be viewed in theonline issue, which is available at http://www.arthritisrheum.org.
828 BLADES ET AL
the Kruskal-Wallis nonparametric analysis of variance or 1-wayanalysis of variance was used. Post hoc significance testing wascarried out using Dunn’s multiple comparison test for non-parametric data or Dunnett’s test for parametric data.
RESULTS
Expression of SDF-1 and CXCR4 in RA and OAsynovium. To establish the level of expression of SDF-1protein and its receptor CXCR4 in RA synovium, 10 SM
samples obtained after joint replacement surgery wereanalyzed immunohistologically. The results were com-pared with findings in 10 SM samples from patients withOA. The expression of ICAM-1 was also analyzed, as acomparative positive control. Representative examplesand the results for all samples are shown in Figures 1Aand B, respectively. SDF-1, CXCR4, and ICAM-1 wereall significantly more highly expressed in RA comparedwith OA samples, in the perivascular inflammatory foci
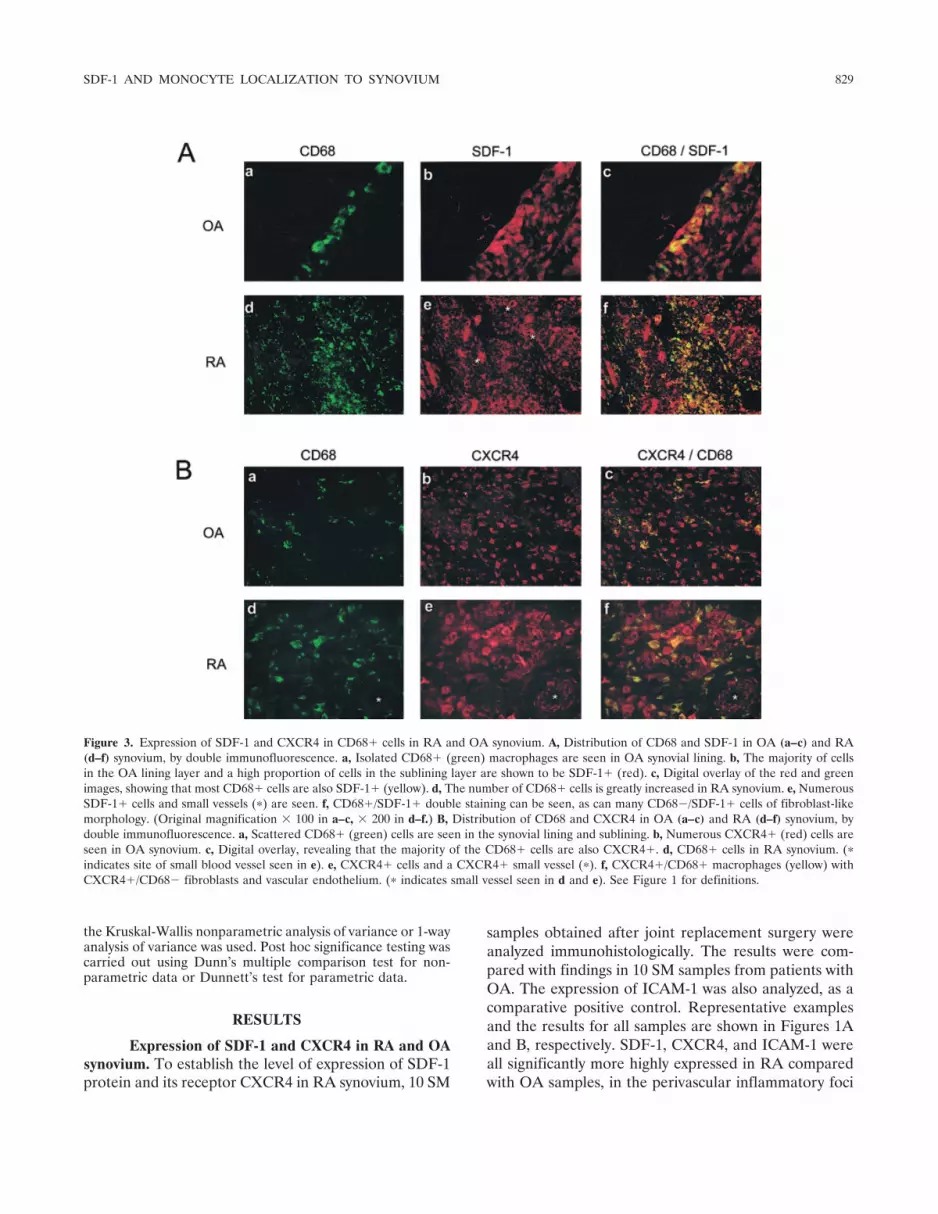
Figure 3. Expression of SDF-1 and CXCR4 in CD68� cells in RA and OA synovium. A, Distribution of CD68 and SDF-1 in OA (a–c) and RA(d–f) synovium, by double immunofluorescence. a, Isolated CD68� (green) macrophages are seen in OA synovial lining. b, The majority of cellsin the OA lining layer and a high proportion of cells in the sublining layer are shown to be SDF-1� (red). c, Digital overlay of the red and greenimages, showing that most CD68� cells are also SDF-1� (yellow). d, The number of CD68� cells is greatly increased in RA synovium. e, NumerousSDF-1� cells and small vessels (�) are seen. f, CD68�/SDF-1� double staining can be seen, as can many CD68�/SDF-1� cells of fibroblast-likemorphology. (Original magnification � 100 in a–c, � 200 in d–f.) B, Distribution of CD68 and CXCR4 in OA (a–c) and RA (d–f) synovium, bydouble immunofluorescence. a, Scattered CD68� (green) cells are seen in the synovial lining and sublining. b, Numerous CXCR4� (red) cells areseen in OA synovium. c, Digital overlay, revealing that the majority of the CD68� cells are also CXCR4�. d, CD68� cells in RA synovium. (�
indicates site of small blood vessel seen in e). e, CXCR4� cells and a CXCR4� small vessel (�). f, CXCR4�/CD68� macrophages (yellow) withCXCR4�/CD68� fibroblasts and vascular endothelium. (� indicates small vessel seen in d and e). See Figure 1 for definitions.
SDF-1 AND MONOCYTE LOCALIZATION TO SYNOVIUM 829
as well as in the diffuse inflammatory infiltrate charac-teristically seen in the sublining layer. When inflamma-tory foci were present in OA samples (3 of 10), theexpression of these antigens was significantly lower thanthat seen in RA. ICAM-1 expression was also signifi-cantly increased in the lining layer of RA SM comparedwith OA SM. In contrast, although the thickness of thelining layer in RA samples was much greater than in OASM, the expression of SDF-1 and CXCR4 in this com-partment was similar in the 2 diseases. In addition,SDF-1 and CXCR4 were strongly expressed in thesynovial vasculature in both RA and OA. A representa-tive example of RA synovium is shown in Figure 2.
Expression of SDF-1 and CXCR4 in CD68�cells. Since strong SDF-1 and CXCR4 immunoreactivitywas seen in cells with macrophage and fibroblast mor-phology, their cellular distribution was further charac-terized by examining their expression on CD68� cells,by double immunofluorescence. As shown in the repre-sentative example in Figure 3, most CD68� cells(�98%) were also positive for both SDF-1 and CXCR4.Aggregates of CD68/SDF-1/CXCR4� cells were seenmore frequently in association with blood vessels withHEV morphology. In addition, clusters of positive cellswere found scattered in the sublining, but more promi-nently in the lining layer. This suggests that SDF-1–CXCR4 interactions may be involved not only in theprocess of CD68� cell extravasation into the joint, butalso in providing directional signals for their final local-ization in the lining layer.
SDF-1–induced U937 cell migration into SMtransplanted into SCID mice. The abundance of SDF-1and CXCR4 in CD68� cells suggested to us that SDF-1plays an important role in the recruitment of monocyte/macrophage cells to the joint. To seek direct functionalevidence in vivo for this hypothesis, we examined thecapacity of SDF-1 to induce the migration of the pro-myelomonocytic U937 cell line into HuSM transplantedinto SCID mice. SDF-1 or TNF� was injected intragraft,while U937 cells were injected intravenously into theanimals.
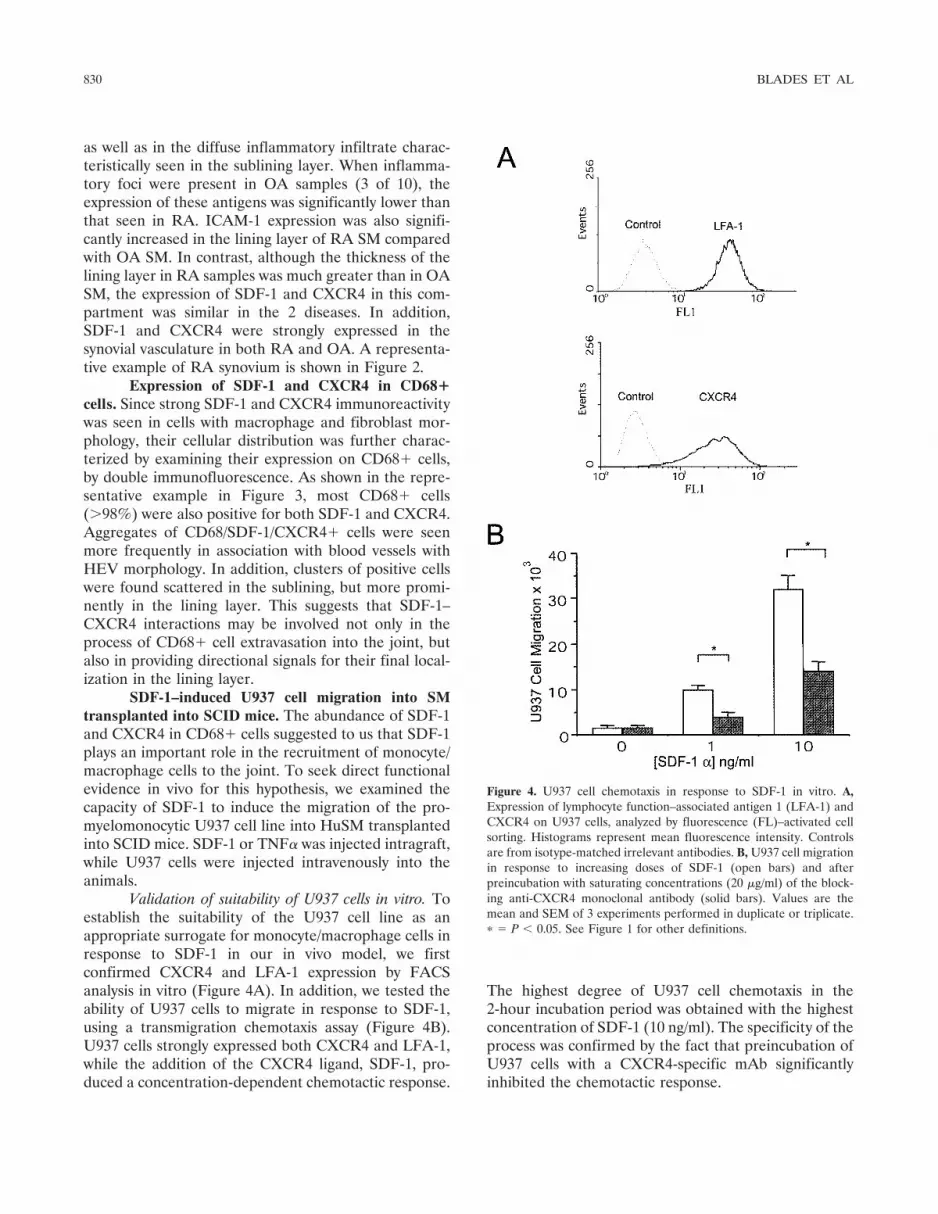
Validation of suitability of U937 cells in vitro. Toestablish the suitability of the U937 cell line as anappropriate surrogate for monocyte/macrophage cells inresponse to SDF-1 in our in vivo model, we firstconfirmed CXCR4 and LFA-1 expression by FACSanalysis in vitro (Figure 4A). In addition, we tested theability of U937 cells to migrate in response to SDF-1,using a transmigration chemotaxis assay (Figure 4B).U937 cells strongly expressed both CXCR4 and LFA-1,while the addition of the CXCR4 ligand, SDF-1, pro-duced a concentration-dependent chemotactic response.
The highest degree of U937 cell chemotaxis in the2-hour incubation period was obtained with the highestconcentration of SDF-1 (10 ng/ml). The specificity of theprocess was confirmed by the fact that preincubation ofU937 cells with a CXCR4-specific mAb significantlyinhibited the chemotactic response.
Figure 4. U937 cell chemotaxis in response to SDF-1 in vitro. A,Expression of lymphocyte function–associated antigen 1 (LFA-1) andCXCR4 on U937 cells, analyzed by fluorescence (FL)–activated cellsorting. Histograms represent mean fluorescence intensity. Controlsare from isotype-matched irrelevant antibodies. B, U937 cell migrationin response to increasing doses of SDF-1 (open bars) and afterpreincubation with saturating concentrations (20 �g/ml) of the block-ing anti-CXCR4 monoclonal antibody (solid bars). Values are themean and SEM of 3 experiments performed in duplicate or triplicate.� � P � 0.05. See Figure 1 for other definitions.
830 BLADES ET AL
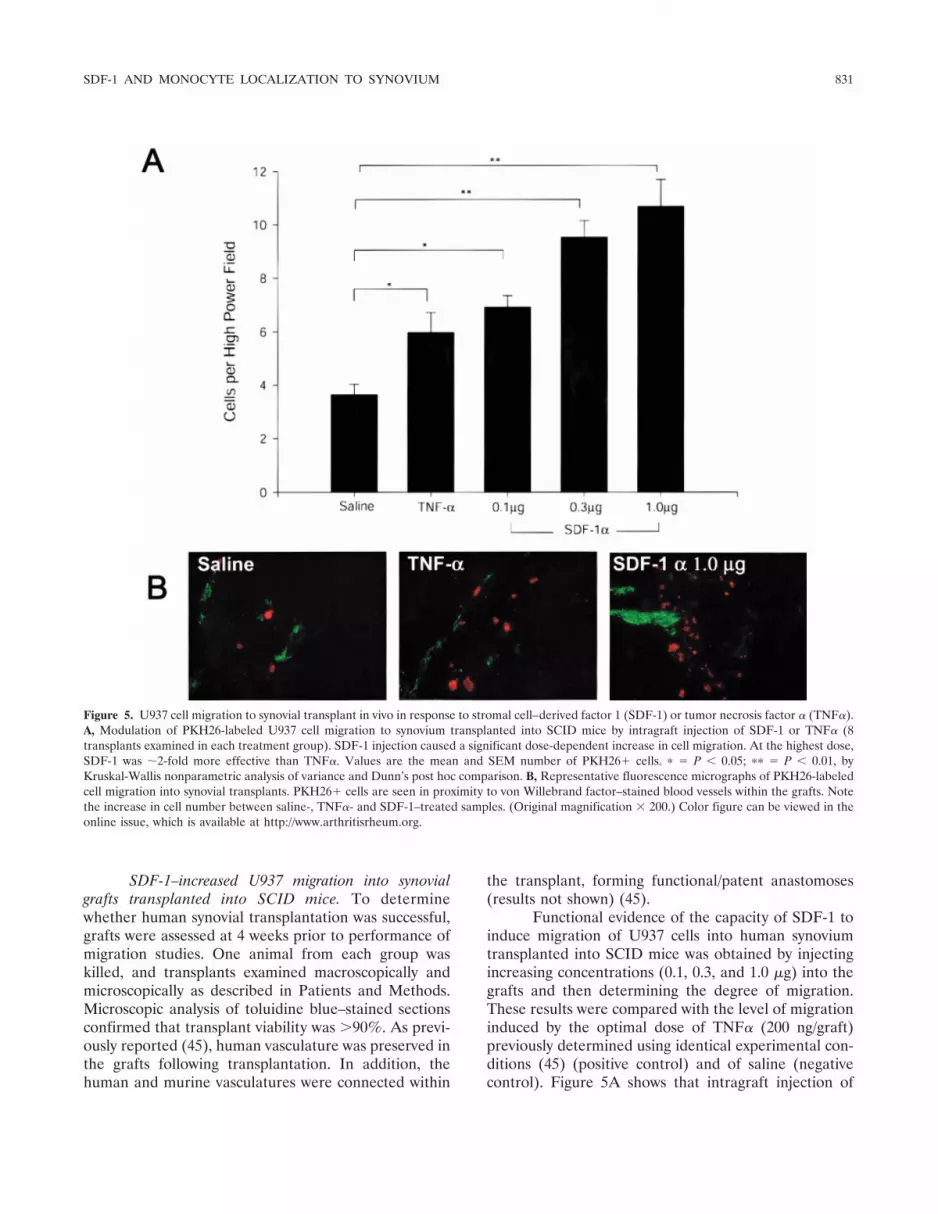
SDF-1–increased U937 migration into synovialgrafts transplanted into SCID mice. To determinewhether human synovial transplantation was successful,grafts were assessed at 4 weeks prior to performance ofmigration studies. One animal from each group waskilled, and transplants examined macroscopically andmicroscopically as described in Patients and Methods.Microscopic analysis of toluidine blue–stained sectionsconfirmed that transplant viability was �90%. As previ-ously reported (45), human vasculature was preserved inthe grafts following transplantation. In addition, thehuman and murine vasculatures were connected within
the transplant, forming functional/patent anastomoses(results not shown) (45).
Functional evidence of the capacity of SDF-1 toinduce migration of U937 cells into human synoviumtransplanted into SCID mice was obtained by injectingincreasing concentrations (0.1, 0.3, and 1.0 �g) into thegrafts and then determining the degree of migration.These results were compared with the level of migrationinduced by the optimal dose of TNF� (200 ng/graft)previously determined using identical experimental con-ditions (45) (positive control) and of saline (negativecontrol). Figure 5A shows that intragraft injection of
Figure 5. U937 cell migration to synovial transplant in vivo in response to stromal cell–derived factor 1 (SDF-1) or tumor necrosis factor � (TNF�).A, Modulation of PKH26-labeled U937 cell migration to synovium transplanted into SCID mice by intragraft injection of SDF-1 or TNF� (8transplants examined in each treatment group). SDF-1 injection caused a significant dose-dependent increase in cell migration. At the highest dose,SDF-1 was 2-fold more effective than TNF�. Values are the mean and SEM number of PKH26� cells. � � P � 0.05; �� � P � 0.01, byKruskal-Wallis nonparametric analysis of variance and Dunn’s post hoc comparison. B, Representative fluorescence micrographs of PKH26-labeledcell migration into synovial transplants. PKH26� cells are seen in proximity to von Willebrand factor–stained blood vessels within the grafts. Notethe increase in cell number between saline-, TNF�- and SDF-1–treated samples. (Original magnification � 200.) Color figure can be viewed in theonline issue, which is available at http://www.arthritisrheum.org.
SDF-1 AND MONOCYTE LOCALIZATION TO SYNOVIUM 831

SDF-1 caused a dose-dependent increase in the migra-tion of U937 cells to the synovial grafts. SDF-1 was moreeffective than TNF� at up-regulating U937 cell migra-tion, causing an increase of 3-fold in the number ofmigrating cells compared with saline-treated animals,and a 2-fold increase over the TNF�-injected group.Figure 5B shows fluorescence micrographs from repre-sentative experiments. It is worth noting that U937 cells(red fluorescence) were mostly found in close relation-ship to graft vessels stained with human specific vWF-related antigen mAb (green fluorescence).
Mechanisms of action of SDF-1 in inducing U937cell migration into SM transplanted into SCID mice. Toinvestigate potential mechanisms of action of SDF-1versus TNF�, we concentrated on 3 areas. First, weexplored whether the effects of SDF-1 were mediatedvia the induction of TNF�, and vice versa. Second, weexamined the variation in ICAM-1 expression and therelationship of this to the degree of migration of U937cells into the grafts. Third, we analyzed the degree ofhuman and mouse vascularity in treated and untreatedtransplants.
Failure of SDF-1 and TNF� to induce reciprocalexpression in synovial grafts. As expected, SDF-1 immu-noreactivity was detected at much higher levels in theSDF-1–injected grafts compared with the saline-injectedcontrols. Similarly, greater TNF�-specific staining wasfound in the TNF�-injected transplants. However,SDF-1 injection did not cause increased expression ofTNF� and, conversely, TNF� injection caused no in-crease in SDF-1 immunoreactivity (Table 2). Thisstrongly indicates that human SDF-1 expression andTNF� expression are regulated in a nonreciprocal man-ner, similar to published findings in mice (49). Inaddition, this suggests that the mechanisms by whichthey increase cell migration into the grafts are indepen-dent from one another.
SDF-1–induced migration into synovial grafts isindependent of ICAM-1 expression. We have previouslydemonstrated that TNF�-induced cell migration intosynovial transplants correlated with increased expression
Figure 6. Modulation of ICAM-1 expression in synovial transplantsby tumor necrosis factor � (TNF�) or SDF-1. A, Representativeexamples of ICAM-1 expression in the original RA tissue and insaline-, TNF� (200 �g)–, and SDF-1 (1 �g)–injected synovial grafts (4weeks posttransplantation). ICAM-1 staining was prominent in thevasculature of the original preimplantation synovium. Following trans-plantation, ICAM-1 was markedly down-regulated. Intragraft injectionof TNF� resulted in a significant increase in vascular ICAM-1 and alsoan increase in diffuse ICAM-1 staining when compared with saline-injected controls. Intragraft injection of SDF-1 did not significantlyincrease either “soluble” or vascular ICAM-1 staining. B, Semiquan-titative assessment of ICAM-1 staining shown in A. The intensity ofICAM-1 staining was arbitrarily scored from 0 (minimal) to 3 (strong)on 3 sections per transplant (8 transplants per group). Analysis wascarried out under blinded conditions by 2 independent observers.Values are the mean and SEM staining intensity. � � P � 0.05, byKruskal-Wallis nonparametric analysis of variance and Dunn’s posthoc comparison. See Figure 1 for other definitions. Color figure can beviewed in the online issue, which is available at http://www.arthritisrheum.org.
Table 2. Immunohistochemical assessment of SDF-1 and TNF�immunoreactivity in cytokine/chemokine-treated synovial transplants*
Treatment SDF-1 TNF�
Control (saline) 0.24 � 0.10 0 � 0TNF�, 200 ng intragraft 1.00 � 0.50 2.5 � 0.29†SDF-1, 1.0 �g intragraft 2.00 � 0.50† 0.7 � 0.58
* Staining intensity was scored arbitrarily on a 0–3 scale, where 0 � nostaining and 3 � strong staining intensity. Values are the mean � SEMimmunohistochemical score from 8 transplants per treatment exam-ined. SDF-1 � stromal cell–derived factor 1; TNF� � tumor necrosisfactor �.† P � 0.05 versus control, by Kruskal-Wallis nonparametric analysis ofvariance and Dunn’s post hoc test.
832 BLADES ET AL

of vascular ICAM-1 and vascular cell adhesion molecule1 (VCAM-1) (45). To investigate whether SDF-1 acts ina similar manner considering that endothelial cells ex-press CXCR4 (refs. 50 and 51 and Figure 2), weexamined ICAM-1 expression in the SDF-1–injectedgrafts as well as in the TNF�- and saline-injectedtransplants. Immunohistochemical analysis showed asignificant increase of ICAM-1 expression in TNF�-injected grafts, confirming our previous results (45), butnot in SDF-1–injected transplants (Figure 6). This sug-gests that the increased U937 migration induced bySDF-1 is independent of ICAM-1 expression and mayoperate through the classic chemokine mechanisms byactivating surface integrins through binding to the spe-cific CXCR4 receptor. Activated integrins would bindmore avidly to endothelial ICAM-1 expressed at basallevels.
SDF-1– and TNF�-induced U937 migration tosynovial transplants does not correlate with the degree ofhuman or murine vascular surface within the grafts. BothSDF-1 (52) and TNF� (53) have been shown to induceneo-angiogenesis; therefore, to determine whether theeffects of SDF-1 and TNF� on U937 cell migration intothe transplant were due to an increase in the vascularbeds feeding the graft, we determined the surface areaof human and mouse vasculature, using species-specificmarkers. Graft vascularity was quantified by determin-ing the volume fraction of immunostained vessels micro-scopically, using a point counting method as described inPatients and Methods. Scatterplots of vascular Vv versusPKH26 migration revealed no discernible pattern ofassociation of the 2 variables. Transplant vascularityshowed no correlation with U937 migration (P � 0.05throughout, by Spearman’s rank correlation) when ei-ther individual transplants or treatment group meanswere compared (data not shown) (45), indicating thatthe degree of migration is largely independent of theextent of graft vascularity.
DISCUSSION
In this study, we investigated the role of SDF-1and its receptor, CXCR4, in the migration of cells of themonocyte/macrophage lineage to the RA synovium.Examining first the general distribution of SDF-1 andCXCR4, we found abundant and much higher expres-sion in RA compared with OA synovium. This wasparticularly prominent in CD68� macrophages in thelining layer, but was also observed in cells in thesublining areas and in endothelial cells. In addition,clusters of CXCR4� cells were found in proximity toSDF-1� vessels.
This suggests that SDF-1 and CXCR4 are in-volved in the process of extravasation of CD68� cellsfrom the circulation into the synovium. Similar mecha-nisms have been postulated for T cells, in reports ofrecent studies that have demonstrated strong expressionof CXCR4 and SDF-1 in T cells in the synovium (43,44).Further support for this concept comes from the obser-vation that SDF-1 associates with heparan sulfates onthe vascular surface (54) and stimulates integrin-mediated arrest on vascular endothelium or purifiedICAM-1 under shear flow (55,56). This is similar tofindings in other chemokines found in RA vessels, suchas MIP-1� and MIP-1�, that express high levels ofheparan sulfate, which is capable of immobilizing thesechemokines on the endothelial surface and increasing Tcell–endothelial cell binding through GTP-mediated sig-naling (57).
Once extravasated in the synovial membrane,monocytes differentiate into tissue macrophages andbegin their journey to their final destination, the lininglayer. From the initial perivascular location, they arefound scattered in the subintima but most abundantly inthe intima, where they represent �80% of the cells. Themechanisms involved in their accumulation in the inti-mal layer are likely to involve chemo- and haptotacticphenomena. The considerable presence of SDF-1 thatwe have demonstrated in the lining layer and the strongexpression of CXCR4 by CD68� cells make thischemokine–chemokine receptor interaction a primecandidate for providing navigational and spatial orien-tation signals necessary for the final positioning of thiscell type within the SM. Even if, by immunohistologicmethods, it is not possible to precisely ascertain whetherSDF-1 is produced by or bound to its receptor CXCR4on the macrophages, the presence of these 2 moleculeson CD68� cells is likely to be of biologic relevance.
However, our data can provide only indirectsupport for this hypothesis. The main problem in obtain-ing direct evidence from patients is that there areobvious technical difficulties and ethical considerationsin performing migration studies in vivo following theinjection of SDF-1. For this reason, to test directly thecapacity of SDF-1 to induce the migration of macro-phage precursors to RA synovium, we have used apreviously validated HuSM-SCID chimera model (45).We used RA or OA tissue interchangeably for trans-plantation since, as previously demonstrated, 4 weeksposttransplantation there is very little histologic differ-ence between the 2 grafted tissues (45). In particular, theinflammatory infiltrate typical of RA has virtually dis-appeared from the grafts. In addition, endothelial celladhesion molecules such as ICAM-1, VCAM-1, and
SDF-1 AND MONOCYTE LOCALIZATION TO SYNOVIUM 833

E-selectin, normally expressed at higher levels in RAversus OA tissue, are down-modulated, with little ex-pression in both RA and OA synovial grafts. However,the inflammatory conditions present in RA synoviumcan be mimicked by intragraft injection of TNF�, whichcan up-regulate the expression of adhesion molecules ineither RA or OA grafts to the levels seen in the RAtissue (45).
Using the HuSM-SCID chimera model, we exam-ined the capacity of SDF-1 injected directly into thesynovial grafts to modulate the migration of U937 cellsto the transplants. This cell line was considered to be theideal surrogate for monocyte/macrophage precursorssince it is a pro-myelomonocytic line, expresses highlevels of CXCR4 (the selective SDF-1 receptor), and hasshown, in vitro, prompt migration in response to SDF-1.
In vivo, SDF-1 also induced a dose-dependentincrease in U937 cell migration into synovial transplants.Moreover, SDF-1 was found to be more effective thanTNF�, causing an 2-fold increase in migration overthat observed in TNF�-injected grafts. Although it ispossible that TNF� may be inducing a pro-apoptoticeffect on U937 cells, this was still a surprising result sinceTNF� is considered to be a very powerful stimulus. Wethen investigated whether SDF-1, in addition to itsintrinsic effect, was acting via induction of TNF�. Asexpected, SDF-1 injection into the grafts caused anincrease in detectable SDF-1- but not TNF�-specificimmunoreactivity compared with saline controls. Simi-larly, intragraft injection of TNF� caused an increase inTNF�-specific immunostaining, but no increase inSDF-1. This is consistent with the finding, in studies ofgene-targeted animals, that SDF-1 production is notinfluenced by either TNF� or lymphotoxin � deletion(49). The injection of single biologically active moleculesinto HuSM grafts allows the observation of downstreameffects and provides robust evidence that, similar tofindings in mice, SDF-1 production and TNF� produc-tion in humans are autonomously regulated. In addition,these data suggest that the mechanisms by which SDF-1and TNF� increase cell migration into the grafts, and byextension in other inflammatory tissues, are indepen-dent from one another.
To investigate this further, we examined thecapacity of SDF-1 and TNF� to modulate ICAM-1expression in the graft synovial microvascular endothe-lium. As previously reported (45), TNF� strongly up-regulated ICAM-1 expression; however, no significantincrease in ICAM-1 expression was detected in theSDF-1–injected grafts. This indicates that the effects ofSDF-1 were mediated separately from the increasedtranscription/mobilization of endothelial surface
ICAM-1 seen following TNF� injection. The lack ofICAM-1 up-regulation was not due to the absence of theSDF-1 receptor on the vascular endothelium. Indeed, inthis study we demonstrated abundant expression ofCXCR4 by the synovial microvasculature, confirmingthe previously reported in vitro findings in culturedendothelial cells (50,51). Therefore, the likely explana-tion for our observations is that in the HuSM-SCIDchimera model, SDF-1 may be acting via the classicchemokine mechanism, namely, the activation of surfaceintegrins (58) of the U937 cells circulating within thegrafts. Activated integrins would then be able to bindmore avidly to endothelial ICAM-1 expressed at basallevels. Indirect evidence for this comes from our in vitroexperiments, which confirmed that, in a transmigrationassay using unstimulated endothelial cells that expressICAM-1 at basal levels, SDF-1 could induce chemotaxisof U937 cells (ref. 59, and Blades MC, et al: unpublishedobservations).
Finally, since both SDF-1 (52) and TNF� (53)have been shown to play a role in angiogenesis, weinvestigated the degree of graft vascularization. This wasdone in order to exclude the possibility that the level ofU937 cell localization to the transplants was influencedby the number of blood vessels feeding the grafts. Wefound no significant difference between treated anduntreated grafts. In addition, there was no correlationbetween the number of cells infiltrating the grafts andthe level of human or mouse vascularity (expressed asthe Vv).
In summary, to our knowledge, this is the firststudy to examine the expression of SDF-1 and CXCR4by CD68� cells in the joint. The physical distributionwithin the synovium strongly supports the concept thatSDF-1–CXCR4 interactions not only facilitate the ex-travasation of monocyte/macrophage cells, but also sup-ply spatial orientation signals to direct them from theperivascular tissue to their final localization in thesynovial intima through the sublining layer. Using theHuSM-SCID chimera model, we obtained functionalevidence that SDF-1 can induce the migration of my-elomonocytic cells into synovial grafts and that thisoperates with different mechanisms from those observedwith TNF�. In addition, we confirmed that SDF-1production and TNF� production are autonomously andindependently regulated. A possible implication of this isthat SDF-1 may play an important role in the RA joint,acting independently from the multiple inflammatorypathways driven by TNF�. This is of particular relevanceconsidering that, although TNF� blockade has beenenormously successful therapeutically, some 40% ofpatients do not respond to this treatment (60). It is
834 BLADES ET AL

possible that sustained production of SDF-1 in the RAjoint, independent of TNF�, may contribute (amongother molecules) to the mechanisms involved in TNF�blockade “resistance.” On this basis, it is also possibleSDF-1 may represent an important target for therapy.Further studies in patients whose disease has beenresistant to anti-TNF� treatment will be needed in orderto clarify these issues.
REFERENCES
1. Pitzalis C, Kingsley G, Lanchbury JS, Murphy J, Panayi GS.Expression of HLA-DR, DQ and DP antigens and interleukin-2receptor on synovial fluid T lymphocyte subsets in rheumatoidarthritis: evidence for “frustrated” activation. J Rheumatol 1987;14:662–6.
2. Pitzalis C, Kingsley G, Murphy J, Panayi G. Abnormal distributionof the helper-inducer and suppressor-inducer T-lymphocyte sub-sets in the rheumatoid joint. Clin Immunol Immunopathol 1987;45:252–8.
3. Weyand CM, Goronzy JJ. The molecular basis of rheumatoidarthritis. J Mol Med 1997;75:772–85.
4. Yanni G, Whelan A, Feighery C, Bresnihan B. Analysis of cellpopulations in rheumatoid arthritis synovial tissues. Semin Arthri-tis Rheum 1992;21:393–9.
5. Firestein GS. Rheumatoid synovitis and pannus. In: Klippel JH,Dieppe PA, editors. Rheumatology. London: Mosby; 1998. p.5.13.1–24.
6. Koch AE. Angiogenesis: implications for rheumatoid arthritis.Arthritis Rheum 1998;41:951–62.
7. Edwards JCW. The synovium. In: Klippel JH, Dieppe PA, editors.Rheumatology. London: Mosby; 1998. p. 1–8.
8. Zvaifler NJ, Firestein GS. Pannus and pannocytes: alternativemodels of joint destruction in rheumatoid arthritis. ArthritisRheum 1994;37:783–9.
9. Yanni G, Whelan A, Feighery C, Bresnihan B. Synovial tissuemacrophages and joint erosion in rheumatoid arthritis. AnnRheum Dis 1994;53:39–44.
10. Edwards JC, Willoughby DA. Demonstration of bone marrowderived cells in synovial lining by means of giant intracellulargranules as genetic markers. Ann Rheum Dis 1982;41:177–82.
11. Athanasou NA. Synovial macrophages. Ann Rheum Dis 1995;54:392–4.
12. Butcher EC. Leukocyte-endothelial cell adhesion as an active,multi-step process: a combinatorial mechanism for specificity anddiversity in leukocyte targeting. Adv Exp Med Biol 1992;323:181–94.
13. Springer TA. Traffic signals for lymphocyte recirculation andleukocyte emigration: the multistep paradigm. Cell 1994;76:301–14.
14. Butcher EC, Picker LJ. Lymphocyte homing and homeostasis.Science 1996;272:60–6.
15. Pitzalis C. Adhesion, migration and cell trafficking. In: FiresteinGS, Panayi GS, Wollheim FA, editors. Rheumatoid arthritis: newfrontiers in pathogenesis and treatment. Oxford: Oxford Univer-sity Press; 2000. p. 137–46.
16. Baggiolini M. Chemokines and leukocyte traffic. Nature 1998;392:565–8.
17. Schall TJ, Bacon KB. Chemokines, leukocyte trafficking, andinflammation. Curr Opin Immunol 1994;6:865–73.
18. Baggiolini M, Dewald B, Moser B. Human chemokines: an update.Annu Rev Immunol 1997;15:675–705.
19. Kelvin DJ, Michiel DF, Johnston JA, Lloyd AR, Sprenger H,Oppenheim JJ, et al. Chemokines and serpentines: the molecularbiology of chemokine receptors. J Leukoc Biol 1993;54:604–12.
20. Neote K, DiGregorio D, Mak JY, Horuk R, Schall TJ. Molecularcloning, functional expression, and signaling characteristics of aC-C chemokine receptor. Cell 1993;72:415–25.
21. Premack BA, Schall TJ. Chemokine receptors: gateways to inflam-mation and infection. Nat Med 1996;2:1174–8.
22. Bargatze RF, Butcher EC. Rapid G protein-regulated activationevent involved in lymphocyte binding to high endothelial venules.J Exp Med 1993;178:367–72.
23. Laudanna C, Campbell JJ, Butcher EC. Role of Rho in chemoat-tractant-activated leukocyte adhesion through integrins. Science1996;271:981–3.
24. Endo H, Akahoshi T, Takagishi K, Kashiwazaki S, Matsushima K.Elevation of interleukin-8 (IL-8) levels in joint fluids of patientswith rheumatoid arthritis and the induction by IL-8 of leukocyteinfiltration and synovitis in rabbit joints. Lymphokine CytokineRes 1991;10:245–52.
25. Koch AE, Kunkel SL, Harlow LA, Johnson B, Evanoff HL, HainesGK, et al. Enhanced production of monocyte chemoattractantprotein-1 in rheumatoid arthritis. J Clin Invest 1992;90:772–9.
26. Koch AE, Kunkel SL, Harlow LA, Mazarakis GK, Burdick MD,Pope RM, et al. Macrophage inflammatory protein-1�: a novelchemotactic cytokine for macrophages in rheumatoid arthritis.J Clin Invest 1994;93:921–8.
27. Kunkel SL, Lukacs N, Kasama T, Strieter RM. The role ofchemokines in inflammatory joint disease. J Leukoc Biol 1996;59:6–12.
28. Robinson E, Keystone EC, Schall TJ, Gillett N, Fish EN. Chemo-kine expression in rheumatoid arthritis (RA): evidence ofRANTES and macrophage inflammatory protein (MIP)-1 betaproduction by synovial T cells. Clin Exp Immunol 1995;101:398–407.
29. Szekanecz Z, Strieter RM, Kunkel SL, Koch AE. Chemokines inrheumatoid arthritis. Springer Semin Immunopathol 1998;20:115–32.
30. Akahoshi T, Endo H, Kondo H, Kashiwazaki S, Kasahara T,Mukaida N, et al. Essential involvement of interleukin-8 in neu-trophil recruitment in rabbits with acute experimental arthritisinduced by lipopolysaccharide and interleukin-1. LymphokineCytokine Res 1994;13:113–6.
31. Barnes DA, Tse J, Kaufhold M, Owen M, Hesselgesser J, StrieterR, et al. Polyclonal antibody directed against human RANTESameliorates disease in the Lewis rat adjuvant-induced arthritismodel. J Clin Invest 1998;101:2910–9.
32. Halloran MM, Woods JM, Strieter RM, Szekanecz Z, Volin MV,Hosaka S, et al. The role of an epithelial neutrophil-activatingpeptide-78-like protein in rat adjuvant-induced arthritis. J Immu-nol 1999;162:7492–500.
33. Ogata H, Takeya M, Yoshimura T, Takagi K, Takahashi K. Therole of monocyte chemoattractant protein-1 (MCP-1) in thepathogenesis of collagen-induced arthritis in rats. J Pathol 1997;182:106–14.
34. Endo H, Akahoshi T, Nishimura A, Tonegawa M, Takagishi K,Kashiwazaki S. Experimental arthritis induced by continuousinfusion of IL-8 into rabbit knee joints. Clin Exp Immunol1994;96:31–5.
35. Zlotnik A, Yoshie O. Chemokines: a new classification system andtheir role in immunity. Immunity 2000;12:121–7.
36. Bleul CC, Farzan M, Choe H, Parolin C, Clark-Lewis I, SodroskiJ, et al. The lymphocyte chemoattractant SDF-1 is a ligand forLESTR/fusin and blocks HIV-1 entry. Nature 1996;382:829–33.
37. McGrath KE, Koniski AD, Maltby KM, McGann JK, Palis J.Embryonic expression and function of the chemokine SDF-1 andits receptor, CXCR4. Dev Biol 1999;213:442–56.
38. Aiuti A, Webb IJ, Bleul C, Springer T, Gutierrez-Ramos JC. Thechemokine SDF-1 is a chemoattractant for human CD34� hema-topoietic progenitor cells and provides a new mechanism toexplain the mobilization of CD34� progenitors to peripheralblood. J Exp Med 1997;185:111–20.
39. Bleul CC, Fuhlbrigge RC, Casasnovas JM, Aiuti A, Springer TA.
SDF-1 AND MONOCYTE LOCALIZATION TO SYNOVIUM 835

A highly efficacious lymphocyte chemoattractant, stromal cell-derived factor 1 (SDF-1) [see comments]. J Exp Med 1996;184:1101–9.
40. Pablos JL, Amara A, Bouloc A, Santiago B, Caruz A, Galindo M,et al. Stromal-cell derived factor is expressed by dendritic cells andendothelium in human skin. Am J Pathol 1999;155:1577–86.
41. Seki T, Selby J, Haupl T, Winchester R. Use of a differentialsubtraction method to identify genes that characterize the pheno-type of cultured rheumatoid arthritis synoviocytes. ArthritisRheum 1998;41:1356–64.
42. Burger JA, Zvaifler NJ, Tsukada N, Firestein GS, Kipps TJ.Fibroblast-like synoviocytes support B-cell pseudoemperipolesisvia a stromal cell-derived factor-1- and CD106 (VCAM-1)-depen-dent mechanism. J Clin Invest 2001;107:305–15.
43. Nanki T, Hayashida K, El Gabalawy HS, Suson S, Shi K, GirschickHJ, et al. Stromal cell-derived factor-1-CXC chemokine receptor 4interactions play a central role in CD4� T cell accumulation inrheumatoid arthritis synovium. J Immunol 2000;165:6590–8.
44. Buckley CD, Amft N, Bradfield PF, Pilling D, Ross E, Arenzana-Seisdedos F, et al. Persistent induction of the chemokine receptorCXCR4 by TGF-�1 on synovial T cells contributes to theiraccumulation within the rheumatoid synovium. J Immunol 2000;165:3423–9.
45. Wahid S, Blades MC, De Lord D, Brown I, Blake G, Yanni G, etal. Tumour necrosis factor � (TNF�) enhances lymphocyte migra-tion into rheumatoid synovial tissue transplanted into severecombined immunodeficient (SCID) mice. Clin Exp Immunol2000;122:133–42.
46. Arnett FC, Edworthy SM, Bloch DA, McShane DJ, Fries JF,Cooper NS, et al. The American Rheumatism Association 1987revised criteria for the classification of rheumatoid arthritis.Arthritis Rheum 1988;31:315–24.
47. Altman R, Asch E, Bloch D, Bole G, Borenstein D, Brandt K, etal. Development of criteria for the classification and reporting ofosteoarthritis: classification of osteoarthritis of the knee. ArthritisRheum 1986;29:1039–49.
48. Pitzalis C, Cauli A, Pipitone N, Smith C, Barker J, Marchesoni A,et al. Cutaneous lymphocyte antigen–positive T lymphocytes pref-erentially migrate to the skin but not to the joint in psoriaticarthritis. Arthritis Rheum 1996;39:137–45.
49. Ngo VN, Korner H, Gunn MD, Schmidt KN, Riminton DS,Cooper MD, et al. Lymphotoxin �/� and tumor necrosis factor arerequired for stromal cell expression of homing chemokines in Band T cell areas of the spleen. J Exp Med 1999;189:403–12.
50. Feil C, Augustin HG. Endothelial cells differentially expressfunctional CXC-chemokine receptor-4 (CXCR-4/fusin) under thecontrol of autocrine activity and exogenous cytokines. BiochemBiophys Res Commun 1998;247:38–45.
51. Volin MV, Joseph L, Shockley MS, Davies PF. Chemokinereceptor CXCR4 expression in endothelium. Biochem BiophysRes Commun 1998;242:46–53.
52. Mirshahi F, Pourtau J, Li H, Muraine M, Trochon V, Legrand E,et al. SDF-1 activity on microvascular endothelial cells: conse-quences on angiogenesis in in vitro and in vivo models. ThrombRes 2000;99:587–94.
53. Paleolog EM, Young S, Stark AC, McCloskey RV, Feldmann M,Maini RN. Modulation of angiogenic vascular endothelial growthfactor by tumor necrosis factor � and interleukin-1 in rheumatoidarthritis. Arthritis Rheum 1998;41:1258–65.
54. Amara A, Lorthioir O, Valenzuela A, Magerus A, Thelen M,Montes M, et al. Stromal cell-derived factor-1� associates withheparan sulfates through the first �-strand of the chemokine.J Biol Chem 1999;274:23916–25.
55. Peled A, Grabovsky V, Habler L, Sandbank J, Arenzana-Seis-dedos F, Petit I, et al. The chemokine SDF-1 stimulates integrin-mediated arrest of CD34(�) cells on vascular endothelium undershear flow. J Clin Invest 1999;104:1199–211.
56. Campbell JJ, Hedrick J, Zlotnik A, Siani MA, Thompson DA,Butcher EC. Chemokines and the arrest of lymphocytes rollingunder flow conditions. Science 1998;279:381–4.
57. Tanaka Y, Fujii K, Hubscher S, Aso M, Takazawa A, Saito K, etal. Heparan sulfate proteoglycan on endothelium efficiently in-duces integrin-mediated T cell adhesion by immobilizing chemo-kines in patients with rheumatoid synovitis. Arthritis Rheum1998;41:1365–77.
58. Peled A, Kollet O, Ponomaryov T, Petit I, Franitza S, GrabovskyV, et al. The chemokine SDF-1 activates the integrins LFA-1,VLA-4, and VLA-5 on immature human CD34(�) cells: role intransendothelial/stromal migration and engraftment of NOD/SCID mice. Blood 2000;95:3289–96.
59. Wheller SK, Perretti M. Dexamethasone inhibits cytokine-inducedintercellular adhesion molecule-1 up-regulation on endothelial celllines. Eur J Pharmacol 1997;331:65–71.
60. Lipsky PE, van der Heijde DMFM, St. Clair EW, Furst DE,Breedveld FC, Kalden JR, et al, for the Anti–Tumor NecrosisFactor Trial in Rheumatoid Arthritis with Concomitant TherapyStudy Group. Infliximab and methotrexate in the treatment ofrheumatoid arthritis. N Engl J Med 2000;343:1594–602.
836 BLADES ET AL





























