Embed Size (px)
Citation preview

Cell, Vol. 18, 1145-l 151, December 1979, Copyright 0 1979 by MIT
Specificity of the Bacteriophage Lambda N Gene Product (p/V): Nut Sequences Are Necessary and Sufficient for Antitermination by pN
Benoit de Crombrugghe, Maria Mudryj, Roberto DiLauro* and Max Gottesman Laboratory of Molecular Biology National Cancer Institute National Institutes of Health Bethesda, Maryland 20205
Summary
We have cloned the nut* site together with the fR, site of bacteriophage lambda in the E. toll galactose operon to examine whether the X promoter s8- quences PR and PL are involved in the recognition specificity of the X N gene product (pN). We first constructed a derivative of plasmld pBR322 in which the expression of the tetracycline genes (tet) is controlled by the gel promoter (P,.,). This new plasmid contains a unique Hind III site between Pa., and tet into which the nutR and fR1 sites were intro- duced. The order of the relevant genetic markers in this second plasmid is Po.,-nut&&et. Ceils trans- formed with this plasmid express tet only if pN is provided and if the plasmid contains an intact gal promoter. Our data suggest that transcription which originates at Ppal is modified by pN at nufR, enabling it to pass through fR1 into tet. We conclude that promoters do not play a specific role in pN recog- nition and that nut sequences are both necessary and sufficient for p N action.
Introduction
The bacteriophage lambda N gene product (pN) con- trols lambda gene expression by suppressing tran- scription termination (Roberts, 1969; Luzzati, 1970; Lozeron, Dahlberg and Szybalski, 1976). In the pres- ence of pN, transcription originating at the phage promoters, pL and pR, can extend beyond the termi- nators tLI and tRt and into the lambda chromosome. In induced lysogens, the pL transcript can also pass the bacterial-prophage junction and continue into the neighboring E. coli galactose (gal) operon (Adhya. Gottesman and de Crombrugghe, 1974). While pN suppressed the termination of transcripts from pL or pR, it fails to act on transcripts from other promoters such as lambda pc17 (Friedman, Wilgus and Mural, 1973b), the promoter of the tryptophan operon (Franklin, 1974) or the gal promoter (Adhya et al., 1974).
The specificity of pN involves nucleotide sequences on the phage chromosome which are not present in gal or in other operons. A phage sequence, nutL (for N utilization site), which lies near pL, has been impli- cated in pN-induced antitermination; when nutL is
l Present address: lstituto di Patologia Generale. II Faculta di Medi- cina y Chirurg’ia, Napoli, Italia.
mutant, pL transcripts always terminate at tLl (Salstrom and Szybalski, 1976). A sequence of 16 bp, 17 of which are identical to nutL, is located some 250 bp distal to pR and about 50 bp before tR1 (Rosenberg et al., 1976); mutations in nuk, are not yet available. It has been proposed that nut sequences are the sites at which pN modifies the transcription machinery, making it resistant to termination (Friedman, Jolly and Mural, 1973a; Adhya et al., 1974).
Are the nut sequences sufficient for the action of pN? If so, transcription originating at any promoter could respond to pN, provided a nut sequence was located between that promoter and a terminator. Al- ternatively, sequences at pL or pR might also be in- volved in the recognition of p N, and only transcription initiated at these promoters would be anti-termi- nated.We have distinguished between these possibil- ities by cloning nutR into the gal operon. We now find that gal-promoted transcription will cross a termination signal when pN is provided. We conclude that the nut sequence is both necessary and sufficient for the action of pN.
Results
Construction of pBdC1, a Vector for Termination Sequences We first constructed a plasmid derived from pBR322 (Bolivar et al., 1977) in which the promoter for the tetracycline resistance genes (tet) was replaced by the promoters of the galactose operon of E. coli. We have previously shown that the expression of the gal genes is controlled by two overlapping promoters that are differentially affected by cyclic AMP (CAMP) (Muss0 et al., 1977). The experiments described in this study concern only the CAMP-stimulated Pl pro- moter.
When the DNA of the transducing X phage pgal6 (Feiss, Adhya and Court, 1972) is digested with both Eco Rl and Hind Ill endonucleases, a fragment of approximately 1000 bp, which contains the gal oper- ator (0) and promoters (Pl and P2), is generated. The fragment is bounded on one side by the Eco Rl cleavage site between the galOP region and the bac- teriai-prophage junction and on the other by the Hind Ill site in the promoter-proximal part of the first gal cistron, ga/E (see Figure 1). The DNA of pBR322 contains a unique Hind Ill site, which lies within the promoter for tet, and a unique Eco Rl site, some 25 bp from the Hind Ill site (Bolivar et al., 1977; Rodriguez et al., 1979). A recombinant plasmid, pBdC1, carrying the ga/OP fragment in place of the 25 bp Eco Rl -Hind Ill fragment of pBR322, was isolated as follows.
After Xpgal6 and pBR322 DNAs were digested by Eco Rl and Hind Ill endonucleases and fragments were joined randomly with T4 DNA ligase, C600 cells were transformed with the ligated mixture and the

Cell 1146
Figure 1. Construction of pBdC1
Letters on upper line indicate reference genes in X gal.
cells were plated on minimal medium containing methyl-/3-D-thiogalactoside (TMG), galactose and ampicillin (50pg/ml). This selects for cells constitutive for gal operon enzymes, since TMG is an anti-inducer of gal, and blocks the growth of wild-type E. coli when galactose is the sole carbon source (Buttin, 1963a; Nakanishi et al., 1973). We predicted that cells har- boring a many-copy plasmid carrying the gal operator would grow in TMG-galactose as a result of titration of the cellular gal repressor (Yarmolinsky and Wies- mayer, 1960; Buttin, 1963b).
Several additional lines of evidence indicate that pBdC1 contains the 1000 bp ga/OP fragment. Cleav- age of pBdC1 by EGO Rl and Hind Ill yields a 1000 bp fragment and a larger fragment corresponding in size to pBR322. The sizes of the fragments obtained by digestion of pBdC1 with Hinf I (see Figure 41, Hap II and Hha I are all consistent with the results of a previous restriction enzyme analysis on a 900 bp Hae III fragment from Apgal8 (Muss0 et al., 1977). Finally, several DNA fragments covering the ga/OP region can conveniently be prepared from pBdC1. They have been used for functional and structural studies of gal, including DNA sequencing, directly demonstrating the presence of galOP region in pBdC1 (data not shown) (Taniguchi, O’Neill and de Crombrugghe, 1979).
Cells containing pBdC1 are completely resistant to 10 pg/ml tetracycline (Figure 2), grow slowly in me- dium containing 25 pg/ml tetracycline and are sensi- tive to a concentration of 50 pg/ml of the antibiotic. This level of tetracycline resistance is less than for cells harboring the parent plasmid, pBR322, which are resistant to 50 pg/ml tetracycline.
The expression of the ret genes in pBdC1 could derive from the gal promoter, which is appropriately oriented for tet transcription (Figure 1). Alternatively, a new promoter might have been created at the boundary between the parent plasmid and the inserted DNA fragment. To distinguish between these possibil- ities, we constructed plasmid pBdC5, which is identi- cal to pBdC1 except that it contains the gal promoter mutation, 8-P3. The 8-P3 mutation results from an A/
T- to -T/A transition in the second base of the Pribnow heptamer corresponding to the Pl gal promoter (Musso et al., 1977). The pBdC5 plasmid was ob- tained with the same selection as described above for pBdC1, using a digest of Xpga18-P3 DNA, and is indistinguishable from pBdC1 by restriction analysis (data not shown).
As shown in Figure 2, cells harboring pBdC5 are much less resistant to tetracycline than cells carrying the gal+ plasmid, pBdC1. We conclude that the tet- racycline resistance conferred by pBdC1 results from transcription of the ret genes from the gal promoter.
In pBdC1, the gal promoter is separated from the ret structural genes by a DNA segment carrying a unique Hind Ill site. The plasmid thus constitutes a vector for the cloning of termination sequences and for studying their effect on ret expression.
Introduction of the Lambda fUutR and tRl Sites into pBdC1 The structure of pBdC1 made it possible for us to introduce the lambda nuts site, as well as the lambda terminator fs,, between the gal promoter and ret. We could then ask whether expression of fet from the gal promoter was affected by lambda N gene product.
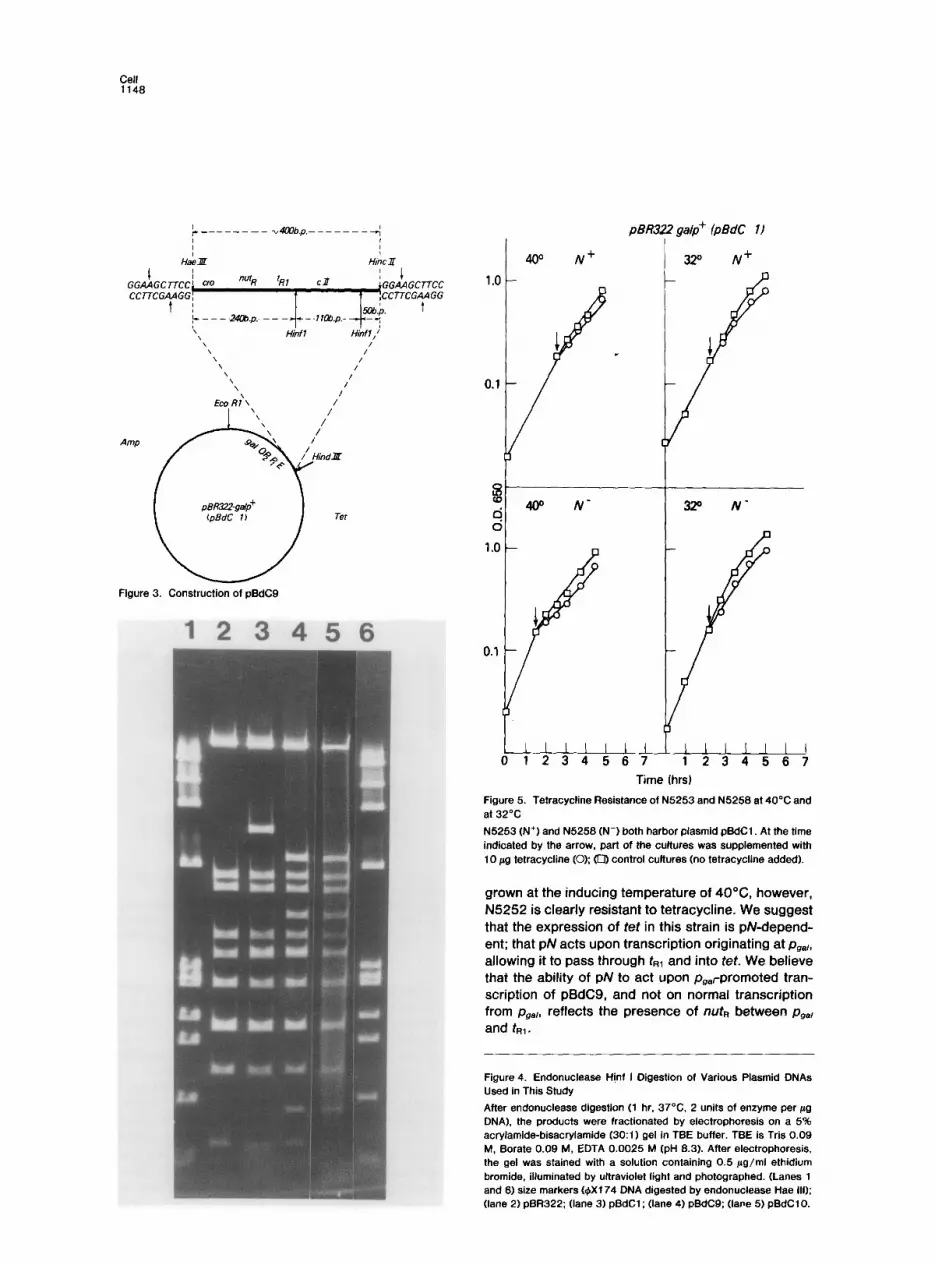
A 397 bp lambda DNA fragment bearing the nute and fR1 sites was inserted at the Hind Ill site of pBdC1. The fragment extends from nucleotide 129 to nucleo- tide 526 with reference to the start of the PR transcript, and contains the carboxyl terminus of gene cro, nutR, fs, and the amino terminal portion of gene c/l (Figure 3; Rosenberg et al., 1978). The 397 bp fragment was a Hint II cleavage product of a -1100 bp fragment obtained by endonuclease Hae Ill digestion of lambda DNA. Hint II cleavage of this 1100 bp fragment yielded flush-ended fragments 397 and -700 bp in length (data not shown), which were ligated directly to syn- thetic deoxynucleotide decamers containing the Hind Ill recognition site. The fragment mixture was then digested with Hind Ill and introduced into the Hind Ill site of pBdC1. The ligated mixture was used to trans- form C600, selecting for ampicillin resistance, and the transformed clones were analyzed by marker rescue for cl 7, ,as described in Experimental Procedures. Those clones which produced cl 7+ recombinants must contain the 397 bp fragment.
To determine the orientation of the lambda fragment with respect to the gal promoter and ret, the recom- binant plasmids were analyzed by digestion with Hinf I endonuclease. The plasmids are expected to yield three new Hinf I fragments and to lose the 740 bp fragment containing the Hind Ill site of pBdC1. The length of the new Hinf I fragments indicates the ori- entation of the 397 bp nuf~ insert. Size measurements of the Hinf I digestion products demonstrate that pBdC9 yields a 340 bp fragment which represents a fusion of a portion of galE and the left part of the 397 bp IX.& fragment (Figure 4). a 100 bp fragment which
Nut Sequences are Necessary and Sufficient for PN Action 1147
Table 1. E. coli Strains
Strain Structure
MR93 ga/E
G104 MRS3/pBdCl
GIOS MR93/pBdC5
N5242 MR93gal’A8 (Acl857ABAMAHl)
N5243 MR93gal+A8 (hc1857N7, 53ABAMAHl)
N5252 N5242/pBdCQ
N5257 N5243/pBdCQ
N5253 N5242/pBdCl
N5258 N5243/pBdCl
G149 N5242/pBdClO
G150 N5243/pBdClO
l AP = Ampicillin resistance.
Source Selection
MR93 + pBdC1 AP’
MR93 + pBdC5 AP ’
MR93 + PI N4830 Gal+, immh. Bio-
MR93 + PI N4831 Gal+, immh, Bio-
N5242 + pBdC9 AP’
N5243 + pBdC9 AP*
N5242 + pBdC1 AP’
N5243 + pBdCl AP’
N5242 + pBdCl0 AP l
N5243 + pBdCl0 AP.
is internal to the 397 bp nutR fragment and a 650 bp fragment which is a fusion of the right part of the nutR fragment with the tet gene of pBR322. Hence the relevant genetic elements of pBdC9 are in the follow- ing order: galp-nut,&,-tet.
A similar plasmid, pBdC10, was constructed, in which the 397 bp nutR fragment was inserted in the Hind Ill side of pBdC5, which is mutant in the gal promoter. The data presented in Figure 4, lanes 4 and 5, show that pBdC9 and pBdC10 are indistinguishable by restriction enzyme analysis, thus indicating that both plasmids contain the 397 bp nutR fragment in the same orientation.
pN Suppresses Termination of Transcription Initiated at the gal Promoter Plasmids pBdC1, BdC5 and BdC9 were introduced into two E. coli strains which are lysogenic for lambda. Strain N5242 carries X cl857ABamAH1; strain N5243 carries the Nam7,53 derivative of the same prophage. The use of these prophage offers several advantages. First. the cl857 mutation produces a thermolabile lambda repressor. Lysogens grown at 32’C are re- pressed; prophage induction occurs at 42°C. Second, ABam and AH1 remove all phage functions lethal to E. coli. The lysogens can therefore be grown indefi- nitely at the inducing temperature. Third, AH1 elim- inates the lambda cro gene, which limits gene N expression. Cultures of N5242, grown at 42”C, pro- duce high levels of pN and no other known phage product.
The analysis of plasmid-bearing derivatives of strains N5242 and N5243 is presented in Figures 5, 6 and 7.
Strains carrying plasmid pBdC1 are expected to be resistant to tetracycline; no terminator lies between pgal and the tet structural genes. The gal promoter is not subject to repression when present on a plasmid, since the levels of gal repressor are insufficient to
I I I I I I I I I 1 2 3 4 1 2 3 4
9 1 3
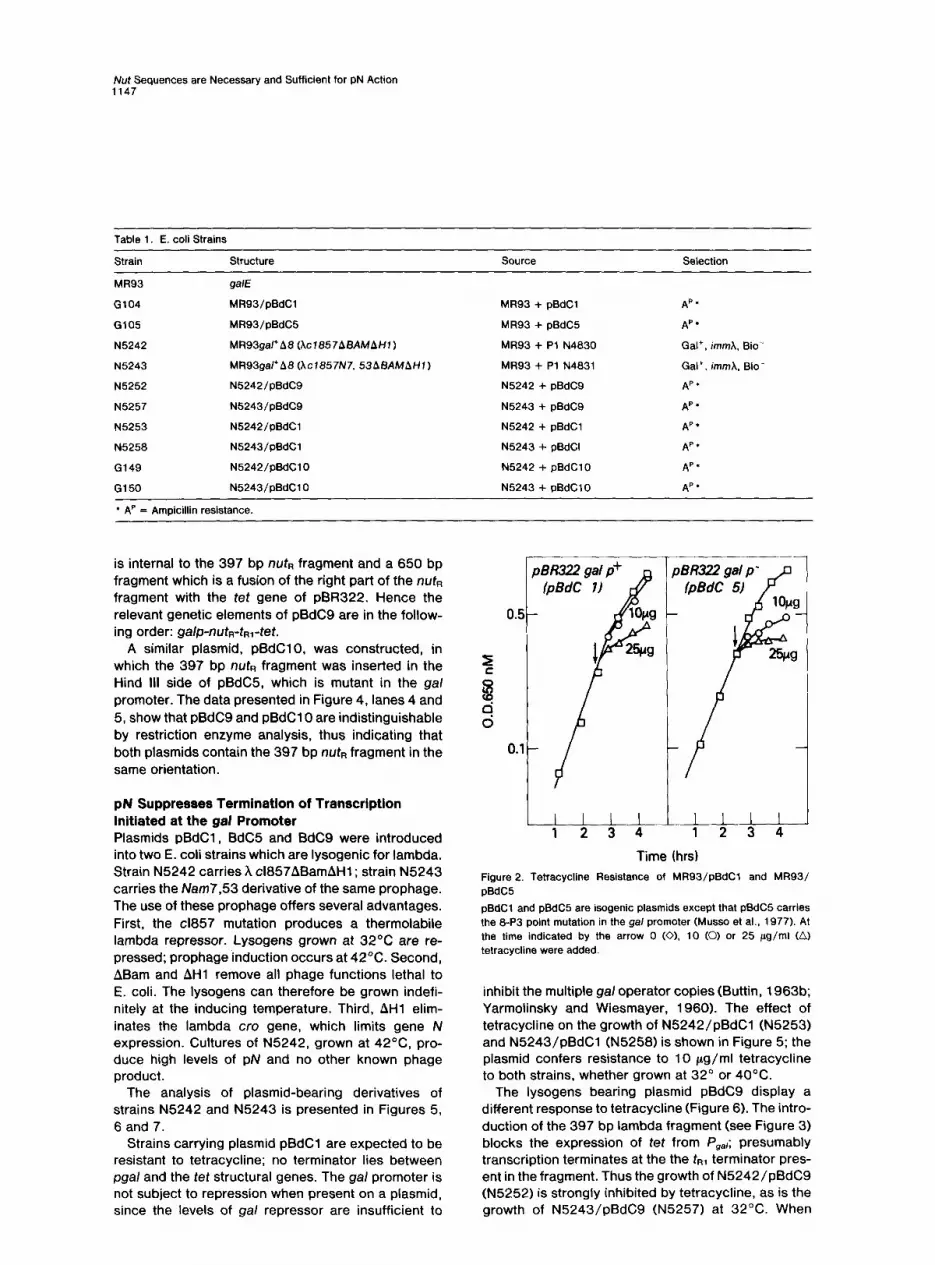
Time (hrs) Figure 2. Tetracycline Resistance of MR93/pBdCl and MR93/ pBdC5
pBdC1 and pBdC5 are isogenic plasmids except that pBdC5 carries the 8-P3 point mutation in the gal promoter (Musso et al., 1977). At the time indicated by the arrow 0 (0). 10 (0) or 25 pgg/ml (A) tetracycline were added.
inhibit the multiple gal operator copies (Buttin, 1963b; Yarmolinsky and Wiesmayer, 1960). The effect of tetracycline on the growth of N5242/pBdCl (N5253) and N5243/pBdCl (N5258) is shown in Figure 5; the plasmid confers resistance to 10 pg/ml tetracycline to both strains, whether grown at 32’ or 40°C.
The lysogens bearing plasmid pBdC9 display a different response to tetracycline (Figure 6). The intro- duction of the 397 bp lambda fragment (see Figure 3) blocks the expression of tet from P,,,; presumably transcription terminates at the the t,=,, terminator pres- ent in the fragment. Thus the growth of N5242/pBdC9 (N5252) is strongly inhibited by tetracycline, as is the growth of N5243/pBdC9 (N5257) at 32°C. When
Cell 1148
c--------~4wb.~.-------~ I , I I
H&U Him I
““tR 571 cx I 1 GGAAGCTTCC
t t LCGAAGG
‘\ \ Hhf 1 Hihfl,i
\ \ /I
\ \ If \
Eco R;\ I \
1’ //
paRm-g8lp+ IpBdC 71
Figure 3. Constructlon of pBdC9
0.1
pBR3 ! galp+ lpBdC 1)
40’= N+ 32O N+
1 1 I I I I I I I I I I I I I 01234567 1234567
320 N-
Time (hrs) Figure 5. Tetracycline Resistance of N5253 and N5258 at 40°C and at 32OC
N5253 (N+) and N5258 (N-I both harbor plasmid pBdC1. At the time indicated by the arrow, part of the cultures was supplemented with 10 Fg tetracycline (0): (iI control cultures (no tetracycline added).
grown at the inducing temperature of 4O”C, however, N5252 is clearly resistant to tetracycline. We suggest that the expression of tet in this strain is pN-depend- ent; that pN acts upon transcription originating at pse,, allowing it to pass through fR1 and into fet. We believe that the ability of pN to act upon p,rpromoted tran- scription of pBdC9, and not on normal transcription from pget, reflects the presence of nutR between pse, and tR1.
Figure 4. Endonuclease Hjnf I Digestion of Various Plasmid ONAs Used in This Study
After endonuclease digestion (1 hr. 37°C. 2 units of enzyme per Fg DNA), the products were fractionated by electrophoresis on a 5% acrylamide-bisacrylamide (3O:l) gel in TEE buffer. TBE is Tris 0.09 M, Borate 0.09 M, EDTA 0.0025 M (pH 8.3). After electrophoresis. the gel was stained with a solution containing 0.5 j.rg/ml ethidium bromide, illuminated by ultraviolet light and photographed. (Lanes 1 and 6) size markers @Xl 74 DNA digested by endonuclease Hae Ill): (lane 2) pER322; (lane 3) pBdC1; (lane 4) pBdC9; (lane 5) pBdCl0.
Nut Sequences are Necessary and Sufficient for pN Action 1149
1.0
0.1
Q ci 6
1.0
0.1
pBR32;
40° N+
400 N’ 320 N’
f
rslp+notR+ @BdC 91
I-L.1 1234567
Time fhrs)
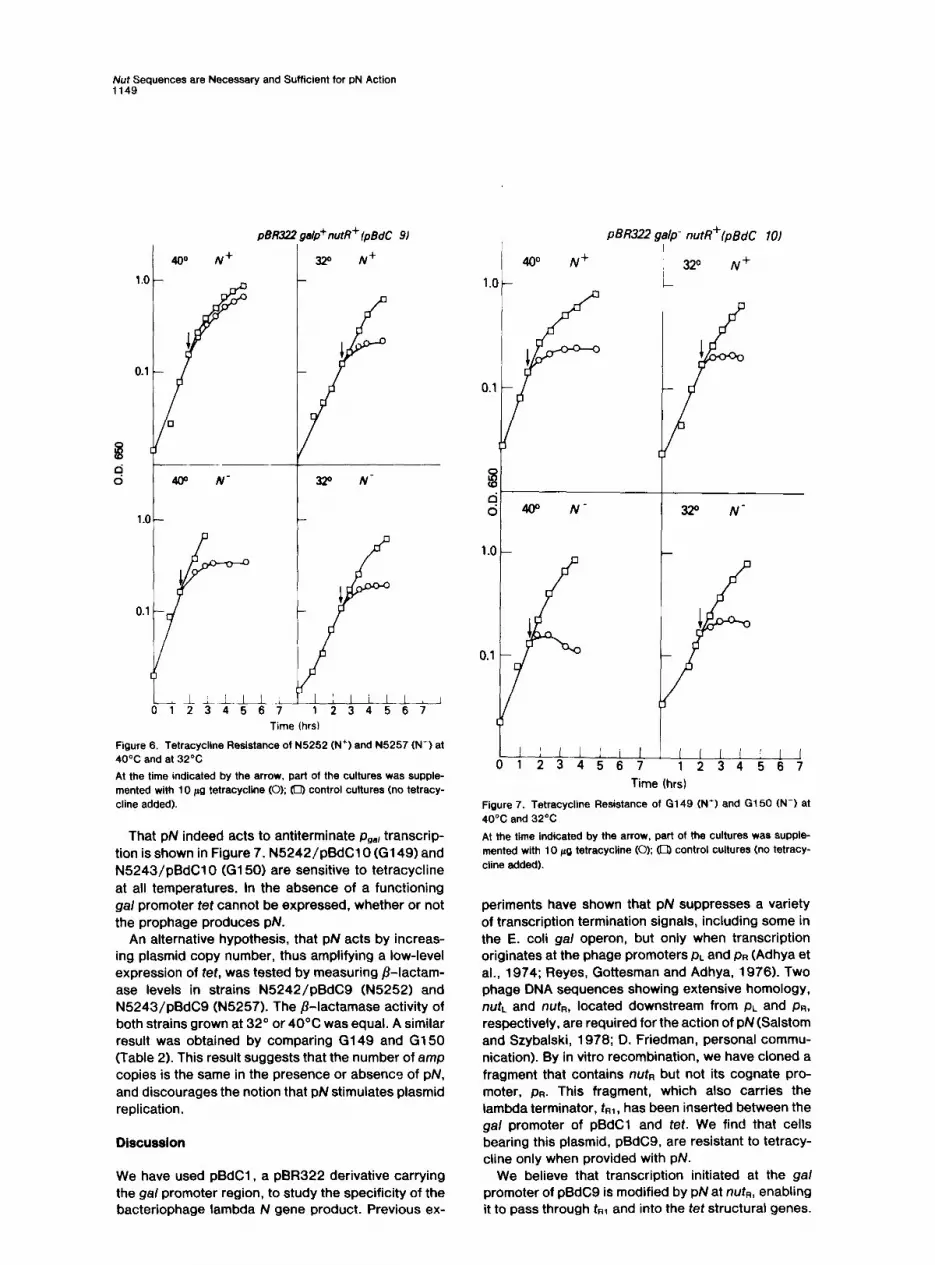
Figure 6. Tetracycline Resistance of N5252 (N+) and N6257 (N-1 at 40% and at 32’C
At the time indicated by the arrow, part of the cultures was supple- mented with 10 Ag tetracycline (0); 0 control cultures (no tetracy- cline added).
That pN indeed acts to antiterminate pge, transcrip- tion is shown in Figure 7. N5242/pBdClO (G149) and N5243/pBdClO (G150) are sensitive to tetracycline at all temperatures. In the absence of a functioning gal promoter tet cannot be expressed, whether or not the prophage produces pN.
An alternative hypothesis, that pN acts by increas- ing plasmid copy number, thus amplifying a low-level expression of tet, was tested by measuring /3-lactam- ase levels in strains N5242/pBdC9 (N5252) and N5243/pBdC9 (N5257). The /I-lactamase activity of both strains grown at 32’ or 40% was equal. A similar result was obtained by comparing G149 and G150 (Table 2). This result suggests that the number of amp copies is the same in the presence or absence of pN, and discourages the notion that pN stimulates plasmid replication.
Discussion
We have used pBdC1, a pBR322 derivative carrying the gal promoter region, to study the specificity of the bacteriophage lambda N gene product. Previous ex-
pBR.322 !
40= N+ 1.0 -
lp- nutR+lpBdC 101
32O N+ -
320 N-
,p’ I
I I I I I I [ 1234567
Time (hrs)
Figure 7. Tetracycline Resistance of G149 (N’) and G150 (N-I at 4O’C and 32°C
At the time indicated by the arrow, part of the cultures was Supple- mented with 10 pg tetracycline (0); (0) control cultures (no tetracy- cline added).
periments have shown that pN suppresses a variety of transcription termination signals, including some in the E. coli gal operon, but only when transcription originates at the phage promoters pL and pR (Adhya et al., 1974; Reyes, Gottesman and Adhya, 1976). Two phage DNA sequences showing extensive homology, nutL and nr.&, located downstream from pL and pR, respectively, are required for the action of pN (Salstom and Szybalski, 1978; D. Friedman, personal commu- nication). By in vitro recombination, we have cloned a fragment that contains nutk but not its cognate pro- moter, pA. This fragment, which also carries the lambda terminator, fR1, has been inserted between the gal promoter of pBdC1 and ret. We find that cells bearing this plasmid, pBdC9, are resistant to tetracy- cline only when provided with pN.
We believe that transcription initiated at the gal promoter of pBdC9 is modified by pN at nub, enabling it to pass through fR1 and into the tet structural genes.

Cell 1150
Table 2. Absence of pN Effect on b-Lactamase Levels
f9-Lactamase (Units)
Experiment Number 32°C 40°c
1 N5242/pBdC9 (N’) 137 392
N5243/pBdC9 (N-) 170 410
2 N5242/pBdCfO (N+) 202 364
N5243/pBdCtO (N-I 262 376
Assays for /3-lactamase were performed as described in Experimental Procedures.
That p,.,-promoted transcription is required for ret expression is shown by the fact that a derivative of pBdC9, pBdC10, mutant in the gal promoter, is sen- sitive to tetracycline both in the presence and absence of pN. Although we have not yet constructed plasmids mutant in nutR or fR1 to confirm that these elements function in the manner ascribed, we believe that these results strongly suggest that promoters do not play a specific role in the recognition of pN. It appears that the nut sequences provide the information that is both necessary and sufficient for the interaction between pN and the transcription complex.
We have described the construction of plasmids which may be quite useful as vectors for the cloning of DNA fragments. DNA fragments, carrying structural gene sequences but no promoters, could be ex- pressed under the control of the gal promoter of pBdC1. Termination signals can also be cloned in pBdC1; they would eliminate ret expression. Muta- tions which restored the ability of these plasmids to confer tetracycline resistance could be readily se- lected.
Plasmid pBdC9, which carries nutR, may be useful in cloning fragments in which terminators block the expression of structural gene sequences. Cells pro- viding pN to these plasmids could suppress transcrip- tion termination and activate the inhibited genes. Fi- nally, in cases where the cloned sequences encode functions lethal to E. coli, the expression of these functions can be turned on or off by induction of a lambda N+ prophage. The possibility then exists of isolating mutations affecting the expression or the lethality of these functions.
The site specificity of other bacteriophage lambda regulatory proteins like pQ could be studied using methods similar to those described here-that is, by cloning in pBdC1 the presumed recognition sites for these factors without their physiological cognate pro- moters.
Experimental Procedures
Enzymes Restriction endonucleases were purchased from New England Bio- labs and Bethesda Research Laboratory. Reactions were performed in 100 mM Tris-Cl (pfi 7.5), 50 mM NaCI. 10 mM MgCl* for endo-
nucleases Eco Rl , Hind Ill and Hint II and in 6 mM Tris-Cl (PH 7.9), 6 mM MgC12. 6 mM /7-mercaptoethanol for endonuclease Hinf I. T4 DNA ligase came from New England Biolabs. Ligase reactions were as described by Ullrich et al. (1977).
Transformation E. coli were grown in L broth to an optical dens&m of -1, centrifuged and resuspended in four tenths of the original volume of 50 mM Car&, recentrifuged and resuspended in one tenth the original volume of 50 mM Car& at 0-2°C. DNA was added and the mixture was left at 0°C for 25 min. The reaction was brought to 37’C for 2 min. then left for 10 min at room temperature. Cells were plated on LB plates containing 50 pg/ml ampicillin or on TMG (10 mM)galactose (0.2%)- M56 (Reyes et al., 1976) plates containing 50 pg/ml ampicillin (Ap).
Marker Rescue Experiment Overnight bacterial cultures grown in tryptone broth-O.01 M MgSO.,- 0.2% maltose-50 r.rg/ml ampicillin were centrifuged and resuspended in Tris-HCI, 0.01 M (pH 7.4). 0.01 M MgSO, and then infected with h cl9Ocl7 at an moi of -5. After absorption for 10 min. the infected cells were subjected to brief ultraviolet irradiation (400 ergs/mm’) and then plated on the nusA1 host, K95, at 32’C. K95 is nonpermis- sive for A cl 7; cl 7+ recombinants form normal plaques. Positive values varied from 3-7 fold above background levels (spontaneous c17+j.
Construction of Lambda Lysogen Plasmid-bearing lambda lysogenic strains were constructed as fol- lows: MR93galE was Pl-transduced to gal+ with Pl CmclrIOOgrown lytically on N4830 or N4831. These donor strains, described in detail elsewhere (M. Gottesman. manuscript in preparation) are defective lambda lysogens of the structure: W31 Olgal+(ch/D-hlo)A8 (A cl857ABAMAHl). N4830 is N+; N4831 carries two amber mutations in gene N, Nam7,53. The A8 deletion increases the co-transduction frequency of gal and prophage lambda. The BAM deletion removes lambda DNA from 58.0-71.3% (Kourilsky et al., 1978). No pL-operon gene but N is present. The Hl deletion extends from within lambda gene cfo through bio and into or beyond uvrB (Castellazzi, Brachet and Eisen, 1972). Gal’ transductants were tested for lambda immu- nity and for bio. Immune, bio- cells were then transformed with plasmid DNA, selecting for ampicillin resistance as described above.
Assey for Tetracycline Resistance Cells were grown in LB broth containing 50 Fg/ml ampicillin. At an optical density,, of -0.2, tetracycline In the amounts indicated in the legends to Figures 5, 6 and 7) was added and the growth of the cells was monitored by their optical densityam for several hours.
Assay for @Lactamase (O’Callaghan et al., 1972) Cells were grown to a density of lO’/ml in LB broth with 50 pg/ml ampicillin, centrifuged, resuspended in l/20 vol 0.1 M KPO,- (pH 7.0) sonicated (3 X 5 set) with a Branson sonifier. and diluted 1000 fold with 0.1 M KPO,- (pH 7.0). Nitrocefin (provided by Glaxo- Allenburys Research) was dissolved in 50% DMSO in 0.1 M KPOd- (pH 7.0) at a concentration of 1 mg/ml. Cell extract and the above nitrocefin solution were mixed in a ratio of 20 to 1, incubated at 25’C. and the optical densityrs2 was monitored during the first 4 min. Units of /3-lactamase are expressed as:
AO.D.,ez
cell O.D. x 1000.
Adhya. S.. Gottesman. M. and de Crombrugghe. 6. (1974). Release of polarity in Escherichia co/i by gene N of phage A: termination and antitermination of transcription. Proc. Nat. Acad. Sci. USA 77. 2534.
Eolivar, F.. Rodriguez. R. L.. Greene, P. J.. Betlach. M. D.. Heynecker. H. L.. Bayer. H. W.. Cross. J. H. and Falkow. S. (1977). Construction and characterization of new cloning vehicles. Gene 2. 95.

Nut Sequences are Necessary and Sufficient for pN Action 1151
Buttin, G. (I 963a). Mecanismes regulateurs dans la biosynthese des enzymes du metabolisme du galactose chez Escherichie co/i K12. J. Mol. Biol. 7, 183.
Buttin, G. (I 963b). Mecanismes regulateurs dans la biosynthese des enzymes du metabolisme du galactose chez Escherichia co/i K12. J. Mol. Biol. 7. 610.
Castellazzi, M., Brachet. P. and Eisen. H. (1972). Isolation and characterization of deletions in bacteriophage lambda residing as prophage in E. co/i K12. Mol. Gen. Genet. 7 17, 211.
Feiss, M., Adhya. S. and Court, D. (I 972). Isolation of plaque forming galactose transducing strains of phage lambda. Genetics 71, 189.
Franklin, N. C. (I 974). Altered reading of genetic signals fused to the N operon of bacteriophage h: genetic evidence for modification of polymerase by the protein product of the N gene. J. Mol. Biol. 89, 33.
Friedman, D. I., Jolly, C. T. and Mural, Ft. J. (1973a). Interference with the expression of the N gene function of phage h in a mutant of Escherichia co/i. Virology 5 1, 216.
Friedman, D.. Wilgus. G. and Mural, Ft. (1973b). Gene N regulator function of phage XimmPl: evidence that a site of N action differs from a site of N recognition. J. Mol. Viol. 81. 505.
Kourilsky. P.. Perricaudet, M.. Gros. D., Garapin. A., Gottesman. M. E.. Fritsch, A. and Tiollais, P. (1978). Description and properties of bacteriophage lambda vectors useful for the cloning of EcoRl DNA fragments. Biochimie 60, 183.
Lozeron. t-f. A., Dahlberg. J. E. and Szybalski. N. (1976). Processing of the major leftward mRNA of coliphage lambda. Virology 71, 262.
Luuati. D. (I 970). Regulation of lambda exonuclease synthesis: role of the N gene product and h repressor. J. Mol. Biol. 49, 515.
Musso, R.. Di Lauro, R.. Adhya. S. and de Crombrugghe. B. (1977). Dual control for transcription of the galactose operon by cyclic AMP and its receptor protein at two interspersed promoters. Cell 12, 847.
Nakanishi, S.. Adhya. S., Gottesman. M. and Pastan, I. (1973). In vitro repression of the transcription of gal operon by purified gal repressor. Proc. Nat. Acad. Sci. USA 70. 334.
O’Callaghan. C. H.. Morris, A., Kirby, S. M. and Shingler, A. H. (1972). Novel method for detection of p-lactamases by using a chromogenic cephalosporin substrate. Antimicrobial Agents and Chemotherapy 1, 283.
Reyes. 0.. Gottesman. M. and Adhya. S. (1976). Suppression of polarity of insertion mutations in the gal operon and N mutations in bacteriophage lambda. J. Bacterial. 126. 1108.
Roberts, J. W. (1969). Termination factor for RNA synthesis. Nature 224, 1188.
Rodriguez, R. L.. West, R. W., Heyneker, H. L.. Bolivar. F. and Boyer. H. W. (1979). Characterizing wild-type and mutant promoters of the tetracycline resistance gene in pBR313. Nucl. Acids Res. 6, 3267.
Rosenberg, hf.. Court, D.. Shimatake, H.. Brady, C. and Wulff. D. L. (1978). The relation between function and DNA sequence in an intercistronic regulatory region in phage A. Nature 272, 414.
Salstrom. J. S. and Szybalski, W. (1978). Coliphage hnuft-: a unique class of mutants defective in the site of gene N product utilization for antitermination of leftward transcription. J. Mol. Biol. 124, 195.
Taniguchi. T.. O’Neill. M. and de Crombrugghe. B. (1979). The interaction site of the E. coli cyclic AMP receptor protein on the DNA of the promoters of the galactose operon. Proc. Nat. Acad. Sci. USA. 76, 5090.
Ullrich. A., Shine, J.. Chirgwin. J., Pictet, R., Tisher. E.. Rutter. W. and Goodman, H. M. (1977). Rat insulin genes: construction of plasmids containing the coding sequences. Science 196. 1313.
Yarmolinsky. M. and Wiesmeyer. H. (1960). Regulation by coliphage lambda of the expression of the capacity to synthesize a sequence of host enzymes. Proc. Nat. Acad. Sci. USA 46, 1626.





























