Embed Size (px)
Citation preview

ORIGINAL ARTICLE
A. Lametschwandtner Æ U. Lametschwandtner
Ch. Radner Æ B. Minnich
Spatial growth and pattern formation in the small intestine microvascularbed from larval to adult Xenopus laevis: a scanning electron microscopestudy of microvascular corrosion casts
Accepted: 16 May 2006 / Published online: 8 August 2006� Springer-Verlag 2006
Abstract The microvascular anatomy of the smallintestine of metamorphosing tadpoles of the SouthAfrican Clawed Toad, Xenopus laevis (Daudin) is stud-ied from developmental stages 55 to 65 and in adults byscanning electron microscopy (SEM) of vascular corro-sion casts (VCCs) and light microscopy. Up to stage 62,VCCs reveal a dense two-dimensional vascular networkensheating the intestinal tube, whose proximal portionforms a clockwise spiralling outer and its distal portionan anti-clockwise spiralling inner coil. Vessels of theintestinal network impose flat and run circularly toslightly obliquely. Locally, dense capillary plexus withsmall ‘‘holes’’ indicating ongoing intussusceptivemicrovascular growth (IMG) and vessel maturation, arepresent. The typhlosole, an invagination along theproximal portion of the small intestine, reveals a densecapillary bed with locally ongoing IMG. VCCs of stages62/63 for the first time reveal a three-dimensional vas-cular bed with longitudinal intestinal folds of varyingsize and heights greatly enlarging the luminal exchangearea of the intestinal tube. From stage 65 onwards,longitudinal intestinal folds undulate and, thoughsmaller in size and less mature as indicated in VCCs bythe presence of wider, sinus-like vessels with small‘‘holes’’ interposed between, closely resemble the intes-tinal folds present in the small intestine of adult Xeno-pus. Our data suggest that maturation of the vascularpattern in the small intestine of X. laevis tadpoles takesplace successively after stages 62–63, and growth duringthis period is preferentially by intussusception.
Keywords Xenopus Æ Small Intestine Æ Microvascularanatomy Æ Intestinal fold Æ Corrosion casting ÆScanning electron microscopy
Introduction
The gross development of the small intestine of theSouth African Clawed Toad, Xenopus laevis Daudin(Bles 1905; Weisz 1945a, 1945b; Nieuwkopp and Faber1967) and its blood vessel system (Millard 1945, 1949)) iswell described. In a detailed account on Xenopus gutdevelopment (including pancreas, liver, and gall bladder)Chalmers and Slack (1998) focussed on histology andspatial arrangement of the intestine from larval days 3 to7 (stages 40–47 according to Nieuwkopp and Faber1967), while Shi and Ishizuya-Oka (1996) reviewed thebiphasic intestinal development.
The blood vessel system and its abnormalities andvariations in adult Xenopus have been described byMillard (1941, 1942). So far all authors confined to themain supplying and draining vessels, and descriptions ofthe vessels end when they or their main branches arriveat or depart from the target organ(s). None of thesestudies pays attention to the intestinal microvasculature.
Recently, fluorescent labelling of endothelial cells,wide field microscopy and confocal microscopy havebeen used to perform a continuous characterization ofthe vascular development of living embryos from stages33 through 46 (Levine et al. 2003).
To our best knowledge there is neither a detailedstudy done on the spatial growth and pattern formationof the microvasculature of the small intestine of tadpolestages 47 through 66, nor on the three-dimensionalanatomy of the intestinal microvasculature of adultXenopus.
Here we analyse resin corrosion casts of the smallintestine microvasculature of larval stages 55 and 56 and60–65, and adult Xenopus using the excellent resolution
A. Lametschwandtner (&) Æ U. LametschwandtnerCh. Radner Æ B. MinnichDepartment of Organismic Biology, Division of Zoologyand Functional Anatomy, Vascular and Muscle Research Unit,University of Salzburg, Hellbrunnerstrasse 34,5020 Salzburg, AustriaE-mail: [email protected].: +43-662-80445602Fax: +43-662-80445698
Anat Embryol (2006) 211: 535–547DOI 10.1007/s00429-006-0104-2

and high depth of focus of the scanning electronmicroscope (Murakami 1971; Lametschwandtner et al.1990; Aharinejad and Lametschwandtner 1992) andpresent the spatiotemporal changes which the micro-vascular bed of the small intestine undergoes while itshortens its length dramatically to approximately 20%of its initial value (Schreiber et al. 2005) and the animalchanges its life-style from herbivorous (tadpole) to car-nivorous (postmetamorphic animal). Correlative lightmicroscopy of stained paraplast embedded, transversetissue sections serves to attribute cast blood vessels todefined anatomical layers within the intestine wall.
We present evidence that non-sprouting angiogenesis(intussusceptive microvascular growth; IMG) and itsfacets (for review on intussusceptive angiogenesis seeDjonov and Makanya 2004) are heavily involved inlarval to adult intestinal vascular pattern formation.
With respect to the main feeding and draining bloodvessels of the small intestine, we use the nomenclatureproposed by Millard (1941, 1945, 1949)).
Materials and methods
Animals
Tadpoles of X. laevis Daudin were raised in aerated tapwater at 23 ± 0.5�C and fed nettle powder. AdultXenopus were housed in aquaria at 19 ± 0.5�C and awater depth around 15 cm. They were fed grinded beefheart or beef liver twice a week.
Histology
Tadpoles
After staging, weighting (mg) and measuring (mm; totallength, body length; for data see Table 1), tadpoles werekilled by immersion in an overdose of an aqueoussolution of tricaine methansulfonate (MS 222; 0.03%),
fixed by immersion in BOUIN‘s solution, dehydrated ina graded series of ethanol, and embedded in paraplastvia methylbenzoate, benzole and benzole-paraplast. Se-ven to ten micrometer thick sections were stained witheither hematoxyline-eosine (H.E.), or Goldners orBrooks trichrome staining (for details see Adam andCzihak 1964).
Adults
Two adult males (body weights 46 and 50 gm; bodylengths 5.5 and 6.0 cm) were killed by submersion inan overdose of an aqueous solution of MS 222 (0.5%). After weighting and measuring, animals werepinned in a supine position onto a wax plate, the skinoverlaying the sternal region was cut by a medianincision, and the thorax was opened by sternotomy.After opening the pericardium, the heart with bulbuscordis and truncus arteriosus was exposed and athread ligature placed around the bulbus. Thereafter, ahole was punched into the apex of the single ventricleby fine-tipped scissors and a tubing (outer diameter1.2 mm, inner diameter 0.6 mm) was introducedmanually via the ventricle into the bulbus cordis, andfirmly tied in place by the ligature. Finally, both atriawere cut open and rinsing with cold (20�C) AmphibianRinger solution (for recipe see Adam and Czihak1964) started with manual pressure. After clear refluxfrom the opened atria, 10 ml of BOUIN‘s solution(Adam and Czihak 1964) was injected with an infuserpump (40 ml/h). After fixation, the abdominal cavitywas opened, defined segments of the small intestinewere excised, and fixed for another 12–24 h in freshfixative at room temperature (20�C). Specimens thenwere dehydrated in a graded series of ethanol andembedded in paraplast or Epon 812. Seven micrometerthick sections or semithin sections (0.5 lm thick) weremade and stained with AZAN stain (Romeis 1989) ormethylenblue II (semithin sections). Micrographstaken with a digital camera (Nikon DS-5 M) were
Table 1 Quantitative data onintestinal vessels of larval andadult Xenopus laevis gained by3D-morphometry of vascularcorrosion casts
NM not measuredSignificant data (p £ 0.5) aremarked by an asterisk
536

imported to Photoshop 7.0 (Adobe) and brightnessand contrast were adjusted.
Vascular casting
Tadpoles
After staging, weighting (mg) and measuring (mm; to-tal length, body length), tadpoles were killed byimmersion in an overdose of an aqueous solution oftricaine methansulfonate (MS 222; 0.03%), and pinnedin a supine position on a wax plate submerged inamphibian Ringer solution. After excision of the skinoverlaying the heart with microscissors, the pigmentedpericardium was opened, heart and bulbus cordis wereexposed, and a ligature from 11–0 surgical suturematerial (diameter 25 lm) was placed around the bul-bus. Then the apex of the ventricle was cut open by asmall incision with microscissors and a thin glass can-nula guided by a micromanipulator was insertedthrough the ventricle into the bulbus cordis and fixed inplace by closing the ligature. Thereafter, the venoussinus was opened and rinsing with amphibian Ringersolution started. As soon as clear reflux came from theopened venous sinus Mercox CL-2B (Ladd ResearchInc., Burlington, Vermont, USA), diluted with mono-meric methylmethacrylate (4 + 1, v + v, 10 mlmonomeric methylmethacrylate contained 0.85 g initi-ator paste MA) was injected with an infuser pump (5–7 ml/h) until the effluent resin became highly viscous.Animals were left for at least 30 min at room temper-ature (20�C) to allow polymerization of the injectedresin, then placed into a water bath (60�C, 12 h) fortempering, and finally macerated in KOH (7.5%, 12–24 h). When maceration was completed, vascular castswere transferred via several passages of distilled waterinto formic acid (5%, 5–10 min, 20�C), rinsed again inseveral passages of distilled water, frozen in distilledwater and freeze-dried in a Lyovac GT2 (LeyboldHeraeus, Cologne, FRG). Dry specimens comprisingmost of the digestive tract were mounted on stubsusing the ‘‘conductive bridge method’’ (Lametschw-andtner et al. 1980) and sputtered with a thin layer ofgold. Coated specimens were investigated with a scan-ning electron microscope (Stereoscan 250, CambridgeLtd., Cambridge, UK) at an accelerating voltage of5–10 kV.
After a first documentation and analysis of thesuperficial (external) vascular layers of the small intes-tine and common bile duct, specimens were removedfrom specimen stubs, submerged into bidistilled water,orientated properly, and frozen at �20�C in a deep-freezer. Ice-embedded specimens were frozen on woodenspecimen holders and sectioned transversely, longitudi-nally, tangentially or horizontally with a mini-wheel-sawplaced in the cryo-chamber of a freeze-microtome
(Lametschwandtner and Lametschwandtner 1992) Sec-tioned ice-embedded specimens were allowed to thaw indistilled water, cleaned, and refrozen in bidistilled waterfor subsequent freeze-drying in the Lyovac GT. Furthersteps in specimen preparation were as described above.According to the needs, specimens were repeatedly sec-tioned to expose interesting vascular territories.
Adults
For preparatory steps preceding resin injection see his-tology (above). After clear reflux from the opened atria,5–7 ml of Mercox-Cl-2B was injected with manualpressure. All further procedures were the same as de-scribed for tadpoles.
Quantitative analyses
To gain insights into dimensional changes of the grow-ing intestinal vascular network we performed 3D-mor-phometry (for details see Minnich et al. 1999) onvascular corrosion casts of stages 56, 59 and 60 and inthe adult stage. Using a grid, we first counted thenumber of ‘‘holes’’ (meshes) per 100 · 100 lm2 fromprinted SEM micrographs (final magnification ·400)thereby neglecting those ‘‘holes’’ hitting the upper hor-izontal and the right vertical borderlines of the grid.Then we measured (1) diameters of arterioles, venulesand capillaries, (2) lengths and widths of vascular me-shes, and (3) branching angles of arterial, capillary andvenous vessels by 3D-morphometry. Data was subjectedto one-way analysis of variance (ANOVA) and pair-wisemultiple comparison procedures (Dunn’s method;Holm-Sidak method) were performed to test data forsignificant differences (P < 0.05) between stages.
Results
Tadpoles
Anatomy
In tadpoles of stage 55 (Fig. 1a), the small intestineforms an inner and an outer coil, whereby the loops ofthe outer coil, which vary in number with the develop-mental stage, spiral clockwise around the loops of theanti-clockwise running inner spiral (Fig. 1b, c). At thebase of the coils the small intestine abruptly changes intothe large intestine (Fig. 1b, c; arrows).
Histology
In stage 56, the small intestine consists of a simplelocally ciliated columnar epithelium with single mucous
537

glands scattered between ciliated epithelial cells. Theepithelium rests on a thin layer of connective tissue.Between the epithelium and the connective tissue, bloodvessels are located. The muscular layer is made up by 1to 2 layers of smooth muscle cells only. A prominentinvagination, the typhlosole, bulges into the intestinallumen (Fig. 1d).
During stages 57–59, the muscular layer increases inthickness whereby the inner circular layer becomes muchthicker than the outer longitudinal layer. Intestinal foldsare still absent. In stage 60, the muscular layer furtherincreases in thickness and intestinal folds start to formand bulge into the intestinal lumen. From stages 62/63onwards, distinct longitudinal intestinal folds of varyingheights are present which then greatly enlarge theluminal surface of the intestine.
Vascular architecture
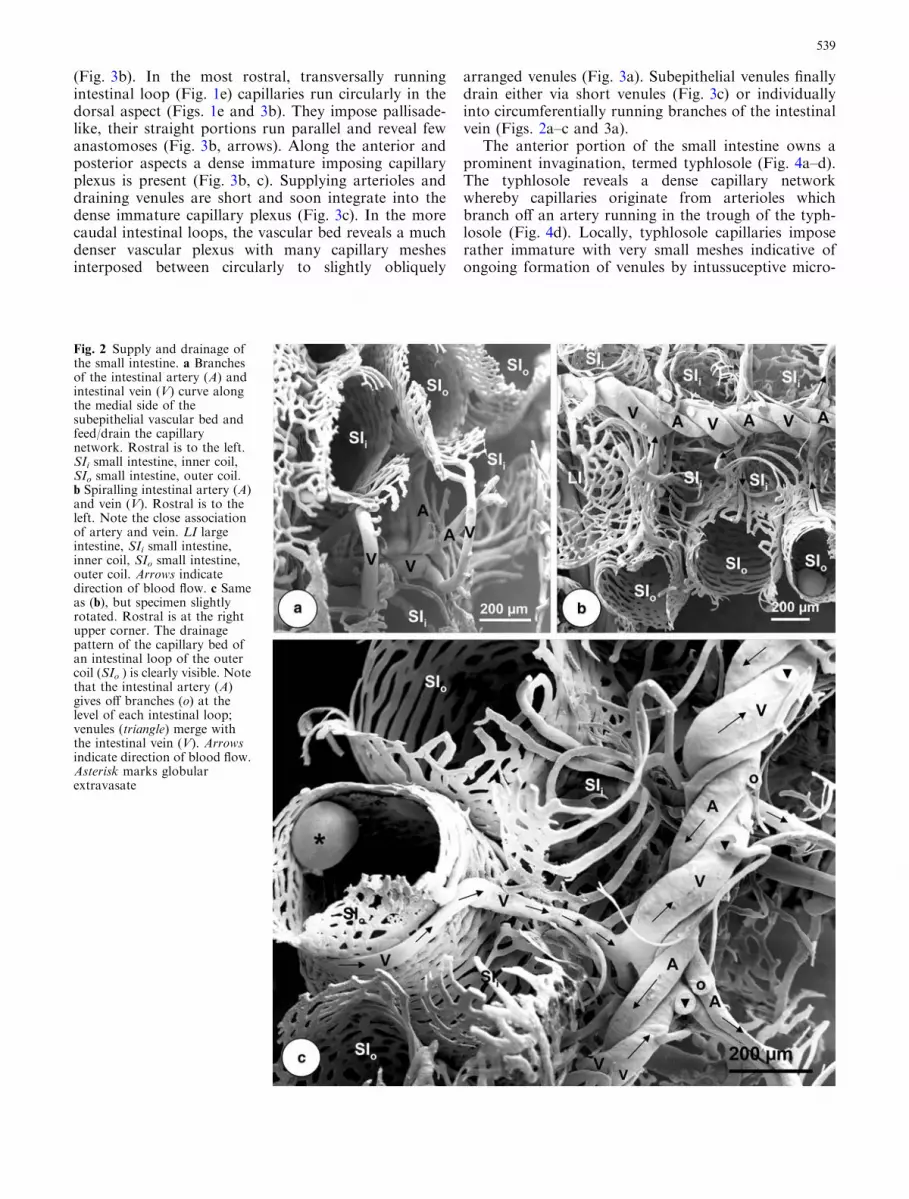
Stages 56–59 In stage 56, small and large intestines re-veal a single-layered vascular bed (Fig. 1e). The arterialsupply of the intestinal vasculature is via the intestinalartery which originates from the mesenteric-celiac arteryand ascends along the inner coil of the small intestinetowards the coils apex. The artery twists clockwisearound the helically descending intestinal vein (Fig. 2a–c) and gives off branches at the level of the coil loops(Fig. 2a–c). Branches run along the inner aspect of theintestinal loops of outer and inner coil and give off shortarterioles which feed the single-layered subepithelialcapillary bed (Fig. 3a–c). Intestinal capillary networkpatterns vary along the proximal-distal extension(Fig. 3a) as well as along the intestinal circumference
Fig. 1 Anatomy, histology and vasculature of the tadpole smallintestine in developmental stages 55 to 56 (staging according toNieuwkopp and Faber 1967). a Tadpole. Ventral view. Note thesilvery pigmented abdomen (A). b Anatomy of the tadpoleintestines (stage 55). Ventral view. Rostral is at the top. LI largeintestine, S stomach, SI small intestine. Note the five clockwiserunning intestinal loops (1–5) of the outer coil which enclose theloops of the inner coil. The arrowhead marks the turning point ofthe outer coil into the inner coil; the arrow marks the end of thesmall intestine (= ileum) and the beginning of the large intestine.Asterisk corresponds to that in (e). c Same as in b, but intestinalcoils are torn apart to show course and spatial relations of internaland external coil loops. Note the intestinal vessels ascending withinthe inner coil (arrow). Arrowhead marks the end of the smallintestine. d Microscopic anatomy of the tadpole small intestine
(duodenum) (stage 55). Transverse section. Goldners Trichromestaining. E intestinal epithelium, L lumen, Ty typhlosole. Note thatthe lamina propria is very thin and the muscular layer consists of 1–2 smooth muscle cell layers only. Triangle corresponds to that in(e). e Microvascular bed of the tadpole intestines (stage 56).Vascular corrosion cast (VCC). Scanning electron microscope(SEM) image. Rostral is to the top. For orientation compare with(b). To enable visualization of subepithelial vascular patterns of theintestines the ventral circumference of the intestinal vascularcorrosion casts has been removed. A artery, LI large intestine,SIo small intestine, outer coil, SIi small intestine, inner coil, Tytyphlosole, V vein. Note the merging of the small intestine (largearrow) into the large intestine. Asterisk corresponds to that in b,triangle to that in (d)
538
(Fig. 3b). In the most rostral, transversally runningintestinal loop (Fig. 1e) capillaries run circularly in thedorsal aspect (Figs. 1e and 3b). They impose pallisade-like, their straight portions run parallel and reveal fewanastomoses (Fig. 3b, arrows). Along the anterior andposterior aspects a dense immature imposing capillaryplexus is present (Fig. 3b, c). Supplying arterioles anddraining venules are short and soon integrate into thedense immature capillary plexus (Fig. 3c). In the morecaudal intestinal loops, the vascular bed reveals a muchdenser vascular plexus with many capillary meshesinterposed between circularly to slightly obliquely
arranged venules (Fig. 3a). Subepithelial venules finallydrain either via short venules (Fig. 3c) or individuallyinto circumferentially running branches of the intestinalvein (Figs. 2a–c and 3a).
The anterior portion of the small intestine owns aprominent invagination, termed typhlosole (Fig. 4a–d).The typhlosole reveals a dense capillary networkwhereby capillaries originate from arterioles whichbranch off an artery running in the trough of the typh-losole (Fig. 4d). Locally, typhlosole capillaries imposerather immature with very small meshes indicative ofongoing formation of venules by intussuceptive micro-
Fig. 2 Supply and drainage ofthe small intestine. a Branchesof the intestinal artery (A) andintestinal vein (V) curve alongthe medial side of thesubepithelial vascular bed andfeed/drain the capillarynetwork. Rostral is to the left.SIi small intestine, inner coil,SIo small intestine, outer coil.b Spiralling intestinal artery (A)and vein (V). Rostral is to theleft. Note the close associationof artery and vein. LI largeintestine, SIi small intestine,inner coil, SIo small intestine,outer coil. Arrows indicatedirection of blood flow. c Sameas (b), but specimen slightlyrotated. Rostral is at the rightupper corner. The drainagepattern of the capillary bed ofan intestinal loop of the outercoil (SIo ) is clearly visible. Notethat the intestinal artery (A)gives off branches (o) at thelevel of each intestinal loop;venules (triangle) merge withthe intestinal vein (V). Arrowsindicate direction of blood flow.Asterisk marks globularextravasate
539

vascular growth (IMG) (Fig. 4c). Capillaries of thetyphlosole drain via distinct venules towards adjacentareas of the intestinal vascular bed (Fig. 4b).
Stages 60–65 Intestinal folds start to form in stage 60.The still one-layered capillary bed follows the inbulgingintestinal epithelium and outlines the longitudinallyorientated forming folds (Fig. 5). Some folds are short(Fig. 5a), others are longer and run parallel (Fig. 5b).From stage 62/63 onwards folds increase in size andnumber (Fig. 6a–e). In stage 64, folds are ratherstraight and of varying heights with smaller foldsinterposed between larger ones (Fig. 6b, d–e). Locallyfolds show already a zig-zag course. Mucosal foldcapillaries form a narrow-meshed network with vesselsorientated from the crest of the fold to the trough.Fold venules drain into circularly running venules lo-cated within the lamina propria of the intestinal mu-cosa (Fig. 6a–c). Locally, longitudinally runningcapillaries of the muscular layer can be seen (Fig. 6a,arrows). Thus already at stage 64–65, a mucosal foldvascular pattern resembling that of the adult smallintestine is established.
Adult
Anatomy The small intestine of the adult is rather shortand consists of a proximal (= duodenum) and a distalportion (= ileum). The duodenum runs close to the
medial aspect of the stomach towards rostrally and thenbends towards laterally where it may form 1 to 2 smallloopings before it changes into the colon.
Histology
The small intestine (from inside to outside) consists of anepithelium and a lamina propria forming the mucosa, amuscularis composed of an inner circular and an outerlongitudinal layer of smooth muscle cells, a thin subse-rosa, and a serosa forming the boundary layer to theabdominal cavity. The mucosa forms longitudinallyorientated, zig-zag running mucosal folds with locallytransverse connecting folds; villi are lacking.
Vascular architecture Anterior and medial intestinalarteries, right gastric artery, and haemorrhoidal arter-y—all branches of the celiac-mesenteric artery supplythe small intestine (Millard 1941). Arteries pierce themuscular layer and run within the lamina propria(Fig. 7a). Arteries give off few centrifugally directedarterioles to supply the capillary bed of the muscularlayer (Fig. 7b, c). Capillaries of the muscular layer locatebetween the outer longitudinal and the inner circularsmooth muscle cell layers. Capillaries run parallel tosmooth muscle cells and thus form a rectangular mesh-work (Fig. 7a, b). Capillaries drain by many smallpostcapillary venules into veins of the lamina propria(Fig. 7b; arrows). The lamina propria is very rich in
Fig. 3 Vascular patterns of theexternal coils of the tadpole(stage 56) small intestine.Subepithelial view. aEnlargement of (1e). Two-dimensional vascular networkcomposed of circularly toslightly obliquely running flatvessels. Note the rather matureimposing venules (V) with adense meshwork of immaturecapillaries (c) interposedbetween. A artery. Arrowsindicate direction of blood flow.b Detail from (1e). Note theparallel running capillaries(arrows) at the rostralcircumference of the intestineand the dense meshwork at thedorsal and ventralcircumferences. c Drainage ofthe intestinal capillary bed via ashort venule (V). Note that thevenule forms within a shortdistance
540

blood vessels whereby large veins by far dominate thefew supplying smaller arteries (Fig. 7a). Intestinalarteries run transversally to slightly obliquely (Fig. 7a)and traverse longitudinally arranged veins (Fig. 7a).Arterioles give off capillaries which feed the subepithelialcapillary network at the trough of the intestinal folds
and then bend towards deeper layers of the laminapropria to ascend within the longitudinally runningintestinal folds (Fig. 8). Ascending arterioles give offproximal capillaries which feed the supepithelial capil-lary network of the trough areas and the most ventralareas of the intestinal folds (Fig. 9a). Ascending arteri-
Fig. 5 Intestinal fold formation in tadpoles of Xenopus laevis atstage 60. Luminal views. Double-headed arrows mark long axis ofsmall intestine. a Initial stage. Note the wide, dilated capillariesgradually elevating into the small intestine lumen (L) representingthe vascular bed of the ingrowing longitudinally arranged intestinal
fold. b More advanced stage revealing the capillary beds of twolongitudinally running intestinal folds. Note the wide marginalvenule (ve) at the crest of one fold and the one-layered capillary bed(c) in the trough area between folds. A artery, V vein
Fig. 4 Vascular patterns of thetyphlosole. Stage 56. Part of thevascular bed is removed toexpose the inbulging typhlosole.a Location and course of thetyphlosole (Ty) in the proximalintestine (= duodenum); Largeintestine (LI). b Dense capillaryplexus of the typhlosole withvery small mesh holes (smallarrows) indicative of ongoingintussusceptive microvasculargrowth (non-sproutingangiogenesis). Note thattyphlosole capillaries merge toform individual venules (largearrows) which capillarize intothe adjacent intestinal capillarybed. c Portion of the typhlosolewith an immature imposingvascular bed. The small holes(arrows) indicate non-sproutingangiogenesis leading to theformation of more maturevessels (dashed lines). Note thelarger meshes at the left sideand the more mature imposingcapillaries (c). d Arterial supplyof the typhlosole. Note theartery (A) in the trough of thetyphlosole. Arrow indicatesdirection of blood flow
541

oles repeatedly give off side-branches which capillarizeand form the subepithelial capillary bed of medial andapical areas of both sides of the folds (Fig. 9a, b;arrowheads). The terminal arteriole finally joins themarginal venule of the intestinal fold which runs alongthe crest of intestinal folds without becoming a capillary(Fig. 9a–c). Generally, subepithelial postcapillary foldvenules drain into a more centrally located venule(Fig. 9a). The marginal venule of the intestinal fold crestdrains by and by via a prominent venule into the centralvenule (Fig. 9a; arrows). Locally, arterio-venous transi-tion distances are rather short. Central venules emptyinto a wide-meshed venous network located in the outerregion of the lamina propria (Figs. 8 and 9a). Veins ofthe proximal small intestine (= duodenum) (Fig. 9d)
drain via the medial gastric vein (pyloric region) and thegastro-duodenal vein (duodenum) into the left branch ofthe abdominal vein or via the hepatic portal vein into theliver (Millard 1941).
In adult Xenopus, the mucosa of the small intestine isstrongly folded (Fig. 10a–d). Folds start at the pyloricregion where the characteristic microvascular pattern ofthe stomach changes rather abruptly into the intestinalpattern of the duodenum where folds bifurcate or con-nect with closeby neighbouring folds via circularly run-ning folds (Fig. 10a and b). At the transition zoneglomerular vessel formations can be found (Fig. 10a,insert). Along the small intestine, pattern and height offolds vary locally as seen best in longitudinally ortransversally sectioned specimens (Fig. 10c and d).
Fig. 6 Intestinal microvascular anatomy in tadpoles of stages 60–65. a Microvascularization of the stomach (ST) and proximalsmall intestine (= duodenum, DD). Portions of the vascularcorrosion casts of the pyloric region of the stomach and the initialregion of the duodenum are removed to enable a luminal view atthe gastric mucosa (GM) and intestinal folds (IF). Note tinylongitudinal folds of the gastric mucosa (large arrows). Locallycapillaries of the muscular layer of the duodenum can be seen(small arrows). Dashed arrows mark veins within the laminapropria of the duodenum which drain mucosal folds. P pylorus. bLongitudional mucosal folds of the small intestine (IF) of atadpole at stage 64. Lumenal view. Note the marginal venule atthe crests of the folds (arrows). c Drainage pattern of an intestinal
fold (IF) at stage 64. Note that capillaries (c) drain by descendingvenules into larger veins (V) located in the lamina propria. Arrowsindicate direction of blood flow. d Microanatomy of the smallintestine at stage 65. Transverse section. Goldners Trichromestaining. Many longitudinal folds of varying height are seen.Arrows indicate the location of the intestinal fold (IF) vascularbeds. E intestinal epithelium, Lp lamina propria ascending intothe intestinal folds, L lumen, M muscular layer (with innercircular and outer longitudinal layers of smooth muscle cells), Sserosa, T trough between intestinal folds. e Microangioarchitec-ture of the small intestine at stage 65. Transverse section.Vascular corrosion cast. Note the large intestinal folds (IF) givingthe intestinal lumen (L) a star-like appearance
542

Quantitative data
In the flat two-dimensional intestinal vascular networkof stages 56, 59 and 60 we counted an average of 2.0, 4.3,5.0 and 0.9 ‘‘holes’’ per unit area (100 x 100 lm).
Mean diameters of capillaries were 10.1 lm (stage56), 20.7 lm (stage 59), 29.2 lm (stage 60), and 26.7 mm(adult) while those of arterioles were 32.4 lm (stage 56),45.8 lm (stage 60) and 46.1 lm (adult). For further datasee Table 1.
Mean diameters of venules were 71.8 lm (stage 59),33.5 lm (stage 60) and 37.9 lm (adult). For furtherdata see Table 1. The pair-wise multiple comparisonprocedures (Dunn’s method) revealed that overalldiameters of intestinal vessels differed significantly be-tween stages 56 and 60, but not between stages 56 and59, and the adult stage. Holm-Sidak’s method revealedsignificant differences in capillary diameters betweenstages 56 and 60 and between stage 56 and the adultstage only.
Mean branching angles of capillaries were 89.1�(stage 56), 68.1� (stage 59), 82.3� (stage 60), and 74.9�(adult), while those of venules in corresponding stages
were 63.4�, 59.1�, 75.0�, and 66.4�. For further data seeTable 1. There were no significant differences inbranching angles of neither vessels.
Discussion
While anatomical, morphological, fine structural andphysiological changes of the digestive tract duringamphibian metamorphosis are studied by several au-thors (e.g. Kordylewski 1983; Ishizuya-Oka and Shi-mozawa 1987; Chalmers and Slack 1998; Shi 2000;Schreiber et al. 2005) existing knowledge on the intesti-nal vascularization is limited to the gross arterial supplyand venous drainage, both in the tadpole (Weisz 1945a,b; Nieuwkopp and Faber 1967; Millard 1945, 1949) andin the adult (Millard 1941, 1942)). Neither the larval northe juvenile or adult intestinal microvascular architec-ture is studied nor is it known by which of the existingmechanisms of angiogenesis, i.e. sprouting, non-sprout-ing (= intussuceptive microvascular growth) and/orbridging the microvascular bed of the intestine formsfrom a single-layered, dense capillary network into ahighly structured three-dimensional network with
Fig. 7 Microvascular anatomyof the small intestine in adultXenopus laevis. a Segment of theproximal duodenum.Doubleheaded arrow marks thelongitudinal axis of theintestinal segment. Vascularcorrosion cast. Serosal view.Arteries (A) run transversally toslightly obliquely and revealtypical dichotomousbranchings. Veins (V) are moreabundant and their initialsegments run longitudinally.Veno-venous anastomoses arenumerous (dashed lines). Thevascular bed of the muscularlayer imposes as delicaterectangular vascular network(arrows) overlaying arteries andveins. b Capillary bed of themuscular layer overlayingvessels of the lamina propria.Note rare arteriolar feeder(large arrow) but multiplevenular drainers (small arrows).c Origin of an arteriolesupplying the capillary bed ofthe muscular layer (arrow). Aartery. Dashed line arrowindicates direction of bloodflow
543

prominent intestinal folds to be found from late climaxstages onwards.
In larval Xenopus, the development of the subintes-tinal vein from the left omphalomesenteric vein in stage40 and its positional change from the left side of theintestine (stage 40), to the ventral aspect (stage 41) to itsfinal position on the right side (stage 43) as well as itsfunction as draining vessel for the gastric vein are de-scribed after labelling of the endothelial cells with DiI-labeled acetylated low-density lipoprotein and confocalmicroscopy (Levine et al. 2003). The potential of thismethod is in the continuous in-vivo observation ofangiogenic processes over time and space. Thoughscanning electron microscopy of resin corrosion castscan just depict the microvascular anatomy present at asingle time-point and relies on a continuous series ofpreparations, and on correlative light microscopy ofcorresponding stained tissue sections, it is a powerfultechnique. Due to its high depth of focus, its excellentspatial resolution, and the possibility to take stereopaired images and to analyse them by 3D-morphometry(Malkusch et al. 1995; Minnich et al. 1999) it allows toanalyse spatial growth and pattern formation withinmicrovascular networks in great detail (Patan et al.1992; Burri and Djonov 2002; Djonov and Makanya2004).
Our data on the number of ‘‘holes’’ which representthe sites of pilar formation during intussusceptivemicrovascular growth and on capillary diameters fromstages 56 to 60 and in the adult strengthen the mor-
phological observations of a maturing vascular networkwith increasing mesh sizes and decreasing capillarydiameters. The flattened vessels which made up thetwo-dimensional vascular bed of the small intestine instages 55 and 56 are considered real structures ratherthen artefacts. Though only 1 to 2 layers of smoothmuscle cells are present, they are obviously strong en-ough to ‘‘squeeze’’ the immature blood vessels resultingin the observed ‘‘ribbon-like’’, flattened cast bloodvessels.
While much is known on genetic and epigeneticmechanisms in the early development of the vascularsystem (Ribatti 2006) and morphology and duration ofthe phases of intussusceptive microvascular growth andits significance for vascular network formation are welldescribed (Djonov and Makanya 2004), we still lackconclusive information on the factors which driveintussusceptive microvascular growth and lead to theformation of a highly structured three-dimensional net-work as represented in the small intestine of adultXenopus and in many other tissues and organs.
In her comprehensive work on the vascular anat-omy of adult X. laevis which is based on dissections of48 animals injected with Ranvier’s carmine-gelatinemass or Ranvier’s Prussian blue through the conusarteriosus, Millard (1941) gives an excellent descriptionand depiction of all arteries and veins and theirbranches, but does not refer to the intrinsic micro-vasculature. Moreover, to enable a clear demonstra-tion of the vascular anatomy of the gastro-intestinal
Fig. 8 Microvascular anatomyof the small intestine of adultXenopus laevis. Serosal view.Detail. Terminal portion of anintestinal artery (A) with severalbifurcations. Note that theterminal arterioles (AA) feedthe subepithelial one-layeredcapillary network (c) at theintestinal fold troughs and thenpierce into the depth to ascendwithin the intestinal folds(encircled areas). Venules (VV)drain intestinal folds into largerveins (V) located within thelamina propria. Vessels of themuscular layer (asterisks) runabove arteries and veins.Dashed line arrows indicatedirection of blood flow inarterial vessels. Arrows indicatedirection of blood flow invenous vessels
544

tract, the author documents the vessels after havingpulled the digestive tract to the left (Millard 1941;Fig. 6) thereby disturbing the in-vivo topography ofthe vascular anatomy as well as the spatial relationsbetween vessels. The vascular corrosion casting meth-od (Murakami 1971), however, enables to documentthe true spatial arrangement of the vessels with thetopographical relations between blood vessels ofneighbouring tissues and organs strictly conserved.Furthermore, after having micrographed all interestingdetails in the SEM, vascular corrosion casts can beremoved from the specimen stub, submerged in dis-tilled water, frozen, and sectioned in any desireddirection to expose and link deeper vascular territorieswith superficial ones in order to trace individualsupplying arteries and/or draining veins over time
and space down to the microvascular level (see e.g.Figs. 9a, 10c).
Intestinal mucosa fold formation is reported to beginat stage 60 (Chalmers and Slack 1998; Chalmers et al.2000). Corrosion casts of the intestinal vasculature ofstage 60 tadpoles reveal how the capillary bed followsthe inbulging intestinal epithelium. Tadpoles of stage 62/63 already revealed conspicuous longitudinal mucosalfolds which increased in heights and complexity towardsstages 64 and 65.
In adult Xenopus there exists abnormalities andvariations in the vascular system (Millard 1942). Withrespect to the intestinal vessels, abnormal branches ofthe right arteria and vena cutanea femoralis lateralissupplied/drained the posterior (distal) intestine (Millard1942). Moreover, the medial gastric vein and the gastro-
Fig. 9 Vascular patterns of intestinal folds in adult Xenopus laevis.a Supply and drainage pattern of an intestinal fold. A terminalarteriole (AA) ascends within the fold towards the apex of the fold(dashed line). The arteriole gives off branches (arrows) at variouslevels and supplies troughs (T) between folds as well as the vascularbed of the proper intestinal fold. Fold capillaries drain via acentrally located venule (VV) into veins (V) within the laminapropria. Note the marginal venule running along the crest of thelongitudinal folds (large arrows). Small arrows indicate smallvenules draining into the central venule. b Same as (a), but capillarybed of one side of the fold removed to expose the course of the
supplying terminal arteriole (AA). Note that the arteriole continuesinto the marginal venule (small arrows) at the crest of the intestinalfold (IF) without becoming a capillary. Arrows indicate direction ofblood flow within the terminal arteriole and draining venule (VV).L lumen, Lp lamina propria T trough between intestinal folds. cRelations between terminal portion of the supplying arteriole (AA)and marginal venule (MV) at the crest of an intestinal fold. Arrowsindicate direction of blood flow. d Drainage of the proximalduodenum (DD) into a branch of the medial gastric vein (GV).DGV duodenal-gastric vein, GA branches of gastric artery, Ppancreas, V vein. Arrows mark direction of blood flow
545

duodenal vein may either drain individually into the leftroot of the abdominal vein or join before and empty as asingle vein into the left root of the abdominal vein(Millard 1941). In the present study we did not payattention to existing abnormalities.
Acknowledgments This work was supported by a grant from theStiftungs- und Forderungsgesellschaft der Paris-Lodron-Universi-tat Salzburg (A.L.). The authors thank Ms. Synnove Tholo forexcellent technical assistance, Mr. Andreas Zankl for photograhicwork, and OR Dr. W.D. Krautgartner for providing excellentworking conditions in the SEM-facility.
References
Adam H, Czihak G (1964) Arbeitsmethoden der makroskopischenund mikroskopischen Anatomie. G. Fischer Verlag, Stuttgart
Aharinejad S, Lametschwandtner A (1992) Scanning electronmicroscopy of vascular sorrosion casts. Springer-Verlag, Wien-Berlin
Bles EJ (1905) The life history of X. laevis Daudin. Trans Roy SocEdin 41:789
Burri PH, Djonov V (2002) Intussusceptive angiogenesis—thealternative to capillary sprouting. Mol Aspects Med 23:1–27
Chalmers AD, Slack JMW (1998) Development of the gut inXenopus laevis. Dev Dyn 212:509–521
Chalmers AD, Slack JMW, Beck CW (2000) Regional geneexpression in the epithelia of the Xenopus gut. Mech Dev96:125–128
Djonov V, Makanya AN (2004) New insights into intussusceptiveangiogenesis. In: Clauss M, Breier G (eds) Mechanisms ofangiogenesis. Birkhauser, Basel-Boston-Berlin, pp 17–33
Ishizuya-Oka A, Shimozawa A (1987) Development of the con-nective tissue in the digestive tract of the larval and metamor-phosing Xenopus laevis. Anat Anz 164:81–93
Kordylewski L (1983) Light and electron microscopic observationsof the development of intestinal musculature in Xenopus. ZMikroskop Anat Forsch 97:719–734
Lametschwandtner A, Simonsberger P, Adam H (1980) On theprevention of specimen charging in scanning electron micros-copy of vascular corrosion casts by attaching conductivebridges. Mikroskopie 36:270–273
Lametschwandtner A, Lametschwandtner U, Weiger T (1990)Scanning electron microscopy of vascular corrosioncasts—technique and applications: updated review. ScanningMicrosc 4:889–941
Lametschwandtner A, Lametschwandtner A (1992) Historical re-view and technical survey of vascular casting and scanningelectron microscopy. In: Motta PM, Murakami T, Fujita H(eds) Scanning electron microscopy of vascular casts: methodsand applications. Kluwer, Dordrecht, London, pp 1–11
Levine AJ, Munoz-Sanjuan I, Bell E, North AJ, Brivanlou AH(2003) Fluorescent labeling of endothelial cellws allows in vivo,continous characterization of the vascular development ofXenopus laevis. Dev Biol 254:50–67
Malkusch W, Konerding MA, Klapthor B, Bruch J (1995) Asimple and accurate method for 3-D measurements in micro-corrosion casts illustrated with tumour vascularization. AnalCell Pathol 9:69–81
Millard N (1941) The vascular anatomy of Xenopus laevis (Dau-din). Trans Roy Soc S Africa 28:387–439
Fig. 10 Microvascular patternsof intestinal folds in adultXenopus laevis. a Transitionzone from stomach (ST) toproximal duodenum (DD).Note the undulating intestinalfolds (left; IF) and theglomerular vascular formationsat the gastro-duodenaltransition zone (arrows; see alsoinsert). b Vascular patterns ofintestinal folds. Lumenal view.Large arrow indicateslongitudinal axis of theduodenum. Note thatlongitudinally undulating foldsinterconnect with neighbouringfolds by numerous transversallyrunning, slightly undulatingfolds (arrows). c Microvascularpattern of longitudinallyrunning intestinal folds.Longitudinal section. A artery,V vein. Note the marginalvenules at the crest of parallelrunning folds (arrows). dMicrovascular patterns ofintestinal folds. Transversesection. Note the luminalextension of the intestinal foldsgiving the lumen (L) a slit-likeappearance. Note the singlelayer of capillaries (arrows) atthe depth of the troughs (T)(enboxed areas). V vein
546

Millard N (1942) Abnormalities and variations in the vascularsystem of Xenopus laevis (Daudin). Trans Roy Soc S Africa29:9–28
Millard N (1945) The development of the arterial system ofXenopus laevis (Daudin) including experiments on the destruc-tion of the larval aortic arches. Trans Roy Soc S Africa 30:217–234
Millard N (1949) The development of the venous system of Xeno-pus laevis (Daudin). Trans Roy Soc S Africa 32:55–97
Minnich B, Leeb H, Bernroider EW, Lametschwandtner A (1999)Three-dimensional morphometry in scanning electron micros-copy: a technique for accurate dimensional and angular mea-surements of microstructures using stereopaired digitizedimages and digital image analysis. J Microsc 195:23–33
Murakami T (1971) Application of the scanning electron micro-scope to the study of the fine distribution of the blood vessels.Arch histol Jpn 32:445–454
Nieuwkopp PD, Faber J (1967) A normal table of Xenopus devel-opment (Daudin). North Holland, Amsterdam
Patan S, Alvarez MJ, Schittny C, Burri PH (1992) Intussusceptivemicrovascular growth: a common alternative to capillarysprouting. Arch Histol Cytol 55:65–75
Ribatti D (2006) Genetic and epigenetic mechanisms in the earlydevelopment of the vascular system. J Anat 208:139–152
Romeis B (1989) Mikroskopische Technik. 17. neubearb. Aufl.,Urban and Schwarzenberg. Munchen, Wien, Baltimore
Shi YB, Ishizuya-Oka A (1996) Biphasic intestinal development inamphibians: embryogenesis and remodeling during metamor-phosis. Curr Top Dev Biol 32:205–235
Shi YB (2000) Amphibian metamorphosis. From morphology tomolecular biology. Wiley, New York
Schreiber AM, Cai L, Brown DD (2005) Remodelling of theintestine during metamorphosis of Xenopus laevis. PNAS102:3720–3725
Weisz P (1945a) The development and morphology of the larva ofthe South African Clawed Toad, Xenopus laevis. I. The thirdform tadpole. J Morphol 77:163–192
Weisz P (1945b) The development and morphology of the larva ofthe South African Clawed Toad, Xenopus laevis. II. Thehatching and the first and second-form tadpole. J Morphol77:193–217
547





























