Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Sox2 expression in breast tumours and activation in breast cancer stem cells
O Leis1, A Eguiara1, E Lopez-Arribillaga1, MJ Alberdi2, S Hernandez-Garcia3, K Elorriaga2,A Pandiella3, R Rezola2 and AG Martin1
1Regulation of Cell Growth Laboratory, Fundacion Inbiomed, San Sebastian, Spain; 2Department of Pathology, Onkologikoa,San Sebastian, Spain and 3Instituto de Biologia Molecular y Celular del Cancer, Salamanca, Spain
The cancer stem cell (CSC) model does not imply thattumours are generated from transformed tissue stem cells.The target of transformation could be a tissue stem cell, aprogenitor cell, or a differentiated cell that acquires self-renewal ability. The observation that induced pluripotencyreprogramming and cancer are related has lead to thespeculation that CSCs may arise through a reprogram-ming-like mechanism. Expression of pluripotency genes(Oct4, Nanog and Sox2) was tested in breast tumours byimmunohistochemistry and it was found that Sox2 isexpressed in early stage breast tumours. However,expression of Oct4 or Nanog was not found. Mammo-sphere formation in culture was used to reveal stem cellproperties, where expression of Sox2, but not Oct4 orNanog, was induced. Over-expression of Sox2 increasedmammosphere formation, effect dependent on continuousSox2 expression; furthermore, Sox2 knockdown pre-vented mammosphere formation and delayed tumourformation in xenograft tumour initiation models. Induc-tion of Sox2 expression was achieved through activationof the distal enhancer of Sox2 promoter upon sphereformation, the same element that controls Sox2 transcrip-tion in pluripotent stem cells. These findings suggest thatreactivation of Sox2 represents an early step in breasttumour initiation, explaining tumour heterogeneity byplacing the tumour-initiating event in any cell along theaxis of mammary differentiation.Oncogene advance online publication, 8 August 2011;doi:10.1038/onc.2011.338
Keywords: Sox2; breast cancer; cancer stem cell;mammosphere
Introduction
Many solid tumours, including breast cancer, exhibit afunctional hierarchy of cells of which only a smallsubpopulation of stem-like cells give rise to the differ-entiated cells that make the bulk of the tumour (Al-Hajjet al., 2003). Cancer stem cells (CSCs) occur as mimeticof normal tissue stem cells based on germ-line variation,
epigenetic change and somatic mutation of stem cellsignalling components, and then acquire more malig-nant phenotype based on accumulation of additionalepigenetic and genetic alterations, and tumour-stromalinteraction at the invasion front (Katoh, 2010). How-ever, the target cell for transformation that originatesCSCs remains an unknown issue. In solid tumours thephenotypic definition of CSCs is rather loose with nosingle marker that unequivocally identifies that popula-tion. Such poor definition has prevented so far theneeded study of clinical relevance of CSC content intumour management. There are a number of reportsusing in vitro culture of tumour cells and animal modelsshowing that CSCs are more resistant to conventionalcancer therapies, therefore, placing these cells at the rootof tumour recurrence and metastases. Several prelimin-ary reports have indeed shown that this is the casewith human cancer patients. In breast cancer, Li et al.(2008) showed that conventional chemotherapy in-creased the fraction of CD44!CD24" cells in aneoadjuvant setting of advanced breast cancer patientsand increased mammosphere formation in vitro. Taneiet al. (2009) have shown that paclitaxel and epirubicin-based chemotherapy enriches for aldehyde dehydrogen-ase-1-positive cells in breast tumours, another markerfor CSCs (Ginestier et al., 2007). In an attempt toassociate CSC content and prognosis, the presence ofCD44!CD24" cells in clinical samples of breast cancerwas not associated with aggressive tumour behaviourand poor clinical outcome (Abraham et al., 2005),indicating that the definition of CSCs must be firstclearly established before it can have clinical use. It ispossible that different breast cancer subtypes rely onspecific CSC populations and, therefore, more precisefunctional identification must be developed.
Reprogramming of adult terminally differentiatedhuman cells into pluripotent stem cells capable ofgenerating all cells in the body has attracted a greatdeal of attention. The production of these inducedpluripotent stem cells requires the over-expression offour transcription factors Oct4, Sox2, Klf4 and c-Myc(Takahashi et al., 2007), although Klf4 and c-Myc canbe replaced by Lin28 and Nanog, and may even bedispensable (Meissner et al., 2007; Nakagawa et al.,2008; Okita et al., 2008; Kim et al., 2009). The efficiencyof this reprogramming process is extremely low andremains so far as an in vitro phenomenon as thereis no evidence that it can naturally occur in vivo.Received 17 January 2011; revised 16 June 2011; accepted 3 July 2011
Correspondence: Dr AG Martin, Regulation of Cell GrowthLaboratory, Fundacion Inbiomed, Paseo Mikeletegi 61, San Sebastian,Gipuzkoa 20009, Spain.E-mail: [email protected]
Oncogene (2011) 1–12& 2011 Macmillan Publishers Limited All rights reserved 0950-9232/11
www.nature.com/onc

The mechanisms underlying the reprogramming processare not well understood yet, however, the three maintranscription factors Oct4, Sox2 and Nanog, calledmaster regulators of pluripotency, have probed respon-sible for maintaining the undifferentiated state (Yama-naka, 2008). Recently, the process of reprogrammingand tumourigenesis (Kawamura et al., 2009; Li et al.,2009; Marion et al., 2009; Utikal et al., 2009) have beenlinked as the p53 tumour suppressor, one of the mainregulators of oncogenic transformation, controls theinduction of pluripotency.
Sox genes encode a family of high-mobility grouptranscription factors that have critical roles in organo-genesis. The functional specificity of different Soxproteins and the tissue specificity of a particular Soxfactor are largely determined by the differential partner-ship with other transcription regulators, many of whichhave not yet been discovered. Recently, a number oflinks have been found between the Sox2 and humancancers. Sox2 has been found to be an immunogenicantigen in 41% of small cell lung cancer patients (Gureet al., 2000) and in 29% of meningioma patients(Comtesse et al., 2005). Immunohistochemistry resultssuggest that Sox2 is involved in invasion and metastasisof pancreatic intraepithelial neoplasia (Sanada et al.,2006), and may also be involved in gastric carcinogen-esis (Li et al., 2004) and prostate cancers (Sattler et al.,2000). Furthermore, Sox2 expression has been observedin 43% of basal cell-like breast carcinomas, suggesting arole in conferring a less differentiated phenotype(Rodriguez-Pinilla et al., 2007). How Sox2 exerts itsoncogenic potential is currently unknown.
In this work we investigated the relationship betweenpluripotency induction and breast cancer by examiningthe presence of pluripotency-related genes in humanbreast tumours. We did not observe expression of Oct4or Nanog but we found expression of Sox2, which inturn was associated with early stages of tumourformation. We observed induction of Sox2 expressionin tumour spheres from natural breast tumour culturesand breast carcinoma cell lines, suggesting a role intumour stem cells. This role was further investigated byover-expressing and downregulating Sox2 expression,and it was found that Sox2 was necessary and sufficientto induce tumour sphere formation and tumour initia-tion in vivo. Regulation of Sox2 promoter activationprobed to be dependent on the upstream enhancer andnot on the core promoter, the same element thatcontrols Sox2 expression in pluripotent stem cells.
Results
Expression of pluripotency-associated genes in naturalbreast tumoursOct4, Sox2 and Nanog are considered the masterregulator genes of pluripotency and are switched off inadult tissues, except for Sox2 that is expressed in neuralprogenitors (Zappone et al., 2000). However, reexpres-sion of some or all of these genes has been found in
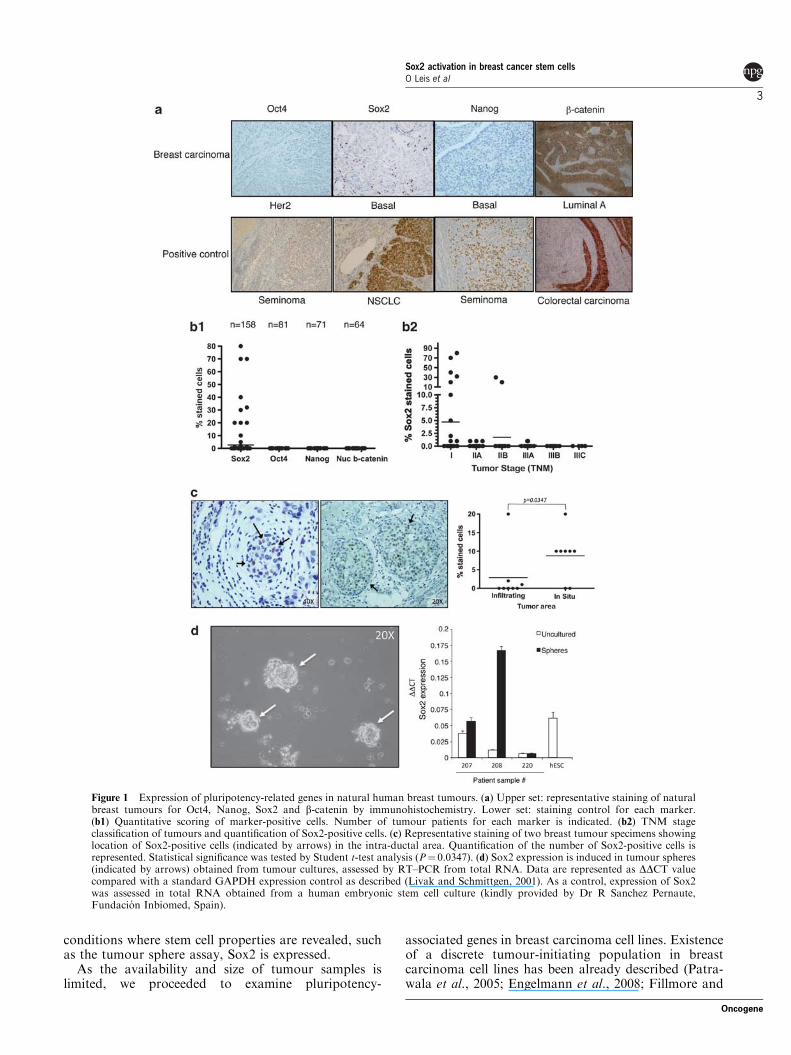
several forms of human cancer. Using immunohisto-chemistry we checked expression of these genes in apanel of human breast tumour samples representative ofthe main breast cancer subtypes (Figure 1a). Weincluded staining for b-catenin as activation of thecanonical Wnt signalling pathway, measured by nuclearlocalization of b-catenin, has been linked to mammaryorganogenesis (Zeng and Nusse, 2010). For positivecontrols, we used sections of seminoma (for Oct4 andNanog), non-small cell lung carcinoma (for Sox2) andcolorectal carcinoma (for nuclear b-catenin). We did notobserve expression of Oct4 or Nanog in any sample ofour series (n# 81 for Oct4 and n# 71 for Nanog), nornuclear b-catenin localization (n# 64). However, we didobserve Sox2-positive staining in B16% of the samplesobserved (n# 158; Figure 1b1). Sox2 expression waspreviously reported in basal type breast cancer (Ro-driguez-Pinilla et al., 2007); in our tumour series weobserved Sox2-positive staining in tumour samples of allthe major breast cancer subtypes (Table 1). For acomplete description of tumour features see Supplemen-tary File 1. When tumours were classified according totheir TNM stage, Sox2-positive tumours fell into theearly stages of tumour progression: 57.9% in stage I,26.3% in stage IIA, 10.5% in stage IIB, 5.3% in stageIIIA and no Sox2-positive tumours in more advancedstages (IIIB or beyond; Figure 1b2). Out of the 25 Sox2-positive tumours found, 13 showed infiltrating tissuestructure by histological examination, while 12 tumoursstill retained in situ intra-ductal tumour component(Figure 1c). Interestingly, those tumours that retainedintra-ductal tumour component showed significantly(P# 0.0347 by paired t-test) higher numbers of Sox2-positive cells there than in the infiltrating part, suggest-ing the presence of Sox2-positive cells in primitive areasof the tumour. Collectively, these data indicate thatSox2 may be expressed in the initial stages of tumourformation and it is lost as the tumour progressestowards advanced stages.
Sox2 is expressed in tumour spheresOwing to the relevant role of Sox2 in pluripotencymaintenance, we reasoned that Sox2 expression mightbe related to the presence of CSCs. We utilized thetumour sphere assay as a surrogate in vitro culture assayfor tumour formation to investigate Sox2 expression inCSCs. In these culture conditions, CSCs are able tomaintain stem-like properties such as tumour initiation,self-renewal and limited differentiation potential (Dontuet al., 2003; Ponti et al., 2005). In our hands, B40% oftumour specimens produced viable sphere cultures witha frequency of sphere forming cells ranging between0.1% and 1% of the total cell suspension (data notshown). We examined the expression of Sox2 inmammospheres from three tumour specimens usingreal-time PCR (RT–PCR). As seen in Figure 1d, Sox2expression was observed in two out of three spherecultures (specimens #207 and #208). We did not detectSox2 expression in specimen #220 in uncultured tumourcells nor in tumour spheres. This result suggests that in
Sox2 activation in breast cancer stem cellsO Leis et al
2
Oncogene
conditions where stem cell properties are revealed, suchas the tumour sphere assay, Sox2 is expressed.
As the availability and size of tumour samples islimited, we proceeded to examine pluripotency-
associated genes in breast carcinoma cell lines. Existenceof a discrete tumour-initiating population in breastcarcinoma cell lines has been already described (Patra-wala et al., 2005; Engelmann et al., 2008; Fillmore and
Figure 1 Expression of pluripotency-related genes in natural human breast tumours. (a) Upper set: representative staining of naturalbreast tumours for Oct4, Nanog, Sox2 and b-catenin by immunohistochemistry. Lower set: staining control for each marker.(b1) Quantitative scoring of marker-positive cells. Number of tumour patients for each marker is indicated. (b2) TNM stageclassification of tumours and quantification of Sox2-positive cells. (c) Representative staining of two breast tumour specimens showinglocation of Sox2-positive cells (indicated by arrows) in the intra-ductal area. Quantification of the number of Sox2-positive cells isrepresented. Statistical significance was tested by Student t-test analysis (P# 0.0347). (d) Sox2 expression is induced in tumour spheres(indicated by arrows) obtained from tumour cultures, assessed by RT–PCR from total RNA. Data are represented as DDCT valuecompared with a standard GAPDH expression control as described (Livak and Schmittgen, 2001). As a control, expression of Sox2was assessed in total RNA obtained from a human embryonic stem cell culture (kindly provided by Dr R Sanchez Pernaute,Fundacion Inbiomed, Spain).
Sox2 activation in breast cancer stem cellsO Leis et al
3
Oncogene
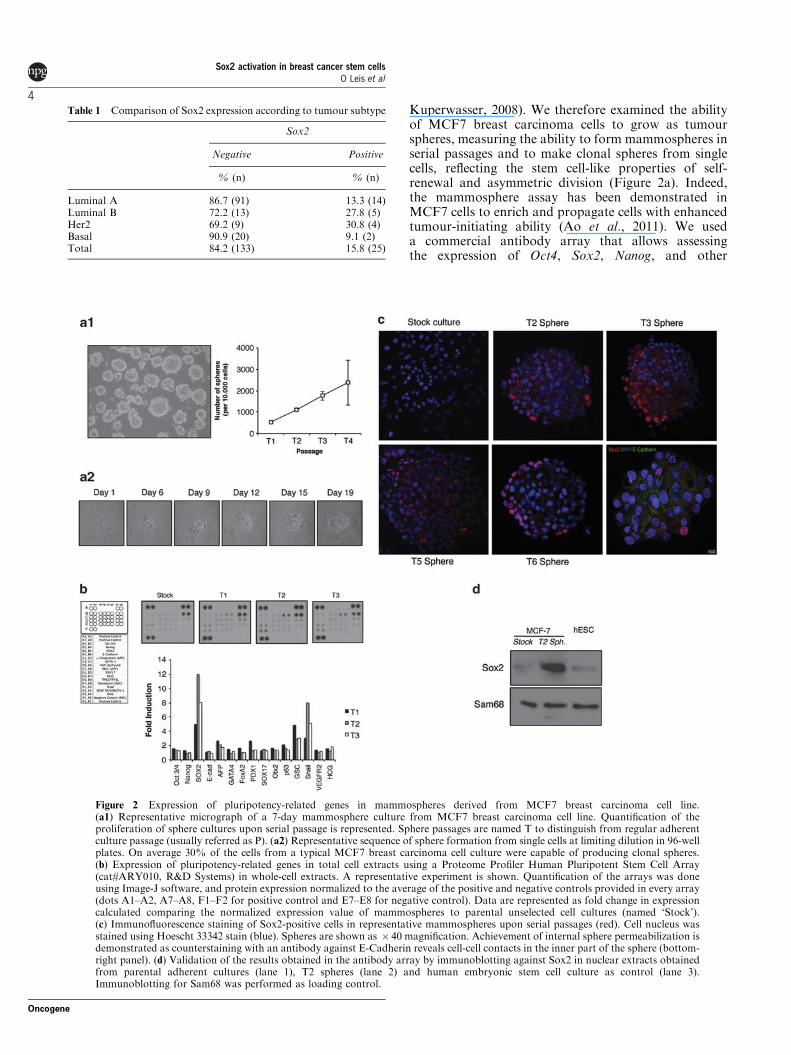
Kuperwasser, 2008). We therefore examined the abilityof MCF7 breast carcinoma cells to grow as tumourspheres, measuring the ability to form mammospheres inserial passages and to make clonal spheres from singlecells, reflecting the stem cell-like properties of self-renewal and asymmetric division (Figure 2a). Indeed,the mammosphere assay has been demonstrated inMCF7 cells to enrich and propagate cells with enhancedtumour-initiating ability (Ao et al., 2011). We useda commercial antibody array that allows assessingthe expression of Oct4, Sox2, Nanog, and other
Table 1 Comparison of Sox2 expression according to tumour subtype
Sox2
Negative Positive
% (n) % (n)
Luminal A 86.7 (91) 13.3 (14)Luminal B 72.2 (13) 27.8 (5)Her2 69.2 (9) 30.8 (4)Basal 90.9 (20) 9.1 (2)Total 84.2 (133) 15.8 (25)
Figure 2 Expression of pluripotency-related genes in mammospheres derived from MCF7 breast carcinoma cell line.(a1) Representative micrograph of a 7-day mammosphere culture from MCF7 breast carcinoma cell line. Quantification of theproliferation of sphere cultures upon serial passage is represented. Sphere passages are named T to distinguish from regular adherentculture passage (usually referred as P). (a2) Representative sequence of sphere formation from single cells at limiting dilution in 96-wellplates. On average 30% of the cells from a typical MCF7 breast carcinoma cell culture were capable of producing clonal spheres.(b) Expression of pluripotency-related genes in total cell extracts using a Proteome Profiler Human Pluripotent Stem Cell Array(cat#ARY010, R&D Systems) in whole-cell extracts. A representative experiment is shown. Quantification of the arrays was doneusing Image-J software, and protein expression normalized to the average of the positive and negative controls provided in every array(dots A1–A2, A7–A8, F1–F2 for positive control and E7–E8 for negative control). Data are represented as fold change in expressioncalculated comparing the normalized expression value of mammospheres to parental unselected cell cultures (named ‘Stock’).(c) Immunofluorescence staining of Sox2-positive cells in representative mammospheres upon serial passages (red). Cell nucleus wasstained using Hoescht 33342 stain (blue). Spheres are shown as $ 40 magnification. Achievement of internal sphere permeabilization isdemonstrated as counterstaining with an antibody against E-Cadherin reveals cell-cell contacts in the inner part of the sphere (bottom-right panel). (d) Validation of the results obtained in the antibody array by immunoblotting against Sox2 in nuclear extracts obtainedfrom parental adherent cultures (lane 1), T2 spheres (lane 2) and human embryonic stem cell culture as control (lane 3).Immunoblotting for Sam68 was performed as loading control.
Sox2 activation in breast cancer stem cellsO Leis et al
4
Oncogene

pluripotency-associated genes in a single test (Figure 2b)to compare protein extracts obtained from serialpassages of tumour spheres. Extracts from humanembryonic stem cells (kindly provided by R. SanchezPernaute, Inbiomed, Spain) were used as control whereexpression of Oct4, Nanog and Sox2 was readilydetected (data not shown). We did not observe expres-sion of Oct4, Nanog or Sox2 in the stock controlpopulation, however, induction of Sox2 protein but notOct4 or Nanog upon sphere formation was observed.Detection of Sox2 by immunoblotting in nuclearextracts from mammospheres confirmed the resultsobtained with the antibody array (Figure 2d). Theseresults are consistent with the observations we obtainedusing the fresh breast tumour specimens.
Next we stained mammospheres by immunofluores-cence in different passages to locate Sox2-positive cells(Figure 2c). We did not observe Sox2 staining fromparental adherent cultures despite detecting positiveexpression in control extracts by immunoblotting, likelydue to insufficient sensitivity of the immunofluorescencetechnique. We observed that only some of the cells in thetumour spheres stained positive for Sox2 (2.9% for T1,5.7% for T2, 17.7% for T3, 12.1% for T4, 24.7% for T5and 32.0% for T6) and those cells located preferentiallyto the periphery of the spheres. This effect was not dueto insufficient access of the antibody to the centre of thespheres, as the inner cells were readily detected byE-Cadherin staining (Figure 2c lower right panel).Therefore, it is tempting to speculate that those fewSox2-positive cells represent the CSCs and/or theirimmediate progeny. Essentially, the same results wereobtained using T47D cells, a different breast carcinomacell line (Supplementary Figure 1).
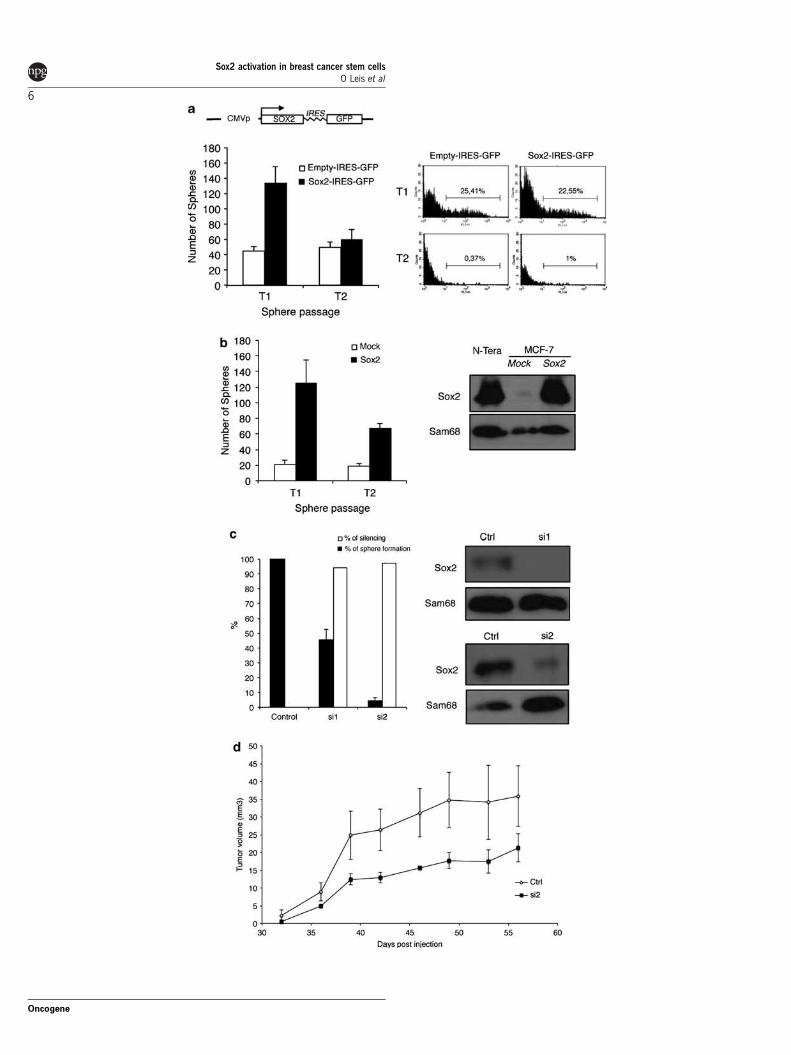
Sox2 is necessary and sufficient for sphere formationIn order to test whether Sox2 was conferring stem cellproperties to the breast carcinoma cells, we used thepCAGSS-Sox2-GFP construct to transiently expressSox2 in MCF7 cells. This vector allows the monitoringof plasmid activity by expression of the green fluorescentprotein controlled by an IRES element. Expression ofSox2 induced the generation of approximately three-foldmore spheres than the empty vector control (Figure 3a).Interestingly, this effect was reversible and contingentto the activity of the plasmid, suggesting that the ectopicexpression of Sox2 was responsible for the increase insphere number. To test this possibility, we generatedMCF7 cells with stable expression of Sox2 throughlentiviral transfer (mCitrinie-P2A-Sox2 lentiviral vector)and observed a marked increase in sphere formationcompared with mock-infected cells and those main-tained in subsequent sphere passages (Figure 3b).We then used RNA interference silencing with double-stranded RNA oligonucleotides (s13294 -si1-, s13295-si2- and controls, Applied Biosystems, Carlsbad, CA,USA) to knockdown Sox2 expression for testing whetherSox2 expression was necessary to induce tumour spheres.Over 90% silencing was achieved with both siRNAsused, as confirmed by immunoblotting (Figure 3c).
When tested for mammosphere formation, we observedan evident reduction of sphere formation compared withcontrol siRNA (50% reduction for -si1- and almostcompete blockage for -si2-) indicating that Sox2expression is necessary for mammosphere formation.We then tested the tumour initiation ability of MCF7cells silenced for Sox2 expression in mouse xenograftassays (Figure 3d and Supplementary Figure 2). Asobserved in Figure 3d, silencing of Sox2 expression withsiRNA reduced the size of the tumours produced, ascompared with siRNA control cells, indicating thatSox2 facilitates tumour initiation.
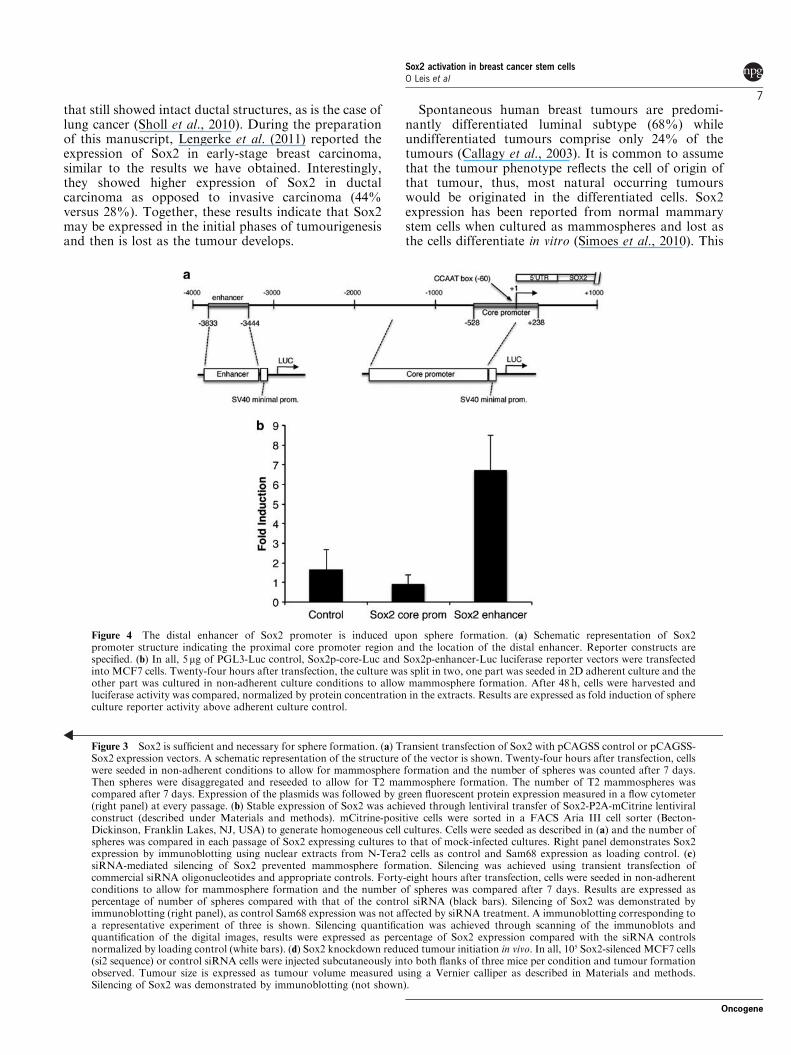
Sox2 promoter is induced upon sphere formationRegulation of Sox2 expression is poorly understood. Anarea located between positions "528 and ! 238 fromthe transcription start site is considered as the coreproximal promoter region (Miyagi et al., 2006).Additionally, an upstream enhancer centred between"3444 and "3833 has an active role in controllingexpression of Sox2 in the reprogramming of oligoden-drocyte precursors (Kondo and Raff, 2004) and inpluripotent stem cells (Tomioka et al., 2002; seeFigure 4a for a schematic representation of the Sox2promoter). We transfected MCF7 and T47D cells withluciferase reporter vectors for the upstream distalenhancer and the core promoter to test whetherinduction of Sox2 protein was achieved throughtranscriptional activation of Sox2 promoter. As seen inFigure 4b and Supplementary Figure 2, the Sox2 corepromoter region reporter was not significantly activatedupon sphere formation whereas the distal enhancerregion was strongly activated (seven-fold on average).Taken together, these results suggest that reactivation ofSox2 expression in breast carcinoma cells upon sphereformation may be controlled at the promoter level,similarly as it is activated in pluripotent stem cells.
Discussion
The presence of CSCs in breast tumours is likely one ofthe main reasons why current oncologic therapies arepoorly effective in preventing tumour progression,metastasis and recurrence (Shafee et al., 2008; Taneiet al., 2009; Cirenajwis et al., 2010), so elimination ofCSCs may become a necessary step for an effective cure.The target of malignant transformation could be a tissuestem cell, a progenitor cell, or a terminally differentiatedcell that acquires, through mutations and epigeneticchanges, the stem cell self-renewing property. Toinvestigate this issue we examined the presence ofstem-like cells by assessing the expression of the majorstem cell pluripotency regulators, Oct4, Sox2 and Nanog(Jaenisch and Young, 2008), in human natural breasttumours. We observed that Sox2 is expressed at theprotein level in breast tumours, as expected (Rodriguez-Pinilla et al., 2007; Chen et al., 2008). However, Sox2expression was more prevalent in the early stages oftumour development and in the ductal areas of tumours
Sox2 activation in breast cancer stem cellsO Leis et al
5
Oncogene
Sox2 activation in breast cancer stem cellsO Leis et al
6
Oncogene
that still showed intact ductal structures, as is the case oflung cancer (Sholl et al., 2010). During the preparationof this manuscript, Lengerke et al. (2011) reported theexpression of Sox2 in early-stage breast carcinoma,similar to the results we have obtained. Interestingly,they showed higher expression of Sox2 in ductalcarcinoma as opposed to invasive carcinoma (44%versus 28%). Together, these results indicate that Sox2may be expressed in the initial phases of tumourigenesisand then is lost as the tumour develops.
Spontaneous human breast tumours are predomi-nantly differentiated luminal subtype (68%) whileundifferentiated tumours comprise only 24% of thetumours (Callagy et al., 2003). It is common to assumethat the tumour phenotype reflects the cell of origin ofthat tumour, thus, most natural occurring tumourswould be originated in the differentiated cells. Sox2expression has been reported from normal mammarystem cells when cultured as mammospheres and lost asthe cells differentiate in vitro (Simoes et al., 2010). This
Figure 3 Sox2 is sufficient and necessary for sphere formation. (a) Transient transfection of Sox2 with pCAGSS control or pCAGSS-Sox2 expression vectors. A schematic representation of the structure of the vector is shown. Twenty-four hours after transfection, cellswere seeded in non-adherent conditions to allow for mammosphere formation and the number of spheres was counted after 7 days.Then spheres were disaggregated and reseeded to allow for T2 mammosphere formation. The number of T2 mammospheres wascompared after 7 days. Expression of the plasmids was followed by green fluorescent protein expression measured in a flow cytometer(right panel) at every passage. (b) Stable expression of Sox2 was achieved through lentiviral transfer of Sox2-P2A-mCitrine lentiviralconstruct (described under Materials and methods). mCitrine-positive cells were sorted in a FACS Aria III cell sorter (Becton-Dickinson, Franklin Lakes, NJ, USA) to generate homogeneous cell cultures. Cells were seeded as described in (a) and the number ofspheres was compared in each passage of Sox2 expressing cultures to that of mock-infected cultures. Right panel demonstrates Sox2expression by immunoblotting using nuclear extracts from N-Tera2 cells as control and Sam68 expression as loading control. (c)siRNA-mediated silencing of Sox2 prevented mammosphere formation. Silencing was achieved using transient transfection ofcommercial siRNA oligonucleotides and appropriate controls. Forty-eight hours after transfection, cells were seeded in non-adherentconditions to allow for mammosphere formation and the number of spheres was compared after 7 days. Results are expressed aspercentage of number of spheres compared with that of the control siRNA (black bars). Silencing of Sox2 was demonstrated byimmunoblotting (right panel), as control Sam68 expression was not affected by siRNA treatment. A immunoblotting corresponding toa representative experiment of three is shown. Silencing quantification was achieved through scanning of the immunoblots andquantification of the digital images, results were expressed as percentage of Sox2 expression compared with the siRNA controlsnormalized by loading control (white bars). (d) Sox2 knockdown reduced tumour initiation in vivo. In all, 105 Sox2-silenced MCF7 cells(si2 sequence) or control siRNA cells were injected subcutaneously into both flanks of three mice per condition and tumour formationobserved. Tumour size is expressed as tumour volume measured using a Vernier calliper as described in Materials and methods.Silencing of Sox2 was demonstrated by immunoblotting (not shown).
Figure 4 The distal enhancer of Sox2 promoter is induced upon sphere formation. (a) Schematic representation of Sox2promoter structure indicating the proximal core promoter region and the location of the distal enhancer. Reporter constructs arespecified. (b) In all, 5mg of PGL3-Luc control, Sox2p-core-Luc and Sox2p-enhancer-Luc luciferase reporter vectors were transfectedinto MCF7 cells. Twenty-four hours after transfection, the culture was split in two, one part was seeded in 2D adherent culture and theother part was cultured in non-adherent culture conditions to allow mammosphere formation. After 48 h, cells were harvested andluciferase activity was compared, normalized by protein concentration in the extracts. Results are expressed as fold induction of sphereculture reporter activity above adherent culture control.
Sox2 activation in breast cancer stem cellsO Leis et al
7
Oncogene

assay is used as a stem-like functional assay that allowsthe propagation of mammary epithelial and breasttumour cells in an undifferentiated state based on theirability to proliferate in suspension (Dontu et al., 2003;Ponti et al., 2005; Farnie et al., 2007) and has been usedfor cancer stem-like specific drug screening in MCF7cells (Zhou et al., 2007; Ao et al., 2011). We did observeinduction of Sox2 expression in mammospheres ob-tained from human breast tumour cell cultures, andfrom MCF7 and T47D cells. Moreover, we did notobserve expression of any of the other pluripotency-associated genes in MCF7-derived spheres, consistentlywith the results obtained in human tumours. Despitesome reports claiming expression of Oct4 and Nanog inbreast carcinoma cell lines, expression of severalpseudogenes have been reported for both Oct4 andNanog (Suo et al., 2005; Zhang et al., 2006; Cantz et al.,2007; Ambady et al., 2010), therefore, particular caremust be taken to avoid PCR amplification of thosepseudogenes. We base our conclusions in antibody-basedassays, thus, looking specifically at the protein level toavoid pseudogene detection. As opposed to spheresderived from glioma cell lines (Ghods et al., 2007), only afew cells in the mammospheres were stained positivefor Sox2, thus it is tempting to speculate that thetumour-initiating property resides in those cells. Furtherexperiments will be needed to specifically isolate Sox2-positive cells and test their tumour generating ability.
In order to verify the contribution of Sox2 tomammosphere formation, we over-expressed Sox2 andobserved that Sox2 expression increased the ability togenerate spheres. Interestingly, this effect was dependenton the continuous expression of Sox2 indicating that Sox2expression was sufficient to induce this stem-like feature.Knockdown of Sox2 expression prevented the formationof spheres; thus, Sox2 expression was not only sufficientbut also necessary to form spheres. Furthermore, whenSox2-silenced MCF7 cells were tested for tumourinitiation in mouse xenograft models, we observed amarked reduction in the size of the tumours formed,suggesting that Sox2 participates in tumour initiationin vivo. This result is consistent with data reported onglioblastoma where Sox2 knockdown prevents gliomastem cells to initiate tumours (Gangemi et al., 2009).
The process of in vitro pluripotency induction presentsseveral parallelisms with tumour generation. Fivegroups independently (Hong et al., 2009; Kawamuraet al., 2009; Li et al., 2009; Marion et al., 2009; Utikalet al., 2009) observed that p53 poses a barrier forpluripotency induction. Mutations that inactivate thep53 tumour suppressor network occur in most naturalhuman tumours, thus, remarkable similarities must existbetween the genetic and epigenetic processes involved inthe generation of cancer, and the acquisition of a lessdifferentiated phenotype and self-renewal ability ininduced pluripotency. Loss of p53 confers stem-likeproperties in committed progenitors in the haemato-poietic system (Akala et al., 2008), and loss of p53increases the size of the mammary repopulatingcompartment in mice by 10-fold, suggesting that p53may act as a regulator of stem cell self-renewal in vivo
(Cicalese et al., 2009). Therefore, it is possible that theacquisition of the self-renewal ability of CSCs arises bysome of the mechanisms shared with those involved inpluripotency reprogramming, in cells that harbourpermissive mutations, such as inactivation of the p53pathway. This mechanism explains the generation ofCSCs from committed progenitors with limited differ-entiation potential responsible for the heterogeneoussubtypes observed in natural tumours. We did notobserve induction of Oct4 or Nanog, therefore, a fullpluripotency programme cannot be established. Theability of non-stem cells to convert to CSCs impliesintrinsic plasticity at the level of the cell of origin of thetumours. In this regard, transformation of an immorta-lized breast epithelial cell line (MCF10A) by theoncogene Src generated cells with features of CSCs onlywhen interleukin-6 was added to the medium, suggestingthat CSCs can be originated from non-CSC transformedcells (Iliopoulos et al., 2011). Similar results wereobtained attending to in vitro culture of floating humanmammary epithelial cells, where cells with stem-likefeatures arise from FACS (fluorescence-activated cellsorting)-sorted differentiated epithelial cells (Chafferet al., 2011). We propose that spontaneous dediffer-entiation in vivo may involve reactivation of one or morepluripotency-associated factors, such as Sox2 expressionin breast cancer, consistent with our observations. Whatthe mechanisms for reactivation are, how are theydifferent from a full pluripotency programme and howcontextual signals from the microenvironment (such asinduction of EMT (Mani et al., 2008) participate in thisprocess, will shed light into how tumours are initiatedand how to control this process.
How Sox2 expression is reactivated remains unclear.Sox2 promoter consists of two main regulatory regions,the core promoter located at the transcription initiationsite and an upstream distal enhancer. Our resultsindicate that it is precisely this upstream distal enhancerthat is activated in mammospheres, as a luciferasereporter with this enhancer is strongly induced uponsphere formation. The upstream enhancer was stronglyactivated in undifferentiated embryonic stem cells andits activity was lost upon differentiation (Tomioka et al.,2002), it is the same behaviour as observed in multi-potent neural stem cells (Miyagi et al., 2004; Miyagiet al., 2006). In embryonic stem cells, a non-consensusoctamer-binding site in the enhancer is occupied byOct3/4-Sox2 dimers, however, as we did not observeOct4 expression in mammospheres, a different mechan-ism must operate. Recently, a possible implication of thetranscription factor NF (nuclear factor)-kB has beenproposed, although no direct regulation was reported(Liu et al., 2010). Kondo and Raff (Kondo and Raff,2004) demonstrated that Sox2 reactivation in theconversion of oligodendrocyte precursors depends onthe recruitment of the protein Brca1 to this enhancer,together with the chromatin remodelling protein Brm.These complexes have a critical role in the maintenanceof the pluripotent state by regulating occupancy ofpluripotency-related gene promoters (Kidder et al.,2009). As Brca1 is a well-known tumour suppressor
Sox2 activation in breast cancer stem cellsO Leis et al
8
Oncogene

gene that when mutated predisposes women to breastand ovarian cancer (Miki et al., 1994), a possibility isthat loss of Brca1 mediates tumour initiation by losingthe control on Sox2 expression, establishing a direct linkbetween Sox2 expression and breast cancer initiation.Nevertheless, this model needs further testing.
Materials and methods
Tumour sample collectionA total of 158 cases of breast tumour samples were collected atthe Pathology Department of Onkologikoa (San Sebastian,Spain) from patients that underwent curative surgical resection(for tumour data see Supplementary File 1). Institutionalreview board approval and expressed informed consent wereobtained from all patients before sample collection. Formalin-fixed paraffin-embedded sections of tumours were stainedroutinely with haematoxylin-eosin and reviewed by twosenior pathologists (RR and KE) to determine the histologicaltype according to WHO breast carcinoma histologicalclassification criteria (2003), and clinical stage according tothe UICC TNM classification (2003). Tumours were sub-divided as: basal-like (ER, PR and HER2-negative and CK5/6and/or EGFR-positive), luminal A (ER and/or PR-positiveand HER2-negative), luminal B (ER and/or PR-positive andHER2-positive) and HER2-positive subgroups. The data andtumour samples used were provided by the Basque Biobankfor Research (http://www.basquebiobank.org), supported byBasque Foundation for Health Innovation and Research.
Cell cultureFresh tumour samples were cut up into small pieces and mincedcompletely using scissors. Tumour pieces were enzymaticallydigested with Collagenase IA (Sigma, St Louis, MO, USA) atthe concentration of 2mg/ml and incubated at 37 1C for 1h.Then, the digested sample was filtered through a 40-mmcell strainer and washed with Hank’s balanced salt solution.For sphere formation, cells were cultured at an initial densityof 50 000 cells/ml in serum-free Dulbecco’s modied Eagle’smedium supplemented with 1% L-glutamine, 1% penicillin/streptomycin, 30% F12 (Sigma), 2% B27 (Invitrogen, Carlsbad,CA, USA), 20ng/ml EGF (Sigma) and 20ng/ml FGFb(Invitrogen) in non-adherent culture plates. For subsequentsphere formation, cells were seeded at a density of 5 000 cells/ml.MCF-7 and T47D breast carcinoma cell lines were obtaineddirectly from the ATCC (Manassas, VA, USA) and grown inDulbecco’s modied Eagle’s medium (Gibco, Carlsbad, CA,USA) supplemented with 10% fetal bovine serum (Sigma) at37 1C in a 5% CO2 incubator. For mammosphere formation,single-cell suspensions were plated in six-well tissue cultureplates covered with poly-2-hydroxyethyl-methacrylate (Sigma)to prevent cell attachment at a density of 1000 cells/ml inthe medium described above. The medium was made semi-solidby the addition of 0.5% Methylcellulose (R&D Systems,Minneapolis, MN, USA) to prevent cell aggregation. Mammo-spheres were collected by gentle centrifugation after 7 days, anddissociated enzymatically (5min in 1:1 trypsin/Dulbecco’smodied Eagle’s medium solution at 37 1C) and mechanicallyby passing through a 25G needle. Single cells were replated at adensity of 1000 cells/ml for subsequent passages.
Plasmids and antibodiespCAGSS-Sox2 and empty pCAGSS control expression vectorswere kindly provided by A Matheu (Medical Research
Council, London, UK). For immunoblotting, immunofluores-cence and immunohistochemistry anti-human Sox2 polyclonalantibody (Neuromics GT15098), anti-human Oct4 polyclonalantibody (Santa Cruz sc-8629, Santa Cruz, CA, USA), anti-human Nanog polyclonal antibody (R&D Systems AF1997),anti-human b-catenin monoclonal antibody (Millipore 05665,Billerica, MA, USA), anti-human E-cadherin polyclonal anti-body (Cell Signaling FAB18381A, Danvers, MA, USA) andanti-human Sam68 polyclonal antibody (Santa Cruz sc-333)were used as indicated.
ImmunohistochemistryImmunohistochemical staining on tumour sections was per-formed using Dako Autostainer automatic universal slidestainer. Antibodies, dilutions, antigen-retrieval methods andscoring systems for ER, PR, HER2, CK5/6, EGFR and ki67are part of the routine histopathological determination at thePathology Department of Onkologikoa (San Sebastian,Spain). Cases were considered positive for Sox2 when anyunequivocal neoplastic cell displayed definite nuclear staining.Positive (as indicated) and negative controls were included ineach slide run.
Real-time PCRTotal RNA was obtained using RNAqueous-Micro kit (Am-bion, Austin, TX, USA) following manufacturer’s instructions.Power SYBER Green PCR Master Mix (Applied Biosystems)was used to amplify the corresponding genes with primers forhuman Sox2 (forward: 50-GGGAAATGGGAGGGGTGCAAAAGAGG-30, reverse: 50-TTGCGTGAGTGTGGATGGGGATTGGTG-30). Amplification of human GAPDH (Hs_GAPDH_2_SG QuantiTect Primer Assay QT01192646, Qiagen,Hilden, Germany) was used as loading control. RT–PCR wasperformed in a StepOnePlus Real-Time PCR System.
ImmunofluorescenceMammospheres were collected by centrifugation in aCytospin (Thermo, Waltham, MA, USA) onto glass slides.Slides were fixed in 4% paraformaldehyde for 1 h at roomtemperature in wet chamber, washed twice in phosphate-buffered saline (PBS) and allowed to permeabilize for 1 h atroom temperature with PBS ! 0.3% Triton X-100 andblocked. Slides were then incubated with primary antibodiesovernight and washed thrice in PBS, and appropriatefluorofore labelled secondary antibodies were added at adilution of 1/250. Hoescht 33342 dye (Sigma, St Louis, MO,USA) was used to reveal nuclear DNA. Immunofluorescencewas visualized in a Zeiss LSM510 confocal laser-scanningmicroscope (Zeiss, Jena, Germany).
ImmunoblottingNuclear extracts (prepared as in Martin et al. (2001)) wereblotted onto a polyvinylidene fluoride membrane (Millipore)and blocked in Tris-buffered saline with 0.2% Tween 20 and5% milk powder. Immunoblotting for Sox2 and Sam68 wascarried out using primary antibodies at a 1:2000 dilution andappropriate secondary antibody coupled to horseradishperoxidase (1:10 000 dilution). SuperSignal West Pico Chemi-luminescent Substrate developing kit (Thermo) was used toreveal the relevant bands.
Transfection, gene silencing and lentiviral gene transferFor transient transfection, MCF-7 cells were cultured in six-well tissue culture plates and transfected when at 70%confluence using Lipofectamine Plus or Oligofectamine
Sox2 activation in breast cancer stem cellsO Leis et al
9
Oncogene

(Invitrogen) according to manufacturer’s instructions. Genesilencing experiments were performed using specific commer-cial siRNA sequences for human Sox2 (s13294 and s13295,Applied Biosystems), and appropriate siRNA controls. Forstable expression of Sox2, lentiviral transfer was used usingthe mCitrinie-P2A-Sox2 lentiviral vector (kindly provided byE Papapetrou, Memorial Sloan-Kettering Cancer Center, NY,USA; Papapetrou et al., 2009).
Mouse xenograft assaysFemale 6-week-old athymic nude mice (Balb/c Nu/Nu) werepurchased from Charles River, and were housed in specificallydesigned pathogen-free isolation animal facility. All animalprocedures were performed in accordance with institutionalanimal care and use guidelines. For injection, MCF7 cells ofeach condition were resuspended in 10ml of the same media ata concentration of 1$ 106 cells/ml. Mice were subcutaneouslyinoculated with the MCF7 cell line in 200 ml of culture mediumwith matrigel in both mammary fat pads. In all, 105 MCF7cells with Sox2 knockdown were inoculated in the rightmammary fat pad, with their respective control in the leftmammary fat pad. Mice were weighed and the inoculation siteswere inspected by palpation at weekly intervals. Whentumours become detectable manually, the growth rates weredetermined by weekly measurement of two diameters of thetumour with a Vernier calliper. The tumour volume wasestimated as the volume of an ellipse using the followingformula: V# 4/3$ (a/2)$ (b/2)2, where ‘a’ and ‘b’ correspondto the longest and shortest diameter, respectively. Animalswere euthanized when their tumours were harvested, fixed andembedded in paraffin.
Luciferase reporter assaysMCF-7 cells were transfected with 5 mg of pGL3 Luc control(Promega, Madison, WI, USA), pGL3-Sox2p-enhancer-Luc(kindly provided by T Kondo, RIKEN, Japan) or pGL3-Sox2p-core-Luc (kindly provided by JM Sanchez-Puelles, CIB,Madrid, Spain) reporter plasmids using Lipofectamine Plus(Invitrogen). Luciferase activity was measured in duplicatewith the Glomax 20/20 luminometer (Promega) and the resultswere expressed as fold induction above control.
Conflict of interest
The authors declare no conflict of interest.
Acknowledgements
The Regulation of Cell Growth Laboratory is supported bygrants from Obra Social Kutxa, Fundacion Medica MutuaMadrilena, Gobierno Vasco (Saiotek program and Consejerıade Educacion PI2010-25) and Instituto de Salud Carlos IIIAccion Estrategica en Salud (PI2010-01035). We thankIzaskun Beloqui, Andres Pavon and Maria Diaz for technicalsupport. We thank the Flow Cytometry Unit at Inbiomed forextensive aid in flow cytometric analysis of fresh tumour cells.We thank our colleagues at Inbiomed for helpful discussion.We thank Dr R Sanchez-Pernaute and Dr L Vellon for criticalreading of the manuscript.
References
Abraham BK, Fritz P, McClellan M, Hauptvogel P, Athelogou M,Brauch H. (2005). Prevalence of CD44+/CD24"/low cells in breastcancer may not be associated with clinical outcome but may favordistant metastasis. Clin Cancer Res 11: 1154–1159.
Akala OO, Park IK, Qian D, Pihalja M, Becker MW, Clarke MF.(2008). Long-term haematopoietic reconstitution by Trp53-/-p16Ink4a-/-p19Arf-/- multipotent progenitors. Nature 453: 228–232.
Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, ClarkeMF. (2003). Prospective identification of tumorigenic breast cancercells. Proc Natl Acad Sci USA 100: 3983–3988.
Ambady S, Malcuit C, Kashpur O, Kole D, Holmes WF, Hedblom Eet al. (2010). Expression of NANOG and NANOGP8 in a variety ofundifferentiated and differentiated human cells. Int J Dev Biol 54:1743–1754.
Ao A, Morrison BJ, Wang H, Lopez JA, Reynolds BA, Lu J. (2011).Response of estrogen receptor-positive breast cancer tumorspheresto antiestrogen treatments. PLoS ONE 6: e18810.
Callagy G, Cattaneo E, Daigo Y, Happerfield L, Bobrow LG, PharoahPD et al. (2003). Molecular classification of breast carcinomas usingtissue microarrays. Diagn Mol Pathol 12: 27–34.
Cantz T, Key G, Bleidissel M, Gentile L, Han DW, Brenne A et al.(2007). Absence of OCT4 expression in somatic tumor cell lines.Stem Cells 26: 692–697.
Chaffer CL, Brueckmann I, Scheel C, Kaestli AJ, Wiggins PA,Rodrigues LO et al. (2011). Normal and neoplastic nonstem cellscan spontaneously convert to a stem-like state. Proc Natl Acad SciUSA 108: 7950–7955.
Chen Y, Shi L, Zhang L, Li R, Liang J, Yu W et al. (2008). Themolecular mechanism governing the oncogenic potential of SOX2 inbreast cancer. J Biol Chem 283: 17969–17978.
Cicalese A, Bonizzi G, Pasi CE, Faretta M, Ronzoni S,Giulini B et al. (2009). The tumor suppressor p53 regulates polarity
of self-renewing divisions in mammary stem cells. Cell 138:1083–1095.
Cirenajwis H, Smiljanic S, Honeth G, Hegardt C, Marton LJ,Oredsson SM. (2010). Reduction of the putative CD44+CD24"breast cancer stem cell population by targeting thepolyamine metabolic pathway with PG11047. Anticancer Drugs21: 897–906.
Comtesse N, Zippel A, Walle S, Monz D, Backes C, Fischer U et al.(2005). Complex humoral immune response against a benign tumor:frequent antibody response against specific antigens as diagnostictargets. Proc Natl Acad Sci USA 102: 9601–9606.
Dontu G, Abdallah WM, Foley JM, Jackson KW, Clarke MF,Kawamura MJ et al. (2003). in vitro propagation and transcrip-tional profiling of human mammary stem/progenitor cells. GenesDev 17: 1253–1270.
Engelmann K, Shen H, Finn OJ. (2008). MCF7 side population cellswith characteristics of cancer stem/progenitor cells express thetumor antigen MUC1. Cancer Res 68: 2419–2426.
Farnie G, Clarke RB, Spence K, Pinnock N, Brennan K, AndersonNG et al. (2007). Novel cell culture technique for primary ductalcarcinoma in situ: role of Notch and epidermal growth factorreceptor signaling pathways. J Natl Cancer Inst 99: 616–627.
Fillmore CM, Kuperwasser C. (2008). Human breast cancer celllines contain stem-like cells that self-renew, give rise to phenotypi-cally diverse progeny and survive chemotherapy. Breast Cancer Res10: R25.
Gangemi RM, Griffero F, Marubbi D, Perera M, Capra MC,Malatesta P et al. (2009). SOX2 silencing in glioblastoma tumor-initiating cells causes stop of proliferation and loss of tumorigeni-city. Stem Cells 27: 40–48.
Ghods AJ, Irvin D, Liu G, Yuan X, Abdulkadir IR, Tunici P et al.(2007). Spheres isolated from 9L gliosarcoma rat cell line possess
Sox2 activation in breast cancer stem cellsO Leis et al
10
Oncogene

chemoresistant and aggressive cancer stem-like cells. Stem Cells 25:1645–1653.
Ginestier C, Hur MH, Charafe-Jauffret E, Monville F, Dutcher J,Brown M et al. (2007). ALDH1 is a marker of normal andmalignant human mammary stem cells and a predictor of poorclinical outcome. Cell Stem Cell 1: 555–567.
Gure AO, Stockert E, Scanlan MJ, Keresztes RS, Jager D, Altorki NKet al. (2000). Serological identification of embryonic neural proteinsas highly immunogenic tumor antigens in small cell lung cancer.Proc Natl Acad Sci USA 97: 4198–4203.
Hong H, Takahashi K, Ichisaka T, Aoi T, Kanagawa O, NakagawaMet al. (2009). Suppression of induced pluripotent stem cellgeneration by the p53-p21 pathway. Nature 460: 1132–1135.
Iliopoulos D, Hirsch HA, Wang G, Struhl K. (2011). Inducibleformation of breast cancer stem cells and their dynamic equilibriumwith non-stem cancer cells via IL6 secretion. Proc Natl Acad SciUSA 108: 1397–1402.
Jaenisch R, Young R. (2008). Stem cells, the molecular circuitry ofpluripotency and nuclear reprogramming. Cell 132: 567–582.
Katoh M. (2010). Network of WNT and other regulatory signalingcascades in pluripotent stem cells and cancer stem cells. Curr PharmBiotechnol 12: 160–170.
Kawamura T, Suzuki J, Wang YV, Menendez S, Morera LB, Raya Aet al. (2009). Linking the p53 tumour suppressor pathway to somaticcell reprogramming. Nature 460: 1140–1144.
Kidder BL, Palmer S, Knott JG. (2009). SWI/SNF-Brg1 regulates self-renewal and occupies core pluripotency-related genes in embryonicstem cells. Stem Cells 27: 317–328.
Kim JB, Greber B, Arauzo-Bravo MJ, Meyer J, Park KI, Zaehres Het al. (2009). Direct reprogramming of human neural stem cells byOCT4. Nature 461: 649–643.
Kondo T, Raff M. (2004). Chromatin remodeling and histonemodification in the conversion of oligodendrocyte precursors toneural stem cells. Genes Dev 18: 2963–2972.
Lengerke C, Fehm T, Kurth R, Neubauer H, Scheble V, Muller F et al.(2011). Expression of the embryonic stem cell marker SOX2 inearly-stage breast carcinoma. BMC Cancer 11: 42.
Li H, Collado M, Villasante A, Strati K, Ortega S, Canamero M et al.(2009). The Ink4/Arf locus is a barrier for iPS cell reprogramming.Nature 460: 1136–1139.
Li X, Lewis MT, Huang J, Gutierrez C, Osborne CK, Wu MF et al.(2008). Intrinsic resistance of tumorigenic breast cancer cells tochemotherapy. J Natl Cancer Inst 100: 672–679.
Li XL, Eishi Y, Bai YQ, Sakai H, Akiyama Y, Tani M et al. (2004).Expression of the SRY-related HMG box protein SOX2 in humangastric carcinoma. Int J Oncol 24: 257–263.
Liu M, Sakamaki T, Casimiro MC, Willmarth NE, Quong AA,Ju X et al. (2010). The canonical NF-\{kappa\}B pathwaygoverns mammary tumorigenesis in transgen. Cancer Res 70:10464–10473.
Livak KJ, Schmittgen TD. (2001). Analysis of relative gene expressiondata using real-time quantitative PCR and the 2(-Delta Delta C(T))method. Methods 25: 402–408.
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan A, Zhou AY et al.(2008). The epithelial-mesenchymal transition generates cells withproperties of stem cells. Cell 133: 704–715.
Marion RM, Strati K, Li H, Murga M, Blanco R, Ortega Set al. (2009). A p53-mediated DNA damage response limitsreprogramming to ensure iPS cell genomic integrity. Nature 460:1149–1153.
Martin AG, San-Antonio B, Fresno M. (2001). Regulation of nuclearfactor kappa B transactivation. Implication of phosphatidylinositol3-kinase and protein kinase C zeta in c-Rel activation by tumornecrosis factor alpha. J Biol Chem 276: 15840–15849.
Meissner A, Wernig M, Jaenisch R. (2007). Direct reprogramming ofgenetically unmodified fibroblasts into pluripotent stem cells. NatBiotechnol 25: 1177–1181.
Miki Y, Swensen J, Shattuck-Eidens D, Futreal PA, Harshman K,Tavtigian S et al. (1994). A strong candidate for the breast andovarian cancer susceptibility gene BRCA1. Science 266: 66–71.
Miyagi S, Nishimoto M, Saito T, Ninomiya M, Sawamoto K, OkanoH et al. (2006). The Sox2 regulatory region 2 functions as a neuralstem cell-specific enhancer in the telencephalon. J Biol Chem 281:13374–13381.
Miyagi S, Saito T, Mizutani K, Masuyama N, Gotoh Y, Iwama Aet al. (2004). The Sox-2 regulatory regions display their activitiesin two distinct types of multipotent stem cells. Mol Cell Biol 24:4207–4220.
Nakagawa M, Koyanagi M, Tanabe K, Takahashi K, Ichisaka T,Aoi T et al. (2008). Generation of induced pluripotent stem cellswithout Myc from mouse and human fibroblasts. Nat Biotechnol 26:101–106.
Okita K, Nakagawa M, Hyenjong H, Ichisaka T, Yamanaka S. (2008).Generation of mouse induced pluripotent stem cells without viralvectors. Science 322: 949–953.
Papapetrou EP, Tomishima MJ, Chambers SM, Mica Y, Reed E,Menon J et al. (2009). Stoichiometric and temporal requirements ofOct4, Sox2, Klf4, and c-Myc expression for efficient humaniPSC induction and differentiation. Proc Natl Acad Sci USA 106:12759–12764.
Patrawala L, Calhoun T, Schneider-Broussard R, Zhou J, Claypool K,Tang DG. (2005). Side population is enriched in tumorigenic, stem-like cancer cells, whereas ABCG2+ and ABCG2" cancer cells aresimilarly tumorigenic. Cancer Res 65: 6207–6219.
Ponti D, Costa A, Zaffaroni N, Pratesi G, Petrangolini G, Coradini Det al. (2005). Isolation and in vitro propagation of tumorigenicbreast cancer cells with stem/progenitor cell properties. Cancer Res65: 5506–5511.
Rodriguez-Pinilla SM, Sarrio D, Moreno-Bueno G, Rodriguez-Gil Y,Martinez MA, Hernandez L et al. (2007). Sox2: a possible driver ofthe basal-like phenotype in sporadic breast cancer. Mod Pathol 20:474–481.
Sanada Y, Yoshida K, Ohara M, Oeda M, Konishi K, Tsutani Y.(2006). Histopathologic evaluation of stepwise progression ofpancreatic carcinoma with immunohistochemical analysis of gastricepithelial transcription factor SOX2: comparison of expressionpatterns between invasive components and cancerous or nonneo-plastic intraductal components. Pancreas 32: 164–170.
Sattler HP, Lensch R, Rohde V, Zimmer E, Meese E, Bonkhoff Het al. (2000). Novel amplification unit at chromosome 3q25-q27 inhuman prostate cancer. Prostate 45: 207–215.
Shafee N, Smith CR, Wei S, Kim Y, Mills GB, Hortobagyi GN et al.(2008). Cancer stem cells contribute to cisplatin resistance inBrca1/p53-mediated mouse mammary tumors. Cancer Res 68:3243–3250.
Sholl LM, Barletta JA, Yeap BY, Chirieac LR, Hornick JL. (2010).Sox2 protein expression is an independent poor prognosticindicator in stage I lung adenocarcinoma. Am J Surg Pathol 34:1193–1198.
Simoes BM, Piva M, Iriondo O, Comaills V, Lopez-Ruiz JA, Zabalza Iet al. (2010). Effects of estrogen on the proportion of stem cells inthe breast. Breast Cancer Res Treat 129: 23–35.
Suo G, Han J, Wang X, Zhang J, Zhao Y, Zhao Y et al. (2005). Oct4pseudogenes are transcribed in cancers. Biochem Biophys ResCommun 337: 1047–1051.
Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda Ket al. (2007). Induction of pluripotent stem cells from adult humanfibroblasts by defined factors. Cell 131: 861–872.
Tanei T, Morimoto K, Shimazu K, Kim SJ, Tanji Y, Taguchi T et al.(2009). Association of breast cancer stem cells identified by aldehydedehydrogenase 1 expression with resistance to sequential paclitaxeland epirubicin-based chemotherapy for breast cancers. Clin CancerRes 15: 4234–4241.
Tomioka M, Nishimoto M, Miyagi S, Katayanagi T, Fukui N, NiwaH et al. (2002). Identification of Sox-2 regulatory region which isunder the control of Oct-3/4-Sox-2 complex. Nucleic Acids Res 30:3202–3213.
Utikal J, Polo JM, Stadtfeld M, Maherali N, Kulalert W, Walsh RMet al. (2009). Immortalization eliminates a roadblock during cellularreprogramming into iPS cells. Nature 460: 1145–1148.
Sox2 activation in breast cancer stem cellsO Leis et al
11
Oncogene

Yamanaka S. (2008). Induction of pluripotent stem cells frommouse fibroblasts by four transcription factors. Cell Prolif41(Suppl 1): 51–56.
Zappone MV, Galli R, Catena R, Meani N, De Biasi S, Mattei E et al.(2000). Sox2 regulatory sequences direct expression of a (beta)-geotransgene to telencephalic neural stem cells and precursors of themouse embryo, revealing regionalization of gene expression in CNSstem cells. Development 127: 2367–2382.
Zeng YA, Nusse R. (2010). Wnt proteins are self-renewal factors formammary stem cells and promote their long-term expansion inculture. Cell Stem Cell 6: 568–577.
Zhang J, Wang X, Li M, Han J, Chen B, Wang B et al. (2006). NANOGP8is a retrogene expressed in cancers. FEBS J 273: 1723–1730.
Zhou J, Zhang H, Gu P, Bai J, Margolick JB, Zhang Y. (2007). NF-kappaB pathway inhibitors preferentially inhibit breast cancer stem-like cells. Breast Cancer Res Treat 111: 419–427.
Supplementary Information accompanies the paper on the Oncogene website (http://www.nature.com/onc)
Sox2 activation in breast cancer stem cellsO Leis et al
12
Oncogene





























