Embed Size (px)
Citation preview

John G. FleagleDepartment of AnatomicalSciences, Health SciencesCenter, State University ofNew York, Stony Brook, NewYork 11794-8081, U.S.A.E-mail:[email protected]
W. Scott McGrawDepartment of Anthropology,The Ohio State University,1680 University Dr.,Mansfield, Ohio 44907,U.S.A. E-mail:[email protected]
Received 3 July 2000Revision received 1 October2001 and accepted10 October 2001
Keywords: Cercocebus,Lophocebus, Papio,Mandrillus, phylogeny,anatomy.
Skeletal and dental morphology of Africanpapionins: unmasking a cryptic clade
One of the more perplexing problems in primate systematics concernsthe phyletic relationships of the large African monkeys—Mandrillus(including drills), Papio, Lophocebus and Cercocebus. For over twentyyears, there has been molecular evidence that mangabeys are anunnatural group and that the terrestrial forms—Cercocebus—are thesister taxon of Mandrillus, while the arboreal forms—Lophocebus—aremore closely allied with Papio. Nevertheless, most systematistshave been reluctant to accept this scheme due to the lack of morpho-logical evidence. In this paper, we undertake a detailed analysis ofthe scapula, humerus, radius, ulna, pelvis, femur and dentition ofpapionin primates. We identify a host of features shared by Cercocebusand Mandrillus to the exclusion of Lophocebus and Papio. The polarityof characters is established by examining an outgroup comprised ofseveral species of Macaca. The features shared by Cercocebus andMandrillus are functionally related to specific feeding and locomotorbehaviors that include aggressive manual foraging, the processing ofhard-object foods and the climbing of vertical trunks. We hypothesizethat the ability to subsist on hard seeds and nuts gleaned from theforest floor is a key adaptation for the Cercocebus–Mandrillus clade.
� 2002 Elsevier Science Ltd.
Journal of Human Evolution (2002) 42, 267–292doi:10.1006/jhev.2001.0526Available online at http://www.idealibrary.com on
IntroductionSince their initial discovery by westernscientists 200 years ago, the larger Africancercopithecine monkeys have been placed intwo seemingly natural groups—mangabeysand baboons (including mandrills andgeladas). Indeed many authors have onlyrecognized two genera—Cercocebus for themangabeys and Papio for the baboons,geladas and mandrills (Thorington &Groves, 1970; Szalay & Delson, 1979).However, most recent authors tend to rec-ognize more taxonomic diversity within thetwo groups. While all mangabeys are long-limbed monkeys with hollow cheeks andlong tails, mangabeys are clearly divided intotwo species groups that show consistent dif-ferences in cranial anatomy (Groves, 1978)and ecology (e.g., Chalmers, 1968; Jones &Sabater Pi, 1968; Quiris, 1975; Waser,
0047–2484/02/030267+26$35.00/0
1977, 1984; Homewood, 1978; Horn,1987; Mitani, 1989; Olupot et al., 1994,1997; Kingdon, 1997). One group, thegaleritus–torquatus–atys–agilis group retainedin the genus Cercocebus, contains predomi-nantly terrestrial monkeys that are reportedto live in large groups with large homeranges (Jones & Sabater Pi, 1968; Quiris,1975; Homewood, 1978; Mitani, 1989).The other species group, the albigena–aterrimus group, contains more slender,strictly arboreal monkeys with smaller homeranges (Chalmers, 1968; Waser, 1977,1984; Horn, 1987; Olupot et al., 1994,1997). These mangabeys are frequentlyplaced in a separate genus, Lophocebus (e.g.,Groves, 1978; Rowe, 1996; Fleagle, 1999).
Likewise, the baboons are generallydivided into three different genera: Papio forthe savannah baboons found throughout
� 2002 Elsevier Science Ltd.

268 . . .
Cercocebus Mandrillus Lophocebus Theropithecus Papio
Figure 1. Phylogeny of African papionin monkeys as evinced by molecular data (Disotell, 1994; Harris &Disotell, 1998; Harris, 2000).
sub-Saharan Africa (Jolly, 1967; Groves,1993); Theropithecus for the geladas of theEthiopian plateau (Jolly, 1972; Jablonski,1993), and Mandrillus for the forest-dwelling mandrill and drill of western andwestern Central Africa (Grubb, 1973).
In 1976, Cronin & Sarich presentedimmunological data showing that Cercocebus(=Lophocebus) albigena was more closelyrelated to baboons and geladas than to C.galeritus and Mandrillus. Dutrilleaux et al.(1982) and Stanyon et al. (1988) demon-strated that Mandrillus and Cercocebus spp.uniquely shared a rearrangement of chromo-some 10 to the exclusion of Lophocebus.More recently, Disotell and colleagueshave demonstrated from both mitochon-drial genes (Disotell, 1994) and nucleargenes (Harris & Disotell, 1998; Harris,2000) that C. galeritus and Mandrillusform one clade, while L. albigena formsa separate clade with baboons (Papio
and Theropithecus), although the detailedrelationships within this later clade areunresolved (Figure 1).
Despite the consistent molecular evidencefor over 20 years supporting a diphyleticorigin of mandrills and baboons from separ-ate groups of mangabeys, morphological evi-dence in support of such a phylogeny has,until recently, been very limited. Thus,while several workers (e.g., Groves, 1978;Nakatsukasa, 1994a,b, 1996) have docu-mented cranial and postcranial differ-ences within mangabeys and others havedocumented differences among mandrills,baboons and geladas (e.g., Jolly, 1970;Szalay & Delson, 1979; Jablonski, 1993;Ciochon, 1994), there has been remarkablylittle morphological evidence put forth link-ing Cercocebus uniquely with Mandrillus (butsee Hill, 1970; Disotell, 1994; Groves,2000) and Lophocebus uniquely with Papioand/or Theropithecus.

269
In the course of a comparative study oflocomotor behavior and skeletal anatomywithin mangabeys, we uncovered numerousosteological similarities that link Cercocebusand Mandrillus, and contrast with theconditions found in Lophocebus and Papio,thus supporting the molecular phylogeny(Fleagle & McGraw, 1998). Moreover, areview of the ecological literature demon-strates that Cercocebus (the drill mangabeysof Kingdon, 1997) and mandrills sharemany characteristics of their feeding ecologyand locomotor behavior that accord with thedental and skeletal features uniting thesegenera. In an earlier paper (Fleagle &McGraw, 1999), we identified features ofthe postcranial anatomy and dentition thatunite Cercocebus and Mandrillus and distin-guish them from Lophocebus and Papio. Inthe present paper, we expand our compara-tive sample in several ways: we include drillsand geladas as well as mangabeys, mandrillsand baboons; we compare females of thetwo clades as well as males; and we includedata on macaques, the outgroup to theAfrican papionins, in order to distinguishprimitive and derived states. In addition, weoffer functional hypotheses for these mor-phological differences based on what isknown about the behavior and ecology ofmangabeys and mandrills. Finally, wepresent an adaptive scenario to explainthe evolution of this clade of large Africancercopithecines in terms of a unique setof ecological adaptations for exploitingresources on the forest floor of Africantropical forests.
Materials and methods
This study is based on examination ofapproximately 150 individual skeletons ofthe genera Cercocebus, Lophocebus, Papio,Mandrillus, Theropithecus and Macaca fromcollections in the American Museum ofNatural History, The Natural History
Museum of London, Powell–CottonMuseum (Birchington, U.K.), Museum ofComparative Zoology (Harvard), RoyalMuseum of Central Africa (Tervuren,Belgium), The Randall L. SusmanCollection at SUNY—Stony Brook, andspecimens collected from the Ivory Coast’sTai Forest.
We have compared the bones and teethof these monkeys using descriptive compari-sons, quantitative, continuous measure-ments, and nonmetric assessments of crestdevelopment. The quantitative measure-ments on individual bones and teeth areillustrated in Figure 2. Measurements weretaken with digital calipers.
These monkeys are very sexually dimor-phic, and comparing mixed samples ofsexually dimorphic species can oftenobscure species-specific differences (seeNakatsukasa, 1994a,b, 1996). Therefore,we report several different quantitativecomparisons of the dental and postcranialskeleton. Initial comparisons are betweenthe larger mixed species samples ofmales from the two clades (i.e., Cercocebusand Mandrillus vs. Lophocebus, Papio, andTheropithecus). Subsequently, we compareall individuals of the two clades by addingthe measurements from the smaller (andmore variable) samples of females of eachclade. Finally, in order to estimate which ofthe character states is likely to be primitivefor papionins as a group, we compare thesame measurements from a small sampleof mixed species and sexes of Macaca tothe mixed species and mixed sex samplesof Cercocebus–Mandrillus clade and theLophocebus, Papio, Theropithecus clade.
Univariate dimensions were convertedinto ratios to permit comparisons ofdifferent sized individuals and species. In allcases, means of the individual speci-mens from each of the two clades werecompared using Student’s t-test for differ-ences between the means of the pooledsamples.

270 . . .
(a) (b) (c)
(d)
(e)
(f)
Figure 2. Measurements of (a) scapula, (b) humerus, (c) ulna, (d) pelvis, (e) tibial shaft and (f) premolars.
Comparative skeletal and dentalanatomy
Osteological differences between speciesof mangabeys and between baboons,mangabeys, and geladas have beendocumented by other studies (e.g., Jolly,1970, 1972; Ciochon, 1994; Nakatsukasa,1994a,b, 1996; see also Gebo & Sargis,1994). In most cases, these studies haveidentified features that distinguish terrestrial
and arboreal taxa. In many of these fea-tures, the arboreal Lophocebus differs fromthe more terrestrial Cercocebus, as well asfrom Mandrillus, Papio and Theropithecus(Nakatsukasa, 1994a,b, 1996; Fleagle &McGraw, 1999). In this study we do notdiscuss those arboreal–terrestrial features;rather our goal is to document features thatseem diagnostic of the Cercocebus–Mandrillusclade and distinguish them from the
271
morphological features of Lophocebus, Papioand Theropithecus. In addition, we attempt toidentify synapomorphies and symplesio-morphies by comparisons with an outgroupcomposed of specimens of several species ofMacaca.
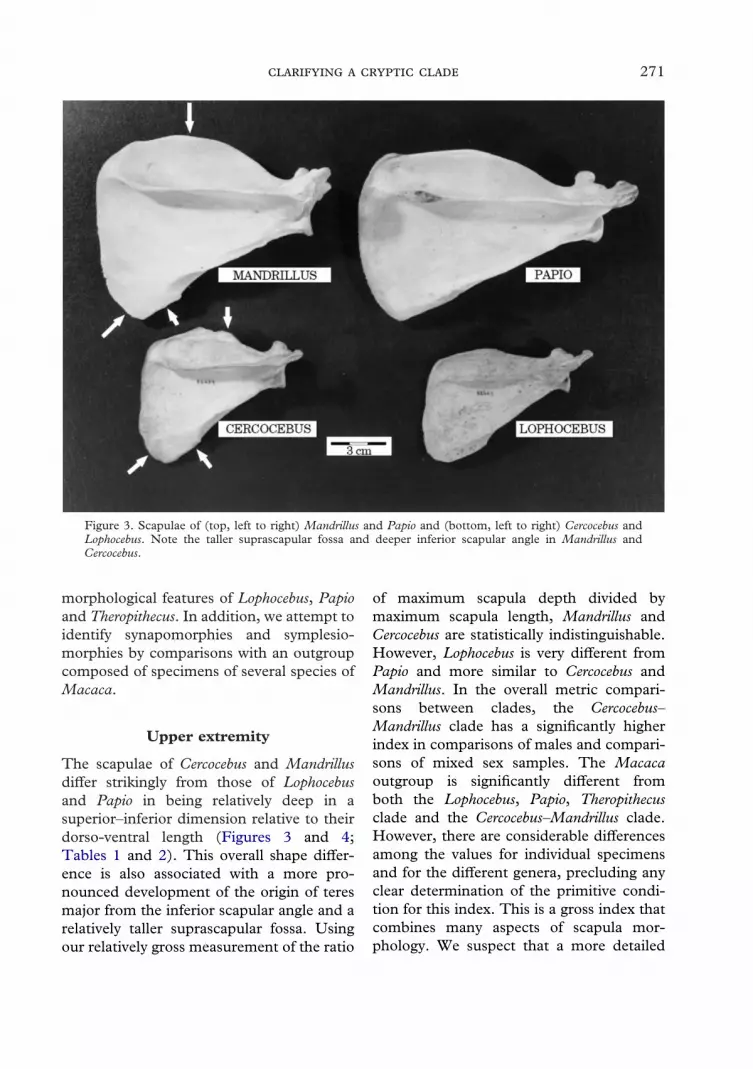
Figure 3. Scapulae of (top, left to right) Mandrillus and Papio and (bottom, left to right) Cercocebus andLophocebus. Note the taller suprascapular fossa and deeper inferior scapular angle in Mandrillus andCercocebus.
Upper extremity
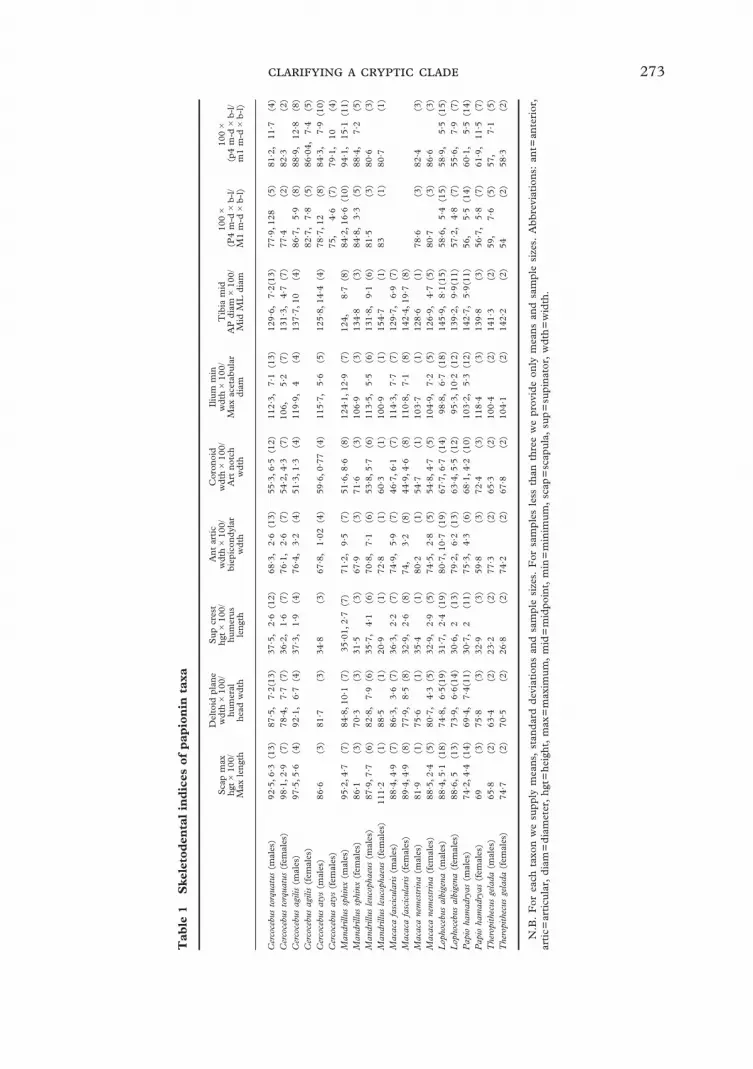
The scapulae of Cercocebus and Mandrillusdiffer strikingly from those of Lophocebusand Papio in being relatively deep in asuperior–inferior dimension relative to theirdorso-ventral length (Figures 3 and 4;Tables 1 and 2). This overall shape differ-ence is also associated with a more pro-nounced development of the origin of teresmajor from the inferior scapular angle and arelatively taller suprascapular fossa. Usingour relatively gross measurement of the ratio
of maximum scapula depth divided bymaximum scapula length, Mandrillus andCercocebus are statistically indistinguishable.However, Lophocebus is very different fromPapio and more similar to Cercocebus andMandrillus. In the overall metric compari-sons between clades, the Cercocebus–Mandrillus clade has a significantly higherindex in comparisons of males and compari-sons of mixed sex samples. The Macacaoutgroup is significantly different fromboth the Lophocebus, Papio, Theropithecusclade and the Cercocebus–Mandrillus clade.However, there are considerable differencesamong the values for individual specimensand for the different genera, precluding anyclear determination of the primitive condi-tion for this index. This is a gross index thatcombines many aspects of scapula mor-phology. We suspect that a more detailed

272 . . .
60 70 80 90 100 110 120
(13)(7)(4)(3)(7)(3)(6)(1)(7)(8)(1)(5)(18)(13)(14)(3)(2)(2)
Cercocebus torquatusC. torquatus
C. agilisC. atys
Mandrillus sphinxM. sphinx
M. leucophaeusM. leucophaeus
Macaca fascicularisM. fascicularisM. nemestrinaM. nemestrina
Lophocebus albigenaL. albigena
Papio hamadryasP. hamadryas
Theropithecus geladaT. gelada
Figure 4. Plot of scapula height index. In this and all subsequent plots, sample sizes for each sex are givenin parentheses. For each taxon and sex the index mean�1 S.D. is depicted. For samples consisting ofthree individuals or less, we have plotted each individual value.
analysis of scapular shape in these taxawould reveal more functional differencesamong these taxa.
Similar differences in scapular shapehave been reported in several other pri-mate groups, notably between Presbytis mel-alophos and Presbytis (Trachypithecus) obscura(Fleagle, 1977), and also between Pitheciapithecia and Chiropotes satanas (Fleagle &Meldrum, 1988). In these primates, theshort and deep scapula is associated withclinging and climbing behavior and thelonger scapula with more quadrupedalbehavior. The deep scapular shape increasesthe lever arm of teres major which is a majorextensor of the humerus. Both Mandrillusand Cercocebus have been reported to regu-larly engage in vertical climbing up treetrunks as they ascend from the forest floor insearch of arboreal foods (as noted by areviewer, this is evident in the Nature tele-vision program ‘‘Mask of the Mandrill’’).Similarly, the much lower value of this indexfor Papio and Theropithecus almost certainlyreflects the less frequent use of vertical
climbing habits in these open-countryterrestrial genera.
The most striking feature of the proximalpart of the humeri of Cercocebus andMandrillus is the exceptional development ofthe deltoid plane which is relatively broadwith prominent, projecting crests medially(deltopectoral crest) and laterally (delto-triceps crest) (Figures 5 and 6; Table 1).Lateral to the deltotriceps crest, bothCercocebus and Mandrillus show a relativelybroad excavation for the upper fibers ofbrachialis that is not seen in Lophocebus andPapio.
An index of the width of the deltoidplane relative to the width of the headof the humerus shows significantly highervalues for the Cercocebus–Mandrillus cladein comparisons of either males or themixed sex samples (Table 2, Figure 7).The Macaca sample is not significantlydifferent from the Cercocebus–Mandrillusclade in this index, suggesting that thebroad deltoid plane is primitive forpapionins.
273
Tab
le1
Ske
leto
den
tal
ind
ices
ofp
apio
nin
taxa
Sca
pm
axhg
t�10
0/M
axle
ngth
Del
toid
plan
ew
dth�
100/
hum
eral
head
wdt
h
Sup
cres
thg
t�10
0/hu
mer
usle
ngth
Ant
arti
cw
dth�
100/
biep
icon
dyla
rw
dth
Cor
onoi
dw
dth�
100/
Art
notc
hw
dth
Iliu
mm
inw
dth�
100/
Max
acet
abul
ardi
am
Tib
iam
idA
Pdi
am�
100/
Mid
ML
diam
100�
(P4
m-d
�b-
l/M
1m
-d�
b-l)
100�
(p4
m-d
�b-
l/m
1m
-d�
b-l)
Cer
coce
bus
torq
uatu
s(m
ales
)92
·5,6
·3(1
3)87
·5,
7·2(
13)
37·5
,2·
6(1
2)68
·3,
2·6
(13)
55·3
,6·5
(12)
112·
3,7·
1(1
3)12
9·6,
7·2(
13)
77·9
,128
(5)
81·2
,11
·7(4
)C
erco
cebu
sto
rqua
tus
(fem
ales
)98
·1,2
·9(7
)78
·4,
7·7
(7)
36·2
,1·
6(7
)76
·1,
2·6
(7)
54·2
,4·3
(7)
106,
5·2
(7)
131·
3,4·
7(7
)77
·4(2
)82
·3(2
)C
erco
cebu
sag
ilis
(mal
es)
97·5
,5·6
(4)
92·1
,6·
7(4
)37
·3,
1·9
(4)
76·4
,3·
2(4
)51
·3,1
·3(4
)11
9·9,
4(4
)13
7·7,
10(4
)86
·7,
5·9
(8)
88·9
,12
·8(8
)C
erco
cebu
sag
ilis
(fem
ales
)82
·7,
7·8
(5)
86·0
4,7·
4(5
)C
erco
cebu
sat
ys(m
ales
)86
·6(3
)81
·7(3
)34
·8(3
)67
·8,
1·02
(4)
59·6
,0·7
7(4
)11
5·7,
5·6
(5)
125·
8,14
·4(4
)78
·7,1
2(8
)84
·3,
7·9
(10)
Cer
coce
bus
atys
(fem
ales
)75
,4·
6(7
)79
·1,
10(4
)M
andr
illus
sphi
nx(m
ales
)95
·2,4
·7(7
)84
·8,1
0·1
(7)
35·0
1,2·
7(7
)71
·2,
9·5
(7)
51·6
,8·6
(8)
124·
1,12
·9(7
)12
4,8·
7(8
)84
·2,1
6·6
(10)
94·1
,15
·1(1
1)M
andr
illus
sphi
nx(f
emal
es)
86·1
(3)
70·3
(3)
31·5
(3)
67·9
(3)
71·6
(3)
106·
9(3
)13
4·8
(3)
84·8
,3·
3(5
)88
·4,
7·2
(5)
Man
drill
usle
ucop
haeu
s(m
ales
)87
·9,7
·7(6
)82
·8,
7·9
(6)
35·7
,4·
1(6
)70
·8,
7·1
(6)
53·8
,5·7
(6)
113·
5,5·
5(6
)13
1·8,
9·1
(6)
81·5
(3)
80·6
(3)
Man
drill
usle
ucop
haeu
s(f
emal
es)
111·
2(1
)88
·5(1
)20
·9(1
)72
·8(1
)60
·3(1
)10
0·9
(1)
154·
7(1
)83
(1)
80·7
(1)
Mac
aca
fasc
icul
aris
(mal
es)
88·4
,4·9
(7)
86·3
,3·
6(7
)36
·3,
2·2
(7)
74·9
,5·
9(7
)46
·7,6
·1(7
)11
4·3,
7·7
(7)
129·
7,6·
9(7
)M
acac
afa
scic
ular
is(f
emal
es)
89·4
,4·9
(8)
77·9
,8·
5(8
)32
·9,
2·6
(8)
74,
3·2
(8)
44·9
,4·6
(8)
110·
8,7·
1(8
)14
2·4,
19·7
(8)
Mac
aca
nem
estr
ina
(mal
es)
81·9
(1)
75·6
(1)
35·4
(1)
80·2
(1)
54·7
(1)
103·
7(1
)12
8·6
(1)
78·6
(3)
82·4
(3)
Mac
aca
nem
estr
ina
(fem
ales
)88
·5,2
·4(5
)80
·7,
4·3
(5)
32·9
,2·
9(5
)74
·5,
2·8
(5)
54·8
,4·7
(5)
104·
9,7·
2(5
)12
6·9,
4·7
(5)
80·7
(3)
86·6
(3)
Lop
hoce
bus
albi
gena
(mal
es)
88·4
,5·1
(18)
74·8
,6·
5(19
)31
·7,
2·4
(19)
80·7
,10·
7(1
9)67
·7,6
·7(1
4)98
·8,
6·7
(18)
145·
9,8·
1(15
)58
·6,
5·4
(15)
58·9
,5·
5(1
5)L
opho
cebu
sal
bige
na(f
emal
es)
88·6
,5(1
3)73
·9,
6·6(
14)
30·6
,2
(13)
79·2
,6·
2(1
3)63
·4,5
·5(1
2)95
·3,1
0·2
(12)
139·
2,9·
9(11
)57
·2,
4·8
(7)
55·6
,7·
9(7
)P
apio
ham
adry
as(m
ales
)74
·2,4
·4(1
4)69
·4,
7·4(
11)
30·7
,2
(11)
75·3
,4·
3(6
)68
·1,4
·2(1
0)10
3·2,
5·3
(12)
142·
7,5·
9(11
)56
,5·
5(1
4)60
·1,
5·5
(14)
Pap
ioha
mad
ryas
(fem
ales
)69
(3)
75·8
(3)
32·9
(3)
59·8
(3)
72·4
(3)
118·
4(3
)13
9·8
(3)
56·7
,5·
8(7
)61
·9,
11·5
(7)
The
ropi
thec
usge
lada
(mal
es)
65·8
(2)
63·4
(2)
23·2
(2)
77·3
(2)
65·3
(2)
100·
4(2
)14
1·3
(2)
59,
7·6
(5)
57,
7·1
(5)
The
ropi
thec
usge
lada
(fem
ales
)74
·7(2
)70
·5(2
)26
·8(2
)74
·2(2
)67
·8(2
)10
4·1
(2)
142·
2(2
)54
(2)
58·3
(2)
N.B
.F
orea
chta
xon
we
supp
lym
eans
,st
anda
rdde
viat
ions
and
sam
ple
size
s.F
orsa
mpl
esle
ssth
anth
ree
we
prov
ide
only
mea
nsan
dsa
mpl
esi
zes.
Abb
revi
atio
ns:
ant=
ante
rior
,ar
tic=
arti
cula
r,di
am=
diam
eter
,hg
t=he
ight
,m
ax=
max
imum
,m
id=
mid
poin
t,m
in=
min
imum
,sc
ap=
scap
ula,
sup
=su
pina
tor,
wdt
h=
wid
th.

274 . . .
Tab
le2
Sig
nifi
can
ceva
lues
for
com
par
ison
sof
pap
ion
inta
xa
Cer
coce
bus–
Man
drill
usvs
.L
opho
cebu
s–P
apio
–The
ropi
thec
us(b
oth
sexe
s)
Cer
coce
bus–
Man
drill
usvs
.L
opho
cebu
s–P
apio
–The
ropi
thec
us(m
ales
only
)M
acac
avs
.C
erco
cebu
s–M
andr
illus
Mac
aca
vs.
Lop
hoce
bus–
Pap
io–T
hero
pith
ecus
Sca
pula
max
.he
ight
�10
0/m
ax.
leng
th<
<0·
001
<<
0·00
10·
009
0·01
Del
toid
plan
ew
idth
�10
0/hu
mer
alhe
adw
idth
<<
0·00
1<
<0·
001
0·38
4<
<0·
001
Sup
.cr
est
heig
ht�
100/
hum
erus
leng
th<
<0·
001
<<
0·00
10·
078
<<
0·00
1A
nt.
arti
cula
rw
idth
�10
0/bi
epic
onyl
arw
idth
<0·
001
<0·
001
0·00
50·
23C
oron
oid
wid
th�
100/
arti
cula
rno
tch
wid
th<
<0·
001
<<
0·00
1<
0·00
1<
<0·
001
Iliu
mm
in.
wid
th�
100/
max
.ac
etab
ular
diam
eter
<<
0·00
1<
<0·
001
0·10
70·
0004
Tib
iam
idA
Pdi
amet
er�
100/
mid
ML
diam
eter
<<
0·00
1<
<0·
001
0·29
50·
002
100�
(p4
m-d
�b-
l/m1
m-d
�b-
l)<
<0·
001
<<
0·00
10·
41<
<0·
001
100�
(P4
m-d
�b-
l/M1
m-d
�b-
l)<
<0·
001
<<
0·00
10·
610·
001
275
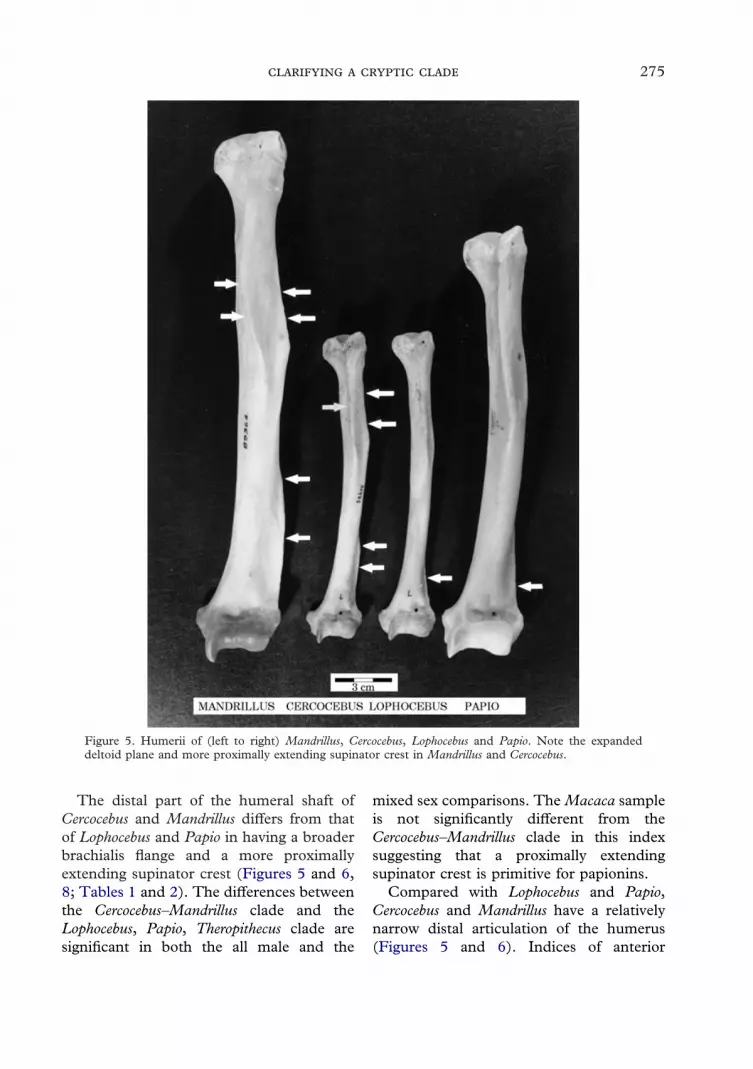
Figure 5. Humerii of (left to right) Mandrillus, Cercocebus, Lophocebus and Papio. Note the expandeddeltoid plane and more proximally extending supinator crest in Mandrillus and Cercocebus.
The distal part of the humeral shaft ofCercocebus and Mandrillus differs from thatof Lophocebus and Papio in having a broaderbrachialis flange and a more proximallyextending supinator crest (Figures 5 and 6,8; Tables 1 and 2). The differences betweenthe Cercocebus–Mandrillus clade and theLophocebus, Papio, Theropithecus clade aresignificant in both the all male and the
mixed sex comparisons. The Macaca sampleis not significantly different from theCercocebus–Mandrillus clade in this indexsuggesting that a proximally extendingsupinator crest is primitive for papionins.
Compared with Lophocebus and Papio,Cercocebus and Mandrillus have a relativelynarrow distal articulation of the humerus(Figures 5 and 6). Indices of anterior

276 . . .
Figure 6. Posterior view of (left to right) Mandrillus, Cercocebus, Lophocebus and Papio humerii. Note thelateral projection of the delto-triceps crest and the narrower olecranon in Mandrillus and Cercocebus.
articular width divided by either thebiepicondyler width or humerus lengthshow significant differences between theCercocebus–Mandrillus clade and theLophocebus, Papio, Theropithecus clade forboth the male only sample and the mixedsex sample (Figure 9; Tables 1 and 2).However, there is considerable variation
among individuals and species within eachclade. The Macaca sample is not signifi-cantly different from the Lophocebus, Papio,Theropithecus clade in these indices, suggest-ing that the Cercocebus–Mandrillus conditionis derived.
On the humerus of Cercocebus andMandrillus there is a relatively more

277
55 65 75 85 95 105
(13)(7)(4)(3)(7)(3)(6)(1)(7)(8)(1)(5)(19)(14)(11)(3)(2)(2)
Cercocebus torquatusC. torquatus
C. agilisC. atys
Mandrillus sphinxM. sphinx
M. leucophaeusM. leucophaeus
Macaca fascicularisM. fascicularisM. nemestrinaM. nemestrina
Lophocebus albigenaL. albigena
Papio hamadryasP. hamadryas
Theropithecus geladaT. gelada
Figure 7. Plot of deltoid plane index.
20 25 30 35 40 45
(12)(7)(4)(3)(7)(3)(6)(7)(8)(1)(5)(19)(13)(11)(3)(2)(2)
Cercocebus torquatusC. torquatus
C. agilisC. atys
Mandrillus sphinxM. sphinx
M. leucophaeusMacaca fascicularis
M. fascicularisM. nemestrinaM. nemestrina
Lophocebus albigenaL. albigena
Papio hamadryasP. hamadryas
Theropithecus geladaT. gelada
Figure 8. Plot of relative height of supinator crest.
prominent medial lip to the trochlea, and arelatively narrower olecranon fossa with asharp, and deep lateral margin (Figures 5and 6; Table 1). In an index of depth of themedial trochlear lip divided by the width ofthe olecranon fossa, the Cercocebus–Mandrillus clade is significantly differentfrom the Lophocebus, Papio, Theropithecus
clade in both the male only comparisons andmixed sex comparisons (Table 2). TheMacaca sample is significantly different fromthe Cercocebus–Mandrillus clade, but notfrom the Lophocebus, Papio, Theropithecusclade, suggesting that the Cercocebus–Mandrillus condition for this index is derivedfor papionins.

278 . . .
50 60 70 80 90 100
(13)(7)(4)(4)(7)(3)
(7)(8)(2)(1)(5)(19)(13)(6)(3)(2)(2)
Cercocebus torquatusC. torquatus
C. agilisC. atys
Mandrillus sphinxM. sphinx
Macaca fascicularisM. fascicularisM. assamensisM. nemestrinaM. nemestrina
Lophocebus albigenaL. albigena
Papio hamadryasP. hamadryas
Theropithecus geladaT. gelada
(6)(1)
M. leucophaeusM. leucophaeus
Figure 9. Plot of relative anterior articular width of humerus.
The distinctive features of the proximalpart of the ulna of Cercocebus and Mandrillusreflect those of the distal humerus. There isa narrower coronoid process with a promi-nent medially facing surface for articulationwith the prominent trochlear lip comparedto the relatively broader coronoid process inLophocebus and Papio (Figure 10; Table 1).In an index of coronoid width divided bywidth of the radial notch, the values for theCercocebus–Mandrillus clade are significantlylower than those for the Lophocebus, Papio,Theropithecus clade in both the males onlycomparisons and the mixed sex comparisons(Table 2; Figure 11). The Macaca sample issignificantly different from either Africanclade, but overlaps extensively with theCercocebus–Mandrillus clade suggesting anarrow coronoid is primitive for papionins.
In Cercocebus and Mandrillus the interos-seus line is prominent and flares ventrally asfar as the ventral surface of the ulnar shaft,whereas in Lophocebus and Papio the interos-seus line is less distinct and confined to thelateral margin of the shaft (Figure 12; Table3). Although not as distinctive as the ulnar
features, in Cercocebus and Mandrillus, theinterosseus (medial) border and the pos-terior border of the radius are more promi-nently developed than in Lophocebus andPapio, and these sharp borders give theradius a triangular rather than a roundedcross-sectional shape (Figure 12; Table 3).
The forelimb bones of Cercocebus andMandrillus show a scapula with featuresfound in other anthropoid species that clingand climb on vertical supports, a humeruswith a large brachialis muscle suggestingadaptations for powerful elbow flexion, anda narrow, stable elbow region. The forearmshows pronounced crest development thatsuggests prominent wrist or digital flexormusculature. This osteological evidenceaccords well with suggestions by Jolly(1970) that mandrills seem to have muchlarger forelimb flexors than other papionins.Jolly (1967:437) further noted that man-drills and drills ‘‘are usually said to be forest-floor animals, but both show anatomicalsigns of considerable greater adaptation totree climbing than in typical baboons(Papio).’’

279
In the absence of more detailed infor-mation about the musculature of these mon-keys, we can nevertheless make somehypotheses about the adaptive significanceof the unusual features of the forelimb ofCercocebus and Mandrillus, based on theirnaturalistic behavior that would seem torequire powerful forelimb flexion. Both areterrestrial foragers that spend much of theirdaily activity rummaging through leaf litteron the forest floor in search of fallen fruits,nuts and seeds, as well as animal material.They are described as aggressive manualforagers that rip apart rotten logs, open largefruits, and harvest terrestrial herbaceousvegetation (Hoshino, 1985; Harrison, 1988;Schaaf et al., 1990; Bergmueller, 1998).While this manual foraging may account forthe digital, carpal, and elbow flexion theyseem insufficient to account for the scapula
features. Both of these monkeys also climbvertical tree trunks in search of food. Itseems likely that this activity would requirepowerful humeral retraction as well aselbow, carpal, and digital flexion.
Figure 10. Proximal ulnae of (left to right) Mandrillus, Cercocebus, Lophocebus and Papio. Note the narowercoronoid in Mandrillus and Cercocebus.
Lower extremity
The pelvis of Cercocebus and Mandrillus dif-fers from that of Lophocebus and Papio inhaving an ilium that is relatively muchbroader at its base (Figures 13 and 14;Tables 1 and 2). The mean values of basalilium breadth relative to acetabulumbreadth are significantly higher for theCercocebus–Mandrillus clade than for theLophocebus, Papio, Theropithecus clade forboth the male only comparisons and themixed sex comparisons. However, thereis considerable spread in the values for

280 . . .
individuals and genera. The values forMacaca are not significantly different fromthose of the Cercocebus–Mandrillus clade,suggesting that a relatively broad basal iliumis primitive for papionins.
The femur of Cercocebus and Mandrillus isgenerally characterized by a more prominentgluteal tuberosity than in Lophocebus orPapio (Table 3). Distally, the femur ofCercocebus and Mandrillus has a patellargroove in which the medial lip is prominentand more similar in form and size to thelateral tip, whereas in Lophocebus and Papiothe lateral lip is more prominent (Figure 15;Table 2). Among other primates, climbingspecies such as Pan troglodytes are character-ized by a patellar groove with equal medialand lateral margins or prominent medialmargins (Ward et al., 1995).
The tibial shaft of Cercocebus andMandrillus is relatively rounder than that ofLophocebus and Papio in which the tibial shaftis more elongate antero-posteriorly (Tables 1and 2; Figure 16). The mean values of rela-tive tibial compression are significantly lowerfor the Cercocebus–Mandrillus clade than for
the Lophocebus, Papio, Theropithecus clade incomparisons for both males only and mixedsex samples. The values for Macaca are notsignificantly different from those of theCercocebus–Mandrillus clade, suggesting thata more rounded tibia shaft is primitive forpapionins.
The features of the hindlimb that distin-guish Cercocebus and Mandrillus are lessextensive than those of the forelimb and lessamenable to interpretation by comparisonwith other primates. Of the behaviors thatseem characteristic of the monkeys, it seemsmost likely that vertical climbing up treetrunks could select for a robust ilium, arelatively more prominent medial patellarmargin and a more rounded tibial shaft.
30 40 50 60 70 9080
(12)(7)(4)(4)(8)(3)(6)
(7)(8)(1)(5)(14)(12)(10)(3)(2)(2)
Cercocebus torquatusC. torquatus
C. agilisC. atys
Mandrillus sphinxM. sphinx
M. leucophaeus
Macaca fascicularisM. fascicularisM. nemestrinaM. nemestrina
Lophocebus albigenaL. albigena
Papio hamadryasP. hamadryas
Theropithecus geladaT. gelada
(1)M. leucophaeus
Figure 11. Plot of index of coronoid width divided by radial notch width.
Premolar morphology
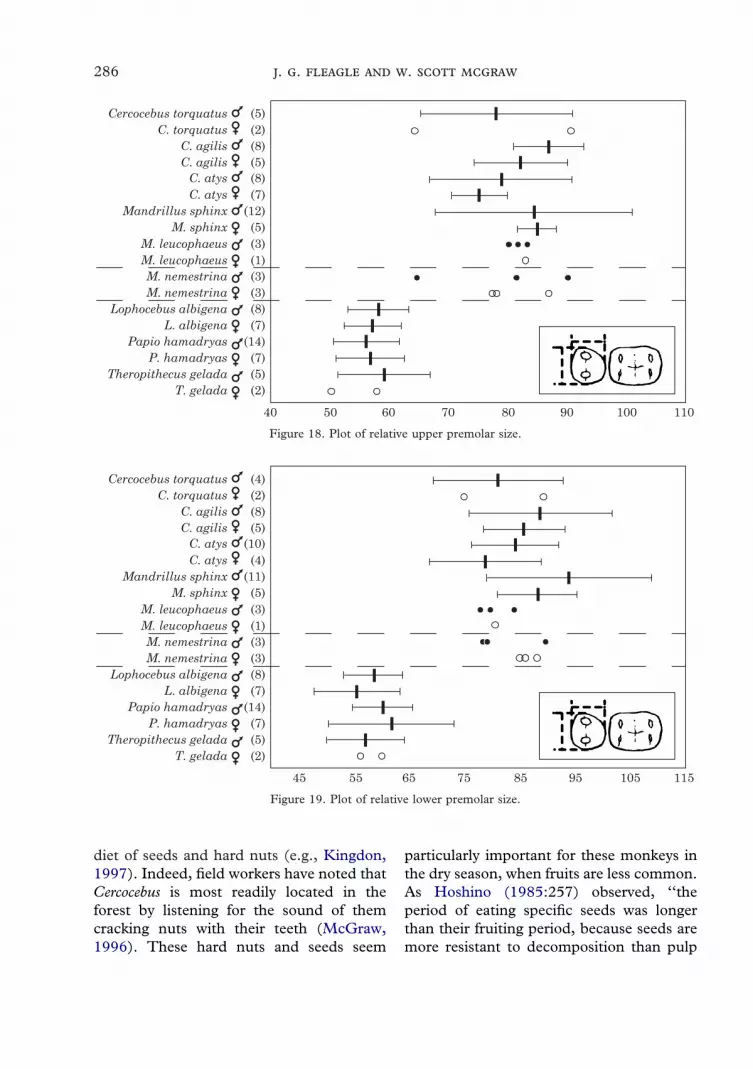
Compared with virtually all other Old Worldmonkeys, Cercocebus and Mandrillus areunusual in having relatively large upper andlower posterior premolars that approach thefirst molar in size (Figures 17–19; Table 1).The ratio of posterior premolar area to first

281
Figure 12. Top: ulnae of (top to bottom) Mandrillus, Cercocebus, Lophocebus and Papio. Bottom: radii of(top to bottom) Mandrillus, Cercocebus, Lophocebus and Papio. Note the more prominent interosseouscrests in the Mandrillus and Cercocebus ulnae and radii.

282 . . .
Table 3 Distribution of nonmetric characters among papionins
Cercocebus Mandrillus Papio Lophocebus
Interosseus crest (ulna) Strong Strong Weak WeakRadial shaft shape Triangular Triangular Rounded RoundedGluteal tuberosity Prominent Prominent Weak WeakPatellar groove: medial lip Weak/round Weak/round Moderate ModeratePatellar groove: lateral lip Strong/sharp Moderate Weak Moderate
Figure 13. Bony pelvis of (left to right) Mandrillus, Cercocebus, Lophocebus and Papio. The ilium ofMandrillus and Cercocebus is much broader at its base (arrows) than those of Lophocebus and Papio.
molar area (both uppers and lowers) issignificantly higher for the Cercocebus–Mandrillus clade than for the Lophocebus,Papio, Theropithecus clade in comparisonsof either male only or mixed sexsamples (Table 2). Our relatively smallsample of Macaca (mostly M. nemestrina) isnot significantly different from theCercocebus–Mandrillus clade in this index,
suggesting that large premolars are primitivefor papionins.
Both Cercocebus and Mandrillus rely exten-sively on hard nuts and seeds that theycollect on the forest floor. Hoshino (1985;see also Rogers et al., 1996) found thatthese were the main food items of mandrillsduring the period of fruit scarcity, andCercocebus in West Africa rely extensively on

283
hard nuts that other primates cannot open(Bergmueller, 1998; Range, 1998; Rutte,1998). It seems most likely that the largepremolars in these species are adaptationsfor cracking open hard nuts and seeds. Inaddition, both Cercocebus and Mandrillushave been reported to eat the pith andbark of various grasses and they may alsouse their premolars for stripping thesefoods (Harrison, 1988). Lophocebus are alsoreported to eat hard nuts such as palmnuts, but they clearly lack the enlargedpremolars of Cercocebus. Unfortunately,there are too few behavioral observations ofthe details of food processing in thesespecies.
80 90 100 110 120 140130
(13)(7)(4)(5)
(7)(3)(6)
(7)(8)(1)(5)(18)(12)(12)(3)(2)(2)
Cercocebus torquatusC. torquatus
C. agilisC. atys
Mandrillus sphinxM. sphinx
M. leucophaeus
Macaca fascicularisM. fascicularisM. nemestrinaM. nemestrina
Lophocebus albigenaL. albigena
Papio hamadryasP. hamadryas
Theropithecus geladaT. gelada
(1)C. atys
(1)M. leucophaeus
Figure 14. Plot of relative basal ilium width.
The adaptive niche of Cercocebus andMandrillus
In the sections above we have documentedmorphological features that distinguishCercocebus and Mandrillus from Lophocebusand Papio and support the molecular studiesthat link these genera as sister taxa. Theformer two genera are among the most
poorly known of all African primatesbecause they move rapidly on the groundthrough dense vegetation and have largeranges. Nevertheless, on the basis of studiesover many years, it is clear that thesemonkeys share many unique behavioral andecological features that unite them and dis-tinguish them from Lophocebus, Papio, andTheropithecus.
Cercocebus has a broad distribution overmuch of central Africa, while Mandrillus isrestricted to areas of Western and westerncentral Africa (Gartlan, 1970; Grubb, 1973;Kingdon, 1997). Both Cercocebus andMandrillus are large, predominantly terres-trial forest-dwelling cercopithecines thatspend much of their daily activity foragingthrough leaf litter on the forest floor. Indescribing the foraging behavior of a hordeof mandrills in Gabon, Rogers et al.(1996:304–305) noted, ‘‘When followingmandrill trails through the forest, it wasobvious that the leaf litter had been exten-sively disturbed, termite nests broken openand rotten wood explored.’’ Likewise, indescribing Cercocebus agilis in Gabon, Quiris

284 . . .
Figure 15. Femora of (left to right) Mandrillus, Cercocebus, Lophocebus and Papio. Note the more equalmedial and lateral lips of the patellar groove in Mandrillus and Cercocebus.
(1975:366) observed, ‘‘Pour terminer,notons que si C. galeritus (agilis) a eteobserve frequemment au sol, fouillant lavase ou la litiere, il n’a jamais ete possible decouvrir ce qu’il cherchait . . .’’. Similarly,Cercocebus atys in the Tai forest, Ivory Coastspends most of its foraging time sortingthrough leaf litter and fallen trees on
the forest floor (McGraw, 1996; personalobservation).
Although Cercocebus and Mandrillus areprimarily terrestrial foragers, they alsoclimb trees to forage on ripe fruits. Whenthey ascend trees Cercocebus often do so byvertical climbing up large trunks (McGraw,1996; see also Quiris, 1975). Similar
285
100 110 120 130 140 170160150
(13)(7)(4)(4)(8)(3)(6)
(7)(8)(1)(5)(15)(11)(11)(3)(2)(2)
Cercocebus torquatusC. torquatus
C. agilisC. atys
Mandrillus sphinxM. sphinx
M. leucophaeus
Macaca fascicularisM. fascicularisM. nemestrinaM. nemestrina
Lophocebus albigenaL. albigena
Papio hamadryasP. hamadryas
Theropithecus geladaT. gelada
(1)M. leucophaeus
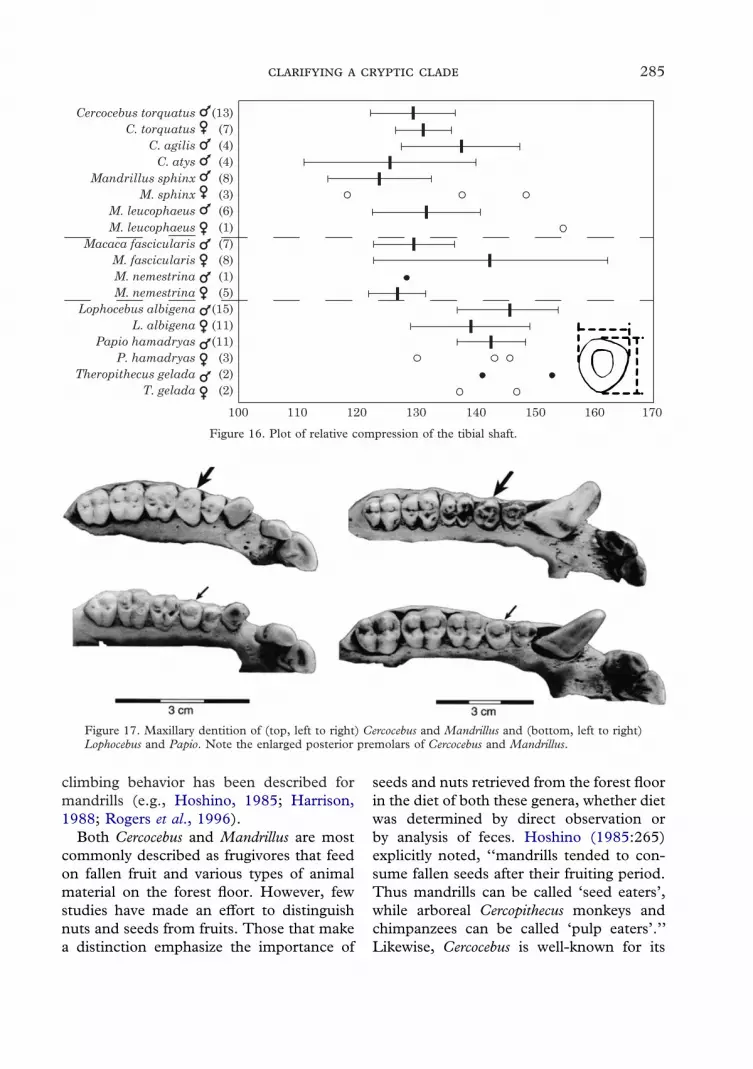
Figure 16. Plot of relative compression of the tibial shaft.
Figure 17. Maxillary dentition of (top, left to right) Cercocebus and Mandrillus and (bottom, left to right)Lophocebus and Papio. Note the enlarged posterior premolars of Cercocebus and Mandrillus.
climbing behavior has been described formandrills (e.g., Hoshino, 1985; Harrison,1988; Rogers et al., 1996).
Both Cercocebus and Mandrillus are mostcommonly described as frugivores that feedon fallen fruit and various types of animalmaterial on the forest floor. However, fewstudies have made an effort to distinguishnuts and seeds from fruits. Those that makea distinction emphasize the importance of
seeds and nuts retrieved from the forest floorin the diet of both these genera, whether dietwas determined by direct observation orby analysis of feces. Hoshino (1985:265)explicitly noted, ‘‘mandrills tended to con-sume fallen seeds after their fruiting period.Thus mandrills can be called ‘seed eaters’,while arboreal Cercopithecus monkeys andchimpanzees can be called ‘pulp eaters’.’’Likewise, Cercocebus is well-known for its
286 . . .
40 50 60 70 80 11090 100
(5)(2)(8)(5)
(7)(12)(5)
(1)(3)(3)(8)(7)(14)(7)(5)(2)
Cercocebus torquatusC. torquatus
C. agilisC. agilis
C. atysMandrillus sphinx
M. sphinx
M. leucophaeusM. nemestrinaM. nemestrina
Lophocebus albigenaL. albigena
Papio hamadryasP. hamadryas
Theropithecus geladaT. gelada
(8)C. atys
(3)M. leucophaeus
Figure 18. Plot of relative upper premolar size.
45 55 65 75 85 11595 105
(4)(2)(8)(5)
(4)(11)(5)
(1)(3)(3)(8)(7)(14)(7)(5)(2)
Cercocebus torquatusC. torquatus
C. agilisC. agilis
C. atysMandrillus sphinx
M. sphinx
M. leucophaeusM. nemestrinaM. nemestrina
Lophocebus albigenaL. albigena
Papio hamadryasP. hamadryas
Theropithecus geladaT. gelada
(10)C. atys
(3)M. leucophaeus
Figure 19. Plot of relative lower premolar size.
diet of seeds and hard nuts (e.g., Kingdon,1997). Indeed, field workers have noted thatCercocebus is most readily located in theforest by listening for the sound of themcracking nuts with their teeth (McGraw,1996). These hard nuts and seeds seem
particularly important for these monkeys inthe dry season, when fruits are less common.As Hoshino (1985:257) observed, ‘‘theperiod of eating specific seeds was longerthan their fruiting period, because seeds aremore resistant to decomposition than pulp

287
and, thus, edible for a longer period. Inother words, mandrills could feed on fruit inthe form of fallen seeds, even when theavailability of fresh fruit (pulp) is poor.’’Thus, it is the ability to eat hard seeds andfruits that seems to permit these large mon-keys to survive in times of low abundance ofripe fruits. In addition, both Cercocebus andMandrillus have been reported to eat terres-trial herbaceous vegetation and variousgrasses, especially during periods of lowfruit abundance. Indeed, there is consider-able dietary overlap between Cercocebus andMandrillus in Cameroon (Caldecott et al.,1996).
Although fallen seeds and nuts are a criti-cal and relatively long lasting resource thatCercocebus and Mandrillus can exploit betterthan other species because of their dentaland locomotor abilities, they neverthe-less seem to be sparsely distributed. Thus,Cercocebus and Mandrillus are unusual in thatboth regularly form supertroops or hordes oflarge numbers of individuals from severaltroops that join together and range verywidely during periods of low fruit availability(Gartlan, 1970; Hoshino, 1985). Furtherstudies of the ecology of these species willundoubtedly reveal additional behavioralsimilarities (Shah, 1996).
Overall, there is abundant evidence thatCercocebus and Mandrillus are a limited andspecialized clade of large cercopithecinescharacterized by shared, unique adaptationsfor terrestrial foraging on the forest floor forfallen fruits, animal material and especiallyhard seeds and nuts. It is this latter specialtycombined with their flexible social organiz-ation that is critical to their survival duringperiods of low fruit availability.
The phylogeny of theCercocebus–Mandrillus adaptation
For each of the morphological featuresthat distinguishes the Cercocebus–Mandrillus
clade from the Lophocebus, Papio, Thero-pithecus clade, we evaluated the conditionin a mixed species, mixed sex sample ofmacaques (M. fascicularis and M. nemest-rina) in order to determine which mor-phologies are likely to be primitive andwhich are derived among papionins. Itturns out that for most of the skeletal fea-tures (broad deltoid plane, high supinatorcrest, broad ilium base, and less medio-laterally compressed tibial shaft), theMacaca mean is not significantly differentfrom that of the Cercocebus–Mandrillus clade.For another feature (coronoid width dividedby radial notch width) the means are statisti-cally different, but there is considerableoverlap and the macaques are clearly moresimilar to the Cercocebus–Mandrillus sample.Likewise, the sample of Macaca nemestrina ismost similar to the Cercocebus–Mandrillussample in having relatively large pre-molars. In only one feature (relative articularwidth of the humerus) is the Lophocebus,Papio, Theropithecus clade more similar tothe macaque morphology. In scapularshape, each of the three groups is distinctive.This suggests that, overall, the morphologi-cal features that distinguish the Cercocebus–Mandrillus clade are mostly primitive forpapionins, and it is the Lophocebus, Papio,Theropithecus clade that is derived.
This simple phylogenetic analysis accordswell with observations of Caldecott et al.(1996), who have noted extensive simi-larities in the ranging behavior and foragingadaptations of pigtail macaques (M. nemest-rina) with those of mandrills and drills, andwith Fooden’s (1980) biogeographic argu-ments that the pigtail macaques are part ofthe initial macaque radiation into Asia. Thiswould suggest that the wide-ranging, forestfloor gleaning adaptation of Cercocebus,mandrills, and pigtail macaques is the basaladaptation of papionins, and that the morearboreal (Lophocebus) and open-country taxa(Papio and Theropithecus) show derivedadaptations.

288 . . .
Discussion
In an earlier paper we described morpho-logical features of the dentition and skeletonof males that characterize Cercocebus andMandrillus as a clade and distinguish themfrom the other clade of African papioninscomposed of Lophocebus, Papio and Thero-pithecus (Fleagle & McGraw, 1999). In thispaper we have provided more extensiveillustrations and broader analyses of thesemorphological features by including femalesand additional species in both clades. Inaddition, we have compared the Africanpapionins with a small sample of macaquesin order to determine whether theCercocebus–Mandrillus features were primi-tive or derived for papionins. There arenumerous aspects of our analysis andresults that merit further discussion. Theseinclude the analysis of female skeletons, thediverse morphology within the Lophocebus,Papio, Theropithecus clade, and the valueof primitive characters in identifyingadaptations.
In our morphological analyses describedabove, the results from comparisons of thelarger, mixed sex samples always accordedwith the results of the all male comparisons(see also Fleagle & McGraw, 1999), andwith the phylogeny of African papioninsbased on biomolecular and genetic studies(Cronin & Sarich, 1976; Hewett-Emett &Cook, 1978; Disotell, 1994, 1996; Harris &Disotell, 1998). However, it is evident fromthe plots of the samples by sex and speciesthat the significant differences were largelydriven by the male sample. The clade-specific, and even species-specific patternswere far less obvious among the femalespecimens than among the males. Similarresults have been reported by others, includ-ing Nakatsukasa (1994a,b, 1996). We can-not explain why females do not show asstrong a pattern of skeletal differences asmales; however, there are several possiblereasons.
One possibility is that the differences arethe result of methodological problems ineither sampling or measuring. For the taxawe measured, complete adult female skel-etons are much less common than adultmale skeletons in the museum collections wevisited. As a result, female samples are morelikely to be a random sample from a varietyof populations rather than a large samplefrom one population. Thus, the apparentdiversity in some of the female samples andthe differences from the males may reflectgeographic diversity. In addition, a singlemisidentified specimen would have a muchgreater effect on the characteristics of asmall sample.
It is also quite possible that the measure-ments of female specimens are less accuratethan those of males. Compared with maleskeletons, female skeletons generally haveless distinctive muscle markings. As a resultmeasurements that depend on identifying aridge caused by a muscle attachment maywell be more difficult to identify in femaleskeletons. However, there is no evidencefrom our results that this is the case. Themeasurements based on muscle attachments(e.g., supinator crest height) are not obvi-ously more variable in the female skeletons,except in Theropithecus. More generally,however, it is well established that the meta-bolic and functional demands on the skel-eton are often different for males andfemales of dimorphic species so thatmany biomechanical features show differentscaling relationships.
Even though we have been able to dem-onstrate significant differences between thetwo clades of African papionins in severalskeletal and dental features, there is alsoconsiderable variation within each of theclades. This is particularly evident for theLophocebus, Papio, Theropithecus clade,which includes taxa with very different pat-terns of substrate use. Lophocebus is a totallyarboreal rainforest monkey; Theropithecus isan almost totally terrestrial monkey from the

289
high plateau of Ethiopia; and the wide-spread Papio is probably best characterizedas predominantly terrestrial, but individualsof many populations regularly forage intrees. In view of the considerable differencesin the ecology and locomotion of extantmembers of this clade, reconstruction of theevolutionary history of adaptations in thisclade is a debated and, in our view, largelyunresolved issue with many alternativescenarios and difficult questions that need tobe addressed in the future.
For example, if the predominantly terres-trial locomotor habits of M. nemestrina,Cercocebus, and Mandrillus are primitive forAfrican papionins, under what conditionsdid the characteristic postcranial features ofLophocebus and Papio evolve? Is Lophocebussecondarily arboreal? This is a scenariofavored by Nakatsukasa (1996) in his studyof skeletal differences among mangabeys.How does Theropithecus fit into the picture?Delson & Dean (1993) have argued fromcranial evidence for similarities betweenTheropithecus and Mandrillus, implying thatthese taxa share features that are primitivefor African papionins. As with our compari-sons of African papionins and macaques, itseems almost certain that any scenario willinvolve some type of mosaic of primitive andderived features in all lineages. Moreover, asthe study by Delson & Dean (1993) empha-sizes, attempts to answer these questions willrequire a broader consideration of the fossilrecord of this group, including Parapapio,fossil baboons, and fossil Theropithecus aswell as more detailed functional studies ofthe biomechanical significance of themorphological features.
In this study we have attempted to ident-ify the functional significance of the featuresshared by Cercocebus and Mandrillus withoutdistinguishing between those features thatare primitive retentions from a commonpapionin ancestry and those that are derivedfor that clade. Our comparisons with a smallsample of Macaca, the outgroup to African
papionins, suggest that many of the featuresshared by Cercocebus and Mandrillus areprobably primitive for all papionins. Theseresults need to be corroborated byadditional outgroup comparisons with alarger sample of macaques, with fossilmacaques such as Procynocephalus, and witha broader sample of other extant and fossilcercopithecines.
While an appreciation of the phylogeny ofindividual morphological and behavioralfeatures is valuable for enabling us to recon-struct the evolutionary history of papionins,the phylogenetic status of individual mor-phological features as either primitive orderived is not critical for our ability tointerpret their functional significance ortheir status as adaptations. There is ampleevidence that both primitive and derivedfeatures have mechanical and physiologicalfunctions that are amenable to interpret-ation through comparative methods (e.g.,Anthony & Kay, 1993; Ross et al., 2001).Thus, as discussed above, it seems mostlikely that the features shared by Cercocebus,Mandrillus and Macaca nemestrina are part ofa longstanding, primitive papionin foragingadaptation related to forest floor locomotionand gleaning.
Summary
Although biomolecular studies of primatesystematics over the past 20 years have con-sistently demonstrated that mandrills are thesister group of Cercocebus mangabeys ratherthan being closely related to baboons andgeladas, there has been little morphologicalor behavior evidence to support thisphylogeny.
Detailed comparisons of the skeletalanatomy of Cercocebus, Mandrillus, Lopho-cebus and Papio reveal osteological featuresof the scapula, humerus, ulna, radius,pelvis, femur and tibia that unite Cercocebusand Mandrillus, and distinguish themfrom Lophocebus and Papio. In addition,

290 . . .
Cercocebus and Mandrillus have greatlyenlarged posterior premolars. The enlargedpremolars are an adaptation for crackingopen hard nuts that both of these monkeysretrieve from the forest floor litter. Theforelimb features are best interpreted asadaptations for extensive and forcefulmanual foraging in forest floor litter as wellas vertical climbing of tree trunks. The hind-limb features are hypothesized as adap-tations for vertical climbing. Comparisonswith an outgroup comprised of Macacaspecies reveals that many features shared byCercocebus and Mandrillus including a broaddeltoid plane, proximally extending supina-tor crest, narrow coronoid, broad basalilium, less mediolaterally compressed tibialshaft and expanded premolars, are primitivefor papionins.
Identification of these shared morphologi-cal features and a review of the litera-ture show that mandrills and Cercocebusmangabeys are similar in many aspects oftheir behavior and ecology. Both are pri-marily wide-ranging terrestrial gleaners onthe forest floor where they manually searchthrough leaf litter and rotten logs in searchof arthropods, fallen fruit, nuts, and seeds.They also ascend the trunk of trees in searchof arboreal food items.
Acknowledgements
We thank the following institutions andindividuals for allowing access to specimensin their care: American Museum of NaturalHistory (Ross MacPhee), The NaturalHistory Museum of London (PaulaJenkins), Field Museum of Natural History(Bill Stanley), UC-Berkeley Laboratoryof Human Evolution (Clark Howell),Museum of Comparative Zoology, HarvardUniversity (Maria Rutzmoser), Powell-Cotton Museum (John Harrison), RoyalMuseum of Central Africa, Tervuren(Wim Van Neer), SUNY-Stony Brook(Randall Susman). We thank Ronald Noe,
the assistants and students of the Tai ForestMonkey project for enabling a betterunderstanding of mangabey biology. BillJungers provided valuable statistical adviceand Luci Betti-Nash skillfully produced thefigures. We thank the various ministries forpermission to carry out fieldwork in theIvory Coast; the Centre Suisse de RechercheScientific and its director, Dr OliverGirardin, provided logistical assistance inAfrica. This work was generously sup-ported by a grant from the LSB LeakeyFoundation.
References
Anthony, M. R. L. & Kay, R. F. (1993). Tooth formand diet in ateline and alouattine primates: Reflectionson the comparative method. Am. J. Science 293A,356–382.
Bergmueller, R. (1998). Nahrungsoekologie undKonkurrenz um Nahrungressourcen bei derRaunchgrauen Mangabe (Cercocebus atys). Diploma,University of Regensburg, Germany.
Caldecott, J. O., Feistner, A. T. C. & Gadsby, E. L.(1996). A comparison of ecological strategies ofpig-tailed macaques, mandrills and drills. In (J. E. Fa& D. G. Lindburg, Eds) Evolution and Ecologyof Macaque Societies, pp. 73–94. Cambridge:Cambridge University Press.
Chalmers, N. R. (1968). Group composition, ecologyand daily activity of free living mangabeys in Uganda.Folia primatol. 8, 247–262.
Ciochon, R. L. (1994). Evolution of the CercopithecoidForelimb: Phylogenetic and Functional Implicationsfrom Morphometric Analyses. Berkeley: University ofCalifornia Press.
Cronin, J. E. & Sarich, V. M. (1976). Molecularevidence for dual origin of mangabeys among OldWorld monkeys. Nature 260, 700–702.
Delson, E. & Dean, D. (1993). Are Papio baringensis R.Leakey, 1969, and P. quadratirostris Iwamoto, 1982,species of Papio or Theropithecus? In (N. Jablonski,Ed.) Theropithecus: The Rise and Fall of a PrimateGenus, pp. 125–156. Cambridge: CambridgeUniversity Press.
Disotell, T. R. (1994). Generic-level relationships ofthe Papionini (Cercopithecoidea). Am. J. phys.Anthrop. 94, 47–57.
Disotell, T. R. (1996). The phylogeny of Old Worldmonkeys. Evol. Anthrop. 5, 18–24.
Dutrillaux, B., Courturier, J., Muleris, M., Lombard,M. & Chauvier, G. (1982). Chromosomal phylogenyof forty-two species or subspecies of cercopithecoids(Primates Catarrhini). Ann. Genet. 25, 96–109.
Fleagle, J. G. (1977). Locomotor behavior and skeletalanatomy of sympatric Malaysian leaf monkeys

291
(Presbytis obscura and Presbytis melalophos). Yearb.phys. Anthrop. 20, 440–453.
Fleagle, J. G. (1999). Primate Adaptation and Evolution.2nd edn. New York: Academic Press.
Fleagle, J. G. & Meldrum, D. J. (1988). Locomotorbehavior and skeletal morphology of two sympatricPitheciine monkeys, Pithecia pithecia and Chiropotessatanas. Am. J. Primatol. 16, 227–249.
Fleagle, J. G. & McGraw, W. S. (1998). Skeletalanatomy of African Papionins: function, phylogenyor both? Am. J. phys. Anthrop. 105, 82–83.
Fleagle, J. G. & McGraw, W. S. (1999). Skeletal anddental morphology supports diphyletic origin ofbaboons and mandrills. Proc. natl. Acad. Sci. 96,1157–1161.
Fooden, J. (1980). Classification and distribution ofliving macaques (Macaca Lacepede, 1799). In (D. G.Lindberg, Ed.) The Macaques: Studies in Ecology,Behavior and Evolution, pp. 1–9. New York: VanNostrand-Reinhold.
Gartlan, J. S. (1970). Preliminary notes on the ecologyand behavior of the drill Mandrillus leucophaeusRitgen, 1824. In (J. R. Napier & P. H. Napier, Eds)Old World Monkeys: Evolution, Systematics andBehavior, pp. 445–480. New York: Academic Press.
Gebo, D. L. & Sargis, E. J. (1994). Terrestrial adap-tations in the postcranial skeletons of guenons. Am.J. phys. Anthrop. 93, 341–371.
Groves, C. P. (1978). Phylogenetic and popu-lation systematics of the mangabeys (Primates:Cercopithecoidea). Primates 19, 1–34.
Groves, C. P. (1993). Order Primates. In (D. E. Wilson& D. M. Reader, Eds) Mammalian Species of theWorld: A Taxonomic and Geographic Reference,pp. 243–277. Washington, DC: SmithsonianInstitute Press.
Groves, C. P. (2000). The phylogeny of theCercopithecoidea. In (P. F. Whitehead & C. J. Jolly,Eds) Old World Monkeys, pp. 77–98. Cambridge:Cambridge University Press.
Grubb, P. (1973). Distribution, divergence andspeciation of the drill and mandrill. Folia primatol. 20,161–177.
Harris, E. E. (2000). Molecular systematics of the OldWorld monkey tribe Papionini: analysis of the totalavailable genetic sequences. J. hum. Evol. 38, 235–256.
Harris, E. E. & Disotell, T. R. (1998). Nuclear genetrees and the phylogenetic relationship of themangabeys (Primates: Papionini). Mol. Biol. Evol.15, 892–900.
Harrison, M. J. (1988). The mandrill in Gabon’s rainforest—ecology, distribution and status. Oryx 22,218–228.
Hewett-Emmett, D. & Cook, C. N. (1978). Atypicalevolution of papionin �-haemoglobins and indicationthat Cercocebus may not be a monophyletic genus. In(D. J. Chivers & K. A. Joysey, Eds) Recent Advancesin Primatology, pp. 291–294. London: AcademicPress.
Hill, W. C. O. (1970). Primates: Comparative Anatomyand Taxonomy, Vol. VIII, Cynopithecinae: Papio,
Mandrillus, Theropithecus. Edinburgh: EdinburghUniversity Press.
Homewood, K. M. (1978). Feeding strategy ofthe Tana mangabey (Cercocebus galeritus galeritus)(Mammalia: Primates). J. Zool. Lond. 186, 375–391.
Horn, A. D. (1987). The socioecology of the blackmangabey (Cercocebus aterrimus) near Lake Tumba,Zaire. Am. J. Primatol. 12, 165–180.
Hoshino, J. (1985). Feeding ecology of mandrills(Mandrillus sphinx) in Campo Animal Reserve,Cameroon. Primates 26, 248–273.
Jablonski, N. G. (1993). The phylogeny ofTheropithecus. In (N. G. Jablonski, Ed.) Theropithecus:The Rise and Fall of a Primate Genus, pp. 209–224.Cambridge: Cambridge University Press.
Jolly, C. J. (1967). The evolution of the baboons.In (H. Vagborg, Ed.) The Baboon in MedicalResearch, Vol. II, pp. 23–50. Austin: University ofTexas Press.
Jolly, C. J. (1970). The large African monkeys as anadaptive array. In (J. R. Napier & P. H. Napier, Eds)Old World Monkeys; Evolution, Systematics, andBehavior, pp. 139–174. New York: Academic Press.
Jolly, C. J. (1972). The classification and naturalhistory of Theropithecus (Simopithecus) (Andrews,1916), baboons of the African Plio-Pleistocene. Bull.Br. Mus. (nat. Hist.), Geol. 22, 1–123.
Jones, C. & Sabater-Pi, J. (1968). Comparative ecologyof Cercocebus albigena (Gray) and Cercocebus torquatus(Kerr) in Rio Muni, West Africa. Folia Primatol. 9,99–113.
Kingdon, J. (1997). The Kingdon Field Guide to AfricanMammals. New York: Academic Press.
McGraw, W. S. (1996). The positional behavior andhabitat use of six sympatric monkeys in the TaiForest, Ivory Coast. Ph.D. Dissertation, SUNY atStony Brook.
Mitani, M. (1989). Cercocebus torquatus: adaptivefeeding and ranging behaviors related to seasonalfluctuations of food resources in the tropical rainforest of south-western Cameroon. Primates 30, 307–323.
Nakatsukasa, M. (1994a). Intrageneric variation oflimb bones and implications for positional behaviorin Old World monkeys. Z. Morph. Anthrop. 80,125–136.
Nakatsukasa, M. (1994b). Morphology of the humerusand femur in African mangabeys and guenons:functional adaptations and implications for theevolution of positional behavior. Afr. Study Monogr.21, 1–61.
Nakatsukasa, M. (1996). Locomotor differentiationand different skeletal morphologies in mangabeys(Lophocebus and Cercocebus). Folia primatol. 66,15–24.
Olupot, W., Chapman, C. A., Brown, C. H. & Waser,P. M. (1994). Mangabey (Cercocebus albigena) popu-lation density, group size, and ranging: a twenty yearcomparison. Am. J. Primatol. 32, 197–205.
Olupot, W., Chapman, C. A., Waser, P. M. & Isabirye-Basuta, G. (1997). Mangabey (Cercocebus albigena)ranging patterns in relation to fruit availability and

292 . . .
the risk of parasite infection in Kibale National park,Uganda. Am. J. Primatol. 43, 65–78.
Quiris, R. (1975). Ecologie et organisation sociale deCercocebus galeritus agilis dans le Nord-Est du Gabon.Terre et Vie 29, 337–398.
Range, F. (1998). Sozialsystem and Konkurrenztypweiblicher Raugrauer mangaben (Cercocebus torquatusatys). Masters Thesis, University Bayreuth,Germany.
Rogers, M. E., Abernathy, K. A., Fontaine, B.,Wickings, E. J., White, L. J. & Tutin, C. E. G.(1996). Ten days in the life of a mandrill horde in theLope Reserve, Gabon. Am. J. Primatol. 40, 297–313.
Ross, C. F., Lockwood, C. A., Fleagle, J. G. & Jungers,W. L. (2001). Adaptation and behavior in the pri-mate fossil record. In (J. M. Plavcan, C. P. vanSchaik, R. F. Kay & W. L. Jungers, Eds) Reconstruct-ing Behavior in the Primate Fossil Record, pp. 1–41.New York: Plenum.
Rowe, N. (1996). The Pictorial Guide to the LivingPrimates. East Hampton: Pogonias Press.
Rutte, C. (1998). Die Nahrungssuchstrategie derRauchgrauen Mangabe (Cercocebus torquatus atys).Masters Thesis, University Erlangen, Germany.
Schaaf, C. D., Butynski, T. M. & Hearn, G. W. (1990).The drill (Mandrillus leucophaeus) and other primatesin the Gran Caldera Volcanica de Luba: Results of asurvey conducted March 7–22, 1990. Unpublishedreport, Government of the Republic of EquatorialGuinea.
Shah, N. F. (1996). Preliminary study of two sympatricmangabey species (Cercocebus galeritus agilis and
Cercocebus albigena) in the Dzanga-Sangha Reserve,Central African Republic. XVIth Congress of theInternational Primatological Society.
Stanyon, R., Fantini, C., Camperio-Ciani, A.,Chiarelli, B. & Ardito, G. (1988). Banded karyo-types of 20 Papionini species reveal no necessarycorrelation with speciation. Am. J. Primatol. 16,3–17.
Szalay, F. S. & Delson, E. (1979). EvolutionaryHistory of the Primates. New York: AcademicPress.
Thorington, R. W. & Groves, C. P. (1970). An anno-tated classification of the Cercopithecoidea. In (J. R.Napier & P. H. Napier, Eds) Old World Monkeys:Evolution, Systematics and Behavior, pp. 629–648.New York: Academic Press.
Ward, C. V., Ruff, C. B., Walker, A., Teaford,M. F., Rose, M. D. & Nengo, I. O. (1995).Functional morphology of Proconsul patellas fromRusinga Island, Kenya, with implications for otherMiocene–Pliocene catarrhines. J. hum. Evol. 29,1–20.
Waser, P. (1977). Feeding, ranging and group size inthe mangabey C. albigena. In (T. H. Clutton-Brock,Ed.) Primate Ecology: Studies of Feeding and RangingBehavior in Lemurs, Monkeys and Apes, pp. 183–222.New York: Academic Press.
Waser, P. (1984). Ecological differences and behavioralcontrasts between two mangabey species. In (P. S.Rodman & J. G. H. Cant, Eds) Adaptations forForaging in Non-human Primates, pp. 195–216. NewYork: Columbia University Press.





























