Embed Size (px)
Citation preview

Appl Microbiol Biotechnol (1995) 43: 1044-1049
ORIGINAL PAPER
K.-P. Choi . I. Molnar' Y. Murooka
cg Springer-Verlag 1994
Secretoryoverproductionof Arthrobactersimplex3.ketosteroidA1. dehydrogenasebyStreptomyceslividanswitha multi.copyshuttlevector
Received: 29 August 1994jReceived last revision: 21 December 1994jAccepted: 23 January 1995
Abstract The gene for 3-ketosteroid ~ i-dehydrogenase(ksdD) of Arthrobacter simplex was expressed in Strep-tomyces lividans and the secreted enzyme was over-produced by using a multi-copy shuttle vector com-posed of pIJ702 and pUC19. Deletional analysis of therecombinant plasmid showed that the entire codingsequence of the ksdD gene was located within a 7-kbsegment of the chromosomal DNA obtained from theenzyme-producing strain of A. simplex. When S.lividans carrying the recombinant plasmid was grownin an appropriate medium, the cells produced about100-fold more 3-ketosteroid ~ i-dehydrogenase thanthe original strain. Although the percentage of enzymesecreted was changed during cultivation, a maximum55% of the enzyme was secreted into the culturedmedium of S. lividans, while A. simplex did not producethe enzyme extracellularly. Secretory overproductionof 3-ketosteroid ~ i-dehydrogenase in S. lividans wasalso identified by sodium dodecyl sulfate/polyacrylam-ide gel electrophoresis and on native gel, and the en-zyme reaction was confirmed by reverse-phase HPLCusing 4-androstene-3,17-dione as a substrate.
Introduction
3-Ketosteroid ~ i-dehydrogenase (KS1DH) [4-ene-3-oxosteroid:( acceptor) 1-ene-oxidoreductase, EC 1.3.99.4]catalyzes the reaction of 4-ene-3-oxosteroids to 1,4-diene derivatives by the trans-axial elimination of the
K.-P. Choi' Y. Murooka (B)Department of Biotechnology, Faculty of Engineering,Osaka University, Suita, Osaka 565, Japan.Fax: (81) 6879-7418
1. Molnar
Department of Biochemistry, University of Cambridge,Tennis Court Road, Cambridge CB2 lQW, UK
C1(1X)and C2([3) hydrogens (Itagaki et al. 1990b).KS1DH enzymes have been isolated and characterizedfrom many bacterial sources, including Pseudomonas,Nocardia, Rhodococus, Mycobacterium, and Arthrobac-ter (Levy and Talalay 1990; Sih and Bennet 1962;Itagaki et al. 1990a; Kaufmann et al. 1992; Wovchaet al. 1979; Penasse and Peyre 1968). These enzymes aremonomers of 56-62 kDa (Kaufmann et al. 1990; Plesiatet al. 1991), containing one mole of FAD per mole ofprotein as a prosthetic group, while the natural electronacceptor appears to be vitamin K2 (Abul-Hajj 1978),and they can transfer electrons to N-methylphenazoli-um sulfate (Penasse and Peyre 1968). KS1DH enzymescan be classified into two groups based on their abilitiesto use Cll-substituted steroids (for example, cortisoneand cortisol) as substrate: the enzyme fromPseudomonas testosteroni can not desaturate Cll hy-droxy- or oxosteroids (Levy and Talalay 1959), whileKS1DH from several gram-positive bacteria can de-saturate these substrates (Sih and Bennet 1962; Itagakiet al. 1990a).
Arthrobacter simplex, a gram-positive bacteriumshowing a rod/coccus growth cycle, has been extensive-ly used in the pharmaceutical industry to introduceC1-C2 double bonds into Cll-substituted steroids(Kondo 1962). The overproduction of KS1DH fromthis species is thus of considerable importance for theproduction of steroid drugs.
Streptomyces host-vector systems have generated in-terest as useful expression systems for the secretoryoverproduction of foreign proteins. These species nat-urally secrete a large amount of protein directly intothe culture fluid and the techniques for the cultivationof this organism are well established on an industrialscale (Molnar 1994). During the course of our studieson cholesterol metabolism in Streptomyces, weachieved the secretory overproduction of cholesteroloxidase from Streptomyces sp. in S. lividans (Molnar etal. 1991). Recently we have cloned the 3-ketosteroid~ i-dehydrogenase gene (ksdD) from A. simplex.

In this work, we investigated the secretory over-production of KS1DH from A. simplex in S. lividans bya multi-copy shuttle vector.
Materialsandmethods
Bacterial strains, plasmids and cultivation conditions
S. lividans TK64 (Hopwood et al. 1985) and Escherichia coli XUBlue (Bullock et al. 1987) were used as hosts for various plasmids. A.simplex IFO12069 is a producer of KSIDH. Plasmid pUC713 wascreated by converting the single Pstl site of plasmid vector pUC702(Molnar et al. 1991) into a unique BglII site by linker insertion, andcarries the lac promoter ofpUC19 (Yanisch et al. 1985), which is alsoactive in S. lividans. The pPDH series of plasmids were constructed,based on pUC713. Streptomyces and Arthrobacter were grown inyeast extract/malt extract (YEME) medium, trypticase/soy broth(TSB) medium, and Fructose/Bennett (FB)/agar medium (Hopwoodet al. 1985) supplemented with 10 ~g/ml (liquid cultures) or 50 ~g/ml(agar cultures) thiostrepton for the plasmid-containing strains.A 1O0-~g/ml sample of 4-androstene-3,17-dione was added as aninducer if applicable. Streptomyces or Arthrobacter strains weregrown at 28°C with shaking in 10 ml medium (test-tube) and 80 mlmedium (Sakaguchi flask). Luria Bertani (LB) medium (Sambrook etal. 1989) supplemented with ampicillin and tetracycline, if necessary,was used for E. coli strains.
Materials
Restriction endonucleases, T4 DNA ligase, the large fragment ofDNA polymerase I and RNase A were purchased from TakaraShuzo Co. Ltd. (Kyoto, Japan), or Toyobo Co. Ltd. (Osaka, Japan).A Gene-Clean DNA purification kit was obtained from Bio 101 Inc.(LaJolla, Calif, USA). Thiostrepton was provided by Asahi KaseiCo. Ltd. (Tokyo, Japan). Phenazine methosulfate and nitrotet-razolium blue were purchased from Nacalai Tesque Ltd. (Kyoto,Japan). Steroids used as substrates or chromatographic standardswere 4-androstene-3,17-dione and 1,4-androstadiene-3,17-dione(from Sigma Chemical Co., St. Louis, Mo., USA).
Recombinant DNA technology
Recombinant DNA techniques for E. coli and Streptomyces werecarried out according to Sambrook et al. (1989), and Hopwood et al.(1985) respectively.
KSIDH assay
KSIDH was assayed by following the increase of absorbance at570 nm reflecting the formation of diformazan (Chiriacy 1975).4-Androstene-3,17-dione (100 mM dissolved in ethanol) was addedto reaction mixtures containing 66.7 mM TRIS/HCI (pH 8.5),150 ~M phenazine methosulfate, 550 ~M nitrotetrazolium blue, andan appropriate amount of enzyme for a final volume of 3 ml. Oneunit of KSIDH was defined as the amount of enzyme needed to form1 ~mol diformazan in 1 min at 28°C. This formation rate of difor-mazan is equivalent to that of 1 ~mol 1,4-androstadiene-3,17 -dionefrom 4-androstene-3,17 -dione. The cells were harvested and washedtwice in buffer (0.03 M TRIS pH 7.5, 0.1 mM EDTA, 0.1 mMdithiothreitol). The washed cells were sonicated and centrifuged toobtain intracellular enzyme. The resultant precipitates were used for
determining membrane-bound enzyme activity. The culture super-natant was used for determining extracellular enzyme activity.
In vitro steroid transformation
4-Androstene-3,17 -dione (300 mmol) was transformed in an in vitroreaction by appropriate dilutions of extra- or intracellular proteinextracts, in reaction mixtures containing 66.7 mM TRIS/HCI (pH8.5) and 15 ~M phenazine methosulfate as the electron acceptor withincubation at 37°C for 30 min. The reactions were stopped byboiling at 5 min, and the reaction mixtures were extracted by ethylacetate. After the solvent had been evaporated, the residue wasdissolved in 0.1 ml 50% acetonitrile solution (v/v distilled water).Analysis was done by reverse-phase HPLC on a Japan Spectro-scopic Co. (Tokyo, Japan) chromatograph, model 88-PU, with anODS-2 column (GL Sciences) at 240 nm using 50% acetonitrilesolution as the eluent.
Electrophoresis of protein
Sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE) was performed in 7.5% (w/v) polyacrylamide slab gels usingthe TRIS/glycine buffer system (Laemmli 1970), and the molecularmass of standard proteins was a low-molecular-mass standard kitobtained from Pharmacia Co. (Uppsala, Sweden). Native PAGEwas performed in 12.5% (w/v) polyacrylamide slab gels according tothe method of Laemmli (1970) except that SDS and 2-mercap-toethanol were omitted. The gels were stained either for total pro-teins with Coomassie brilliant blue G250, or for enzymatic activitywith 0.1 mM phenazine methosulfate, 0.5 mM nitrotetrazolium blueand 0.1 mM 4-androstene-3,17-dione in 66.5 mM TRIS/HCI buffer(pH 8.5). After developing a blue color, the reaction of gel forenzymatic activity was stopped by 0.2 M acetic acid.
Results
Deletional analysis of the 3-ketosteroidL11-dehydrogenase gene
Recently we cloned the 3-ketosteroid L11-dehydrogen-ase gene (ksdD) from A. simpex into S. lividans (Molnaret al. 1995). The plasmid pPDH1 (Fig. 1) of S. lividansstrain PDH1 carries an insert of 9.1 kb cloned at theBglII site of pUC713. Deletional analysis localized thegenetic determinant for KS1DH on the 7-kb genomicDNA fragment, and indicated that BamHI 1 and SphI2 are within the open-reading frame for KS1DH.
Production of KS1DH by S. lividans strains carryingrecombinant plasmids
S. lividans TK64 cells carrying plasmids were grownaerobically at 28°C for 5 days in 10 ml YEME and TSBmedium, supplemented with 10 J.lg/ml thiostrepton.The original producing strain, A. simplex, was culti-vated in the same conditions without thiostrepton for1 day in YEME, and 2 days in TSB medium with orwithout 4-androstene-3,17-dione induction. In S.

1046
BglIllSau 3AI \~BglIlISau3AI
pPDHl i1B
~
2
Sp
~ P6
P3
pPDH2
pPDH3 I
pPDH4 I
pPDHS
pPDH7 I
Fig. 1 Restriction endonuclease map of pPDHI and its derivativeplasmids. Deletion plasmids were isolated in E. coli and transformedto S. lividans TK64 for detection of the 3-ketosteroid ~ I-dehydro-genase (KSIDH) activity. The open bar indicates an insertion frag-ment from A. simplex. The thick line indicates the region for theexpression of KSIDH+ phenotype. Tsr thiostrepton-resistancegene; B BamBI, Sp SphI, H HindIII. +, - Activity of KSIDH
lividans TK64 cells carrying pUC713 or A. simplexwithout 4-androstene-3,17-dione, no enzyme activitieswere detected, whereas S. lividans TK64 carryingpPDH1 or pPDH3 and A. simplex with 4-androstene-3,17-dione produced KS1DH (Table 1). The highestenzyme activity was observed intracellularly when S.lividans cells carrying pPDH3 were grown in YEMEmedium. Interestingly, when S. lividans (pPDH3) weregrown in TSB medium, although the total enzymeproduction was almost the same in YEME medium,about 41 % of KS1DH was produced extracellularly. InS. lividans carrying pPDH1 or pPDH3, the mem-brane-bound KS1DH was not detected, whereas A.simplex with 4-androstene-3,17-dione produced about30% of the membrane-bound enzyme and 70% of theintracellular enzyme. The total production of KS1DHby S. lividans carrying pPDH3 in YEME or TSB me-dium increased about 100-fold more than that of A.simplex. The cells carrying pPDH1 produced less
Table 1 Enzyme activities of 3-ketosteroid ~ I-dehydrogenase invarious strains Cells were grown and assayed as described in Mater-ials and methods. (Extra extracellular enzyme, Intra intracellularenzyme, Membrane membrane-bound enzyme, ND not detected,Y EM E yeast extract/malt extract, TSB trypticase/soy broth)
Strain
A. simplex(+ 4-AD)aS. lividans
a 4-Androstene-3,17-dione added
+9
~7
.....--------.....--
/
, /" /" /'-_/5
5
~4u>--€;'03'".§~2J:Cl
iiil~
3
E2"0,
£Q;
1 u>-0
000 4 53
Days
2
Fig.2 Production of the intracellular and membrane-boundKSIDH in the producing strain A. simplex. The cells were grown inyeast extract/malt extract (YEME; DOO) or trypticase/soy broth(TSB; .A..e) medium with 4-androstene-3, 17-dione at 28°C. D.A.Intracellular activity, oe membrane-bound activity, O. cellgrowth; --- pH in YEME medium; -, pH in TSB medium
KS1DH than did those with pPDH3. We found thatpPDH1 was unstable during cultivation.
Time course of KS1DH production in S. lividanscarrying pPDH3
The time courses of KS1DH production by A. simplexwith 4-androstene-3,17-dione and S. lividans TK64(pPDH3) in two media are shown in Figs. 2, 3. When A.
KS1DH
3 4 5 "H B B
I :I I +
9.1kb
Plasmid Medium Enzyme activity(mU/mg dry cells)
Extra In tra Membrane
YEME ND 4 1.7TSB ND 3.4 1.7
pUC713 YEME ND ND NDTSB ND ND ND
pPDHI YEME ND 81 NDTSB 34 49 ND
pPDH3 YEME 1.3 600 NDTSB 210 307 ND

9
:r: 7c.
~~
/'~" .-" .-" /'- - --_/5
800
0
4
'"u>--uaC'>..§ 400=>
S
..§C'>E
2=a:;u>-0:r:
0(j):.::
00 3 4
Days
Fig.3 Production of the extracellular and intracellular KSIDH inS. /ividans (pPDH3). The cells were grown in YEME (OLD) or TSB(.~.) mediumat 28°C. O. Extracellular activity, L~ intracellu-lar activity,O. cellgrowth,--- pH in YEME medium; - pH inTSB medium
2 5 6
simplex cells were cultured in YEME medium with4-androstene-3,17-dione, production of the intracellu-lar and membrane-bound KS1DH increased at 1 dayand then decreased (Fig. 2). When A. simplex was cul-tured in TSB medium with 4-androstene-3,17-dione,production of the intracellular KS1DH increased at1 day and after 2.5 days the enzyme activity decreased.Maximum production of intracellular (4 mU/mg drycells) and membrane-bound (1.8 mU/mg dry cells)KS1DH enzymes was obtained during cultivation inYEME medium. However, no extracellular enzyme wasdetected (Fig. 2).
In S. lividans cells carrying pPDH3 cultured inYEME medium (Fig. 3), production of the intracellularKS1DH started at 2 days and after 5 days the enzymeactivity sharply decreased, whereas only a smallamount of extracellular enzyme was produced betweendays 2 and 5. In TSB medium (Fig. 3), production ofthe intracellular KS1DH occurred for 3-4 days andthen decreased, while production of the extracellularKS1DH was observed for 3-5 days and subsequentlydecreased. Highest activities of the intracellular andextracellular KS1DH were about 800 mU/mg and300 mU/mg dry cells respectively. Throughout the ex-periment, approximately 23%-55% of the total activ-ity of KS1DH was secreted into the culture broth.
Analysis of the extracellular products from S. lividanscarrying pPDH3 by SDS-PAGE and native gel
Secretory overproduction of KS1DH in S. lividans(pPDH3) was identified by SDS-PAGE (Fig.4A) and
1047
4 5 6 4' 5' 6'
kDa94 ~
c: KS10H67 ~
43 ~
A B c
Fig.4A-C Analysis of extracellular proteins in various strains bySDS-PAGE and native gel. A SDS-PAGE. Lane 1,A. simplex; lane 2,S. /ividans (pUC713); lane 3, S. lividans (pPDH3). B Native gelelectrophoresis stained with Coomassie brilliant blue. Lane 4, S.lividans (pUC713); lane 5, S. /ividans (pPDH3); lane 6, A. simplex.C Native gel electrophoresis stained by phenazine methosulfate andnitrotetrazolium blue. Lane 4', S. lividans (pUC713); lane 5', S./ividans (pPDH3); lane 6', A. simplex
native gel (Fig. 4B, C). The molecular mass of KS1DHfrom A. simplex was calculated to be 54329Da accord-ing to the DNA sequence (Molnar et al. 1995). Theextracellular KS1DH produced by S. lividans (pPDH3)was observed in SDS-PAGE at about the 55-kDa posi-tion (Fig. 4A, lane 3), however no such protein bandwas observed in the supernatant of S. lividans carryingpUC713 (Fig. 4A, lane 2) or A. simplex (Fig. 4A, lane 1).In native PAGE, the extracellular KS1DH was detec-ted (Fig. 4B, lane 5) and the enzyme activity was alsoshown by active staining (Fig. 4C, lane 5').
Identification of the extracellular and intracellularKS1DH by HPLC
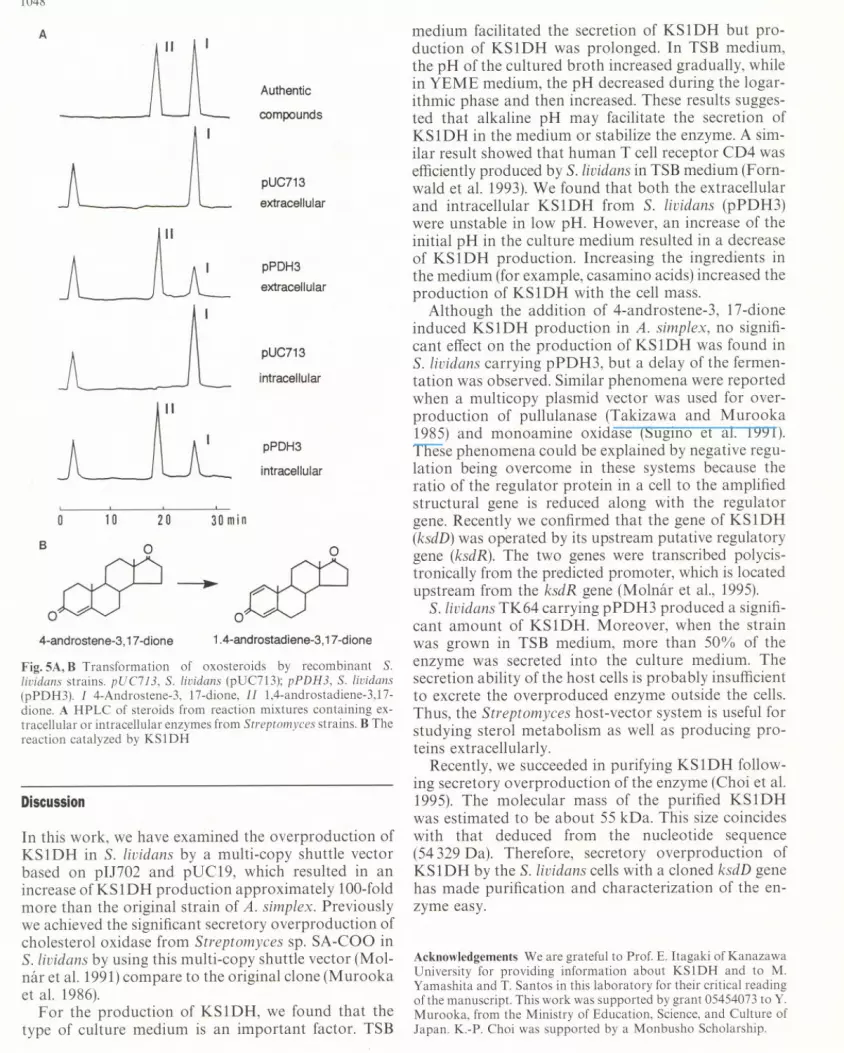
Extracellular (100 mU/ml) and intracellular KS1DH(100 mU/ml) were incubated with the 4-androstene-3,17-dione at 37°C for 30 min, and the reaction mixtureswere analyzed by HPLC. The retention time of auth-entic 4-androstene-3, 17-dione was 26.5 min and thatof authentic 1,4-androstadiene-3,17-dione 19.2 min(Fig. 5A). In the reaction mixtures of the extracellularand intracellular KS1DH from S. lividans (pPDH3), thesame peaks were observed but not in S. lividans(pUC713). From these results, we concluded that theextracellular and intracellular KS1DH enzyme catalyzethe formation of 1,4-androstadiene-3, 17-dione from4-androstene-3, 17-dione in reactions in vitro.
lU'I1\
A
~L-lLk~JLlL
10 20 30min
Authentic
compounds
pUC713extracellular
pPDH3extracellular
pUC713
intracellular
pPDH3
intracellular
a
B asP -. /X)Pa4-androstene-3,17 -dione 1.4-androstadiene-3,17-dione
Fig. SA, B Transformation of oxosteroids by recombinant S.lividans strains. pUC713, S. lividans (pUC713); pPDH3, S. lividans(pPDH3). I 4-Androstene-3, 17-dione, II 1,4-androstadiene-3,17-dione. A HPLC of steroids from reaction mixtures containing ex-tracellular or intracellular enzymes from Streptomyces strains. B Thereaction catalyzed by KS1DH
Discussion
In this work, we have examined the overproduction ofKSIDH in S. lividans by a multi-copy shuttle vectorbased on pIn02 and pUCI9, which resulted in anincrease ofKSIDH production approximately 1O0-foldmore than the original strain of A. simplex. Previouslywe achieved the significant secretory overproduction ofcholesterol oxidase from Streptomyces sp. SA-COO inS. lividans by using this multi-copy shuttle vector (Mol-nar et aI. 1991) compare to the original clone (Murookaet aI. 1986).
For the production of KSIDH, we found that thetype of culture medium is an important factor. TSB
medium facilitated the secretion of KSIDH but pro-duction of KSIDH was prolonged. In TSB medium,the pH of the cultured broth increased gradually, whilein YEME medium, the pH decreased during the logar-ithmic phase and then increased. These results sugges-ted that alkaline pH may facilitate the secretion ofKSIDH in the medium or stabilize the enzyme. A sim-ilar result showed that human T cell receptor CD4 wasefficiently produced by S. lividans in TSB medium (Forn-wald et aI. 1993). We found that both the extracellularand intracellular KSIDH from S. lividans (pPDH3)were unstable in low pH. However, an increase of theinitial pH in the culture medium resulted in a decreaseof KSIDH production. Increasing the ingredients inthe medium (for example, casamino acids) increased theproduction of KSIDH with the cell mass.
Although the addition of 4-androstene-3, 17-dioneinduced KSIDH production in A. simplex, no signifi-cant effect on the production of KSIDH was found inS. lividans carrying pPDH3, but a delay of the fermen-tation was observed. Similar phenomena were reportedwhen a multicopy plasmid vector was used for over-production of pullulanase (Takizawa and Murooka1985) and monoamine oxidase (Sugino et aI. 1991).These phenomena could be explained by negative regu-lation being overcome in these systems because theratio of the regulator protein in a cell to the amplifiedstructural gene is reduced along with the regulatorgene. Recently we confirmed that the gene of KSIDH(ksdD)was operated by its upstream putative regulatorygene (ksdR). The two genes were transcribed polycis-tronically from the predicted promoter, which is locatedupstream from the ksdR gene (Molnar et aI., 1995).
S. lividans TK64 carrying pPDH3 produced a signifi-cant amount of KSIDH. Moreover, when the strainwas grown in TSB medium, more than 50% of theenzyme was secreted into the culture medium. Thesecretion ability of the host cells is probably insufficientto excrete the overproduced enzyme outside the cells.Thus, the Streptomyces host-vector system is useful forstudying sterol metabolism as well as producing pro-teins extracellularly.
Recently, we succeeded in purifying KSIDH follow-ing secretory overproduction of the enzyme (Choi et aI.1995). The molecular mass of the purified KSIDHwas estimated to be about 55 kDa. This size coincideswith that deduced from the nucleotide sequence(54329 Da). Therefore, secretory overproduction ofKSIDH by the S. lividans cells with a cloned ksdD genehas made purification and characterization of the en-zyme easy.
Acknowledgements We are grateful to Prof. E. Itagaki of KanazawaUniversity for providing information about KS1DH and to M.Yamashita and T. Santos in this laboratory for their critical readingof the manuscript. This work was supported by grant 05454073 to Y.Murooka, from the Ministry of Education, Science, and Culture ofJapan. K.-P. Choi was supported by a Monbusho Scholarship.

References
Abul-Hajj Y (1978) Isolation of vitamin k2 (35) from Nocardiarestrictus and Corynebacterium simplex. J Bioi Chern253:2356-2360
Bullock WO, Fernandez JM, Short JM (1987) A high efficiencyplasmid transforming recA Escherichia coli strain with betagalac-tosidase selection. Biotechniques, 5:376
Chiriacy M (1975) Genetics of alcohol dehydrogenase in Sacchar-omyces cerevisiae. I. Isolation and genetic analysis of adh mu-tants. Mutat Res 29:315-326
Choi KP, Molnar I, Yamashita M, Murooka Y (1995) Purificationand characterization of the 3-ketosteroid-~' -dehydrogenase ofArthrobacter simplex produced in Streptomyces /ividans. J Bio-chern 115:1043-1049
Fornwald JA, Donovan MJ, Gerber R, Keller J, Taylor DP, ArcuriEJ, Brawner ME (1993) Soluble forms of the human T cellreceptor CD4 are efficiently expressed by Streptomyces lividans.Biotechnology 11: 1031-1036
Hopwood DA, Bibb MJ, Charter KF, Kieser T, Thompson CJ,Kieser HM, Lydiate DJ, Schrempf H (1985) Genetic manipula-tion of Streptomyces, a laboratory manual. John Innes Founda-tion, Norwich, UK
Itagaki E, Wakabayashi T, Hatta T (1990a) Purification and charac-terization of 3-ketosteroid-~I-dehydrogenase from Nocardiacorallina. Biochim Biophys Acta 1038:60-67
Itagaki E, Hatta T, Wakabayashi T, Suzuki K (1990b) Spectralproperties of 3-ketosteroid-~I-dehydrogenase from Nocardiacolallina. Biochim Biophys Acta 1040:281-286
Kaufmann G, Schuman G, Wollweber L, Huller E, Atrat P (1990)Steroid-l-dehydrogenases in nocardioform bacteria studied byelectrophoresis and immunoblotting techniques. J Basic Micro-biaI30:415-423
Kaufmann G, Thole H, Kraft R, Atrat P (1992) Steroid-I-de-hydrogenase of Rhodococus erythropo/is: purification and N-terminal amino acid sequence. J Steroid Biochem Mol Bioi43:297-301
Kondo E (1962) Steroid I-dehydrogenation by a crude enzymepreparation from Arthrobacter simplex. Agric Bioi Chern27:69-70
Laemmli UK (1970) Cleavage of structural proteins during assemblyof the head of bacteriophage T4. Nature 224:680-685
Levy H, Talalay P (1959) Bacterial oxidation of steroids I:ring A dehydrogenations by intact cells. J Bioi Chern234:2009-2013
1049
Molnar I (1994) Secretory production of homologous and hetero-logous proteins by recombinant Streptomyces: what has beenaccomplished') In: Murooka Y, Imanaka T (eds) Recombinantmicrobes for industrial and agricultural applications. Dekker,New York, pp 81-104
Molnar I, Choi KP, Hayashi N, Murooka Y (1991) Secretory over-production of Streptomyces cholesterol oxidase by Streptomyceslividans with a multi-copy shuttle vector. J Ferment Bioeng72:368-372
Molnar I, Choi KP, Yamashita M, Murooka Y (1995) Molecularcloning, expression in Streptomyces /ividans, and nucleotide se-quence analysis of a gene cluster from Arthrobacter simplexencoding 3-ketosteroid-~ I-dehydrogenase, 3-ketosteroid-~ 5-isomerase, and a hypothetic regulatory protein. Mol Microbiol15:895-905
Murooka Y, Ishizaki T, Nimi 0, Maekawa N (1986) Cloning andexpression of a Streptomyces cholesterol oxidase gene in Strepto-myces lividans with plasmid plJ702. Appl Environ Microbiol52: 1382-1385
Penasse L, Peyre M (1968) Studies of 3-oxo steroid ~ I-oxydo reduc-tase of Arthrobacter simplex. Steroids 12:525-544
Plesiat P, Grandquillot H, Harayama S, Vragar S, Michael-BriandY (1991) Cloning, sequencing, and expression of the Pseudomonastestosteroni gene encoding 3-oxosteroid-~ I-dehydrogenase.J BacterioI173:7219-7229
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning:a laboratory manual, 2nd edn. Cold Spring Harbor Laboratory,Cold Spring Harbor, NY
Sih CJ, Bennet RE (1962) Steroid-I-dehydrogenation of Nocardiarestrictus. Biochim Biophys Acta 56:584-592
Sugino H, Ishibashi K, Sakaue M, Yamashita M, Murooka Y (1991)Gene cloning of the maoA gene and overproduction of a solublemonoamine oxidase from Klebsiella aerogenes. Appl MicrobiolBiotechnoI35:606-610
Takizawa N, Murooka Y (1985) Cloning of the pullulanase gene andoverproduction of pullulanase in Escherichia coli and Klebsiellaaerogenes. Appl Environ Microbiol 49:294-298
Wovcha MG, Brook KE, Komonek LA (1979) Evidence for twosteroid 1, 2-dehydrogenase activities in Mycobacteriumfortuitum.Biochym Biophys Acta 574:471-479
Yanisch C, Viera J, Messing J (1985) Improved M13 phage cloningvectors and pUC19 vectors. Gene 33: 103-119













![A new type of glycolipid, 1-[α-mannopyranosyl-(1α-3)-(6-O-acyl-α-mannopyranosyl)]-3-O-acylglycerol, from Arthrobacter atrocyaneus](https://img.dokumen.tips/doc/110x75/636047f0fa7cd8ab3f0d3549/a-new-type-of-glycolipid-1-mannopyranosyl-1-3-6-o-acyl-mannopyranosyl-3-o-acylglycerol.jpg)





