Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, May 2004, p. 2576–2585 Vol. 186, No. 90021-9193/04/$08.00�0 DOI: 10.1128/JB.186.9.2576–2585.2004Copyright © 2004, American Society for Microbiology. All Rights Reserved.
ScaC, an Adaptor Protein Carrying a Novel Cohesin That Expands theDockerin-Binding Repertoire of the Ruminococcus flavefaciens
17 CellulosomeMarco T. Rincon,1* Jennifer C. Martin,1 Vincenzo Aurilia,2 Sheila I. McCrae,1 Garry J. Rucklidge,3
Martin D. Reid,3 Edward A. Bayer,4 Raphael Lamed,5 and Harry J. Flint1
Microbial Genetics Group1 and Proteomics Unit,3 Rowett Research Institute, Aberdeen, United Kingdom; Institute of ProteinBiochemistry, National Research Council, Naples, Italy2; and Department of Biological Chemistry, The Weizmann Institute
of Science, Rehovot,4 and Department of Molecular Microbiology and Biotechnology, Tel Aviv University,Ramat Aviv,5 Israel
Received 25 November 2003/Accepted 26 January 2004
A new gene, designated scaC and encoding a protein carrying a single cohesin, was identified in thecellulolytic rumen anaerobe Ruminococcus flavefaciens 17 as part of a gene cluster that also codes for thecellulosome structural components ScaA and ScaB. Phylogenetic analysis showed that the sequence of the ScaCcohesin is distinct from the sequences of other cohesins, including the sequences of R. flavefaciens ScaA andScaB. The scaC gene product also includes at its C terminus a dockerin module that closely resembles thosefound in R. flavefaciens enzymes that bind to the cohesins of the primary ScaA scaffoldin. The putative cohesindomain and the C-terminal dockerin module were cloned and overexpressed in Escherichia coli as His6-taggedproducts (ScaC-Coh and ScaC-Doc, respectively). Affinity probing of protein extracts of R. flavefaciens 17separated in one-dimensional and two-dimensional gels with recombinant cohesins from ScaC and ScaArevealed that two distinct subsets of native proteins interact with ScaC-Coh and ScaA-Coh. Furthermore,ScaC-Coh failed to interact with the recombinant dockerin module from the enzyme EndB that is recognizedby ScaA cohesins. On the other hand, ScaC-Doc was shown to interact specifically with the recombinantcohesin domain from ScaA, and the ScaA-Coh probe was shown to interact with a native 29-kDa protein spotidentified as ScaC by matrix-assisted laser desorption ionization—time of flight mass spectrometry. Theseresults suggest that ScaC plays the role of an adaptor scaffoldin that is bound to ScaA via the ScaC dockerinmodule, which, via the distinctive ScaC cohesin, expands the range of proteins that can bind to the ScaA-basedenzyme complex.
Since the first description of cellulosomes in the anaerobicthermophile Clostridium thermocellum (3, 5), enzyme com-plexes responsible for plant cell wall breakdown have beenfound in many species of cellulolytic gram-positive bacteria (2).In general, different species of clostridia have fairly minorvariations of the paradigm established with C. thermocellum, inwhich a noncatalytic scaffolding protein (the primary scaffol-din), which carries multiple cohesin domains and a cellulose-binding module, provides the framework for assembling arange of different enzymes. Enzyme subunits belonging to thecellulosome each carry a single type of dockerin that pairsspecifically with the cohesin domains. Attachment of the com-plex to the bacterial cell surface occurs either via hydrophilicdomains in the scaffolding proteins, as in mesophilic clostridia(6, 15, 18, 31), or via a C-terminal dockerin that links theprimary scaffoldin to one of several cell wall anchoring proteins(anchoring scaffoldins) (16) via a second type of cohesin-dock-erin specificity, as in C. thermocellum (4, 21, 22).
Investigations with organisms that do not belong to the ge-nus Clostridium are, however, now revealing some potentiallymore fundamental variations in the organization of cellulo-
some complexes. For example, the cellulosome system of Bac-teroides cellulosolvens has a two-component scaffoldin arrange-ment similar to that of C. thermocellum, except that the typesof cohesins carried by the primary and anchoring scaffoldinsare reversed (13, 33). In Acetivibrio cellulolyticus, the numberof enzymes incorporated into the cellulosome complex is am-plified by the involvement of a multi-cohesin-bearing adaptorscaffoldin which mediates between the primary and anchoringscaffoldins (12, 34).
Ruminococcus flavefaciens is a major cellulolytic species inthe rumen of herbivores (19, 25). This bacterium produces acomplex in which many plant cell wall-degrading enzymes areattached to a structural protein, ScaA, via specific dockerininteractions with the three conserved cohesin repeats of ScaA(28). ScaA, however, lacks an identifiable cellulose-bindingmodule (CBM), although a CBM was identified in the cellulaseEndB (29). In addition, previous studies revealed at least twodistinct classes of dockerins among enzymes from R. flavefa-ciens 17 (1), and only one of these groups of dockerins, foundin the EndB, EndA, XynB, and XynD enzymes, interacts withScaA (28). ScaA interacts via its C-terminal dockerin with alarger protein, ScaB, which contains seven distinctive cohesinrepeats and is believed to mediate the anchoring of the cellu-losome complex to the cell surface via an as-yet-unknownmechanism (14, 28). The scaA and scaB genes are locatedadjacent to each other in the chromosome.
* Corresponding author. Mailing address: Microbial GeneticsGroup, The Rowett Research Institute, Greenburn Road, Bucksburn,Aberdeen AB21 9SB, United Kingdom. Phone: 44-(0)1224-712751.Fax: 44-(0)1224-716687. E-mail: [email protected].
2576
on January 19, 2015 by guesthttp://jb.asm
.org/D
ownloaded from

In our quest for novel cohesin-dockerin interactions amongproteins expressed by R. flavefaciens 17, we found a new struc-tural protein encoded by a gene upstream of the scaA gene.This protein, designated ScaC, contains a unique cohesin do-main with a binding specificity distinct from that of ScaA andScaB cohesins, as well as a dockerin module that interacts withScaA. We propose a role for the newly characterized ScaCprotein as an adaptor molecule that greatly expands the rep-ertoire of proteins able to participate in the R. flavefacienscellulosome.
MATERIALS AND METHODS
Strains and growth conditions. R. flavefaciens 17 was grown anaerobically (8)either in modified M2 medium (26) or in Hungate-Stack medium (17) with 0.4%microcrystalline cellulose (Avicel PH101; Honeywill & Stein, London, UnitedKingdom), 0.4% birchwood xylan (Sigma, St. Louis, Mo.), or 0.4% cellobiose(Sigma) as an energy source, as described previously (28). E. coli Solopack GoldXL10 and BL21(DE3) (supplied by Stratagene, La Jolla, Calif.) were used ashosts for transformation and protein expression, respectively, with constructsmade in pET30 Ek/LIC (Novagen, Madison, Wis.). E. coli strains were routinelygrown on Luria-Bertani medium with appropriate antibiotic selection.
Sequence of scaC. The scaC coding sequence was obtained from PCR exten-sion of the scaA gene with the primers listed in Table 1 (Fig. 1). The reverseScaAcoh9r and M13 forward (specific for the vector sequence) primers yieldeda 1.8-kb DNA fragment from a pUC18 library of R. flavefaciens 17 chromosomalDNA partially digested with Sau3AI (28). Sequencing of the 1.8-kb DNA frag-ment revealed a new open reading frame. The scaC gene sequence was com-pleted in both DNA strands by using the additional internal primers ScaAcoh10r,ScaAcoh11r, and ScaAcoh10f.
Cloning and expression of ScaC His6-tagged protein constructs. The putativemature peptide of ScaC (ScaC-mat) after removal of the signal peptide (aminoacids 30 to 308; GETVQ-LIGVKT), the cohesin domain (ScaC-Coh) (aminoacids 30 to 207; GETVQ-NPQPQVT), and the C-terminal dockerin domain(ScaC-Doc) (amino acids 197 to 308; VTVGN-LIGVKT) were all cloned in thepET30 Ek/LIC expression vector (Novagen) after PCR amplification of the DNAfragments with primers designed to contain 14 or 15 bp of sequence specific forthe ligation-independent cloning site of the vector (see Table 1 for primerdetails). After amplification the DNA product was resolved in an agarose gel andpurified with a QIAquick gel extraction kit (Qiagen, Hilden, Germany) usedaccording to the manufacturer’s instructions. The DNA fragment was thentreated with 5 U of T4 DNA polymerase (Promega, Madison, Wis.) in a 100-�l(total volume) mixture containing 2.5 mM dATP, 1 mM dithiothreitol (DTT), 33mM Tris-acetate (pH 7.9), 66 mM potassium acetate, 10 mM magnesium acetate,and 0.1 mg of bovine serum albumin ml�1. The reaction mixture was incubatedat 37°C for 1 h and heat inactivated at 70°C for 15 min. The T4 DNA polymerase-
treated DNA fragment was then purified and concentrated with a QIAquickPCR purification kit (Qiagen). Ligation was carried out by using the pET30Ek/LIC vector as recommended by the manufacturer, and the resulting recom-binant plasmid was transformed in E. coli SoloPack Gold XL10 chemicallycompetent cells, plated on Luria-Bertani agar supplemented with 30 �g ofkanamycin ml�1, and incubated at 37°C for 16 h.
Double His6-S-tagged proteins from pET30 Ek/LIC constructs were overex-pressed following transformation in E. coli BL21(DE3). Cells were recoveredand lysed by sonication (Soniprep; Sanyo) after growth at 37°C on Luria-Bertanimedium supplemented with 1% (vol/vol) glycerol, 1% (wt/vol) glucose, and 30 �gof kanamycin ml�1. Induction was carried out by adding 0.5 mM IPTG (isopro-pyl-�-D-thiogalactopyranoside) to a culture at an optical density at 600 nm of 0.8to 1.0 and then incubating the culture either at 37°C for 4 h or at 16°C for 16 hin a refrigerated incubator. Protein constructs were purified by nickel affinitychromatography as described previously (14)
Cell surface-associated protein extraction from R. flavefaciens. Protein extractsused in this study were prepared by the method of Devillard et al. (11), with somemodifications. Briefly, bacterial cell pellets were harvested from a 200-ml cultureof R. flavefaciens at the stationary phase by centrifugation at 10,000 � g for 10min at 4°C. The pellet was washed twice with 200 ml of TBS buffer (25 mMTris-HCl buffer [pH 7.0], 150 mM NaCl) and resuspended in 10 ml of TBS buffercontaining 2% (wt/vol) N-laurylsarcosine. Proteases were inhibited by addition of1 mM phenylmethylsulfonyl fluoride and 250 �l of reconstituted protease inhib-itor cocktail (Sigma). After incubation for 1 h in ice, the cell suspension wasvortexed briefly and centrifuged at 15,000 � g for 15 min at 4°C. The supernatantfluids (cell surface-associated protein fraction) were then collected and ultracen-trifuged at 150,000 � g for 1 h at 4°C (L8-70 M ultracentrifuge; BeckmanCoulter, Buckinghamshire, United Kingdom). The supernatant was collected,and proteins were concentrated by ultrafiltration (VivaSpin6; 10,000-Da cutoff;VivaScience, Lincoln, United Kingdom)
Western blotting and detection of cohesion-dockerin interaction. Protein ex-tracts were separated by sodium dodecyl sulfate-polyacrylamide gel electro-phoresis (SDS-PAGE) by the Laemmli method (20). After electrophoresis, pro-teins were blotted onto a polyvinylidene difluoride (PVDF) membrane(Immobilon-P; Millipore, Bedford, United Kingdom) by following the manufac-turer’s instructions and using a semidry blotting system (Biometra, Gottingen,Germany) at 0.8 mA/cm2. Membranes were blocked for 1 h at room temperatureby rocking in 50 ml of blocking buffer (TBS buffer [pH 7.0] containing 3%[wt/vol] skim milk, 0.05% [vol/vol] Tween 20, and 1 mM CaCl2). Incubation withthe recombinant His6-tagged protein probes was performed in the same blockingbuffer at 4°C with rocking overnight. The following day the membranes werewashed three times in CaTTBS buffer (TBS buffer with 0.05% [vol/vol] Tween 20and 1 mM CaCl2) and then incubated for 1 h with a 1:5,000 dilution of nickel-activated horseradish peroxidase (HRP) (India HisProbe HRP; Pierce, Rock-ford, Ill.) in CaTTBS buffer containing 1% skim milk. The bands were visualizedby enhanced chemiluminescence (Supersignal West Pico chemiluminescent sub-strate; Pierce) by following the manufacturer’s instructions after the membraneswere washed five times at 5-min intervals in CaTTBS buffer. Spot tests were
TABLE 1. Oligonucleotide primers used in this study
Primer Sequencea Domain amplifiedb
ScaAcoh9r GGAGCTACCACGCTAAC ScaA N-terminal X domain (rev)ScaAcoh10f GCTGTTAAGTATGACAGCAGC ScaC cohesin domain (fwd)ScaAcoh10r TCATCAACATCGCCATTTG ScaC dockerin domain (rev)ScaAcoh11r GCCTGAATGCCGGAAGATGG ScaC cohesin domain (rev)M13f GTAAAACGACGGCCAGT pUC18-specific sequence (fwd)ScaCstart/LIC-f GACGACGACAAGATGGGCGAAACAGTGCAGATATC ScaC cohesin domain (fwd)ScaCEnd/LIC-r GAGGAGAAGCCCGGTTTAGGTTTTCACGCCTATTAATA ScaC dockerin domain (rev)ScaCCoh/LIC-r GAGGAGAAGCCCGGTTTAAGTAACCTGAGGCTGCGGA ScaC cohesin domain (rev)ScaCDoc/LIC-f GACGACGACAAGATGGTAACAGTGGGTAATCCGCA ScaC dockerin domain (fwd)XynXDoc/LIC-f GACGACGACAAGATGCCTGTTGGATTCTATATAGAT XynX dockerin domain (fwd)XynXDoc/LIC-r GAGGAGAAGCCCGGTTTAGCCATTGATTCTGAAGGAG XynX dockerin domain (rev)EndBDoc/LIC-f GACGACGACAAGATGTCTGTTGCCACGACTACTTCT EndB dockerin domain (fwd)EndBDoc/LIC-r GAGGAGAAGCCCGGTTTATTCGGGAAGCTTGTCTATG EndB dockerin domain (rev)XynBDoc/LIC-f GACGACGACAAGATGGCCACCACTACAACTACAACG XynB dockerin domain (fwd)XynBDoc/LIC-r GAGGAGAAGCCCGGTTTATAAGCTTGCACCTTTACCGAT XynB dockerin domain (rev)
a Residues that are underlined were added to the 5� ends to generate cloning sites for ligation-independent cloning. Residues in boldface type were added in additionto the ligation-independent cloning sequence as a stop codon for the cloning of internal domains.
b Abbreviations: fwd, forward; rev, reverse.
VOL. 186, 2004 NOVEL CELLULOSOMAL ADAPTOR IN R. FLAVEFACIENS 2577
on January 19, 2015 by guesthttp://jb.asm
.org/D
ownloaded from
2578 RINCON ET AL. J. BACTERIOL.
on January 19, 2015 by guesthttp://jb.asm
.org/D
ownloaded from

carried out in the same way as the Western blotting analysis on a nylon mem-brane spotted with 1 �l of His6-tagged recombinant proteins that were previouslypurified (14, 28, 29) and probed either with an HRP-conjugated S-protein (No-vagen) or biotinylated His6-tagged ScaC-Coh.
Sample preparation for two-dimensional (2D) gel electrophoresis. A total of300 �g of protein extracts was precipitated by using Perfect-FOCUS (GenoTechnology, St. Louis, Mo.) as recommended by the manufacturer. Each proteinsample was suspended in a solution containing 9 M urea, 4% 3-[(3-cholamido-propyl)-dimethylammonio]-1-propanesulfonate (CHAPS), 0.5% BioLyte (pH 3to 10; Bio-Rad), and 3% DTT and was homogenized gently with a plastic pestle.The sample was sonicated twice for 2 min with cooling for 5 min between thecycles and was allowed to sit for 20 min to facilitate protein solubilization.Finally, the sample was cleared of debris by centrifugation at 14,000 � g before340 �l (total volume) was loaded into the sample tray. Immobilized pH gradient(IPG) strips were carefully placed face down on the sample so that there were notrapped air bubbles beneath the strip.
Isoelectric focusing. Bio-Rad IPG strips (pH 3 to 6) were used for separationof proteins in the first dimension. Rehydration was performed at 20°C for 1 hwithout an applied voltage. After 1 h, each lane was overlaid with mineral oil toprevent the strips from drying out. The rehydration procedure was switched tothe active mode for a further 16 h (50 V/strip). A wick wetted with 10 �l of H2Oand another wick wetted with 10 �l of 3.5% DTT were placed at the anode andcathode, respectively, in a fresh focusing tray before an IPG strip was transferredfrom the rehydration tray. The strip was overlaid with mineral oil, and the initialstart-up and ramping protocol used was the protocol described in the Bio-Radmanual; after this the strips were subjected to a total of 60 kV � h. After 1 h offocusing the strips were moved to a tray containing fresh wicks and overlaid withmineral oil, and the focusing was continued.
Reduction and alkylation of IPG strips. After focusing, the strips were re-moved from the focusing tray and were placed face up and equilibrated in asolution containing 6 M urea, 2% SDS, 0.375 M Tris-HCl (pH 8.8), 20% glycerol,and 130 mM DTT for 10 min, after which each strip was drained and placed intoa solution containing 0.375 M Tris-HCl (pH 8.8), 20% glycerol, and 135 mMiodoacetamide for 10 min.
Second dimension (SDS-PAGE). The second-dimension procedure was per-formed by using an IsoDalt tank apparatus (Hoefer Scientific) essentially asdescribed by Laemmli (20), with the following modifications. The strips wererinsed in Laemmli electrode buffer, drained, and placed on top of an 8 to 16%SDS–polyacrylamide gradient gel (18 by 18 cm), and each strip was sealed inposition with 2% agarose in Laemmli electrode buffer. Standard Mr proteins(Bio-Rad Protein Plus standards) were loaded onto paper wicks (5 by 5 mm),dried, and loaded on top of the gel alongside the IPG strips, and each standardwick was also sealed in position with agarose. The gels (up to 10 gels per run)were electrophoresed at 200 V for 9.5 h before removal from the cassettes andstaining.
Colloidal Coomassie blue staining. Fixation and staining steps were per-formed in 200-ml solutions in Nunclon plastic dishes (23 by 23 cm; FisherScientific). The gels were fixed in 50% ethanol–2% o-phosphoric acid for at least3 h, after which they were rigorously washed in distilled H2O for at least 1 h. Thegels were placed in 34% methanol–2% o-phosphoric acid containing 17%(NH4)2SO4 and 0.05% Coomassie blue G (Sigma) for 3 days, after which theywere rinsed thoroughly in distilled H2O to remove adherent particulate Coo-massie blue before imaging. The gels were imaged with a Bio-Rad GS-710scanner and were analyzed by using the Bio-Rad PD Quest image analysissoftware.
Spot cutting and acquisition of MALDI spectra. Spots of interest were excisedfrom a gel manually, placed in a 96-well V-bottom plate, and trypsinized by usinga MassPrep station. Tryptic peptides were spotted onto a matrix-assisted laserdesorption ionization (MALDI) target plate (Teflon 2 � 96 plate; AppliedBiosystems), mixed with recrystallized �-cyano-4-hydroxycinnamic acid in aceto-
nitrile-water-trifluoroacetic acid (70:30:0.1), and dried before acquisition of spec-tra with an Applied Biosystems Voyager DE MALDI-time of flight (TOF)instrument; the machine was calibrated with a peptide standard for capillaryelectrophoresis (Sigma).
Phylogenetic analysis. Phylogenetic trees and a sequencing alignment weregenerated by using the ClustalW program (http://www2.ebi.ac.uk/clustalw/).Dockerin sequences were obtained from the GenBank website (http://www.ncbi.nlm.nih.gov/) or the Carbohydrate-Active Enzymes server (CAZy website;http://afmb.cnrs-mrs.fr/�pedro/CAZY/db.html) designed by Coutinho and Hen-rissat (7, 10). Sources of the cohesins and enzyme-borne dockerin sequencesused in this work have been described previously (28, 33, 34).
Miscellaneous. Protein concentrations were determined by the Bradfordmethod. Biotinylation of the His6-tagged ScaC-Coh recombinant protein for thespot test was carried out as described previously (28)
Nucleotide sequence accession numbers. The DNA sequence of scaC and apartial sequence of xynX have been deposited in the EMBL nucleotide sequencedatabase under accession numbers AJ585075 and AJ586790, respectively.
RESULTS
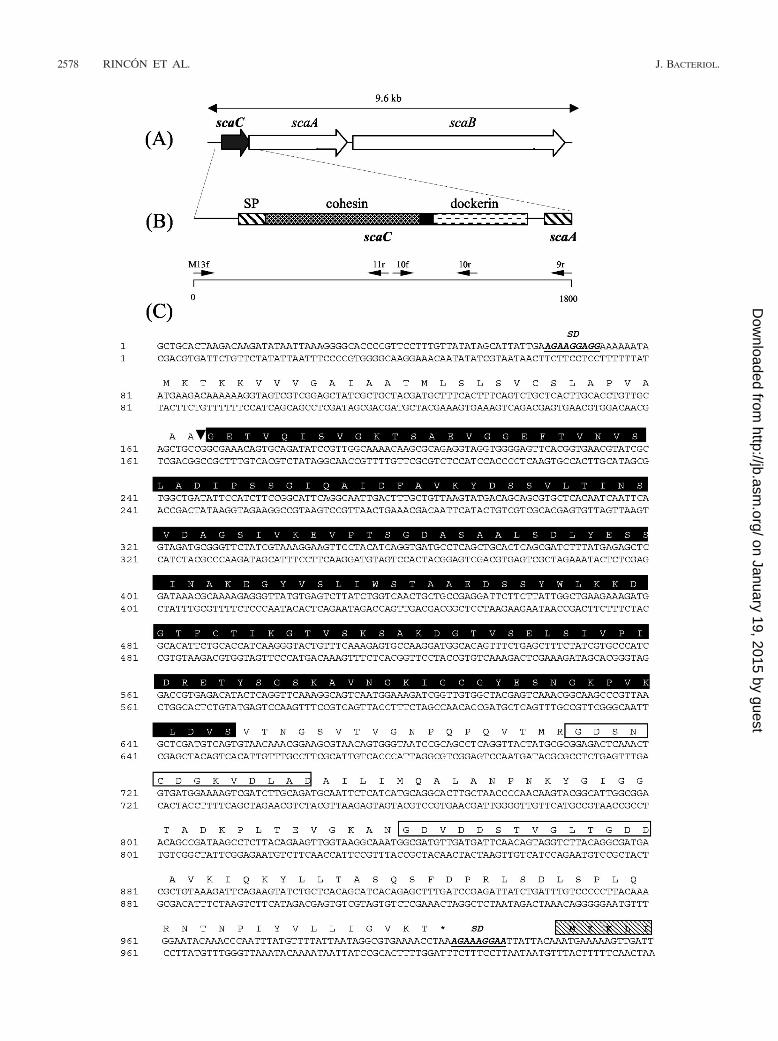
Sequencing and domain structure of scaC. scaC was discov-ered by genome walking upstream of the previously reportedscaA and scaB genes (Fig. 1A) (14, 28). The scaC start codon(ATG) is preceded by a Shine-Delgarno sequence (AGAAAGGAGG) (Fig. 1C). The intergenic spacer region betweenscaC and scaA is only 18 bp long and lacks any obvious rho-independent hairpin-like secondary structure, suggesting thatthe two genes might be cotranscribed.
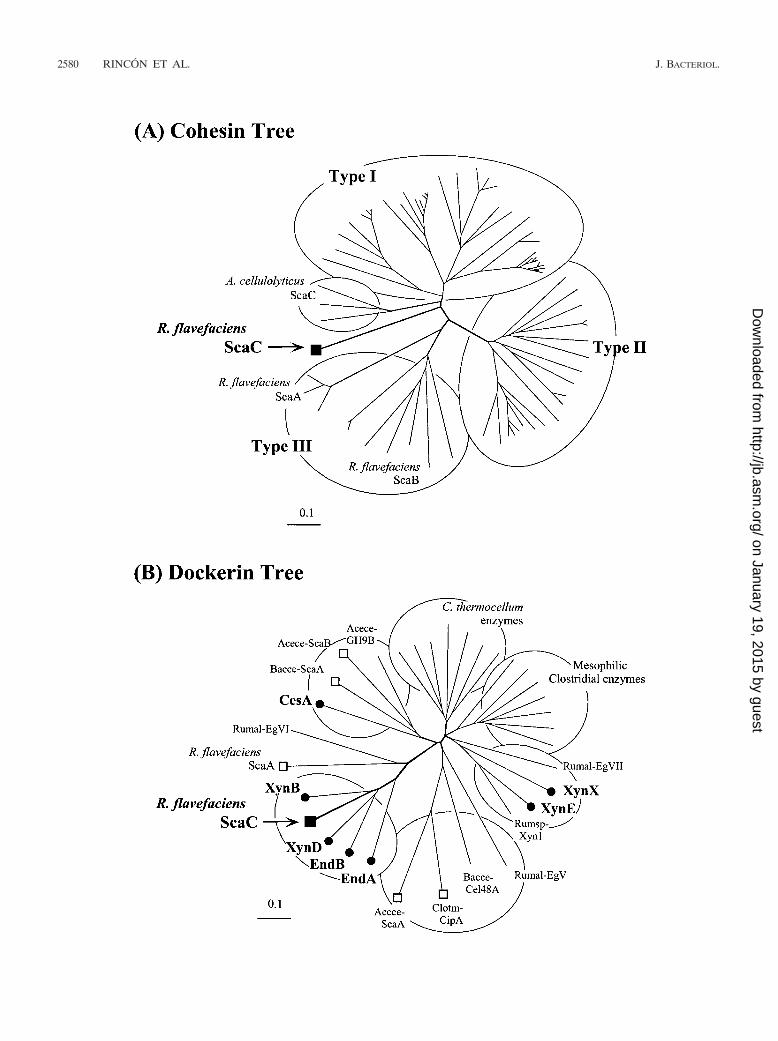
scaC encodes a 308-amino-acid protein with an estimatedmolecular mass of 31.8 kDa (Fig. 1C). Sequencing analysis ofthe gene product of scaC with the program SignalP (http://www.cbs.dtu.dk/services/SignalP/) resulted in prediction of atypical prokaryote gram-positive signal-sorting peptide se-quence consisting of 29 amino acids with the estimated cleav-age site at AA-GE. A 160-amino-acid region follows the sort-ing signal, and sequence analysis with the SIB BLAST networkservice (http://ca.expasy.org/tools/blast/) revealed significanthomology to cohesin domains from scaffolding and anchoringproteins previously identified in C. thermocellum (CipA, CipB,AncA), Clostridium cellulovorans (CbpA), Clostridium acetobu-tylicum, Clostridium cellulolyticum (CipC, CipCCA), Clostrid-ium josui (CipA), B. cellulosolvens (CipBC), A. cellulolyticus(CipV), and R. flavefaciens (ScaA, ScaB). Furthermore, asearch analysis of the Pfam database (http://www.sanger.ac.uk/cgi-bin/Pfam/getacc?PF00963) also confirmed that the N-ter-minal 160-residue region of ScaC is probably a cohesin do-main. Phylogenetic analysis (Fig. 2A) and multiple-sequencealignment (Fig. 3) revealed that the ScaC sequence is novel.Like cohesins from A. cellulolyticus ScaC (34), the cohesinfrom R. flavefaciens ScaC does not fall into any of the threecurrently recognized groups of bacterial cohesins (Fig. 2A).
At the C terminus of ScaC there are two recognizable 22- to
FIG. 1. Diagrams showing the disposition on the genome, the domain organization, and the sequence of the R. flavefaciens scaC gene. (A) Geneorganization of the structural scaffoldin proteins in R. flavefaciens 17. (B) Sequence strategy and domain architecture. scaC was discovered to beupstream of scaA by PCR walking extension with primers ScaAcoh9r (9r) and M13f (see Table 1 and Materials and Methods for details) from apUC18 R. flavefaciens 17 plasmid library. The sequence was completed by using the additional internal primers ScaAcoh11r (11r), ScaAcoh10f(10f), and ScaAcoh10r (10r). Domains within ScaC are indicated. The solid box indicates a linker sequence, and the striped boxes indicateN-terminal signal peptides (SP). The positions of the ScaC cohesin and dockerin are indicated. (C) Nucleotide sequence and sequence of the geneproduct (top) of scaC. Shine-Dalgarno sequences (SD) and the predicted cleavage site that removes the signal peptide (�) are indicated. TheF-hand repeat motifs typical of the Ca2�-binding loop of dockerin domains are indicated by open boxes. The putative cohesin domain is indicatedby a solid box. The initial sequence coding for ScaA is indicated by a striped box.
VOL. 186, 2004 NOVEL CELLULOSOMAL ADAPTOR IN R. FLAVEFACIENS 2579
on January 19, 2015 by guesthttp://jb.asm
.org/D
ownloaded from
2580 RINCON ET AL. J. BACTERIOL.
on January 19, 2015 by guesthttp://jb.asm
.org/D
ownloaded from
24-residue F-hand variations of the EF-hand motif that arecharacteristic of loops involved in the binding of calcium (Fig.1C), which is typical of dockerin modules (9, 27). Phylogeneticanalysis (Fig. 2B) placed the ScaC sequence on a branch of thedockerin tree together with the EndB-like dockerins of R.
flavefaciens 17 glycoside hydrolases EndA, EndB, XynB, andXynD, which have previously been shown to interact with theScaA type III cohesins (14, 28). Moreover, the predicted rec-ognition residues (23, 24, 27) of the ScaC dockerin are remark-ably similar to those of the EndB-like enzymes. On this basis
FIG. 2. Relationship of R. flavefaciens ScaC-borne cohesin and dockerin domains to previously described cellulosomal domains. (A) Phylo-genetic analysis of the ScaC cohesin relative to the known type I, II, and III cohesins. The ScaC cohesin maps on a separate branch, distinct fromall other cohesins. The ScaC cohesin emanates from the central branch close to the branching point of the three A. cellulolyticus ScaC cohesinsand approximately equidistant from the type I cohesins and the point of deflection that separates the type II cohesins from the type III cohesins.(B) Phylogenetic analysis of the ScaC dockerin relative to other enzyme and scaffoldin-borne dockerins. The ScaC dockerin (solid square) mapsamong the EndB-like dockerins from R. flavefaciens enzymes (solid circles) but distinct from the dockerins of CesA, XynE, and XynX. The otherknown R. flavefaciens scaffoldin-borne dockerin (ScaA) and the dockerins of C. thermocellum (Clotm), A. cellulolyticus (Acece), and B. cellulo-solvens (Bacce) are indicated by open squares. Other dockerin-borne enzymes include enzymes from Ruminococcus albus (Rumal-EgV, Rumal-EgVI, Rumal-EgVII) and Ruminococcus sp. (Rumsp-Xyn1) and a selection of enzymes from C. thermocellum and mesophilic clostridia (C.cellulolyticum, C. cellulovorans, and C. josui). For a list of the proteins and their accession numbers, see references 28, 33, and 34. Scale bars 0.1% amino acid substitutions.
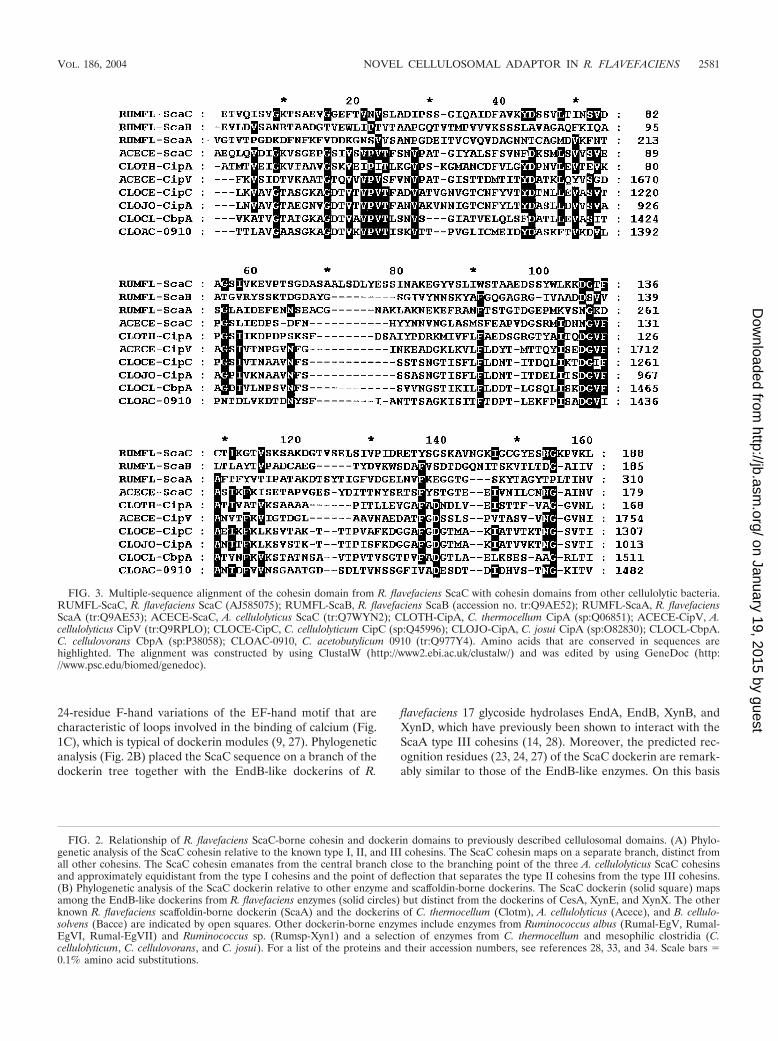
FIG. 3. Multiple-sequence alignment of the cohesin domain from R. flavefaciens ScaC with cohesin domains from other cellulolytic bacteria.RUMFL-ScaC, R. flavefaciens ScaC (AJ585075); RUMFL-ScaB, R. flavefaciens ScaB (accession no. tr:Q9AE52); RUMFL-ScaA, R. flavefaciensScaA (tr:Q9AE53); ACECE-ScaC, A. cellulolyticus ScaC (tr:Q7WYN2); CLOTH-CipA, C. thermocellum CipA (sp:Q06851); ACECE-CipV, A.cellulolyticus CipV (tr:Q9RPLO); CLOCE-CipC, C. cellulolyticum CipC (sp:Q45996); CLOJO-CipA, C. josui CipA (sp:O82830); CLOCL-CbpA.C. cellulovorans CbpA (sp:P38058); CLOAC-0910, C. acetobutylicum 0910 (tr:Q977Y4). Amino acids that are conserved in sequences arehighlighted. The alignment was constructed by using ClustalW (http://www2.ebi.ac.uk/clustalw/) and was edited by using GeneDoc (http://www.psc.edu/biomed/genedoc).
VOL. 186, 2004 NOVEL CELLULOSOMAL ADAPTOR IN R. FLAVEFACIENS 2581
on January 19, 2015 by guesthttp://jb.asm
.org/D
ownloaded from

we predicted that ScaC dockerin should interact specificallywith cohesin domains from ScaA.
Cloning and Western blot analysis of recombinant proteinfrom ScaC. The mature peptide of ScaC (full-length sequenceexcluding the putative signal peptide), as well as the cohesindomain and the dockerin module, were all cloned as doubleHis6-S-tagged proteins. Recombinant ScaC-matl and ScaC-Coh cohesin were overexpressed at 37°C after IPTG induction.The protein yield obtained from the soluble fraction afterNi-nitrilotriacetic acid affinity purification (Fig. 4) revealedthat both recombinant proteins are highly soluble and fairlystable under the expression conditions used. In contrast, theScaC dockerin was overexpressed at 16°C. The latter condi-tions were chosen because of the low yields obtained at 37°C,apparently due to the formation of inclusion bodies when theprotein was expressed at 37°C.
The purified His6-S-tagged ScaC-Coh recombinant proteinwas used as a probe for a cell surface-associated protein frac-tion of R. flavefaciens 17 grown on birchwood xylan or crystal-line cellulose (Avicel) following one-dimensional SDS-PAGEseparation and Western blotting. This approach revealed pos-itive interactions with several proteins whose sizes ranged from45 to 90 kDa (Fig. 5). For comparison, the same protein frac-tion was probed with His6-tagged ScaA-Coh. ScaA-Coh recog-nized a different set of proteins in xylan-grown cells than incellulose-grown cells, but in both cases the proteins differedfrom the proteins recognized by ScaC-Coh. On average, thepolypeptides recognized by ScaA-Coh were larger than those
recognized by ScaC-Coh (Fig. 5). To further ascertain thedifference in protein recognition between cohesins from ScaCand ScaA, a similar Western blot analysis was carried out withthe cell surface-associated protein fraction of R. flavefaciens 17grown on birchwood xylan after separation by 2D SDS-PAGE(Fig. 6A). One protein preparation was divided into threealiquots, and each aliquot was resolved by 2D gel electrophore-sis. One gel was subjected to Coomassie blue staining in orderto reveal the total number of spots. Proteins from the twoother gels were transferred to a blotting membrane; the pro-teins from one of these gels were subsequently probed withScaA-Coh, and the proteins from the other gel were probedwith ScaC-Coh. The results revealed �20 positive spots thatinteracted with ScaA-Coh with a pI range of 4 to 5 and amolecular weight range consistent with the one-dimensionalWestern blot analysis (Fig. 5). ScaC cohesin, on the otherhand, was able to recognize 11 different spots with a muchwider pI range and an average molecular mass of 37 to 90 kDa(Fig. 6C), as predicted from the one-dimensional Western blotanalysis (Fig. 5).
Among the R. flavefaciens native proteins recognized byHis6-tagged ScaA-Coh was a 29-kDa protein, which is consis-tent with the estimated molecular mass of ScaC. Although notshown in Fig. 5, this protein was clearly one of the prominentspots in the 2D Western blot analysis (Fig. 6B). A proteinprofile analysis of the 29-kDa protein spot recovered from 2Dgel separation (Fig. 6A) was carried out by trypsinization fol-lowed by MALDI-TOF analysis, and the resulting peptide fin-gerprint was compared with the theoretical trypsinized patternfor ScaC generated by using the computer-assisted program
FIG. 4. SDS-PAGE of recombinant protein probes. His6-taggedproteins were overexpressed in E. coli (see Materials and Methods fordetails). The recombinant product was purified by nickel affinity chro-matography and was separated by SDS-PAGE. Proteins were stainedwith Coomassie blue. Lane M contained a protein molecular sizemarker. The predicted molecular masses of the protein constructs are16.7, 23.2, and 33.9 kDa for His6-ScaC-Doc, His6-ScaC-Coh, and His6-ScaC-mat, respectively. The faint bands above the protein constructsappear to be products of dimerization. The numbers on the left indi-cate the positions of the protein molecular mass markers (in kilodal-tons).
FIG. 5. One-dimensional Western blot analysis of recombinant co-hesin domains from ScaC and ScaA. Cell surface-associated proteinsfrom R. flavefaciens 17 grown on birchwood xylan (lanes X) or crys-talline cellulose (Avicel) (lanes C) were separated by SDS-PAGE.Proteins were blotted onto a PVDF membrane and incubated afterblocking with recombinant His6-ScaC-Coh or His6-ScaA-Coh. Cohe-sin-dockerin interactions were exposed by incubating the membraneswith Ni-conjugated peroxidase to detect the polyhistidine tag of therecombinant proteins following enhanced chemiluminescence. Thenumbers on the left indicate the positions of the protein molecularmass markers (in kilodaltons).
2582 RINCON ET AL. J. BACTERIOL.
on January 19, 2015 by guesthttp://jb.asm
.org/D
ownloaded from
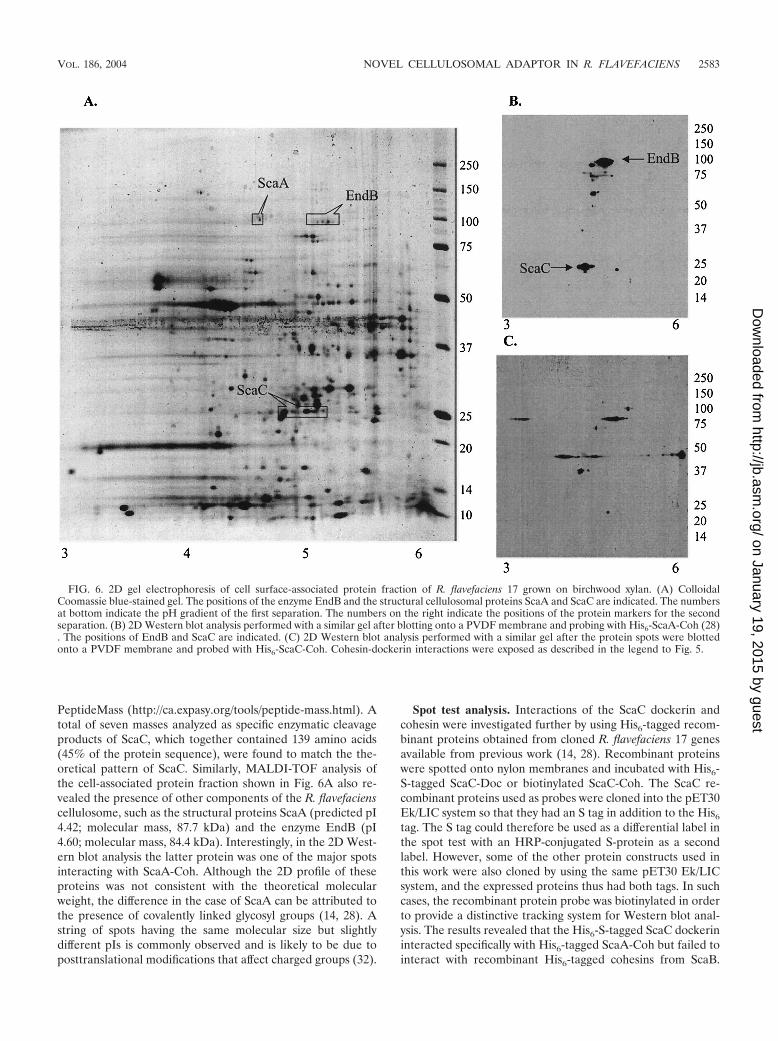
PeptideMass (http://ca.expasy.org/tools/peptide-mass.html). Atotal of seven masses analyzed as specific enzymatic cleavageproducts of ScaC, which together contained 139 amino acids(45% of the protein sequence), were found to match the the-oretical pattern of ScaC. Similarly, MALDI-TOF analysis ofthe cell-associated protein fraction shown in Fig. 6A also re-vealed the presence of other components of the R. flavefacienscellulosome, such as the structural proteins ScaA (predicted pI4.42; molecular mass, 87.7 kDa) and the enzyme EndB (pI4.60; molecular mass, 84.4 kDa). Interestingly, in the 2D West-ern blot analysis the latter protein was one of the major spotsinteracting with ScaA-Coh. Although the 2D profile of theseproteins was not consistent with the theoretical molecularweight, the difference in the case of ScaA can be attributed tothe presence of covalently linked glycosyl groups (14, 28). Astring of spots having the same molecular size but slightlydifferent pIs is commonly observed and is likely to be due toposttranslational modifications that affect charged groups (32).
Spot test analysis. Interactions of the ScaC dockerin andcohesin were investigated further by using His6-tagged recom-binant proteins obtained from cloned R. flavefaciens 17 genesavailable from previous work (14, 28). Recombinant proteinswere spotted onto nylon membranes and incubated with His6-S-tagged ScaC-Doc or biotinylated ScaC-Coh. The ScaC re-combinant proteins used as probes were cloned into the pET30Ek/LIC system so that they had an S tag in addition to the His6
tag. The S tag could therefore be used as a differential label inthe spot test with an HRP-conjugated S-protein as a secondlabel. However, some of the other protein constructs used inthis work were also cloned by using the same pET30 Ek/LICsystem, and the expressed proteins thus had both tags. In suchcases, the recombinant protein probe was biotinylated in orderto provide a distinctive tracking system for Western blot anal-ysis. The results revealed that the His6-S-tagged ScaC dockerininteracted specifically with His6-tagged ScaA-Coh but failed tointeract with recombinant His6-tagged cohesins from ScaB.
FIG. 6. 2D gel electrophoresis of cell surface-associated protein fraction of R. flavefaciens 17 grown on birchwood xylan. (A) ColloidalCoomassie blue-stained gel. The positions of the enzyme EndB and the structural cellulosomal proteins ScaA and ScaC are indicated. The numbersat bottom indicate the pH gradient of the first separation. The numbers on the right indicate the positions of the protein markers for the secondseparation. (B) 2D Western blot analysis performed with a similar gel after blotting onto a PVDF membrane and probing with His6-ScaA-Coh (28). The positions of EndB and ScaC are indicated. (C) 2D Western blot analysis performed with a similar gel after the protein spots were blottedonto a PVDF membrane and probed with His6-ScaC-Coh. Cohesin-dockerin interactions were exposed as described in the legend to Fig. 5.
VOL. 186, 2004 NOVEL CELLULOSOMAL ADAPTOR IN R. FLAVEFACIENS 2583
on January 19, 2015 by guesthttp://jb.asm
.org/D
ownloaded from

Furthermore, there was no detectable interaction between bi-otinylated ScaC-Coh and either of the two previously charac-terized types of dockerins derived from known R. flavefaciensproteins, including EndB, XynB, XynX, XynE, CesA, andScaA. (Fig. 7)
DISCUSSION
The results of the present study established that a newlyidentified protein, ScaC, interacts with the primary ScaA scaf-foldin via its C-terminal dockerin module. ScaC is thus the firstnoncatalytic protein in R. flavefaciens that has been shown topair with ScaA cohesins, which otherwise recognize dockerinsborne by the enzymes EndA, EndB, XynD, and XynB. Theunique cohesin domain of ScaC appears to mediate interac-tions with a range of native R. flavefaciens 17 proteins thatexhibit no overlap with the subset of proteins that is recognizedby the ScaA cohesins. This conclusion is supported by thefailure of the ScaC cohesin to recognize representative recom-binant dockerins from the enzymes EndB and XynB or fromthe scaffolding protein ScaA. It appears, therefore, that ScaCacts as a type of adaptor, allowing many proteins that are notrecognized directly by the ScaA cohesins to be bridged into thecellulosome assembly.
We assume that the subset of proteins that is recognized bythe ScaC cohesin must have a distinct type of dockerin domainthat has yet to be identified. In previous work it has been foundthat the dockerins present in the enzymes CesA and XynE,whose sequences differ from the sequences of the dockerins inEndA, EndB, XynB, and XynD (1), failed to recognize ScaAcohesins (28). However, we found no evidence in this studythat these dockerins were recognized by ScaC. This argues thatthere should be at least the following four distinct cohesin-dockerin binding specificities in R. flavefaciens 17: (i) betweenthe ScaB cohesins and ScaA-Doc; (ii) between the ScaA co-hesins and the EndB-type enzyme dockerins; (iii) betweenCesA-XynE dockerins and an unknown cohesin(s); and (iv)between ScaC-Coh and an unknown type of dockerin.
The role proposed here for ScaC further emphasizes thedifferences in cellulosome organization between R. flavefaciensand cellulolytic Clostridium spp. In the cellulosome model de-scribed for C. thermocellum the catalytic subunits are organizedalong the primary scaffoldin protein, CipA. CipA in turn in-teracts with at least three other anchoring scaffoldins (SdbA,OlpA, and OlpB) (4, 30) through its dockerin domain. More-over, CipA possesses a family 3a CBM with high affinity towardcellulose fibrils and is thought to mediate the specific interac-tions of the bacterium with its substrate. A similar overallarrangement, with distinctive variations on the theme, exists inthe cellulosome systems of A. cellulolyticus and B. cellulosolvens(12, 13, 33, 34). In contrast, in R. flavefaciens the catalyticsubunits are organized in a different way. The dockerin of theprimary scaffoldin ScaA interacts with any of the seven cohesinrepeats in the putative anchoring scaffoldin ScaB, and certain(EndB-like) catalytic subunits are then able to interact with thethree cohesin repeats present in ScaA. ScaC, however, intro-duces another dimension into the interaction with ScaA,whereby an additional subset of proteins, which we assumepossess distinctive dockerin modules, are recruited into thecellulosome. The single ScaC cohesin could thus have a trueadaptor function, which is distinct from the proposed adaptorfunction described for A. cellulolyticus cellulosomal ScaB (34).In the latter case, the major function of the ScaB scaffoldinseems to be to amplify the number of enzymes that can beincorporated into the A. cellulolyticus ScaC anchoring scaffol-din (34).
The functional significance of the R. flavefaciens ScaC adap-tor mechanism has yet to be fully established. If production ofthe ScaC protein were subject to regulation independent ofScaA, then it might provide an effective mechanism for regu-lating the subunit composition of the cellulosome. We can alsospeculate that the additional cohesin specificity provided byScaC could allow the incorporation of plant cell wall-degradingenzymes acquired by horizontal gene transfer from phyloge-netically distant organisms into the R. flavefaciens cellulosome.
One interesting observation from this work is that ScaC-Cohseems to interact with proteins whose molecular sizes aresmaller than those of the range of proteins recognized byScaA-Coh. The enzymes so far known to interact with ScaAcohesins all have complex multimodular structures, but itseems likely, judging by their small molecular sizes, that pro-teins interacting with ScaC have a simpler modular organiza-tion. This might reflect their evolutionary origin (e.g., acquisi-tion from different source organisms via horizontal transfer) or
FIG. 7. Spot test analysis of ScaC-mediated cohesin-dockerin in-teractions. (A and B) Recombinant His6-tagged proteins were spottedonto a nylon membrane and revealed by interaction with Ni-conju-gated peroxidase. (C) The membrane was incubated with the biotin-ylated ScaC-mat construct, and cohesin-dockerin interactions revealedby a second incubation with peroxidase-conjugated streptavidin.(D) The membrane was incubated with the ScaC-Doc construct, andcohesin-dockerin interactions were revealed by a second incubationwith HRP-conjugated S protein, which recognized the S tag presentonly in the recombinant ScaC-Doc construct. Spots 1 to 6, dockerindomains from EndB, XynB, CesA, XynE, XynX, and ScaA, respec-tively; spots 7 and 8, X domains from ScaA and ScaB, respectively;spots 9 to 14, cohesins 1, 2, 3, 4, 6, and 7 from ScaB, respectively; spot15, ScaA-Coh2.
2584 RINCON ET AL. J. BACTERIOL.
on January 19, 2015 by guesthttp://jb.asm
.org/D
ownloaded from

might indicate entirely different roles (e.g., nonenzymatic) onthe cell surface. The functions of the proteins that bind ScaC,including the question of whether they are in fact enzymes orare structural components, have yet to be established.
The combination of Western blot analysis and 2D gel elec-trophoresis proved to be a valuable tool for the study of pro-tein interactions in the cellulosome organization of R. flavefa-ciens 17 cell wall-degrading enzymes. The main priority forfuture research on this system is to use proteomic analysis toidentify proteins separated by 2D gel electrophoresis that canbe pinpointed as structural components of the cellulosome byprotein affinity analysis.
ACKNOWLEDGMENTS
R.R.I. is supported by the Scottish Executive Environment andRural Affairs Department. M.T.R. is employed under the EU frame-work V program GEMINI. This research was supported by the IsraelScience Foundation (grants 394/03 and 771/01), by the U.S.-IsraelBinational Agricultural Research and Development Fund (BARD re-search grant 3106-99C), and by a grant from the United States-IsraelBinational Science Foundation, Jerusalem, Israel.
REFERENCES
1. Aurilia, V., J. C. Martin, S. I. McCrae, K. P. Scott, M. T. Rincon, and H. J.Flint. 2000. Three multidomain esterases from the cellulolytic rumen anaer-obe Ruminococcus flavefaciens 17 that carry divergent dockerin sequences.Microbiology 146:1391–1397.
2. Bayer, E. A., H. Chanzy, R. Lamed, and Y. Shoham. 1998. Cellulose, cellu-lases and cellulosomes. Curr. Opin. Struct. Biol. 8:548–557.
3. Bayer, E. A., and R. Lamed. 1986. Ultrastructure of the cell surface cellulo-some of Clostridium thermocellum and its interaction with cellulose. J. Bac-teriol. 167:828–836.
4. Bayer, E. A., L. J. Shimon, Y. Shoham, and R. Lamed. 1998. Cellulosomes—structure and ultrastructure. J. Struct. Biol. 124:221–234.
5. Beguin, P., and M. Lemaire. 1996. The cellulosome: an exocellular, multi-protein complex specialized in cellulose degradation. Crit Rev. Biochem.Mol. Biol. 31:201–236.
6. Belaich, J. P., C. Tardif, A. Belaich, and C. Gaudin. 1997. The cellulolyticsystem of Clostridium cellulolyticum. J. Biotechnol. 57:3–14.
7. Bourne, Y., and B. Henrissat. 2001. Glycoside hydrolases and glycosyltrans-ferases: families and functional modules. Curr. Opin. Struct. Biol. 11:593–600.
8. Bryant, M. P. 1973. Nutritional requirements of the predominant rumencellulolytic bacteria. Fed. Proc. 32:1809–1813.
9. Chauvaux, S., P. Beguin, J. P. Aubert, K. M. Bhat, L. A. Gow, T. M. Wood,and A. Bairoch. 1990. Calcium-binding affinity and calcium-enhanced activityof Clostridium thermocellum endoglucanase D. Biochem. J. 265:261–265.
10. Coutinho, P. M., and B. Henrissat. 1999. Carbohydrate-active enzymes: anintegrated database approach, p. 3–12. In H. J. Gilbert, G. J. Davies, B.Henrissat, and B. Svensson (ed.), Recent advances in carbohydrate bioengi-neering. The Royal Society of Chemistry, Cambridge, United Kingdom.
11. Devillard, E., D. B. Goodheart, S. K. Karnati, E. A. Bayer, R. Lamed, J.Miron, K. E. Nelson, and M. Morrison. 2004. Ruminococcus albus 8 mutantsdefective in cellulose degradation are deficient in two processive endocellu-lases, Cel48A and Cel9B, both of which possess a novel modular architec-ture. J. Bacteriol. 186:136–145.
12. Ding, S. Y., E. A. Bayer, D. Steiner, Y. Shoham, and R. Lamed. 1999. A novelcellulosomal scaffoldin from Acetivibrio cellulolyticus that contains a family 9glycosyl hydrolase. J. Bacteriol. 181:6720–6729.
13. Ding, S. Y., E. A. Bayer, D. Steiner, Y. Shoham, and R. Lamed. 2000. Ascaffoldin of the Bacteroides cellulosolvens cellulosome that contains 11 typeII cohesins. J. Bacteriol. 182:4915–4925.
14. Ding, S. Y., M. T. Rincon, R. Lamed, J. C. Martin, S. I. McCrae, V. Aurilia,Y. Shoham, E. A. Bayer, and H. J. Flint. 2001. Cellulosomal scaffoldin-likeproteins from Ruminococcus flavefaciens. J. Bacteriol. 183:1945–1953.
15. Doi, R. H., A. Kosugi, K. Murashima, Y. Tamaru, and S. O. Han. 2003.Cellulosomes from mesophilic bacteria. J. Bacteriol. 185:5907–5914.
16. Fujino, T., P. Beguin, and J. P. Aubert. 1993. Organization of a Clostridiumthermocellum gene cluster encoding the cellulosomal scaffolding proteinCipA and a protein possibly involved in attachment of the cellulosome to thecell surface. J. Bacteriol. 175:1891–1899.
17. Hungate, R. E., and R. J. Stack. 1982. Phenylpropanoic acid: growth factorfor Ruminococcus albus. Appl. Environ. Microbiol. 44:79–83.
18. Kosugi, A., K. Murashima, Y. Tamaru, and R. H. Doi. 2002. Cell-surface-anchoring role of N-terminal surface layer homology domains of Clostridiumcellulovorans EngE. J. Bacteriol. 184:884–888.
19. Krause, D. O., B. P. Dalrymple, W. J. Smith, R. I. Mackie, and C. S.McSweeney. 1999. 16S rDNA sequencing of Ruminococcus albus and Rumi-nococcus flavefaciens: design of a signature probe and its application in adultsheep. Microbiology 145:1797–1807.
20. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 227:680–685.
21. Leibovitz, E., H. Ohayon, P. Gounon, and P. Beguin. 1997. Characterizationand subcellular localization of the Clostridium thermocellum scaffoldin dock-erin binding protein SdbA. J. Bacteriol. 179:2519–2523.
22. Lemaire, M., H. Ohayon, P. Gounon, T. Fujino, and P. Beguin. 1995. OlpB,a new outer layer protein of Clostridium thermocellum, and binding of itsS-layer-like domains to components of the cell envelope. J. Bacteriol. 177:2451–2459.
23. Mechaly, A., H. P. Fierobe, A. Belaich, J. P. Belaich, R. Lamed, Y. Shoham,and E. A. Bayer. 2001. Cohesin-dockerin interaction in cellulosome assem-bly: a single hydroxyl group of a dockerin domain distinguishes betweennon-recognition and high affinity recognition. J. Biol. Chem. 276:9883–9888.
24. Mechaly, A., S. Yaron, R. Lamed, H. P. Fierobe, A. Belaich, J. P. Belaich, Y.Shoham, and E. A. Bayer. 2000. Cohesin-dockerin recognition in cellulo-some assembly: experiment versus hypothesis. Proteins 39:170–177.
25. Michalet-Doreau, B., I. Fernandez, and G. Fonty. 2002. A comparison ofenzymatic and molecular approaches to characterize the cellulolytic micro-bial ecosystems of the rumen and the cecum. J. Anim. Sci. 80:790–796.
26. Miyazaki, K., J. C. Martin, R. Marinsek-Logar, and H. J. Flint. 1997.Degradation of xylans by the rumen anaerobe Prevotella bryantii (formerlyPrevotella ruminicola subsp. brevis) B14. Anaerobe 3:373–381.
27. Pages, S., A. Belaich, J. P. Belaich, E. Morag, R. Lamed, Y. Shoham, andE. A. Bayer. 1997. Species-specificity of the cohesin-dockerin interactionbetween Clostridium thermocellum and Clostridium cellulolyticum: predictionof specificity determinants of the dockerin domain. Proteins 29:517–527.
28. Rincon, M. T., S. Y. Ding, S. I. McCrae, J. C. Martin, V. Aurilia, R. Lamed,Y. Shoham, E. A. Bayer, and H. J. Flint. 2003. Novel organization anddivergent dockerin specificities in the cellulosome system of Ruminococcusflavefaciens. J. Bacteriol. 185:703–713.
29. Rincon, M. T., S. I. McCrae, J. Kirby, K. P. Scott, and H. J. Flint. 2001.EndB, a multidomain family 44 cellulase from Ruminococcus flavefaciens 17,binds to cellulose via a novel cellulose-binding module and to another R.flavefaciens protein via a dockerin domain. Appl. Environ. Microbiol. 67:4426–4431.
30. Schwarz, W. H. 2001. The cellulosome and cellulose degradation by anaer-obic bacteria. Appl. Microbiol. Biotechnol. 56:634–649.
31. Tamaru, Y., and R. H. Doi. 1999. Three surface layer homology domains atthe N terminus of the Clostridium cellulovorans major cellulosomal subunitEngE. J. Bacteriol. 181:3270–3276.
32. Vercoe, P. E., S. A. Kocherginskaya, and B. A. White. 2003. Differentialprotein phosphorylation-dephosphorylation in response to carbon source inRuminococcus flavefaciens FD-1. J. Appl. Microbiol. 94:974–980.
33. Xu, Q., E. A. Bayer, M. Goldman, R. Kenig, Y. Shoham, and R. Lamed. 2004.Architecture of the Bacteroides cellulosolvens cellulosome: description of acell surface-anchoring scaffoldin and a family 48 cellulase. J. Bacteriol. 186:968–977.
34. Xu, Q., W. Gao, S. Y. Ding, R. Kenig, Y. Shoham, E. A. Bayer, and R. Lamed.2003. The cellulosome system of Acetivibrio cellulolyticus includes a noveltype of adaptor protein and a cell surface anchoring protein. J. Bacteriol.185:4548–4557.
VOL. 186, 2004 NOVEL CELLULOSOMAL ADAPTOR IN R. FLAVEFACIENS 2585
on January 19, 2015 by guesthttp://jb.asm
.org/D
ownloaded from











