Embed Size (px)
Citation preview

Plant Molecular Biology 9: 343- 353 (1987) © Martinus Nijhoff Publishers, Dordrecht - Printed in the Netherlands 343
Differential changes in the synthesis and steady-state levels of thylakoid proteins during bean leaf senescence
D. R. Roberts, 1 J. E. Thompson, 1 E. B. Dumbroff, 1 S. Gepstein 1. and A. K. Mattoo 2 1Department of Biology, University of Waterloo, Waterloo, Ontario, Canada N2L 3G1; 2Beltsville Agricultural Research Center, Plant Hormone Laboratory, USDA, Beltsville, MD 20705, USA; * on leave from the Department of Biology, Technion-Israel Institute of Technology, Haifa, Israel
Received 12 January 1987; accepted in revised form 15 June 1987
Key words: bean, leaf senescence, protein degradation, protein synthesis, thylakoids
Abstract
During senescence of primary bean leaves (Phaseolus vulgaris), there are differential changes in the rates at which thylakoid proteins are synthesized. In particular, synthesis of the 32 kD herbicide-binding protein con- tinues throughout senescence, whereas formation of the a and 3 subunits of ATPase, the 68 kD photosystem I reaction center polypeptide, cytochrome f, cytochrome b 6 and the structural apoprotein of the light- harvesting chlorophyll protein complex (LHCP) declines. Pulse-chase experiments with intact leaves indicated rapid degradation of the 32 kD protein, which is consistent with its known rapid rate of turnover. This degrada- tion was light-dependent and inhibited by DCMU, and the kinetics of degradation were similar for young and senescent membranes. In Coomassie-stained gels, the 68 kD reaction center polypeptide of photosystem I, the oL and 3 subunits of ATPase and the LHCP were the dominant proteins for all ages of membranes. Western blot analysis indicated that cytochrome f and cytochrome b 6 are selectively depleted during senescence. The data have been interpreted as indicating that translational disruptions in both the cytoplasmic and chloroplastic compartments may contribute to the decline in photosynthetic electron transport in the senescing leaf.
Introduction
Photosynthetic capability and net production of carbohydrate decline with advancing leaf senescence in a temporal pattern that parallels the loss of chlo- rophyll and protein. This can be attributed in part to curtailed synthesis and rapid degradation of Cal- vin cycle enzymes and a consequent decline in the dark reactions of photosynthesis. Ribulose- bisphosphate carboxylase activity in particular decreases dramatically during the early stages of leaf senescence [2].
There is also loss of photosynthetic electron trans- port capability in senescing leaves. Moreover, the de-
crease in non-cyclic electron transport is greater than the decline in the activities of photosystem I or pho- tosystem II [11], which suggests that intersystem electron transport becomes rate-limiting. The pho- tosystems are spatially separated in the thylakoids, photosystem II being localized predominantly in the appressed granal membranes and photosystem I in the non-appressed stromal membranes [1]. Inter- system electron transport is thought to be achieved by lateral diffusion of the mobile carriers, plastoqui- none and plastocyanin, through the plane of the thylakoids [1, 27]. Such diffusion would be restricted and intersystem electron transport constrained if there were a decrease in thylakoid fluidity during

344
senescence. However, unlike the plasmalemma and microsomal membranes, which sustain a large de- crease in bilayer fluidity in a variety of senescing tis- sues [5, 13, 19, 291, chloroplast membranes show no significant change in fluidity with advancing senes- cence [20]. Rather, the decrease in intersystem elec- tron transport in senescing leaves has been attributed to a decline in the functional concentration of cytochrome f /b 6 [9].
These observations collectively suggest that cer- tain thylakoid proteins are selectively inactivated or degraded as senescence progresses. On the basis of immunological studies, Ben-David et al. [3] suggest- ed that the cytochrome f /b 6 complex is depleted from the thylakoids more rapidly than photosystem I complex proteins. In the present study, we have ex- amined the turnover of thylakoid proteins in senes- cing primary leaves of bean in order to determine whether selective depletion of proteins reflects
-differential changes in thylakoid protein synthesis or enhanced degradation of specific proteins.
Materials and methods
Labeling o f plant material and membrane isolation
Bean seedlings (Phaseolus vulgqris L. cv. Kinghorn) were grown in a mixture of sand, peat and soil (1:1:2) under a 16-hour photoperiod, fluorescent illumina- tion of 269 #E s -~ m -2 and day/night temperatures of 27 and 22°C. Primary leaves were severed from 7, 14, 21 and 28-day-old seedlings at the leaf base, trimmed to approximately 18 cm 2, rinsed with dis- tilled water, blotted dry and pulse-labeled at 24 °C by floating them with the abaxial surface up on 60 ml of Hutner's nutrient solution (1/2 strength, pH 6.2) [24] containing 240 #1 Tween 20 and 1000/~Ci [35S]-methionine (1000 mCi/mmole) for 2-4 .5 hours under fluorescent lighting (63/zE s -1 m-2). At the end of this incubation period, four 18-cm z leaf segments were rinsed and homogenized in 40 ml of isolation buffer (0.3 M sorbitol, 50 mM tricine, 0.05 M MgC12 and 1% bovine serum albu- min, pH 8.0) for one 4-second period in an Om-
nimixer.* The brei was filtered through 8 layers of cheesecloth, and the filtrate was centrifuged at 2000 g for 2 min. The pellet was resuspended in wash buffer (0.3 M sorbitol, 50 mM tricine, pH 8.0) and recentrifuged at 2000 g for 2 min. The resulting crude chloroplast fraction was lysed by resuspension in 50 mM tricine (pH 8.0), and the plastid mem- branes were pelleted by centrifugation at 6000 g for 5 min and resuspended in 3 ml of wash buffer. Radi- olabeled thylakoid proteins were also isolated from Spirodela oligorrhiza as previously described [25].
In some experiments, the leaves were labeled with 35S-methionine using a pulse-chase regime. After 4.5 hours of floating on Hutner's nutrient solution containing labeled methionine, the leaves were rinsed with distilled water and floated for additional 2, 4, 8 and 24 hour periods under fluorescent lighting (63/xE s -~ m -2) on Hutner's nutrient solution con- taining 10 mM unlabeled methionine before isola- tion of chloroplast membranes.
Protein, chlorophyll and radioactivity determinations
For measurements of protein, 300 #1 of the thylak- oid suspension were precipitated over a 24-hour peri- od at 4 °C in the presence of 1.2 ml of 30% trichlo- roacetic acid and centrifuged at 16000 g for 6 min. The pellet was washed 3 times by resuspension in 80% acetone and centrifugation at 16000 g for 6 min and was then dissolved in 300/~1 of 1 M NaOH by heating to 90°C for 5 min. Protein levels were determined as described by Lowry et al. [14].
For measurements of radioactivity, 50 #1 of the thylakoid suspension were vortexed in 10/~I of Bio- Solv solubilizer (Beckman) and after 2 hours, 5 ml of ACS aqueous counting scintillant (Amersham) were added, and the samples were counted in a Searle Scintillation Counter.
Chlorophyll was measured by extracting leaf discs with dimethyl sulfoxide and measuring the absor-
* Mentionofspecificinstruments, tradenames, or manufacturers is for the purpose of identification and does not imply an endorse- ment by the United States Government.

345
bance of the extract at 645 and 663 nm as described by Hiscox and Israelstam [8].
Electrophoresis and western blotting
Thylakoids (20 #g protein) were pelleted by centrifu- gation at 16000 g for 6 min and suspended in 20/~I of solubilizing buffer (562.5 izl of 0.5 M Tris, pH 6.8, 0.09 g sodium dodecyl sulfate, 225/~1 mercap- toethanol, 225/~1 glycerol; diluted 1:1 with distilled water) by gentle vortexing for 10 rain at 24 °C. The resulting protein solution was stored at - 7 0 °C for 24 hours and then thawed prior to fractionation on gradient (10-20%) polyacrylamide gels with a 5°70 stacking gel [12]. The gels were loaded with either constant protein (10 #g/lane) or constant radioac- tivity (6800 dpm/lane). Following fixation and fluorography using Autofluor (National Diagnos- tics) as an autoradiographic image enhancer, the gels were dried and exposed to Kodak (XAR-Z) X-ray film at -70°C.
For western blotting, the proteins were transferred from the polyacrylamide gels onto nitrocellulose sheets (0.45 #m or 0.1 #m pore size) using a Bio-Rad transblot apparatus. The filters were then treated in sequence with 2°70 bovine serum albumin in phos- phate buffered saline (PBS) for 2 hours and with primary antibody (1:1000 in PBS) for 12 hours, and washed 4 times, each for 10 min, with 2% bovine se- rum albumin in PBS. They were then treated with secondary antibody (goat antirabbit IgG conjugated to peroxidase) (Bio-Rad) in PBS for 1 hour and washed once for 10 min in PBS, again for 10 min in PBS containing 0.02°70 Triton X-100 and twice more, each for 10 min, in PBS. Bound antibody was visual- ized by treatment with peroxidase substrate [23]. Polyclonal antibodies against the 32 kD herbicide- binding protein were a gift from Dr J. Hirschberg, Hebrew University, Jerusalem, Israel. These anti- bodies were prepared by overproducing a portion of the 32 kD protein from Amaranthus hybridus, con- sisting of the region from amino acid residue 52 to the carboxy terminus [7], in E. coli, purifying the polypeptide and using it to raise specific antibodies in rabbit (Hirschberg et al., in preparation [22]). For preparation of other polyclonal antibodies, the
cytochrome f/b 6 complex was isolated as described by Hurt and Hauska [10], the/3 subunit of ATPase as described by Nelson [21], the 68 kD polypeptide of photosystem I as described by Bengis and Nelson [4] and the light-harvesting chlorophyll protein com- plex and the 33 kD polypeptide of photosystem II as described by Yamamoto et al. [31]. Antibodies were raised in rabbits and tested for their specificity as previously described [21].
Results
Temporal pattern of leaf senescence
The temporal pattern of primary leaf senescence was established by measuring changes in leaf chlorophyll over time. From day 7 through day 28, chlorophyll concentrations per unit dry weight and per unit leaf area declined progressively reaching values as low as 11/zg cm -2 of leaf surface and 7 mg g-~ dry weight (Fig. 1).
4 0 4 0 0 0
35
30
> , r - 2 5 r , 0 L. 0
-~ 20 L)
O~ 1 5 /
t 0
1 I I I 7 t 4 2 t 28
Leo f Age (days ofter plonting)
$500
4-- >
~000 u
0
; '500 o n~
._u 2000 "~a.
1500 ~D
0
1ooo _o (-
~00
Fig. 1. Changes in chlorophyll and total thylakoid protein syn- thesis in senescing primary leaves of bean. ( • ), Leaf chlorophyll (mg g- l dry weight); (o) , leaf chlorophyll ~g cm-2); (n), specific radioactivity (dpm/ug protein) for thylakoids isolated from leaves that were pulse-labeled with [35S]-methionine. Stan- dard errors of the means are shown when larger than the symbol; n = 3 .

346
Steady-state levels of thylakoid proteins in senescing leaves
Thylakoid proteins were fractionated on SDS- polyacrylamide gels, and discrete proteins were iden- tified on the basis of previously reported molecular weights [3, 6] and immunologically on western blots (Fig. 2). The proteins identified include subunit I (68 kD polypeptide) of the photosystem I complex, the/3 subunit of ATPase, the 33 kD extrinsic poly- peptide of the photosystem II complex, the 25 and 26 kD apoproteins of the light-harvesting chlo- rophyll a /b complex (LHCP), cytochromes f and b 6 and the 32 kD herbicide-binding protein of pho- tosystem II (Fig. 2). Coomassie-stained polypeptide profiles of the isolated thylakoids were closely simi- lar for all ages of leaves ranging from young (day 7) through extensively senescent (day 35) (Fig. 3). In particular, steady-state levels of the major proteins, including the 68 kD reaction center polypeptide of
photosystem I, the ot and/~ subunits of ATPase and the LHCP complex, remained stable through day
21 (Fig. 3, lanes 3, 4 and 5). By day 28, a small de- crease in the steady-state levels of the ct and /~ subunits of ATPase was apparent (Fig. 3, lane 2). By day 35, the leaves were moribund and beginning to abscise, and decreased steady-state levels of the 68 kD reaction center polypeptide of photosystem I and of the LHCP complex were also evident (Fig. 3, lane 1).
The apparent stability of the major thylakoid pro- teins during the earlier stages of senescence was fur- ther examined by visualizing specific proteins on im- munoblots. Thylakoids were isolated from 7- and 21-day-old leaves and fractionated by SDS-PAGE. Of the proteins depicted immunologically, viz., the 68 kD reaction center polypeptide of photosystem I complex, the/~ subunit of ATPase, cytochromes f and b6, the 33 kD extrinsic protein of photosystem II, the 24 and 26 kD polypeptides of LHCP (Fig. 4)
Fig. 2. Immunological identification ofthylakoid proteins from 7-day-old primary leaves. (A): Lane 1, 68 kD polypeptide of photosystem I complex; Lane 2, 25 kD and 26 kD polypeptides of LHCP; Lane 3, amido black-stained blot; Lane 4,/~ subunit of ATPase; Lane 5, 33 kD extrinsic polypeptide of photosystem II; Lane 6, cytochrome f. Arrows in lane 3 denote the polypeptides detected on immunoblots in lanes 1, 2, 4, 5 and 6. (B): Lane 1, cytochrome b6; Lane 2, fluorogram of newly synthesized proteins separated by SDS-PAGE; Lane 3, 32 kD herbicide-binding protein. Arrows in lane 2 denote the polypeptides detected on immunoblots in lanes 1 and 3.

Fig. 3. Steady-state levels of thylakoid proteins in senescing primary bean leaves as visualized by Coomassie-staining of poly- acrylamide gels. Lane 1, 35-day-old leaves; lane 2, 28-day-old leaves; lane 3, 21-day-old leaves; lane 4, 14-day-old leaves; lane 5, 7-day-old leaves. Molecular weight markers (kD) are specified. Arrows from top to bottom denote proteins corresponding to the 68 kD polypeptide of photosystem I complex, the a and subunits of ATPase and the 25 and 26 kD polypeptides of LHCP, respectively.
and the 32 kD herbicide-binding protein (Fig. 5), only cytochrome f and cytochrome b 6 showed a de- crease in steady-state concentration over this period (Figs. 4 and 5). These observations corroborate a previous report that these two proteins are selectively depleted from senescing thylakoids [3]. Thylakoids from Spirodela oligorrhiza were run as a control in the western blots for the 32 kD herbicide-binding protein since this protein is well characterized in this
347
organism [18]. As well as identifying the 32 kD herbicide-binding protein, the 32 kD-specific anti- body detected a 28-29 kD protein band in thylak- oid samples from primary bean leaves that was not present in the western blot for Spirodela thylakoids (Fig. 5, compare lane 1 with lanes 2-5). This 28- 29 kD protein may be a different conformation of the 32 kD herbicide-binding protein since it mi- grates as a 32 kD protein band when excised from gels and rerun on a second SDS gel (Greenberg, B., Edelman, M., personal communication). However, the possibility that it is a degradation product of the 32 kD protein is not precluded. The 28- 29 kD pro- tein was detectable even when the protease inhibitors PMSF, TLCK, bromphenol acyl bromide or TPCK were included in the isolation medium either singly or in combination at a concentration of 1 mM (data not shown). When the leaves were boiled prior to isolating the thylakoids in an effort to prevent the potential formation of proteolytic artifacts during the isolation procedure, neither the 32 kD protein nor the additional band at 28- 29 kD were discerni- ble. This may be additional support for the sugges- tion that the 28-29 kD protein represents a differ- ent conformation of the 32 kD protein. It is conceivable that the absence of the 32 kD protein and the 28- 29 kD protein in boiled samples reflects the fact that, at least for thylakoids from bean leaf, boiling appears to induce the formation of ag- gregates that are not broken down during solubiliza- tion in SDS.
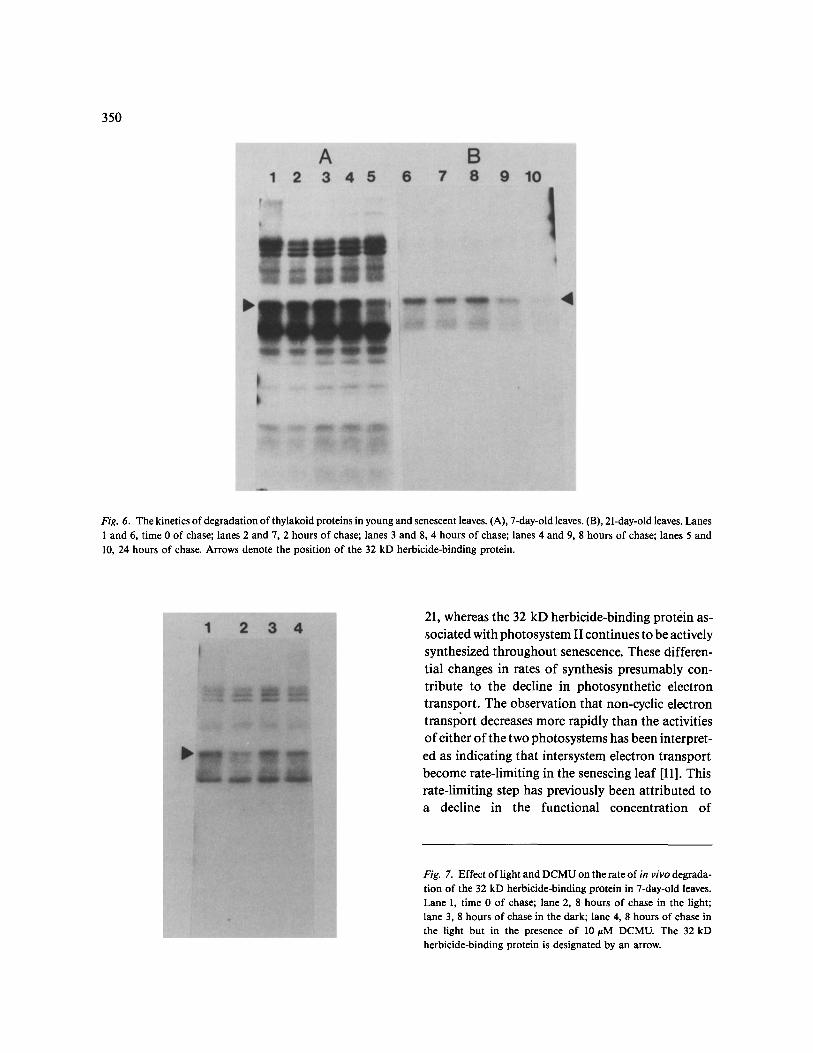
The closely similar Coomassie-banding patterns in SDS-polyacrylamide gels for thylakoids from 7- and 21-day-old leaves (Fig. 3) suggested that there are no pronounced differential changes in the rates at which thylakoid proteins are degraded during the early stages of senescence. This prospect was exam- ined by pulse-chase experiments. Young (7-day-old) and senescing (21-day-old) leaves were pulse-labeled with [35S]-methionine for 4.5 hours, and the radi- oactivity was chased in the presence of excess non- radioactive methionine for 2, 4, 8 and 24 hours be- fore thylakoids were isolated and fractionated by SDS-PAGE. In young 7-day-old bean leaves, as in photosynthetic tissue of other higher plants [15], the 32 kD protein turns over more readily than other thylakoid proteins (Fig. 6A). None of the bean

348
Fig. 4. Western blot analysis of thylakoid proteins from senescing primary leaves of bean. Lanes 1 and 2, the 68 kD polypeptide of pho- tosystem I for 7- and 21-day-old leaves, respectively; lanes 3 and 4,/3 subunit of ATPase for 7- and 21-day-old leaves, respectively; lanes 5 and 6, 33 kD extrinsic polypeptide of photosystems II for 7- and 21-day-old leaves, respectively; lanes 7 and 8, 25 and 26 kD polypeptides of LHCP for 7- and 21-day-old leaves, respectively; lanes 9 and 10, cytochrome f for 7- and 21-day-old leaves, respectively; lanes 11 and 12, cytochrome b 6 for 7- and 21-day-old leaves, respectively. Separate experiments on different gels are shown.
thylakoid proteins apart from the 32 kD herbicide- binding protein showed any marked degradation over the 24-hour chase period (Fig. 6A). In senes- cing 21-day-old leaves, only the 32 kD protein and its putative 2 8 - 2 9 kD conformer were labeled to the point of detection (Fig. 6B). This presumably reflects an overall decline in protein synthesis in the older tissue. Free amino acids are known to be trans- located out of attached senescing leaves [28], and thus, notwithstanding enhanced proteolytic activity, it is unlikely that the decline in [35S]-methionine in- corporation into protein in the older leaves is due to a larger amino acid pool. The kinetics of degrada- tion for the 32 kD herbicide-binding protein ap- peared to be similar for young and senescing leaves (Fig. 6). As previously shown for Spirodela [16], in vivo degradation of the 32 kD protein in bean
thylakoids was light-dependent and inhibited by DCMU, a photosystem II herbicide (Fig. 7).
Synthesis o f thylakoid proteins during senescence
Synthesis of thylakoid proteins, as reflected by meas- urements of specific radioactivity (dpm//~g protein) after pulse-labeling with [35S]-methionine, declined with advancing senescence (Fig. 1). In these experi- ments, leaves of various ages were labeled for 4 hours with [35S]-methionine after which the thylakoids were isolated, washed thoroughly and counted for radioactivity. Specific radioactivity (dpm/gg pro- tein) of membrane proteins decreased by about 55 % between days 7 and 14 and thereafter decreased more slowly through day 28 as senescence progressed

349
teins appeared to be stable throughout the early stages of senescence, differential changes in their rates of synthesis were detectable in pulse-labeling experiments. Leaves of various ages were pulse-la- beled with [35S]-methionine, and thylakoid mem- branes were isolated and fractionated by SDS-PAGE (Fig. 8). Of particular interest are the findings that synthesis of the 32 kD herbicide-binding protein re- mains active throughout senescence, whereas syn- thesis of the ot & ~ subunits of ATPase (57 and 58 kD polypeptides) and of the 26 kD polypeptide of the LHCP complex declines dramatically as the leaves age (Fig. 8). Indeed, by day 21 the ~ &/3 subunits of ATPase and the 26 kD polypeptide of LHCP were not detectable in fluorograms that had been loaded with constant protein or constant radioactivity (Fig. 8). The 68 kD reaction center polypeptide of the photosystem I complex also showed decreased synthesis, particularly between days 7 and 14 (Fig. 8). Two other polypeptides, cytochrome f (38 kD) and cytochrome b 6 (23 kD), which were de- tectable in fluorograms from 7-day-old leaves, were not synthesized to the point of detection in the older membranes (Fig. 8).
Fig. 5. Changes in the steady-state level of 32 kD herbicide- binding protein during natural senescence as visualized by west- ern blot analysis. Lane 1, thylakoids from Spirodela oligorrhiza; lane 2, thylakoids from 7-day-old leaves; lane 3, thylakoids from 14-day-old leaves; lane 4, thylakoids from 21-day-old leaves; lane 5, thylakoids from 28-day-old leaves. Arrows denote the po- sition of the 32 kD herbicide-binding protein.
(Fig. 1). Sacher [26] has reported that senescent leaves take up exogenous amino acids less efficiently than young leaves. If this difference was large, it could account for the apparent decline in thylakoid protein synthesis with advancing senescence. How- ever, this does not appear to have been a major factor contributing to the decline in [35S]-methionine up- take into thylakoid proteins in the present study inas- much as the 32 kD herbicide-binding protein, which is turned over more rapidly than any other thylakoid proteins, was strongly labeled in both young and senescent leaves (Fig. 6).
Although the steady-state levels of thylakoid pro-
Discussion
Previous studies with primary bean leaves have demonstrated that the activities of photosystem I and photosystem II decrease with advancing senes- cence [11, 20]. However, the decline in intersystem electron transport is greater than the decline in ac- tivities of either of the two photosystems [11, 20]. For primary leaves of Phaseolus vulgaris, cv. Kinghorn (the variety used in the present study), photosystem I and photosystem II have been reported to decline by 25 °7o and 15 %, respectively, by 21 days after plant- ing relative to their activities in young 9-day-old leaves, whereas non-cyclic electron transport decreases by 50% [20]. The data from the present study indicate that over this same period there are pronounced differential changes in the rates at which thylakoid proteins are synthesized. In particu- lar, the capability to synthesize the 26 kD polypep- tide of LHCP, the o~ &/3 subunits of ATPase and cytochromes f and b6 declines dramatically by day
350
Fig. 6. The kinetics of degradation of thylakoid proteins in young and senescent leaves. (A), 7-day-old leaves. (B), 21-day-old leaves. Lanes 1 and 6, time 0 of chase; lanes 2 and 7, 2 hours of chase; lanes 3 and 8, 4 hours of chase; lanes 4 and 9, 8 hours of chase; lanes 5 and 10, 24 hours of chase. Arrows denote the position of the 32 kD herbicide-binding protein.
21, whereas the 32 kD herb ic ide-b ind ing pro te in as-
soc ia ted with pho tosys t em II cont inues to be act ively
synthesized t h roughou t senescence. These differen-
t ia l changes in rates o f synthesis p re sumab ly con-
t r ibu te to the decl ine in pho tosyn the t i c e lect ron
t r anspor t . The observa t ion tha t non-cyclic e lect ron
t r anspo r t decreases more rap id ly than the activi t ies
o f e i ther o f the two pho tosys tems has been in terpret -
ed as ind ica t ing tha t in tersystem elect ron t r anspo r t
become ra te- l imi t ing in the senescing leaf [11]. This
ra te- l imi t ing step has previously been a t t r ibu ted to
a decl ine in the func t iona l concen t ra t ion o f
Fig. 7. Effect of light and DCMU on the rate of in vivo degrada- tion of the 32 kD herbicide-binding protein in 7-day-old leaves. Lane 1, time 0 of chase; lane 2, 8 hours of chase in the light; lane 3, 8 hours of chase in the dark; lane 4, 8 hours of chase in the light but in the presence of 10 #M DCMU. The 32 kD herbicide-binding protein is designated by an arrow.

351
Fig. 8. SDS-PAGE of newly synthesized proteins in primary leaves of bean during progressive senescence. (A), pulse-labeled for 2 hours and loaded with constant protein (10 #g/lane). (B), pulse-labeled for 4.5 hours and loaded with constant protein (10 #g/lane). (C), pulse- labeled for 4.5 hours and loaded with constant radioactivity (6800 dpm/lane). Lane 1, 7-day-old leaves; lane 2, 14-day-old leaves; lane 3, 21-day-old le~aves; lane 4, 28-day-old leaves. Arrows from top to bottom denote the positions of the 68 kD reaction center protein of pho- tosystem I, ~ and fl subunits of ATPase, cytochrome f, the 32 kD herbicide-binding protein, the 26 kD polypeptide of LHCP and cytochrome b6, respectively. Molecular weight markers (kD) are specified.
cytochrome f /B 6 [3, 9], and it is clear from the pres- ent study that this reflects decreased synthesis of these components.
Differential changes in the rates of synthesis for thylakoid proteins could be expected to change their relative levels in the membrane. However, the time re- quired for this to be detectable following a decline in synthesis will be influenced by the rates at which the proteins are turned over as well as their relative abundance in the membrane, and this is likely to be different for each protein. For example, although synthesis of the 26 kD polypeptide of LHCP declined between days 7 and 21, its concentration in the thylakoid membrane as visualized in Coomassie stained gels and on western blots did not change per- ceptibly through day 28. This presumably reflects the high abundance of this protein in the thylakoids
as well as relatively slow degradation. Indeed, the half life for the 26 kD polypeptide of LHCP in Spirodela has been estimated to be 2 months (H. Hoffman-Falk, M.Sc. dissertation, Weizmann Insti- tute of Science, Rehovot, Israel). By contrast, cytochromes f and b6, for which there was also a de- cline in synthesis between days 7 and 21, are present in much lower concentrations in thylakoids and showed selective depletion as early as day 21. Changes in rates of protein degradation with ad- vancing senescence would also affect steady-state levels of the proteins in the membranes. However, this could only be evaluated for the 32 kD herbicide- binding protein, because in the older leaves this pro- tein and its putative 2 8 - 2 9 kD conformer were the only thylakoid proteins to become labeled reprodu- cibly to the point of detection after a 4.5-hour pulse

352
with [35S]-methionine. This presumably reflects the pronounced decline in synthesis of thylakoid pro- teins in the older leaves. Pulse labeling for periods longer than 4.5 hours was not attempted because of concern that the leaves would become altered if left in the detached state for periods longer than 28.5 hours (the cumulative duration of the labeling and chase periods). In some experiments with the older leaves, there was slight labeling of the 68 kD reaction center polypeptide of photosystem I (see Fig. 8), but this was not a reproducible feature of the labeling pattern for the older leaves (see Fig. 6).
For the 32 kD protein, which was readily labeled in the older membranes, there was no apparent change in degradation rate with advancing senes- cence. This is noteworthy in the context that pho- tosystem II activity as well as non-cyclic electron transport have been reported to decline by 25 % and 50%, respectively, over the same period [20]. Turno- ver of the 32 kD protein in the senescing leaves proved to be light-dependent and inhibited by DCMU, which is consistent with previous proposals that its rapid turnover reflects its propensity to sus- tain photodamage during normal photosynthetic function [16, 22, 30]. However, the present finding of no major change in its turnover rate in senescing leaves notwithstanding a decline in photosystem ac- tivity and non-cyclic electron transport, suggests that, at least in the senescing leaf, degradation of this protein may not be mediated by electron trans- port. The protective effect of DCMU on 32 kD pro- tein turnover has previously been attributed to bind- ing of the herbicide to its receptor on the 32 kD protein [17]. Mattoo and Edelman [15] have pro- posed that the 32 kD protein may undergo a light- mediated conformational change, which renders it prone to degradation. The contention that a confor- mational change precedes proteolysis is given cre- dence by the finding that in isolated thylakoids the 32 kD protein undergoes a light-dependent transfor- mation that alters its sensitivity to exogenous trypsin [18]. It is conceivable, therefore, that DCMU affords protection against degradation by preventing a light- induced change in conformation.
Thylakoids, unlike the plasmalemma and microsomal membranes, exhibit no change in lipid fluidity or in fatty acid composition with advancing
senescence over a period during which photosystem I, photosystem II and non-cyclic electron transport have been reported to decline by 57%, 74% and 89%, respectively [20]. Thus the decline in photosyn- thetic electron transport may well be due solely to changes in thylakoid proteins as the leaf ages. The present study has provided evidence for differential changes during the early stages of senescence in the rates at which thylakoid proteins are synthesized. This results in early, selective depletion of cytochromes f and b 6 and possibly alters the func- tional status of other proteins for which changes in steady-state levels are not detectable until the later stages of senescence. Of particular interest is the finding that, of the proteins most affected, some, such as cytochrome f, are chloroplast-encoded, whereas others, such as the structural polypeptide of LHCP, are nuclear-encoded. This indicates that translational disruptions in both the cytoplasmic and chloroplastic compartments may contribute to the decline in photosynthetic electron transport in the senescing leaf.
Acknowledgement
This research was supported by the Natural Sciences and Engineering Research Council of Canada.
References
1. Anderson JM: Consequences of spatial separation of pho- tosystem 1 and 2 in thylakoid membranes of higher plant chloroplasts. FEBS Letters 124:1-10 (1981).
2. Batt T, Woolhouse HW: Changing activities and sites of syn- thesis of photosynthetic enzymes in leaves of the Perilla frutescens (L.). J Expt Bot 26: 569-579 (1975).
3. Ben-David H, Nelson N, Gepstein S: Differential changes in the amount of protein complexes in the chloroplast mem- brane during senescence of oat and bean leaves. Plant Physiol 73:507-510 (1983).
4. Bengis C, Nelson N: Subunit structure of chloroplast pho- tosystem I reaction center. J Biol Chem 252:4564-4569 (1977).
5. Borochov A, Halevy AH, Shinitzky M: Increase in microvis- cosity with aging in protoplast plasmalemma of rose petals. Nature 263:158-159 (1976).
6. Haehnel W: Photosynthetic election transport in higher plants. Ann Rev Plant Physiol 35:659-693 (1984).

7. Hirschberg J, Mclntosh L: Molecular basis of herbicide resis- tance in Amaranthus hybridus. Science 222:1346-1349 (1983).
8. Hiscox JD, Israelstam GF: A method for the extraction of chlorophyll from leaf tissue without maceration. Can J Bot 57:1332-1334 (1979).
9. Holloway PJ, Maclean DJ, Scott K J: Rate-limiting steps of electron transport in chloroplasts during ontogeny and senes- cence of barley. Plant Physiol 72:795-801 (1983).
10. Hurt E, Hauska G: Identification of the polypeptides in the cytochrome b6/f complex from spinach chloroplasts with redox-center carrying subunits. J Bioenerg Biomembranes 14:405-424 (1982).
11. Jenkins GI, Woolhouse HW: Photosynthetic electron trans- port during senescence of the primary leaves of Phaseolus vulgaris L. II. The activity of photosystems I and II and a note on the site of reduction of ferricyanide. J Expt Bot 32: 989-997 (1981).
12. Laemmli UK: Cleavage of structural proteins during the as- sembly of the head of the bacteriophage T 4. Nature 227: 680-685 (1970).
13. Legge RL, Thompson JE, Murr DP, Tsujita J: Sequential changes in lipid fluidity and phase properties of microsomal membranes from senescing rose petals. J. Expt Bot 33: 303-312 (1982).
14. Lowry OH, Rosebrough N J, Farr AL, Randall R J: Protein
measurement with the Folin phenol reagent. J Biol Chem 193: 265-275 (1951).
15. Mattoo AK, Edelman M: Photoregulation and metabolism of a thylakoidal herbicide-receptor protein. In: St. John JB, Berlin E, Jackson PC (eds) Frontiers of Membrane Research (Beltsville Symposium 9) pp. 23-34. Totowa: Rowman and Allanheld (1985).
16. Mattoo AK, Hoffman-Falk H, Marder JB, Edelman M: Regulation of protein metabolism: coupling of photosynthet- ic electron transport to in vivo degradation of the rapidly metabolized 32-kilodalton protein of the chloroplast mem- branes. Proc Natl Acad Sci USA 81: 1380-1384 (1984).
17. Mattoo AK, Marder JB, Gaba V, Edelman M: Control of 32 kDa thylakoid protein degradation as a consequence of herbicide binding to its receptor. In: Akoyunoglou G, Senger H (eds) Regulation of Chloroplast Differentiation. Plant Bi- ology Series. New York: Alan R. Liss Inc. (1986) in press.
18. Mattoo AK, Pick U, Hoffman-Falk H, Edelman M: The rapidly metabolized 32000-dalton polypeptide of the chlo- roplast is the proteinaceous shield regulating photosystem II
353
electron transport and mediating diuron herbicide sensitivi- ty. Proc Natl Acad Sci USA 78:1572-1576 (1981).
19. McKersie BD, Lepock JR, Kruuv J, Thompson JE: The ef- fects of cotyledon senescence on the composition and physi- cal properties of membrane lipids. Biochim Biophys Acta 508:197-212 (1978).
20. McRae DG, Chambers JA, Thompson JE: Senescence- related changes in photosynthetic electron transport are not due to alterations in thylakoid fluidity. Biochim Biophys Acta 810:200-208 (1985).
21. Nelson N: Structure and synthesis of chloroplast ATPase. Methods Enzymol 97:510-523 (1983).
22. Ohad I, Kyle DJ, Hirschberg J: Light-dependent degradation of the QB-protein in isolated pea thylakoids. EMBO Journal 4:1655-1659 (1985).
23. Peferoen M, Huybrechts R, DeLoof A: Vacuum-blotting: a new simple and efficient transfer of proteins from sodium dodecyl sulfate-polyacrylamide gels to nitrocellulose. FEBS Letters 145:369-372 (1982).
24. Poser HB: Aquatic vascular plants. In: Wilt FH, Wessels NK (eds) Methods in Developmental Biology, pp. 301- 317. New York: Crowell (1967).
25. Reisfeld A, Mattoo AK, Edelman M: Processing of a chlo- roplast-translated membrane protein in vivo. Europ J Bi- ochem 124:125-129 (1982).
26. Sacher JA: Studies of permeability, RNA and protein turno- ver during ageing of fruit and leaf tissues. In: Aspects of the Biology of Aging, pp. 269- 304. Symposia of the Society for Experimental Biology. Cambridge: Cambridge University Press (1967).
27. Takano M, Takahashi M-A, Asada K: Reduction of pho- tosystem I reaction center, P-700, by plastocyanin in stroma thylakoids from spinach: Lateral diffusion of plastocyanin. Arch Biochem Biophys 218:369-375 0982).
28. Thimann KV, Tetley RM, Tran Thanh Van M: The metabo- lism of oat leaves during senescence. II. Senescence in leaves attached to the plant. Plant Physiol 54:858-862 (1974).
29. Thompson JE, Mayak S, Shinitzky M, Halevy AH: Accelera- tion of membrane senescence in cut carnation flowers by treatment with ethylene. Plant Physiol 69:859-863 (1982).
30. Woolhouse HW: The biochemistry and regulation of senes- cence in chloroplasts. Can J Bot 62:2934-2942 (1984).
31. Yamamoto Y, Doi M, Tamura N, Nishimura M: Release of polypeptides from highly active O2-evolving photosystem 2 preparation by Tris treatment. FEBS Lett 133:265-268 (1981).











![Nora Roberts - A fost odata o stea [ibuc info]](https://img.dokumen.tips/doc/110x75/63248a2a4d8439cb620d6e07/nora-roberts-a-fost-odata-o-stea-ibuc-info.jpg)

















