Embed Size (px)
Citation preview

Ro
GMa
b
c
d
a
ARRAA
KCOPPT
1
sotJ2rbfdtQ2fi
R
0d
Neuropsychologia 48 (2010) 3513–3520
Contents lists available at ScienceDirect
Neuropsychologia
journa l homepage: www.e lsev ier .com/ locate /neuropsychologia
esonance of cortico–cortical connections of the motor system with thebservation of goal directed grasping movements
iacomo Kocha,b,∗, Viviana Versacea, Sonia Bonnìa, Federica Lupoa, Emanuele Lo Gerfoa,assimiliano Oliveri a,d, Carlo Caltagironea,c
Laboratorio di Neurologia Clinica e Comportamentale, Fondazione Santa Lucia IRCCS, Via Ardeatina, 306, 00179 Rome, ItalyStroke Unit, Dipartimento di Neuroscienze, Università di Roma Tor Vergata, Viale Oxford 81, 00133 Rome, ItalyClinica Neurologica, Dipartimento di Neuroscienze, Università di Roma Tor Vergata, Viale Oxford 81, 00133 Rome, ItalyDipartimento di Psicologia, Università di Palermo, Palermo, Italy
r t i c l e i n f o
rticle history:eceived 14 March 2010eceived in revised form 13 July 2010ccepted 28 July 2010vailable online 4 August 2010
eywords:onnectivitybservation
a b s t r a c t
Goal directed movements require the activation of parietal, premotor and primary motor areas. In mon-keys, neurons of these areas become active also during the observation of movements performed byothers, especially for coding the goal of the action (mirror system). Using bifocal transcranial magneticstimulation (TMS) in healthy subjects, we tested whether the observation of goal directed reach to graspactions may lead to specific changes in the short-latency connections linking key areas of the mirrorsystem, such as the anterior intraparietal cortex (AIP) and the ventral premotor cortex (PMv), with theprimary motor cortex (M1). We found that AIP-M1 and PMv-M1 cortico–cortical interactions were specif-ically activated when observing successful reaching to grasp goal directed actions, in which the hand
arietal cortexremotor cortexranscranial magnetic stimulation
posture was congruent with the goal of the action performed by the actor. On the other hand they werenot modified when the same goal directed actions were performed wrongly with an inappropriate grasp-ing posture. A similar profile of excitability was observed when testing specific intracortical facilitatorycircuits in M1 (I2-waves), known to reflect the activity in cortico–cortical pathways transmitting infor-mation from PMv. We conclude that the simple observation of others’ goal directed actions is able toinduce specific neurophysiological changes in some cortico–cortical circuits of the human motor system.
. Introduction
The planning and execution of simple goal directed movementsuch as reaching and grasping a cup to drink require the activationf an interconnected cortical network formed by parietal, premo-or and primary motor areas (Binkofski et al., 1998; Castiello, 2005;eannerod, Arbib, Rizzolatti, & Sakata, 1995; Rizzolatti & Luppino,001). In monkeys, neurons of these areas (area F5 in the infe-ior frontal cortex and area PF/PFG in the inferior parietal cortex)ecome active also during the observation of movements per-ormed by others, leading to the notion that the same system isirectly involved in action understanding, without the interven-
ion of other sophisticated cognitive processes (Kraskov, Dancause,uallo, Shepherd, & Lemon, 2009; Rizzolatti, Fogassi, & Gallese,001; Rizzolatti, Fabbri-Destro, & Cattaneo, 2009). These neuronsre during observation of actions that share the same goal, but∗ Corresponding author at: Fondazione S. Lucia IRCCS, Via Ardeatina 306, 00179ome, Italy. Tel.: +39 06 51501369; fax: +39 06 51501388.
E-mail addresses: [email protected], [email protected] (G. Koch).
028-3932/$ – see front matter © 2010 Elsevier Ltd. All rights reserved.oi:10.1016/j.neuropsychologia.2010.07.037
© 2010 Elsevier Ltd. All rights reserved.
are performed with different effectors, suggesting that actions arerepresented in this cortical region with respect to their goals.
Functional neuroimaging investigations provided evidence thatsuch a mirror system may also exist in humans, reporting asimilar profile of task-related activation in the same parietaland premotor regions during movement observation (Buccino,Binkofski, Fink, Fadiga, & Fogassi, 2001; Calvo-Merino, Glaser,Grèzes, Passingham, & Haggard, 2005; Grafton, Arbib, Fadiga, &Rizzolatti, 1996; Iacoboni et al., 2005; Iacoboni, Woods, Brass,Bekkering, & Mazziotta, 1999).
Furthermore, neurophysiological investigations using transcra-nial magnetic stimulation (TMS) showed that the amplitude ofmotor evoked potentials (MEPs), obtained by stimulating the pri-mary motor cortex (M1), change during the observation of specificmovements performed by others (Aglioti, Cesari, Romani, & Urgesi,2008; Clark, Tremblay, & Ste-Marie, 2004; Fadiga, Craighero, &
Olivier, 2005; Fadiga, Fogassi, Pavesi, & Rizzolatti, 1995; Gangitano,Mottaghy, & Pascual-Leone, 2001; Strafella & Paus, 2000; Urgesi,Candidi, Fabbro, Romani, & Aglioti, 2006). These findings suggestthat the simple observation of movements is able to activate thecortical representation of the same muscles involved in the actual
3 cholog
tagm
fitn(ihaSP
bacwseu2&OMR
owrt2aaroihc&
Pta
2
2
mttC
2
irh
6btw
i
514 G. Koch et al. / Neuropsy
ask. Besides, a recent study was able to demonstrate that the neuralctivity of M1 was specifically enhanced depending on the actionoal, instead of the muscular pattern required to perform a certainovement (Cattaneo, Caruana, Jezzini, & Rizzolatti, 2009).These investigations detected only changes occurring in M1, the
nal output of the motor system, however they did not explorehe functional role of other cortical areas in which the mirroreurons have been found, such as the ventral premotor cortexPMv) and the anterior intraparietal cortex (AIP). Recent TMS stud-es using a “virtual lesion” approach indeed reported that also inumans there is neurophysiological evidence for the role of parietalnd premotor areas in action observation (Cattaneo, Mandrini, &chwarzbach, 2010; Avenanti, Bolognini, Malavita, & Aglioti, 2007;obric & Hamilton, 2006).
Therefore in the current study we aimed to verify, by means ofifocal twin-coil TMS method, if the observation of goal directedctions may lead to specific changes in the excitability of theonnections linking AIP and PMv with M1. With this method,hen a conditioning pulse is applied over these cortical regions,
hortly prior to a test pulse over the hand area of M1, the motorvoked response triggered by the M1 pulse is specifically mod-lated depending on the current task (Davare, Lemon, & Olivier,008; Davare, Montague, Olivier, Rothwell, & Lemon, 2009; KochRothwell, 2009a, 2009b; Koch et al., 2006; Koch, Fernandez Del
lmo, et al., 2008; Koch, Oliveri, et al., 2008; Koch et al., 2010;ars et al., 2009; O’Shea, Sebastian, Boorman, Johansen-Berg, &
ushworth, 2007).Bifocal TMS provides the unique opportunity to investigate
ngoing causal interactions between interconnected cortical areasith a high temporal resolution. In fact, this technique may
eveal whether a specific cortical area exerts task-related inhibi-ion or facilitation over the primary motor cortex (see Koch et al.,006), with important physiological implications, that cannot beddressed using imaging techniques (see Iacoboni et al., 1999). Thedvantage of probing these pathways with TMS methods is that theesponse to a TMS conditioning pulse depends on the excitabilityf the pathway at the time the stimulus is applied. Thus changesn the effectiveness of the conditioning pulse give an indication ofow the excitability of the connection changes over time when theortical networks becomes active during a specific motor task (KochRothwell, 2009a, 2009b).We hypothesized that changes in excitability of AIP-M1 and
Mv-M1 connections should be detected not only during motorasks but also during the simple observation of different types ofctions.
. Materials and methods
.1. Subjects
We studied 12 right-handed (Oldfield, 1971) healthy subjects (6 men; 6 women;ean age: 26.2 years ± 4.3). All participants gave their written informed consent
o take part in the research. The study was conducted according to the Declara-ion of Helsinki and the experiments were approved by the IRCCS “S. Lucia” Ethicalommittee.
.2. Video clips
Each participant was tested in the different experimental session lasting approx-mately 60 min. Participants sat in a comfortable armchair in a dimly illuminatedoom with their right hand resting on a pillow, with their elbows flexed at 90◦ andands pronated in a totally relaxed position.
They were sitting at a distance of 80 cm in front of a 19-inch monitor (resolution,
40 × 480 pixels; refresh frequency, 85 Hz) on which stimuli appeared on a uniformackground and subtended a visual angle of 10.4◦ × 12.6◦ . Stimulus-presentationiming, TMS triggering, and randomization were controlled using PsySchope soft-are running on an Apple Macintosh Computer.They watched several clips that showed three experimental conditions repeatedn a pseudo-random order for sixteen times (96 trials) (see Fig. 1).
ia 48 (2010) 3513–3520
Subjects watched three different types of movie clips showing: a successful goaldirected action (GDA), a goal directed action performed wrongly with inappropriategrasping (iGDA) and a neutral condition in which no movement was performed,although the target object was presented (STATIC).
The GDA consisted in clips in which the actors performed two types of reach tograsp movements in order to grasp a small cup or a big cup. To be successful, theseactions required different hand postures such as precision grip (PG) for the smallcup and whole hand grasp (WHG) for the big cup. In this condition there were twoclips showing: (i) a hand reaching, pinching with the right index and thumb a smallbottle top and then lifting the object to a height of approximately 5 cm (precisiongrip, PG) and (ii) a hand reaching and grasping with the whole hand a big cup andthen lifting the object to a height of approximately 5 cm (whole hand grasp, WHG).
The iGDA clip presented the same objects to be grasped. However the actionswere incongruent, since the actors performed a PG to unsuccessfully grasp the bigcup and a WHG to try to grasp the small cup. There was a clip showing: (i) a handreaching and pinching the cup with an inappropriate movement (PG), without liftingit; (ii) a hand reaching and grasping the bottle top with an inappropriate movementwithout lifting it (WHG).
Finally in the STATIC condition, although both objects were randomly presented,no action was performed by the actors. For the STATIC condition there were twoclips, one showing a resting hand in front of the bottle top and the other a restinghand in front of the cup (see Fig. 1).
All the videos had the same duration, i.e. 5000 ms sampled in 120 frames. Move-ments started 1000 ms after the beginning of the video. This duration was chosenin accordance with previous investigations (Gangitano et al., 2001). The end of thevideo was followed by a full black screen for 5000 ms (see Fig. 1).
2.3. Experiment 1. Changes in the excitability of parieto–motor connectivity andof premotor–motor connectivity during observation of movements
Twelve right-handed subjects took part in this experiments in which we testedpossible changes occurring in the AIP-M1 and PMv-M1 connections during theobservation of different clips. AIP-M1 and PMv-M1 connections were tested in dif-ferent sessions performed at least one week apart. The order of presentation of thedifferent conditions was counterbalanced across subjects.
2.4. Transcranial magnetic stimulation
Electromyographic (EMG) traces were recorded from the right first dorsalinterosseous (FDI) and abductor digiti minimi (ADM) muscles of the right handusing 9-mm diameter, Ag–AgCl surface cup electrodes. For the FDI muscle, theactive electrode was placed over the muscle belly and the reference electrode overthe metacarpophalangeal joint of the index finger. For the ADM muscle, the activeelectrode was placed over the muscle belly and the reference electrode over themetacarpophalangeal joint of the little finger (see Fig. 1). Responses were amplifiedwith a Digitimer D360 amplifier (Digitimer Ltd., Welwyn Garden City, Hertford-shire, UK) through filters set at 20 Hz and 2 kHz with a sampling rate of 5 kHz,then recorded by a computer using SIGNAL software (Cambridge Electronic Devices,Cambridge, UK).
To test AIP-M1 and PMv-M1 connectivity, we used a paired pulse stimulationtechnique with two high-power Magstim 200 machines (Magstim Co., Whitland,Dyfed, UK). The magnetic stimulus had a nearly monophasic pulse configurationwith a rise time of about 100 �s, decaying back to zero over about 0.8 ms.
First the intensity of TMS was adjusted to evoke an MEP of approximately 1 mVpeak-to-peak in the relaxed right FDI and ADM. The hand motor area of left M1was defined as the point where stimulation evoked the largest MEP from bothmuscles. The test stimulator was connected to a small custom-made figure-of-eight-shaped coil (external diameter 50 mm). The coil over M1 was always placedtangentially to the scalp at 45◦ angle from midline of the central sulcus, inducing aposterior–anterior (PA) current flow. The conditioning stimulator was connected toanother small custom-made figure-of-eight-shaped coil (external diameter 50 mm).
We used a neuronavigation system (Softaxic, E.M.S., Bologna, Italy) to preciselyposition the coil over the stimulation sites, using individual anatomical magneticresonance images; this technique has been described in detail previously (Koch et al.,2007). The individual coordinates of each stimulation site were normalized a poste-rior into the Montreal Neurological Institute (MNI) coordinate system and averaged.To target AIP, the coil was positioned in the anterior portion of IPL (supramarginalgyrus), close to the intersection between the intraparietal sulcus and postcentralsulcus. The centre of the coil was positioned tangentially to the skull with the han-dle pointing downward and posteriorly (30◦). Mean normalized MNI coordinatesof AIP were −52.8 ± 4.1, −41.6 ± 3.8, and 56.1 ± 4.5 mm (x, y, z, mean ± SD). On thebasis of our recent published paired pulse TMS work with PPC and M1 sites (Kochet al., 2007; Koch, Fernandez Del Olmo, et al., 2008; Koch, Oliveri, et al., 2008; Kochet al., 2010) we used the following parameters to best activate AIP-M1 interplay.
The intensity of the M1 pulse was adjusted to evoke an MEP of approximately 1 mVpeak-to-peak in the relaxed right FDI. In half of the trials, M1 TMS was preceded bya conditioning pulse delivered 6 ms earlier, over the ipsilateral AIP at an intensity of90% of the resting motor threshold (RMT). We defined RMT as the lowest intensitythat evoked five small responses (about 50 �V) in the contralateral FDI muscle in aseries of ten stimuli when the subject kept the FDI muscles relaxed in both hands.
G. Koch et al. / Neuropsychologia 48 (2010) 3513–3520 3515
F xperi6 ls wer(
og(aaMai2
mgowaasap
2o
ptogpwcetat
ig. 1. (A) Schematic representation of the different clips showed during the TMS ems in half of the trails the test stimulus delivered over M1. Motor evoked potentia
ADM) muscles.
To target PMv, the coil was positioned over the caudal portion of the parspercularis of the inferior frontal gyrus. The centre of the coil was positioned tan-entially to the skull with the handle pointing downward and slightly posteriorly10◦). Mean normalized MNI coordinates of PMv site were −52.8 ± 4.1, 11.6 ± 3.8,nd 25.1 ± 4.5 mm (x, y, z, mean ± SD). We used the following parameters to bestctivate PMv-M1 interplay: the intensity of the M1 pulse was adjusted to evoke anEP of approximately 1 mV peak-to-peak in the relaxed right FDI; in half of the tri-
ls, M1 TMS was preceded by a conditioning pulse delivered 6 ms earlier, over thepsilateral PMv at an intensity of 90% of the RMT (Bäumer et al., 2009; Davare et al.,008, 2009).
The different clips were presented with TMS delivered in a single or paired pulseodality at the end of the reaching phase just before the actors were beginning to
rasp the target object (4000 ms after the onset of the clip, 3000 ms after the onsetf the movement). We choose this late delay in order to apply TMS when the actorsere assuming the appropriate (or inappropriate) posture of the hand to executePG or a WHG movement. There were in each sub-condition eight trials (single
nd pair pulses) for a total of 96 trials. Within each experimental conditions in eachubject, the mean peak-to-peak amplitude of the conditioned MEP was expresseds a percentage of the mean peak-to-peak amplitude size of the unconditioned testulse to use averages of MEPs.
.5. Experiment 2. Changes in the excitability of M1 intracortical circuits duringbservation of movements
In this experiment, performed in six subjects that took part in experiment 1,aired stimuli were applied over left M1, in order to test changes in M1 intracor-ical circuits that are thought to receive the input form PMv (Cattaneo et al., 2005)r AIP (Koch et al., 2007), namely the late I-wave circuits. It is known that a sin-le TMS pulse produces a repetitive discharge of the corticospinal neurons in therimary motor cortex (Amassian & Stewart, 2003; Di Lazzaro et al., 2003). The first
ave of excitation results from direct activation of corticospinal neurons and isalled the direct or D wave. The succeeding indirect waves (I-waves, termed I1, I2, I3,tc.) are thought to depend on transsynaptic excitation of intracortical interneuronshat synapse on the corticospinal neurons (Amassian & Stewart, 2003; Di Lazzaro etl., 2003). Indeed, the later I-waves mediate the input of cortico–cortical pathwaysransmitting information from other cortical areas (Cattaneo et al., 2005; Shimazu,
ments. (B) A conditioning stimulus was applied over the AIP or PMv, preceding bye recorded by the right first dorsal interosseus (FDI) and the abductor digiti minimi
Maier, Cerri, Kirkwood, & Lemon, 2004). To test this circuit, we therefore used ashort intracortical facilitation protocol (SICF) (Ziemann et al., 1998). Test stimulus(TS) was set at 130% RMT and was followed by a subthreshold conditioning stim-ulus (CS) adjusted at an intensity of 90% RMT. The ISIs of 2.5 ms (corresponding tothe peak of the I2-wave) was used on the basis of a previous study showing thatduring preparation for grasp, the modulation of the MEP obtained at ISI 2.5 ms is apredictor of muscle activity that will be used during grasp and that likely reflects theactivation of cortico–cortical inputs to M1, including those from premotor cortex,that represent the object to be grasped in terms of the actions required to grasp it(Cattaneo et al., 2005).
We also verified possible changes involving other inhibitory circuits such as theshort intracortical inhibition (SICI) (Kujirai et al., 1993) that are also involved inmotor preparation (Reynolds & Ashby, 1999). A subthreshold CS at 90% RMT wasapplied 2 ms before the test stimulus (TS). The intensity of the TS was adjusted toevoke an MEP of approximately 1 mV peak-to-peak in the relaxed right FDI. Thedifferent clips were presented with TMS delivered as in experiment 1. Data wereanalyzed as in experiment 1.
3. Data analysis
The effects of paired stimulation on the size of MEP recordedfrom the contralateral FDI and ADM in response to M1 TMS werefirst analyzed as the percentage of the mean peak-to-peak ampli-tude of the unconditioned test M1 pulse. In experiment 1, meanpercentage values were analyzed with a four-way repeated mea-sures analyses of variance (ANOVA) with CONNECTION (AIP-M1 vs.PMv-M1), CLIP (GDA vs. nGDA vs. STATIC), GRASP (PG vs. WHG)
and MUSCLE (FDI vs. ADM) as within-subjects main factors. Inexperiment 2, mean percentage values were analyzed with a three-way repeated measures analyses of variance (ANOVA) with CLIP(GDA vs. nGDA vs. STATIC), GRASP (PG vs. WHG) and MUSCLE(FDI vs. ADM) as within-subjects main factors. Moreover in each
3 chologia 48 (2010) 3513–3520
epApn
4
4
owmmb(piaAof1pnadtmf1pwwtfism
4
sbp(momptoi
Fig. 2. Effects of AIP conditioning on ipsilateral M1 excitability during the observa-
TM
516 G. Koch et al. / Neuropsy
xperiment the same ANOVAs were also performed on the meaneak-to-peak MEPs amplitude obtained with M1 stimulation alone.significant main effect in the ANOVA was followed by Tukey’s
ost hoc analysis. The Greenhouse-Geisser correction was used foron-spherical data. A p value <0.05 was considered significant.
. Results
.1. Experiment 1
Subjects were completely relaxed during presentation of actionbservation videos, since no background EMG activity was detectedhen showing both GDA and nGDA clips. We found in this experi-ent that both AIP-M1 and PMv-M1 connections were selectivelyodulated by the observed type of grasp movement as revealed
y the ANOVA analysis showing a significant main effect of CLIPF(2,22) = 2.93; p < 0.05), significant SITE × MUSCLE (F(1,11) = 7.83;< 0.05) and MUSCLE × GRASP (F(1,11) = 6.19; p < 0.05) double
nteractions and a significant CLIP × GRASP × MUSCLE triple inter-ction (F(2,22) = 3.36; p < 0.05). Post hoc analysis showed thatIP-M1 excitability was selectively enhanced for the FDI musclenly during the observation of GDA clip when actors were per-orming a PG movement towards the small target (GDA vs. nGDA:09.99 vs. 76.66%; p < 0.05; GDA vs. STATIC: 109.99 vs. 86.37%;< 0.05) (Fig. 2). These findings were specific for the AIP-M1 con-ections since when the same analysis was performed on MEPsmplitude obtained for both muscle by TS alone, no significantifference emerged (Table 1). Indeed, post hoc analysis showedhat PMv-M1 excitability was selectively enhanced only for the FDI
uscle during the observation of GDA clip when actors were per-orming a PG movement towards the small target (GDA vs. nGDA:35.66 vs. 104.16%; p < 0.05; GDA vs. STATIC: 135.66 vs. 102.25%;< 0.05). On the contrary, for the ADM muscle, PMv-M1 excitabilityas selectively enhanced only during the observation of GDA cliphen actors were performing a WHG movement towards the big
arget (GDA vs. nGDA: 118.67 vs. 90.52%; p < 0.05) (Fig. 3). Thesendings were specific for the PMv-M1 connections since when theame analysis was performed on MEPs amplitude obtained for bothuscle by TS alone, no significant difference emerged (Table 2).
.2. Experiment 2
We found in this experiment that SICF intracortical circuits wereensitive to the observed type of shown movement as revealedy ANOVA showing a significant CLIP main factor ((F(2,10) = 13.72;< 0.01) and a significant triple CLIP × GRASP × MUSCLE interaction
F(2,10) = 4.81; p < 0.05). Post hoc analysis showed that for the FDIuscle SICF excitability was selectively enhanced only during the
bservation of GDA clip when actors were performing a PG move-
ent towards the small target (GDA vs. nGDA: 191.34 vs. 124.66%;< 0.05; GDA vs. STATIC: 191.34 vs. 127.97%; p < 0.05). On the con-rary for the ADM muscle SICF excitability was selectively enhancednly during the observation of GDA clip when actors were perform-ng a WHG movement towards the big target (GDA vs. nGDA: 161.38
able 1EP amplitude (mean ± S.D.) obtained in different experimental conditions in experimen
Static GDA
M1 AIP-M1 M1
FDIPG (small) 1.39 ± 0.41 1.18 ± 0.32 1.27 ± 0WHG (big) 1.42 ± 0.33 1.33 ± 0.28 1.31 ± 0
ADMPG (small) 0.98 ± 0.32 0.96 ± 0.26 1.02 ± 0WHG (big) 0.92 ± 0.25 0.82 ± 0.24 0.92 ± 0
tion of different clips. Panels A and B show the changes of the mean percentage ofMEP amplitudes (with 100% representing no change) due to left AIP conditioningrecorded from the right FDI (A) or right ADM (B). Asterisks indicate a significantvalue at post hoc analysis p < 0.05. Errors bars indicate S.E.M.
vs. 109.44%; p < 0.05<, GDA vs. STATIC: 161.38 vs. 82.47%; p < 0.05)(Fig. 4).
For SICI circuits no significant differences emerged from theANOVA (Fig. 5).
5. Discussion
The current results demonstrate that, in humans, the sim-ple observation of successful goal directed actions is sufficientto activate short-latency cortico–cortical connections betweennon primary and primary motor areas. The excitability ofcortico–cortical pathways originating from key areas of the mir-ror system, such as the AIP and PMv, does not simply change inresponse to the observation of a specific hand posture, but increasesselectively only when both the hand posture and the goal of theaction performed by the actor are congruent. Furthermore a similarprofile of activation was observed when testing a specific facilita-tory intracortical circuit in M1, that is known to reflect the activityin cortico–cortical pathways transmitting information from other
cortical areas. Taken together, these data demonstrate that observ-ing others’ actions induces specific neurophysiological changes inshort-latency cortico–cortical circuits of the motor system.In both AIP and PMv specific sets of neurons that fire inresponse to the observations of others’ actions have been constantly
t 1.
nGDA
AIP-M1 M1 AIP-M1
.42 1.45 ± 0.45 1.38 ± 0.39 1.22 ± 0.28
.43 1.24 ± 0.32 1.37 ± 0.36 1.25 ± 0.45
.25 0.81 ± 0.38 1.11 ± 0.31 0.96 ± 0.26
.36 1.02 ± 0.26 0.96 ± 0.31 0.85 ± 0.33

G. Koch et al. / Neuropsychologia 48 (2010) 3513–3520 3517
Fig. 3. Effects of PMv conditioning on ipsilateral M1 excitability during the obser-vMrv
isngs
5
seipwwlEAcLmTeg
Fig. 4. Changes in the excitability of the I2-waves intracortical circuits during theobservation of different clips. Panels A and B show the changes of the mean per-centage of MEP amplitudes (with 100% representing no change) recorded from the
TM
ation of different clips. Panels A and B show the changes of the mean percentage ofEP amplitudes (with 100% representing no change) due to left PMv conditioning
ecorded from the right FDI (A) or right ADM (B). Asterisks indicate a significantalue at post hoc analysis p < 0.05. Errors bars indicate S.E.M.
dentified (Rizzolatti et al., 2009). Thus, as discussed below, weuggest that the increased excitability of AIP-M1 and PMv-M1 con-ections that emerges during observation of others’ reaching torasp movements could reflect the neural activity of these specificet of neurones.
.1. AIP-M1 connection
In a recent study, using combined tc-TMS and Diffusion Ten-or Imaging (DTI) methods (Koch et al., 2010), we found that thexcitability of the short-latency AIP-M1 connection change dramat-cally during planning of PG movements, and marginally duringreparation of WHG movements. These results are in agreementith recent fMRI investigations, revealing that the AIP is activehen subjects perform a PG to reach and grasp a small object, with
ess activity for WHG (Begliomini, Wall, Smith, & Castiello, 2007;hrsson et al., 2000) and with previous literature showing thatIP is involved in both hand configurations and object-size basedoding (Biagi et al., 2010; Baumann et al., 2009; Murata, Gallese,
uppino, Kaseda, & Sakata, 2000 JN). Moreover, in monkeys AIP,irror neurons have lately been described (Rizzolatti et al., 2009).hese neurons seems to show the same features of those discov-red in the premotor cortex, being crucially involved in coding theoals of observed motor acts (Rizzolatti et al., 2009).
able 2EP amplitude (mean ± S.D.) obtained in different experimental conditions in experimen
Static GDA
M1 PMv-M1 M1
FDIPG (small) 1.40 ± 0.43 1.51 ± 0.36 1.47 ± 0WHG (big) 1.48 ± 0.32 1.50 ± 0.35 1.34 ± 0
ADMPG (small) 0.91 ± 0.36 0.84 ± 0.29 0.88 ± 0WHG (big) 0.80 ± 0.31 0.71 ± 0.24 0.80 ± 0
right FDI (A) or right ADM (B) when a conditioning pulse was applied over M1 2.5 msafter the test pulse. Asterisks indicate a significant value at post hoc analysis p < 0.05.Errors bars indicate S.E.M.
Consistent with this background, in the current study we foundthat AIP-M1 was also activated during the observation of goaldirected grasping movements, specifically for PG movements, andonly marginally for WHG movements, thus implying a specificmodulation depending both on the goal of the action and on thetype of grasp.
Nevertheless, the activation of AIP-M1 connection during theobservation of WHG movements could have been masked becausein the experimental clips the observed grasp was viewed from theside. The observer was not able to fully see the little finger, andthis could explain the finding that only the PG influenced AIP-M1interactions, since AIP is more visually dependent than PMv, wherethe grasp is encoded more in motor reference frame (Murata etal., 1997; Murata et al., 2000). For both the AIP-M1 and PMv-M1interactions we used the same ISI of 6 ms, on the basis of previ-ous investigations performed with subjects at rest (Bäumer et al.,2009; Koch et al., 2007), in which it was demonstrated that for bothconnections a peak of activation was found around these latencies.Given that the effects of AIP stimulation on M1 could involve eitherdirect projections, or more likely the activity of an indirect pathway
that involves the ipsilateral PMv and then M1 (Koch et al., 2010),one could expect that a longer ISI would be more appropriate totest AIP-M1 interactions. However the true latency of the interac-tion is complicated by the fact that a single suprathreshold TMSt 2.
nGDA
PMv-M1 M1 PMv-M1
.35 1.88 ± 0.49 1.49 ± 0.41 1.54 ± 0.33
.42 1.48 ± 0.38 1.47 ± 0.35 1.51 ± 0.31
.22 0.84 ± 0.31 0.95 ± 0.38 0.94 ± 0.34
.39 1.05 ± 0.29 0.82 ± 0.28 0.75 ± 0.38
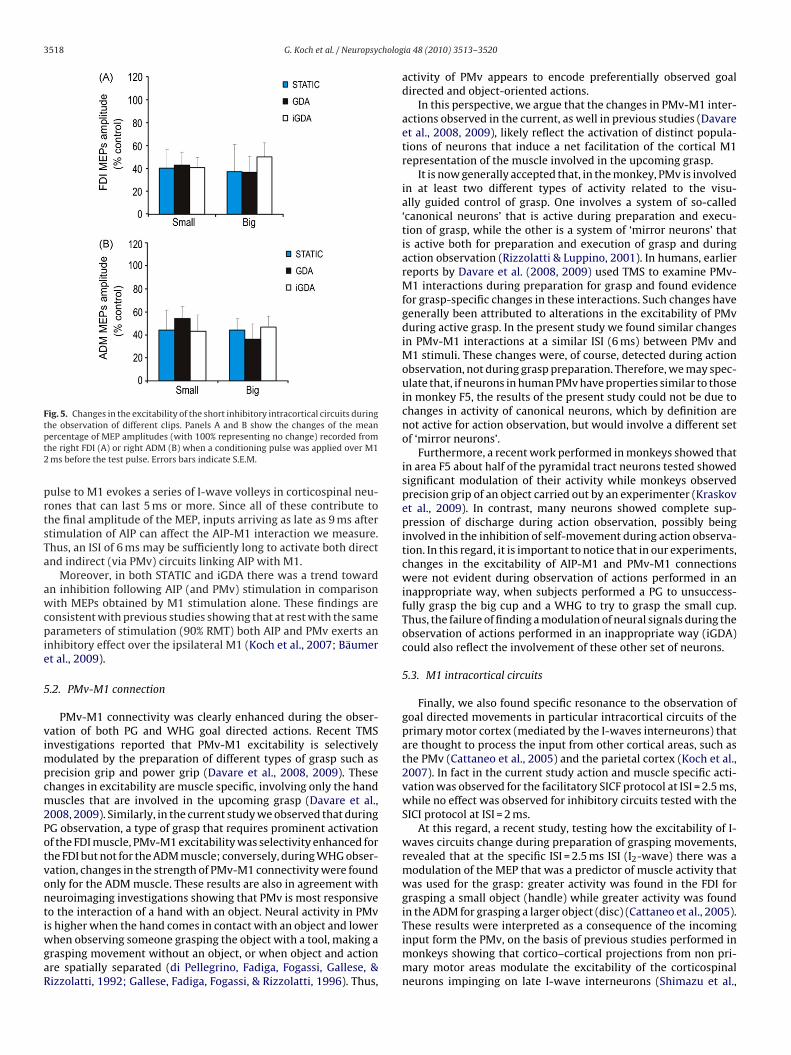
3518 G. Koch et al. / Neuropsycholog
Fig. 5. Changes in the excitability of the short inhibitory intracortical circuits duringtpt2
prtsTa
awcpie
5
vimpcm2PotvontiwgaR
he observation of different clips. Panels A and B show the changes of the meanercentage of MEP amplitudes (with 100% representing no change) recorded fromhe right FDI (A) or right ADM (B) when a conditioning pulse was applied over M1ms before the test pulse. Errors bars indicate S.E.M.
ulse to M1 evokes a series of I-wave volleys in corticospinal neu-ones that can last 5 ms or more. Since all of these contribute tohe final amplitude of the MEP, inputs arriving as late as 9 ms aftertimulation of AIP can affect the AIP-M1 interaction we measure.hus, an ISI of 6 ms may be sufficiently long to activate both directnd indirect (via PMv) circuits linking AIP with M1.
Moreover, in both STATIC and iGDA there was a trend towardn inhibition following AIP (and PMv) stimulation in comparisonith MEPs obtained by M1 stimulation alone. These findings are
onsistent with previous studies showing that at rest with the samearameters of stimulation (90% RMT) both AIP and PMv exerts an
nhibitory effect over the ipsilateral M1 (Koch et al., 2007; Bäumert al., 2009).
.2. PMv-M1 connection
PMv-M1 connectivity was clearly enhanced during the obser-ation of both PG and WHG goal directed actions. Recent TMSnvestigations reported that PMv-M1 excitability is selectively
odulated by the preparation of different types of grasp such asrecision grip and power grip (Davare et al., 2008, 2009). Thesehanges in excitability are muscle specific, involving only the handuscles that are involved in the upcoming grasp (Davare et al.,
008, 2009). Similarly, in the current study we observed that duringG observation, a type of grasp that requires prominent activationf the FDI muscle, PMv-M1 excitability was selectivity enhanced forhe FDI but not for the ADM muscle; conversely, during WHG obser-ation, changes in the strength of PMv-M1 connectivity were foundnly for the ADM muscle. These results are also in agreement witheuroimaging investigations showing that PMv is most responsiveo the interaction of a hand with an object. Neural activity in PMv
s higher when the hand comes in contact with an object and lowerhen observing someone grasping the object with a tool, making arasping movement without an object, or when object and actionre spatially separated (di Pellegrino, Fadiga, Fogassi, Gallese, &izzolatti, 1992; Gallese, Fadiga, Fogassi, & Rizzolatti, 1996). Thus,
ia 48 (2010) 3513–3520
activity of PMv appears to encode preferentially observed goaldirected and object-oriented actions.
In this perspective, we argue that the changes in PMv-M1 inter-actions observed in the current, as well in previous studies (Davareet al., 2008, 2009), likely reflect the activation of distinct popula-tions of neurons that induce a net facilitation of the cortical M1representation of the muscle involved in the upcoming grasp.
It is now generally accepted that, in the monkey, PMv is involvedin at least two different types of activity related to the visu-ally guided control of grasp. One involves a system of so-called‘canonical neurons’ that is active during preparation and execu-tion of grasp, while the other is a system of ‘mirror neurons’ thatis active both for preparation and execution of grasp and duringaction observation (Rizzolatti & Luppino, 2001). In humans, earlierreports by Davare et al. (2008, 2009) used TMS to examine PMv-M1 interactions during preparation for grasp and found evidencefor grasp-specific changes in these interactions. Such changes havegenerally been attributed to alterations in the excitability of PMvduring active grasp. In the present study we found similar changesin PMv-M1 interactions at a similar ISI (6 ms) between PMv andM1 stimuli. These changes were, of course, detected during actionobservation, not during grasp preparation. Therefore, we may spec-ulate that, if neurons in human PMv have properties similar to thosein monkey F5, the results of the present study could not be due tochanges in activity of canonical neurons, which by definition arenot active for action observation, but would involve a different setof ‘mirror neurons’.
Furthermore, a recent work performed in monkeys showed thatin area F5 about half of the pyramidal tract neurons tested showedsignificant modulation of their activity while monkeys observedprecision grip of an object carried out by an experimenter (Kraskovet al., 2009). In contrast, many neurons showed complete sup-pression of discharge during action observation, possibly beinginvolved in the inhibition of self-movement during action observa-tion. In this regard, it is important to notice that in our experiments,changes in the excitability of AIP-M1 and PMv-M1 connectionswere not evident during observation of actions performed in aninappropriate way, when subjects performed a PG to unsuccess-fully grasp the big cup and a WHG to try to grasp the small cup.Thus, the failure of finding a modulation of neural signals during theobservation of actions performed in an inappropriate way (iGDA)could also reflect the involvement of these other set of neurons.
5.3. M1 intracortical circuits
Finally, we also found specific resonance to the observation ofgoal directed movements in particular intracortical circuits of theprimary motor cortex (mediated by the I-waves interneurons) thatare thought to process the input from other cortical areas, such asthe PMv (Cattaneo et al., 2005) and the parietal cortex (Koch et al.,2007). In fact in the current study action and muscle specific acti-vation was observed for the facilitatory SICF protocol at ISI = 2.5 ms,while no effect was observed for inhibitory circuits tested with theSICI protocol at ISI = 2 ms.
At this regard, a recent study, testing how the excitability of I-waves circuits change during preparation of grasping movements,revealed that at the specific ISI = 2.5 ms ISI (I2-wave) there was amodulation of the MEP that was a predictor of muscle activity thatwas used for the grasp: greater activity was found in the FDI forgrasping a small object (handle) while greater activity was foundin the ADM for grasping a larger object (disc) (Cattaneo et al., 2005).
These results were interpreted as a consequence of the incominginput form the PMv, on the basis of previous studies performed inmonkeys showing that cortico–cortical projections from non pri-mary motor areas modulate the excitability of the corticospinalneurons impinging on late I-wave interneurons (Shimazu et al.,
cholog
2isi
MTtmtibwmccdtsptnsatsso
Spbtww
tAnp(iUiifiscrs
to
R
A
A
A
B
G. Koch et al. / Neuropsy
004; Prabhu et al., 2009). We therefore speculate that also dur-ng the observation of meaningful actions, the activation of theseet of interneurons is likely due to the processing of the incomingnput from PMv.
It has to be noticed that no specific modulation was found whenEPs by TS alone over M1 were analyzed in the different conditions.
his is partially in contrast with previous investigations reportinghat during the observation of a clip showing a reach to grasp move-
ent, there was an increase of MEP amplitude for the FDI musclehat was modulated by the amount of the observed finger aperture,ncreasing at the end of the reaching phase on comparison with theeginning (Gangitano et al., 2001). However, in the current studye focused in the critical late phase of reaching to grasp move-ents, and therefore we did not directly test these time-locked
hanges in excitability. Nevertheless other methodological issuesould explain this difference. First, it is possible that this could beue to the low number of trials per condition. In each conditionhere were 16 trials, of which only half (eight trials) were withingle pulses. Though this number can be enough for conditionedaradigms such as bifocal stimulation or paired-pulses (becausehe normalization to single pulses gets rid of noise), it is clearlyot enough for a noisy paradigm such as modulation of MEPs fromingle pulses. Second, subjects were performing a passive viewing,nd were not performing other concomitant tasks in order to con-rol for their attendance to the stimuli. In this case the power of thetimulus could have been reduced. Third, the clips were always theame in each condition and this could have reduced the reactivityf subjects to action observation.
Moreover we did not observe any specific changes occurring inICI circuits during action observation such as those reported in arevious study (Strafella & Paus, 2000). This discrepancy could alsoe due to the different experimental procedure adopted, since inhe study by Strafella and Paus (2000), clips were showing hand-riting and simple lateral arm movement while no grasping actionsere displayed.
To the best of our knowledge, we demonstrated for the firstime that in humans specific interactions occur between PMv andIP with M1 during action observation. Thus, these short-latencyeurophysiological connections become active not only duringlanning or execution of specific reaching to grasp movementsKoch & Rothwell, 2009a, 2009b; Koch et al., 2010), but also dur-ng the observation of goal directed actions (Rizzolatti et al., 2009).sing direct recording of extracellular neural activity in patients, it
s becoming possible to demonstrate mirroring spiking activity dur-ng action-execution and action observation in the human medialrontal cortex (Mukamel et al., 2010). Thus, recording neural activ-ty of single neurons in the same regions explored in the currenttudy, such as the lateral wall of the frontal cortex and the parietalortex could be important to investigate more in depth the neu-ophysiological interactions described here at the level of corticalystems.
Finally, we believe that the current findings could be relevanto extend neuro-rehabilitation approaches based on movementbservation in stroke patients (Rizzolatti et al., 2009).
eferences
glioti, S. M., Cesari, P., Romani, M., & Urgesi, C. (2008). Action anticipation and motorresonance in elite basketball players. Nature Neuroscience, 11, 1109–1116.
massian, V. E., & Stewart, M. (2003). Motor cortical and other cortical interneu-ronal networks that generate very high frequency waves. Supplements to ClinicalNeurophysiology, 56, 119–142.
venanti, A., Bolognini, N., Malavita, A., & Aglioti, S. M. (2007). Somatic and motorcomponents of action simulation. Current Biology, 17, 2129–2135.
äumer, T., Schippling, S., Kroeger, J., Zittel, S., Koch, G., Thomalla, G., et al. (2009).Inhibitory and facilitatory connectivity from ventral premotor to primary motorcortex in healthy humans at rest—A bifocal TMS study. Clinical Neurophysiology,120, 1724–1731.
ia 48 (2010) 3513–3520 3519
Begliomini, C., Wall, M. B., Smith, A. T., & Castiello, U. (2007). Differential corticalactivity for precision and whole-hand visually guided grasping in humans. TheEuropean Journal of Neuroscience, 25, 1245–1252.
Binkofski, F., Dohle, C., Posse, S., Stephan, K. M., Hefter, H., Seitz, R. J., et al. (1998).Human anterior intraparietal area subserves prehension: A combined lesion andfunctional MRI activation study. Neurology, 50, 1253–1259.
Buccino, G., Binkofski, F., Fink, G. R., Fadiga, L., Fogassi, L., et al. (2001). Action obser-vation activates premotor and parietal areas in a somatotopic manner: An fMRIstudy. The European Journal of Neuroscience, 13, 400–404.
Calvo-Merino, B., Glaser, D. E., Grèzes, J., Passingham, R. E., & Haggard, P. (2005).Action observation and acquired motor skills: An FMRI study with expertdancers. Cerebral Cortex, 15, 1243–1249.
Castiello, U. (2005). The neuroscience of grasping. Nature Reviews. Neuroscience, 6,726–736.
Cattaneo, L., Voss, M., Brochier, T., Prabhu, G., Wolpert, D. M., & Lemon, R. N. (2005).A cortico–cortical mechanism mediating object-driven grasp in humans. Pro-ceedings of the National Academy of Sciences of the United States of America, 102,898–903.
Cattaneo, L., Caruana, F., Jezzini, A., & Rizzolatti, G. (2009). Representation of goaland movements without overt motor behavior in the human motor cortex:A transcranial magnetic stimulation study. The Journal of Neuroscience, 29,11134–11138.
Cattaneo, L., Mandrini, M., & Schwarzbach, J. (2010). State-Dependent TMS revealsa hierarchical representation of observed acts in the temporal, parietal, andpremotor cortices. Cerebral Cortex [Epub ahead of print].
Clark, S., Tremblay, F., & Ste-Marie, D. (2004). Differential modulation of corticospinalexcitability during observation, mental imagery and imitation of hand actions.Neuropsychologia, 42, 105–112.
Davare, M., Lemon, R., & Olivier, E. (2008). Selective modulation of interactionsbetween ventral premotor cortex and primary motor cortex during precisiongrasping in humans. The Journal of Physiology, 586, 35–42.
Davare, M., Montague, K., Olivier, E., Rothwell, J. C., & Lemon, R. N. (2009). Ventralpremotor to primary motor cortical interactions during object-driven grasp inhumans. Cortex, 45, 1050–1057.
Di Lazzaro, V., Oliviero, A., Mazzone, P., Pilato, F., Saturno, E., Di leone, M., et al.(2003). Generation of I waves in the human: Spinal recordings. Supplements toClinical Neurophysiology, 56, 143–152.
di Pellegrino, G., Fadiga, L., Fogassi, L., Gallese, V., & Rizzolatti, G. (1992). Understand-ing motor events: A neurophysiological study. Experimental Brain Research, 91,176–180.
Ehrsson, H. H., Fagergren, A., Jonsson, T., Westling, G., Johansson, R. S., & Forssberg,H. (2000). Cortical activity in precision- versus power-grip tasks: An fMRI study.Journal of Neurophysiology, 83, 528–536.
Fadiga, L., Craighero, L., & Olivier, E. (2005). Human motor cortex excitability duringthe perception of others’ action. Current Opinion in Neurobiology, 15, 213–218.
Fadiga, L., Fogassi, L., Pavesi, G., & Rizzolatti, G. (1995). Motor facilitation duringaction observation: A magnetic stimulation study. Journal of Neurophysiology,73, 2608–2611.
Gallese, V., Fadiga, L., Fogassi, L., & Rizzolatti, G. (1996). Action recognition in thepremotor cortex. Brain, 119, 593–609.
Gangitano, M., Mottaghy, F. M., & Pascual-Leone, A. (2001). Phase-specific modu-lation of cortical motor output during movement observation. Neuroreport, 12,1489–1492.
Grafton, S. T., Arbib, M. A., Fadiga, L., & Rizzolatti, G. (1996). Localization of grasprepresentations in humans by positron emission tomography. 2. Observationcompared with imagination. Experimental Brain Research, 112, 103–111.
Iacoboni, M., Molnar-Szakacs, I., Gallese, V., Buccino, G., Mazziotta, J. C., & Rizzo-latti, G. (2005). Grasping the intentions of others with one’s own mirror neuronsystem. PLoS Biology, 3(3), e79.
Iacoboni, M., Woods, R. P., Brass, M., Bekkering, H., Mazziotta, J. C., et al. (1999).Cortical mechanisms of human imitation. Science, 286, 2526–2528.
Jeannerod, M., Arbib, M. A., Rizzolatti, G., & Sakata, H. (1995). Grasping objects: Thecortical mechanisms of visuomotor transformation. Trends in Neurosciences, 18,314–320.
Koch, G., Cercignani, M., Pecchioli, C., Versace, V., Oliveri, M., Caltagirone, C., et al.(2010). In vivo definition of parieto–motor connections involved in planning ofgrasping movements. Neuroimage, 51, 300–312.
Koch, G., Fernandez Del Olmo, M., Cheeran, B., Ruge, D., Schippling, S., Caltagirone,C., et al. (2007). Focal stimulation of the posterior parietal cortex increasesthe excitability of the ipsilateral motor cortex. The Journal of Neuroscience, 27,6815–6822.
Koch, G., Fernandez Del Olmo, M., Cheeran, B., Schippling, S., Caltagirone, C., Driver,J., et al. (2008). Functional interplay between posterior parietal and ipsilateralmotor cortex revealed by twin-coil transcranial magnetic stimulation duringreach planning toward contralateral space. The Journal of Neuroscience, 28,5944–5953.
Koch, G., Franca, M., Del Olmo, M. F., Cheeran, B., Milton, R., Alvarez Sauco, M.,et al. (2006). Time course of functional connectivity between dorsal premo-tor and contralateral motor cortex during movement selection. The Journal ofNeuroscience, 26, 7452–7459.
Koch, G., Oliveri, M., Cheeran, B., Ruge, D., Lo Gerfo, E., Salerno, S., et al. (2008).Hyperexcitability of parietal-motor functional connections in the intact left-hemisphere of patients with neglect. Brain, 131, 3147–3155.
Koch, G., & Rothwell, J. C. (2009a). TMS investigations into the task-dependent func-tional interplay between human posterior parietal and motor cortex. BehaviouralBrain Research, 202, 147–152.

3 cholog
K
K
K
M
M
M
M
O
O
520 G. Koch et al. / Neuropsy
och, G., & Rothwell, J. C. (2009b). Transcranial magnetic stimulation investigationsof reaching and grasping movements. In D. A. Novak, & J Hermsdorfer (Eds.),Sensorimotor control of grasping: Physiology and pathophysiology (pp. 72–83).Cambridge University Press.
raskov, A., Dancause, N., Quallo, M. M., Shepherd, S., & Lemon, R. N. (2009). Corti-cospinal neurons in macaque ventral premotor cortex with mirror properties:A potential mechanism for action suppression? Neuron, 64(6), 922–930.
ujirai, T., Caramia, M. D., Rothwell, J. C., Day, B. L., Thompson, P. D., Ferbert, A.,et al. (1993). Corticocortical inhibition in human motor cortex. The Journal ofPhysiology, 471, 501–519.
ars, R. B., Klein, M. C., Neubert, F. X., Olivier, E., Buch, E. R., Boorman, E. D., etal. (2009). Short-latency influence of medial frontal cortex on primary motorcortex during action selection under conflict. The Journal of Neuroscience, 29,6926–6931.
ukamel, R., Ekstrom, A. D., Kaplan, J., Iacoboni, M., & Fried, I. (2010). Single-neuronresponses in humans during execution and observation of actions. Current Biol-ogy,. April 7 [Epub ahead of print].
urata, A., Fadiga, L., Fogassi, L., Gallese, V., Raos, V., & Rizzolatti, G. (1997). Objectrepresentation in the ventral premotor cortex (area F5) of the monkey. Journalof Neurophysiology, 78, 2226–2230.
urata, A., Gallese, V., Luppino, G., Kaseda, M., & Sakata, H. (2000). Selectivity forthe shape, size, and orientation of objects for grasping in neurons of monkey
parietal area AIP. Journal of Neurophysiology, 83, 2580–2601.ldfield, R. C. (1971). The assessment and analysis of handedness: The Edinburghinventory. Neuropsychologia, 9, 97–113.
’Shea, J., Sebastian, C., Boorman, E. D., Johansen-Berg, H., & Rushworth, M. F. (2007).Functional specificity of human premotor–motor cortical interactions duringaction selection. The European Journal of Neuroscience, 26, 2085–2095.
ia 48 (2010) 3513–3520
Pobric, G., & Hamilton, A. F. (2006). Action understanding requires the left inferiorfrontal cortex. Current Biology, 16, 524–529.
Prabhu, G., Shimazu, H., Cerri, G., Brochier, T., Spinks, R. L., Maier, M. A., et al. (2009).Modulation of primary motor cortex outputs from ventral premotor cortex dur-ing visually guided grasp in the macaque monkey. The Journal of Physiology, 587,1057–1069.
Reynolds, C., & Ashby, P. (1999). Inhibition in the human motor cortex is reducedjust before a voluntary contraction. Neurology, 53(4), 730–735.
Rizzolatti, G., & Luppino, G. (2001). The cortical motor system. Neuron, 31, 889–901.Rizzolatti, G., Fogassi, L., & Gallese, V. (2001). Neurophysiological mecha-
nisms underlying action understanding and imitation. Nature Reviews, 2,661–670.
Rizzolatti, G., Fabbri-Destro, M., & Cattaneo, L. (2009). Mirror neurons and theirclinical relevance. Nature Clinical Practice: Neurology, 5, 24–34.
Shimazu, H., Maier, M. A., Cerri, G., Kirkwood, P. A., & Lemon, R. N. (2004). Macaqueventral premotor cortex exerts powerful facilitation of motor cortex outputs toupper limb motoneurons. The Journal of Neuroscience, 24, 1200–1211.
Strafella, A. P., & Paus, T. (2000). Modulation of cortical excitability duringaction observation: A transcranial magnetic stimulation study. Neuroreport, 11,2289–2292.
Urgesi, C., Candidi, M., Fabbro, F., Romani, M., & Aglioti, S. M. (2006). Motor facil-itation during action observation: Topographic mapping of the target muscle
and influence of the onlooker’s posture. The European Journal of Neuroscience,23, 2522–2530.Ziemann, U., Tergau, F., Wassermann, E. M., Wischer, S., Hildebrandt, J., & Paulus,W. (1998). Demonstration of facilitatory I wave interaction in the human motorcortex by paired transcranial magnetic stimulation. The Journal of Physiology,511, 181–190.





























