Embed Size (px)
Citation preview

3
Journal of Foraminiferal Research, v. 30, no. 1, p. 3–28, January 2000
REMARKS ON WEST PACIFIC NUMMULITIDAE (FORAMINIFERA)
JOHANN HOHENEGGER1, ELZA YORDANOVA1, AND AKIO HATTA2
ABSTRACT
Living Nummulitidae achieve their highest diversity inthe subtropical and tropical West Pacific. Although allhouse symbiotic microalgae, they avoid highly illumi-nated areas near the water surface, since their flat testscould be easily damaged by the hydrodynamic regime.The preference for calm water conditions extends theirdepth distribution down to the base of the photic zone.West Pacific Nummulitidae can be differentiated into tenspecies belonging to six genera according to an ecologicalspecies concept. The genus Operculina d’Orbigny is rep-resented by three species. While O. discoidalis(d’Orbigny) prefers a fine-grained bottom under medi-um light conditions (10% surface intensity), O. ammo-noides (Gronovius) prefers a coarser substrate andsometimes can be found on hard bottoms. Light depen-dence ranges from 1.5% to 68% surface intensity. Lessilluminated coarse sands are inhabited by Operculina cf.O. complanata (Defrance), which is the dominant sym-biont-bearing foraminifer between light intensities of0.2% to 12% surface illumination. The genus Planoste-gina Banner and Hodgkinson demonstrates transitionsto the genus Operculina in test form and surface, whilethe division into chamberlets is similar to Heterostegina.Planostegina operculinoides (Hofker) is distinguished byflat tests and delicate chamberlets. It lives on sandy bot-toms restricted to light intensities between 0.45% and26% surface illumination. The more robust Plansteginaaff. P. operculinoides (Hofker) prefers light intensitiesbetween 0.4% and 2.7% surface illumination. Planoper-culina heterosteginoides (Hofker) shows morphologicaltransition to Operculina cf. O. complanata in developingincomplete septula. This species lives in low illuminatedareas (0.3% to 2.5% surface intensity) and prefers me-dium to fine-grained sands. Heterostegina depressad’Orbigny spans a broad range in light intensities (2%to 70% surface illumination), and is protected againstirradiation by thick tests and a cryptic life mode nearthe surface. Test construction enables life under stronghydrodynamic regimes. This species lives firmly attachedto hard substrates, thus counteracting transportation bywater movement. Nummulites venosus (Fichtel and Moll)differs from H. depressa in having undivided chambers.It lives exclusively on coarse sand and avoids high sed-iment movement, thus starting its distribution beneaththe fair weather wave base. According to light intensi-ties, the upper limit may be similar to O. ammonoides(80%), while the lower limit is 2.5% surface illumina-tion. Operculinella cumingii (Carpenter) inhabits coarseto medium sand in deeper regions between 1.2% and25% surface illumination. Tests of the cyclic, large-sized
1 Institut fur Palaontologie, Geozentrum Universitat Wien, Althan-straße 14, A-1010 Wien, Austria.
2 Faculty of Education, Kagoshima University, Koritomo, Kagoshi-ma 890, Japan.
species Cycloclypeus carpenteri Brady are easily trans-ported due to the thin, plate-like form. The upper dis-tribution limit correlates with the storm wave base, re-stricting C. carpenteri to depths below 50 m. The lowerdistribution limit depends on light intensity and is lo-cated near the base of the photic zone (0.4% surfaceillumination).
INTRODUCTION
Members of the family Nummulitidae (de Blainville) arecharacterized by planispirally-coiled, multilocular tests.Each chamber wall, consisting of secreted low magnesiumcalcite, covers the total test including all former chambersduring the process of chamber construction; this leads tolamellar tests (e.g., Hansen 1979). Formation of new cham-bers was studied in detail on the living nummulitid Heter-ostegina depressa (Spindler and Rottger, 1974; Rottger andInstitut fur den Wissenschaftlichen Film, 1982). A speciallamination form (bilamellar tests) gives rise to a complexcanal system that is characteristic for all living and fossilnummulitids (e.g., Hottinger, 1977a).
Transparency of the calcareous, lamellar test walls en-ables light penetration, which is necessary for the photo-synthesizing symbiotic microalgae. All investigated livingnummulitids house diatoms exclusively, but each speciesharbors a mixture of different diatom species (e.g., Lee andothers, 1989; Lee and Anderson, 1991; Lee, 1994). Thisdependency on microalgae restricts the nummulitids to theeuphotic zone. Hyaline or glassy test regions can projectover the surface in form of interseptal piles or septal ele-vations, and are regarded as focusing mechanisms enablinglight collection in the deepest parts of the euphotic zone(Hottinger, 1997).
Members of this family became very abundant in youngergeological history. Starting in the early Paleogene, high evo-lutionary rates—reflected in complex and large tests—makethis group important index fossils for shallow marine pa-leoenvironments (Schaub, 1981; see reference list in Pig-natti, 1998). The shallow benthic zones of the Paleoceneand Eocene are based largely on nummulitids, especiallyrepresentatives of the genera Assilina and Nummulites (Ser-ra-Kiel and others, 1998). In the Oligocene and Neogene,genera such as Planostegina, Heterostegina, Spiroclypeus,and Cycloclypeus show evolutionary lines that are also use-ful for age determination of shallow warm-water sediments(Hottinger, 1977a; Banner and Hodgkinson, 1991; Drooger,1993), but these genera are less abundant and diverse thantheir Paleogene relatives.
Except the Caribbean species Heterostegina antillarum,which seems to be a synonym of H. depressa according tomolecular genetic investigation (Holzmann and others,1998), all nummulitid species live in the tropical Indo-Pa-cific. Highest diversity can be found in the West Pacific(Hallock 1984, Hohenegger 1994), but investigations onthese deep-dwelling organisms are rare. Nevertheless, com-parison of nummulitid diversities by workers using similar

4 HOHENEGGER, YORDANOVA, AND HATTA
FIGURE 1. Study areas in the West Pacific. 1. Amakusa Jima. 2.Sesoko Jima (location details in Hohenegger and others, 1999). 3. Sek-isei-Sho (location details in Ujiie and Hatta, 1995), 4. Belau (locationdetails in Hohenegger, 1996).
species concepts revealed differences between the Red Sea(Hottinger and others, 1993; Haunold and others, 1998) andthe Ryukyu Archipelago (Hatta and Ujiie, 1992; Hoheneg-ger, 1994). Four species—Operculina ammonoides, Heter-ostegina depressa, H. operculinoides, and Heterocyclinatuberculata—are characteristic in the Red Sea. The firstthree species can also be found in the Ryukyus, while theannular H. tuberculata is replaced by its homeomorph Cy-cloclypeus carpenteri. Nummulites venosus and Operculinacf. O. complanata are additional typical elements of theWest Pacific foraminiferal fauna. The northern and southerndistribution borders of nummulitids in the West Pacific areunclear. O. ammonoides is found in shallow (15 m), finesediments north of the Tokara Strait (Oki, 1989), which sep-arates the northern part of the Ryukyu Archipelago from theislands Tanegashima and Yaku located south of Kyushu, Ja-pan. This species is reported from Tanabe Bay at a northernlatitude of 338 429, where Heterostegina depressa is alsonoted (Uchio, 1962, 1968; Chiji and Lopez, 1968). Nothingis known about the geographical boundaries of deeper-dwelling nummulitids.
The distribution of West Pacific nummulitids is stronglydependent on the environmental factors of temperature, lightintensity, water energy, and substrate, which are non-line-arly correlated with the complex environmental gradient ofwater depth (Hohenegger, 1994; Hohenegger and others,1999). Depth dependence of larger foraminifers is mani-fested in two ways.
First, test flattening promoting light penetration is pro-vided by changing chamber proportions within one species,as is evident in the larger miliolids Amphisorus hemprichiiand Parasorites orbitolitoides (J. Hohenegger, personal ob-servation). Thinning of the lamellae is the primary mecha-nism of test flattening in hyaline species (Hallock and Han-sen, 1979; Hottinger, 1997). This is also coupled in somespecies with altered chamber proportions (e.g., Operculinaammonoides; Hottinger 1977b; Pecheux, 1995). Addition-ally, the surface area occupied by light-collecting mecha-nisms like pustules, interseptal piles or elevated sutures willincrease with depth in hyaline species with broad depthranges.
Second, the ranges of species can be restricted to smallintervals along the depth gradient, leading to a successionof closely related species (e.g., Amphistegina lessonii-group, Hansen and Buchardt, 1977). Such species keep theirtest variability small, but trends in test flattening can beobserved between species (e.g., from very thick lenticularAmphistegina lobifera to lenticular A. lessonii and thin len-ticular A. bicirculata; Larsen, 1976). These successions ofspecies along an environmental gradient are termed coeno-clines (Whittaker, 1973), and can be used for gradient de-termination based on species proportions in the form oftransfer equations (Hohenegger, 1995). Depth determinationbased on species proportions is useful for ancient environ-ments. Using taxonomic uniformitarianism (Dodd and Stan-ton, 1990), depth estimation by larger foraminifers is pos-sible only for younger geological ages, i.e., those not ex-ceeding the Upper Miocene, due to the high evolutionaryrates. The functional morphological approach may extendthis estimation of paleodepth to the phylogenetic start of
modern larger foraminifers in the basal Upper Cretaceous(Hottinger, 1978, 1983a, 1997).
Depth dependence of living Nummulitidae through en-largement of test size combined with flattening is found inOperculina ammonoides (Hottinger, 1977b; Pecheux, 1995)and Heterostegina depressa (Hottinger, 1977b) from theRed Sea. In the same region, Fermont (1977a,b) describedvarious depth trends in size parameters of the embryonicchambers in O. ammonoides and H. depressa. Althoughboth Fermont (1977a) and Pecheux (1995) noted two mor-phologically distinct groups in O. ammonoides distin-guished by evolute and involute tests occupying the samedepths in shallower areas, they hesitated to reflect these dis-tinctions by naming two species. On the contrary, both in-terpreted these differences as ecophenotypes of a single spe-cies, without, however, presenting any genealogical evi-dence. Hohenegger (1994, 1995, 1996), adhering to a sim-ilar species concept used by the Red Sea workers, founddepth successions of the five Northwest Pacific nummulitidspecies in the form of coenoclines.
10,215 living specimens were collected during the workon nummulitid distribution in the West Pacific (Fig. 1) usingsampling and investigation methods described in Hoheneg-ger (1994) and Hohenegger and others (1999). Ten morpho-logically distinct species could be detected based on sub-jective criteria. The high variability in test construction be-tween geographically isolated groups led in the past to the

5WEST PACIFIC NUMMULITIDAE
erection of numerous species, especially in the genus Oper-culina d’Orbigny; in fact, these merely represent geograph-ical variants. Two species—Heterostegina depressa and Cy-cloclypeus carpenteri—are morphologically clearly sepa-rated: they showed no transitions and could be easily rec-ognized. Thus, they represent good ‘ecological’ and also‘biological’ species, which are genealogically proven by re-production (Rottger and others, 1990; Kruger and others,1996). The other groups were separated into homogeneousmorphological classes based on subjective criteria, and sta-tistical proof will have to be provided in the future.
The following article concentrates on detecting consistentspecies within the Nummulitidae of the West Pacific withoutmaking a detailed systematic revision based on a statisticalinvestigation of populations. New aspects of morphometricinvestigation are introduced. This is followed by an expla-nation of ecological restrictions, especially relations of num-mulitids to the factors of hydrodynamic forces, light inten-sity, and substrate.
THE ECOLOGICAL SPECIES CONCEPT
Explaining depth dependencies of nummulitids eitherbased on test changes within a single species or based oncoenoclines depends on the species concept used. Detectingand explaining the distribution and habitats of living num-mulitids therefore requires a consistent species concept.Since the proof of genealogical connections between differ-ent morphogroups is complicated in larger foraminifers, dif-ficulties arise in using the ‘biological species concept’,which is based on sexual reproduction resulting in fertileoffspring (e.g., Mayr and Ashlock, 1991). A trimorphic cy-cle of generations is postulated for nummulitids, but thecomplete life cycle has been described only for H. depressa(Rottger and others, 1986, 1990). The interpretation of mor-phological homogeneity as an expression of a common genepool (e.g., Falconer, 1960; Hartl, 1980), which is the prac-tical aspect for recognizing biological species, is often bi-ased by ecological, seasonal, and generation variations re-sulting in distinct morphogroups (Mayr and Ashlock, 1991).Rottger and others (1986) regarded the schizonts of H. de-pressa as a new species as long as the genealogical con-nections to the agamonts were unknown (Rottger and others,1990).
Homogeneity of genealogically related organism groupsalong an ecological gradient is expressed as unimodal fre-quency distributions; this is used to determine the locationand range of the realized ecological niche (Roughgarden,1974) according to the environmental factor under consid-eration (Kendeigh, 1974). This ecological niche is the maincriterion of the ‘evolutionary species concept’ (Simpson,1951, 1961) by identifying the ‘own unitary evolutionaryrole’ (Wiley, 1981) with the ‘ecological niche’ (Hartl, 1980).An ‘ecological species’ can thus be defined as a ‘genealog-ically related group of organisms fitting an own particularecological niche in a biotic community’ (Grant, 1971 inpart; similar definition by van Valen, 1976).
Groups of genealogically related organisms that are dis-tinguished by continuous transitions of morphotypes havinga unimodal frequency distribution along an environmentalgradient can thus be interpreted as ecological variants of a
single species. One example is test flattening and increaseof papillae number with depth in Amphistegina radiata(Hohenegger, in press). In contrast, heterogeneity in fre-quencies along the gradient, expressed as multimodal dis-tributions in combination with continuously changing mor-phology, points to different species (e.g., Amphistegina les-sonii-species group; Hohenegger, in press).
Thus, morphology can also be used as the practical aspectfor recognizing and differentiating between closely related‘ecological species’. Morphologically slightly distinctgroups, which occupy identical or strongly overlapping eco-logical niches (i.e., the same distribution range along anenvironmental gradient), should be regarded as genealogi-cally related morphotypes of a single species as long asthese relationships remain unproven. When occupying sep-arate niches as expressed in multimodal frequency distri-butions along a gradient that are consistent within marinebiomes (Longhurst, 1998), morphologically slightly distinctgroups must be interpreted as different species according tothe ecological species concept mentioned above (for a com-prehensive discussion of the ecological species concept seeHohenegger, in press).
The generic position of most nummulitid species is alsoconfused (Hottinger, 1977a; Loeblich and Tappan, 1988;Banner and Hodgkinson, 1991), and needs revision. Clas-sification of genera and higher categories in the Nummuli-tidae is mainly based on morphology (Banner and Hodg-kinson, 1991) and does not directly represent phylogeneticrelationships. Hottinger (1977a) classified the nummulitidsin morphogroups, but did not assign taxonomic rank. Wemethodologically follow Hottinger in grouping the speciesaccording to test construction. After detailed examination ofphylogenetic relationships, these groups of genera may beestablished as subfamilies in future work.
INVOLUTE NUMMULITIDS
The main characteristic trait of the three species belong-ing to this morphogroup is the completely involute test,where chamber lumina cover the former test parts. Thisleads to thick lenticular tests in small individuals. In largespecimens, the chambers of the last whorl become flat andthin. In that case, they do not completely progress to theumbonal part. While one species belonging to this morpho-group—Heterostegina depressa—can be differentiated bythe division of chambers into chamberlets, morphologicaldistinction between the others—Nummulites venosus andOperculinella cumingii—is difficult and has led to taxo-nomic difficulties.
Detailed investigation on a large set of living individualsfrom the West Pacific (Fig. 1; 1304 specimens of N. veno-sus, 676 specimens of O. cumingii) revealed morphologicaldifferences that are supported by distinct habitat preferenc-es. While O. cumingii is characterized by large tests devel-oping an expansive, flat last whorl, N. venosus tests showsuch test construction only in very large specimens. Thesmaller sizes of the latter led to the interpretation of bothspecies as different generations, where the smaller formswere regarded as A-generation (gamonts or schizonts), andthe larger as B-generation or agamonts (Hofker, 1933; Hot-tinger, 1977a; Rogl and Hansen, 1984; Hohenegger, 1994).

6 HOHENEGGER, YORDANOVA, AND HATTA
FIGURE 2. Depth distributions of nummulitids with similar tests at two transects NW of Sesoko Island, Okinawa, Japan (location details inHohenegger and others, 1999). Fit of empirical data by power transformed normal distributions.
Frequency distributions along the depth gradient showclear differences between both ‘generations’. While postu-lated ‘megalospheres’ (N. venosus) are abundant at shallow-er depths, ‘agamonts’ (O. cumingii) dominate in the deeperregions, exceeding by far the number of ‘gamonts’ (Fig. 2).This is in extreme contrast to past experiences with fora-miniferal reproduction, where either gamonts predominate(.95%) in natural populations showing alternating genera-tions (Boltovskoy and Wright, 1976; Zohary and others,1980; Hallock, 1985; Pecheux, 1995) or agamonts are ab-sent in asexually reproducing generations (schizonts; Rott-ger and others, 1989, Fujita, 1997). Furthermore, bothgroups also differ in geographical distribution. The so-called‘agamonts’ could not be found in the western South Pacific(J-P. Debenay, personal communication), while ‘gamonts’(N. venosus, named Operculinella cumingii in Debenay,1988) are common.
Hanzawa (1939) first demonstrated morphological differ-ences between both species in picturing equatorial thin sec-tions of O. cumingii, all possessing large initial chambers
with a maximum diameter of 93 mm. The interpretation asagamonts thus became invalid. The author placed the small-er-sized forms, now determined as N. venosus, into the syn-onymy of Operculina ammonoides (Hanzawa, 1939, Pl.XV, figs. 4, 5). An open nomenclature (Operculina sp.) wasused by Hanzawa (1939, Pl. XV, figs. 9, 11) for a singleagamont of N. venosus that shows a microspheric proloculuswith 22 mm diameter.
Differences between the three involute nummulitids wereinvestigated in living species, all obtained from a singlesample taken at 50 m depth east of the Nakanose reef nearSesoko Island (Hohenegger and others, 1999). Thus, theyrepresent time-equivalent populations inhabiting a single lo-cation. The illustrated specimens of N. venosus in Pl. 1, figs.1–10 show different proloculus sizes. While two specimens(figs. 3–6) are characterized by large initial chambers (115mm and 131 mm), one specimen (Pl. 1, figs. 9, 10) that hasbegun to flatten the last whorl shows a small proloculus (55mm). The size of the initial chamber of the largest individualpictured in Pl. 1, figs 1, 2 could not be measured; it is

7WEST PACIFIC NUMMULITIDAE
PLATE 11–10 Nummulites venosus (Fichtel and Moll). 1, 9 agamonts, photomicrographs. 2, 10 agamonts, X-ray micrographs. 3, 5, 7 megalospheres,
photomicrographs. 4, 6, 8 megalospheres, X-ray micrographs. 11–18 Operculinella cumingii (Carpenter). 11 agamont, photomicrograph. 12 agamont,X-ray micrograph. 13, 15, 17 megalospheres, photomicrographs. 14, 16, 18 megalospheres X-ray micrographs. All specimens from 50 m at NakanoseReef (Hohenegger and others, 1999), 3 12.

8 HOHENEGGER, YORDANOVA, AND HATTA
FIGURE 3. Growth of the test margin in dependence of the revolution angle. Broken lines indicate normal exponential growth, while full linesmark the modified growth function (formula 2) with changing expansion rate (function parameters in Table 1).
interpreted as an agamont because it is completely similarin size and shape to Operculina sp. as figured in Hanzawa(1939, Pl. XV, figs. 9, 11).
Proloculus size also varies in O. cumingii specimens (Pl.1, figs. 11–18). While three individuals (figs. 13–18) havesimilar diameters (116 mm, 115 mm, and 98 mm), the initialchamber of the remaining specimen (figs. 11, 12) is signif-icantly smaller (38 mm). Interpreting the latter as an agam-ont of O. cumingii, the generation differences in test sizebetween gamonts (?schizonts) and agamonts seem to be lesspronounced than in N. venosus or H. depressa, since onefigured gamont (?schizont; Pl. 1, figs. 17, 18) is much largerthan the agamont.
Morphological differences between N. venosus and O.cumingii are primarily based on four character groups. Bothspecies are distinguished by planispirally-coiled tests. The
course of the test margin can thus be modeled by a loga-rithmic function of the form
r 5 r0b1a (1)
where r represents the distance from the center of the pro-loculus to the outer margin. The first constant (r0) of equa-tion (1) characterizes the initial radius at the revolution an-gle a 5 0 (measured in radians) and the constant b1 marksthe expansion rate (Raup, 1966). This parameter is the mainform factor of a logarithmic spiral, since it determines thegrade of expansion during revolution around the origin.Small-sized specimens of all three involute species that lacka flattened final part can significantly be fitted using thislogarithmic function (Fig. 3). The expansion rate of N. ven-osus is small (mean 5 1.07), leading to low chamber heightsthat slowly increase during growth (Pl. 1, figs. 4, 6, 8). O.

9WEST PACIFIC NUMMULITIDAE
cumingii tests are distinguished by significantly larger ex-pansion rates (mean 5 1.11; Pl. 1, figs. 12, 14, 16), resultingin higher whorls than in N. venosus. The largest expansionrate b1 (mean 5 1.12) is found in the smaller tests or ininitial parts of H. depressa (Pl. 4, figs. 2, 4), but differencesin comparison to O. cumingii are insignificant.
Large tests of all three species positively deviate in theirfinal parts from the ideal logarithmic spiral. This deviationis coupled with strong test flattening. Growth can be mod-eled in this test part by the following modified logarithmicspiral with changing expansion rate b 5 b2 1 ca, yielding
r 5 r0 (b2 1 ca)a. (2)
N. venosus demonstrates clear differences between genera-tions in test growth (Table 1; Fig. 3). Gamonts (?schizonts)follow the logarithmic function, while spirals of the agam-onts (Pl. 1, figs. 2, 10) can be fitted significantly by themodified function (2). Differences in growth between gen-erations could also be detected for O. cumingii (Table 1;Fig. 3). The contours of megalospheric individuals (e.g., Pl.1, fig. 18) are best fitted by the modified growth function(Table 1; Fig. 3). All investigated specimens of H. depressarepresent a single generation of megalospheres (gamonts orschizonts). They demonstrate optimal fit by the modifiedlogarithmic function (Table 1; Fig. 3). Therefore, test growthin megalospheric generations of O. cumingii and H. de-pressa is similar. Differences between N. venosus and O.cumingii again are manifested in agamonts by the form pa-rameter c of the modified growth function (Table 1), whereN. venosus (c 5 0.0003) significantly differs from O. cum-ingii (c 5 0.0012). Therefore, the expansion of the flat lastwhorl is much stronger in the latter than in the former spe-cies.
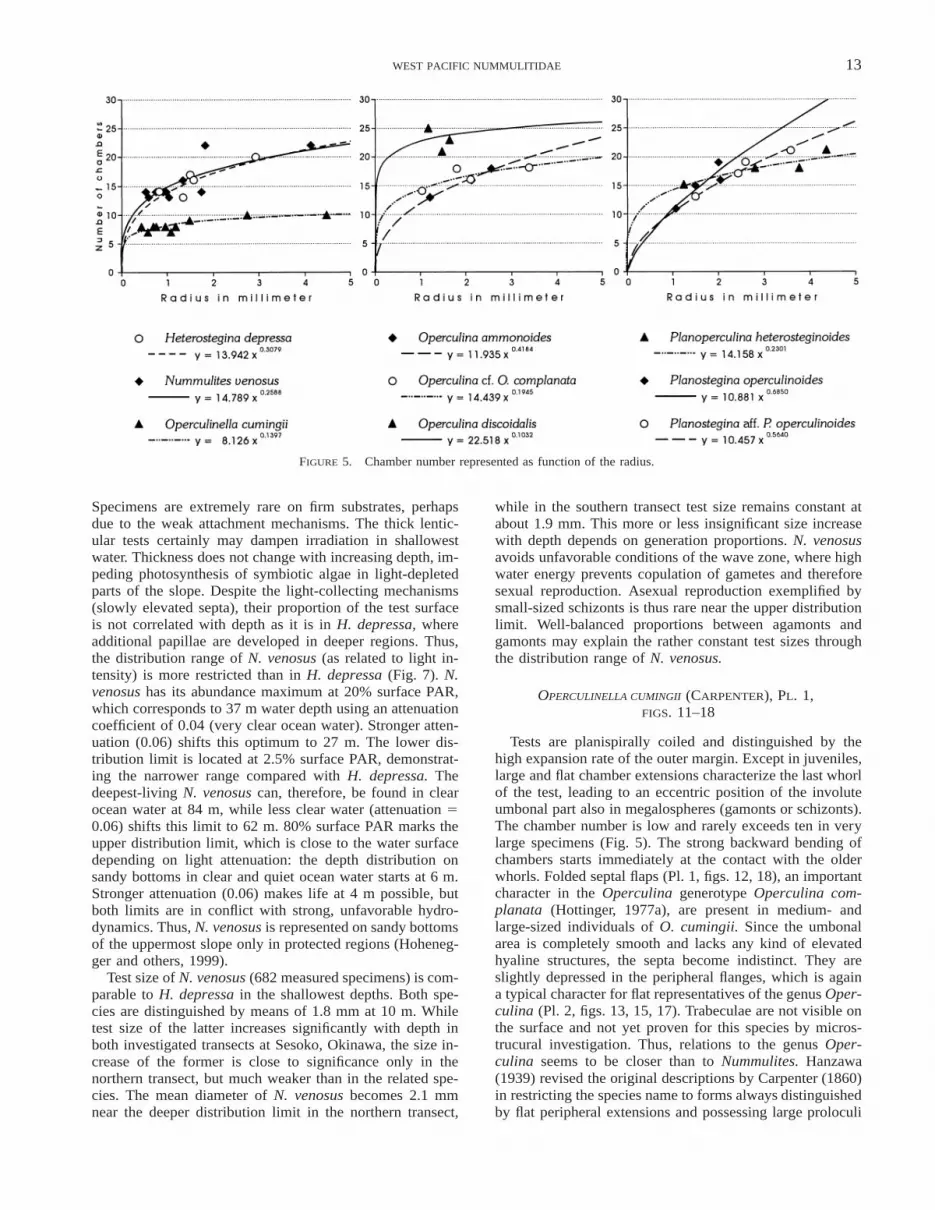
The second characteristic morphological trait for differ-entiating the three involute nummulitids is the chambernumber within one whorl. This character changes with sizeand thus depends on the radius. While O. cumingii and H.depressa are similar in spiral form and N. venosus differsfrom both by low whorl heights, O. cumingii is now sepa-rated from both species by a significantly lower chambernumber. Measurements on the last and penultimate whorlsof the illustrated specimens in Plates 1 and 4 yielded sig-nificant regressions between radii and chamber numbers forthe three species (Fig. 5). The chamber number increases inO. cumingii from 7 at a radius of 0.5 mm to 10 in whorlspossessing a final radius of 4 mm. N. venosus starts with12 chambers at 0.5 mm radius. A strong increase leads toan average of 21 chambers in whorls with a 4 mm radius,which is more than twice the number of O. cumingii. Cham-ber numbers in the first whorls of H. depressa are similarto N. venosus, starting with 11 chambers at 0.5 mm andattaining 21 chambers per whorl with a 4 mm final radius.
Chamber form is the third morphological character thatallows differentiation between the three involute nummuli-tids. Hanzawa (1939) again recorded differences between N.venosus and O. cumingii. The septa of N. venosus remainstraight in equatorial section until a region located betweenthe half and the second third of chamber height, and thenturn backwards (Pl. 1, figs. 2, 4, 6, 8). These curved partsare short and can be characterized by the angle between twovectors with a common origin situated in the center of the
proloculus. The endpoint of the first vector is the interio-marginal onset of the preceding chamber septum, while thesecond vector connects the origin with the contact pointbetween the septum and the preceding chamber (Fig. 6).This angle is constant (mean 5 258) in N. venosus (Pl. 1,figs. 4, 6, 8) and does not significantly increase with size.It never exceeds 458 in agamonts (Fig. 6; Pl. 1, figs. 2, 10).In small O. cumingii the interval where radial septa start toturn backwards is located within the first half of the equa-torial chamber height (Pl. 1, figs. 14, 16). This region shiftswith increasing size to the interiomarginal onset of the sep-tum; thus, chambers immediately bend backwards in flatextensions of the last whorl, starting with their contact tothe older test (Pl. 1, figs. 12, 18). The angle that describeschamber bending is a function of size in O. cumingii. Thisparameter starts with a minimum of 308 in the first whorlsand rapidly increases up to a maximum of 1608 in final partsof large specimens (Fig. 6). H. depressa is distinguished bysimilar chamber forms as O. cumingii, also increasing withsize (minimum 5 308), but its narrower chambers lead toinsignificantly smaller angles (maximum 5 1408) character-izing the backward bending chamber parts in large speci-mens.
The septa and their courses on the umbonal area representthe last characteristic traits for species differentiation usinglight-optical microscopes. While septa are hyaline and ele-vated in N. venosus and H. depressa, they can be recog-nized in O. cumingii only by their slight depressions in theexpanded part of final whorls (Pl. 1, figs. 11, 17), or in smallspecimens (Pl. 1, fig. 15). Septa of the alar prolongationsare always visible and of sigmoidal form both in H. de-pressa and N. venosus. The straight courses in the umbonalarea of O. cumingii tests can be detected only by X-rayinvestigation (Pl. 1, figs. 14, 16, 18), since optically indis-tinct septa are typical for the lenticular test part, as men-tioned first in Hanzawa (1939).
These three species of involute, West Pacific nummulitidswill briefly be treated with regard to their morphology, ge-neric position, biology, and habitat.
HETEROSTEGINA DEPRESSA D’ORBIGNY, PL. 4, FIGS. 1–6
Thick involute tests are characteristic for juveniles andsmaller-sized individuals. Peripheral flattening of involutechambers is found in large specimens, especially agamonts.Septula completely divide the chambers into chamberlets.The undivided chamber extensions on the involute part ofthe older test are of sigmoidal form. Raised hyaline suturesare the only surface ornaments in the shallow-living speci-mens, and flat papillae become additional ‘light collecting’structures in the deeper-living forms. The tests lack aper-tures; a complex channel system with a marginal cord, in-vestigated in detail by Hottinger (1977a) and Spindler(1978), enable direct connections between chamberlets andthe protoplasm outside the test. A detailed description oftest morphology as well as the synonymy list can be foundin Hottinger and others (1993). Starting with the work ofLutze and others (1971), this species is the best-studied larg-er foraminifer in respect to biology, especially growth, nu-trition, and reproduction, which is based on the huge workof Rottger and coworkers (Dettmering and others, 1998;

10 HOHENEGGER, YORDANOVA, AND HATTA
PLATE 21–6 Operculina discoidalis (d’Orbigny). 1, 3, 5 photomicrographs. 2, 4, 6 X-ray micrographs, specimens from 19 m in the lagoon NE of Sesoko
Island (Hohenegger and others, 1999). 7–12 Operculina ammonoides (Gronovius). 7, 9, 11 photomicrographs. 8, 10, 12 X-ray micrographs,specimens from 30 m in the transect NW of Sesoko Island (Hohenegger and others, 1999). 13–18 Operculina cf. O. complanata (Defrance). 13,15, 17 photomicrographs. 14, 16, 18 X-ray micrographs, specimens from 90 m in the transect NW of Sesoko Island. 312.

11WEST PACIFIC NUMMULITIDAE
TABLE 1. Function parameters of the logarithmic spirals according to formulas 1 and 2.
Initial radius r0
Normal logarithmicspiral expansion
rate b1
Modified logarithmicspiral expansion rate b2 Changing factor c
Heterostegina depressaNummulites venosusNummulites venosus, agamontOperculinella cumingiiOperculinella cumingii, agamont
0.1280.2210.0260.1320.034
1.1261.0711.0961.1111.145
1.0911.0711.0791.0651.113
0.001900.00030.00280.0012
Operculina ammonoidesOperculina cf. O. complanataOperculina discoidalisPlanoperculina heterosteginoidesPlanostegina operculinoidesPlanostegina aff. P. operculinoides
0.3270.1410.4590.1220.0680.205
1.1381.2091.1011.2561.2511.207
1.1051.1671.1011.1731.1771.153
0.00320.003400.00790.00680.0051
Kruger, 1994; Rottger, 1972a,b, 1973, 1974, 1976, 1990;Rottger and Berger, 1972; Rottger and Inst.Wiss.Film, 1982,1984; Rottger and Spindler, 1976; Rottger and others, 1980,1984, 1986, 1989, 1990, 1998).
In the West Pacific, H. depressa is the only nummulitidthat lives in shallow pools on the reef crest, although spec-imen numbers are low (Kuwano, 1956; Rottger, 1976; Ho-henegger, 1994; Hohenegger and others, 1999). Highest in-dividual numbers are found on hard substrates in upper partsof the reef slope (Hallock, 1984; Debenay, 1988; Hoheneg-ger, 1994, 1996; Hohenegger and others, 1999). They attachto coral rubble or reef rock, where they hide in smallgrooves that protect against water energy and irradiation,which is extreme in the upper wave zone. H. depressa isless common on coarse sand, but it becomes abundant onthis substrate in deeper habitats, where larger componentsfor attachment (e.g., macroids; Hottinger, 1983b) are rare.
The frequency optimum and both the upper and lowerdistribution limits depend primarily on light intensity, whichis necessary for photosynthesis of symbiotic algae. Labo-ratory investigations at constant light intensities (illumina-tion time 12 h per day) revealed optimum growth between7.81 and 11.07 mE m22 s21 (Rottger and others, 1980). Thiscorrelates with the growth optima for its symbiotic diatomsat 8.78 mE m22 s21 (Lee and others, 1982). Since light in-tensities in nature are not constant during daytime and alsochange with solar altitude or weather conditions dependingon season, a comparison and correlation between laboratorycultures and natural habitats is difficult. Thus, photobiolo-gists prefer the percentage of surface photosynthetic activeradiation (PAR; e.g., Titlyanov and Latypov, 1991) for mea-suring light intensity. This measure is independent of day-time, weather, or season. Using this measure, H. depressashows an abundance maximum at 31% surface irradiation(Fig. 7). Considering an attenuation coefficient of 0.04,which is typical for very clear ocean water as experiencedat the transect W off Sesoko Island, Okinawa (Hoheneggerand others, 1999), this maximum corresponds with 27 mwater depth. A slightly stronger attenuation of 0.06, asfound in the second transect NW off Sesoko Island (Hohen-egger and others, 1999), shifts this maximum upwards to 19m. The lower distribution limit is located at 2% surfacePAR, extending the depth distribution of H. depressa invery clear ocean water (attenuation coefficient 5 0.04) to89 m. Again, slightly stronger attenuation (0.06) shifts thislimit to 65 m. On the other side, H. depressa resists extreme
surface irradiation on the reef crest and in the uppermostmeters of the slope by protecting itself beneath coral rubbleor in tiny grooves of the reef rock (Rottger, 1976).
Test size increases significantly with depth, starting witha mean value of 1.4 mm at the surface. Statistical investi-gation is based on 1391 living individuals from the twotransects NW off Sesoko Island, Okinawa (Hohenegger andothers, 1999). Both regression lines are nearly identical, thusmanifesting the depth trend in increasing size, ending withsizes of 3.5 mm at the lower distribution limit (Yordanova,1998). A similar trend was documented in H. depressa fromHawaii (Rottger and Hallock, 1982), but not confirmed sincethe data came from different sites and samples were takenat different times through the year. All living individualsfrom Okinawa were collected within two months (June andJuly 1996) at two depth transects, thus representing similargrowth stages. An explanation of the significant size in-crease can be given by the trimorphic generation cycle ofH. depressa. Asexually reproducing schizonts dominate inshallow, high-energy waters, because the large juveniles areat lower risk (e.g., Leutenegger, 1977b; Rottger and others,1990). Small-sized zygotes that are less protected againstturbulence prefer calm regions below the fair-weather base.Gamonts and schizonts of H. depressa do not differ in pro-loculus size, but the tests of the first are significantly largerthan the latter (Rottger and others, 1990). The higher pro-portion of gamonts in combination with the few, but gigan-tic, agamonts in the deeper parts of the distribution rangemay cause the significant increase in test size.
NUMMULITES VENOSUS (FICHTEL AND MOLL), PL. 1,FIGS. 1–10; PL. 4, FIG. 10
The planispirally-coiled tests are completely involute insmall- and medium-sized individuals with low chamberheight, which is caused by low expansion rates of the log-arithmic spirals (Table 1; Fig. 3). Thus, the course of thetest margin becomes similar to a ‘spiral of Archimedes’(Batschelet, 1971). Alar prolongations incompletely coverthe older test in large specimens, where chambers of the lastwhorls start flattening. These parts are pronounced in thefew ‘adult’ agamonts, while only large megalospheres dem-onstrate slight marginal flattening in the last whorl that leadsto a keel-like structure. The test surface is smooth, and el-evated hyaline sutures are always visible. On the umbonaltest part, sigmoidal and sometimes branched courses of sep-

12 HOHENEGGER, YORDANOVA, AND HATTA
FIGURE 4. Proportion of involute and semi-involute test parts in the Operculina individuals figured in Pl. 2. Initial whorl starts at radians 5 0;whorl length 5 6.282 radians.
ta are caused by the high chamber number within one whorl(.12), which is a typical character for the genus Nummu-lites (Fig. 5). Interiomarginal foramina allow connectionsbetween chambers, and the canal system with a thick mar-ginal cord opens passages between chambers and the exter-nal test. Transverse trabeculae that are diagnostic for thegenus Nummulites mark the branches of sutural canals onthe surface (Pl. 4, fig. 10). A detailed description of testmorphology and the canal system can be found in Hottinger(1977a) and in the description of the holotype by Rogl andHansen (1984).
The biology of N. venosus is less known than that of H.
depressa, but reproduction of agamonts (Kruger, 1994) aswell as gamonts (Rottger and others, 1998) has been ob-served in laboratory cultures. Multiple fission is very similarto processes found in H. depressa (Kruger, 1994). The lon-gevity of one gamont of N. venosus that released triflagel-lated gametes was 569 days (Rottger and others, 1998),which is three times the life span of H. depressa gamonts(Kruger, 1994).
The main factors determining distribution boundaries ofN. venosus are light intensity, substrate, and hydrodynamics.This species prefers coarse sand, but avoids high-energy re-gions of the wave zone (Hohenegger and others, 1999).
13WEST PACIFIC NUMMULITIDAE
FIGURE 5. Chamber number represented as function of the radius.
Specimens are extremely rare on firm substrates, perhapsdue to the weak attachment mechanisms. The thick lentic-ular tests certainly may dampen irradiation in shallowestwater. Thickness does not change with increasing depth, im-peding photosynthesis of symbiotic algae in light-depletedparts of the slope. Despite the light-collecting mechanisms(slowly elevated septa), their proportion of the test surfaceis not correlated with depth as it is in H. depressa, whereadditional papillae are developed in deeper regions. Thus,the distribution range of N. venosus (as related to light in-tensity) is more restricted than in H. depressa (Fig. 7). N.venosus has its abundance maximum at 20% surface PAR,which corresponds to 37 m water depth using an attenuationcoefficient of 0.04 (very clear ocean water). Stronger atten-uation (0.06) shifts this optimum to 27 m. The lower dis-tribution limit is located at 2.5% surface PAR, demonstrat-ing the narrower range compared with H. depressa. Thedeepest-living N. venosus can, therefore, be found in clearocean water at 84 m, while less clear water (attenuation 50.06) shifts this limit to 62 m. 80% surface PAR marks theupper distribution limit, which is close to the water surfacedepending on light attenuation: the depth distribution onsandy bottoms in clear and quiet ocean water starts at 6 m.Stronger attenuation (0.06) makes life at 4 m possible, butboth limits are in conflict with strong, unfavorable hydro-dynamics. Thus, N. venosus is represented on sandy bottomsof the uppermost slope only in protected regions (Hoheneg-ger and others, 1999).
Test size of N. venosus (682 measured specimens) is com-parable to H. depressa in the shallowest depths. Both spe-cies are distinguished by means of 1.8 mm at 10 m. Whiletest size of the latter increases significantly with depth inboth investigated transects at Sesoko, Okinawa, the size in-crease of the former is close to significance only in thenorthern transect, but much weaker than in the related spe-cies. The mean diameter of N. venosus becomes 2.1 mmnear the deeper distribution limit in the northern transect,
while in the southern transect test size remains constant atabout 1.9 mm. This more or less insignificant size increasewith depth depends on generation proportions. N. venosusavoids unfavorable conditions of the wave zone, where highwater energy prevents copulation of gametes and thereforesexual reproduction. Asexual reproduction exemplified bysmall-sized schizonts is thus rare near the upper distributionlimit. Well-balanced proportions between agamonts andgamonts may explain the rather constant test sizes throughthe distribution range of N. venosus.
OPERCULINELLA CUMINGII (CARPENTER), PL. 1,FIGS. 11–18
Tests are planispirally coiled and distinguished by thehigh expansion rate of the outer margin. Except in juveniles,large and flat chamber extensions characterize the last whorlof the test, leading to an eccentric position of the involuteumbonal part also in megalospheres (gamonts or schizonts).The chamber number is low and rarely exceeds ten in verylarge specimens (Fig. 5). The strong backward bending ofchambers starts immediately at the contact with the olderwhorls. Folded septal flaps (Pl. 1, figs. 12, 18), an importantcharacter in the Operculina generotype Operculina com-planata (Hottinger, 1977a), are present in medium- andlarge-sized individuals of O. cumingii. Since the umbonalarea is completely smooth and lacks any kind of elevatedhyaline structures, the septa become indistinct. They areslightly depressed in the peripheral flanges, which is againa typical character for flat representatives of the genus Oper-culina (Pl. 2, figs. 13, 15, 17). Trabeculae are not visible onthe surface and not yet proven for this species by micros-trucural investigation. Thus, relations to the genus Oper-culina seems to be closer than to Nummulites. Hanzawa(1939) revised the original descriptions by Carpenter (1860)in restricting the species name to forms always distinguishedby flat peripheral extensions and possessing large proloculi

14 HOHENEGGER, YORDANOVA, AND HATTA
FIGURE 6. Chamber form represented as the angle of backward bend. This form parameter is a function of the revolution angle. Note differentscale intervals along the axes of revolution angle indicating the whorl number (length of one whorl 5 6.282 radians, initial whorl starts at radians5 0).

15WEST PACIFIC NUMMULITIDAE
FIGURE 7. Dependence of nummulitid species on light intensities measured in percentages of photosynthetic active radiation (PAR). Theoreticalfrequency distributions as power transformed normal curves are obtained by fitting the empirical data (Hohenegger, in press). Note the right-sideskewness of most functions. The corresponding depths to light intensities with different attenuation coefficients (0.04 and 0.06) are typical for cleartropical water.

16 HOHENEGGER, YORDANOVA, AND HATTA
PLATE 31–6 Planostegina aff. P. operculinoides (Hofker). 1, 3, 5 photomicrographs. 2, 4, 6 X-ray micrographs. 7–12 Planostegina operculinoides
(Hofker). 7, 9, 11 photomicrographs. 8, 10, 12 X-ray micrographs. 13–18 Planoperculina heterosteginoides (Hofker). 13, 15, 17 photomicrographs.14, 16, 18 X-ray micrographs. All specimens from 90 m in the transect NW of Sesoko Island (Hohenegger and others, 1999), 310.

17WEST PACIFIC NUMMULITIDAE
(Carpenter, 1860, Pl. 5, figs. 16–17). He used the genusname Operculinella as introduced by Yabe (1918) in de-scribing these recent nummulitids that are homeomorphs ofthe Paleogene Nummulites possessing smooth, involute, andthick lenticular tests.
The biology of these deep-dwelling organisms is com-pletely unknown. Like N. venosus, this species preferscoarse sands as substrate. O. cumingii did not demonstratetest flattening in the umbonal part with increasing depth,again a parallelism to the morphological relative N. venosus.Since the distribution range of the former is located deeperthan that of the latter (Fig. 2), light supply becomes difficultto obtain for the symbiotic microalgae in those regions. Thismay explain the extreme dimensions of the thin and flatperipheral extensions in both micro- and megalospheres,which enables penetration of photons for the symbionts. O.cumingii has its distribution optimum at 4.5% surface PAR(Fig. 7), corresponding to 70 m in clear ocean water (atten-uation coefficient 5 0.04). Less clear water (attenuation 50.06) shifts this maximum to 52 m. The lower distributionlimit at 1.2% surface PAR coincides with 100 m depth inclear water and 74 m in less clear water. The upper limit islocated at 25% surface PAR. Thus, shallowest O. cumingiican be detected on sandy bottoms at 31 m in very clearwater, while stronger attenuation (0.06) enables onset at 23m. An attenuation coefficient of 0.1 shifts this limit to 13m, which is inconvenient for the large and flat O. cumingii,since they will be transported and destroyed by high waveenergy.
Mean test size of O. cumingii (200 measured individuals)is constant through its distribution range. No significant re-lations to water depth could be detected at either transectinvestigated. While in the first transect the mean sizes re-main constant (varying around a 4 mm mean diameter), onlythe shallowest samples in the second transect show suchlarge sizes. The deeper samples are characterized by smallertests with a mean diameter of 2.5 mm. Constant test sizesagain are an argument for well-balanced proportions be-tween agamonts and gamonts, whereby a trimorphic gen-eration cycle with a sequence of schizonts need not neces-sarily be assumed.
SEMIINVOLUTE AND EVOLUTE NUMMULITIDSWITHOUT CHAMBER PARTITIONS
Besides the genus Heterostegina, nummulitids with un-divided chambers and semiinvolute to evolute tests are thebest-documented living forms. Semiinvolute coiling is oftencombined with thick tests, while evolute coiling correspondsto flat tests. Most initial whorls of thick tests are completelyinvolute. The first typical traits that characterize this groupare hyaline septa without trabeculae, which enable a cleardifferentiation from the porous chamber walls. Septa aresometimes slightly elevated in the form of papillae that in-crease in size and number with age and become knobs.These linearly arranged surface structures follow the septalcourses and sometimes get large in size (bosses). The num-ber of knobs increases with depth, leading to the interpre-tation as light-focusing mechanisms for symbiotic microal-gae in deeper water (e.g., Hottinger, 1997).
High variability in test form and size reflects both eco-
logical factors and geographical differences. Using a mor-phological species concept, numerous species were de-scribed in the past, leading to confusion in the taxonomy ofliving and fossil forms (see synonymy lists for Operculinaand Planoperculina in Hottinger, 1977a). Furthermore, thegeneric position is unclear. According to the modern statusof genus classification in Loeblich and Tappan (1988), thishomogeneous species group has to be subdivided into thegenera Assilina d’Orbigny and Operculina d’Orbigny. Onlytwo test characters differentiate the two genera. First, thegenus Operculina is characterized by folded septal flaps,which are smooth in Assilina. Second, the arrangement ofstolons that connect the chambers is irregular in Assilinaand linear in Operculina. The latter character depends ontest thickness and is subsequently correlated with the widthof the septal area. Changes in stolon arrangement can bedemonstrated for the species ‘Assilina’ ammonoides, whereshallow-living specimens with thick tests possess wide sep-tal areas and irregularly arranged stolons. Increasing depthleads to test flattening (Hottinger, 1977b): septal flaps be-come narrow inducing a linear arrangement of stolons.Therefore, only the folding of septal flaps remains as thegenerotypic character for Operculina—a weak character forthe differentiation of genera. Septa of initial whorls aresmooth in living representatives of the genus Operculina,for example Operculina cf. O. complanata (Cushman,1914), and they become folded in the last whorls of largespecimens (Pl. 2, figs. 14, 16). This is also significant forfossil representatives of O. complanata, which representsthe generotype, where even large specimens lack septalfolds (Hottinger, 1977a, Fig. 39). Thus, differentiation be-tween the genera Assilina and Operculina is impossible,especially in small-sized individuals with evolute tests.Since this character is weak for generic differentiation, thename Operculina d’Orbigny 1826 has priority over Assilinad’Orbigny 1839 at least in classifying living members ofthis group.
Even though two distinct morphotypes of Operculina arefound in the Red Sea (Hottinger, 1977b; Reiss and Hottin-ger, 1984; Haunold and others, 1998), both were regardedas ecophentotypes of a single species (O. ammonoides).This opinion is based on more or less continuous morpho-logical transitions from thick semiinvolute specimens to flatevolute individuals, yet with rare transitional forms (Pe-cheux, 1995). The thick specimens prefer shallow, fine sub-strates, and evolute individuals mainly live on coarse sand.Fermont (1977a) and Pecheux (1995) interpreted these eco-phenotypes as caused by differences in feeding or symbiontcomposition. O. ammonoides thus occupies a broad envi-ronmental range. Two additional species in the West Pacificare clearly separated from O. ammonoides by morphologi-cal differences. The first, Operculina cf. O. complanata,differs in having evolute tests with folded septal flaps andwas often found in deeper samples (Cushman, 1914). Thefrequency distributions along the depth gradient are alsoquite distinct, as expressed in constant and significant mul-timodal frequency distributions (Fig. 2). The second Pacificspecies, O. discoidalis, is characterized by complete invo-lute coiling in the first whorls, narrow chambers, a largehyaline umbonal plug, rare knobs, and a rhombic axial sec-

18 HOHENEGGER, YORDANOVA, AND HATTA
tion. It prefers soft substrates and calm conditions, i.e., fine-grained sediments below the fair-weather wave base.
The first group of test characters for separating the threespecies is the form of growth as expressed in the expansionrate of the logarithmic spiral (formula 1) modeling the outertest margin (Table 1; Fig. 3). The three illustrated specimensof O. discoidalis (Pl. 2, figs. 1–6) are best fitted by thelogarithmic spiral and show low mean expansion rates (b1
5 1.10), which is similar to the low rates found in N. ven-osus (b1 5 1.07). Expansion rates are significantly higher(b1 5 1.14) in semivolute, thick representatives of O. am-monoides (Pl. 2, figs. 11, 12) and in the inner whorls of flatevolute forms (Pl. 2, figs. 7–10). The outer margins of innerwhorls in Operculina cf. O. complanata can also be fittedby a simple logarithmic spiral (Pl. 2, figs. 13–18) possessingthe highest expansion rate (b1 5 1.21) of all living Oper-culina species. Similar to species belonging to the involutegroup of nummulitids (H. depressa, N. venosus, and O.cumingii), the expansion rate of the last whorl significantlydeviates in most Operculina species from the logarithmicspiral, following the modified equation of formula (2). De-viations from the ‘normal’ logarithmic spiral are insignifi-cant in O. discoidalis (Table 1; Fig. 3). Flat evolute speci-mens of O. ammonoides show significant deviations fromthe normal spiral function in the last whorl, as is the casefor all Operculina cf. O. complanata specimens. Again, thechanging expansion rate as a function of size is significantlyhigher in Operculina cf. O. complanata (b2 5 1.17) thanin the flat evolute O. ammonoides (b2 5 1.11), which ex-plains the stronger expansions of the final whorl in the for-mer species despite the similar changing factors c (Table 1;Fig. 3).
The transfer from involute to semiinvolute and evolutetest parts within a specimen can also be demonstrated usingthe modified logarithmic growth spiral functions. Relatingthe inner margin of the last whorl to the outer margins ofboth last whorls characterizes the proportion of overlapping(Fig. 4); coincidence of the outer margin of the penultimatewhorl with the inner margin is typical for evolute coiling.The three investigated specimens of O. dicoidalis are dis-tinguished by a large involute test part until a mean outerradius of 0.85 mm. Total evolute whorls would theoreticallyoccur at a mean radius of 2 mm. The mean limits of involuteand semiinvolute test parts are located at a radius of 0.4 mmand 1 mm in the flat specimens of O. ammonoides, whileOperculina cf. O. complanata specimens shifts these meanlimits to 0.3 mm and 0.7 mm radii (Fig. 4). In combinationwith the high expansion rates, the involute and semiinvolutetest parts are restricted to the first whorl (radians: 0 to 6.828)in O. complanata, whereas complete evolute coiling dom-inates the remaining test.
Axial section and test thickness is the second charactergroup that enables morphological differentiation in Oper-culina species. While in axial section the evolute morpho-types of O. ammonoides and all individuals of Operculinacf. O. complanata are distinguished by virtually parallelwalls slowly decreasing from the central part, differencesbetween O. discoidalis and the thick semiinvolute O. am-monoides become significant. All representatives of O. dis-coidalis show rhombic axial sections (Pl. 5, figs. 1–6) alsoin the extremely flat forms (Pl. 5, fig. 4), while the thick O.
ammonoides demonstrate parallel sides in axial view (Pl. 5,figs. 7–9). Test thickness also differs between the fourgroups of morphotypes. Since thickness is a function of size,the ratio (T/D) between maximum thickness and the largestdiameter is used to characterize this trait (e.g., Hallock1979). The ratio of the single, thick, and semiinvolute rep-resentative of O. ammonoides (Pl. 2., figs. 11, 12) is similarto the mean value of 0.394 in O. discoidalis, while threeinvestigated flat forms of the former species are distin-guished by a slightly lower mean ratio of 0.281. This differsconsiderably from the three figured specimens (Pl. 2, figs.13–18) of Operculina cf. O. complanata (T/D 5 0.112).
The chamber number in the last whorl also allows dif-ferentiation between the Operculina species (Fig. 5). Whilethe investigated semiinvolute as well as evolute specimensof O. ammonoides show a strong increase starting with ninechambers at a radius of 0.5 mm and attaining 21 chambersat a final radius of 4 mm, Operculina cf. O. complanatahas a slower increase. It starts with 13 chambers and reaches19 chambers at 4 mm. The number of chambers per whorlis rather constant in O. discoidalis and varies around a meanvalue of 23.
Chamber form can be characterized by the angle of back-ward bending as defined in the chapter on involute num-mulitids. This character is a function of the revolution angle,and can be used to differentiate all Operculina species (Fig.6). In the thick and semiinvolute O. ammonoides specimen(Pl. 2, fig. 12), this angle remains more or less constant(308). The flat and evolute forms of O. ammonoides, in con-trast, are distinguished by an increase starting with a meanangle of 248 after the first whorl and attaining an angle of808 after the third whorl. This angle is significantly largerin every whorl of Operculina cf. O. complanata, where amean of 398 characterizes the end of the first whorl, becom-ing 1208 at the end of the third whorl. The distinct chamberform of O. discoidalis is well-documented by this parame-ter. All three investigated specimens show a mean initialangle of 218 after the first whorl. While the thick specimen(Pl. 2, fig. 2) exhibits a weak increase (268 after the 3rdwhorl), which is similar to the mean in the semiinvolute O.ammonoides specimen, both other specimens show a de-crease starting with the 3rd whorl. The mean angle at theend of the 4th whorl is 78. Thus, chambers become narrowin the last whorl of O. dicoidalis and are distinguished bystraight or convex septa (Pl. 2, figs. 4, 6).
OPERCULINA AMMONOIDES (GRONOVIUS), PL. 2, FIGS. 7–12;PL. 5, FIGS. 7–12
Two distinct morphogroups can be separated using thegrade of chamber embracing and test thickness. Both mor-photypes are distinguished by hyaline septa, which areslightly elevated in the inner test part. Disintegration of sep-tal elevations into small and rounded papillae is possible inlateral parts and transgresses to the center. Higher elevationleads to small knobs, which are arranged like a string ofpearls. Septal elevations and knobs can attain extremeheight and become sharp ridges in geographic variants ofthis species living near the northern distribution boundary(Pl. 5, figs. 10–12). Some knobs also become larger, form-

19WEST PACIFIC NUMMULITIDAE
PLATE 41–6 Heterostegina depressa d’Orbigny. 1, 3, 5 photomicrographs. 2, 4, 6 X-ray micrographs, specimens from 50 m depth at Nakanose Reef
(Hohenegger and others, 1999), 312. 7 Cycloclypeus carpenteri Brady, photomicrograph, specimen from 70 m depth in the transect W of SesokoIsland, 310. 8–9 Planostegina operculinoides (Hofker), details of the surface, SEM micrographs. 8 34658. 9 3122, specimen from 70 m depthin the transect NW of Sesoko. 10 Nummulites venosus (Fichtel and Moll), details of specimen Pl. 1, fig. 9 demonstrating elevated trabeculae,photomicrograph, 337. 11 Planoperculina heterosteginoides (Hofker), details of the surface, SEM micrograph, specimen from 90 m depth in thetransect NW of Sesoko Island, 384.

20 HOHENEGGER, YORDANOVA, AND HATTA
ing bosses in the central test. Often they are not restrictedto septal courses. The thick, semiinvolute morphotypes aredistinguished by low expansion rates and a mean T/D-ratio(max. thickness/max. diameter) of 0.35. All evolute, flatmorphotypes show high expansion rates, where septa be-come depressed in the final chambers of large-sized speci-mens. The mean T/D-ratio of 0.28 is more than twice thatof the form-related Operculina cf. O. complanata. Canalsystems as well as chamber connections such as stolons andforamina are comprehensively described by Hottinger(1977a). Detailed information on test morphology can befound in Hottinger (1977b) and Hottinger and others (1993).Synonymy lists are also given in Hottinger (1977a) and Hot-tinger and others (1993).
With the exception of studies on symbiotic algae (Leu-tenegger, 1977a; Lee and others, 1989), nothing is knownabout the biology—especially nutrition, growth, and repro-duction—of this morphologically well-studied species. Re-production biology is concluded from size differences of theinitial chambers and mixed with ecological interpretationsthat are mainly based on the distribution of empty tests (Pe-cheux, 1995).
O. ammonoides prefers coarse sand in both transects NWoff Sesoko, Okinawa, and is found in low numbers on mud-dy bottom here. Specimens also inhabit well-structured coralrubble, where they find protection against irradiation anderosion in small grooves (Hohenegger and others, 1999).Life just below the sand surface in not shaded areas, asnoted by Lee (1994) from the Red Sea, could not be verifiedfor the investigated Pacific forms, but a semi-infaunal lifemode of the thick specimens was observed in muddy sedi-ments. Transitions in frequency distributions from evoluteto semiinvolute specimens are more or less continuous inthe upper part of the investigated distribution range, but thelatter morphotypes avoid living on coarse-grained sedi-ments. Fine sand and silt is the preferred substrate of thesethick forms in the Ryuku Archipelago (Pl. 5, figs. 7–9),while the geographic variants with sharp septal elevations(Pl. 5, figs. 10–12) are the monospecific larger foraminifersliving on muddy substrate at Amakusa Island (Fig. 1) nearthe northern distribution boundary of O. ammonoides. Onlya few specimens of this ‘northern’ morphotype were rep-resented in slope samples from Okinawa.
Light intensity is the main factor influencing the distri-bution of O. ammonoides (Fig. 7). The abundance optimumis at 14% surface PAR corresponding to 45 m in very clearseawater (attenuation 5 0.04). Slightly stronger (0.06) orhigh attenuation (0.13) shifts this optimum to 35 m and 15m, respectively. Attenuation in the lagoon W of MotobuPeninsula (Hohenegger, 1994; Hohenegger and others,1999) is 0.13 due to suspended organic and inorganic par-ticles. This leads to the occurrence of thick O. ammonoidesin muddy sediments of shallow lagoons (maximum depth19 m). The input of fine-grained sediments by rivers intocalm areas around Kyushu and South Honshu, Japan, is cou-pled with a relatively high attenuation (; 0.10), and enablessettlement of O. ammonoides. The geographic morphotypeat these latitudes has flat tests with highly elevated suturesand is the monospecific larger foraminifer on muddy bottomfrom 9 m to 40 m (Chiji and Lopez, 1968; Oki, 1989). Thelower distribution limit of O. ammonoides is located at
1.5% surface PAR, which extends the distribution to 95 min very clear ocean-water. Lower transparency (attenuation5 0.06) shifts this limit upwards to 70 m, and strong atten-uation of 0.13 as found in muddy lagoons makes life below32 m impossible for this species. The upper limit of irra-diation tolerance is apparentely 68% surface PAR, coincid-ing with 9 m depth in very clear seawater. Attenuation of0.06 shifts this limit to 6 m and the first few O. ammonoidescan be found at 3 m in calm lagoons with fine-clastic andbiogene suspension, which raises attenuation to .0.10.
The mean test size of 780 measured living individualsfrom both transects at Sesoko Island reveals a slight increasefrom 1.5 mm at 20 m to 2 mm at 60 m in the northerntransect. This trend is weakly significant in the northern andinsignificant in the southern transect, where size variesaround a mean of 1.8 mm. Hohenegger and others (1999)demonstrated the consistency of means and variances of testsizes in two subgroups with increasing depth. The propor-tions also remain constant in both groups, which compli-cates an interpretation as different generations as long asproloculus size is not measured. Nevertheless, the few mea-sured specimens (seven individuals) belong to two sizegroups. The one is distinguished by initial chambers with amean 105 mm diameter, while the other measures 45 mm.Fermont (1977a) and Pecheux (1995) noted differences inproloculus size between the semiinvolute and evolute mor-photypes, but could not explain these distinctions biologi-cally. Measurement of proloculus size from all specimensof the investigated transects, in combination with reproduc-tion studies, may shed light on the problem of different gen-erations in O. ammonoides.
OPERCULINA CF. O. COMPLANATA (DEFRANCE), PL. 2, FIGS.13–18
The tests are very similar in size and coiling to the flat,evolute O. ammonoides. Differences can be found in themore pronounced expansion rate (Table 1; Fig. 3). Hyalinesepta are slightly elevated, especially in the deeper-livingforms. The test surface is completely smooth in some spec-imens living near the upper distribution limit. In most in-dividuals, narrow knobs cover the surface of inner whorls,but become rare in the flat expanded final part (O. com-planata var. granulosa in Cushman, 1914). Depressed su-tures often characterize these final whorls. The main differ-ence from the flat, evolute O. ammonoides is the intensivelyfolded septal flaps that start after the first whorl. In com-paring these specimens with typical O. complanata fromthe Oligocene and Early Miocene, the homeomorphism andagreement in most test characters is remarkable (Hottinger,1977a). The missing connecting links in the Upper Mioceneand Pliocene prevent a definite integration of the livingforms within O. complanata, since homeomorphism couldbe caused by iterative evolution.
The biology of this species is mostly unknown, exceptreproduction, where the release of biflagellated gametes wasobserved for the first time within the genus Operculina(Rottger and others, 1998). A small number of mastigo-nemes, so far proven for ‘primitive’ foraminifers and Am-phistegina gibbosa (Hedley and others, 1968; Goldstein,

21WEST PACIFIC NUMMULITIDAE
1997; Dettmering and others, 1998), covers the flagellae ofthis species (Rottger and others, 1998).
Operculina cf. O. complanata is abundant on sandy bot-toms in the investigated transects NW off Sesoko, wheremiddle-grained to fine-grained sediments dominate. Waterenergy influences the distribution inasmuch as this speciesavoids strong water movement. The abundance optimum isachieved at 3% surface PAR, which explains the distributionof Operculina cf. O. complanata in deeper, light-depletedwater (Fig. 7). This optimum corresponds to 80 m at anattenuation of 0.04 and 60 m in less transparent water (at-tenuation 5 0.06). The lower limit at 0.2% surface PARcorrelates with 140 m depth in very clear ocean water, mak-ing this one of the deepest-living larger foraminifers. Stron-ger attenuation of 0.06 shifts this limit to 105 m. The upperdistribution boundary is located at 12% surface PAR (50 min very clear seawater, 40 m in less clear water). Suspendedsediment particles that cause strong attenuation elevate theupper distribution limit to 16 m. Thus, Operculina cf. O.complanata can be expected on fine-sand bottoms in shal-low water under calm conditions and low transparency, evenat 30 m.
Test size (2451 measured living specimens) does not sig-nificantly change with water depth. Variation around meansizes of 1.8 mm (SD 5 0.18) and 2.1 mm (SD 5 0.26),respectively is characteristic for the investigated transects ofSesoko Island, Okinawa. Test flattening with depth was not-ed, but not proven statistically. The mean T/D-ratio of Oper-culina cf. O. complanata measured on seven individualsfrom a 70 m sample is 0.169, whereas this parameter be-comes 0.103 at a 90 m sample (eight measured specimens).Therefore, test flattening in combination with the high num-ber of septal piles and knobs seems to enable life near thebase of the photic zone. Proloculus size was also measuredon the 15 individuals mentioned above, and can be groupedmuch like the initial chambers of O. ammonoides. Despitethe small number, two size groups were recognized. Largeproloculi vary between 77 mm and 114 mm, while the smallsizes measure 29 mm to 56 mm. Differences between depthsare highly insignificant in small-sized proluculi (mean at 70m 5 43.6 mm, mean at 90 m 5 45.0 mm), but become closeto significant in the group with large initial chambers ac-cording to one-way analysis of variance and the Mann-Whitney U-Test. The initial chambers are slightly larger inthe shallower (mean 5 96.2 mm) than in the deeper sample(mean 5 81.4 mm). These differences in proloculus sizecannot be explained biologically. They may signify gener-ations (agamonts and gamonts), but this remains to be prov-en in reproduction studies under laboratory conditions.
OPERCULINA DISCOIDALIS (D’ORBIGNY), PL. 2, FIGS. 1–6;PL. 5, FIGS. 1–6
Tests are completely involute in the first whorls and be-come semiinvolute to evolute starting with the 3rd or 4thwhorl. Transitions may be so rapid that the change frominvolute to evolute coiling takes place within one whorl(Fig. 4; see individuals Pl. 2, fig. 1; Pl. 5, fig. 2). Expansionrates of the logarithmic spiral are low (mean b1 5 1.10), butsignificantly higher than in Nummulites venosus. A largeand thick hyaline central plug characterizes the umbilical
region and becomes the thickest test part. Thus, axial sec-tions are rhombic (Pl. 5, figs. 1–6), which is a good diag-nostic character for separating O. discoidalis from the re-lated semiinvolute O. ammonoides, which differs in havingparallel lateral sides (Pl. 5, figs. 7–9). The septa are slightlyelevated in geographical variants from Belau, Western Car-olines, and disintegrate into tiny pearl-like papillae, but nev-er become knobs (Pl. 5, figs. 1–3). The pearl-like arrange-ment of stronger septal papillae that change in the centralpart into small knobs is typical for the northern geographicvariants as represented in the Ryukyus (Pl. 5, figs. 4–6). Allseptal flaps are smooth, again similar to O. ammonoides.Nothing is known about the test microstructure, especiallythe canal system, but no trabeculae were detected. The nameO. discoidalis seems to be correct, since the type specimenof d’Orbigny (1826) as well as the variant O. discoidalisvar. involuta as introduced by Cushman (1921) fall into thevariation range.
The biology of O. discoidalis is completely unknown,and substrate preferences are also unclear. This species isvery abundant on muddy substrates in tropical regions,where it is often the monospecific larger foraminifer. Thus,the foraminiferal fauna of lagoons or small inter-island ba-sins that are characterized by calm water conditions andmuddy bottom as represented in Sesoko (Hohenegger, 1994)and Belau (Hohenegger, 1996) is dominated by this species.Besides this predominance, some O. discoidalis can befound on coarse- to medium-grained sands of the reef slope.Well-structured basins or lagoons behind the fringing reefsof Belau create different environments that are reflected inthe composition of the larger foraminiferal fauna. Despitesimilar position (200 m behind the reef crest), identicaldepth (30 m), and similar sediments (coarse to mediumsand), the larger foraminiferids differed. In one sample ofnormal diversity, N. venosus and H. depressa are abundantalongside O. ammonoides. The other sample, located 500m to the west of the first sample, is dominated by O. dis-coidalis. Only a single living specimen of O. ammonoideswas found, while 512 living individuals of O. discoidalisgave the impression of a monospecific habitat (Hohenegger,1996). No other larger foraminifer was detected at this lo-cation.
Beside hydrodynamics, substrate, and nutrition, light in-tensity seems to be the fourth factor influencing the habitatof this species. Only two measurements of surface PARwere taken in the lagoon and on the reef slope near SesokoIsland, Japan. Light intensity is 9% surface PAR at 18 m inthe lagoon, where O. discoidalis is abundant. Two livingindividuals were collected at 50 m on the reef slope, whichcorresponds to 7% surface PAR. Individuals from Belau, onthe one hand, are abundant on black mud at 11 m depth inthe bay of Malakal Port. On the other hand, they inhabitmedium sand at 30 m south of Kwannon Island (Hoheneg-ger, 1996). Although light intensity could not be measuredat the sampling stations in Belau, the high transparency ofseawater, especially in the locality south of Kwannon Island,indicates a higher PAR tolerance of O. discoidalis than wasmeasured at Sesoko Island.
EVOLUTE NUMMULITIDS WITH COMPLETE ORINCOMPLETE CHAMBER PARTITIONS
Three species of West Pacific nummulitids belong to thismorphogroup, which is characterized by evolute, planispiral,

22 HOHENEGGER, YORDANOVA, AND HATTA
and flat tests. As in the evolute Operculina species, knobscover the surface of inner whorls at least, but papillae fol-lowing the septal courses are not developed. Strong foldingof the septal flaps changes into undulation that starts thedivision of chambers into chamberlets. The emended genusname Planoperculina Hottinger (1977a) is used for thesenummulitids with incomplete chamber partitions (Bannerand Hodgkinson, 1991). Evolute nummulitids that are dis-tinguished by complete chamber partitions (5chamberlets)through septula were given the genus name Planostegina(Banner and Hodgkinson, 1991) in consideration of the ho-meomorphism with Heterostegina.
While differentiation of Planoperculina heterosteginoi-des (Hofker, 1933) from the Planostegina species is easybased on the incomplete development of septula, the mor-phological differentiation between both Planostegina spe-cies will be described in the following passage section. Mor-phological separation is based on specimens taken at 90 mW off Sesoko Island, Okinawa. In every sample from thisdepth, the two Planostegina species are morphologicallydifferentiated without transitions. Additionally, Planoper-culina heterosteginoides is represented at this depth andalso lacks transitional forms to both Planostegina species.Naming of the two Planostegina species is difficult, sincetheir taxonomy was confused with the first discovery ofthese deep-dwelling forms (Hofker, 1927; see Hottinger1977a). Here, the name Planostegina aff. P. operculinoidesis used for the more robust forms, while the extreme flatforms with granules are typical Planostegina operculinoi-des as described by Hofker (1933) and Hottinger (1977a);an exact determination requires investigation of the typespecimens.
All investigated tests follow the exponential spiralgrowth, at least in the inner whorls (Table 1; Fig. 3). Theexpansion rate is identical and extremely high both in P.operculinoides and P. heterosteginoides (b1 5 1.25), whilea slightly lower rate of 1.21—which is the same as in Oper-culina cf. O. complanata—characterizes Planostegina aff.P. operculinoides. All final whorls of large-sized individu-als are distinguished by significant deviations from the nor-mal logarithmic spiral in possessing a flexible expansionrate (formula 2). Again, Planostegina aff. P. operculinoidesis characterized by the lowest expansion rate (c 5 0.0051),while P. heterosteginoides shows a slightly stronger expan-sion (c 5 0.0079) than P. operculinoides (c 5 0.0068).According to these results, differences in spiral growth areinsignificant between Planostegina operculinoides andPlanoperculina heterosteginoides, but obvious for bothPlanostegina species.
Test thickness is the second trait separating the three spe-cies. The mean ratio between maximum thickness and max-imum diameter (T/D-ratio) was measured using four indi-viduals from each species. This ratio is virtually identical inPlanostegina aff. P. operculinoides (T/D 5 0.126) andPlanoperculina heterosteginoides (T/D 5 0.120). Both areslightly higher than the ratio of Operculina cf. O. complan-ata (T/D 5 0.112). P. operculinoides is distinguished byextremely flat tests, reflected in a T/D-ratio of 0.077. There-fore, thickness is a good character for separating the Plan-ostegina species operculinoides and aff. P. operculinoides.
Chamber number per whorl is also a good diagnostic
character within this morphogroup (Fig. 5). Both Planos-tegina species demonstrate a similar increase, starting witha mean of seven chambers per whorl at a 0.5 mm radius.This number increases in P. operculinoides to 28 chambers,and in Planostegina aff. P. operculinoides to 23 chambersat a 4 mm radius. Again, Planoperculina heterosteginoidesis similar to Operculina cf. O. complanata in this character:12 chambers per whorl are present at a radius of 0.5 mmand the slow increase leads to 19 chambers at 4 mm, whichis the identical number in the related Operculina species.
Chamber form is characterized by the angle between theinteriomarginal onset of the chamber at the older test andthe contact of the marginal wall with the preceding chamber,as described in the chapter on involute nummulitids. Again,this character strongly differs among the three species (Fig.6). Planostegina aff. P. operculinoides has the lowest val-ues, with a mean angle of 588 after the first whorl and 1028after the third whorl. Embracing of the marginal chambersis stronger in P. heterosteginoides, starting with a slightlylower mean angle of 488 after the first whorl, but attaining1538 after the third whorl. Strongest embracing can be foundin P. operculinoides (768 at the end of the first whorl). Thestrong increase to 1808 after the second whorl demonstratesthe tendency to total embracing and annular arrangement ofchamberlets (Pl. 3, figs. 8, 12).
The chamberlets in Planostegina act as compartments en-abling information transfer and the regulation of physiolog-ical processes. Their area does not always remain constantduring growth, as supposed for larger foraminifers by Reissand Hottinger (1984). Chamberlets show a slow but signif-icant increase with growth in all measured specimens of P.operculinoides as tested by linear regression. The area ofchamberlets is low in P. operculinoides (mean 5 13,924mm2, SD 5 8,849), and does not significantly differ betweenthe three measured specimens (Pl. 3, figs. 7–12) as testedby one-way analysis of variance (Table 2). Chamberlets aresignificantly larger in Planostegina aff. P. operculinoides(mean 5 32,417 mm2, SD 5 27,833) as proven by ANOVA(Table 2), again demonstrating homogeneous size in thethree measured specimens (Pl. 3, figs. 1–6). Chamber sizeis therefore more than two times larger in Planostegina aff.P. operculinoides, and serves as a further diagnostic char-acter to differentiate the two Planostegina species in theWest Pacific (Table 2).
PLANOPERCULINA HETEROSTEGINOIDES (HOFKER), PL. 3,FIGS. 13–18; PL. 4, FIG. 11
The flat, evolute, and planispirally coiled tests deviatestrongly from the logarithmic growth spiral in the finalwhorl of large individuals. In combination with the highexpansion rate (b1 5 1.26) of the ‘normal’ logarithmic spiralat the inner whorls, chambers of the final test part may ap-proximate uniserial arrangement (Pl. 3, figs. 17, 18). Theinner lamella of the chamber wall that covers the previouschamber is intensively folded and occasionally touches thefrontal septum. Thus, undulations demonstrate tendencies todivide the chambers completely, which is realized in themarginal chamber part (Pl. 3, figs. 14, 16, 18). The arrange-ment of numerous, small, cone-shaped knobs on the surfacefollows the septal courses like strings of pearls. Septal un-

23WEST PACIFIC NUMMULITIDAE
PLATE 51–3 Operculina discoidalis (d’Orbigny), lateral and axial views, specimens from 30 m depth south of Kwannon Island, Belau (Hohenegger,
1996). 4–6 Operculina discoidalis (d’Orbigny), lateral and axial views, specimens from 19 m in the lagoon NE of Sesoko Island (Hohenegger andothers, 1999). 7–9 Operculina ammonoides (Gronovius), semiinvolute form, lateral and axial views, specimens from 19 m in the lagoon NE ofSesoko Island. 10–12 Operculina ammonoides (Gronovius), evolute form with strong sculptures, specimens from 30 m depth south of AmakusaIsland (Fig. 1). All micrographs; 315.

24 HOHENEGGER, YORDANOVA, AND HATTA
TABLE 2. Testing of homogeneity and differences in chamberlet area within and between the Planostegina-species by one-way analysis of variance(ANOVA).
Number ofchamberlets
Mean area inmicrometer2
Standarddeviation
Planostegina operculinoides, plate 3Specimen figs. 7, 8Specimen figs. 11, 12Specimen figs. 9, 10Total
285279148712
14,53913,90812,77013,924
8,8738,8028,8378,850
ANOVA Mean square df F-value Significance
Variance between groupsVariance within groups
152,363,65778,106,701
2709
1.951 0.143
Number ofchamberlets
Mean area inmicrometer2 Standard deviation
Planostegina aff. P. operculinoides, plate 3Specimen figs. 5, 6Specimen figs. 1, 2Specimen figs. 3, 4Total
200339181720
30,15334,59330,84632,418
23,55231,69224,04427,833
ANOVA Mean square df F-value Significance
Variance between groupsVariance within groups
1,538,485,786772,557,459
2717
1.991 0.137
ANOVA Mean square df F-value Significance
Comparison between Planostegina operculinoides and Planostegina aff. P. operculinoidesVariance between groupsVariance within groups
25,163,463,577427,280,048
51,426
58.892 0.000
dulations are also marked on the test surface by small knobs(Pl. 4, fig. 11). In the final test part, all knobs fuse at theirbase, resulting in structured ridges that trace the inner teststructures. Knobs become larger and dome-shaped in thecentral test. Their arrangement and form is then similar toknobs in the umbonal part of Operculina cf. O. complanata.A comprehensive description of the test morphology is giv-en in Hottinger (1977a), where the canal system is picturedand described in detail. Short synonymy lists can be foundin Hottinger (1977a) as well as Loeblich and Tappan (1994).
The biology of P. heterosteginoides is completely un-known. Proloculus size, measured on four specimens, variesbetween 62 mm and 86 mm. These measures coincide withOperculina cf. O. complanata specimens possessing largerproloculi, and homogeneity does not indicate any generationdifferences. P. heterosteginoides prefers medium to fine-grained sand and calm water. The mean test size of P. het-erosteginoides, based on 221 individuals, is 2.8 mm at 90m and 3.7 mm at 100 m depth. Fixation mechanisms to thesubstrate seem to be weak, as is concluded in analogy tothe related species Cycloclypeus carpenteri (Kruger andothers, 1996), where large and flat individuals are easilytransported by slow water motion.
Most individuals live at weak light intensities of 0.8%surface PAR (Fig. 7), corresponding to 110 m in very clearwater (attenuation 5 0.04). Slightly unclear water (attenu-ation 5 0.06) shifts this limit upwards to 80 m, and weaktransparency (attenuation 5 0.13) transposes this optimumto 37 m. The lower distribution limit is located at 0.3%surface PAR, and the corresponding depths for different at-tenuation coefficients are 132 m (0.04), 97 m (0.06), and 45
m (0.13). 2.5% surface PAR is the upper limit of light in-tensity for P. heterosteginoides; this correlates with 87 m(0.04), 63 m (0.06), and 29 m (0.13) at various seawatertransparencies. Beside Operculina cf. O. complanata, thisspecies is the deepest-dwelling nummulitid in the West Pa-cific.
PLANOSTEGINA OPERCULINOIDES (HOFKER), PL. 3, FIGS. 7–12;PL. 4, FIGS. 8, 9
This species differs from Planoperculina in having com-pletely subdivided chambers. Tests are extremely flat, evenin large-sized individuals, which is exemplified by a T/D-ratio below 0.1. Septal and septula elevations strongly riseabove the test surface, especially in the final whorl (Pl. 4,fig. 9). A few dome-shaped knobs are located on the surfaceof the central test part, but they become obscured by a veryhigh number of granules. Granulation decreases withgrowth, but sparsely scattered granules are also present inthe final test parts (Pl. 4, fig. 9). In some specimens inten-sive granulation spreads over the whole test. Beside thesegranules, fine spikes of 3.5 mm maximum length denselycover the lateral wall surface in the umbonal regions (Pl. 4,fig. 8). Their function is unclear. A detailed description oftest morphology that includes the canal system is given inHottinger (1977a) and Hottinger and others (1993). Theshort synonymy list in Hottinger and others (1993) must becompleted with Heterostegina longisepta Zheng (1979).This name is also used by Hatta and Ujiie (1992) for P.operculinoides.
The biology of this species is unknown. Hofker (1933)

25WEST PACIFIC NUMMULITIDAE
noted three generations in his emendation of H. operculi-noides. The two A-generations are characterized by largeproloculi (;70 mm), while the initial chamber of the B-generation is much smaller (;20 mm). All measured pro-loculi (four specimens) from the lower distribution bound-ary of P. operculinoides at 90 m range between 37 mm and45 mm, and are thus situated between both generation clas-ses mentioned by Hofker (1933). Detailed investigation mayshed light on the problem, if these small proloculus sizesresult from diminution of the embryonic apparatus corre-sponding to depth as described for O. ammonoides and H.depressa (Fermont 1977a,b). P. operculinoides lives oncoarse to fine-grained sand and avoids hard substrates. Thisspecies prefers calm water, but is not disturbed by weakwater motion.
Light intensity influences the habitat range of P. oper-culinoides (Fig. 7). The frequency distribution reaches anoptimum at 2.7% surface PAR, which is the highest of allevolute, flat nummulitids in this morphogroup. Differingtransparency (attenuation 5 0.04, 0.06, and 0.13) corre-sponds to optima at 82 m, 60 m, and 28 m, respectively.The upper distribution limit of 26% surface PAR shifts thislimit from 31 m in clear water (attenuation 5 0.04) to 22m (attenuation 5 0.06) to 10 m in waters of low transpar-ency. This high limit explains why living P. operculinoidesare sometimes represented in more or less shallow water.0.45% surface PAR signalizes the lower distribution limit(123 m in very clear ocean water; attenuation 5 0.04). Low-er transparency (attenuation 5 0.06 and 0.13) moves thislimit upwards to 100 m and 42 m, respectively.
Since the depth distribution of P. operculinoides showsa broad range, tendencies in test size could be checked basedon 125 living individuals. W off Sesoko, test size decreasedsignificantly in the southern transect. The largest mean di-ameters (4.4 mm and 2.6 mm) are observed in both transectsbetween 40 m and 60 m depth, coinciding with the distri-bution optimum. Towards the deeper parts, the decrease intest size is significant in both transects, attaining a finalmean size of 1.5 mm at 90 m. Competition with Planoper-culina heterosteginoides and Planostegina aff. P. opercu-linoides, that become abundant in these deeper regions, mayexplain the size decrease in P. operculinoides. Also the lowproportion of large-sized agamonts, which are abundant atthe niche optimum and completely disappear at the distri-bution borders (Pecheux, 1995; Hottinger, 1997), may ex-plain depth-correlated diminution in test size.
PLANOSTEGINA AFF. P. OPERCULINOIDES (HOFKER), PL. 3,FIGS. 1–6
The evolute, flat and planispirally coiled tests deviate infinal test parts from the logarithmic spiral. All chambers arecompletely subdivided into chamberlets by secondary septa.Dome-shaped knobs of variable size cover the surface ofthe inner whorls, while the largest test part is completelysmooth. Septal elevations in the form of low ridges are onlydeveloped in final chambers of very large tests. The smoothlateral walls of chamberlets are subdivided into hyaline ar-eas that are separated by narrow, band-like fields. Pores con-centrate in these bands, while the hyaline regions are im-perforate. These transparent areas cannot be found in other
living nummulitids and are interpreted as mechanisms toreceive light for the symbiotic algae. The test microstruc-ture—especially the canal system—is unknown, but it seemsto be similar to P. operculinoides as described in Hottinger(1977a).
Since this is the first record of this species, nothing isknown about its biology. Proloculus size was measured inthe three figured individuals (Pl. 3, figs. 1–6). While twospecimens show sizes (82 mm and 84 mm) similar to pro-loculi in Planoperculina heterosteginoides, the third is sig-nificantly smaller (52 mm) and resembles diameters in P.operculinoides. Similar to P. heterosteginoides, Planoste-gina aff. P. operculinoides prefers medium- to fine-grainedsand deposited under calm conditions. The mean test sizeof the 104 measured individuals is 2.3 mm.
Based on light intensity, the optimum is located at 1.2%surface PAR, which is slightly higher than in P. heteroste-ginoides (Fig. 7). This maximum corresponds in very clearocean water (attenuation 5 0.04) to 100 m depth, and isshifted under less transparent conditions (attenuation 50.06) to 74 m or 34 m (attenuation 5 1.13). The tolerancerange is narrow, with a lower limit at 0.4% and an upperlimit at 2.7% surface PAR. Differing transparency of theocean water moves for the lower limit to 126 m (0.04), 102m (0.06), and 43 m (1.13). The corresponding depths to theupper limit of surface PAR are 82 m, 60 m, and 28 m,respectively. Based on light intensity, the optimum andrange of Planostegina aff. P. operculinoides are very sim-ilar to P. heterosteginoides.
ANNULAR NUMMULITIDS
Two species of nummulitids belonging to different generaare distinguished from the others by an annular arrangementof chamberlets. Heterocyclina tuberculata Mobius is theonly nummulitid that is not reported from the Pacific, whileCycloclypeus carpenteri inhabits deeper bottoms (60–150m) in the tropical Central and West Pacific (Koba, 1978).Generic differences are based on the embryonic apparatus,where H. tuberculata resembles Planostegina in initialchambers. Additionally, the canal systems and stolons thatconnect the chamberlets differ between Heterocyclina andCycloclypeus (Hottinger, 1977a), thus becoming diagnosticcharacters.
CYCLOCLYPEUS CARPENTERI BRADY, PL. 4, FIG. 7
The annular arrangement of chamberlets is characteristicfor the sexual and asexual generations. Chamberlets aremuch longer than wide and seem to retain their size withgrowth, but this was not proven statistically. No interseptalpillars–the typical surface structure of Heterocyclina tuber-culata–can be found in C. carpenteri. Septa as well as sep-tula are slightly elevated over the glassy, hyaline, and in-tensively perforated walls of chamberlets. In contrast to oth-er nummulitids with chamber partitions and pronounced su-tures, elevations are higher in the septula than in the septa.Septula elevation starts with low height at the precedingchamber and instantaneously becomes higher. It ends witha rounded knob at the frontal septum. This leads to an an-nular arrangement of equally sized knobs on the flat surfaceof the lateral test. Large knobs also cover the thick central

26 HOHENEGGER, YORDANOVA, AND HATTA
part of C. carpenteri, but retain their annular arrangement,which traces the courses of central chambers and chamber-lets. Test microstructure, especially stolons and the canalsystem, is described in detail by Hottinger (1977a), whileHofker (1927) explained the embryonic apparatus and dif-ferentiation between generations.
C. carpenteri is the second nummulitid, in which someaspects of biology and reproduction are known (Kruger andothers, 1996). The size difference between the giganticagamonts and the much smaller gamonts is obvious. Sexualreproduction is documented by the release of biflagellatedgametes from gamonts (Rottger and others, 1998), whileasexual reproduction was proven by multiple fission. Theseprocesses are very similar to the reproduction modes in H.depressa (Kruger, 1994). The life span of gamonts was mea-sured in laboratory cultures and doubles H. depressa witha mean of 360 days (Kruger and others, 1996). The longev-ity of agamonts was estimated by relating growth rates oflaboratory cultures to maximum test sizes in natural popu-lations. Thus, a life span of several to many years is sup-posed for agamonts (Kruger and others, 1996).
The geographical as well as depth distribution of C. car-penteri was studied by Koba (1978). He found this speciesrestricted to the slope of coral reefs, with an optimumaround 90 m. Investigations of the present authors revealeda preference of C. carpenteri for medium- to fine-grainedsand. Strong water motion transports the individuals, sincethey live loosely attached to the grains on the sandy surface.Depending on bottom topography, currents can be intensive(50 cm/s) at 200 m depth (Yamazato and others, 1968). Onthe other hand, C. carpenteri is not found on muddy bot-toms in deep lagoons (Koba, 1978).
Again, beside water energy, light intensity seems to bethe dominant environmental gradient affecting the distribu-tion of C. carpenteri (Fig. 7). The optimum is located at2.8% surface PAR (81 m, attenuation 5 0.04; 60 m, atten-uation 5 0.06). 0.4% surface PAR signalizes the lower dis-tribution limit, corresponding to 126 m in very clear sea-water and 102 m in less transparent water. The upper dis-tribution limit is masked by the dependence of C. carpenterion water motion, but apparently located at 10% surfacePAR. The corresponding depths in clear and less clear waterare 51 m and 42 m, respectively.
Test size distribution along the depth gradient was mea-sured on 70 specimens of small-sized gamonts. No signifi-cant size change with depth was detected; thus, a mean of8.23 mm (SD 5 3.28) seems to be characteristic for thisspecies, while the agamonts collected by SCUBA measuredbetween 3 cm and 6 cm.
ACKNOWLEDGMENTS
Thanks are due to the whole scientific and technical staffof the Tropical Biosphere Research Center, Sesoko Station,University of the Ryukyus, especially Kiyoshi Yamazatoand Kazunori Takano, who made field work for J.H. as aninvited foreign researcher in the years 1992, 1993, and 1996possible. A.H. and J.H. collected samples while participat-ing in the 1995 cruise on the vessel ‘Keiten Maru’ to Belau,Micronesia, which was initiated by the Kagoshima Univer-sity Research Center for the South Pacific (director Akio
Inoue) as a research project entitled ‘Man and the Environ-ment in Micronesia’. J.H. spent seven months as a visitingprofessor at the Research Center for the South Pacific start-ing with September 1997, where he collected samples at thesouthern coasts of Kyushu. Kimihiko Oki from KagoshimaUniversity was helpful in sampling and discussing sedimen-tological and environmental problems. J.H. further dis-cussed questions on the biology, ecology, and taxonomy oflarger foraminifers with Lukas Hottinger (Basel), Fred Rogl(Wien), and Rudolf Rottger (Kiel). Robert Bolland (Univer-sity of Maryland, Okinawa) collected agamonts of C. car-penteri by SCUBA. Preliminary results were presented as aposter during the 24th European Micropaleontological Col-loquium in Bilbao (September 1998). The main part of thispaper is based on results from the research project ‘Rela-tions between biocoenoclines and taphocoenoclines’ of theAustrian Science Foundation (FWF Project P10946-GEO),where E.Y. and J.H. were financially supported. M. Sta-chowitsch (Wien) revised the English text.
REFERENCES
BANNER, F.T., and HODGKINSON, R.L., 1991, A revision of the fora-miniferal subfamily Heterosteginidae: Revista Espanola de Micro-paleontologıa, v. 23, p. 101–140.
BATSCHELET, E., 1971, Introduction to Mathematics for Life Scien-tists: Springer-Verlag, Berlin-Heidelberg-New York, 495p.
BOLTOVSKOY, E., and WRIGHT, R., 1976, Recent Foraminifera: Dr. W.Junk, The Hague, 515p.
CARPENTER, W.B., 1860, Researches on the foraminifera. Part III. Onthe genera Peneroplis, Operculina, and Amphistegina: Philosoph-ical Transactions of the Royal Society London, v. 149 (1859), p.1–41.
CHIJI, M., and LOPEZ, S.M., 1968, Regional foraminiferal assemblagesin Tanabe Bay, Kii Peninsula, Central Japan: Publications of theSeto Marine Biological Laboratory, v. 16, p. 85–125.
CUSHMAN, J.A., 1914, A monograph of the Foraminifera of the NorthPacific Ocean. Part IV. Chilostomellidae, Globigerinidae, Num-mulitidae: Smithonian Institution, United States National Muse-um, Bulletin 71, p. 1–46.
, 1921, Foraminifera of the Philippines and adjacent seas: Unit-ed States Natural Museum, Bulletin, v. 100, p. 1–608.
DEBENAY, J.-P., 1988, Foraminifera larger than 0.5mm in the south-western lagoon of New Caledonia: distribution related to abioticproperties: Journal of Foraminiferal Research, v. 18, p. 158–175.
DETTMERING, CH., ROTTGER, R., HOHENEGGER, J., and SCHMALJO-HANN, R., 1998, The trimorphic life cycle in foraminifera: Obser-vations from cultures allow new evaluation: European Journal ofProtistology, v. 34, p. 363–368.
DODD, J.R., and STANTON, R.J., 1990, Paleoecology, Concepts andApplications, 2nd Edition: John Wiley, New York, 502p.
DROOGER, C.W., 1993, Radial Foraminifera; Morphometrics and Evo-lution: Verhandelingen der Koninklijke Akademie van Weten-schappen, Afd. Natuurkunde, eerste Reeks, deel 41, Amsterdam,242p.
FALCONER, D.S., 1960, Introduction to Quantitative Genetics: Oliverand Boyd, Edinburgh, 365p.
FERMONT, W.J.J., 1977a, Biometrical investigation of the genus Oper-culina in Recent sediments of the Gulf of Elat: Utrecht Micropa-leontological Bulletins, v. 15, p. 111–147.
, 1977b, Depth-gradients in internal parameters of Heterosteginain the Gulf of Elat: Utrecht Micropaleontological Bulletins, v. 15,p. 149–163.
FUJITA, K., 1997, Annual variation of Marginopora kudakjimaensisGudmundsson assemblages in shallow sea areas surrounding coralreefs (in Japanese): Abstract, 11th Meeting of the Japanese As-sociation of Benthology.
GOLDSTEIN, S.T., 1997, Gametogenesis and antiquity of reproductivepattern in the Foraminiferida: Journal of Foraminiferal Research,v. 27, p. 319–328.

27WEST PACIFIC NUMMULITIDAE
GRANT, V., 1971, The evolutionary species, in Grant, V. (ed.), PlantSpeciation: Columbia University Press, New York, p. 37–45.
HALLOCK, P., 1979, Trends in test shape with depth in large, symbiontbearing foraminifera: Journal of Foraminiferal Research, v. 9, p.61–69.
, 1984, Distribution of selected species of living algal symbiontbearing Foraminifera on two Pacific coral reefs: Journal of Fora-miniferal Research, v. 14, p. 250–261.
, 1985, Why are larger Foraminifera large?: Paleobiology, v.11, p. 195–208.
, and HANSEN, H.J., 1979, Depth adaptation in Amphistegina:change in lamellar thickness: Bulletin of the Geological Societyof Denmark, v. 27, p. 99–104.
HANSEN, H.J., 1979, Test structure and evolution in the Foraminifera:Lethaia, v. 12, p. 173–182.
, and BUCHARDT, B., 1977, Depth distribution in Amphisteginain the Gulf of Elat: Utrecht Micropaleontological Bulletins, v. 15,p. 205–224.
HANZAWA, S., 1939, Revision of ‘Nummulites’ cumingii (Carpenter):Japanese Journal of Geology and Geography, v. 16, p. 223–232.
HARTL, D.L., 1980, Principles of Population Genetics: Sinauer Asso-ciates, Sunderland, Massachusetts, 488p.
HATTA, A., and UJIIE, H., 1992, Benthic Foraminifera from coral seasbetween Ishigaki and Iriomote Islands, Southern Ryukyu IslandArc, Northwestern Pacific. Part 2. Systematic descriptions of Ro-taliina: Bulletin of the College of Science, University of the Ryu-kyus, v. 54, p. 163–287.
HAUNOLD, TH.G., BAAL, CH., and PILLER, W.E., 1998, Larger Fora-minifera, in Piller, W.E., and Haunold, Th.G. (eds.), The NorthernBay of Safaga (Red Sea, Egypt): An actuopalontological ap-proach. V. Foraminifera: Abhandlungen der senckenbergischennaturforschenden Gesellschaft, v. 548, 155–180.
HEDLEY, R.H., PARRY, D.M., and WAKEFIELD, J.S.J., 1968, Repro-duction in Boderia turneri (Foraminifera): Journal of Natural His-tory, v. 2, p. 147–151.
HOFKER, J., 1927, The Foraminifera of the Siboga Expedition. Part 1.Families Tinoporidae, Rotaliidae, Nummulitidae, Amphistegini-dae: E.J. Brill Publishers, Leiden, 78p.
, 1933, Papers from Dr. Th. Mortensen’s Pacific Expedition1914–16. LXII. Foraminifera of the Malay Archipelago: Viden-skabelige Meddelelser fra Dansk naturhistorik Foreningi, v. 93, p.71–167.
HOHENEGGER, J., 1994., Distribution of living larger Foraminifera NWof Sesoko-Jima, Okinawa, Japan: P.S.Z.N. I: Marine Ecology, v.15, p. 291–334.
, 1995, Depth estimation by proportions of living larger fora-minifera: Marine Micropaleontology, v. 26, p. 31–47.
, 1996, Remarks on the distribution of larger foraminifera (Pro-tozoa) from Belau (Western Carolines): Kagoshima University Re-search Center for the South Pacific, Occasional Papers, v. 30, p.85–90.
, in press, Coenoclines of larger Foraminifera, in Lee, J.J., andHallock, P. (eds.): Biology of Foraminifera II, MicropaleontologyPress.
, YORDANOVA, E., NAKANO, Y., and TATZREITER, F., 1999,Habitats of larger foraminifera on the upper reef slope of SesokoIsland, Okinawa, Japan: Marine Micropaleontology, v. 36, p. 109–168.
HOLZMANN, M., HOHENEGGER, J., PILLER, W.E., and PAWLOWSKI, J.,1998, Molecular phylogeny of large Foraminifera based on partialSSU rDNA sequences, in Longoria, J.F., and Gamper, M.A. (eds.),Forams ’98. International Symposium on Foraminifera: SociedadMexicana de Paleontologıa, A.C., Special Publication: Monterrey,Nuevo Leon, Mexico, p. 50.
HOTTINGER, L., 1977a, Foraminiferes operculiniformes: Memoires duMuseum d’Histoire Naturelle, Nouvelle Serie, Serie C, Sciencesde la Terre, v. 40, 159pp.
, 1977b, Distribution of larger Peneroplidae, Borelis and Num-mulitidae in the Gulf of Elat, Red Sea: Utrecht Micropaleontolog-ical Bulletin, 15, p. 35–110.
, 1978, Comparative anatomy of elementary shell structures inselected larger foraminifera, in Hedley, R.H., and Adams, C.G.(eds.), Foraminifera, Volume 3: Academic Press, London, p. 203–266.
, 1983a, Processes determining the distribution of larger Fora-minifera in space and time: Utrecht Micropaleontological Bulle-tins, v. 30, p. 239–253.
, 1983b, Neritic macroid genesis, an ecological approach, inPeryt, T. (ed.), Coated Grains: Springer Verlag, Berlin, p. 38–55.
, 1997, Shallow benthic foraminiferal assemblages as signalsfor depth of their deposition and their limitations: Bulletin SocieteGeologie de France, v. 168, p. 491–505.
, HALICZ, E., and REISS, Z., 1993, Recent Foraminifera fromthe Gulf of Aqaba, Red Sea: Academia Scientiarum et ArtiumSlovenica, Classis IV: Historia Naturalis, v. 33, Ljubljana, 179 p.
KENDEIGH, S.C., 1974, Ecology with Special Reference to Animalsand Man: Prentice Hall, Englewood Cliffs, New Jersey.
KOBA, M., 1978, Distribution and environment of Recent Cyclocly-peus: Science Reports of the Tohoku University, 7th Series (Ge-ography), v. 28, p. 283–311.
KRUGER, R., 1994, Untersuchungen zur Entwicklung rezenter Num-mulitiden: Heterostegina depressa, Nummulites venosus und Cy-cloclypeus carpenteri: Sc.D. Thesis, University of Kiel, Germany,Uni Press, Hochschulschriften, v. 70, Literatur Verlag, Munsterund Hamburg, 97p.
, ROTTGER, R., LIETZ, R., and HOHENEGGER, J., 1996, Biologyand reproductive processes of the larger foraminiferan Cyclocly-peus carpenteri (Protozoa, Nummulitidae): Archiv fur Protisten-kunde, v. 147, p. 307–321.
KUWANO, Y., 1956, Invertebrate fauna of the intertidal zone of theTokara Islands. XII. Foraminifera: Publications of the Seto MarineBiological Laboratory, v. 5, p. 273–285.
LARSEN, A.R., 1976, Studies of Recent Amphistegina, taxonomy andsome ecological aspects: Israel Journal of Earth-Sciences, v. 25,p. 1–26.
LEE, J.J., 1994, Diatoms, or their chloroplasts, as endosymbiotic part-ners for foraminifera: Proceedings of the 11th International Dia-tom Symposium, Memoirs of the California Academy of Science,no 17, p. 21–36.
, and ANDERSON, O.R., 1991, Symbiosis in foraminifera, inLee, J.J., and Anderson, O.R. (eds.), Biology of Foraminifera:Academic Press, London, p. 157–220.
, MCENERY, M.E., TER KUILE, B., EREZ, J., ROTTGER, R.,ROCKWELL, R.F., FABER, W.W. JR., and LAGZIEL, A., 1989, Iden-tification and distribution of endosymbiotic diatoms in larger Fo-raminifera: Micropaleontology, v. 35, p. 353–366.
LEE, M.J., ELLIS, R., and LEE, J.J., 1982, A comparative study ofphotoadaptation in four diatoms isolated as endosymbionts fromlarger foraminifera: Marine Biology, v. 68, p. 193–197.
LEUTENEGGER, S., 1977a, Ultrastructure des foraminiferes perfores etimperfores ainsi que de leurs symbiotes: Cahiers de Micropaleon-tologie, v. 3, 52p.
, 1977b, Reproduction cycles of larger foraminifera and depthdistribution of generations: Utrecht Micropaleontological Bulletin,v. 15, p. 27–34.
LOEBLICH, A.R., and TAPPAN, H., 1988, Foraminiferal Genera andtheir Classification: Van Nostrand Reinhold Company, New York,2 vol., 970p.
, and , 1994, Foraminifera of the Sahul Shelf and TimorSea: Cushman Foundation for Foraminiferal Research, SpecialPublication No. 31, 661p.
LONGHURST, A., 1998, Ecological Geography of the Sea: AcademicPress, San Diego, 398p.
LUTZE, G.F., GRABERT, B., and SEIBOLD, E., 1971, Lebendbeobach-tungen an Groß-Foraminiferen (Heterostegina) aus dem Persi-schen Golf: Meteor-Forschungs-Ergebnisse, series C, no. 6, p. 21–40.
MAYR, E., and ASHLOCK, P.D., 1991, Principles of Systematic Zool-ogy, 2nd edition: McGraw-Hill, New York, 475p.
OKI, K., 1989, Ecological analysis of benthonic Foraminifera in Ka-goshima Bay, South Kyushu, Japan: South Pacific Study, v. 10,1–191.
D’ORBIGNY, A.D., 1826, Tableau methodique de la classe des Cephal-opodes: Annales des Sciences Naturelles, v. 7, p. 245–314.
, 1839, Foraminiferes, in Ramon de la Sagra, Histoire physique,politique et naturelle de l’Isle de Cuba, Arthus Bertrand, Paris.
PECHEUX, M.J.-F., 1995, Ecomorphology of a recent large foraminifer,Operculina ammonoides: Geobios, v. 28, p. 529–566.

28 HOHENEGGER, YORDANOVA, AND HATTA
PIGNATTI, J.S., 1998, Paleogene Larger Foraminifera. Reference List,in Drobne, K., and Hottinger, L. (eds.), Paleogene Shallow Ben-thos of the Tethys, 1: Academia Scientiarum et Artium Slovenica,Classis IV: Historia Naturalis, v. 34/1, Ljubljana, 296p.
RAUP, D.M., 1966, Geometric analysis of shell coiling: general prob-lems: Journal of Paleontology, v. 40, p. 1178–1190.
REISS, Z., and HOTTINGER, L., 1984, The Gulf of Aqaba, EcologicalMicropaleontology: Ecological Studies 50, Springer Verlag, Ber-lin, 354p.
ROGL, F., and HANSEN, H.J., 1984, Foraminifera described by Fichtel& Moll in 1798. A revision of Testacea Microscopica: Neue Denk-schriften des Naturhistorischen Museums in Wien, v. 3, 143 p.
ROTTGER, R., 1972a, Die Kultur von Heterostegina depressa (Fora-minifera: Nummulitidae): Marine Biology, v. 15, p. 150–159.
, 1972b, Analyse von Wachstumskurven von Heterostegina de-pressa (Foraminifera: Nummulitidae): Marine Biology, v. 17, p.228–242.
, 1973, Die Ektoplasmahulle von Heterostegina depressa (Fo-raminifera: Nummulitidae): Marine Biology, v. 21, p. 127–138.
, 1974, Larger Foraminifera: reproduction and early stages ofdevelopment in Heterostegina depressa: Marine Biology, v. 26,p. 5–12.
, 1976, Ecological observation of Heterostegina depressa (Fo-raminifera, Nummulitidae) in the laboratory and in its natural hab-itat: First International Symposium on Benthonic Foraminifera ofContinental Margins, Part A. Ecology and Biology, Maritime Sed-iments, Special Publication 1, p. 75–79.
, 1990, Biology of Larger Foraminifera: present status of thehypothesis of trimorphism and ontogeny of the gamont of Het-erostegina depressa: Studies in Benthic Foraminifera. Proceed-ings of the Fourth International Symposium on Benthic Forami-nifera, Sendai 1990, p. 43–54.
, and BERGER, W., 1972, Benthic foraminifera: morphology andgrowth in clone cultures of Heterostegina depressa: Marine Bi-ology, v. 15, p. 89–94.
, and HALLOCK, P., 1982, Shape trends in Heterostegina de-pressa (Protozoa, Foraminiferida): Journal of Foraminiferal Re-search, v. 12, p. 197–204.
, and INSTITUT FUR DEN WISSENSCHAFTLICHEN FILM, 1982,Die Großforaminifere Heterostegina depressa. Organisation undWachstum der megalospharischen Generation: Film 1451 of IWF,Gottingen. Publikationen zu wissenschaftlichen Filmen, SektionBiologie, ser. 15, no. 21, 15p.
, and , 1984, Die Großforaminifere Heterostegina de-pressa. Vielteilung der mikrospharischen und der megalosphar-ischen Generation: Film 1506 of IWF, Gottingen. Publikationenzu wissenschaftlichen Filmen, Sektion Biologie, ser. 16, no. 28,20p.
, and SPINDLER, M., 1976, Development of Heterostegina de-pressa individuals (Foraminifera, Nummulitidae) in laboratorycultures: First International Symposium on Benthonic Foraminif-era of Continental Margins, Part A. Ecology and Biology, Mari-time Sediments, Special Publication 1, p. 81–87.
, DETTMERING, CH., KRUGER, R., SCHMALJOHANN, R., andHOHENEGGER, J., 1998, Gametes in Nummulitids (Foraminifera):Journal of Foraminiferal Research, v. 28, p. 345–348.
, FLADUNG M., SCHMALJOHANN, R., SPINDLER, M., and ZACH-ARIAS, H., 1986, A new hypothesis: The so-called megalosphericschizont of the larger foraminifer, Heterostegina depressad’Orbigny, 1826, is a separate species: Journal of ForaminiferalResearch, v. 16, p. 141–149.
, IRWAN, A., SCHMALJOHANN, R., and FRANZISKET, L., 1980,Growth of the symbiont-bearing foraminifera Amphistegina les-sonii d’Orbigny and Heterostegina depressa d’Orbigny (Proto-zoa), in Schwemmler, W., and Schenk, H.E.A. (eds.), Endocyto-biology, Endosymbiosis and Cell Biology: Walter de Gruyter, Ber-lin, p. 125–132.
, KRUGER, R., and DE RIJK, S., 1990, Trimorphism in Fora-minifera (Protozoa) — Verification of an old hypothesis: EuropeanJournal of Protistology, v. 25, p. 226–228.
, SCHMALJOHAN, R., and ZACHARIAS, M., 1989, Endoreplica-tion of zygotic nuclei in the larger Foraminifera Heterostegina
depressa (Nummulitidae): European Journal of Protistology, v. 25,p. 226–228.
, SPINDLER, M., SCHMALJOHAN, R., RICHWIEN, M., and FLAU-DUNG, M., 1984, Functions of the canal system in the rotaliidforaminifera Heterostegina depressa: Nature, v. 309, p. 789–791.Erratum note, 1985: Nature, v. 315, p. 77.
ROUGHGARDEN, J., 1974, The fundamental and realized niche of asolitary population: The American Naturalist, v. 108, p. 232–235.
SCHAUB, H., 1981, Nummulites et Assilines de la Tethys paleogene.Taxonomie, phylogenese, biostratigraphie: Memoires Suisses dePaleontologie, vols. 104–106, 236p.
SERRA-KIEL, J., HOTTINGER, L., CAUS, E., DROBNE, K., FERRANDEZ,C., JAUHRI, A.K., LESS, G., PAVLOVEC, R., PIGNATTI, J., SAMSO,J.M., SCHAUB, H., SIREL, E., STROUGO, A., TAMBAREAU, Y., TO-SOUELLA, J., and ZAKREVSKAYA, E., 1998, Larger foraminiferalbiostratigraphy of the Tethyan Paleocene and Eocene: Bulletin So-ciete Geologie de France, v. 169, p. 281–299.
SIMPSON, G.G., 1951, The species concept: Evolution, v. 5, p. 285–298.
, 1961, Principles of Animal Taxonomy: Columbia UniversityPress, New York, 247p.
SPINDLER, M., 1978, Anatomy of the test of Heterostegina depressa(Foraminiferida): Journal of Foraminiferal Research, v. 8, p. 319–331.
, and ROTTGER, R., 1974, Der Kammerbauvorgang derGroßforaminifere Heterostegina depressa (Nummulitidae): Ma-rine Biology, v. 18, 146–159.
TITLYANOV, E.A., and LATYPOV, Y.Y., 1991, Light-dependence inscleractinian distribution in the sublittoral zone of South ChinaSea Islands: Coral Reefs, v. 10, p. 133–138.
UCHIO, T., 1962, Recent foraminifera thanatocoenoses of beach andnearshore sediments along the coast of Wakayama-Ken, Japan:Publications of the Seto Marine Biological Laboratory, v. 10, p.133–144.
, 1968, Foraminiferal assemblages in the vicinity of the SetoMarine Biological Laboratory, Shirahama-Cho, Wakayama-Ken,Japan (Part 1): Publications of the Seto Marine Biological Labo-ratory, v. 150, p. 399–417.
UJIIE, H., and HATTA, A., 1995, Quantitative analyses of benthic fo-raminiferal assemblages from southern Ryukyu Island Arc, sub-tropical northwestern Pacific Ocean: Journal of Foraminiferal Re-search, 25, p. 334–349.
VAN VALEN, L., 1976, Ecological species, multispecies and oaks: Tax-on, v. 25, p. 233–239.
WHITTAKER, R.H., 1973, Direct gradient analysis: Techniques, inWhittaker, R.H. (ed.), Handbook of Vegtation Science 5. Ordina-tion and Classification of Communities: Dr. W. Junk, The Hague,p. 9–51.
WILEY, E.O., 1981, Phylogenetics. The Theory and Practice of Phy-logenetic Systematics: John Wiley and Sons, New York-Chich-ester-Brisbane-Toronto, 439p.
YABE, H., 1918, Notes on Operculina-rocks from Japan, with remarkson ‘Nummulites’ cumingii CARPENTER: Scientific Reports ofthe Tohoku Imperial University, 2nd series (Geology), v. 4, p.105–126.
YAMAZATO, K., KAMURA, M, and NISHIJIMA, N., 1968, The distri-bution of benthos on the coast of the Ryukyu Islands, in An In-vestigative Report of Ryukyu Fisheries Resources: Agricultureand Forestry Bureau, Ryukyu Government, Naha, p. 164–192.
YORDANOVA, E., 1998, Beziehungen zwischen lebenden Individuenund Totengemeinschaften von Großforaminiferen am RiffhangNW von Sesoko, Okinawa, Japan: Sc.D. Thesis, University ofVienna, Austria, 217p.
ZHENG, S.Y., 1979, The Recent Foraminifera of the Xisha Island,Guangdong Province, China, II: Studia Marina Sinica, v. 15, p.1–27 (in Chinese).
ZOHARY, T., REISS, Z., and HOTTINGER, L., 1980, Population dynamicsof Amphisorus hemprichii (Foraminifera) in the Gulf of Elat (Aq-aba), Red Sea: Eclogae Geologicae Helveticae, v. 73, p. 1071–1094.
Received 22 April 1999Accepted 27 August 1999





























