Embed Size (px)
Citation preview

ORIGINAL PAPER
Mark I. Stevens Æ Arne Fjellberg Æ Penelope Greenslade
Ian D. Hogg Æ Paul Sunnucks
Redescription of the Antarctic springtail Desoria klovstadiusing morphological and molecular evidence
Received: 22 June 2005 / Revised: 13 December 2005 / Accepted: 27 January 2006 / Published online: 25 April 2006� Springer-Verlag 2006
Abstract Isotoma klovstadi Carpenter, 1902 was one ofthe first Collembola described from the Antarctic con-tinent. It was first collected in November 1899 during theBritish Antarctic Expedition on the north coast of Vic-toria Land in the Ross Sea region. It is now known tooccur in an extensive area of northern Victoria Land,including the offshore Possession, Coulman, and FoynIslands. More recently, I. klovstadi was moved to thegenus Gnathisotoma Cassagnau, 1957 and has been in-cluded in this genus in an unpublished checklist (online)of all described Collembola. Here, we redescribe thespecies and use morphological and molecular (COI and18S genes) evidence to investigate its affinities within theIsotominae. We show that it does not belong to Gnath-isotoma, or Isotoma s. str. (the viridis group) as currentlyconceived, but is likely to be part of the species complexof Isotoma s. lat. We discuss reasons for placing it in thegenus Desoria Nicolet, 1841. Our results reinforce the
already high level of endemicity in the Antarctic faunaand emphasise the value of both morphological andmolecular studies in examining relict Gondwanan taxaand their evolutionary relationships with those of otherSouthern Hemisphere continents.
Abbreviations Ant.: Antenna or antennal ÆAbd.: abdomen Æ PAO: postantennal organ
Introduction
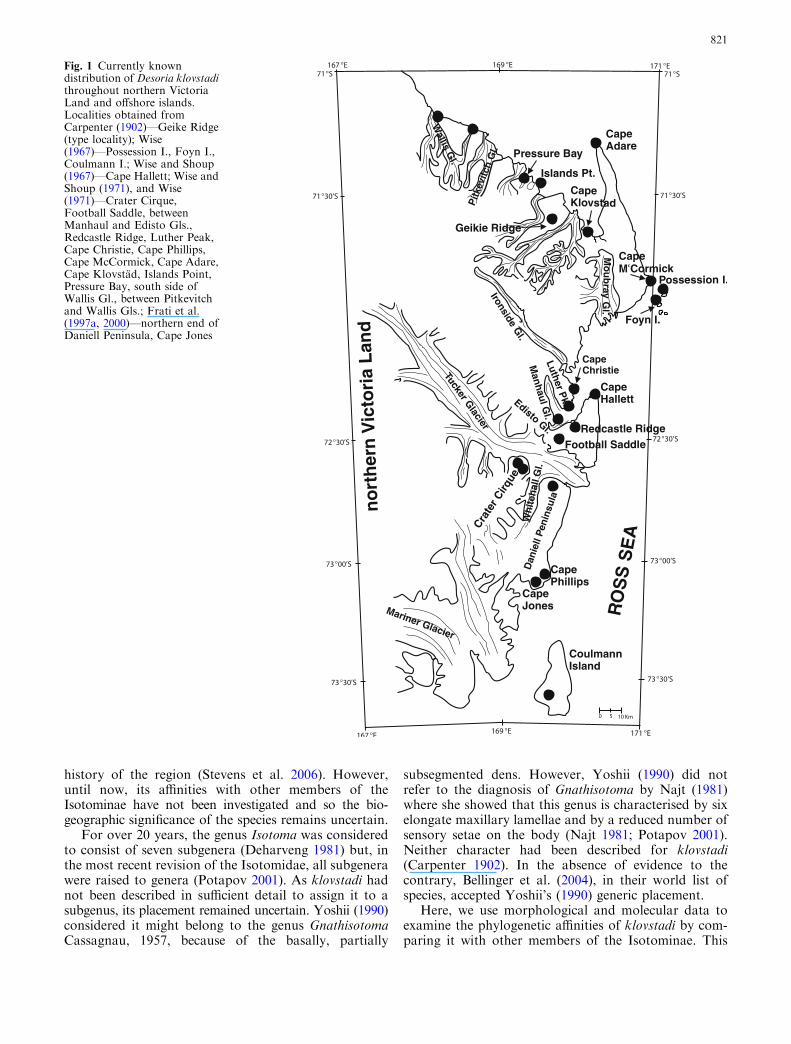
In November 1899, during the Southern Cross BritishAntarctic Expedition, 1898–1900, Herluf Klovstadcollected Collembola on Geikie Ridge at the head ofRobertson Bay on the northern coast of Victoria Landin the Ross Sea region (Fig. 1). Carpenter (1902)assigned the specimens to the genus Isotoma based onthe ‘forwardly positioned head, the close equality inlength between the third and fourth abdominal segmentsand the entire absence of scales’, and described thespecies as Isotoma klovstadi n. sp. (Fig. 2). Numerouscollections of the species have been made since 1899.The species is now known to be restricted to northernVictoria Land and the offshore Possession, Coulman,and Foyn Islands, covering an area 250·150 km with itssouthern limit marked by the Mariner Glacier (Wise1967; Wise and Shoup 1967; Wise 1971; Wise and Shoup1971; Frati and Carapelli 1999) (Fig. 1). The species isconsidered endemic to this small region of EasternAntarctica where 60% (6) of collembolan genera and90% (9) of species are endemic (Wise 1967, 1971;Greenslade 1995; Stevens and Hogg 2006a). Like mostCollembola in this region, it is likely to be a relict thathas survived glaciation in refugia (see Wise 1971; Fratiet al. 2001; Stevens and Hogg 2006a, b; Stevens et al.2006). As one of the six endemic isotomid species inEastern Antarctica, it has the potential to increase ourunderstanding of the biogeography and evolutionary
M. I. Stevens (&)Allan Wilson Centre for Molecular Ecology & Evolution,Massey University, Private Bag 11-222, Palmerston North,New ZealandE-mail: [email protected].: +64-6-3569099Fax: +64-6-3505626
P. Sunnucks Æ M. I. StevensDepartment of Genetics, La Trobe University,3086 Bundoora, VIC, Australia
Present address: M. I. Stevens Æ P. SunnucksSchool of Biological Sciences, Monash University,3800 Clayton, VIC, Australia
A. FjellbergZoological Museum, Lund University, 223 62 Lund, Sweden
P. GreensladeDepartment of Botany and Zoology, Australian NationalUniversity, 0200 Australian Capital Territory, Australia
I. D. HoggCentre for Biodiversity and Ecology Research,University of Waikato, Private Bag 3105,Hamilton, New Zealand
Polar Biol (2006) 29: 820–830DOI 10.1007/s00300-006-0120-y
history of the region (Stevens et al. 2006). However,until now, its affinities with other members of theIsotominae have not been investigated and so the bio-geographic significance of the species remains uncertain.
For over 20 years, the genus Isotoma was consideredto consist of seven subgenera (Deharveng 1981) but, inthe most recent revision of the Isotomidae, all subgenerawere raised to genera (Potapov 2001). As klovstadi hadnot been described in sufficient detail to assign it to asubgenus, its placement remained uncertain. Yoshii (1990)considered it might belong to the genus GnathisotomaCassagnau, 1957, because of the basally, partially
subsegmented dens. However, Yoshii (1990) did notrefer to the diagnosis of Gnathisotoma by Najt (1981)where she showed that this genus is characterised by sixelongate maxillary lamellae and by a reduced number ofsensory setae on the body (Najt 1981; Potapov 2001).Neither character had been described for klovstadi(Carpenter 1902). In the absence of evidence to thecontrary, Bellinger et al. (2004), in their world list ofspecies, accepted Yoshii’s (1990) generic placement.
Here, we use morphological and molecular data toexamine the phylogenetic affinities of klovstadi by com-paring it with other members of the Isotominae. This
Fig. 1 Currently knowndistribution of Desoria klovstadithroughout northern VictoriaLand and offshore islands.Localities obtained fromCarpenter (1902)—Geike Ridge(type locality); Wise(1967)—Possession I., Foyn I.,Coulmann I.; Wise and Shoup(1967)—Cape Hallett; Wise andShoup (1971), and Wise(1971)—Crater Cirque,Football Saddle, betweenManhaul and Edisto Gls.,Redcastle Ridge, Luther Peak,Cape Christie, Cape Phillips,Cape McCormick, Cape Adare,Cape Klovstad, Islands Point,Pressure Bay, south side ofWallis Gl., between Pitkevitchand Wallis Gls.; Frati et al.(1997a, 2000)—northern end ofDaniell Peninsula, Cape Jones
821

combined approach is critical in cases when phenotypesare shared because of convergence (homoplasy) ratherthan homology. We selected two genes that have beenused extensively for phylogenetic reconstructions, 18Snuclear ribosomal RNA (18S rRNA) and mitochondrialDNA cytochrome c oxidase I (COI). The 18S rRNAgene has been used for analysing the phylogenetic rela-tionships of organisms, and the V4 region has beenfound to provide useful phylogenetic information attaxonomic levels similar to those under investigationhere (e.g. Hillis and Dixon 1991; Black et al. 1997;Giribet and Ribera 2000). Sequence analysis of the COIgene has been a cornerstone of phylogenetic and phy-logeographic studies for the past decade (e.g. Folmeret al. 1994; Simon et al. 1994; Soto-Adames 2002;Stevens and Hogg 2003; Garrick et al. 2004). Further-more, protein coding genes such as COI offer easy align-ment and taxonomic analysis of nucleotide sequences (e.g.Hogg and Hebert 2004). The combination of a bi-parentally inherited nuclear gene (18S rRNA) and amaternally-inherited mtDNA gene (COI) is particularlyinformative as it provides independent sequences withseparate modes of inheritance and different expectedrates of evolution and lineage sorting (Hillis and Dixon1991; Avise 2004).
Our data suggest that Isotoma klovstadi does notbelong either to Gnathisotoma or to Isotoma s. str. (theviridis group) as currently conceived. Reasons for nowplacing the species klovstadi in the genus Desoria Nico-let, 1841 are discussed and the species will be referred toas Desoria klovstadi.
Morphological description
Family ISOTOMIDAE Schaffer, 1896Sub-family ISOTOMINAE Schaffer, 1896Genus Desoria Nicolet, 1841
Diagnosis (after Potapov 2001): Isotominae with 6–8ocelli; thickened apical setae on the manubrium absent;abdomen (Abd.) V and VI separate, rarely fused; ante-rior of ventral tube with setae, tibiotarsus without cla-vate hairs; claw usually with inner tooth; furca long withtapering crenulated dens, mucro with three or four teeth,with or without setae; normally with six or more accp s’setae (sensilla) on body segments (Fig. 3a).
Desoria klovstadi (Carpenter, 1902)Isotoma klovstadi Carpenter, 1902Gnathisotoma klovstadi (Carpenter, 1902)
Specimens examined: Several specimens collected byB. Reid (in 1959), three adults and three juveniles(probably second instar) collected by T.G.A. Green, R.Seppelt, and C. Beard (31 January 2004). All specimensfrom Cape Hallett (72�19¢11.5¢¢S, 170�14¢2.56¢¢E) andvouchers stored in the A. Fjellberg Collection, Tjome,Norway.
Description of species
Habitus colour and clothing
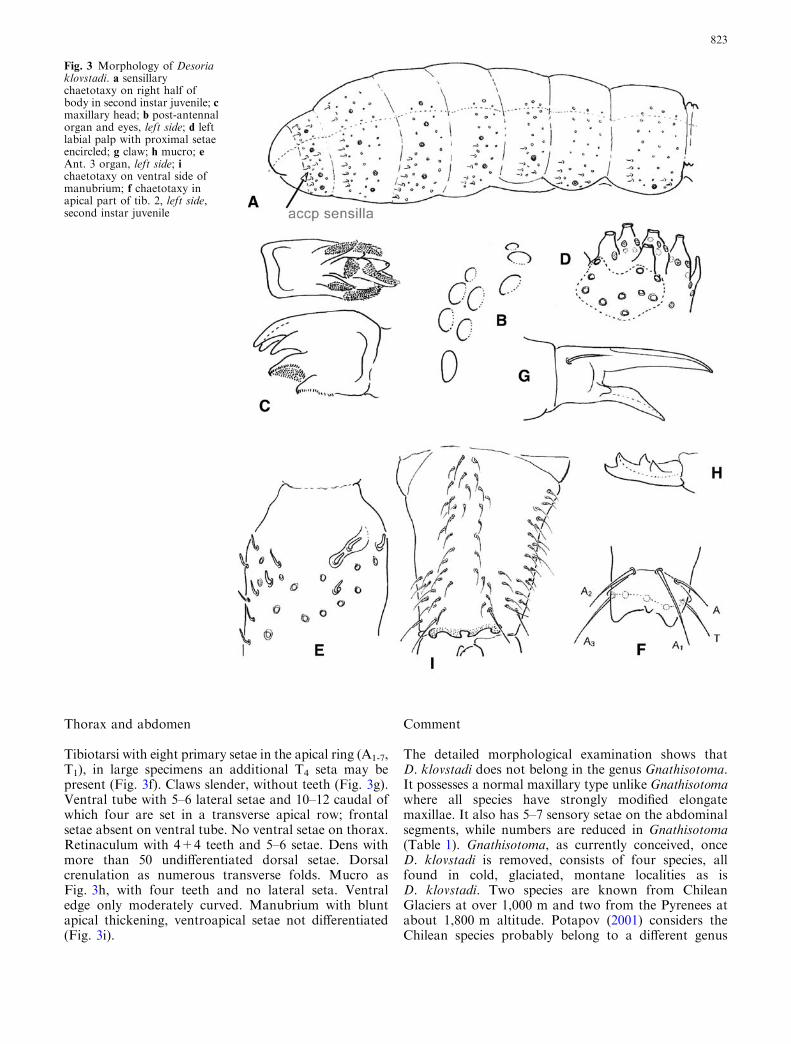
Largest specimen 2.0 mm. Body brownish or oliveblack, ventral side of head and extremities paler. Bodyshape elongate, Isotoma-like (Fig. 2). Body covered withfine, smooth setae. Macrochaetae moderately developed,those of Abd. 4–6 twice as long as surrounding ordinarysetae. Median macrochaeta on abd.5 about 0.7–0.8 aslong as tergite. Abd. 5–6 not fused. Sensilla (s’ setae) oftergites short, like the short sensilla on antennae. Sens-illary formula variable, normal condition of small juve-niles (0.5 mm) appears to be 66/56676 of which 11/1111are spine-like microsensilla (Fig. 3a).
Head and antennae
Antenna (Ant.) 1 with 4-5 hair-like ventral sensilla andone short blunt sensillum. Ventral and lateral sides ofAnt. 2 and 3 with many short, blunt, curved sensilla.Ant. 3 organ as Fig. 3e. Ant. 4 with many short and longhair-like sensilla. Subapical pin-seta of Ant. 4 simple.Head with 6+6 postlabial setae along ventral line. Eyes8+8, G & H smaller than the other six. PAO oval, 1.5 aslong as diameter of nearest ocellus (Fig. 3b). Maxillaryouter lobe bifurcate, 4 sublobal hairs. Maxilla with3-toothed capitulum, lamellae short, undifferentiated, alldensely packed with small denticles. Marginal ciliationshort, not much longer than denticles (Fig. 3c). Man-dibles normal. Labrum with 5-5-4 undifferentiated setae,apical edge with four roundish folds, ventroapical cili-ation brush-like, composite. Labial palp with a full set ofpapillae and guards, e7 present. Proximal setae 8–10(Fig. 3d). Basomedian field with seven setae, basolateralfield with five setae. Frontoclypeal field with more than30 setae, prelabral setae 4.
Fig. 2 Syncroscopy image of an adult Desoria klovstadi, length ofspecimen is 1.9 mm (image by B. O’Brien). This specimen wascollected at Cape Hallett in January 2002
822
Thorax and abdomen
Tibiotarsi with eight primary setae in the apical ring (A1-7,T1), in large specimens an additional T4 seta may bepresent (Fig. 3f). Claws slender, without teeth (Fig. 3g).Ventral tube with 5–6 lateral setae and 10–12 caudal ofwhich four are set in a transverse apical row; frontalsetae absent on ventral tube. No ventral setae on thorax.Retinaculum with 4+4 teeth and 5–6 setae. Dens withmore than 50 undifferentiated dorsal setae. Dorsalcrenulation as numerous transverse folds. Mucro asFig. 3h, with four teeth and no lateral seta. Ventraledge only moderately curved. Manubrium with bluntapical thickening, ventroapical setae not differentiated(Fig. 3i).
Comment
The detailed morphological examination shows thatD. klovstadi does not belong in the genus Gnathisotoma.It possesses a normal maxillary type unlike Gnathisotomawhere all species have strongly modified elongatemaxillae. It also has 5–7 sensory setae on the abdominalsegments, while numbers are reduced in Gnathisotoma(Table 1). Gnathisotoma, as currently conceived, onceD. klovstadi is removed, consists of four species, allfound in cold, glaciated, montane localities as isD. klovstadi. Two species are known from ChileanGlaciers at over 1,000 m and two from the Pyrenees atabout 1,800 m altitude. Potapov (2001) considers theChilean species probably belong to a different genus
Fig. 3 Morphology of Desoriaklovstadi. a sensillarychaetotaxy on right half ofbody in second instar juvenile; cmaxillary head; b post-antennalorgan and eyes, left side; d leftlabial palp with proximal setaeencircled; g claw; h mucro; eAnt. 3 organ, left side; ichaetotaxy on ventral side ofmanubrium; f chaetotaxy inapical part of tib. 2, left side,second instar juvenile
823

because only one maxillary lamella is elongated com-pared to six in the species from the Pyrenees.
Desoria klovstadi does not show strong similarities toIsotoma in the narrow sense (i.e. the viridis group,Fjellberg 2003; see also Simonsen et al. 1999) notably inthe absence of anterior setae on the ventral tube, the lackof teeth on the claw and the absence of spines on theventroapical face of the manubrium (Table 1). Desoriaklovstadi has a long and slender mucro with four sub-equal teeth, while in the viridis group it is short, compactwith three subequal subapical teeth and a smaller apicaltooth. Also the claw inner edge in the viridis group hastwo inner teeth while D. klovstadi claw has no teeth. Theabdominal macrochaeta on abd. V are much shorterthan the median length of the tergite and smooth inD. klovstadi, whereas in the viridis group they are abouttwice as long as the tergite and serrated.
The Antarctic species is most similar to Desoria inthese characters, as currently conceived, except in theabsence of anterior setae on the ventral tube and thediffering number of tibiotarsal setae (Table 1). MostDesoria species where this character is known, includingthe type of the genus Desoria saltans Nicolet, 1841, havea complete set of apical tibiotarsal setae (11 setae,including T1–4), but D. klovstadi has only eight primarysetae, with an additional ninth (T4) in large specimens.In this character D. klovstadi differs from Desoria s. str.,but this setal pattern is found in several other genera;Isotoma s. str., Halisotoma, Isotomurus, Marisotoma,Heteroisotoma, and some Parisotoma, for instanceP. trispinata MacGillivray, 1896. This latter species isincluded in Desoria by Potapov (1991). Most Parisotoma,including the southern Parisotoma insularis Deharveng,1981, have only seven apical setae (T-setae absent) asdoes the southern Folsomotoma punctata Wahlgren,
1906. The significance of the increased number ofproximal setae in the labial palp in D. klovstadi isuncertain. This character is shared with Archisotoma,Agrenia, Hydroisotoma and Isotomurus and is possiblyan adaptive character related to living in a moist habitat,so may not reflect a close phylogenetic relationship.
Molecular analyses
Taxa examined
We selected several morphologically appropriate generafrom the Isotominae Schaffer, 1896 to include in the in-group (Isotomurus Borner, 1903, PseudisotomaHandschin,1924, Isotomiella Bagnall, 1939, Isotoma Bourlet, 1839,Desoria Nicolet and Desor, 1841, Parisotoma Bagnall,1940, Folsomotoma Bagnall, 1949, Vertagopus Borner,1906) in order to compare their relationship withD. klovstadi (Table 2). Phylogenetic analyses were rootedusing the genus Subisotoma Stach, 1947 (Subisotoma sp.,location Tasmania, Australia) an isotomid in the sub-family Anurophorinae. All sequences are accessible fromGenBank (Table 2).
DNA extraction and analyses
Total genomic DNA was extracted from entire individ-uals (preserved in 95% ethanol) using a ‘salting out’technique (Sunnucks and Hales 1996). Polymerase chainreaction (PCR) amplification (Saiki et al. 1988) wascarried out using a 25 ll reaction volume consisting of3 ll extracted DNA, 1· PCR buffer (Roche), 2.2 mMMgCl2, 0.2 mM of each dNTP (Boehringer Mannheim),
Table 1 Morphological characters that differ between Desoria klovstadi, Desoria s. str., Isotoma s. str. (viridis group), and Gnathisotoma(Chile and Pyrenees spp.)
Characters Desoriaklovstadi
Desoria s. str. Isotoma s. str.(viridis group)
Gnathisotoma(Chile)
Gnathisotoma(Pyrenees)
Source This work Fjellberg Fjellberg, Potapov Yosii NajtNo. of ocelli 8+8 6+6–8+8 8+8 5+5 (in text) 6+6
(in figures)6+6
PAO shape Size cf. ocellus Oval 1.5 Variable Smaller thanocellus, roundish
Elliptical 1.5 Elliptical 1.5
Labial papillae Normal Variable Normal Curved, strong Normal?Number of elongated maxillary lamellae None Variable None 1 6Maxillary teeth elongated No Variable No Yes, 2 slightly Yes, 2External lobe of maxilla (shape of palp+ number of sublobal hairs)
Bifurcate+4 Variable Bifurcate 2/i+3+1 long seta 1 long seta
Ventral tube, anterior setae None Present Present None? NoneTibiotarsal setae (distal row) 8–9 11 8–10 Unknown UnknownTeeth on claw Absent Variable 2 inner, 2 outer Absent VariableSetae on ventro-apicalface of manubrium
Many simple Many, simple Spine-like, thickened Many, simple Few, simple
Mucronal shape Long, slender Variable Short compact Medium Short, compactFusion of abdominal segments V and VI Separate Variable Separate Fused laterally SeparateLength of macrochaetae cf. toLength of abdominal segment V
Shorter Shorter or longer Longer·2 Shorter Shorter?
Sensory setae per tergite 5–7 accp Variable >6 accp Unknown 2–3 accp 1–2 al
824
1.0 lM of each primer, and 0.2 U of Taq DNA poly-merase (Roche) on a Biometra T1 thermocycler(Whatman Biometra). PCR amplification of a 710 bpfragment of COI was carried out using the universalprimers LCO1490 (5¢-ggt caa caa atc ata aag ata ttg g-3¢)and HCO2198 (5¢-taa act tca ggg tga cca aaa aat ca-3¢)(Folmer et al. 1994). The thermal cycling conditionswere: 94�C for 1 min followed by five cycles of dena-turation and polymerase amplification (94�C for 1 min,45�C for 1.5 min and then 1 min at 72�C) and followedby 35 cycles of 94�C for 1 min, 51�C for 1.5 min andthen 1 min at 72�C, followed by 5 min at 72�C.Amplification of a 500 bp fragment of rDNA 18S wasconducted using the primers 18S-1F (5¢-ata ttg gag ggcaag tct gg-3¢) and 18S-1R (5¢-tgg cat cgt tta tgg tta g-3¢)(Otto and Wilson 2001). The thermal cycling conditionswere: 94�C for 1 min followed by 40 cycles of 94�C for20 s, 50�C for 30 s and then 90 s at 72�C, followed by5 min at 72�C. All reaction products were purified byusing SAPEXO (USB Corp.), or if necessary band-excised using the Perfectprep gel cleanup kit (Eppendorf).Purified PCR products were sequenced (utilising theforward and reverse primers) directly using the BigDyeTerminator chemistry (Applied Biosystems). Sequencingwas performed on a capillary ABI3730 genetic analyser(Applied Biosystems) at the Allan Wilson CentreGenome Service, Massey University.
Sequences were verified as being consistent with col-lembolan DNA using the GenBank BLAST algorithm,which showed high similarity with other collembolanand insect sequences. Cytochrome oxidase sequenceswere also checked for open reading frames, the presenceof which counts against the occurrence of nuclear copies(Sunnucks and Hales 1996). Sequences were alignedusing SEQUENCHER (Gene Codes ver. 4.5) sequenceeditor, and analysed using PAUP* 4.0b10 (Swofford2002). We used v2-tests, as implemented in PAUP*to determine whether the assumption of equal basefrequencies among sequences was violated for allsites, parsimony-informative sites, and for the mtDNA(COI) data set we also tested third codon positions.Modeltest ver. 3.7 (Posada and Crandall 1998) wasused to determine the appropriate model parameters formaximum likelihood (ML) heuristic searches. Themodel selected for the COI data set was TVM + I + C(�ln L = 4,333.27; rate matrix: A–C=0.0001, A–G=4.9023, A–T=0.2921, C–G=1.0174, C–T=4.9024, G–T=1.0000; I=0.2232, C=0.1910; with base frequencies setto A=0.3342, C=0.1920, G=0.1175, T=0.3564), andthe model selected for the rRNA (18S) data set wasK80+C (-ln L=957.22; with equal base frequencies;ti/tv=3.2108; C=0.1559); all other options in PAUP*remained as default for the ML and maximum-parsi-mony (MP) (branch and bound) analyses. Confidence in
Table 2 Collembola (Isotomidae) used in the COI or 18S analyses (or both)
Species Location Habitat NCBI accession numbers
COI 18S
Desoria klovstadi (Carpenter, 1902) Cape Hallett, northernVictoria Land
Moss DQ285297, DQ365783–DQ365786
DQ365770–DQ365772
Desoria tshernovi (Martynova, 1974) Somerset I., Canadian arctic Wet moss AY665341a DQ365773Desoria germanica (Huther &Winter, 1961)
Norway, Vestfold, Tjome On snow, meadows DQ365787, DQ365788
Desoria tigrina (Nicolet, 1842) Norway, Vestfold, Tjome Compost DQ365789, DQ365790Desoria olivacea (Tullberg, 1871) Norway, Akershus,
TappenbergvannSphagnum bog DQ365792 DQ365775
Desoria olivacea (Tullberg, 1871) Greenland, Nigerdlikasik,Kvanefjord
Snowbed DQ365791 DQ365774
Folsomotoma punctata(Wahlgren, 1906)
Macquarie Island, Garden Cove Colobanthus DQ365777
Isotoma anglicana Lubbock, 1862 Cornwallis I., Canadian arctic Rock AY383534a
Isotoma viridis Bourlet, 1839 Braunschweig, Germany Soil sample AJ605706b
Isotomiella minor (Schaffer, 1896)Yosii, 1939
Greenland, Equaliut, Bjornsund Snowbed DQ365793 DQ365781
Isotomurus antennalisBagnall, 1940
Sweden, Oland Wet moss DQ365776
Isotomurus sp. Igloolik I., Canadian arctic Lake strand line AY665352a
Parisotoma insularisDeharveng, 1981
Macquarie Island, Garden Cove Colobanthus DQ365794 DQ365780
Parisotoma notabilis(Schaffer, 1896)
Norway, Vestfold, Tjome Beach meadow DQ365779
Pseudisotoma sensibilis(Tullberg, 1871)
Greenland, Maniitsoq Snowbed DQ285296 DQ365778
Subisotoma sp. Pier Tier, Tasmania Moss DQ365795 DQ365782Vertagopus brevicaudus(Carpenter, 1900)
Cornwallis I., Canadian arctic Wet moss AY665354a
aGenBank (NCBI) accession numbers from Hogg and Hebert (2004)bGenBank (NCBI) accession numbers from Czarnetzki and Tebbe (2004)
825

the cladistic analyses was assessed by estimation of theg1 skewness statistic from 100,000 random tree lengths(Hillis and Huelsenbeck 1992), and bootstrap analyseswere conducted using 500 pseudoreplicates (Felsenstein1985).
Results
Mitochondrial (COI) gene
A 586-bp fragment (195 codons) was used in theseanalyses. No insertions, deletions, or stop codons weredetected. The nucleotide composition of all sequenceswas biased towards A and T (A=27%, T=34%,C=21%, G=18%). Among the 19 Isotomidae sequencesthere were 318 constant sites, 34 variable sites that wereparsimony uninformative, 234 parsimony-informativenucleotide substitutions, and sequence divergence(uncorrected distances) ranged from 15.4 to 27.5%between species (Table 3). We were unable to reject thehypothesis of homogeneity of base frequencies amongsequences for all sites (v254=42.28, P>0.05), but thiswas rejected using only the 234 parsimony-informativesites (v254=112.62, P<0.001), and using only thirdcodon positions (v254=138.37, P<0.05). The logDet/Paralinear distance tree (using a proportion of variablesites sensu Lockhart et al. 1994) had similar topology toa neighbour-joining phenogram (using all sites and alsoby removing the third codon positions), as well as to thetopologies constructed using MP and ML analyses (besttree 4,055.41). We also examined both transitions andtransversions relative to sequence divergence, in additionto the first and second codon positions relative to thethird position, which indicated that saturation does notappear to significantly affect the results (at the thirdcodon position or for ti or tv). Because base compositiondid not affect the topologies, we present only the MPanalyses using all three codon positions. The parsimony
analysis found ten most parsimonious trees. The MPphylogram (Fig. 4) was found to have similar topologiesto the MP consensus, NJ and ML analyses. There wasevidence for strong phylogenetic signal (g1=�0.816,gcrit=�0.09, P<0.01) but bootstrap values were gener-ally low among genera (Fig. 4).
Three Desoria spp. (tshernovi, germanica, olivacea)with D. klovstadi form a clade that was sister toVertagopus brevicaudus, I. anglicana, and Isotomurus sp.The genera Parisotoma, Isotomiella, Pseudisotoma, andD. tigrina were all more distant to D. klovstadi. Althoughbootstrap support was low, there were no trees (evenunder different model parameters, e.g. (un)correcteddistance (ML, NJ), K2-P, or LogDet) that would sup-port a closer association of D. klovstadi with Isotomaanglicana Lubbock, 1862. In addtion, we tested the MLtree (not shown) against a tree where D. klovstadiwas constrained to form a monophyletic group withI. anglicana. Forcing D. klovstadi with I. anglicana didnot result in a significantly worse log likelihood score forthe ML analyses (Shimodaira–Hasegawa test; P=0.23),but the unconstrained tree was identified with a lowerlog likelihood score.
Nuclear (18S rRNA) gene
The alignment of the 18S rRNA (V4 region) sequenceswas relatively straightforward. Some sequences variedin length, which was neither species- nor genus-specific,and relates to the variable length of loops in the gene. Thealignment used here includes only unambiguous sites (i.e.all sites with no insertions and/or deletions detected).
A 410-bp fragment was used in these analyses. Thenucleotide composition of all sequences was onlyslightly biased for A and T (A=29%, T=24%,C=28%, G=19%). Among the 14 sequences there were357 constant sites, 32 variable sites that were parsimonyuninformative, 21 parsimony-informative nucleotide
Table 3 Sequence divergence values (based on uncorrected distances) for the Collembola (Isotomidae) species used in the COI analysis
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
1 Desoria klovstadi2 0.0053 0.005 0.0034 0.009 0.007 0.0075 0.005 0.003 0.000 0.0076 Desoria tshernovi 0.184 0.179 0.179 0.186 0.1797 Desoria germanica 0.177 0.172 0.172 0.174 0.172 0.1818 Desoria olivacea (G) 0.184 0.181 0.183 0.181 0.183 0.201 0.1559 Desoria olivacea (N) 0.186 0.183 0.184 0.183 0.184 0.200 0.154 0.00310 Desoria tigrina 0.254 0.249 0.249 0.247 0.249 0.270 0.234 0.241 0.23911 Isotoma anglicana 0.189 0.188 0.184 0.189 0.184 0.200 0.186 0.162 0.164 0.23512 Isotomurus sp. 0.213 0.208 0.208 0.212 0.208 0.203 0.210 0.200 0.203 0.275 0.18613 Vertagopus brevicaudus 0.189 0.188 0.184 0.184 0.184 0.198 0.188 0.183 0.179 0.258 0.181 0.19814 Pseudisotoma sensibilis 0.215 0.212 0.212 0.213 0.212 0.229 0.200 0.184 0.181 0.261 0.196 0.206 0.19815 Isotomiella minor 0.235 0.234 0.230 0.232 0.230 0.227 0.210 0.213 0.210 0.247 0.213 0.218 0.241 0.21816 Parisotoma insularis 0.220 0.215 0.218 0.218 0.218 0.229 0.188 0.198 0.198 0.247 0.212 0.217 0.215 0.220 0.22017 Subisotoma sp. 0.217 0.212 0.215 0.213 0.215 0.215 0.203 0.220 0.218 0.275 0.234 0.206 0.217 0.217 0.220 0.210
826

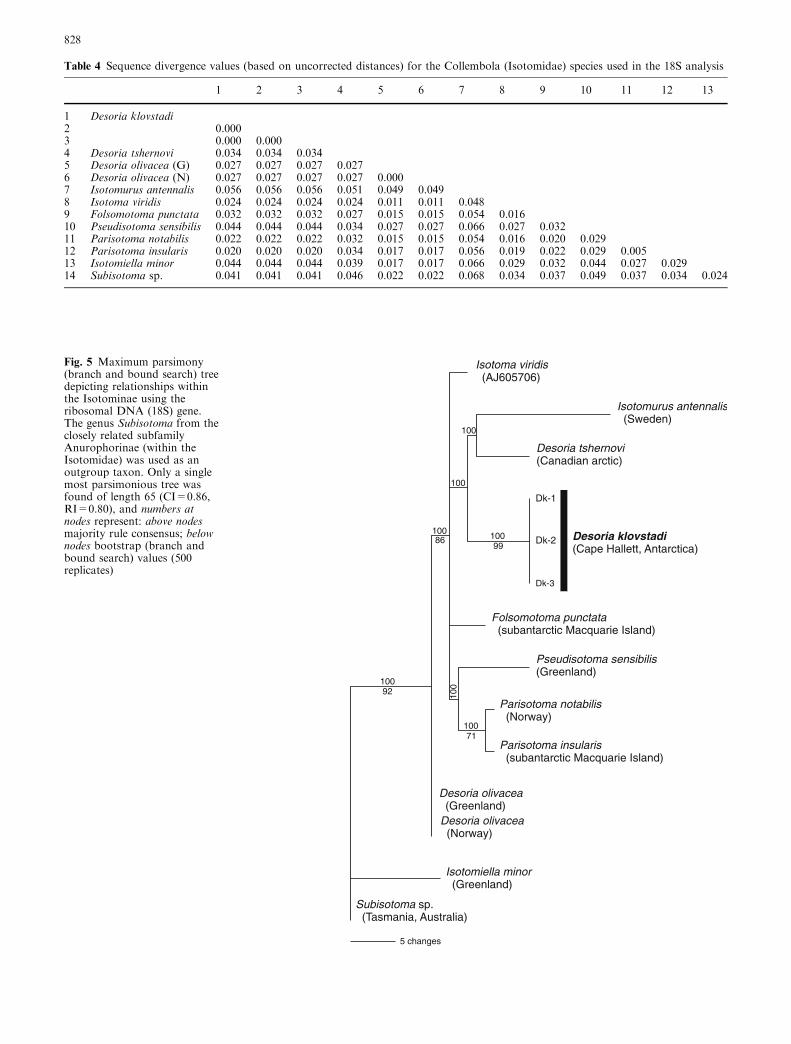
substitutions, and sequence divergence (uncorrecteddistances) ranged from 0.5 to 6.8% between species(Table 4). We were unable to reject the hypothesis ofhomogeneity across base frequencies among sequencesfor all sites (v239=3.31, P=1.00), or for parsimony-informative sites (v239=50.14, P>0.05). The parsimonyanalysis found a single most parsimonious tree (treelength=65, CI=0.86, RI=0.80) (Fig. 5) that had simi-lar topologies to the NJ and ML (best tree 953.79)analyses. Strong phylogenetic signal was indicated(g1=�1.103, gcrit=�0.09, P<0.01), but bootstrapvalues were low (Fig. 5).
Desoria klovstadi was more closely aligned to Desoriatshernovi Martynova, 1974 in the MP analysis, and thesetwo species grouped with I. antennalis (Bagnall 1940)to form an unresolved sister clade to Isotoma and
Folsomotoma, and a sub-clade that included Pseudisotoma,and Parisotoma. The second Desoria sp., olivacea wassister to these genera, but was also distant to D. klovstadifor the COI gene (see Fig. 4). Although bootstrap sup-port was generally low (as found for the mtDNA anal-yses), there were no trees (even under different modelparameters, e.g. uncorrected distance, K2-P, or LogDet)that resulted in a closer relationship between D. klov-stadi and I. viridis Bourlet, 1839. In addition, we furthertested the ML tree (not shown) against a tree whereD. klovstadi was constrained to form a monophyleticgroup with I. viridis. Forcing D. klovstadi with I. viridisdid not result in a significantly worse log likelihood scorefor the ML analyses (Shimodaira–Hasegawa test;P=0.22), but the unconstrained tree was identified witha lower log likelihood score.
100
100
100
100
100
100
100
100
100
100
100
100
100
Desoria klovstadi(Cape Hallett,Antarctica)
Desoria tshernovi(Canadian arctic)
Desoria germanica(Norway)
Desoria germanica(Norway)
Desoria olivacea(Greenland)
Desoria olivacea(Norway)
Isotoma anglicana(Canadian arctic)
Isotomurus sp.(Canadian arctic)
Vertagopus brevicaudus(Canadian arctic)
Pseudisotoma sensibilis(Greenland)
Desoria tigrina(Norway)
Desoria tigrina(Norway)
Parisotoma insularis(subantarctic Macquarie Island)
Subisotoma sp.(Tasmania, Australia)
100
100
100
100
Dk-1
Dk-2
Dk-3
Dk-4
Dk-5
50 changes
Isotomiella minor(Greenland)
Fig. 4 Maximum parsimony(branch and bound search) treedepicting relationships withinthe Isotominae using themitochondrial DNA (COI)gene. The genus Subisotomafrom the closely relatedsubfamily Anurophorinae(within the Isotomidae) wasused as an outgroup taxon. Tenmost parsimonious trees werefound of length 881 (CI=0.50,RI=0.60), and numbers atnodes represent: above nodesmajority rule consensus; belownodes bootstrap (branch andbound search) values (500replicates)
827
Isotomiella minor(Greenland)
Dk-1
Dk-2
Dk-3
5 changes
Desoria klovstadi(Cape Hallett, Antarctica)
Isotoma viridis(AJ605706)
Isotomurus antennalis(Sweden)
Desoria tshernovi(Canadian arctic)
Folsomotoma punctata(subantarctic Macquarie Island)
Pseudisotoma sensibilis(Greenland)
Parisotoma insularis(subantarctic Macquarie Island)
Parisotoma notabilis(Norway)
Desoria olivacea(Greenland)
Desoria olivacea(Norway)
Subisotoma sp.(Tasmania, Australia)
100
100
100
100
100
100
10092
9986
71
Fig. 5 Maximum parsimony(branch and bound search) treedepicting relationships withinthe Isotominae using theribosomal DNA (18S) gene.The genus Subisotoma from theclosely related subfamilyAnurophorinae (within theIsotomidae) was used as anoutgroup taxon. Only a singlemost parsimonious tree wasfound of length 65 (CI=0.86,RI=0.80), and numbers atnodes represent: above nodesmajority rule consensus; belownodes bootstrap (branch andbound search) values (500replicates)
Table 4 Sequence divergence values (based on uncorrected distances) for the Collembola (Isotomidae) species used in the 18S analysis
1 2 3 4 5 6 7 8 9 10 11 12 13
1 Desoria klovstadi2 0.0003 0.000 0.0004 Desoria tshernovi 0.034 0.034 0.0345 Desoria olivacea (G) 0.027 0.027 0.027 0.0276 Desoria olivacea (N) 0.027 0.027 0.027 0.027 0.0007 Isotomurus antennalis 0.056 0.056 0.056 0.051 0.049 0.0498 Isotoma viridis 0.024 0.024 0.024 0.024 0.011 0.011 0.0489 Folsomotoma punctata 0.032 0.032 0.032 0.027 0.015 0.015 0.054 0.01610 Pseudisotoma sensibilis 0.044 0.044 0.044 0.034 0.027 0.027 0.066 0.027 0.03211 Parisotoma notabilis 0.022 0.022 0.022 0.032 0.015 0.015 0.054 0.016 0.020 0.02912 Parisotoma insularis 0.020 0.020 0.020 0.034 0.017 0.017 0.056 0.019 0.022 0.029 0.00513 Isotomiella minor 0.044 0.044 0.044 0.039 0.017 0.017 0.066 0.029 0.032 0.044 0.027 0.02914 Subisotoma sp. 0.041 0.041 0.041 0.046 0.022 0.022 0.068 0.034 0.037 0.049 0.037 0.034 0.024
828

Discussion
Our phylogenetic and morphological analyses confirmthat the closest relatives of D. klovstadi are likely to beIsotoma s. lat. There is no doubt that klovstadi fits thedefinition for Desoria given by Potapov (2001), but ourmorphological and molecular analyses suggest thatklovstadi may not have found its final systematic posi-tion. It is possible that D. klovstadi now represents theonly extant species of an endemic Antarctic genus orsubgenus. This is supported by the position of D. klov-stadi in the mtDNA (COI) and rRNA (18S) phyloge-netic analyses, and also by the high level of sequencedivergence of the COI gene from all other taxa presentedhere (15–28% sequence divergence, Table 3). This rein-forces the ‘‘relic’’ status, rather than of recent (Holoceneor Pleistocene) origin, of this Antarctic Collembolon, ashas also recently been shown for four Antarctic speciesin the sub-family Anurophorinae (Stevens et al. 2006).In addition, using mtDNA (COII) sequences for allIsotomidae from GenBank (data not shown) I. anglicanaand I. viridis were closely allied, but D. klovstadi wasclearly not in the viridis group (no COII sequences wereavailable for Desoria spp.). The position of D. klovstadiin the COI and 18S phylogenetic reconstructions placesit in a position with closest affinities to three Desoria spp.(tshernovi, tigrina, germanica for the mtDNA) and toD. tshernovi (for the 18S gene), although bootstrap supportwas generally low for most branches. A BLASTn search(using 586 bp in the NCBI data base) also indicated thatD. tshernovi (AY665341) had the highest alignmentscore to D. klovstadi (where I. anglicana (AY665326)was ninth). In all the phylogenetic reconstructions (MP,ML, NJ) using various model parameters, D. klovstadidid not group with either I. viridis (18S), or I. anglicana(mtDNA).
Traditional morphological characters examined forD. klovstadi puts the species in an intermediate positionbetween Isotoma s. str. and Desoria s. str., but closer toDesoria. The phylogenetic placement of D. klovstadi inthe mitochondrial and ribosomal analyses also suggestsa closer affinity to Desoria than to any other genus in theIsotominae presented here. It is clear from the evidencehere that Desoria is polyphyletic and warrants furtherexamination to resolve the phylogenetic positions ofother inadequately known species groups in that genus.Accordingly, D. klovstadi is unambiguously placed inDesoria as the genus is defined by Potapov (2001).
The collembolan fauna of Victoria Land and theQueen MaudMountains (Ross Sea Region) of Antarcticacurrently consists of 12 species. Excluding one unde-scribed species (Frati et al. 1997b), ten genera are nowrepresented. All the species are endemic to small areas inthe region except for the widespread Friesea grisea,which also occurs on the Antarctic Peninsula and SouthGeorgia. It has been suggested that F. griesea mayhave been introduced from the Antarctic Peninsula(Rounsevell 1978). Of the remaining nine genera, six are
endemic to the region (Antarctophorus, Biscoia, Gom-phiocephalus, Antarcticinella, Neocyptopygus, Gressitta-cantha) (Wise 1967, 1971; Greenslade 1995; Frati et al.1997a, b; Stevens and Hogg 2002; Stevens and Hogg2003, 2006a). The remaining two genera (Cryptopygusand Tullbergia) are represented by species common tothe Antarctic, subantarctic and extreme southern areasof adjacent continents (Wise 1967, 1971; Greenslade1995; Hogg and Stevens 2002; Stevens and Hogg 2006b),but congeneric representatives have been described fromthe northern hemisphere also. However, Rusek (2002)suggests that Cryptopygus s. str. does not occur there.The tenth genus, Desoria, is mainly holarctic in distri-bution, and D. tshernovi is circum-polar (Spitsbergen,Russian Arctic, Greenland, Alaska, Canada) (Potapov2001). It has few species south of the equator, of whichmost appear to be recently introduced. Thus the geo-graphic range of D. klovstadi (northern Victoria Landand offshore islands) is markedly disjunct from thenative distribution of any of its closest relatives that wehave been able to identify. Our data further reinforcesthe already high level of endemicity resulting from long-term isolation of the Eastern Antarctic collembolanfauna (Wise 1971; Greenslade 1995; Stevens and Hogg2006a, b; Stevens et al. 2006).
Acknowledgements We thank Ken Christiansen, Frans Janssens,and Francesco Frati for discussions that initiated this work, and tothree anonymous reviewers for helpful comments on the manu-script. We are also indebted to Rod Seppelt, Allan Green, Cath-erine Beard, and Brent Sinclair for recent collections from CapeHallett (1999–2004). Arctic collections were supported by anNSERC (Canada) operating grant to P.D.N. Hebert. We are alsoextremely grateful for logistic and/or financial support from DavidPenny, Bryan Gould, (University of Waikato Vice-Chancellor’sfund), Antarctica New Zealand, and Australian Antarctic Division(ASAC grants 2355, 2397).
References
Avise JC (2004) Molecular markers, natural history and evolution,2nd edn. Sinauer, Sunderland, pp 541
Bellinger PF, Christiansen KA, Janssens F (2004) Online checklistof the Collembola of the World (http://www.collembola.org)
Black WC, Klompen JSH, Kierans JE (1997) Phylogenetic rela-tionships among tick subfamilies (Ixodida: Ixodidae: Argasi-dae) based on the 18S nuclear rDNA gene. Mol PhylogenetEvol 7:129–144
Carpenter G (1902) Aptera: Collembola, Insecta, chap 9. Thereport on the collections of natural history made in theAntarctic regions during the voyage of the Southern Cross.British Museum (Natural History), London, pp 221–223
Czarnetzki AB, Tebbe CC (2004) Diversity of bacteria associatedwith Collembola—a cultivation-independent survey based onPCR-amplified 16S rRNA genes. FEMS Microbiol Ecol49:217–227
Deharveng L (1981) Collemboles des Iles Subantarctiques del’Ocean Indien. Comite National Francais des RecherchesAntartiques 48:33–109
Felsenstein J (1985) Confidence limits on phylogenies: an approachusing the bootstrap. Evolution 39:783–791
Fjellberg A (2003) Revision of six northern species of the Isotomaviridis Bourlet, 1839 complex (Collembola, Isotomidae). Nor JEntomol 50:91–98
829

Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R (1994) DNAprimers for the amplification of mitochondrial cytochrome coxidase subunit I from diverse metazoan invertebrates. MolMar Biol Biotechnol 3:294–299
Frati F, Fanciulli PP, Carapelli A, Dallai R (1997a) The Collem-bola of northern Victoria Land Antarctica: distribution andecological remarks. Pedobiologia 41:50–55
Frati F, Fanciulli PP, Carapelli A, DeCarlo L, Dallai R (1997b)Collembola of northern Victoria Land: distribution, populationstructure and preliminary molecular data to study origin andevolution of Antarctic Collembola. In: 3rd meeting on Ant-arctic biology, Santa Margherita Ligure, Italy, Camerino Uni-versity Press, pp 321–330
Frati F, Carapelli A (1999) An assessment of the value of nuclearand mitochondrial genes in elucidating the origin and evolutionof Isotoma klovstadi Carpenter (Insecta, Collembola). AntarctSci 11:160–174
Frati F, Fanciulli PP, Carapelli A, Dell’ampio E, Nardi F, Spin-santi G, Dallai R (2000) DNA sequence analysis to study theevolution of Antarctic Collembola. Ital J Zool Suppl 1:133–139
Frati F, Spinsanti G, Dallai R (2001) Genetic variation of mtCOIIgene sequences in the collembolan Isotoma klovstadi from Vic-toria Land, Antarctica: evidence for population differentiation.Polar Biol 24:934–940
Garrick RC, Sands CJ, Rowell DM, Tait NN, Greenslade P,Sunnucks P (2004) Phylogeography recapitulates topography:very fine-scale local endemism of a saproxylic ‘giant’ springtailat Tallaganda in the Great Dividing Range of south-east Aus-tralia. Mol Ecol 13:3329–3344
Giribet G, Ribera C (2000) A review of arthropod phylogeny: newdata based on Ribosomal DNA sequences and direct characteroptimization. Cladistics 16:204–231
Greenslade P (1995) Collembola from the Scotia Arc and AntarcticPeninsula including descriptions of two new species and noteson biogeography. Pol Pismo Entomol 64:305–319
Hillis DM, Dixon MT (1991) Ribosomal DNA: molecular evolu-tion and phylogenetic inference. Q Rev Biol 66:411–453
Hillis DM, Huelsenbeck JP (1992) Signal, noise, and reliability inmolecular phylogenetic analyses. J Hered 83:189–195
Hogg ID, Hebert PDN (2004) Biological identification of spring-tails (Hexapoda: Collembola) from the Canadian Arctic, usingmitochondrial DNA barcodes. Can J Zool 82:749–754
Hogg ID, Stevens MI (2002) Soil fauna of Antarctic coastallandscapes. In: Beyer L, Bolter M (eds) Geoecology of Ant-arctic ice-free coastal landscapes, vol 154. Ecological studiesanalysis and synthesis, Springer, Berlin Heidelberg New York,pp 265–278
Lockhart PJ, Steel MA, Hendy MD, Penny D (1994) Recoveringevolutionary trees under a more realistic model of sequenceevolution. Mol Biol Evol 11:605–612
Najt J (1981) Contribution a l’etude de la phylogenese et del’ecomorphose chez les Isotoma: le sous-genera Gnathisotoma(Collembole, Isotomidae) et l’espece-souche I. (Desoria) fjell-bergi n. sp. Bulletin Museum Nationale d’Histoire Naturelle,Paris 4 Ser. 3:415–430
Otto JC, Wilson KJ (2001) Assessment of the usefulness ofribosomal 18S and mitochondrial COI sequences in Prostig-mata phylogeny. In: Halliday RB, Walter DE, Proctor HC,Norton RA, Colloff MJ (eds) Acarology: proceedings of the10th international congress, CSIRO publ., Melbourne, pp100–109
Posada D, Crandall KA (1998) Modeltest: testing the model ofDNA substitution. Bioinformatics 14:817–818
Potapov MB (1991) Species of the genus Isotoma subgenus Par-isotoma Bagnall, 1940 and Sericeotoma subgen. nov. (Collem-bola, Isotomidae) of USSR fauna. Acta Zool Cracoviensia34:267–301
Potapov M (2001) Synopses on Palaearctic Collembola: Isotomi-dae. Abhandlungen und Berichte des NaturkundemuseumsGorlitz 73:1–603
Rounsevell DE (1978) Populations of introduced arthropods atAustralian Antarctic stations. Pac Insects 18:199–202
Rusek J (2002) Do we have Cryptopygus—representatives (Col-lembola: Isotomidae) in Europe? Pedobiologia 46:302–310
Saiki RK, Gelfand DH, Stoffel S (1988) Primer-directed enzymaticamplification of DNA with a thermostable DNA polymerase.Science 239:487–491
Simon C, Frati F, Beckenback A, Crespi B, Liu H, Flook P (1994)Evolution, weighting, and phylogenetic utility of the mito-chondrial gene sequences and a complilation of conservedpolymerase chain reaction primers. Ann Entomol Soc Am87:651–701
Simonsen V, Filser J, Krogh PH, Fjellberg A (1999) Three speciesof Isotoma (Collembola, Isotomidae) based on morphology,isozymes and ecology. Zool Scr 28:281–287
Soto-Adames FN (2002) Molecular phylogeny of the Puerto RicanLepidocyrtus and Pseudosinella (Hexapoda: Collembola), avalidation of Yoshii’s ‘‘color pattern species’’. Mol PhylogenetEvol 25:27–42
Stevens MI, Hogg ID (2002) Expanded distributional records ofCollembola and Acari in southern Victoria Land, Antarctica.Pedobiologia 46:485–496
Stevens MI, Hogg ID (2003) Long-term isolation and recent rangeexpansion revealed for the endemic springtail Gomphiocephalushodgsoni from southern Victoria Land, Antarctica. Mol Ecol12:2357–2369
Stevens MI, Hogg ID (2006a) Molecular ecology of Antarcticterrestrial Invertebrates. In: Huiskes A, Convey P, BergstromD, (eds) Trends in Antarctic terrestrial and limnetic ecosystems.Kluwer (in press)
Stevens MI, Hogg ID (2006b) Contrasting levels of mitochondrialDNA variability between mites (Penthalodidae) and springtails(Hypogastruridae) from the Trans-Antarctic Mountains sug-gest long-term effects of glaciation and life history on substi-tution rates, and speciation processes. Soil Biol Biochem 38 (inpress)
Stevens MI, Greenslade P, Hogg ID, Sunnucks P (2006) SouthernHemisphere springtails: could any have survived glaciation ofAntarctica? Mol Biol Evol 23 (in press)
Sunnucks P, Hales DF (1996) Numerous transposed sequences ofmitochondrial cytochrome oxidase I–II in aphids of the genusSitobion (Hemiptera: Aphididae). Mol Biol Evol 13:510–524
Swofford DL (2002) PAUP* Phylogenetic analysis using parsi-mony (*and other methods), ver. 4.0b10. Sinauer Associates,Sunderland
Yoshii R (1990) Report on the cryophilous Collembola , collectedby Dr. S. Kohshima. Contributions from te Biological Labo-raory, Kyotoa University 27:523–533
Wise KAJ (1967) Collembola (Springtails). Antarct Res Ser10:123–148
Wise KAJ (1971) The Collembola of Antarctica. Pac Insects Mo-nogr 25:57–74
Wise KAJ, Shoup J (1967) Distribution of Collembola at CapeHallett. Antarct Res Ser 10:325–330
Wise KAJ, Shoup J (1971) Entomological investigations in Ant-arctica, 1964–1965 season. Pac Insects Monogr 25:27–56
830





























