Embed Size (px)
Citation preview

CLAUDIO GAUCHER AND GERARD J.B. GERMS
SOUTH AFRICAN JOURNAL OF GEOLOGY, 2006, VOLUME 109 PAGE 193-214
193
IntroductionThe Neoproterozoic to early Palaeozoic was an eventfultime in the geological history of the Earth, in that the firstmulticellular animals emerged and that major orogeniesand glaciations occurred (Knoll, 2000). These events andtheir associated sedimentary and igneous rocks can be dated and correlated by radiometric,chemostratigraphic, isotopic and biostratigraphicalstudies. Biostratigraphy entails the study of body andtrace fossils. In Neoproterozoic to early Palaeozoicsuccessions, organic-walled microfossils (includingacritarchs: Evitt, 1963; Downie, 1973) occur, which canbe used for dating of the sediments (Vidal andMoczydlowska-Vidal, 1997; Knoll, 2000; Grey et al.,2003). The aim of this study of organic-walledmicrofossils of the Cango Caves and Gamtoos Groupsand Sardinia Bay Formation of the Saldania Belt in SouthAfrica (Figure 1) is to determine the age relationshipbetween these sedimentary successions in order tounravel the Neoproterozoic to early Palaeozoic geologyof South Africa. The results of this study can alsocontribute to establishing a global biostratigraphy of the
Neoproterozoic to early Palaeozoic. This study formspart of IGCP-Project 478, entitled “Neoproteozoic toearly Palaeozoic events in southwest-Gondwana”.
The Saldania belt is one of the Neoproterozoic toearly Paleozoic orogenic belts in southern Africa thatfringe the Kalahari and Congo cratons (Rozendaal et al.,1999; Kisters et al., 2002). The Saldania beltconsists of a western and southern branch (Figure 1).The western branch was probably formed by theoblique collision of the Kalahari and the Rio de la PlataCratons (Grunow et al., 1996). Two major folding phasescan be distinguished in the western branch of theSaldania Belt, i.e. an older ~580 to ~570 Ma and ayounger ~540 to ~530 Ma phase (Kisters et al., 2002).Strike-slip faulting is common (Rozendaal et al., 1999;Kisters et al., 2002). During the early Paleozoic a shiftfrom collisional to extensional tectonics (circa 530 Ma)took place in the Saldania Belt (Kisters et al., 2002).
The southern branch of the Saldania Belt consistspredominantly of the Cango Caves, Kansa, Gamtoos andKaaimans Groups. Outcrops are restricted to three east-west trending, elongate inliers in the Permo-Triassic
Recent advances in South African Neoproterozoic-Early Palaeozoicbiostratigraphy: correlation of the Cango Caves and Gamtoos Groups
and acritarchs of the Sardinia Bay Formation, Saldania Belt
Claudio Gaucher Departamento de Geología, Facultad de Ciencias. Igua 4225, 11400 Montevideo, Uruguay.
e-mail: [email protected]
Gerard J.B. GermsDepartment of Geology, University of Johannesburg, P.O. Box 524, Auckland Park, 2006
Johannesburg, South Africae-mail: [email protected]
© 2006 March Geological Society of South Africa
ABSTRACT
Organic-walled microfossils (acritarchs) of the Cango Caves and Gamtoos Groups and Sardinia Bay Formation of the Saldania Belt
were studied with the aim to unravel the Neoproterozoic to early Palaeozoic geology of South Africa. Such a micropalaeontological
study will also contribute to establish a world-wide biostratigraphy of the Neoproterozoic to early Paleozoic. Distinct assemblages
of organic-walled microfossils occur in the Cango Caves (CCG) and Gamtoos Groups (GG). Eleven species of organic-walled
microfossils are described from the CCG, six from the GG, and three from the Sardinia Bay Formation. In the CCG, the mixed
clastic-carbonatic Nooitgedagt Member and the overlying, predominantly carbonatic Kombuis Member of the Matjies River
Formation are dominated by Bavlinella and Soldadophycus respectively. The overlying, mainly siliciclastic Groenefontein and Huis
Rivier Formations are leiosphaerid dominated. In the GG, carbonates of the Lime Bank and lower Kaan Formations are Bavlinella-
dominated, the asssemblage of the uppermost Kaan and the lower Van Stadens Formations being dominated by Soldadophycus.
The microflora of the Kleinrivier Formation is dominated by leiosphaerids. The acritarch assemblages and lithostratigraphy indicate
that correlations between units of the GG and CCG become possible and that possible tectonic duplication of lithostratigraphic
units occurs in the GG. The acritarch assemblages of the Lime Bank and most of the Kaan Formations of the GG are similar, and
can be correlated with the Nooitgedagt Member of the CCG. The uppermost Kaan Formation can be correlated with the Kombuis
Member (CCG). The Kleinrivier and the upper Van Stadens Formations are correlative to the Groenefontein Formation and the
overlying Huis Rivier Formation. The generally accepted lithostratigraphic subdivision of the GG does not represent a true
stratigraphic section, but reflects most probably stacking of thrusted sheets. The acritarch assemblages of the CCG and GG indicate
a late Ediacaran age for these units. Correlative successions in southwestern Gondwana include the Nama Group (Namibia), the
Arroyo del Soldado Group (Uruguay) and the Corumbá Group (Brazil). The occurrence of Leiosphaeridia associated with heavily
mineralized, undetermined acritarchs in the Sardinia Bay Formation is also reported.
Cape Fold Belt (Figure 1). The Cango Caves and KansaGroups in the Oudtshoorn inlier (Figures 1 and 3) arepart of a succession which was formally known as theKango Group (Le Roux, 1999). The lithology of
the former Kango Group resembles that of the GamtoosGroup (Figure 5) and therefore it has been suggestedthat they are correlative (Stump, 1976; Tankard et al.,1982).
SOUTH AFRICAN JOURNAL OF GEOLOGY
RECENT ADVANCES IN SOUTH AFRICAN NEOPROTEROZOIC-EARLY PALAEOZOIC BIOSTRATIGRAPHY194
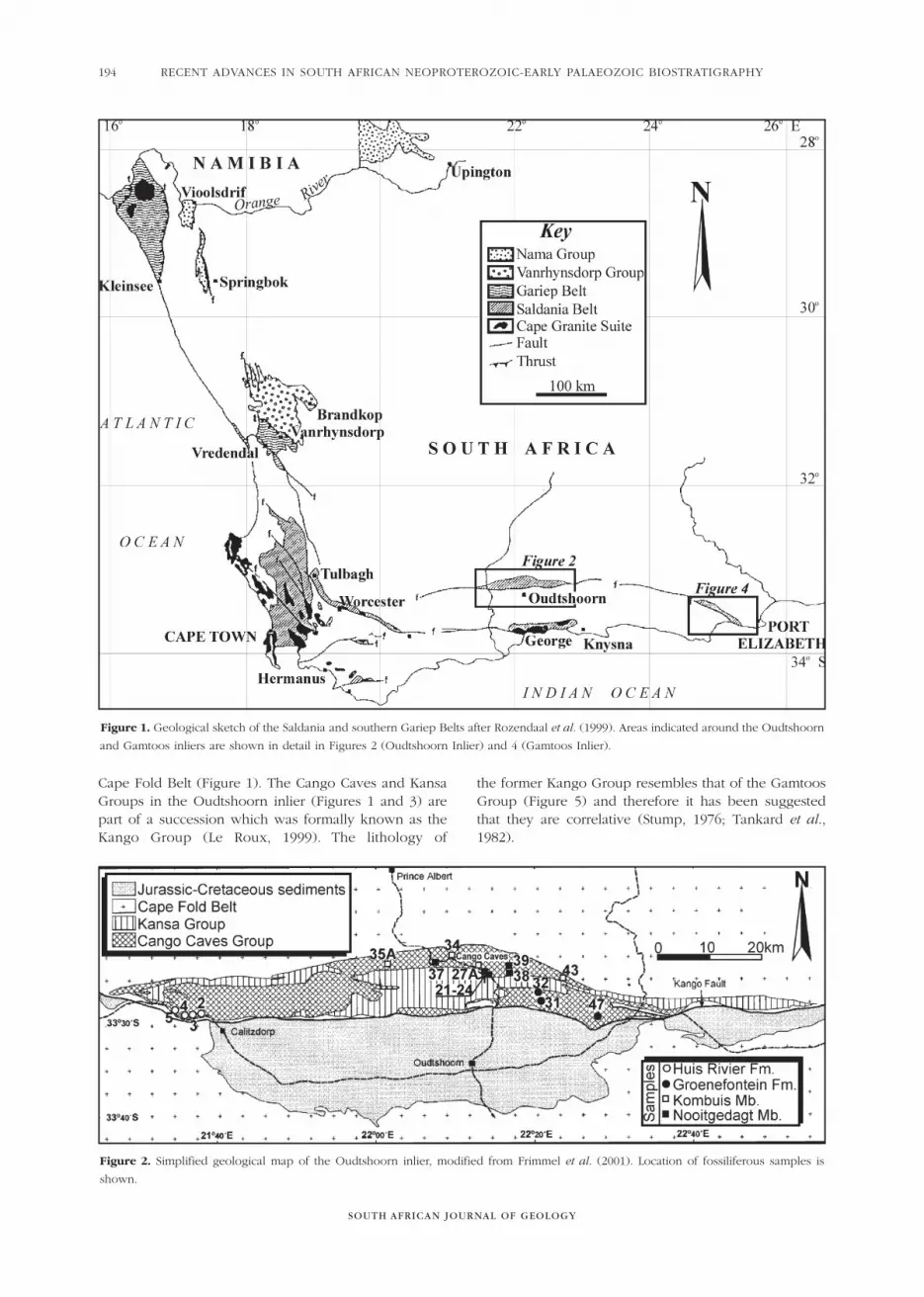
Figure 1. Geological sketch of the Saldania and southern Gariep Belts after Rozendaal et al. (1999). Areas indicated around the Oudtshoorn
and Gamtoos inliers are shown in detail in Figures 2 (Oudtshoorn Inlier) and 4 (Gamtoos Inlier).
Figure 2. Simplified geological map of the Oudtshoorn inlier, modified from Frimmel et al. (2001). Location of fossiliferous samples is
shown.

CLAUDIO GAUCHER AND GERARD J.B. GERMS
SOUTH AFRICAN JOURNAL OF GEOLOGY
195
Various types of granitoids intruded the SaldaniaBelt. The granitoids have been subdivided into an oldersuite of syn-to late-tectonic S-types granites (circa 550 to540 Ma) a younger suite of largely post-tectonic I-typegranites (~540 to ~520 Ma) and a suite of volumetricallysubordinate post-tectonic A-type granites (~520 to ~500Ma) (Scheepers, 1995; Rozendaal et al., 1999; Silva et al.,2000; Kisters et al., 2002). Most of the mentioned ageshave been obtained by the U-Pb SHRIMP method.
Due to poor outcrops, the stratigraphy of theSaldania Belt is difficult to unravel (Hartnady et al, 1974;Theron et al., 1992; Rozendaal et al., 1999). Whereasrecent studies have contributed much to elucidate thestratigraphy of the western Saldania Belt (Belcher andKisters, 2003), the stratigraphy of its southern branch is
still poorly known. Gaucher and Germs (2003) reportedfor the first time organic-walled microfossils from theCango Caves Group, strongly suggesting an upperEdiacaran age for the group. In this paper, furtherstudies of organic-walled microfossils of the CangoCaves and Gamtoos Groups and the Sardinia BayFormation were undertaken to determine thebiostratigraphic correlations of these sedimentarysuccessions.
Lithostratigraphy and available age constraintsCango Caves GroupThe Cango Caves Group (CCG; Le Roux, 1999) isexposed as a basement inlier along the core of a mega-anticline of the Permo-Triassic Cape Fold Belt (Le Roux,
Figure 3. Schematized stratigraphic column of the Cango Caves and Kansa Groups modified after Le Roux and Gresse (1983), Frimmel
et al. (2001) and Gaucher and Germs (2003), showing litho-, chemo- and biostratigraphic data. Sources of � 13C and 87Sr/86Sr-data of Cango
Caves carbonates: Fölling and Frimmel (2002). Pb-Pb dating shown for the Kombuis Member after Fölling et al. (2000).

1997) (Figure 2). The succession, previously namedGoegamma Subgroup (Le Roux, 1999), is part of thePan-African Saldania Belt, which marks the southernmargin of the Kalahari Craton. From base to top, theCCG includes the Matjies River, Groenefontein and HuisRivier Formations (Le Roux, 1997; 1999; Rozendaal et al.,1999) (Figure 3). The succession is made up ofinterbedded siliciclastic and carbonatic deposits at thebase, which pass into siliciclastic turbidites up section(Figure 3). Figure 3 shows the main lithostratigraphicfeatures of this unit, along with chemostratigraphic datareported by Fölling and Frimmel (2002) and microfossiloccurrences first reported in this paper.
Reliable age constraints are scarce for the CCG.Fölling et al. (2000) report a Pb-Pb double-spikeisochron age of 553 ± 30 Ma for carbonates of theKombuis Member (Figure 3). While detrital zircons ofthe Cango Caves Group yield ages around 1100 Ma,those from the overlying Kansa Group also contain a 518± 9 Ma component (Barnett et al., 1997). Theseprovenance ages suggest an Ediacaran age for the CCG,and a late Cambrian or younger age for the overlyingKansa Group. On the basis of C and Sr isotopechemostratigraphy, and a difference of 100°C in thermaloverprint between the Nooitgedagt and Kombuismembers, Frimmel et al. (2001) and Fölling and Frimmel(2002) postulated a pre-Vendian age for the NooitgedagtMember and a Vendian age for the rest of the CCG. Thispre-Vendian age for the Nooitgedagt Member is not inagreement with our micropalaeontological findings (asdiscussed below).
Gamtoos Group The Gamtoos Group (GG) occurs in the Gamtoos inlierof the eastern Saldania Belt in the Permo-Triassic FoldBelt (Figure 1). Its metasedimentary rocks are exposednorthwest of Port Elizabeth in the Eastern Cape Provincein South Africa (Figures 1, 4).
The stratigraphy of the GG was originally establishedby Amm (1935) and later re-interpreted and re-describedby Frankel (1937), Haughton et el. (1937), Bell (1980)and Hill and Nolte (1989). Hill and Nolte (1989)subdivided the group into four formations, bottom totop being the Lime Bank, Klein Rivier, Kaan and VanStadens Formations (Figure 5). The lower boundary ofthe GG is not exposed whereas its upper boundary withthe Sardinia Bay Formation is faulted or unconformable(Le Roux, 2000, Figures 4 and 5). The structuralcomplexity of this group, including many thrust faultsand ubiquitous recumbent folding (Nolte, 1990), andpoor exposures make it difficult to estimate thethickness of the formations reliably. For this reason nostratigraphic columns of the GG indicating thicknesseshave yet been published. Figure 5 shows the generalisedstratigraphy of the GG.
The GG is characterised by predominantlycalcareous units (Lime Bank and Kaan Formations) andpredominantly arkosic and pelitic units (Kleinrivier and Van Stadens Formations, Le Roux, 2000). Phyllite isthe rock type common to all the individual formations(Le Roux, 2000). According to Nolte (1990) thecarbonates were probably deposited in a shallow marineenvironment over which alluvial fans and turbidites
SOUTH AFRICAN JOURNAL OF GEOLOGY
RECENT ADVANCES IN SOUTH AFRICAN NEOPROTEROZOIC-EARLY PALAEOZOIC BIOSTRATIGRAPHY196
Figure 4. Geological map of the Gamtoos inlier, modified after Hill and Nolte (1989). Note location of microfossil-bearing samples.
Due to the complex geology west of the Sardinia Bay, the Sardinia Bay Formation has been included in the Table Mountain Group.

CLAUDIO GAUCHER AND GERARD J.B. GERMS
SOUTH AFRICAN JOURNAL OF GEOLOGY
197
Figure 5. Schematized lithostratigraphic column of the Gamtoos Group, showing stratigraphic distribution of organic-walled microfossils.
No vertical scale implied. Stratigraphic subdivision after Hill and Nolte (1989).

accumulated as basin margin uplift was accelerated. The arkosic rocks are tentatively attributed to a shallowmarine environment (Le Roux, 2000).
Sardinia Bay FormationThe Sardinia Bay Formation is a predominantlyarenaceous succession of folded metasedimentary rockswhich crops out west and northwest of Port Elizabeth(Eastern Cape Province, South Africa) and overlies theGamtoos Group (Figures 4 and 5). In its type areaimmediately east of Sardinia Bay, Shone (1979; 1983)limited the term Sardinia Bay Formation to describe onlythe untectonised outcrops (ca. 180 m thick) (Figure 4).However, Bell (1980) and Toerien and Hill (1991) preferto include all rocks between the Gamtoos Group in thewest and the Peninsula Formation of the Table MountainGroup in the east in the Sardinia Bay Formation. Due tothe complex geology the western Sardinia BayFormation has been included in the Peninsula Formationof the Table Mountain Group, as shown in Figure 4.
The tectonic deformation of the Sardinia BayFormation is complex. Near Port Elizabeth this formationforms a large overturned asymmetric anticlinal structurewith thrust folding (Le Roux, 2000). Some areas of theSardinia Bay Formation are more affected by tectonicdeformation than others. Formal acceptance of the term“Sardinia Bay Formation” by SACS (1980) will have towait until the complex structural deformation of theformation has been unravelled (Le Roux, 2000).
Near Sardinia Bay the basal polymicticmetaconglomerate (ca. 50 m thick) is overlain by thin- tomedium-bedded, predominantly quartzitic sandstonewhich is intercalated with olive-grey to greenish-blackphyllitic shale. Minor small-pebble conglomerate also occur, predominantly comprising vein quartzpebbles (Shone, 1979, 1983; Toerien and Hill, 1991). The sandstone is mostly fine- to medium-grained, lightgrey and locally contains abundant large euhedralmicrocline feldspar crystals probably indicating shorttransport from source (Le Roux, 2000). The shale contentof the Sardinia Bay Formation generally increasesstratigraphically upwards (Le Roux, 2000). The upperboundary of the formation, according to Le Roux (2000)is gradational.
The Sardinia Bay Formation in its type area was mostprobably deposited in a tidal shelf setting (Shone, 1979;1983; Bell, 1980; Nolte, 1985). Le Roux (2000) hassuggested that the formation may represent molasse-type deposits, which accumulated in rift basins formedduring the closing stages of the Saldania Orogen.
Thus far only Paleozoic trace fossils, such asCruziana and burrows resembling Thalassinoides,Ophiomorpha and Diplocraterion and no body fossilshave been found in the Sardinia Bay Formation (Shone,1979; 1983). The above-mentioned trace fossils are notindicative of a specific Palaeozoic age. Some geologistsconsider the Sardinia Bay Formation to be thestratigraphic equivalent of the early to middleOrdovician Graafwater Formation of the Table Mountain
Group in the Western Cape Province (Bell, 1980; Shone,1983; 1988; Le Roux, 2000).
The samples for this micropaleontological studywere collected from the accessible Schoenmakerskoparea, located east of Shone’s (1979; 1983) type area ofthe Sardinia Formation (Figure 4).
Materials and methodsA total of 74 palynological macerations (37 Cango CavesGroup, 29 Gamtoos Group and 8 Sardinia BayFormation) of pelites and carbonates were prepared atthe Micropalaeontology Laboratory of the Facultad deCiencias (Montevideo). Following crushing anddigestion of samples (~150 g) with concentrated HCl,72% HF was applied for 24 hours to the silicate/organicresidues. After neutralization, boiling, concentrated HClwas used to dissolve flourides. Remaining organicresidues were recovered by means of a 5 µm sieve,stored in glass flasks and mounted with glycerin-gelatineon standard glass slides. Throughout the preparation,gravity settling was used instead of centrifugation, toavoid destruction of fragile, large acritarchs. Microfossilswere identified and counted under a Leica DM LPpolarizing microscope, using transmitted, reflected, andcombined reflected-transmitted light (in the latter caseswith oil immersion objectives). This allowed observationof opaque structures, and also to assess the fossil nature of the isolated microstructures. This method,developed by Pflug and Reitz (1992, and referencestherein), makes possible the comparison of thereflectivity and transparency of microfossils. Moderncontaminants are non-reflective, regardless of theirtransparency and colour, because they have notundergone carbonisation. Moreover, the epi-illuminationmethod allows discarding opaque mineral structures thatresemble microfossils (e.g. pyrite framboids). All thestructures described here are clearly reflective andorganic in nature.
PalaeontologyPreservationWhereas twenty-two samples of the CCG (total numberof samples=37) yielded identifiable organic remains, 17 samples of the GG and the Sardinia Formation (totalnumber=37) contain microfossils. In the CCG, a cleardifference in the degree of thermal alteration of fossils inthe different formations was observed. Whereasmicrofossils occuring in samples of the Matjies RivierFormation (Nooitgedagt and Kombuis Members) werefound to be mostly opaque and carbonized, microfossilsof the Groenefontein and Huis Rivier Formations areusually moderate to dark brown in colour. The thermalalteration index (TAI, Staplin in Hunt 1996) according tothe schemes of Hunt (1996) and Teichmüller et al. (1998)is 5 for the Nooitgedagt Member, 4 to 5 for the KombuisMember, and 3 for the Groenefontein and Huis RivierFormations. These TAI values imply temperatures inexcess of 250°C for the Nooitgedagt Member, between170-220°C for the Kombuis Member, and 120 to 170°C
SOUTH AFRICAN JOURNAL OF GEOLOGY
RECENT ADVANCES IN SOUTH AFRICAN NEOPROTEROZOIC-EARLY PALAEOZOIC BIOSTRATIGRAPHY198
CLAUDIO GAUCHER AND GERARD J.B. GERMS
SOUTH AFRICAN JOURNAL OF GEOLOGY
199
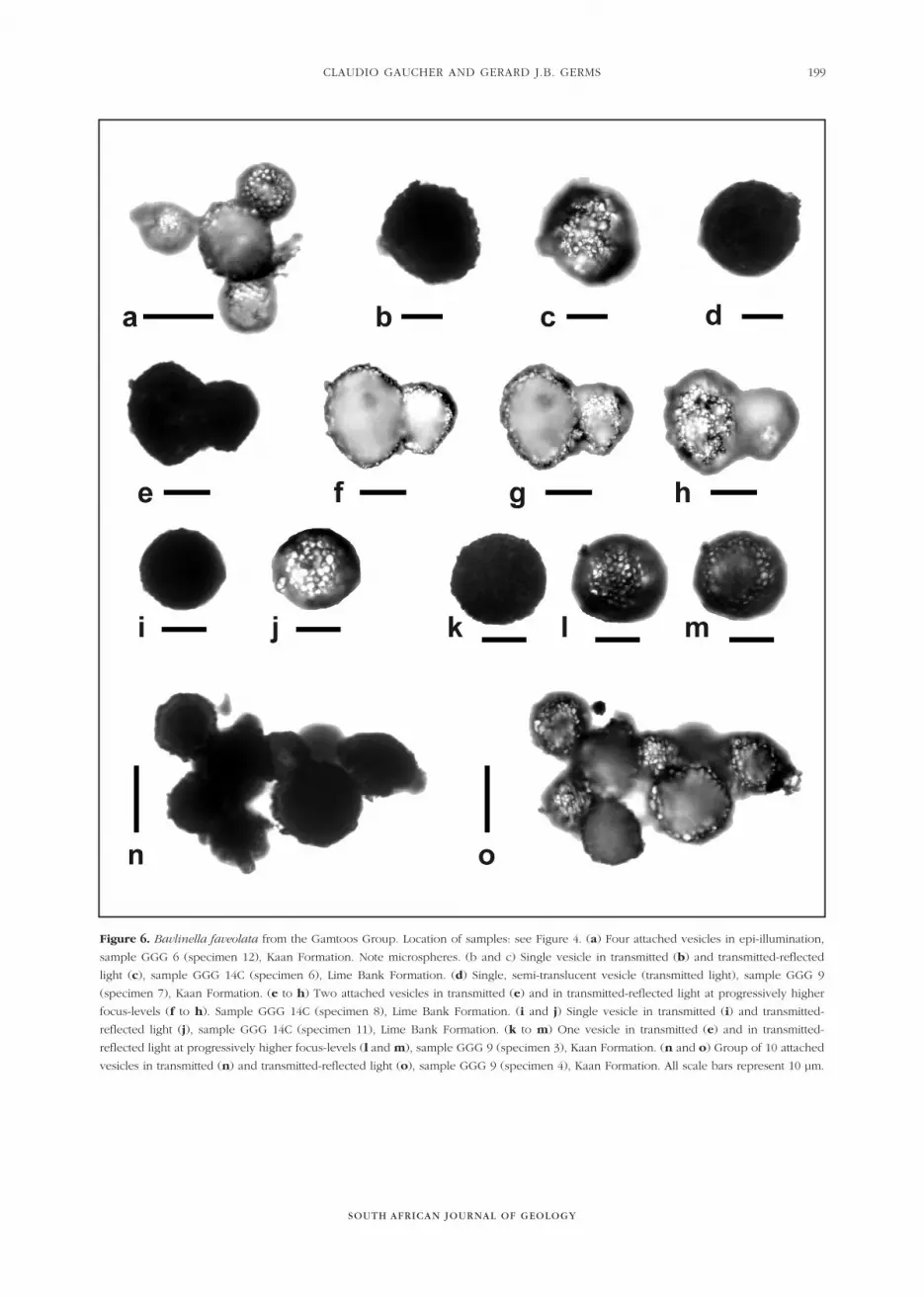
Figure 6. Bavlinella faveolata from the Gamtoos Group. Location of samples: see Figure 4. (a) Four attached vesicles in epi-illumination,
sample GGG 6 (specimen 12), Kaan Formation. Note microspheres. (b and c) Single vesicle in transmitted (b) and transmitted-reflected
light (c), sample GGG 14C (specimen 6), Lime Bank Formation. (d) Single, semi-translucent vesicle (transmitted light), sample GGG 9
(specimen 7), Kaan Formation. (e to h) Two attached vesicles in transmitted (e) and in transmitted-reflected light at progressively higher
focus-levels (f to h). Sample GGG 14C (specimen 8), Lime Bank Formation. (i and j) Single vesicle in transmitted (i) and transmitted-
reflected light (j), sample GGG 14C (specimen 11), Lime Bank Formation. (k to m) One vesicle in transmitted (e) and in transmitted-
reflected light at progressively higher focus-levels (l and m), sample GGG 9 (specimen 3), Kaan Formation. (n and o) Group of 10 attached
vesicles in transmitted (n) and transmitted-reflected light (o), sample GGG 9 (specimen 4), Kaan Formation. All scale bars represent 10 µm.

SOUTH AFRICAN JOURNAL OF GEOLOGY
RECENT ADVANCES IN SOUTH AFRICAN NEOPROTEROZOIC-EARLY PALAEOZOIC BIOSTRATIGRAPHY200
Figure 7. Acritarchs of the Cango Caves Group. Location of samples: see Figure 2. (a) Bavlinella faveolata, Nooitgedagt Member, sample
GGO 24 C (specimen 8). (b) Bavlinella faveolata, translucent, attached vesicles; Nooitgedagt Member, sample GGO 37 (specimen 7).
(c) Bavlinella faveolata, Kombuis Member, sample GGO 27 A (specimen 7). Note translucent microspheres near the edge of the vesicle.
(d) Leiosphaeridia tenuissima, Nooitgedagt Member, sample GGO 24 C (specimen 12). The same specimen could be classified under
Chuaria circularis Walcott, depending on the lower size limit accepted for the taxon (see Butterfield et al. 1994). (e) Chuaria circularis,
Kombuis Member, sample GGO 34 B (specimen 1). Concentric folding of the relatively thick wall is visible in reflected light.
(f) Leiosphaeridia tenuissima, Huis Rivier Formation, sample GGO 2 (specimen 15). Note less thermal alteration than preceding specimens.
(g) Leiosphaeridia tenuissima, Groenefontein Formation, sample GGO 31 B (specimen 1). (h-i) Leiosphaeridia minutissima, intensely
corroded, Kombuis Member, sample 34 B (specimens 3 and 4 respectively). Specimen illustrated in (h) shows possible cell division
(compare with Grey et al. 2003: Figure 4G). (j) Leiosphaeridia minutissima, Huis Rivier Formation, sample GGO 2 (specimen 1). The
specimen shown is less carbonized than L. minutissima from the Kombuis Member and has a distinctly corroded vesicle wall. (k-l)
Asteridium cf. A. tornatum, in reflected (k) and transmitted light (l), Kombuis Member, sample GGO 27 A (specimen 6). Note short and
robust processes (arrowed). Black scale bars represent 10 µm and gray scale bars 50 µm.

CLAUDIO GAUCHER AND GERARD J.B. GERMS
SOUTH AFRICAN JOURNAL OF GEOLOGY
201
for the Groenefontein and lower Huis Rivier Formations(Hunt 1996, Teichmüller et al. 1998). Metamorphismtemperatures calculated by Frimmel et al. (2001) usingcalcite-graphite carbon isotope geothermometry for theNooitgedagt Member are considerably higher (388 ±8°C) than those reported here. On the other hand,temperatures calculated by the same authors for theKombuis Member are only slightly higher (less than 270to 289°C) than alteration temperatures inferred frompalynomorphs. Though there is a difference in TAIbetween the Nooitgedagt and Kombuis Members, a gapof 100°C (Frimmel et al. 2001) seems excessive in viewof TAI estimations reported above. Besides, there issome variation, ranging from slightly less than TAI 4 toTAI 5 in thermal alteration of Kombuis palynomorphsaccording to sample location, which is unrelated tostratigraphic height. Therefore, prudence is suggestedwhile interpreting peak metamorphic temperatures ofthe CCG, especially if the samples were taken fromdifferent sections.
In the GG, thermal alteration is comparable or higherthan that observed in the CCG. For the Lime BankFormation and lower-middle Kaan Formation TAI equals5 (temperature >250°C). Acritarchs of the upper KaanFormation yielded TAI ranging between 4 and 5, similarto the Klein Rivier Formation (temperature between 170 to 250°C). In the Van Stadens Formation, weobserved TAI around 3 to 4, indicating temperaturesaround 170°C for this unit. TAI is dificcult to estimate forthe Sardinia Formation, because a distinct mineralizationof the cell walls occurs (see Figure 11 for reference).However, a few unmineralized specimens suggest aslightly lower TAI around 3. Thrusts may have played animportant role in the observed pattern of thermaloverprint in the GG (see below).
A common feature of all acritarchs occurring in theCCG and GG is an advanced corrosion of vesicle walls,as also reported for the Nama Group (Germs et al.,1986).
RepositoryAll palynological slides, containing specimens describedhere, are kept in the Precambrian collection of theDepartamento de Paleontología, Facultad de Ciencias(Montevideo, Uruguay). Position of specimens in theslides are clearly marked on corresponding duplicates.
Systematic palaeontology
Kingdom Eubacteria Woese and Fox (1977)Phyllum Cyanobacteria Stanier et al. (1978)Classis, Ordo et Fam. indet.
Genus Bavlinella (Schepeleva) Vidal (1976)
Type species. Bavlinella faveolata (Schepeleva) Vidal(1976)
Bavlinella faveolata (Schepeleva) Vidal (1976)
Figures 6 a-o and 7 a-c
1974 Sphaerocongregus variabilis Moorman: plates. 1 to 3
1976 Bavlinella faveolata Vidal: figures. 7A to C1990 Sphaerocongregus variabilis Vidal and Nystuen
figures. 9A-B, D-E, G-L1992 Bavlinella faveolata Schopf: plate 54J1-J3
1996 Bavlinella faveolata Gaucher et al.: figures. 7.1-7.22000 Bavlinella faveolata Gaucher : plate 9, plates 18.1
and 18.22003 Bavlinella faveolata Gaucher et al.: figures 5C to
H, 6F2003 Bavlinella faveolata Gaucher and Germs: figures
6.1 to 6.32005 Bavlinella faveolata Gaucher et al.: figures 6a to i
Type specimen. Vidal (1976) adopted the diagnosisgiven by Moorman (1974) for Sphaerocongregusvariabilis as the valid diagnosis for Bavlinella faveolata.Vidal and Nystuen (1990: p.194) stated that the typespecimen illustrated by Shepeleva (1962, in Vidal, 1976)“is in fact the organic residue after maceration offramboidal pyrite”, and recommend the use of the juniorsynonym instead of Bavlinella faveolata for this species.Nevertheless, German et al. (1989) had alreadydesignated a lectotype for the species from the KotlinFormation of the former USSR. This lectotype has beenalso illustrated by Schopf (1992: pl. 54-J). Therefore, thevalid designation of a lectotype supersedes any previousrestriction of the application of the name of the genusand species Bavlinella faveolata. Sphaerocongregusvariabilis Moorman (1974) is thus to be considered as ajunior synonym (Gaucher et al., 2003).
Material. Hundreds of specimens occur in darklimestones, shales and marls of the Nooitgedagt Member(samples GGO 22A, 23B, 24B, 24C, 37, 38 and 41; Figure2), Kombuis Member (sample GGO 27A; Figure 2), theLime Bank Formation (samples GGG 1A, 1B, 14A’ and14C; Figure 4), Kaan Formation (samples GGG 4, 6 and 9; Figure 4) and a single vesicle in the Klein RivierFormation (sample GGG 39B).
Description. The observed specimens are mainly singlespheroidal vescicles made up of hundreds of tightlypacked, micron-sized (1 to 2 �m) microspheres (Figure 6), thus corresponding to the endosporangiamorphotype of Moorman (1974). Agglomerations of upto 12 attached vesicles also occur (Figures 6 n to o; 7 ato b). Preservation is good despite advancedcarbonisation, which sometimes can only be studied inreflected light. In spite of the fact that a large proportionis opaque (due to carbonisation), the specimens do notrepresent framboidal pyrite, because (i) they are bluish-white under epi-illumination; (ii) they showconsiderable flattening and folding of the vesicle-wall(Figures 6e to h), (iii) many specimens (Figure 6i; n; 7b) are translucent, and (iv) the morphology of the

microspheres is identical to that reported from bona fideBavlinella and distinctly different from pyrite framboids(Figures 6; 7a to c).
Dimensions. Diameter of vesicles ranges in the CCGbetween 4 and 28 �m (mean= 11.9 �m, N=71), andbetween 3 and 30 �m in the GG (mean=12.6 �m, N=82).A single vesicle 44 :m in diameter occurs in theNooitgedagt Member (CCG).
Remarks. Bavlinella faveolata is the dominant taxon in theNooitgedagt Member of the CCG and in the Lime Bankand Kaan Formations of the GG, representing a verycharacteristic microflora mostly associated with organic-rich limestones. The species is long-ranging (Gaucher,2000), but reached its acme in the upper Ediacaran,when it was a dominating component of the biotaworldwide (Moorman, 1974; Mansuy and Vidal, 1983;Knoll and Sweet, 1987), especially associated withorganic-rich sediments (Germs et al. 1986; Palacios,1989; Vidal and Nystuen, 1990; Gaucher, 2000; Gaucheret al., 2003; 2005; Gaucher and Germs, 2003).
Genus Siphonophycus Schopf (1968), emend Knoll et al.(1991)
Type species. Siphonophycus kestron Schopf (1968)
Siphonophycus robustum Schopf (1968), emend Knoll et al. (1991) figure 9 l
1968 Eomycetopsis robusta Schopf: plates 82.2-3; 83.1-41991 Siphonophycus robustum Knoll et al.: figures 10.3;
10.51994 Siphonophycus robustum Butterfield et al.: figures
26A; G1994 Siphonophycus robustum Hofmann and Jackson:
figure 11.52003 Siphonophycus robustum Gaucher et al., figures
6C; D
Type specimen. Specimen of plate 83.1 (a, b) of Schopf(1968).
Material. One large (0.5 mm) filament mat inmacerations of black, laminated limestone of theNooitgedagt Member, CCG (sample GGO 21, Figure 2).
Description. Psilate, unbranched, nonseptate filaments2.5 to 4.0 µm in diameter. The entangled filaments occuras irregular mat-fragments (Figure 9 l). An intensepyritization of the microfossil is observed, with small,subeuhedral pyrite crystals encrusting the filaments(Figure 9 l).
Remarks. The species of the genus Siphonophycus aredistinguished on the basis of trichome-size (Knoll et al.,1991; Butterfield et al., 1994; Hoffman and Jackson,
1994). The material from the Nooitgedagt Member fallswithin the size boundaries of S. robustum. The habit ofthe filament-aggregates described here is similar to S. robustum as well. Butterfield et al. (1994) find that thespecies is the most common shale-facies microbial mat-builder. Our material occurs in fine-grained, laminated,non-stromatolitic limestones deposited in a low-energyenvironment. They are a rare component of themicrobiota, possibly due to a distal depositionalenvironment of the Nooitgedagt Formation.
Incertae sedisGroup Acritarcha Evitt (1963)Genus Asteridium Moczydlowska (1991)
Type species. Asteridium lanatum (Volkova)Moczydlowska (1991)
Asteridium cf. A. tornatum (Volkova, 1968)Moczydlowska (1991) figures 7k; l
1968 Micrhystridium tornatum Volkova: plates. 4.1 to 4.4; 10.8
1991 Asteridium tornatum Moczydlowska: plates. 1A to C
2003 Micrhystridium tornatum Gaucher and Germs:figures. 6.4; 6.5
Material. One group of 10 attached, well-preserved,carbonised vesicles from a limestone sample of theKombuis Member, CCG (sample GGO 27A, Figure 2).
Description. Small spheroidal vesicles covered withconical, short and robust processes. All availablespecimens form a cluster of 10 individuals.Dimensions. Diameter of vesicles ranges between 10and 14 µm (mean=12.9 µm, N=8). Length of processes is1 to 2 µm.
Remarks. This is a subordinate but stratigraphicallyimportant taxon, only occurring in the Kombuis Memberof the CCG. Association of these acritarchs in clusters iscommon for the species (Moczydlowska, 1991).
Genus Chuaria Walcott (1899)
Type species. Chuaria circularis Walcott (1899)
Chuaria circularis Walcott (1899) Figures 7e; 8a
Material. Three carbonised, opaque specimens fromthe lower Groenefontein Member of the CCG (sampleGGO 34B, Figure 2) and Kleinrivier Formation, GG(sample GGG 39C, Figure 4).
Description. Thick-walled, psilate, opaquesphaeromorphs with concentric folds and larger than250 µm in diameter.
SOUTH AFRICAN JOURNAL OF GEOLOGY
RECENT ADVANCES IN SOUTH AFRICAN NEOPROTEROZOIC-EARLY PALAEOZOIC BIOSTRATIGRAPHY202
CLAUDIO GAUCHER AND GERARD J.B. GERMS
SOUTH AFRICAN JOURNAL OF GEOLOGY
203
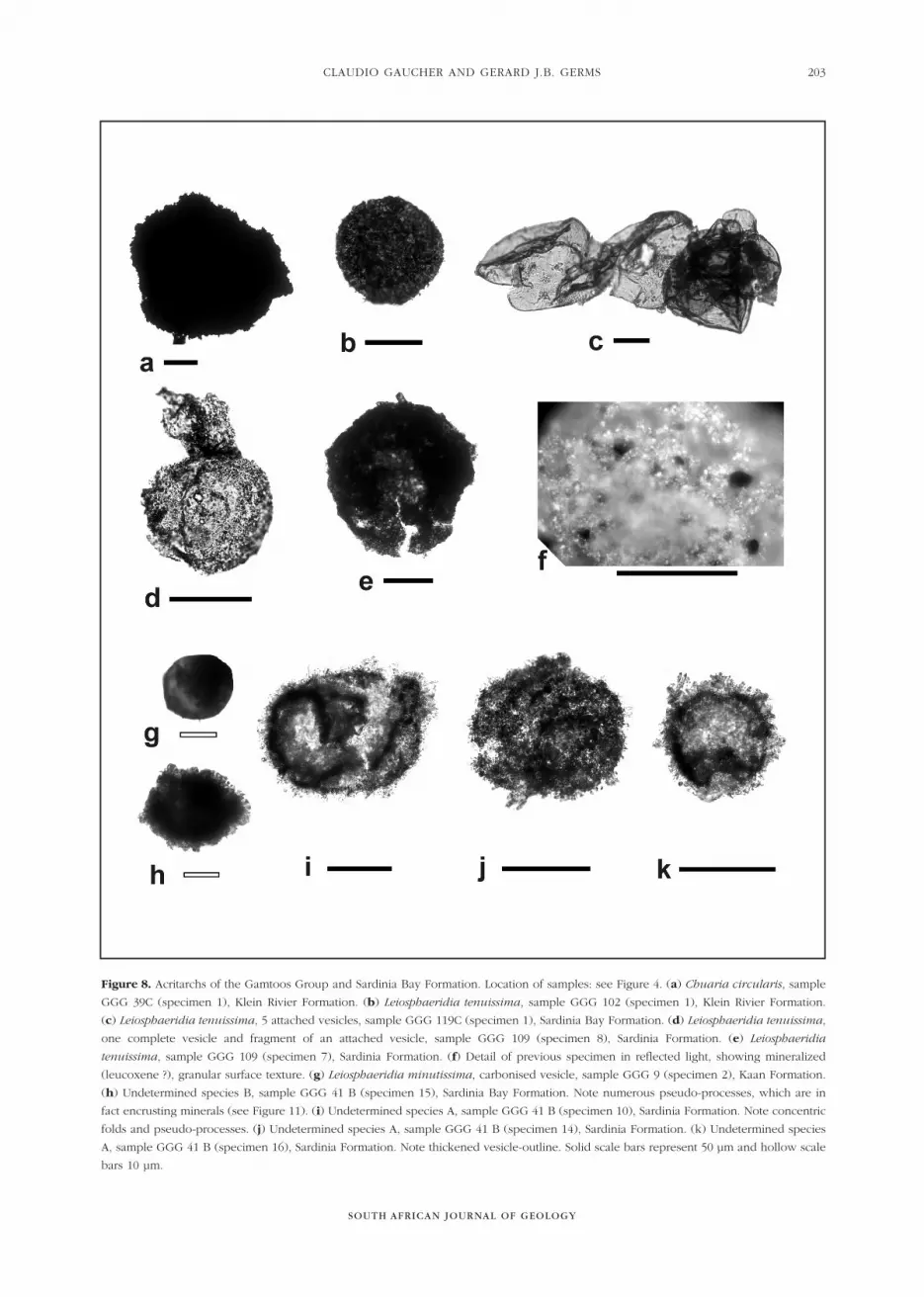
Figure 8. Acritarchs of the Gamtoos Group and Sardinia Bay Formation. Location of samples: see Figure 4. (a) Chuaria circularis, sample
GGG 39C (specimen 1), Klein Rivier Formation. (b) Leiosphaeridia tenuissima, sample GGG 102 (specimen 1), Klein Rivier Formation.
(c) Leiosphaeridia tenuissima, 5 attached vesicles, sample GGG 119C (specimen 1), Sardinia Bay Formation. (d) Leiosphaeridia tenuissima,
one complete vesicle and fragment of an attached vesicle, sample GGG 109 (specimen 8), Sardinia Formation. (e) Leiosphaeridia
tenuissima, sample GGG 109 (specimen 7), Sardinia Formation. (f) Detail of previous specimen in reflected light, showing mineralized
(leucoxene ?), granular surface texture. (g) Leiosphaeridia minutissima, carbonised vesicle, sample GGG 9 (specimen 2), Kaan Formation.
(h) Undetermined species B, sample GGG 41 B (specimen 15), Sardinia Bay Formation. Note numerous pseudo-processes, which are in
fact encrusting minerals (see Figure 11). (i) Undetermined species A, sample GGG 41 B (specimen 10), Sardinia Formation. Note concentric
folds and pseudo-processes. (j) Undetermined species A, sample GGG 41 B (specimen 14), Sardinia Formation. (k) Undetermined species
A, sample GGG 41 B (specimen 16), Sardinia Formation. Note thickened vesicle-outline. Solid scale bars represent 50 µm and hollow scale
bars 10 µm.
SOUTH AFRICAN JOURNAL OF GEOLOGY
RECENT ADVANCES IN SOUTH AFRICAN NEOPROTEROZOIC-EARLY PALAEOZOIC BIOSTRATIGRAPHY204
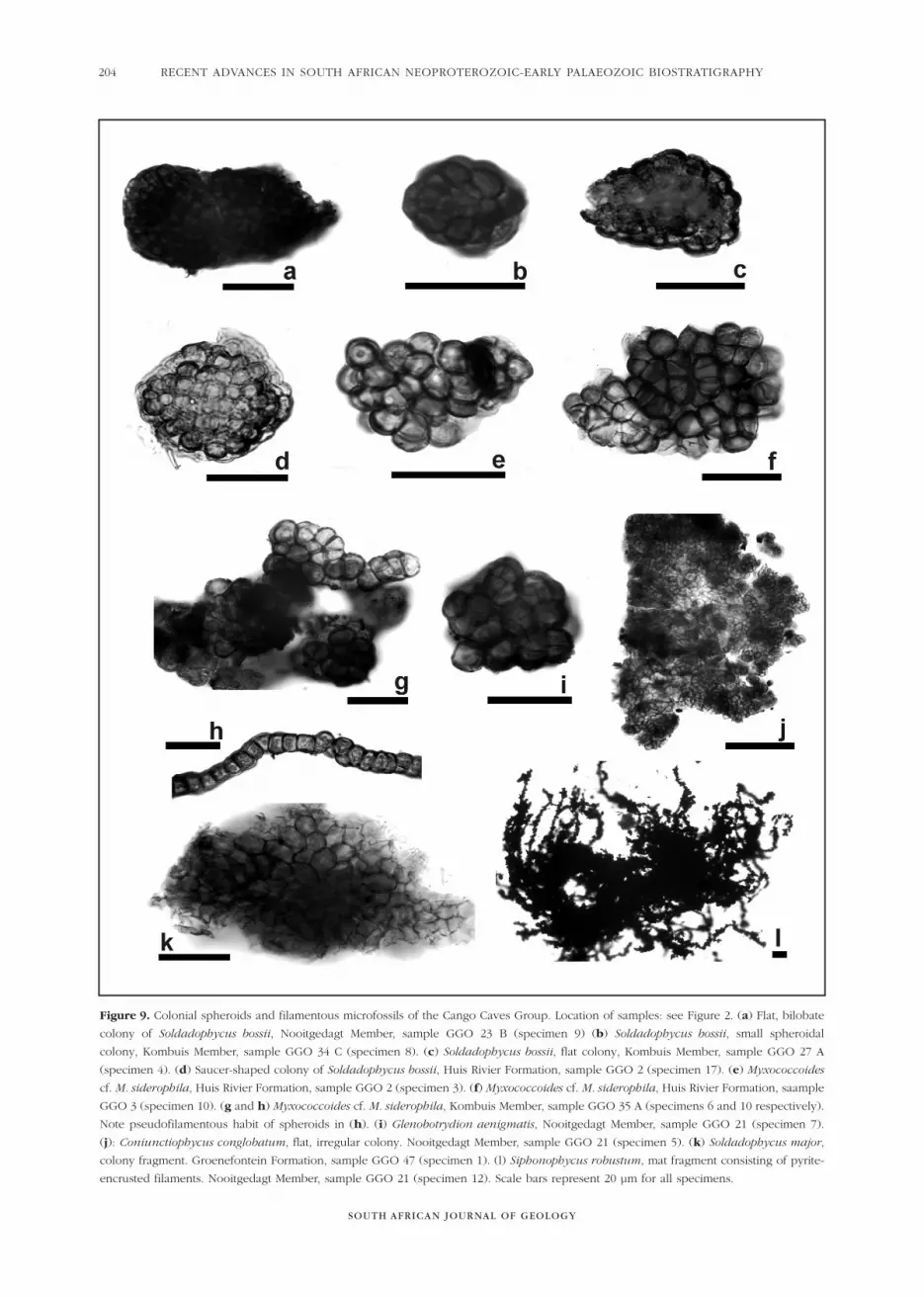
Figure 9. Colonial spheroids and filamentous microfossils of the Cango Caves Group. Location of samples: see Figure 2. (a) Flat, bilobate
colony of Soldadophycus bossii, Nooitgedagt Member, sample GGO 23 B (specimen 9) (b) Soldadophycus bossii, small spheroidal
colony, Kombuis Member, sample GGO 34 C (specimen 8). (c) Soldadophycus bossii, flat colony, Kombuis Member, sample GGO 27 A
(specimen 4). (d) Saucer-shaped colony of Soldadophycus bossii, Huis Rivier Formation, sample GGO 2 (specimen 17). (e) Myxococcoides
cf. M. siderophila, Huis Rivier Formation, sample GGO 2 (specimen 3). (f) Myxococcoides cf. M. siderophila, Huis Rivier Formation, saample
GGO 3 (specimen 10). (g and h) Myxococcoides cf. M. siderophila, Kombuis Member, sample GGO 35 A (specimens 6 and 10 respectively).
Note pseudofilamentous habit of spheroids in (h). (i) Glenobotrydion aenigmatis, Nooitgedagt Member, sample GGO 21 (specimen 7).
(j): Coniunctiophycus conglobatum, flat, irregular colony. Nooitgedagt Member, sample GGO 21 (specimen 5). (k) Soldadophycus major,
colony fragment. Groenefontein Formation, sample GGO 47 (specimen 1). (l) Siphonophycus robustum, mat fragment consisting of pyrite-
encrusted filaments. Nooitgedagt Member, sample GGO 21 (specimen 12). Scale bars represent 20 µm for all specimens.
CLAUDIO GAUCHER AND GERARD J.B. GERMS
SOUTH AFRICAN JOURNAL OF GEOLOGY
205
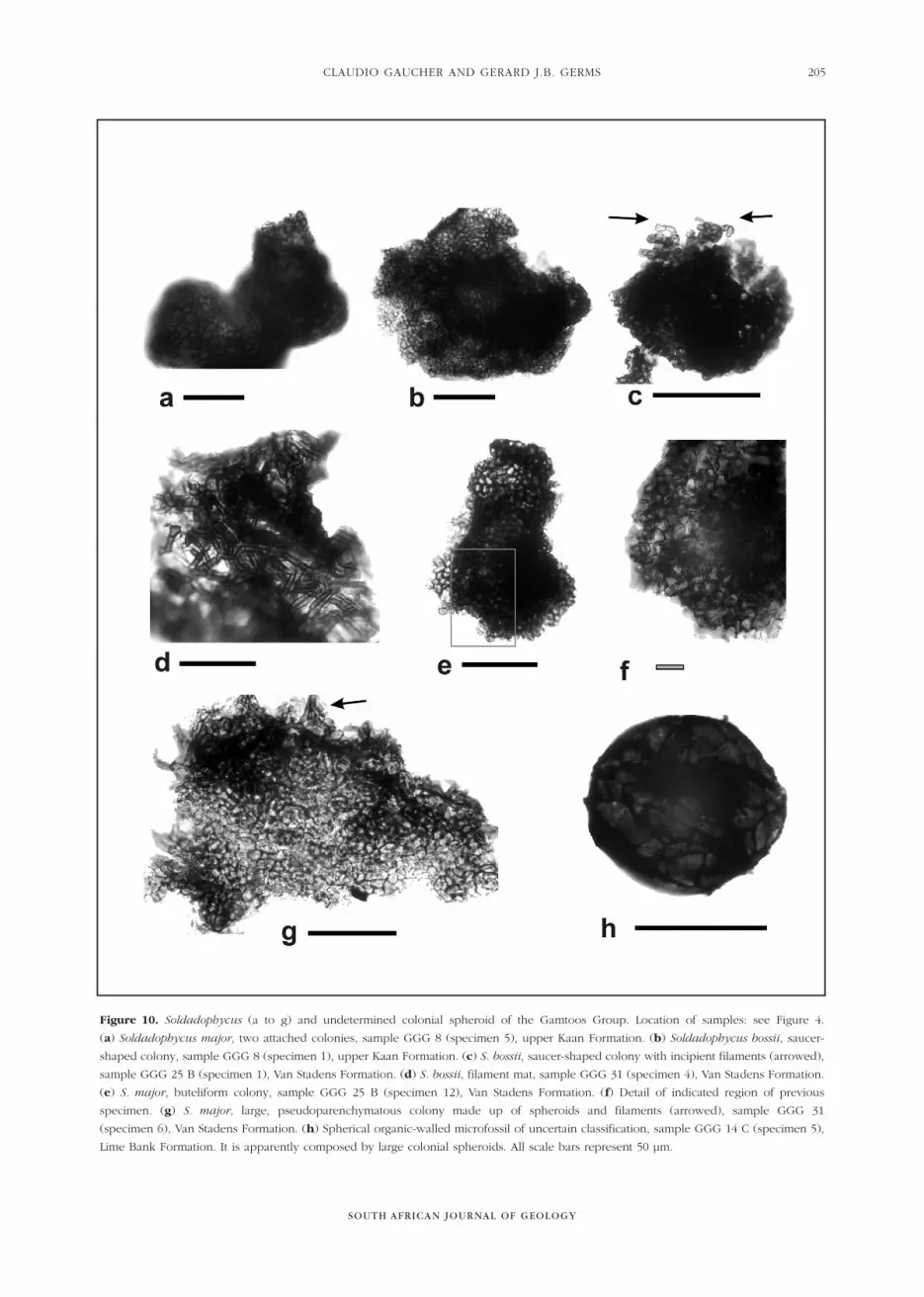
Figure 10. Soldadophycus (a to g) and undetermined colonial spheroid of the Gamtoos Group. Location of samples: see Figure 4.
(a) Soldadophycus major, two attached colonies, sample GGG 8 (specimen 5), upper Kaan Formation. (b) Soldadophycus bossii, saucer-
shaped colony, sample GGG 8 (specimen 1), upper Kaan Formation. (c) S. bossii, saucer-shaped colony with incipient filaments (arrowed),
sample GGG 25 B (specimen 1), Van Stadens Formation. (d) S. bossii, filament mat, sample GGG 31 (specimen 4), Van Stadens Formation.
(e) S. major, buteliform colony, sample GGG 25 B (specimen 12), Van Stadens Formation. (f) Detail of indicated region of previous
specimen. (g) S. major, large, pseudoparenchymatous colony made up of spheroids and filaments (arrowed), sample GGG 31
(specimen 6), Van Stadens Formation. (h) Spherical organic-walled microfossil of uncertain classification, sample GGG 14 C (specimen 5),
Lime Bank Formation. It is apparently composed by large colonial spheroids. All scale bars represent 50 µm.
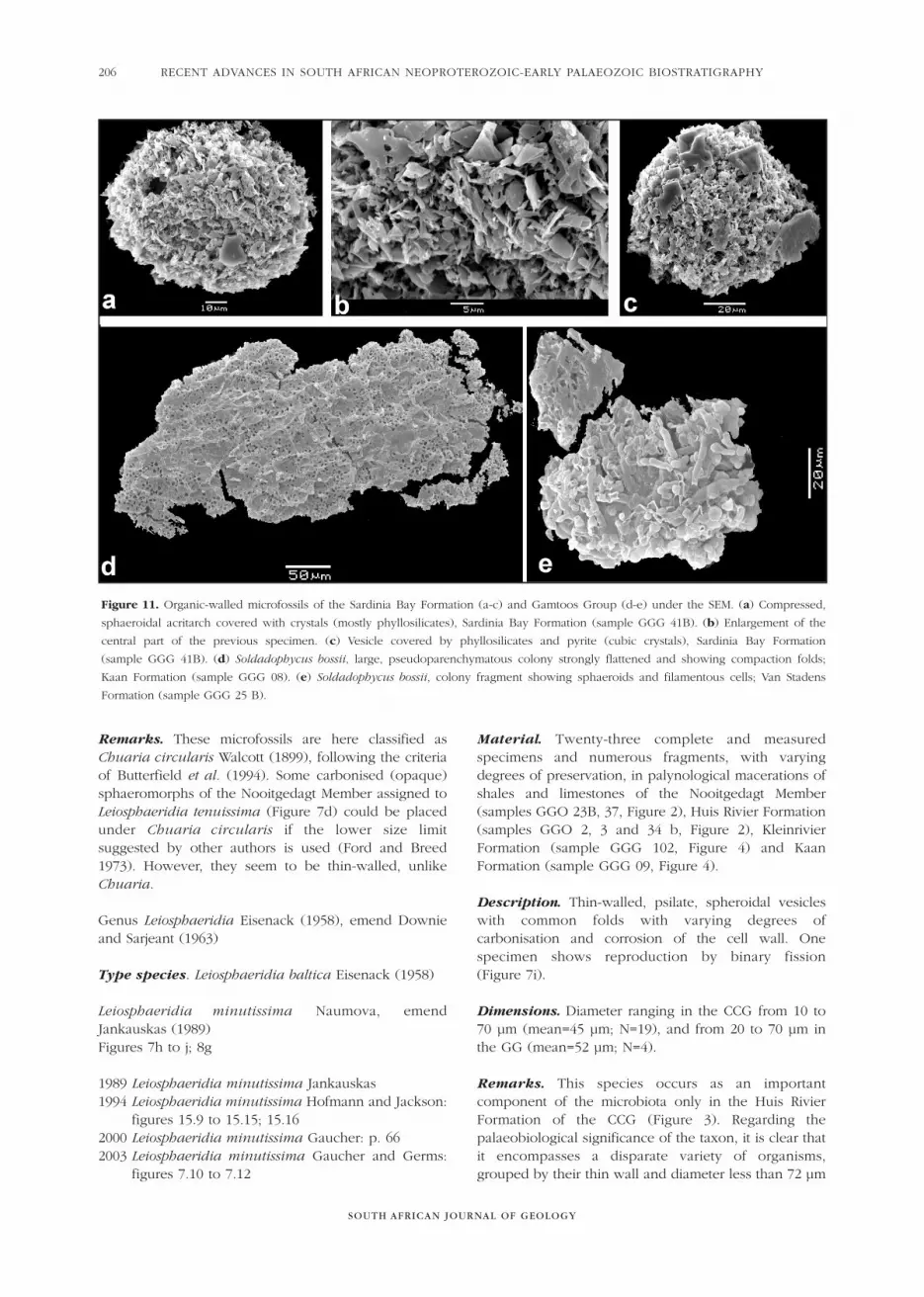
Remarks. These microfossils are here classified asChuaria circularis Walcott (1899), following the criteriaof Butterfield et al. (1994). Some carbonised (opaque)sphaeromorphs of the Nooitgedagt Member assigned toLeiosphaeridia tenuissima (Figure 7d) could be placedunder Chuaria circularis if the lower size limitsuggested by other authors is used (Ford and Breed1973). However, they seem to be thin-walled, unlikeChuaria.
Genus Leiosphaeridia Eisenack (1958), emend Downieand Sarjeant (1963)
Type species. Leiosphaeridia baltica Eisenack (1958)
Leiosphaeridia minutissima Naumova, emendJankauskas (1989)Figures 7h to j; 8g
1989 Leiosphaeridia minutissima Jankauskas1994 Leiosphaeridia minutissima Hofmann and Jackson:
figures 15.9 to 15.15; 15.162000 Leiosphaeridia minutissima Gaucher: p. 662003 Leiosphaeridia minutissima Gaucher and Germs:
figures 7.10 to 7.12
Material. Twenty-three complete and measuredspecimens and numerous fragments, with varyingdegrees of preservation, in palynological macerations ofshales and limestones of the Nooitgedagt Member(samples GGO 23B, 37, Figure 2), Huis Rivier Formation(samples GGO 2, 3 and 34 b, Figure 2), KleinrivierFormation (sample GGG 102, Figure 4) and KaanFormation (sample GGG 09, Figure 4).
Description. Thin-walled, psilate, spheroidal vesicleswith common folds with varying degrees ofcarbonisation and corrosion of the cell wall. Onespecimen shows reproduction by binary fission (Figure 7i).
Dimensions. Diameter ranging in the CCG from 10 to70 µm (mean=45 µm; N=19), and from 20 to 70 µm inthe GG (mean=52 µm; N=4).
Remarks. This species occurs as an importantcomponent of the microbiota only in the Huis RivierFormation of the CCG (Figure 3). Regarding thepalaeobiological significance of the taxon, it is clear thatit encompasses a disparate variety of organisms,grouped by their thin wall and diameter less than 72 µm
SOUTH AFRICAN JOURNAL OF GEOLOGY
RECENT ADVANCES IN SOUTH AFRICAN NEOPROTEROZOIC-EARLY PALAEOZOIC BIOSTRATIGRAPHY206
Figure 11. Organic-walled microfossils of the Sardinia Bay Formation (a-c) and Gamtoos Group (d-e) under the SEM. (a) Compressed,
sphaeroidal acritarch covered with crystals (mostly phyllosilicates), Sardinia Bay Formation (sample GGG 41B). (b) Enlargement of the
central part of the previous specimen. (c) Vesicle covered by phyllosilicates and pyrite (cubic crystals), Sardinia Bay Formation
(sample GGG 41B). (d) Soldadophycus bossii, large, pseudoparenchymatous colony strongly flattened and showing compaction folds;
Kaan Formation (sample GGG 08). (e) Soldadophycus bossii, colony fragment showing sphaeroids and filamentous cells; Van Stadens
Formation (sample GGG 25 B).

CLAUDIO GAUCHER AND GERARD J.B. GERMS
SOUTH AFRICAN JOURNAL OF GEOLOGY
207
(Jankauskas, 1989). In particular, one specimenmeasuring 52 µm is attached to 4 other vesicles largerthan 100 µm which belong into Leiosphaeridiatenuissima (Figure 8c), showing that the size boundaryis in fact arbitrary.
Leiosphaeridia tenuissima Eisenack (1958)Figures 7 d; f to g; 8 b to f
1958 Leiosphaeridia tenuissima Eisenack: plates 1.2; 1.31994 Leiosphaeridia tenuissima Butterfield et al.: figure
16I1994 Leiosphaeridia tenuissima Hofmann and Jackson:
figure 12E1998 Leiosphaeridia tenuissima Gaucher et al.: figure 4.62000 Leiosphaeridia tenuissima Gaucher: plate 11.52003 Leiosphaeridia tenuissima Gaucher and Germs:
figures 6.6; 6.8; 6.92004b Leiosphaeridia tenuissima Gaucher et al.: figure
4D2005 Leiosphaeridia tenuissima Gaucher et al.: figures
8g; h
Material. Twenty-six fairly well-preserved and measuredspecimens and many fragments in macerations of 8 shale samples and one limestone sample of theNooitgedagt Member (sample GGO 24C; Figure 2),Groenefontein Formation (sample GGO 31B; Figure 2),Huis Rivier Formation (samples GGO 2, 4 and 5; Figure2), Kleinrivier Formation (samples GGG 39C and 102)and Sardinia Formation (samples GGG 109 and 119C).
Description. Thin-walled, compressed, psilatespheroidal vesicles with common folds. The degree ofcarbonisation is variable, the specimens from theNooitgedagt Member (CCG, Figure 7d) and theKleinrivier Formation (GG, Figure 8b) being the mostcarbonised. The least thermal alteration is found insamples of the Huis Rivier Formation (Figure 7j) and the Sardinia Formation (Figures 8c-d). Someleiosphaerids of the latter unit (Figures 8d to f), as wellas other acritarchs occurring there (Figures 8h to k)show a distinct diagenetic encrustation of the vesiclewall. The encrusting mineral is acid resistant, highlyreflective and yellowish white in epi-illumination(leucoxene, pyrite and phyllosilicates, see below).
Dimensions. Diameter ranging in the CCG from 72 to200 µm (mean=143 µm; N=7), from 72 to 120 µm(mean=102 µm; N=6) in the GG and between 80 and 170 (mean=117 µm, N=12) in the Sardinia Formation.
Remarks. L. tenuissima is the dominant species of theassemblage preserved in the Groenefontein andKleinrivier Formations (Figures 3; 5); is an importantcomponent of the Huis Rivier assemblage (along with L.minutissima and Soldadophycus bossii, Figure 3) andalso of the microflora of the Sardinia Formation (Figure 5). Many specimens show an irregular granular
Figure 12. Litho- and biostratigraphic correlations between the
successions exposed in the Oudtshoorn and Gamtoos inliers.
(A) Assuming no tectonic duplications and considerable facies
changes; (B) assuming tectonic duplications (preferred here).

surface texture (Figures 8d to f), which is either theresult of mineral encrustation or vesicle corrosion, andnot a primary feature.
Undetermined species AFigures 8i to k; 11a to c
Material. Seventeen measured specimens from a shalesample of the Sardinia Formation (sample GGG 41B,Figure 4).
Description. Vesicles circular to oval in outline,covered by platy phyllosilicate crystals (encrustation?)composed by K, Si and Al according to EDS analyses,and also pyrite and leucoxene (Figure 11). These crystalsresemble short, filiform processes when observed underthe optical microscope. Concentric folds are common(Figure 8i-k), and a thickened vesicle outline is oftenobserved (Figure 8k).
Dimensions. Maximum diameter of vesicles rangesbetween 66 and 150 µm (mean=102 µm, sd=26 µm,N=17).
Remarks. The pseudo-processes give these acritarchsan appearance similar to Comasphaeridium Staplin et al.(1965). The nature of the “processes” is only revealedunder the scanning electron microscope, though theoccurrence of encrusting minerals is also shown byreflective light microscopy. Given the compressed,subcircular shape, occurrence of concentric folds andthe relatively large size, we place these microfossils inthe Acritarcha. Attached crystals hinder the observationof the real surface sculpture, thus preventing a moreprecise assignment of these specimens. Undetermined species BFigure 8h
Material. Three measured specimens from a shale sampleof the Sardinia Formation (sample GGG 41B, Figure 4).
Description. Vesicles circular to oval in outline,apparently covered by numerous, densely-packed,filiform processes. Vesicle outline is obscured by theprocess-like structures in their proximal part.
Dimensions. Maximum diameter of vesicles rangesbetween 26 and 44 µm (mean=37 µm, N=3).
Remarks. This species is considerably smaller than theundetermined species A. No specimens could beisolated for SEM imaging, but the vesicles are most likelycovered by phyllosilicates, because they are distinctlyreflective under epi-illumination (as the previousspecies). Therefore, we envisage that the “processes” arein fact an artifact caused by mineral encrustation, asdemonstrated for the species A. This prevents a moreprecise taxonomic assignment.
Incertae sedisGenus Coniunctiophycus Zhang (1981)Type species: Coniunctiophycus gaoyuzhuangenseZhang (1981)Coniunctiophycus conglobatum Zhang (1981)Figure 9j
1981 Coniunctiophycus conglobatum Zhang: plate 4.2000 Coniunctiophycus conglobatum Gaucher: plates
12.8; 9; 13.52003 Coniunctiophycus conglobatum Gaucher and
Germs: figure 7.102005 Coniunctiophycus conglobatum Gaucher et al.:
figure 9c
Type specimen. Holotype: specimen figured by Zhang(1981: pl. 4, Figure 11), with catalogue number BGP7804.
Material. Six well-preserved colonies occurring inlimestones and shales of the Nooitgedagt to lowermostKombuis Members (samples GGO 21 and 35A, Figure 2).
Description. Colonial, originally spherical-ellipsoidal tosubpolyhedral cells, showing distortion by mutualcompression. Colonies flat and irregular (Figure 9j) tospherical, composed of hundreds of cells. Two smallcolonies are attached to Glenobotrydion, suggesting thatthey are epiphytic, as previously reported for the taxon(Gaucher, 2000).
Dimensions. Cell-diameter varies between 1 and 3 µm(mean=1.7 µm; N=15). Spherical colonies reach 40 µm|in diameter and flat, irregular colonies reach 100 µm inmaximum dimension.
Remarks. These are the smallest colonial spheroids ofthe CCG. It is a common component of latestNeoproterozoic microfloras (e.g. Gaucher, 2000;Gaucher et al., 2005).
Genus Glenobotrydion Schopf (1968)
Type species. Glenobotrydion aenigmatis Schopf(1968)
Glenobotrydion aenigmatis Schopf (1968)Figure 9.i
1968 Glenobotrydion aenigmatis Schopf: plate 81, figure5; plate 83; figure 9; plate 84, figures 4 to 5; text-Figure 6
2000 Glenobotrydion aenigmatis Gaucher: plates. 12.5to 12.7
Type specimen. Holotype: colony with cataloge number58505 (Paleobotoony Collection of Harvard University)figured by Schopf (1968: pl. 84, Figure 5).
SOUTH AFRICAN JOURNAL OF GEOLOGY
RECENT ADVANCES IN SOUTH AFRICAN NEOPROTEROZOIC-EARLY PALAEOZOIC BIOSTRATIGRAPHY208

CLAUDIO GAUCHER AND GERARD J.B. GERMS
SOUTH AFRICAN JOURNAL OF GEOLOGY
209
Material. Fourteen well-preserved colonies occurring inmacerations of a single sample of limestone (GGO 21,Figure 2) of the Nooitgedagt Member.
Description. Spheroidal to ellipsoidal, colonialmicrofossils, showing optically dense, spheroidal toellipsoidal inner bodies. Cell-walls robust and psilate.Colonies are irregular or spheroidal (Figure 9i),sometimes enclosed in a granular sheath, and composedof dozens of cells. Dyads are common, reflectingdifferent stages of vegetative division of cells.
Dimensions. Diameter of cells ranging between 5.0and12 µm (mean=8.3 µm, N=32), and diameter of innerbodies varies between 2 and 4 µm. The spheroidalcolony shown in Figure 9i measures 42 µm.
Comments. The material of the CCG belongs toGlenobotrydion, because of occurrence of typical innerbodies and size-frequency distribution (Schopf, 1968).One difference are the spheroidal colonies occurring inthe CCG, not known from the type-area. Similar colonieswere reported for Glenobotrydion aenigmatis from theArroyo del Soldado Group by Gaucher (2000).
Genus Myxococcoides Schopf (1968)
Type species. Myxococcoides minor Schopf (1968)
Myxococcoides cf. M. siderophilaFigures 9e; g; h
2000 Myxococcoides siderophila Gaucher: plates 13.1 to 5
2003 Myxococcoides sp. A Gaucher and Germs: figures7.5; 7.7; 7.8
2003 Myxococcoides sp. B Gaucher and Germs: figure7.6
2005 Myxococcoides siderophila Gaucher et al.: figure 9g
Materia. A dozen well-preserved colonies withhundreds of cells, recovered from shales (3 samples)and limestones (1 sample) of the Kombuis Member(samples GGO 35A, Figure 2), lower GroenefonteinFormation (sample GGO 34C) and the Huis RivierFormation (samples GGO 2, 3; Figure 2).
Description. Spheroidal, robust-walled microfossils.Walls psilate, up to 1.5 µm thick. Cells commonlyaggregated into loose, irregular colonies of up to severaldozens of cells (Figures 9e; g). Long chains of spheroids,mostly uni- or biserial (Figure 9h) occur
Dimensions. Diameter of spheroids varies between 4.4and 10.8 µm (mean=7.4 µm, N=31).
Remarks. These fossils fit the generic diagnosis ofMyxococcoides, and are here assigned to M. siderophilaGaucher (2000) because: (1) they show robust walls, (2)
their size is within the size-frequency distribution of thetype material (Gaucher, 2000), and (3) they form loose,irregular colonies. It differs from Soldadophycus in itssize, more robust walls, absence of filaments and colonyhabits. It is distinguished from Glenobotrydionaenigmatis by the ubiquitous occurrence in the lattertaxon of a dense internal body and dyads, and by itslarger size.
Genus Soldadophycus Gaucher et al. (1996)
Type species. Soldadophycus bossii Gaucher et al. (1996)
Soldadophycus bossii Gaucher et al. (1996)Figures 9a to d;10b to d; 11d; e
1996 Soldadophycus bossii Gaucher et al.: figures 6.1 to6.5; 6.7
1998 2000 Soldadophycus bossii Gaucher: plates 14; 15;17.4
2003 Soldadophycus bossii Gaucher et al.: figures 5B; 6A;B
2003 Soldadophycus bossii Gaucher and Germs: figures7.1 to 7.4
2004a Soldadophycus bossii Gaucher et al.: figures 4B; C;E
2005 Soldadophycus bossii Gaucher et al.: figures 7a; b;9a; b, d to f
Type specimen. Holotype: specimen FCDP 3188,figured by Gaucher et al. (1996: Figure 6.1).
Material. Dozens of colonies and colony-fragments inpalynological macerations of shales (5 samples) andlimestones (6 samples) of the Nooigedagt Member(samples GGO 22A, 23B, 24C; Figure 2), KombuisMember (samples GGO 27A, 35A, Figure 2), lowerGroenefontein Formation (sample GGO 34C), HuisRivier Formation (samples GGO 2, 3, 4, Figure 2), VanStadens Formation (sample GGG 25B, Figure 4) andupper Kaan Formation (sample GGG 08, Figure 4).Microfossils are very dark brown to black in the GG, andmedium to very dark brown in the CCG. All specimensare clearly reflective under epi-illumination, and most ofthem are strongly compressed (e.g. Figure 11d). Thisproves the fossil nature of the specimens described here.
Description. Soldadophycus bossii is characterized bythe co-occurrence of psilate, spheroidal cells andseptate, branched filaments (Gaucher et al., 1996).Different type of colonies were recognized by Gaucher(2000), which are made up only of spheroidal cells, onlyof filaments or both. The material from the CCG includesmostly spheroidal and saucer-shaped colonies onlycomposed of spheroidal cells (Figures 9a to d).Specimens from the GG include spheroidal, saucer-shaped, buteliform and larger, pseudoparenchymatouscolonies, which are composed by spheroids, filamentsor both type of cells (Figures 10b to d). Typical

transitions from spheroids to filaments and vice versa isobserved in many specimens (e.g. Figure 10c).
Dimensions. Diameter of spheroidal cells ranges in theCCG between 2.2 and 6.0 µm (mean=4.2 µm, N=64). Inthe GG, spheroidal cells are 3.1 to 6.9 µm in diameter(mean=4.4 µm, N=44), width of filaments varyingbetween 2.5 and 3.2 µm. Diameter of saucer-shapedcolonies ranging from 36 to 90 µm in the CCG (mean=55µm, N=7), and from 85 to 90 µm in the GG. Spheroidalcolonies range in diameter between 21 and 54 µm in theCCG, a single spheroidal vesicle from the GG measuring56 µm. These values are within the cell and colony sizestypical for the species (Gaucher, 2000).
Remarks. This species dominates the assemblagepreserved in the Kombuis Member, and that of theupper Kaan and Van Stadens Formation (along withSoldadophycus major). It is an important butsubordinate component of the microbiota of the lowerHuis Rivier Formation. Differences in colony-types and colony-size between the CCG and GG could be due to a more proximal setting of the latter unit. The palaeobiological affinities of Soldadophycus remainuncertain. Similar morphologies have been reported forWengania Zhang (1989) a red alga from the DoushantuoFormation (Yuan et al., 2002; Xiao et al., 2004).However, Wengania has larger and spherical thalli,unknown for S. bossii.
Soldadophycus major Gaucher (2000)Figures 9k; 10a; e to g2000 Soldadophycus major Gaucher: plates 16.1 to
6; 17.62003 Soldadophycus major Gaucher and Germs: figure
7.112004a Soldadophycus major Gaucher et al.: figures 4F; G2005 Soldadophycus major Gaucher et al.: figures 8a
to e
Type specimen. Colony fragment FCDP 3216 figured byGaucher (2000: pl. 16. 2).Material. Five well-preserved colonies and colony-fragments in palynological macerations of siltstones ofthe Quebrada de Viera Member (site LA-31, Figure 1).
Material. Dozens of colonies and colony-fragments inpalynological macerations of shales (3 samples) andlimestones (2 samples) of the Nooitgedagt Member(sample GGO 24C, Figure 2), Groenefontein Formation (sample GGO 47, Figure 2), upper KaanFormation (sample GGG 08, Figure 4) and Van StadensFormation (samples GGG 25B, 31; Figure 4).
Description. Colonial spheroids characterized byhyaline, psilate, flexible walls. Transitions fromspheroidal into filamentous cells and vice-versa areobserved. Filaments are septate and branched, occurringin about 30% of the studied colonies. Colony-types
observed include spheroidal, ellipsoidal, discoidal,buteliform (Figures 10 a; e; f) and fragmentarypseudoparenchymatous (Figures 9k; 10g). They aremade up only of spheroids, only of filaments (filamentmats) or both.
Dimensions. Diameter of spheroids varies in the CCGbetween 3.3 and 8 µm (mean=5.6 µm, N=18) andbetween 3.7 and 11.1 µm (mean=7.4 µm, N=29) in theGG. There, ellipsoidal colonies are 36 to 70 µm inmaximum diameter, discoidal colonies 240 to 300 µm in diameter, and fragments of pseudoparenchymatouscolonies reach 0.5 mm in maximum dimension.Filamentous cells are 2 to 3 µm wide in the CCG and 2 to 5 µm in the GG.
Remarks. The material described under this speciesclearly belongs to Soldadophycus major Gaucher (2000),because of the occurrence of similar colony habits,spheroid-filament transitions and the same size of cellsas the type material from the Arroyo del Soldado Group.S. major is a dominant component of the microflora inthe upper Kaan and in the Van Stadens Formation(together with S. bossii), and an accessory species in therest of the units where it occurs. As for S.bossii, morediverse colony-types and larger colony-size in the GGcould be due to a more proximal setting of this unitcompared to the CCG.
DiscussionBiostratigraphyFour different, low-diversity assemblages can bedistinguished in the CCG, GG and Sardinia BayFormation. These are (Figures 3; 5):A Bavlinella-dominated assemblage occurring in theNooitgedagt Member (CCG), Lime Bank Formation andmost of the Kaan Formation (GG). These intervals arelithologically characterized by organic-rich limestones,marls and shales, and by great abundance ofmicrofossils.
A Soldadophycus-dominated assemblage mainlyoccurring in the Kombuis Member (CCG), upper KaanFormation and lower Van Stadens Formation (GG). Thisinterval is characterized by the transition fromcarbonates into shales.
A Leiosphaeridia-dominated assemblage occurring inthe Groenefontein and Huis Rivier Formations of theCCG, and the Kleinrivier Formation of the GG. Theseunits are characterized by shales and sandtoneinterbeds, mainly of turbiditic origin.
An acritarch assemblage comprising Leiosphaeridiaand undetermined acritarchs with mineral encrustationoccurring in interbedded shales of the Sardinia BayFormation.
The depauperate Bavlinella, Soldadophycus andLeiosphaeridia palynomorph assemblages preserved inthe CCG and GG are typical for the late Ediacaran(Vendian). Similar low-diversity, high-abundancemicrofloras were also reported for the late Ediacaran of
SOUTH AFRICAN JOURNAL OF GEOLOGY
RECENT ADVANCES IN SOUTH AFRICAN NEOPROTEROZOIC-EARLY PALAEOZOIC BIOSTRATIGRAPHY210

CLAUDIO GAUCHER AND GERARD J.B. GERMS
SOUTH AFRICAN JOURNAL OF GEOLOGY
211
Namibia (Germs et al., 1986; Gaucher et al., 2005), theEast European Platform (Volkova, 1985), France (Mansuyand Vidal, 1983), Norway (Vidal and Nystuen, 1990),Spain (Palacios, 1989; Vidal et al. 1994), Spitsbergen(Knoll and Sweet; 1987), Sweden (Vidal, 1976), Uruguay(Gaucher, 2000; Gaucher et al., 2004a); ShackletonRange of Antarctica (Weber, 1991), Australia (Grey et al.2003), Brazil (Gaucher et al. 2003; Teixeira and Gaucher,2004), Canada (Moorman 1974) and China (Zang, 1992).The microflora matches the Kotlin-Rovno assemblage ofVidal and Moczydlowska-Vidal (1997), which ischaracterized by low diversity, abundance of Bavlinellafaveolata, rarity or absence of acanthomorphs andabsence of large (> 500 µm) leiosphaerids (Vidal andMoczydlowska-Vidal, 1997; Knoll, 2000; Grey et al.,2003).
The assemblages that most resemble the microflorasof the CCG and GG are those of SouthwesternGondwana: Nama Group and Holgat Formation ofNamibia (Germs et al., 1986; Gaucher et al., 2005),Arroyo del Soldado Group of Uruguay (Gaucher, 2000;Gaucher et al., 2004a), Corumbá Group of Brazil(Gaucher et al., 2003) and the Eleuterio-Pico de Itapeva-Pouso Alegre basins (Brazil, Teixeira and Gaucher,2004). All these successions are late Ediacaran, post-Gaskiers (582 ± 0.4 Ma: Bowring, in MacGabhann, 2005;Bowring et al., 2003) in age, as demonstrated by U-Pbzircon ages of intercalated ash beds (Grotzinger et al.,1995; Boggiani et al., 2005), C and Sr chemostratigraphy(Saylor et al., 1998; Fölling and Frimmel, 2002; Boggianiet al., 2003; Gaucher et al., 2003; Gaucher et al., 2004b)and skeletal fossils (mainly Cloudina, Namacalathusand Titanotheca: Germs, 1972; Gaucher andSprechmann, 1999; Grotzinger et al., 2000; Gaucher et al., 2003; Teixeira and Gaucher, 2004). A lateEdiacaran age assignment is consistent with availablePb-Pb ages of carbonates of the CCG (553 ± 30 Ma:Fölling et al., 2000) and U-Pb detrital zircons ages of theoverlying Kansa Group (518 ± 9 Ma: Barnett et al., 1997).
No biostratigraphic inferences can be drawn from theacritarchs occurring in the Sardinia Formation yet,because taxonomic assignement of the most commonacritarchs (species A and B) is prevented by an acid-resistant mineral encrustation that obscures surfaceornamentation.
Stratigraphic correlationWhereas our studies show that the assemblagespreserved in the carbonatic Lime Bank and Kaanformations are dominated by Bavlinella, the assemblageoccurring in the Klein Rivier Formation is dominated byleiospheres (Figure 5). Soldadophycus dominates theassemblage recovered from the Van Stadens Formation,which shows a lower degree of thermal overprint. The Bavlinella assemblage from the Lime Bank andKaan formations are identical, and very similar to theassemblage of the Nooitgedagt Member (CCG). The Soldadophycus assemblage of the Van StadensFormation resembles that occurring in the Kombuis
Member and, to a lesser extent, the assemblage of thelower Huis Rivier Formation. Leiospheres dominate boththe Klein Rivier Formation and the stratigraphic intervalconsisting of the Groenefontein and Huis Rivierformations of the CCG. The micropalaeontological dataobtained may be interpreted in two different ways,namely by assuming considerable facies changesbetween the CCG and GG (Figure 12a), or by tectonicduplication in the GG (Figure 12b). By assumingconsiderable facies changes, the carbonatic Lime BankFormation and most of the Kaan Formation can becorrelated to the Nooitgedagt Member of the CCG(Figure 12a). The upper Kaan and lower Van StadensFormation may correlate to the Kombuis Member of theCCG. The upper Van Stadens Formation would becorrelative of the Groenefontein and Huis RivierFormations.
The acritarch assemblages and lithostratigraphy ofthe GG not only can be explained by facies changes butalso by tectonic duplication, and enable a differentcorrelation of the GG and CCG at formational level(Figure 12b). We evisage that the carbonatic, Bavlinella-dominated, carbonaceous Lime Bank and most of theKaan Formations represent the same stratigraphic unit.Likewise, the shale-rich and turbiditic-sandstonecontaining Kleinrivier and upper Van StadensFormations may represent the same lithostratigraphicunit. They can be correlated to the shaley,Leiosphaeridia-dominated Groenefontein Formation andthe turbiditic-sandstone rich Huis Rivier Formation(Figure 12b). The Soldadophycus-dominated uppermostKaan Formation may be a correlate of the KombuisFormation (CCG), both being characterized by blackcarbonates often containing rounded quartz grains.
Previous workers (e.g. Shone et al., 1990) stated thatthe lithostratigraphic subdivision of the GG isimpracticable. Our work shows that the generallyaccepted lithostratigraphic subdivision of Hill and Nolte(1989) does not represent a true stratigraphic section,but probably the stacking of thrust sheets. However, byusing combined biostratigraphic and lithostratigraphicdata, the original stratigraphic succession can bereconstructed and correlations with the CCG becomepossible.
Regardless the correlation at the formation level, it isclear that the CCG and GG yield almost identicalacritarch assemblages, which are typical of lateEdiacaran successions. Similar microfloras weredescribed from the Holgat Formation of the Port NollothGroup (Gaucher et al., 2005) and the overlying Nama Group (Germs et al., 1986) in southern Africa. The Arroyo del Soldado Group (Uruguay) and theCorumbá Group (Brazil) yielded comparable acritarchassemblages (Gaucher et al., 2003). Pb-Pb datings ofupper Kombuis limestones yielding an age of 553 ± 30and available Sr and C isotopic trends recorded in theCCG (Fölling and Frimmel, 2002 and references therein)are consistent with the late Ediacaran age suggested herefor the unit. Thus, an important basin of late Ediacaran

age represented by the Nama, Cango Caves andGamtoos Groups surrounds the western and southernKalahari Craton.
The age and stratigraphic position of the SardiniaBay Formation has been a matter of debate. Itsstratigraphic position beneath the Peninsula Formationof the Table Mountain Group in the Gamtoos and PortElizabeth areas and the occurrence of Phanerozoic tracefossils (Shone, 1979) indicate that it is older than thePeninsula Formation and Phanerozoic in age. Detritalzircon with ages of ~510 Ma occur in the GraafwaterFormation (Armstrong et al., 1998), which underlies thePeninsula Formation and forms the base of the TableMountain Group in the Cape Town region. Based on theabove mentioned data, the Sardinia Bay Formation hasbeen included in the Table Mountain Sandstone Groupand by various geologists considered to be a correlativeof the Graafwater Formation. Ongoing research onshales of the Sardinia Formation may enable a moreaccurate biostratigraphic correlation of this unit.
ConclusionsThe CCG and GG yield almost identical acritarchassemblages, which are typical of late Ediacaransuccessions worldwide (Germs et al., 1986; Vidal andMoczydlowska-Vidal, 1997; Knoll, 2000; Gaucher, 2000;Gaucher et al., 2003; 2005).
Our micropalaeontological data indicate that the VanStadens Formation is late Ediacaran in age and it is nota correlate of the Kansa Group (former upper KangoGroup), as suggested by Tankard et al. (1982).
Two possible correlations at formational level areenvisaged (Figure 12): (a) assuming considerable facieschanges between the CCG and GG; or (b) assumingtectonic duplication of lithostratigraphic units in the GG.Our study renders support to the latter option.
The microfossil assemblages and lithology indicatethat tectonic duplications of lithostratigraphic unitsoccurred in the GG, and that its lithostratigraphic units can be correlated to the CCG (Figure 12).
The generally accepted lithostratigraphic subdivisionof Hill and Nolte (1989) does not appear to represent atrue stratigraphic section, but is probably the result ofthrust sheet stacking.
The microfloras of the CCG and GG are similar tothose of the Holgat Formation of the Port Nolloth Group (Gaucher et al., 2005), the overlying Nama Group(Germs et al., 1986), the Arroyo del Soldado Group of Uruguay (Gaucher, 2000) and the Corumbá Group ofBrazil (Gaucher et al., 2003).
The occurrence of Leiosphaeridia associated withacritarchs showing mineral encrustations in the SardiniaBay Formation is reported for the first time.
AcknowledgementsWe, and especially the second author of this paper, areindebted to Prof. N.J. Beukes, who made it possible tocarry out our research. Leticia Chiglino carried out mostof the palynological macerations. Alta Germs provided
invaluable support with the compilation of this paper.Field expenses and analyses were financed by aresearch grant from the Sasol Fund of the Faculty ofScience, University of Johannesburg, to G.J.B. Germs.Financial support from the Comisión Sectorial deInvestigación Científica (CSIC, Uruguay) to C. Gaucher(Project CSIC-VSP C-40) is gratefully acknowledged. We thank Andrew Knoll and Udo Zimmermann forconstructive reviews that helped improve a first draft ofthis paper. This publication is a contribution to IGCPProject 478: Neoproterozoic to early Palaeozoic Eventsin southwest-Gondwana.
ReferencesAmm, F.L. (1935). The pre-Cape rocks of the Uitenhage Division.
Transactions Geological Society South Africa, 37 (1934), 69-86.
Armstrong, R., De Wit, M.J., Reid, D., York, D. and Zartman, R. (1998). Cape
Town’s Table Mountain reveals rapid Pan-African uplift of its basement
rocks. Journal of African Earth Sciences, 27, 10-11.
Barnett, W., Armstrong, R.A. and De Wit, M.J. (1997). Stratigraphy of the
upper Neoproterozoic Kango and lower Paleozoic Table Mountain Groups
of the Cape Fold Belt revisited. South African Journal of Geology,
100, 237-250.
Belcher, R.W. and Kisters, A.F. M. (2003). Lithostratigraphic correlations in
the western branch of the Pan-African Saldania Belt: The Malmesbury
Group revisited. South African Journal of Geology, 106, 327-342.
Bell, C.M. (1980). Deformation of the Table Mountain Group in the Cape
Fold Belt south of Port Elizabeth. Transactions Geological Society of South
Africa, 83, 115-124.
Boggiani, P.C., Babinski, M., Yamamoto, J.K., Fairchild, T. , Riccomini, C.,
Diragitch, A.A. and Liu, D. (2005). U-Pb SHRIMP investigation of ash beds
in the Corumbá Group (Ediacaran), Paraguai Belt, Brazil. Extended
Abstracts of the 2nd Symposium Neoproterozoic-Early Palaeozoic Events in
southwestern Gondwana, Windhoek, Namibia, 8-10.
Boggiani, P.C., Sial, A.N., Babinski, M. and Ferreira, V.P. (2003). New carbon
isotopic data from the Corumbá Group as a contribution to a composite
section for the Neoproterozoic III in South America. Programme and
Extended Abstracts of the III International Colloquium Vendian-Cambrian
of W-Gondwana, University of Cape Town, South Africa, 13-16.
Bowring, S., Myrow, P., Landing, E., Ramezani, J. and Grotzinger, J. (2003).
Geochronological constraints on terminal Neoproterozoic events and the
rise of metazoans. Geophysical Research Abstracts, 5, 13219.
Butterfield, N.J., Knoll, A.H. and Sweet, K. (1994). Paleobiology of the
Neoproterozoic Svanbergfjellet Formation, Spitsbergen. Fossils and Strata
34, 1-84.
Downie, C. (1973). Observations on the nature of the Acritarchs.
Palaeontology, 16, 239-259.
Downie, C. and Sarjeant, W.A.S. (1963). On the interpretation of some
hystrichosphere genera. Palaeontology, 6, 83- 96.
Eisenack, A. (1958). Tasmanites Newton 1875 und Leiosphaeridia n.g. als
Gattungen der Hystrichosphaeridia. Paleontographica, Abteilung A,
110, 1- 19.
Evitt, W.R. (1963). A discussion and proposals concerning fossil
dinoflagellates, hystrichospheres, and acritarchs, II. Proceedings of the
National Academy of Sciences USA, 49, 298-302.
Fölling, P.G. and Frimmel, H.E. (2002). Chemostratigraphic correlation of
carbonate successions in the Gariep and Saldania Belts, Namibia and South
Africa. Basin Research, 13, 1-37.
Fölling, P.G., Zartman, R.E. and Frimmel, H.E. (2000). A novel approach to
double-spike Pb-Pb dating of carbonate rocks: examples from
Neoproterozoic sequences in southern Africa. Chemical Geology,
171, 97-122.
Ford, T.D. and Breed, W.J. (1973). The problematical Precambrian fossil
Chuaria. Palaeontology, 16, 535-550.
Frimmel, H.E.; Fölling, P.G. and Diamond, R. (2001). Metamorphism of the
Permo-Triassic Cape Fold Belt and its basement, South Africa. Mineralogy
and Petrology, 73, 325-346.
Frankel, J.J. (1937). The geology of a portion of the Gamtoos Valley.
SOUTH AFRICAN JOURNAL OF GEOLOGY
RECENT ADVANCES IN SOUTH AFRICAN NEOPROTEROZOIC-EARLY PALAEOZOIC BIOSTRATIGRAPHY212

CLAUDIO GAUCHER AND GERARD J.B. GERMS
SOUTH AFRICAN JOURNAL OF GEOLOGY
213
Transactions Geological Society of South Africa, 39 (1936), 263-279.
Gaucher, C. (2000). Sedimentology, palaeontology and stratigraphy of the
Arroyo del Soldado Group (Vendian to Cambrian, Uruguay). Beringeria,
26, 1-120.
Gaucher, C., Boggiani, P.C., Sprechmann, P., Sial, A.N. and Fairchild, T.R.
(2003). Integrated correlation of the Vendian to Cambrian Arroyo del
Soldado and Corumbá Groups (Uruguay and Brazil): palaeogeographic,
palaeoclimatic and palaeobiologic implications. Precambrian Research,
120, 241-278.
Gaucher, C., Chiglino, L. and Pecoits, E. (2004a). Southernmost exposures of
the Arroyo del Soldado Group (Vendian to Cambrian, Uruguay):
palaeogeographic implications for the amalgamation of west-Gondwana.
Gondwana Research, 7, 701-714.
Gaucher, C., Frimmel, H.E. and Germs, G.J.B. (2005). Organic-walled
microfossils and biostratigraphy of the upper Port Nolloth Group
(Namibia): implications for the latest Neoproterozoic glaciations.
Geological Magazine, 142, 539-559.
Gaucher, C. and Germs, G.J.B. (2003). Preliminary biostratigraphic
correlation of the Arroyo del Soldado Group (Vendian to Cambrian,
Uruguay) with the Congo Caves and Nama Groups (South Africa and
Namibia). Revista de la Sociedad Uruguaya de Geologia 3, Publicación
Especial, 1, 141-160.
Gaucher, C., Sial, A.N., Blanco, G. and Sprechmann, P. (2004b).
Chemostratigraphy of the lower Arroyo del Soldado Group (Vendian,
Uruguay) and palaeoclimatic implications. Gondwana Research,
7, 715-730.
Gaucher, C. and Sprechmann, P. (1999). Upper Vendian skeletal fauna of the
Arroyo del Soldado Group, Uruguay. Beringeria, 23, 55-91.
Gaucher, C., Sprechmann, P. and Montaña, J. (1998). New advances on the
geology and paleontology of the Vendian to Cambrian Arroyo del Soldado
Group of the Nico Pérez Terrane of Uruguay. Neues Jahrbuch für Geologie
und Paläontologie, Monatshefte, 1998, 106-118.
Gaucher, C., Sprechmann, P. and Schipilov, A. (1996). Upper and Middle
Proterozoic fossiliferous sedimentary sequences of the Nico Pérez Terrane
of Uruguay: Lithostratigraphic units, paleontology, depositional
environments and correlations. Neues Jahrbuch für Geologie und
Paläontologie, Abhandlungen, 199, 339-367.
German, T.N., Mikhajlova, N.S. and Yankauskas, T.V. (1989).
Sistematicheskoe opisanie mikrofossilij [Systematic description of
microfossils]. In: T. V. Jankauskas (Editor), Mikrofossilii Dokembriya SSSR
[Precambrian microfossils of the USSR], Nauka, Leningrad, USSR, 34-151.
Germs, G. J. B. (1972). New shelly fossils from Nama Group, South West
Africa. American Journal of Science, 272, 752-761.
Germs, G.J.B., Knoll, A.H. and Vidal, G. (1986). Latest Proterozoic
microfossils from the Nama Group, Namibia (South West Africa).
Precambrian Research, 32, 45-62.
Grey, K., Walter, M.R. and Calver, C.R. (2003). Neoproterozoic biotic
diversification: Snowball Earth or aftermath of the Acraman impact?
Geology, 31, 459-462.
Grotzinger, J. P., Bowring, S. A., Saylor, B. Z. and Kaufman, A. J. (1995).
Biostratigraphic and Geochronologic Constraints on Early Animal
Evolution. Science, 270, 598- 604.
Grotzinger, J.P., Watters, W.A. and Knoll, A.H. (2000). Calcified metazoans in
thrombolite-stromatolite reefs of the terminal Proterozoic Nama Group,
Namibia. Paleobiology, 26, 334-359.
Grunow, A., Hanson, R. and Wilson, R. (1996). Were aspects of Pan-African
deformation linked to Iapetus opening? Geology, 24, 1063-1066.
Hartnady, C.J.H., Newton, A.R. and Theron, J.N. (1974). The stratigraphy and
structure of the Malmesburg Group in the southwestern Cape. Bulletin of
the Precambrian Research Unit, University of Cape Town, South Africa,
15, 195-213.
Haughton, S.H., Frommurze, H.F. and Visser, D.J.L. (1937). The geology of a
portion of the coastal belt near the Gamtoos Valley, Cape Province.
Explanation of Sheet 151 Gamtoos River, Geological Survey of South Africa.
Hill, R.S. and Nolte, C.C. (1989). Gamtoos Group. In: M. R. Johnson (Editor),
Catalogue of South African Lithostratigraphic Units, SACS, Council for
Geoscience, South Africa, 1, 2pp.
Hunt, J.M. (1996). Petroleum Geochemistry and Geology. Second Edition.
W.H. Freeman and Co., New York, United States, xx + 743pp.
Hofmann, H.J and Jackson, G.D. (1994). Shale- facies microfossils from the
Proterozoic Bylot Supergroup, Baffin Island, Canada. Journal of
Paleontology, Memoir 37, 1-39.
Jankauskas, T.V. (1989). Mikrofossilii Dokembriia SSSR., Nauka, Leningrad,
USSR, 188pp.
Kisters, A.F.M., Belcher, R.W., Scheepers, R., Rozendaal, A. and Smith
Jordaan, L. (2002). Timing and kinematics of the Colenso Fault: The Early
Paleozoic shift from collisional to extensional tectonics in the Pan African
Saldania Belt, South Africa. South African Journal of Geology, 105, 257-270.
Knoll, A.H. (2000). Learning to tell Neoproterozoic time. Precambrian
Research, 100, 3-20.
Knoll, A.H. and Sweet, K. (1987). Micropaleontology across the Precambrian-
Cambrian boundary in Spitsbergen. Journal of Paleontology, 61, 898-926.
Knoll, A.H., Sweet, K. and Mark, J. (1991). Paleobiology of a Neoproterozoic
tidal flat/lagoonal complex: the Draken Conglomerate Formation,
Spitsbergen. Journal of Paleontology, 65, 531-570.
Le Roux, F.G. (2000). The geology of the Port Elizabeth-Uitenhage area,
Explanation of Sheets 3325 DC and DD, 3425 BA Port Elizabeth, 3325 CD
and 3425 AB Uitenhage, 3325 CB Uitenhage Noord and 3325 DA Addo.
55pp. (scale: 1:50 000). Council for Geoscience, South Africa, 55 pp.
Le Roux, J.P. (1997). Cycle hierarchy of a Neoproterozoic carbonate-
siliciclastic shelf: Matjies River Formation of the Kango Group, South
Africa. South African Journal of Geology, 100, 1-10.
Le Roux, J.P. (1999). Cango Caves Group. In: M. R. Johnson (Editor),
Catalogue of South African Lithostratigraphic Units, SACS, Council for
Geoscience, South Africa, 6, 1-2.
Le Roux, J. P. and Gresse, P. G. (1983). The sedimentary-tectonic realm of
the Kango Group. In: A. P. G. Söhnge and I. W. Hälbich (Editors),
Geodynamics of the Cape Fold Belt, Special Publication of the Geological
Society of South Africa, 12, 33-46.
Mansuy, C. and Vidal, G. (1983). Late Proterozoic Brioverian microfossils
from France: taxonomic affinity and implications of plankton productivity.
Nature, 302, 606-607.
MacGabhann, B.A. (2005). Age constraints on Precambrian glaciations and
subdivision of Neoproterozoic time. Abstracts of the International
Conference on Glacial Sedimentary Processes and Products, University of
Wales, United Kingdom, 1-13.
Moczydlowska, M. (1991). Acritarch biostratigraphy of the Lower Cambrian
and the Precambrian-Cambrian boundary in southeastern Poland. Fossils
and Strata, 29, 1-127.
Moorman, M. (1974). Microbiota of the late Proterozoic Hector Formation,
southwestern Alberta, Canada. Journal of Paleontology, 48, 524-539.
Nolte, C.C. (1990). Structure and tectonostratigraphy of the Gamtoos Belt
between Tweewaters and Classen Point, Eastern Cape Province, Republic
of South Africa. Unpublished M.Sc. thesis, Univ. Port Elizabeth, South
Africa, 236pp.
Palacios, T. (1989). Microfósiles de pared orgánica del Proterozoico Superior
(Región Central de la Península Ibérica). Memorias del Museo
Paleontológico de la Universidad de Zaragoza, 3, 1-91.
Pflug, H.D. and Reitz, E. (1992). Palynostratigraphy in Phanerozoic and
Precambrian Metamorphic Rocks. In: M. Schindlowski, S.Golubic, M. M.
Kimberley, D. M. McKirdy and P.A. Trudinger (Editors), Early Organic
Evolution: Implications for Mineral and Energy Resources. Springer, Berlin,
Germany, 509-518.
Rozendaal, A., Gresse, P.G., Scheepers, R. and Le Roux, J.P. (1999).
Neoproterozoic to Early Cambrian crustal evolution of the Pan-African
Saldania Belt, South Africa. Precambrian Research, 97, 303-323.
SACS (South African Committee for Stratigraphy) (1980). Stratigraphy of
South Africa, Part 1 (L.E. Kent, Compiler). Lithostratigraphy of the
Republics of Bophuthatswana, Transkei and Venda. Handbook of the
Geological Survey of South Africa, 8, 690pp.
Saylor, B. Z., Kaufman, A. J., Grotzinger, J. P. and Urban, F. (1998).
A composite reference section for terminal Proterozoic strata of southern
Namibia. Journal of Sedimentary Reserach, 68, 1223-1235.
Scheepers, R. (1995). Geology, geochemistry and petrogenesis of Late
Precambrian S-, I- and A- type granitoids in the Saldania Belt, Western
Cape Province, South Africa. Journal of African Earth Sciences, 21, 35-38.
Schopf, J.W. (1968). Microflora of the Bitter Springs Formation, Late
Precambrian, Central Australia. Journal of Paleontology, 42, 651- 688.
Schopf, J.W. (1992). Atlas of representative Proterozoic microfossils. In: J. W.
Schopf and C. Klein (Editors), The Proterozoic Biosphere-A
Multidisciplinary Study, Cambridge University Press, United Kingdom,
1054-1117.

Shone, R.W. (1979). Sardinia Bay excursion. In: I.C. Rust (Editor), Excursion
Guide Book 1 to 4, Geocongress ’79, Geological Society South Africa,
171-180.
Shone, R.W. (1983). The Geology of the Sardinia Bay Formation,
Unpublished Ph.D. thesis, University of Port Elizabeth, South Africa, 405pp.
Shone, R.W., Nolte, C.C. and Booth, P.W.K. (1990). Pre-Cape rocks of the
Gamtoos area-a complex tectonostratigraphic package preserved as a horst
block. South African Journal of Geology, 93, 616-621.
Silva, L.C.D., Gresse, P.G., Scheepers, R., McNaughton, N.J., Hartmann, L.A.
and Fletcher, I. (2000). U-Pb SHRIMP and Sm-Nd age on the timing and
sources of the Pan African Cape Granite Suite, South Africa. Journal of
African Earth Sciences, 30, 795-815.
Stanier, R.Y.; Sistrom, W.R.; Hansen, T.A.; Whitton, B.A.; Castenholz, R.W.;
Pfenning, N.; Gorlenko, V.N.; Kondratieva, E.N.; Eimhjellen, K.E.;
Whittenbury, R.; Gerna, R.L. and Trüper, H.G. (1978). Proposal to place
nomenclature of the Cyanobacteria (blue-green algae) under the rules of
the International Code of Nomenclature of Bacteria. International Journal
of Systematic Bacteriology, 28, 335-336.
Staplin, F.L., Jansonius, J. and Pocock, S.A.J. (1965). Evaluation of some
acritarchous hystrichosphere genera. Neues Jahrbuch Geologie und
Paläontologie, Abhandlungen, 123, 167-201.
Stump, E. (1976). On the Late Precambrian –Early Paleozoic metavolcanics
and metasedimentary rocks of the Queen Maud Mountains, Antarctica and
a comparison with rocks of similar age from southern Africa. Report of the
Institute for Polar Studies, Ohio State University, United States of America,
62, 1-212.
Tankard, A.J., Jackson, M.P.A., Eriksson, K.A., Hobday, D.K., Hunter, D.R.
and Minter, W.E.L. (1982). Crustal evolution of Southern Africa. Springer-
Verlag, New York, United States of America, 523pp.
Teichmüller, M., Littke, R. and Robert, P. (1998). Coalification and maturation.
In: Taylor, G.H., Teichmüller, M., Davis, A., Diessel, C.F.K., Littke, R. and
Robert, P. (Editors), Organic Petrology, Gebrüder Borntraeger, Berlin,
Germany, 86-174.
Teixeira, A.L. and Gaucher, C. (2004). Bacias do Estágio de Transição
dos setores Meridional (parcial) e Central da Província Mantiqueira.
In: V. Mantesso-Neto, A. Bartorelli, C. D. R. Carneiro and B. B. Brito-Neves
(Editors), Geologia do Continente Sul-Americano: Evolução da Obra de
Fernando Flávio Marques de Almeida, Editora Beca, São Paulo, Brazil,
503-525.
Theron, J.N., Gresse, P.G., Siegfried, H.P. and Rogers, J. (1992). The Geology
of the Cape Town Area: Explanation Sheet 3318. Geological Survey South
Africa, 140pp.
Toerien, D.K. and Hill, R.S. (1991). The geology of the Port Elizabeth area:
Explanations Sheet 3324 (scale: 1:50.000), Geological Survey South Africa,
35pp.
Vidal, G. (1976). Late Precambrian microfossils from the Visingsö Beds in
southern Sweden. Fossils and Strata, 9, 1-57.
Vidal, G. and Moczydlowska-Vidal, M. (1997). Biodiversity, speciation, and
extinction trends of Proterozoic and Cambrian phytoplankton.
Paleobiology, 23, 230-246.
Vidal, G. and Nystuen, J.P. (1990). Micropaleontology, depositional
environment and biostratigraphy of the upper Proterozoic Hedmark
Group, southern Norway.-American Journal of Science, 290-A, 261-294.
Vidal, G., Palacios, T., Gamez-Vintaned, J.A., Diez Balda, M.A. and Grant,
S.W.F. (1994). Neoproterozoic-early Cambrian geology and palaeontology
of Iberia. Geological Magazine, 131, 729- 765.
Volkova, N.A. (1968) Akritarkhi dokembrijskikh i nizhnekembrijskikh
otlozhenij Estonii [Acritarchs from the Precambrian and Cambrian deposits
of Estonia]. In: N.A. Volkova, Z.A. Zhuravleva, V.E. Zabrodin and B. S.
Klinger (Editors), Problematiki pogranichnykh sloev rifeya i kembriya
Russkoj platformy, Urala I Kazakhstana [Problematica of Riphean and
Cambrian layers of the Russian platform, Urals and Kazakhstan], Nauka,
Moskow, p, 8-36.
Volkova, N. A. (1985). Acritarchs and other plant microfossils of the East
European Platform. In: B.S. Sokolov and A.B. Iwanowski (Editors):
The Vendian System, Springer, Berlin, Germany, 155-165.
Weber, B. (1991). Microfossils in Proterozoic sediments from the southern
Shackelton Range, Antarctica: a preliminary report. Zeitschrift für
Geologische Wissenschaften, 19, 185-197.
Walcott, C.D. (1899). Precambrian fossiliferous formations. Geological Society
of America Bulletin, 10, 199-244.
Woese, C.R., Fox, G.E. (1977). Phylogenetic structure of the prokaryotic
domain: The primary kingdoms. Proceedings of the National Academy of
Sciences, United States, 74, 5088-5090.
Xiao, S., Knoll, A.H., Yuan, X. and Pueschel, C.M. (2004). Phosphatized
multicellular algae in the Neoproterozoic Doushantuo Formation, China,
and the early evolution of florideophyte red algae. American Journal of
Botany, 91, 214-227.
Yuan, X., Xiao, S., Yin, L., Knoll, A.H., Zhou, C. and Mu, X. (2002).
Doushantuo fossils: life on the eve of animal radiation. University of
Science and Technology of China Press, Beijing, China, 171pp.
Zang, W. (1992). Sinian and Early Cambrian floras and biostratigraphy on the
South China platform. Palaeontographica Abteilung B, 224, 75-119.
Zhang, Y. (1981). Proterozoic stromatolite microfloras of the Gaoyuzhuang
Formation (Early Sinian: Riphean), Hebei, China. Journal of Paleontology,
55, 485-506.
Zhang, Y. (1989). Multicellular tallophytes with differentiated tissues from
late Proterozoic phosphate rocks of South China. Lethaia, 22, 113-132.
Editorial handling J. Gutzmer
SOUTH AFRICAN JOURNAL OF GEOLOGY
RECENT ADVANCES IN SOUTH AFRICAN NEOPROTEROZOIC-EARLY PALAEOZOIC BIOSTRATIGRAPHY214





























