Embed Size (px)
Citation preview

lable at ScienceDirect
International Biodeterioration & Biodegradation 63 (2009) 334–340
Contents lists avai
International Biodeterioration & Biodegradation
journal homepage: www.elsevier .com/locate/ ib iod
Purified Cladonia verticillaris lichen lectin: Insecticidal activity on Nasutitermescorniger (Isoptera: Termitidae)q
Michele D.C. Silva a, Roberto A. Sa a, Thiago H. Napoleao a, Francis S. Gomes a, Nataly D.L. Santos a,Auristela C. Albuquerque b, Haroudo S. Xavier c, Patrıcia M.G. Paiva a, Maria T.S. Correia a,Luana C.B.B. Coelho a,*
a Laboratorio de Glicoproteınas, Departamento de Bioquımica-CCB, Universidade Federal de Pernambuco, Cidade Universitaria, 50670-420 Recife, Pernambuco, Brazilb Departamento de Biologia, Universidade Federal Rural de Pernambuco, Dois Irmaos, 52171-030 Recife, Pernambuco, Brazilc Laboratorio de Farmacognosia, Departamento de Ciencias Farmaceuticas-CCS, Universidade Federal de Pernambuco, Cidade Universitaria, 50740-521 Recife, Pernambuco, Brazil
a r t i c l e i n f o
Article history:Received 29 August 2008Received in revised form15 October 2008Accepted 3 November 2008Available online 5 December 2008
Keywords:Cladonia verticillarisLectinLichenNasutitermes cornigerTermite
q Scientific relevance: Cladonia verticillaris lichen leeasily purified in milligram quantities that may be dein termite management strategies. The effect of Clavhighlights the potential use of this bioactive protein t
* Corresponding author. Tel.: þ55 8121268540; faxE-mail address: [email protected] (L.C.B.B
0964-8305/$ – see front matter � 2008 Elsevier Ltd.doi:10.1016/j.ibiod.2008.11.002
a b s t r a c t
Cladonia verticillaris lichen lectin (ClaveLL) was isolated through Sephadex G-100 gel filtration chro-matography and characterized as a pure lectin through AKTA-FPLC and HPLC systems. The lichen extract(LE), protein fraction (F1) and ClaveLL were assayed to evaluate their potential insecticidal and/orrepellence activities on termite Nasutitermes corniger. LE, F1 and ClaveLL were evaluated for hemag-glutinating activity (HA), protein concentration and presence of secondary metabolites; preparations andactive ClaveLL, free of secondary metabolites, were able to induce termite mortality. ClaveLL LC50 valuesafter 10 days for workers and soldiers were 0.196 and 0.5 mg ml�1, respectively. C. verticillaris prepara-tions are potential tools for researches involving control of termites (or other insects) of economicinterest to wooden industry or agriculture as well as preservation of plant species that are targets oftermites or other plagues.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
Termites constitute a great group of social insects that areecologically beneficial since they are the major responsible fordecomposition of cellulose, the most abundant carbohydratepolymer in the world, acting in biorecycling promoting carbonturnover in the environment. Termites have the ability to digestcellulose due to the presence of three types of cellulases (endo-glucanases, exoglucanases and b-glucosidases) with endogenousand/or symbiotic origin in their gut (Breznak and Brune, 1994).Endogenous enzymes are encoded by genes in the termite genomeand symbiotic enzymes are produced by hindgut symbionts (Zhouet al., 2008). The order Isoptera is constituted of seven families thathave (except Termitidae) cellulose-fermenting protozoa in thehindguts of termites (Tokuda et al., 1999). The flagellates in Ter-mitidae have been lost and the primary digestive role is displayed
ctin (ClaveLL) is a novel lectinveloped as a biotechnical tooleLL on Nasutitermes cornigero enhance termite resistance.: þ55 8121268576.. Coelho).
All rights reserved.
by a consortia of prokaryotes. According to morphological andmolecular features, the Termitidae family is accepted as the mostrecently evolved and derived from Isoptera order (Miura et al.,1998).
Biodeterioration of wood by termites is a serious problem forwood utilization which has economical and environmentalimpacts. According to Korb (2007), an estimate from 2005 put theannual damage by termites at about US$ 50 billion worldwide.Replacement of deteriorated wood also increases the number oftrees cut and the impact of deforestation (Clausen and Yang, 2007).In addition to wood damage, termites are also responsible forbiodeterioration of paintings, ancient books, monuments andbuildings of historical importance. Due to these negative impacts,termites are considered as plague-insects and new effectivemethods for termite control and monitoring have been searched(Koestler et al., 2000).
The genus Nasutitermes belongs to the family Termitidae, whichcontains around 60% of all termite cataloged species and includeswood, grass or soil feeders (Tokuda et al., 1999; Inward et al., 2007).The Nasutitermes constitutes the largest genus of wood-feeding inthe order Isoptera. The presence of bacteria able to promotecellulose and xylan hydrolysis was detected in the gut of Nasuti-termes species (Warnecke et al., 2007). Representatives of these

M.D.C. Silva et al. / International Biodeterioration & Biodegradation 63 (2009) 334–340 335
insects can be found mainly distributed over tropical regions(Bergamaschi et al., 2007) and Nasutitermes species can invadeurban environment in Brazilian semi-arid region attacking woodemployed in structures of buildings (Paes et al., 2007).
The kingdom Plantae contains secondary metabolites andmacromolecules resultant of the primary metabolism that can actas defensive substances against microorganisms, insects andherbivorous or in response to abiotic stress. The main role of plantsecondary metabolites is believed to protect plants from attack bypathogens or predators (Zhao et al., 2005). There are many proteinsin plants, as reviewed by Carlini and Grossi-de-Sa (2002), involvedin defensive mechanisms such as lectins, ribosome-inactivatingproteins, inhibitors of proteolytic enzymes and glycohydrolases,arcelins, chitinases, hemilectins, canatoxin-like protein as well asmodified forms of storage proteins and ureases.
Lectins are proteins of wide distribution in nature and non-immune origin with the ability to recognize a carbohydrate ora glycosylated molecule through their reversible binding sites(Kennedy et al., 1995; Correia et al., 2008). Lectins stand out inbiotechnology due to many applications and other potential utili-ties that are daily evaluated and studied by researchers in lecti-nology. Several lectins have showed effects in different life stages ofmany insect orders such as Coleoptera (Leite et al., 2005; Sadeghiet al., 2006; Macedo et al., 2007), Diptera (Sa et al., 2008b), Hem-iptera (Sauvion et al., 2004), Homoptera (Bandyopadhyay et al.,2001), Hymenoptera (Couty et al., 2001) and Lepidoptera (Coelhoet al., 2007; Macedo et al., 2007). The unique report of lectintoxicity on an insect of Isoptera order is shown by the lethal effectof lectin from the heartwood of a timber tree (Myracrodruonurundeuva) on N. corniger (Sa et al., 2008a). This lectin inducedmortality of workers and soldiers with LC50 values of0.248 mg ml�1 and 0.199 mg ml�1, respectively. The authorsconcluded that this lectin may play a role in the natural resistanceof M. urundeuva.
Lectins can be isolated from lichens. The involvement of theselectins in the symbiotic establishment of lichens and their possiblephysiological role has been studied and suggested (Elifio et al.,2000; Molina and Vicente, 2000). ClaveLL is a new lectin from C.verticillaris lichen that has been purified in our laboratory. Potentialantimicrobial effects and possible use of ClaveLL as a histochemicalmarker are currently evaluated.
In this work, we evaluated the insecticidal and repellent prop-erties of ClaveLL, a pure lectin sample (free of secondary metabo-lites that can interfere in insecticidal assay), against N. corniger,a wood-damaging termite species. Successful development ofalternative insecticides and repellents will preserve existing woodstructures as well as lower the extinction danger of overexploitedspecies. Additionally, a lichen extract and a protein fraction from C.verticillaris used in ClaveLL purification protocol were also investi-gated for insecticidal and repellent effects.
2. Material and methods
2.1. Protein content
The protein concentration was estimated in all samplesaccording to Lowry et al. (1951) using bovine serum albumin (31–500 mg/mL) as standard.
2.2. Lectin purification
C. verticillaris lichen was collected at Alhandra City, State ofParaıba, northeastern Brazil and taxonomic identification waskindly performed by Dr. Eugenia Cristina Gonçalves Pereira (Uni-versidade Federal de Pernambuco). C. verticillaris lichen lectin(ClaveLL) was obtained through a sequential purification protocol.
Lichen was dried at 28 �C and an overnight extraction (16 h; 28 �C)was performed in 0.15 M sodium phosphate buffer pH 7.0 con-taining 0.15 M NaCl (PBS), followed by filtration, centrifugation(3000 g, 4 �C, 15 min) and protein precipitation with 30% ammo-nium sulphate for 4 h at 28 �C. The 0–30% precipitate fraction (F1)was obtained by centrifugation (3000 g, 4 �C, 15 min), submitted toprotein quantification and loaded (18 mg of protein) onto a gelfiltration Sephadex G-100 (1.4� 63 cm) column aiming to obtainpure and active ClaveLL. The lichen extract (LE), protein fraction (F1)and ClaveLL samples obtained in the purification process had theirhemagglutinating activity (HA) evaluated with 2.5% (v/v) glutaral-dehyde-treated erythrocyte suspension in 0.15 M NaCl (Bing et al.,1967) according to Correia and Coelho (1995). HA (titer) wasdefined as the lowest dilution of the sample that showed hemag-glutination. Specific HA was defined as the ratio between the titerand protein concentration (mg ml�1). HA inhibition assays wereperformed with 200 mM N-acetylglucosamine and 0.5 mg ml�1
solution of glycoproteins from rabbit and bovine fetal serum.
2.3. Phytochemical analysis
Phytochemical evaluation of LE, F1 and ClaveLL samples in PBSand of a lichen methanolic extract (an infusion of 3 g of trituratedlichen in methanol prepared under constant agitation for 30 min)was performed. Samples were analyzed by thin layer chromatog-raphy (TLC) on silica sheet (Merck, Germany). The analyses usedseveral systems of development as mobile phase, reagents foradequate revelation and chromatographic standards.
Each sample was investigated for the presence of: (I) alkaloids(EtOAc–HCOOH–AcOH–H2O [100:11:11:26 v/v] as mobile phase andDragendorff’s reagent for revelation; Wagner and Bladt, 1996); (II)terpenoids and steroids (EtOAc–HCOOH–AcOH–H2O [100:0.5:0.5:0.5v/v] and Liebermann–Burchard’s reagent; Harborne, 1998); (III)saponins (EtOAc–HCOOH–AcOH–H2O [100:11:11:26 v/v] and ani-saldehyde for revelation; Wagner and Bladt, 1996); (IV) iridoids(EtOAc–HCOOH–AcOH–H2O [100:11:11:26 v/v] and vanillin-sulphuric acid for revelation; Wagner and Bladt, 1996); (V) sugars(n-BuOH–Me2CO–phosphate buffer pH 5.0 [40:50:10 v/v] and 2,3,5-triphenyltetrazolium chloride to reveal; Wallenfels, 1950); (VI)coumarins (Et2O–toluene–AcOH 10% [50:50:50 v/v] and UV 365 nmto detect; Wagner and Bladt, 1996); (VII) cinnamic derivatives, phe-nylpropanoglucosides, flavonoids and phenolic acids (EtOAc–HCOOH–AcOH–H2O [100:11:11:26 v/v] and Neu’s reagent to reveal;Wagner and Bladt, 1996; Markhan, 1982; Neu, 1956); (VIII) conden-sed proanthocyanidins and leucoanthocyanidins (EtOAc–HCOOH–AcOH–H2O [100:11:11:26 v/v] and vanillin-chloridric acid to reveal;Roberts et al.,1956); and (IX) hydrolysable tannins (n-BuOH–Me2CO–phosphate buffer pH 5.0 [40:50:10 v/v] and 1% iron alum to reveal;Stiasny, 1912).
2.4. Insecticidal assay
The termite species N. corniger was collected in the campus ofthe Universidade Federal Rural de Pernambuco and identified by Dr.Luiz Roberto Fontes (Superintendencia de Controle de Endemias,SUCEN, Brazil). Termite colonies were kept at vegetation housefrom the Departamento de Agronomia from Universidade FederalRural de Pernambuco.
Termiticidal activity was evaluated by a bioassay based on themethod described by Sa et al. (2008a). Assays were carried out inPetri plates (90�15 mm) with the lower plate covered with filterpaper. Disks of filter paper (4 cm diameter) impregnated with200 mL of LE or F1 samples at 20, 2 or 1 mg ml�1 of protein (cor-responding to 320, 32 or 16 mg of protein cm�2, respectively) wereplaced in plates for feeding by the insects. Treatments with thelectin contained ClaveLL samples at 0.6, 0.4, 0.2 or 0.05 mg ml�1

M.D.C. Silva et al. / International Biodeterioration & Biodegradation 63 (2009) 334–340336
corresponding to 9.6, 6.4, 3.2 or 0.8 mg of protein cm�2, respectively.PBS was used as negative control. The paper disks containing thesamples were dried at 28 �C and then placed in the plates. Twentyinsects (16 workers and 4 soldiers) from termite colonies werecarefully transferred to the plates. All plates were kept in darknessat 28 �C. Monitoring of assays was performed daily to detect deathof insects and to guarantee the humidity of the plates by addinga water drop to the paper covering the plate until all of the insectshad died. Assays were made in quintuplicate. Total experimentalduration was defined as the number of days to reach 100% mortalityrate in negative control.
The survival rates (%) were obtained for each treatment andexpressed as mean� standard deviation (SD). Statistical analysiswas performed using the computer package Statplus� 2006 (Ana-lystSoft, Canada) for determination of the protein concentrationrequired to kill 50% of insects in each treatment (LC50) for LE, F1
(after 4 days) and ClaveLL (after 10 days) by probit analysis methodwith a reliability interval of 95%. Significant differences betweeneach possible pair of treatments were established performingStudent’s t-test (p< 0.05), assuming equal variances (homosce-dastic) and considering the accumulative mortality during allassays using Origin� 6.0 (Microcal, USA) software.
2.5. Repellence assay
The potential repellent property from LE, F1 and ClaveLL wasevaluated using an assay based on Sa et al. (2008a). A 2% (w/v) agarsolution in distilled water was added to Petri plates until the borderof the plates so that there was no space between the superiorsurface of agar and plate covers. After solidification wells weremade in agar by the removal of 1 central cylinder (25 mm diam-eter) and 8 peripheral cylinders (6 mm diameter). Disks of filterpaper (6 mm diameter) were impregnated with LE or F1 samples at300, 150, 30 or 15 mg of total protein (corresponding to 1060, 530,106 or 53 mg cm�2, respectively) and ClaveLL sample at 11.5, 5.75,2.25, 1.5 or 0.75 mg of total lectin (corresponding to 40.6, 20.3, 7.95,5.3 or 2.65 mg lectin cm�2, respectively). PBS was used as negativecontrol. The paper disks were put to dry at 28 �C and placed in theperipheral wells. Each protein concentration was present in doublein plates and each plate was carried out in triplicate, performinga total of 6 replicates for each protein concentration assayed.Twenty insects (16 workers and 4 soldiers) were carefully trans-ferred to central wells in the plates that were maintained indarkness at 28 �C. Plates were observed daily to evaluate absenceor presence of termites in wells, construction standards of tunnelsin agar, closing by insects of constructed galleries and death ofinsects.
3. Results
3.1. Lectin isolation
An amount of 20 g from C. verticillaris flour allowed obtaining1162.50 mg (total protein) in LE, 510.14 mg in F1 and 18.58 mg ofpurified ClaveLL using a simple protocol. ClaveLL represents 0.093%of the dry weight of crude flour, and 1.59% and 3.64% of total proteinpresent in LE and F1, respectively. LE, F1 and ClaveLL agglutinatedglutaraldehyde-treated rat, chicken, quail, rabbit (mostly) andhuman (all ABO system) erythrocytes. LE and F1 showed highprotein concentration (7.75 and 19.85 mg ml�1, respectively), andspecific HA (33 and 412, respectively). HA of ClaveLL (specific HA of780.5) was reduced in the presence of N-acetylglucosamine andabolished in the presence of rabbit and bovine fetal serum. Allprotein samples including active ClaveLL and free of secondarymetabolites were chosen to evaluate their potential insecticidalproperties via lectin effect.
3.2. Phytochemical analysis
Phytochemical evaluation of LE, F1 and ClaveLL samples showednone of the secondary metabolites (alkaloids, terpenoids, steroids,saponins, iridoids and polyphenols as coumarins, cinnamic deriv-atives, phenylpropanoglucosides, flavonoids, phenolic acids,condensed proanthocyanidins, leucoanthocyanidins and hydro-lysable tannins) evaluated.
Phytochemical evaluation of lichen methanolic extract wasperformed aiming to analyze the potential presence of terpenoids.TLC revealed bands corresponding to b-amirin and b-sitosterolstandards and additional bands of smaller polarity that characterizetriterpenes and steroids, which reacted with Liebermann–Burch-ard’s reagent. No other secondary metabolite was detected in themethanolic extract.
3.3. Termiticidal activity of lichen extract and fraction
LE, F1 and ClaveLL in PBS, samples free of secondary metabolites,were submitted to insecticidal assay against N. corniger aiming toevaluate termiticidal activity. LE, F1 and ClaveLL in all testedconcentrations induced 100% mortality of workers and soldiers.Also, in all assays survival rate of 50% was detected in a smaller timethan that obtained for the control.
LE and F1 containing 20, 2 or 1 mg ml�1 of protein (corre-sponding to 320, 32 or 16 mg of protein cm�2, respectively) induced100% mortality of workers after 11, 13 and 14 days (LE) and 7, 10 and12 days (F1), in respective concentrations. Survival rate of 50% for LEtreatment was observed after 2 days (20 mg ml�1) and at fifth andsixth days (2 and 1 mg ml�1, respectively). F1 treatments’ survivalrate of 50% was reached after 2 days (20 mg ml�1) and betweenfourth and fifth days (with 2 and 1 mg ml�1, respectively). Mortalityinduction on workers by LE and F1 was related to protein concen-tration (Fig. 1A and C). In negative control, 100% of mortality wasobserved after 17 days and survival rate was lower than 50% around7 days. There was a significant difference among LE treatmentlevels 20, 2 and 1 mg ml�1 in the induction of worker’s death.
LE and F1 samples also induced 100% mortality of soldiers after 7,12 and 14 days (LE) and 7, 9 and 13 days (F1) at 20, 2 or 1 mg ml�1 ofprotein, respectively. Survival rate of 50% was observed at 2, 6 and 7days for LE and after 2, 5 and 6 days for F1 (concentrations abovecited, respectively). Mortality induction on soldiers by LE and F1
was also related to protein concentration (Fig. 1B and D). In nega-tive control, 100% mortality of soldiers was observed after 16 daysand 50% survival rate after 7 days.
LE and F1 LC50 values after 4 days for workers and soldiers wereshown in Table 1. Although LC50 values indicate that LE was moreefficient than F1, comparison between LE and F1 effects on workersor soldiers showed statistically significant differences only ata concentration of 20 mg ml�1 (320 mg cm�2), as illustrated inmortality data for workers and soldiers compared to negativecontrol.
3.4. Termiticidal activity of ClaveLL
Analysis of worker’s survival revealed that treatment withClaveLL at 0.6 mg ml�1 (9.6 mg cm�2) induced death of all workersafter 15 days. Treatments with 0.4 (6.4 mg cm�2), 0.2 (3.2 mg cm�2)and 0.05 (0.8 mg cm�2) mg ml�1 of lectin promoted 100% mortalityafter 19, 23 and 26 days, respectively. Survival rate of 50% for thesetreatments was observed around 9 and 10 days for treatment with0.6 and 0.4 mg ml�1 ClaveLL, and around 12 and 14 days for treat-ments with 0.2 and 0.05 mg ml�1 ClaveLL. Mortality induction on N.corniger workers by ClaveLL was also proportional to proteinconcentration (Fig. 2A). LC50 value for workers after 12 days was0.196 mg ml�1 (or 3.13 mg cm�2). In negative control 100% of
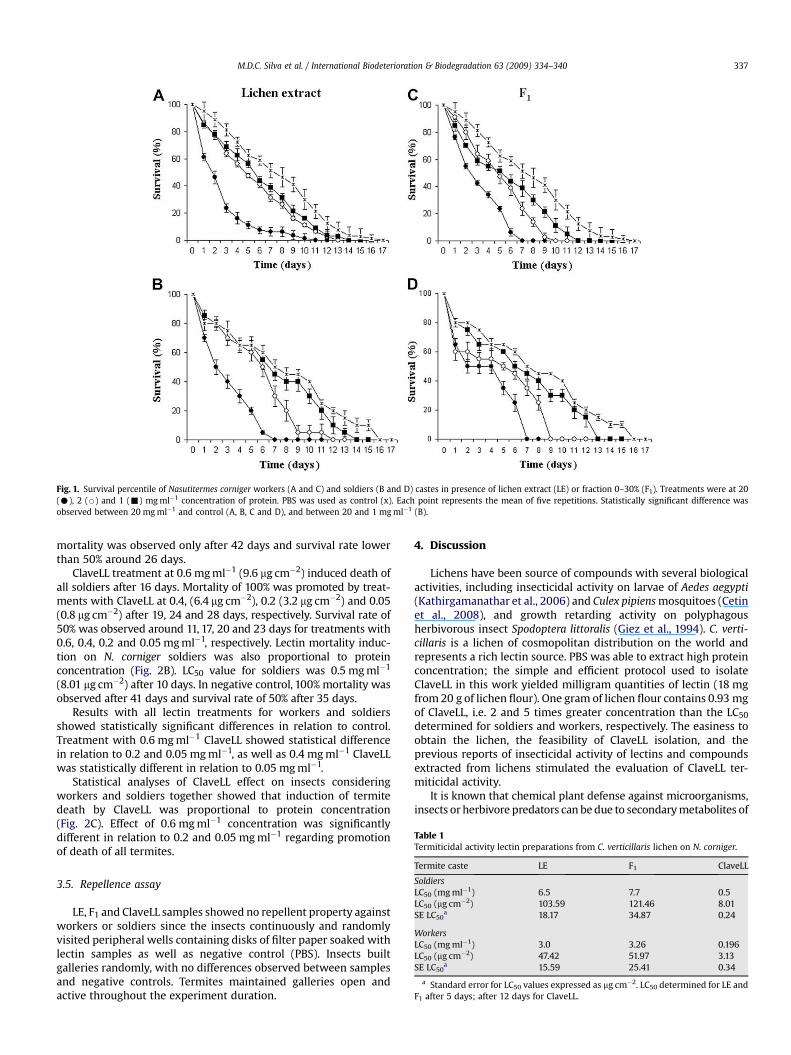
Fig. 1. Survival percentile of Nasutitermes corniger workers (A and C) and soldiers (B and D) castes in presence of lichen extract (LE) or fraction 0–30% (F1). Treatments were at 20(C), 2 (B) and 1 (-) mg ml�1 concentration of protein. PBS was used as control (x). Each point represents the mean of five repetitions. Statistically significant difference wasobserved between 20 mg ml�1 and control (A, B, C and D), and between 20 and 1 mg ml�1 (B).
Table 1Termiticidal activity lectin preparations from C. verticillaris lichen on N. corniger.
M.D.C. Silva et al. / International Biodeterioration & Biodegradation 63 (2009) 334–340 337
mortality was observed only after 42 days and survival rate lowerthan 50% around 26 days.
ClaveLL treatment at 0.6 mg ml�1 (9.6 mg cm�2) induced death ofall soldiers after 16 days. Mortality of 100% was promoted by treat-ments with ClaveLL at 0.4, (6.4 mg cm�2), 0.2 (3.2 mg cm�2) and 0.05(0.8 mg cm�2) after 19, 24 and 28 days, respectively. Survival rate of50% was observed around 11, 17, 20 and 23 days for treatments with0.6, 0.4, 0.2 and 0.05 mg ml�1, respectively. Lectin mortality induc-tion on N. corniger soldiers was also proportional to proteinconcentration (Fig. 2B). LC50 value for soldiers was 0.5 mg ml�1
(8.01 mg cm�2) after 10 days. In negative control, 100% mortality wasobserved after 41 days and survival rate of 50% after 35 days.
Results with all lectin treatments for workers and soldiersshowed statistically significant differences in relation to control.Treatment with 0.6 mg ml�1 ClaveLL showed statistical differencein relation to 0.2 and 0.05 mg ml�1, as well as 0.4 mg ml�1 ClaveLLwas statistically different in relation to 0.05 mg ml�1.
Statistical analyses of ClaveLL effect on insects consideringworkers and soldiers together showed that induction of termitedeath by ClaveLL was proportional to protein concentration(Fig. 2C). Effect of 0.6 mg ml�1 concentration was significantlydifferent in relation to 0.2 and 0.05 mg ml�1 regarding promotionof death of all termites.
Termite caste LE F1 ClaveLL
SoldiersLC50 (mg ml�1) 6.5 7.7 0.5LC50 (mg cm�2) 103.59 121.46 8.01SE LC50
a 18.17 34.87 0.24
WorkersLC50 (mg ml�1) 3.0 3.26 0.196LC50 (mg cm�2) 47.42 51.97 3.13SE LC50
a 15.59 25.41 0.34
a Standard error for LC50 values expressed as mg cm�2. LC50 determined for LE andF1 after 5 days; after 12 days for ClaveLL.
3.5. Repellence assay
LE, F1 and ClaveLL samples showed no repellent property againstworkers or soldiers since the insects continuously and randomlyvisited peripheral wells containing disks of filter paper soaked withlectin samples as well as negative control (PBS). Insects builtgalleries randomly, with no differences observed between samplesand negative controls. Termites maintained galleries open andactive throughout the experiment duration.
4. Discussion
Lichens have been source of compounds with several biologicalactivities, including insecticidal activity on larvae of Aedes aegypti(Kathirgamanathar et al., 2006) and Culex pipiens mosquitoes (Cetinet al., 2008), and growth retarding activity on polyphagousherbivorous insect Spodoptera littoralis (Giez et al., 1994). C. verti-cillaris is a lichen of cosmopolitan distribution on the world andrepresents a rich lectin source. PBS was able to extract high proteinconcentration; the simple and efficient protocol used to isolateClaveLL in this work yielded milligram quantities of lectin (18 mgfrom 20 g of lichen flour). One gram of lichen flour contains 0.93 mgof ClaveLL, i.e. 2 and 5 times greater concentration than the LC50
determined for soldiers and workers, respectively. The easiness toobtain the lichen, the feasibility of ClaveLL isolation, and theprevious reports of insecticidal activity of lectins and compoundsextracted from lichens stimulated the evaluation of ClaveLL ter-miticidal activity.
It is known that chemical plant defense against microorganisms,insects or herbivore predators can be due to secondary metabolites of
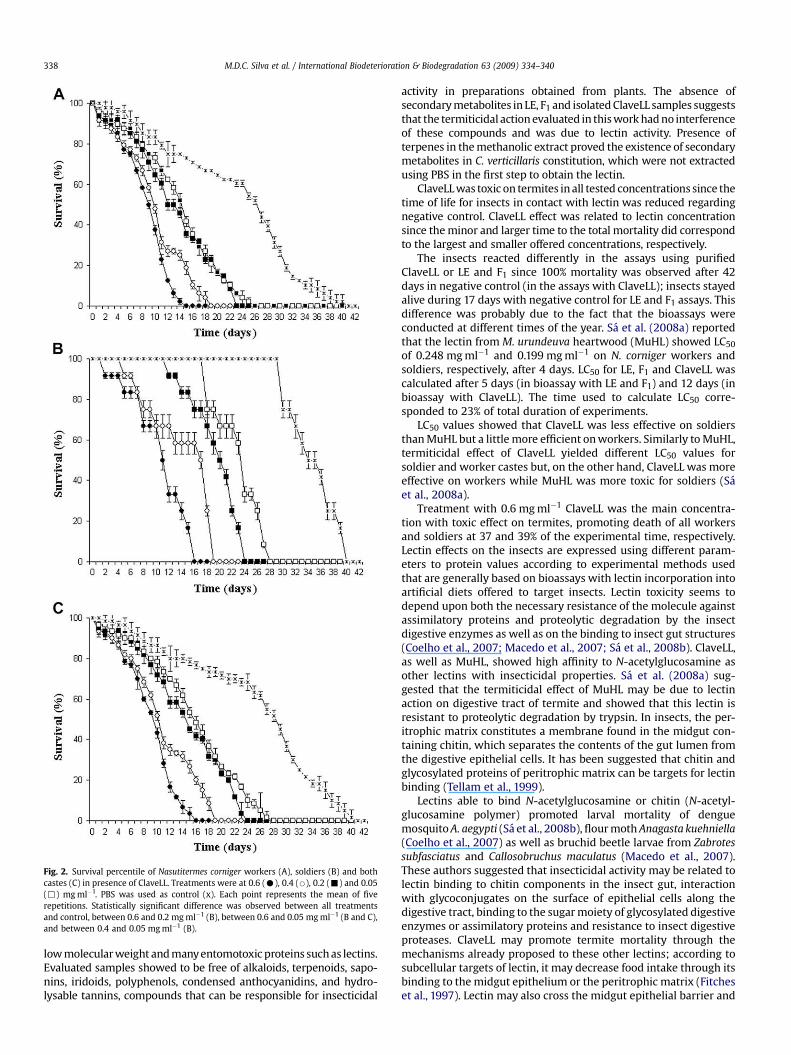
Fig. 2. Survival percentile of Nasutitermes corniger workers (A), soldiers (B) and bothcastes (C) in presence of ClaveLL. Treatments were at 0.6 (C), 0.4 (B), 0.2 (-) and 0.05(,) mg ml�1. PBS was used as control (x). Each point represents the mean of fiverepetitions. Statistically significant difference was observed between all treatmentsand control, between 0.6 and 0.2 mg ml�1 (B), between 0.6 and 0.05 mg ml�1 (B and C),and between 0.4 and 0.05 mg ml�1 (B).
M.D.C. Silva et al. / International Biodeterioration & Biodegradation 63 (2009) 334–340338
low molecular weight and many entomotoxic proteins such as lectins.Evaluated samples showed to be free of alkaloids, terpenoids, sapo-nins, iridoids, polyphenols, condensed anthocyanidins, and hydro-lysable tannins, compounds that can be responsible for insecticidal
activity in preparations obtained from plants. The absence ofsecondary metabolites in LE, F1 and isolated ClaveLL samples suggeststhat the termiticidal action evaluated in this work had no interferenceof these compounds and was due to lectin activity. Presence ofterpenes in the methanolic extract proved the existence of secondarymetabolites in C. verticillaris constitution, which were not extractedusing PBS in the first step to obtain the lectin.
ClaveLL was toxic on termites in all tested concentrations since thetime of life for insects in contact with lectin was reduced regardingnegative control. ClaveLL effect was related to lectin concentrationsince the minor and larger time to the total mortality did correspondto the largest and smaller offered concentrations, respectively.
The insects reacted differently in the assays using purifiedClaveLL or LE and F1 since 100% mortality was observed after 42days in negative control (in the assays with ClaveLL); insects stayedalive during 17 days with negative control for LE and F1 assays. Thisdifference was probably due to the fact that the bioassays wereconducted at different times of the year. Sa et al. (2008a) reportedthat the lectin from M. urundeuva heartwood (MuHL) showed LC50
of 0.248 mg ml�1 and 0.199 mg ml�1 on N. corniger workers andsoldiers, respectively, after 4 days. LC50 for LE, F1 and ClaveLL wascalculated after 5 days (in bioassay with LE and F1) and 12 days (inbioassay with ClaveLL). The time used to calculate LC50 corre-sponded to 23% of total duration of experiments.
LC50 values showed that ClaveLL was less effective on soldiersthan MuHL but a little more efficient on workers. Similarly to MuHL,termiticidal effect of ClaveLL yielded different LC50 values forsoldier and worker castes but, on the other hand, ClaveLL was moreeffective on workers while MuHL was more toxic for soldiers (Saet al., 2008a).
Treatment with 0.6 mg ml�1 ClaveLL was the main concentra-tion with toxic effect on termites, promoting death of all workersand soldiers at 37 and 39% of the experimental time, respectively.Lectin effects on the insects are expressed using different param-eters to protein values according to experimental methods usedthat are generally based on bioassays with lectin incorporation intoartificial diets offered to target insects. Lectin toxicity seems todepend upon both the necessary resistance of the molecule againstassimilatory proteins and proteolytic degradation by the insectdigestive enzymes as well as on the binding to insect gut structures(Coelho et al., 2007; Macedo et al., 2007; Sa et al., 2008b). ClaveLL,as well as MuHL, showed high affinity to N-acetylglucosamine asother lectins with insecticidal properties. Sa et al. (2008a) sug-gested that the termiticidal effect of MuHL may be due to lectinaction on digestive tract of termite and showed that this lectin isresistant to proteolytic degradation by trypsin. In insects, the per-itrophic matrix constitutes a membrane found in the midgut con-taining chitin, which separates the contents of the gut lumen fromthe digestive epithelial cells. It has been suggested that chitin andglycosylated proteins of peritrophic matrix can be targets for lectinbinding (Tellam et al., 1999).
Lectins able to bind N-acetylglucosamine or chitin (N-acetyl-glucosamine polymer) promoted larval mortality of denguemosquito A. aegypti (Sa et al., 2008b), flour moth Anagasta kuehniella(Coelho et al., 2007) as well as bruchid beetle larvae from Zabrotessubfasciatus and Callosobruchus maculatus (Macedo et al., 2007).These authors suggested that insecticidal activity may be related tolectin binding to chitin components in the insect gut, interactionwith glycoconjugates on the surface of epithelial cells along thedigestive tract, binding to the sugar moiety of glycosylated digestiveenzymes or assimilatory proteins and resistance to insect digestiveproteases. ClaveLL may promote termite mortality through themechanisms already proposed to these other lectins; according tosubcellular targets of lectin, it may decrease food intake through itsbinding to the midgut epithelium or the peritrophic matrix (Fitcheset al., 1997). Lectin may also cross the midgut epithelial barrier and

M.D.C. Silva et al. / International Biodeterioration & Biodegradation 63 (2009) 334–340 339
pass into the insect circulatory system resulting in a toxic actioninterfering with endogenous lectins involved in self-defensepresent in the haemolymph (Fitches et al., 2001); it may be inter-nalized by endocytotic vesicles into the epithelial cells and blocksnuclear localization and nuclear sequence-dependent proteinimport inhibiting cell proliferation (Yu et al., 1999).
N-acetylglucosamine specific lectins with chitin-binding ability,in general, appear to own insecticidal property; however, plantlectins with different sugar specificities also acted as insecticidalmolecules suggesting that this property is not exclusive to N-ace-tylglucosamine specific lectins. For example, XCL, a Xerocomuschrysenteron (mushroom) lectin with affinity for D-galactose andlactose and to a lesser extent, N-acetyl-D-galactosamine, had effecton Drosophila melanogaster and pea aphid Acyrthosiphon pisumshowing LC50 values of 0.4 mg ml�1 and 0.023 mg ml�1, respec-tively (Trigueros et al., 2003). Gracilaria ornata (a red alga) lectin,GOL, only inhibited by some glycoproteins, caused a significantreduction (65.1%) in Callosobruchus maculates larval survival at 1%(w/w) in relation to control artificial seeds. Also, GOL significantlyaffected the percentage of adult emergence in relation to control(Leite et al., 2005). In other work, Sadeghi et al. (2006), using 14plant lectins (of four lectin families) with several specificities ata concentration of 0.05% (w/v) solution, showed that all lectins haddeterrent effect against oviposition of C. maculatus adults. Talisiaesculenta seed lectin with chitin-binding properties and inhibitedby mannose and glucose produced ca. 90% mortality to C. maculatusand Zabrotes subfasciatus larvae with LD50 and ED50 (decrease of50% weight) of ca. 1% (w/w) for both insects (Macedo et al., 2002).
The precise mechanism of insecticidal action of lectins is stillunknown and it has been suggested that various modes of actionexist to different lectins at the cellular levels not necessarilyimplying in the disruption of cellular function and lysis. Actionmechanism of lectins as toxic molecules can also be related tointerference with the functions of digestive enzymes and assimi-latory proteins inhibiting digestion and absorption causing nutri-tional deprivation (Coelho et al., 2007; Sa et al., 2008b).
N. corniger are insects that have the ability to build tunnels andgalleries and on stress condition at the presence of possible toxicsubstances they can react closing these spaces to avoid physicalcontact (Su et al., 1982). The investigation of repellent activityrevealed that ClaveLL and other samples (LE and F1) did not haverepellent property on termites; the results suggested that theinsects have eaten the paper disk containing lectin and that theinsecticidal effect was due to action of lectin. Similarly to ClaveLL,MuHL did not show repellent properties (Sa et al., 2008a).
C. verticillaris lichen lectin (ClaveLL) is a novel lectin easilypurified in milligram quantities that may be developed asa biotechnical tool in termite management to facilitate plantspecies preservation and the use of wood in high risk environ-ments. The effect of ClaveLL on N. corniger highlights the potentialuse of this bioactive protein to enhance termite resistance.
Acknowledgements
The authors express their gratitude to the Conselho Nacional deDesenvolvimento Cientıfico e Tecnologico (CNPq) and the Coor-denaçao de Aperfeiçoamento de Pessoal de Nıvel Superior (CAPES)for research grants and fellowship (CNPq, LCBBC). They are alsodeeply grateful to Maria Barbosa Reis da Silva and Joao AntonioVirgınio (for technical assistance).
References
Bandyopadhyay, S., Roy, A., Das, S., 2001. Binding of garlic (Allium sativum) leaflectin to the gut receptors of homopteran pests is correlated to its insecticidalactivity. Plant Science 161, 1025–1033.
Bergamaschi, S., Dawes-Gromadzki, T.Z., Luchetti, A., Marini, M., Mantovani, B.,2007. Molecular taxonomy and phylogenetic relationships among AustralianNasutitermes and Tumulitermes genera (Isoptera, Nasutitermitinae) inferredfrom mitochondrial COII and 16S sequences. Molecular Phylogenetics andEvolution 45, 813–821.
Bing, D.H., Weyand, J.G., Stavinsky, A.B., 1967. Hemagglutination with aldehyde-fixed erythrocytes for assay of antigens and antibodies. Proceedings of theSociety for Experimental Biology and Medicine 124, 1166–1170.
Breznak, J.A., Brune, A., 1994. Role of microorganisms in the digestion of lignocel-lulose by termites. Annual Review of Entomology 39, 453–487.
Carlini, C.R., Grossi-de-Sa, M.F., 2002. Plant toxic proteins with insecticidal prop-erties. A review on their potentialities as bioinsecticides. Toxicon 40, 1515–1539.
Cetin, H., Tufan-Cetin, O., Turk, A.O., Tay, T., Candan, M., Yanigoklu, A., Sumbul, H.,2008. Insecticidal activity of major lichen compounds, (�)- and (þ)-usnic acid,against the larvae of the house mosquito, Culex pipiens L. Parasitology Research102, 1277–1279.
Clausen, C.A., Yang, V., 2007. Protecting wood from mould, decay, and termites withmulti-components biocide systems. International Biodeterioration & Biodeg-radation 59, 20–24.
Coelho, M.B., Marangoni, S., Macedo, M.L.R., 2007. Insecticidal action of Annonacoriacea lectin against the flour moth Anagasta kuehniella and the rice mothCorcyra cephalonica (Lepidoptera: Pyralidae). Comparative Biochemistry andPhysiology C 146, 406–414.
Correia, M.T.S., Coelho, L.C.B.B., 1995. Purification of a glucose/mannose specificlectin, Isoform 1, from seeds of Cratylia mollis Mart (camaratu bean). AppliedBiochemistry and Biotechnology 55, 261–273.
Correia, M.T.S., Coelho, L.C.B.B., Paiva, P.M.G., 2008. Lectins, carbohydrate recogni-tion molecules: are they toxic? In: Siddique, Y.H. (Ed.), (Org.), Recent Trends inToxicology, vol. 37 Transworld Research Network, Kerala, pp. 47–59.
Couty, A., de la Vina, G., Clark, S.J., Kaiser, L., Pham-Delegue, M.H., Poppy, G.M., 2001.Direct and indirect sublethal effects of Galanthus nivalis agglutinin (GNA) on thedevelopment of a potato-aphid parasitoid, Aphelinus abdominalis (Hymenop-tera: Aphelinidae). Journal of Insect Physiology 47, 553–561.
Elifio, S.L., da Silva, M.L.C.C., Iacomini, M., Gorin, P.A.J., 2000. A lectin from thelichenized basidiomycete Dictyonema glabratum. New Phytologist 148,327–334.
Fitches, E., Gatehouse, A.M.R., Gatehouse, J.A., 1997. Effects of snowdrop lectin(GNA) delivered via artificial diet and transgenic plants on the development oftomato moth (Lacanobia oleracea) larvae in laboratory and glasshouse trials.Journal of Insect Physiology 14, 727–739.
Fitches, E., Woodhouse, S.D., Edwards, J.P., Gatehouse, J.A., 2001. In vitro and in vivobinding of snowdrop (Galanthus nivalis agglutinin; GNA) and jackbean (Cana-valia ensiformis; Con A) lectins within tomato moth (Lacanobia oleracea) larvae;mechanisms of insecticidal action. Journal of Insect Physiology 47, 777–787.
Giez, I., Lange, O.L., Proksch, P., 1994. Growth retarding activity of lichen substancesagainst the polyphagous herbivorous insect Spodoptera littoralis. BiochemicalSystematics and Ecology 22, 113–120.
Harborne, J.B., 1998. Phytochemical Methods: A Guide to Modern Techniques ofPlant Analysis. Chapman & Hall, London.
Inward, D.J.G., Vogler, A.P., Eggleton, P., 2007. A comprehensive phylogenetic anal-ysis of termites (Isoptera) illuminates key aspects of their evolutionary biology.Molecular Phylogenetics and Evolution 44, 953–967.
Kathirgamanathar, S., Ratnasooriya, W.D., Baekstrom, P., Andersen, R.J.,Karunaratne, V., 2006. Chemistry and bioactivity of Physciaceae lichens Pyxineconsocians and Geterodermia leucomelos. Pharmaceutical Biology 44, 217–220.
Kennedy, J.F., Paiva, P.M.G., Correia, M.T.S., Cavalcanti, M.S.M., Coelho, L.C.B.B., 1995.Lectins, versatile proteins of recognition: a review. Carbohydrate Polymers 26,219–230.
Koestler, R.J., Sardjono, S., Koestler, D.L., 2000. Detection of insect infestation inmuseum objects by carbon dioxide measurement using FTIR. InternationalBiodeterioration & Biodegradation 46, 285–292.
Korb, J., 2007. Termites. Current Biology 17, R995–R999.Leite, Y.F.M.M., Silva, L.M.C.M., Amorim, R.C.N., Freire, E.A., Jorge, D.M.M.,
Grangeiro, T.B., Benevides, N.M.B., 2005. Purification of a lectin from the marinered alga Gracilaria ornata and its effect on the development of the cowpeaweevil Callosobruchus maculatus (Coleoptera: Bruchidae). Biochimica et Bio-physica Acta 1724, 137–145.
Lowry, O.H., Rosebrough, N., Farr, A.L., Randall, R.J., 1951. Protein measurement withthe folin phenol reagent. Journal of Biological Chemistry 193, 265–275.
Macedo, M.L.R., Freire, M.G.M., Novello, J.C., Marangoni, S., 2002. Talisia esculentalectin and larval development of Callosobruchus maculatus and Zabrotes sub-fasciatus (Coleoptera: Bruchidae). Biochimica et Biophysica Acta 1571, 83–88.
Macedo, M.L.R., Freire, M.G.M., Silva, M.B.R., Coelho, L.C.B.B., 2007. Insecticidalaction of Bauhinia monandra leaf lectin (BmoLL) against Anagasta kuehniella(Lepidoptera: Pyralidae), Zabrotes subfasciatus and Callosobruchus maculatus(Coleoptera: Bruchidae). Comparative Biochemistry and Physiology A 146,486–498.
Markhan, K.R., 1982. Techniques of Flavonoid Identification. Academic Press,London.
Miura, T., Maekawa, K., Kitade, O., Abe, T., Matsumoto, T., 1998. Phylogenetic rela-tionships among subfamilies in higher termites (Isoptera: Termitidae) based onMitochondrial COII gene sequences. Annals of the Entomological Society ofAmerica 91, 515–523.
Molina, M.C., Vicente, C., 2000. Purification and characterization of two isolectinswith arginase activity from the lichen Xanthoria parietina. Journal ofBiochemistry and Molecular Biology 33, 300–307.

M.D.C. Silva et al. / International Biodeterioration & Biodegradation 63 (2009) 334–340340
Neu, R.A., 1956. A new reagent for differentiating and determining flavones onpaper chromatograms. Naturwissenschaften 43, 82.
Paes, J.B., Melo, R.R., Lima, C.R., Oliveira, E., 2007. Resistencia natural de setemadeiras ao cupim subterraneo (Nasutitermes corniger Motsch.) em ensaio depreferencia alimentar. Revista Brasileira de Ciencias Agrarias 2, 57–62.
Roberts, E.H., Cartwright, R.A., Wood, D.J., 1956. The flavones of tea. Journal of theScience of Food and Agriculture 7, 637–646.
Sa, R.A., Napoleao, T.H., Santos, N.D.L., Gomes, F.S., Albuquerque, A.C.,Coelho, L.C.B.B., Bieber, L.W., Paiva, P.M.G., 2008a. Induction of mortality onNasutitermes corniger by Myracrodruon urundeuva heartwood lectin. Interna-tional Biodeterioration & Biodegradation 62, 460–464.
Sa, R.A., Santos, N.D.L., Silva, C.S.B., Napoleao, T.H., Gomes, F.S., Cavada, B.S.,Coelho, L.C.B.B., Navarro, D.M.A.F., Bieber, L.W., Paiva, P.M.G., 2008b. Larvicidalactivity of lectins from Myracrodruon urundeuva on Aedes aegypti. ComparativeBiochemistry and Physiology C, doi:10.1016/j.cbpc.2008.08.004.
Sauvion, N., Nardonb, G., Febvay, G., Gategouse, A.M.R., Rahbe, Y., 2004. Binding ofthe insecticidal lectin Concanavalin A in pea aphid, Acyrthosiphon pisum (Harris)and induced effects on the structure of midgut epithelial cells. Journal of InsectPhysiology 50, 1137–1150.
Sadeghi, A., Van Damme, E.J.M., Peumans, W.J., Smagghe, G., 2006. Deterrentactivity of plant lectins on cowpea weevil Callosobruchus maculatus (F.) ovipo-sition. Phytochemistry 67, 2078–2084.
Stiasny, E., 1912. The Qualitative Detection and Differentiation of Vegetable Tannins.Collegium, Haltingen.
Su, N., Tamashiro, M., Yates, J.R., Haverty, M.I., 1982. Effects of behavior in theevaluation of insecticides for prevention of or remedial control of the Formosansubterranean termite. Journal of Economic Entomology 75, 188–193.
Tellam, R.L., Wijffels, G., Wiladsen, P., 1999. Peritrophic matrix proteins. InsectBiochemistry and Molecular Biology 29, 87–101.
Tokuda, G., Lo, N., Watanabe, H., Slaytor, M., Matsumoto, T., Noda, H., 1999. Meta-zoan cellulase genes from termites: intron/exon structures and sites ofexpression. Biochimica et Biophysica Acta 1447, 146–159.
Trigueros, V., Lougarre, A., Ali-Ahmed, D., Rahbe, Y., Guillot, J., Chavant, L.,Fournier, D., Paquereau, L., 2003. Xerocomus chrysenteron lectin: identificationof a new pesticidal protein. Biochimica et Biophysica Acta 1621, 292–298.
Wagner, H., Bladt, S., 1996. Plant Drug Analysis, second ed. Springer, New York.Wallenfels, K., 1950. Detection of reducing sugars in paper chromatogram and
quantitative evaluation. Naturwissenschaften 37, 491–492.Warnecke, F., Luginbuhl, P., Ivanova, N., Ghassemian, M., Richardson, T.H., Stege, J.T.,
Cayouette, M., McHardy, A.C., Djordjevic, G., Aboushadi, N., Sorek, R., Tringe, S.G.,Podar, M., Martin, G.H., Kunin, V., Dalevi, D., Madejska, J., Kirton, E., Platt, D.,Szeto, E., Salamov, A., Barry, K., Mikhailova, N., Kyrpides, N.C., Matson, E.G.,Ottesen, E.A., Zhang, X., Hernandez, M., Murillo, C., Acosta, L.G., Rigoutsos, I.,Tamayo, G., Green, B.D., Chang, C., Rubin, E.M., Mathur, E.J., Robertson, D.E.,Hugenholtz, P., Leadbetter, J.R., 2007. Metagenomic and functional analysis ofhindgut microbiota of a wood-feeding higher termite. Nature 450, 560–565.
Yu, L.G., Fernig, D.G., White, M.R.H., Spiller, D.G., Appleton, P., Evans, R.C.,Grierson, I., Smith, J.A., Davies, H., Gerasimenko, O.V., Petersen, O.H., Milton, J.D.,Rhodes, J.M., 1999. Edible mushroom (Agaricus bisporus) lectin, whichreversibly inhibits epithelial cell proliferation, blocks nuclear localizationsequence-dependent nuclear protein import. Journal of Biological Chemistry274, 4890–4899.
Zhao, J., Davis, L.C., Verpoorte, R., 2005. Research review paper: elicitor signaltransduction leading to production of plant secondary metabolites. Biotech-nology Advances 23, 283–333.
Zhou, X., Wheeler, M.M., Oi, F.M., Scharf, M.E., 2008. Inhibition of termite cellulasesby carbohydrate-based cellulase inhibitors: evidence from in vitro biochemistryand in vivo feeding studies. Pesticide Biochemistry and Physiology 90, 31–41.























