Embed Size (px)
Citation preview

if Factors Affecting The Growth Of
PSYCHROPHDJC MICRO-ORGANISMS
m FOODS
A Review
Technical Bulletin No. 1320
Agricultural Research Service UNITED STATES DEPARTMENT OF AGRICULTURE

f FACTORS AFFECTING THE GROWTH OF
PSYCHROPHILIC
^ MICRO-ORGANISMS
IN FOODS 4 ^
CO ^ " / ■-—■^ A Review 1>AJ^ ^
LLIOTl
^r R. PAUL/ELLIOTT
and
H. DAVID MICHENER V
Western Utilization Research and Development Division
NATIWW. ÄQRICULTURRI llBWOf
RECEIVED MÀKw) - i36b
U. S. DEPT. Of AGRICULTURE
'7 ^ Technical BuDetin No. 1320 \ ^,
Agricultural Research Service^
UKOBD S'^BBS DEPiCRTaOEKT OF AGRICULTURE //

Acknowledgment
The authors acknowledge the assistance of Anne M. Avakian, Librarian of the Western Utilization Research and Development Division.
Names of companies and trade names are used in this publication solely to provide specific information. Mention of a company or a trade name does not constitute a guarantee or warranty of the com- pany or the product by the U.S. Department of Agriculture or an endorsement by the Department over other companies or products not mentioned.
Washington,/! D.C. Issued January, 1965 ^
For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, D.C. 20402 Price 35 cents

Contents Page
INTEODUCTION 1 Psychrophilic micro-organisms in food spoilage 1 Definitions 1
EFFECT OF TEMPERATURE ON MICROBIAL ACTIVITIES 6 Lag phase 6 Logarithmic phase 8 Temperature coefficient (Qw) and temperature characteristic (/*)— 9 Lipids 19 Enzyme production and activity 20 Cell size and cell crop 24 Adaptation 25
EFFECT OF WATER AVAILABILITY ON MICROBIAL ACTIVITIES 28
SPOILAGE OF MEATS AND FISH 32 Effect of low temperature 32 Floral changes in spoilage 34 Inoculum size 40 Freezing and thawing 45 Heat 49 Type of surface 51 Relative humidity 52 Air movement 55 Oxygen 55 Gas storage 55 Packaging 59 Acid 61 Antemortem treatment 61 Autolysis 62 Antibiotics 62 Salt 64
SPOILAGE OF EGGS 65 Penetration of shell eggs 65 Shell coatings 67 Thermostabilization 67 Gas storage 67 Internal defenses 67 Effect of metals on penetration and growth 69 Effect of aging 70 Liquid egg 70
SPOILAGE OF FRUITS 71
SPOILAGE OF VEGETABLES 72
SUMMARY AND CONCLUSIONS 76
REVIEWS AND BIBLIOGRAPHIES ON LOW-TEMPERATURE MICROBIOLOGY 78
LITERATURE CITED 79

Factors Affecting The Growth Of Psychrophîlic Mîcro-Organîsms In Foods:
A Review By R. PAUL ELLIOTT* and H. DAVID MICHENER, Western Utilization Research
and Development Division, Agricultural Research Service
INTRODUCTION
Psychrophilic micro-organisms in food spoilage Psychrophîlic micro-organisms are of major importance in mod-
em food technology, because their activities limit strictly the time that foods can be stored chilled—that is, at refrigeration temperatures above freezing. However, microbial growth is not a problem in frozen foods held at a suitably low storage tempera- ture. They should be stored well below —12*' C, which is about the lowest temperature permitting growth of psychrophiles (827)^ In fact, to minimize nonmicrobial chemical changes, frozen foods are best held at ^17.8° or below {171, W).
Psychrophilic growth does not result in food poisoning. Food- poisoning organisms rarely grow below 10° C, and none has been reported to grow below 3.3**. A review of minimum tempera- tures for growth of psychrophilic and food-poisoning micro-or- ganisms has been published elsewhere {327). An earlier review {US) covered microbiological standards and handling codes for chilled and frozen foods.
The purpose of this review is to bring together pertinent facts on growth of micro-organisms during decomposition of refrig- erated meats, fish, eggs, fruits, and vegetables. Studies on psy- chrophilic growth in other foods or menstrua are included only when they apply to psychrophilic spoilage of these commodities.
For the benefit of the reader who wishes to study phases of low-temperature microbiology not covered in this report, a list of reviews and bibliographies is presented on page 78.
Definitions For the purposes of this review, psychrophiles are defined as
micro-organisms that grow relatively rapidly at 0° C. {229). Psychrophiles always grow more rapidly at higher temperatures
♦Present address: Division of Field Operations, Food and Drug Ad- ministration, U.S. Dept. of Health, Education, and Welfare, Washington, D.C.
* Italic numbers in parentheses refer to literature cited, p. 79. 1
2 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
than they do at 0°. Optima for most are at 20'' to 30° ; for some at 30° to 45° ; for a few at about 15° or below (232, 327, 5U).
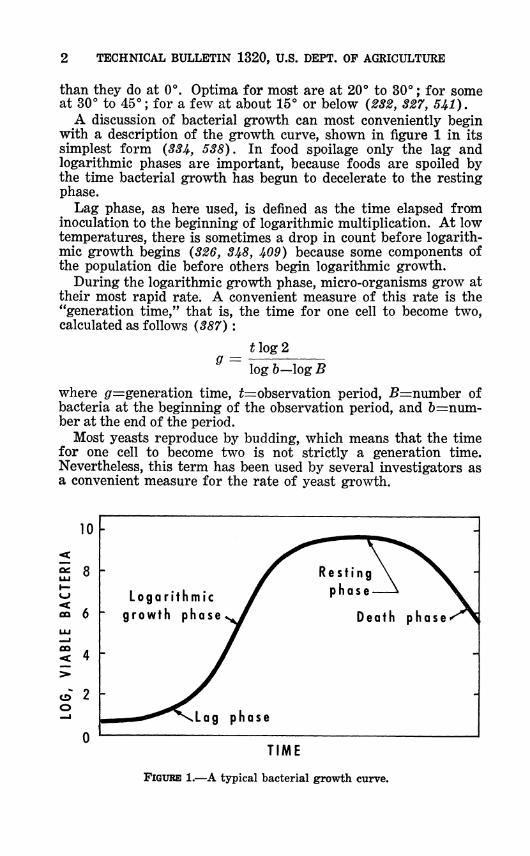
A discussion of bacterial growth can most conveniently begin with a description of the growth curve, shown in figure 1 in its simplest form {33i, 538). In food spoilage only the lag and logarithmic phases are important, because foods are spoiled by the time bacterial growth has begun to decelerate to the resting phase.
Lag phase, as here used, is defined as the time elapsed from inoculation to the beginning of logarithmic multiplication. At low temperatures, there is sometimes a drop in count before logarith- mic growth begins (326, 3^8, Í09) because some components of the population die before others begin logarithmic growth.
During the logarithmic growth phase, micro-organisms grow at their most rapid rate. A convenient measure of this rate is the "generation time," that is, the time for one cell to become two, calculated as follows (387) :
¿log 2 log &—log B
where ^régénération time, inobservation period, ß^number of bacteria at the beginning of the observation period, and ö—num- ber at the end of the period.
Most yeasts reproduce by budding, which means that the time for one cell to become two is not strictly a generation time. Nevertheless, this term has been used by several investigators as a convenient measure for the rate of yeast growth.
10
8 -
tn 6
^ A < 4
o 2 o
Logarithmic growth phase
.Lag phase
TIME
FIGURE 1.—A typical bacterial growth curve.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 3
When growth rates of molds or actinomycetes are to be meas- ured, generation time cannot be used because the colony grows by terminal extension and branching of the hyphae rather than by binary fission of individuals. Haines {179, 182) prepared growth curves by plotting the logarithms of the total lengths of hyphae against time. As a measure of growth rate he used the slope of the steepest part of the resulting curve.
Instead of growth rate, some investigators use rates of related microbial activities such as oxygen consumption or carbon dioxide production.
As the temperature changes, growth rates and activity rates change. As a convenient means of measuring the effect of tem- perature on these rates, three terms are often used : temperature coefficient (Qio), temperature characteristic (/A), and Belehrádek's exponent b.
Temperature coefficient (Qio) can be defined as a ratio of growth rate or activity rate at one temperature to that at a temperature 10° C. lower, or:
where fci and fcg are velocity coefficients of growth, activity rates, or generation times at temperatures differing by lO"" (55). Qio can be calculated for any other temperature interval (AT) from the following formula (S87) :
AT
Temperature characteristic (ju.) is a related figure and is de- rived from the van't Hoff-Arrhenius equation:
4.6 (log fc2-log fci) T2T,
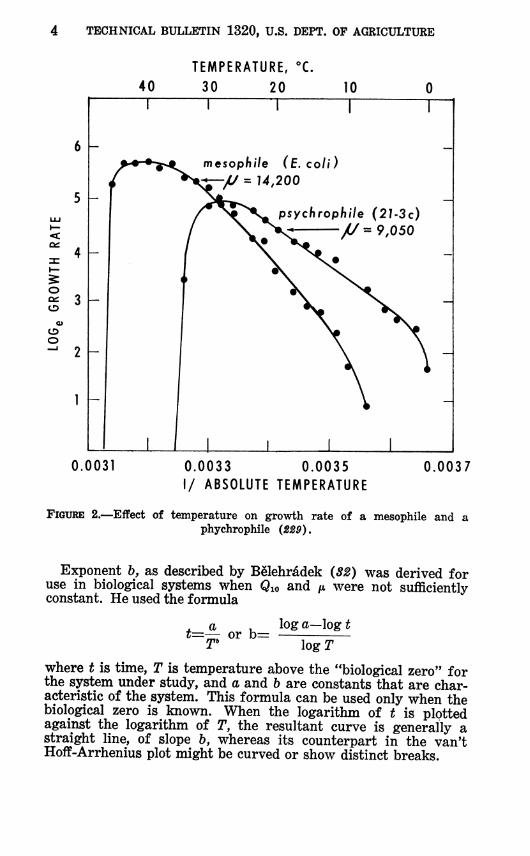
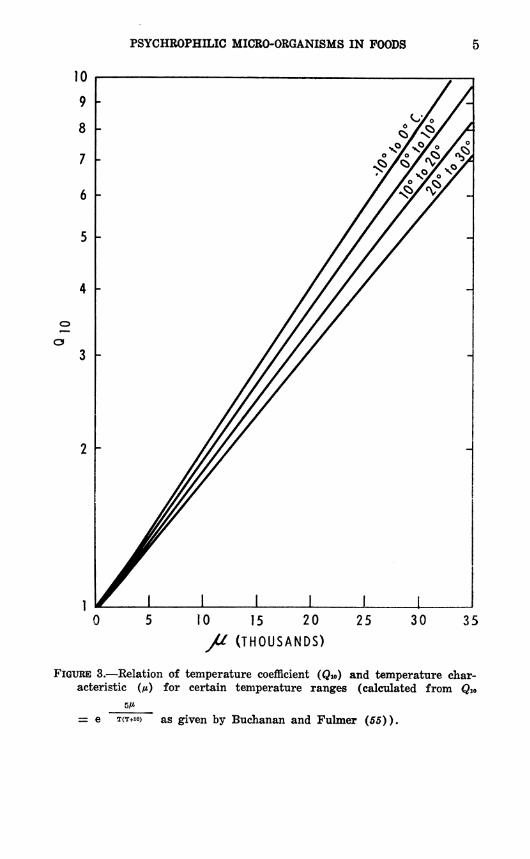
where fei and fe are growth or activity coefficients at absolute temperatures Ti and T2, respectively (55, 387). The temperature characteristic is the energy of activation of the reaction in calories per gram molecule and is represented by the slope of the curve obtained when the logarithm of a growth or activity rate is plotted against the reciprocal of the absolute temperature, as shown in figure 2. As will be discussed later, /x is usually constant with respect to temperature over a considerable temperature range. Qio and /* are related as shown in figure 3, so that if either is known, the other can be found provided the temperature range is given. Figure 3 also demonstrates that if /* is constant, Qio will change slightly with temperature in accordance with theory.
4 TECHNICAL BULLETIN 1320, U.S. DEFT. OP AGRICULTURE
O a: 3 CO
O O
2 -
1 -
40 "T"
0.0031
TEMPERATURE, °C. 30 20 ]0 T
mesophile (E.COI'I)
-// = 14,200
ychrophile (2i-3c) —//= 9,050
0.0033 0.0035 1/ ABSOLUTE TEMPERATURE
0.0037
FIGURE 2.—Effect of temperature on growth rate of a mesophile and a phychrophile (2ê9).
Exponent b, as described by Bëlehradek (32) was derived for use in biological systems when Qi« and ¡j. were not sufficiently constant. He used the formula
i=—- or b: rpb
log a—log t logT
where t is time, T is temperature above the "biological zero" for the system under study, and a and & are constants that are char- acteristic of the system. This formula can be used only when the biological zero is known. When the logarithm of t is plotted against the logarithm of T, the resultant curve is generally a straight line, of slope b, whereas its counterpart in the van't Hoff-Arrhenius plot might be curved or show distinct breaks.
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS
10 15 20 25
JU (THOUSANDS)
35
FIGURE 3.—Relation of temperature coefficient (Qio) and temperature char- acteristic (/*) for certain temperature ranges (calculated from Qio
5M
= e T(T+io) as given by Buchanan and Fulmer (55)).

TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
EFFECT OF TEMPERATURE ON MICROBIAL ACTIVITIES
Lag phase Numerous investigators have found the lag period of microbial
growth to be highly sensitive to temperature; lag is shortest at the optimum temperature {891 ) and is prolonged as temperature is lowered. In the range near the minimum growth temperature, lag increases so rapidly with decreasing temperature that it ap- proaches infinity. Hanson and Fletcher (200) reported consid- erable mold growth on turkey pies at —6.7° C. after 9 months but none after 6 months. Gibbons (152) found a lag period of 50 weeks at —5'' for the natural bacterial ñora on fish. Chistyakov and Bocharova {7i) found a strain of Oospora with a lag period of 414 days at —8°, the longest lag period we have found on record. It is possible that other investigators would have recorded similarly long lag periods with other organisms, had they con- tinued observations for prolonged periods.
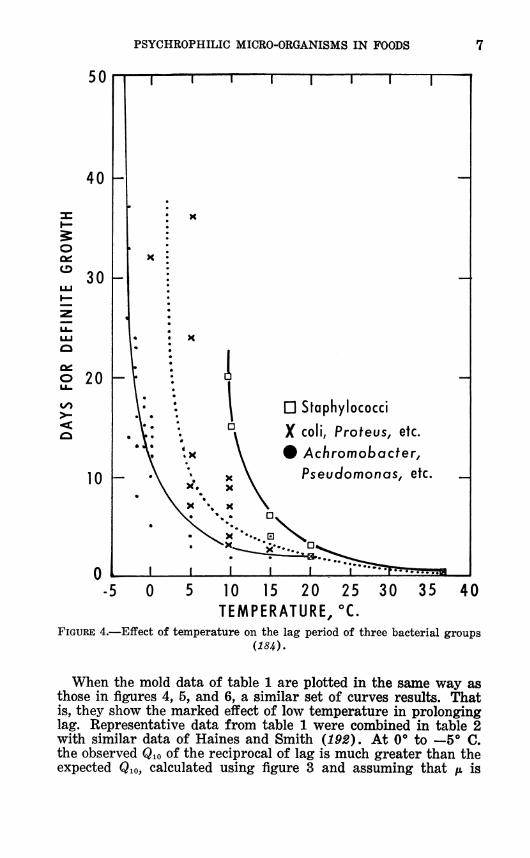
In the temperature range where both psychrophiles and me- sophiles are able to grow (generally between 10° and 30° C), the psychrophiles have a much shorter lag period (fig. 4 and 5). However, lag time varies widely among both the psychrophilic bacteria (fig. 6) and the psychrophilic molds (table 1).
TABLE 1.—Lag periods of various species and strains of molds at various temperatures
Lag period at—
Species -8" C.
-5° C.
-2^ C.
0° c.
+2° c.
+5° C.
+ 10" c.
Room temper- ature
Aspergillus glaucus Mucor recemosus Phycomyces nitens Fusarium sp. I Pénicillium sp Fusarium culmorum Monilia nigra Pénicillium glaucum Mucor sp Fusarium sp. Ill Fusarium sp. IV Fusarium sp. II Pénicillium sp. FusaHum sp. V Botrytis cinérea Oospora sp Cladosporium herbarum. Chaetostylum fresenii— Cladosporium sp. Monilia nigra
Days
C) C) o C) C) C) C) C)
C) C) C) C)
I
Days
8 C) C)
177 138 131 142
42 38 38 38 29 28 42 20 18 13 10
2
Days
n C) 55 18 46 13 49 21 22 12
4 5
20 12 17 13 18 12
8 2
Days 159
17 22
4 52
8 24 15
Days 72
6 14
3 46
6 8 9
Days
4"
Days
5 4 4
11 10 11 11 11 12
8
5 5 3 9 3 5
11 6 5 6
2 1 2 2 6 1 1 2 1 2 2 2 1 2 1 2 1 1 1 1
*No growth observed. SOURCE: Chistyakov, F. M., and Bocharova, Z. Z. (7S),
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS
o
O
i>0
< D Staphylococci
X coli, Profeus, etc. # Ac/iromobacfer,
Pseuc/omonas, etc.
-5 0 10 15 20 25 TEMPERATURE, "C.
30 35 40
FIGURE 4.—Effect of temperature on the lag period of three bacterial groups
When the mold data of table 1 are plotted in the same way as those in figures 4, 5, and 6, a similar set of curves results. That is, they show the marked effect of low temperature in prolonging lag. Representative data from table 1 were combined in table 2 with similar data of Haines and Smith (192). At 0° to —5** C. the observed Qio of the reciprocal of lag is much greater than the expected Qio, calculated using figure 3 and assuming that fi is
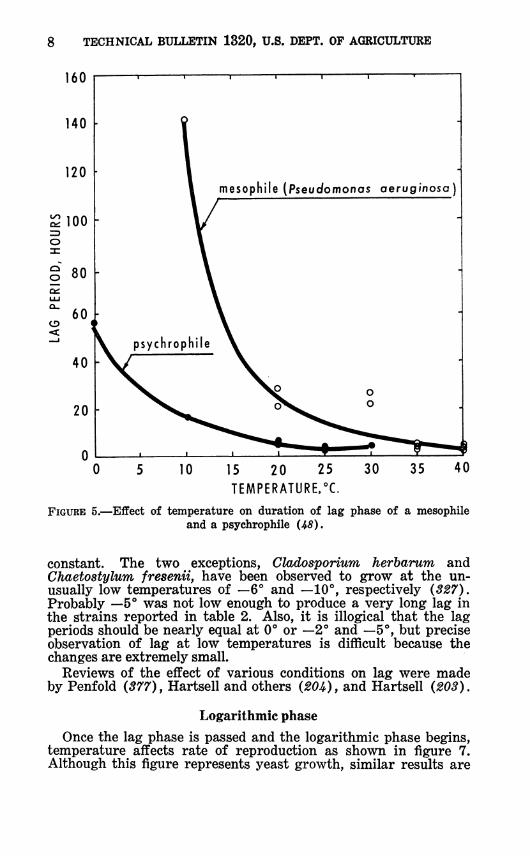
8 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
160
mesophile (Pseudomonas aeruginosa ]
10 15 20 25 30 TEMPERATURE/C.
FIGURE 5.—Effect of temperature on duration of lag phase of a mesophile and a psychrophile (-45).
constant. The two exceptions, Cladosporium herbarum and Chaetostylum fresenii, have been observed to grow at the un- usually low temperatures of —6° and —10°, respectively (327). Probably —5° was not low enough to produce a very long lag in the strains reported in table 2. Also, it is illogical that the lag periods should be nearly equal at 0° or —2° and —5°, but precise observation of lag at low temperatures is difficult because the changes are extremely small.
Reviews of the effect of various conditions on lag were made by Penfold (377), Hartsell and others (20i), and Hartsell (203).
Logarithmic phase Once the lag phase is passed and the logarithmic phase begins,
temperature affects rate of reproduction as shown in figure 7. Although this figure represents yeast growth, similar results are
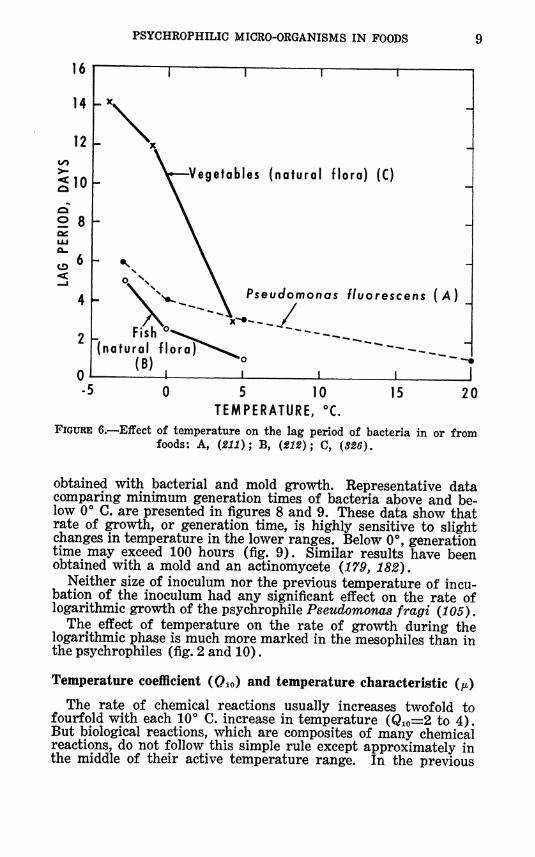
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS
16
14
12
2 8
CD 6
2 -,
1 \ I r
-Vegetables (natural flora) (C)
X^^^N^ \ Pseudomonas fluorescem
Fish <=> (natural floraf
(B) J_
•5 0 5 10 15 20 TEMPERATURE, "C.
FIGURE 6.—Effect of temperature on the lag period of bacteria in or from foods: A, (211); B, (212); C, (326).
obtained with bacterial and mold growth. Representative data comparing minimum generation times of bacteria above and be- low 0° C. are presented in figures 8 and 9. These data show that rate of growth, or generation time, is highly sensitive to slight changes in temperature in the lower ranges. Below O*', generation time may exceed 100 hours (fig. 9). Similar results have been obtained with a mold and an actinomycete (179, 182).
Neither size of inoculum nor the previous temperature of incu- bation of the inoculum had any significant effect on the rate of logarithmic growth of the psychrophile Pseudomonas fragi (105).
The effect of temperature on the rate of growth during the logarithmic phase is much more marked in the mesophiles than in the psychrophiles (fig. 2 and 10).
Temperature coefficient (Qio) and temperature characteristic (fi)
The rate of chemical reactions usually increases twofold to fourfold with each 10° C. increase in temperature (Qio=2 to 4). But biological reactions, which are composites of many chemical reactions, do not follow this simple rule except approximately in the middle of their active temperature range. In the previous
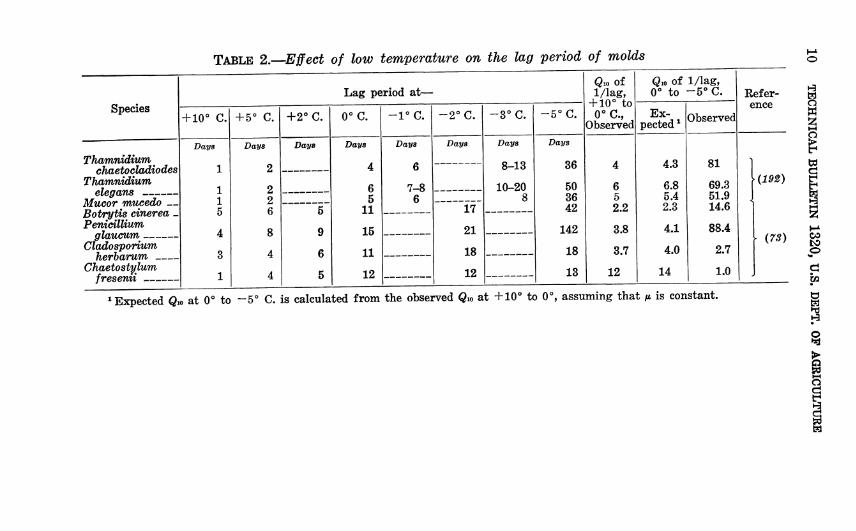
TABLE 2.—Effect of low temperature on the lag period of molds
Lag period at— Qio of 1/lag,
+ 10° to 0°C.,
Observed
Qio of 1/lag, 0° to -5° C. Refer-
Species + 10° C. + 5° C. +2°C. 0°C. -1°C. -2°C. -3°C. -5°C. Ex-
pected ^ Observed ence
Thamnidium chaetocladiodes
Thamnidium elegans
Mucor mucedo — Botrytis cinérea - Pénicillium
glaucum Cladosporium
herbarum Chaetostylum
fresenii
Days
1
1 1 5
4
3
1
Days
2
2 2 6
8
4
4
Days
--
9
6
5
Days
4
6 5
11
15
11
12
Days
6
7-8 6
Days
17'
21
18
12
Days
8-13
10-20 8
Days
36
50 36 42
142
18
13
4
6 5 2.2
3.8
3.7
12
4.3
6.8 5.4 2.3
4.1
4.0
14
81
69.3 51.9 14.6
88.4
2.7
1.0
■(192)
{7S)
»Expected Q» at 0° to -5° C. is calculated from the observed Q.o at +10° to 0°, assuming that /i is constant.
O
s:
to
I—I
CO
O
TO
»Tí
k I d
»
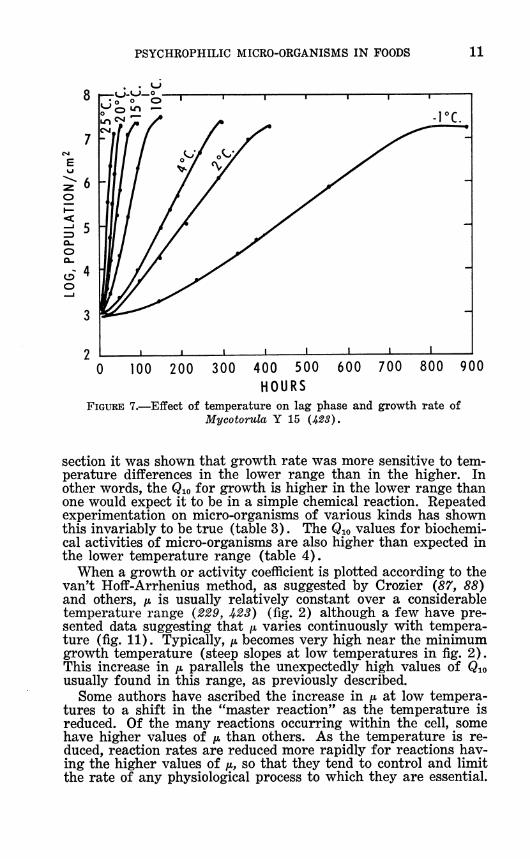
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 11
0 100 200 300 400 500 600 700 800 900 HOURS
FIGURE 7.—Effect of temperature on lag phase and growth rate of Mycotorula Y 15 U23),
section it was shown that growth rate was more sensitive to tem- perature differences in the lower range than in the higher. In other words, the Qw for growth is higher in the lower range than one would expect it to be in a simple chemical reaction. Repeated experimentation on micro-organisms of various kinds has shown this invariably to be true (table 3). The Qio values for biochemi- cal activities of micro-organisms are also higher than expected in the lower temperature range (table 4).
When a growth or activity coefficient is plotted according to the van't Hoff-Arrhenius method, as suggested by Crozier (87, 88) and others, [i is usually relatively constant over a considerable temperature range (229, Í23) (fig. 2) although a few have pre- sented data suggesting that /A varies continuously with tempera- ture (fig. 11). Typically, /* becomes very high near the minimum growth temperature (steep slopes at low temperatures in fig. 2). This increase in p. parallels the unexpectedly high values of Qio usually found in this range, as previously described.
Some authors have ascribed the increase in ¡i at low tempera- tures to a shift in the "master reaction" as the temperature is reduced. Of the many reactions occurring within the cell, some have higher values of [i than others. As the temperature is re- duced, reaction rates are reduced more rapidly for reactions hav- ing the higher values of /x, so that they tend to control and limit the rate of any physiological process to which they are essential.
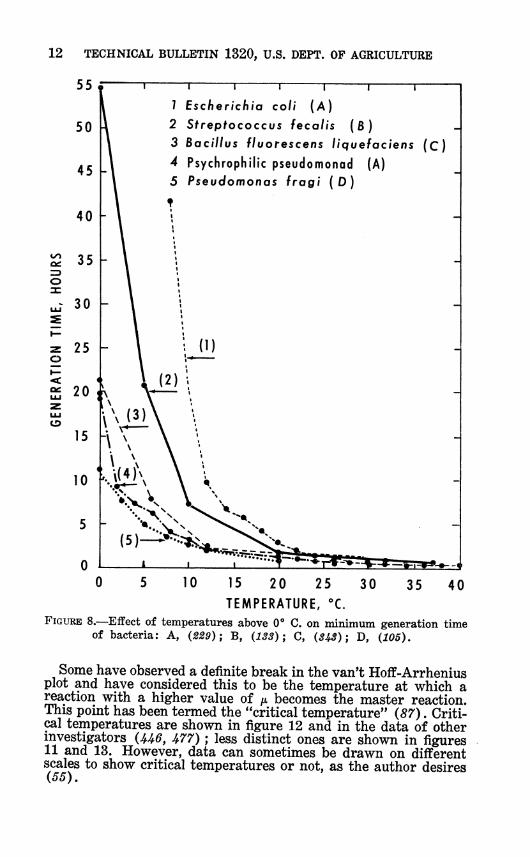
12 TECHNICAL BULLETIN 1320, U.S. DEPT. OP AGRICULTURE
1 1 1 1
1 Escherichia co/i (A)
2 Streptococcus fecalis (ß)
3 Bacillus fluorescens liquefaciens (c)
4 Psychrophilic pseudomonad (A) 5 Pseudomonas fragt (D)
0 5 10 15 20 25 30 35 40
TEMPERATURE, "C.
FIGURE 8.—Effect of temperatures above 0*» C. on minimum generation time of bacteria: A, (229); B, (133); C, (343); D, (105).
Some have observed a definite break in the van't Hoff-Arrhenius plot and have considered this to be the temperature at which a reaction with a higher value of p. becomes the master reaction. This point has been termed the "critical temperature" (87). Criti- cal temperatures are shown in figure 12 and in the data of other investigators (U6, U77) ; less distinct ones are shown in figures 11 and 13. However, data can sometimes be drawn on different scales to show critical temperatures or not, as the author desires (55).
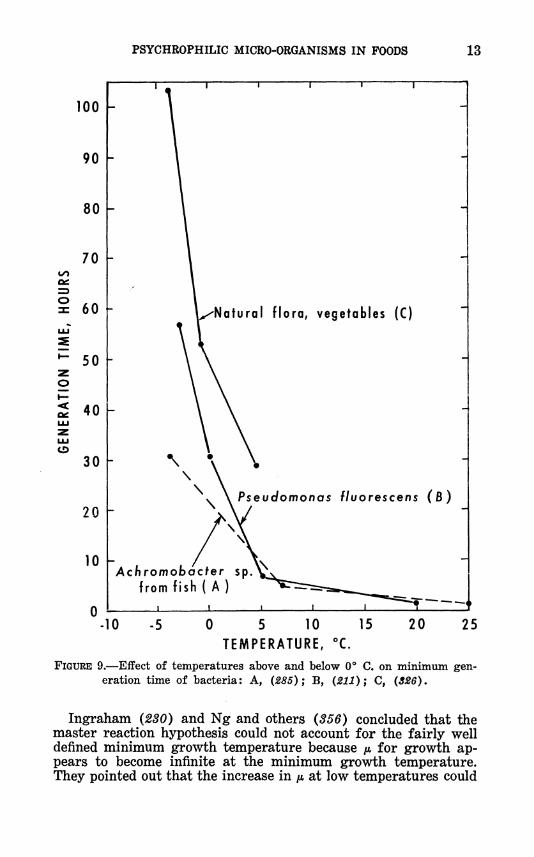
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 13
100
90
80
70
ai 60 - LU
S
- 50 z o
^ 40
CD 30
20
10
\^Natural flora, vegetables (C)
Pseudomonas fluorescens (ß)
Achromobacter sp from fish ( A )
■10 0 5 10 15 20 25
TEMPERATURE, "C.
FIGURE 9.—Effect of temperatures above and below 0** C. on minimum gen- eration time of bacteria: A, (285); B, (211); C, (326).
Ingraham (230) and Ng and others (356) concluded that the master reaction hypothesis could not account for the fairly well defined minimum growth temperature because /* for growth ap- pears to become infinite at the minimum growth temperature. They pointed out that the increase in /* at low temperatures could
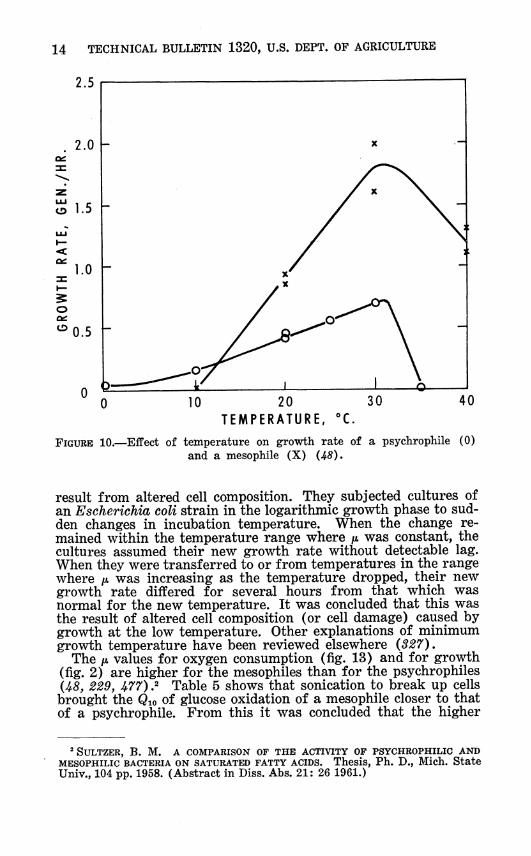
14 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
2.5
10 20 TEMPERATURE,
30 40
FIGURE 10.—EflPect of temperature on growth rate of a psychrophile (0) and a mesophile (X) (45).
result from altered cell composition. They subjected cultures of an Escherichia coli strain in the logarithmic growth phase to sud- den changes in incubation temperature. When the change re- mained within the temperature range where fi was constant, the cultures assumed their new growth rate without detectable lag. When they were transferred to or from temperatures in the range where ju. was increasing as the temperature dropped, their new growth rate differed for several hours from that which was normal for the new temperature. It was concluded that this was the result of altered cell composition (or cell damage) caused by growth at the low temperature. Other explanations of minimum growth temperature have been reviewed elsewhere (327).
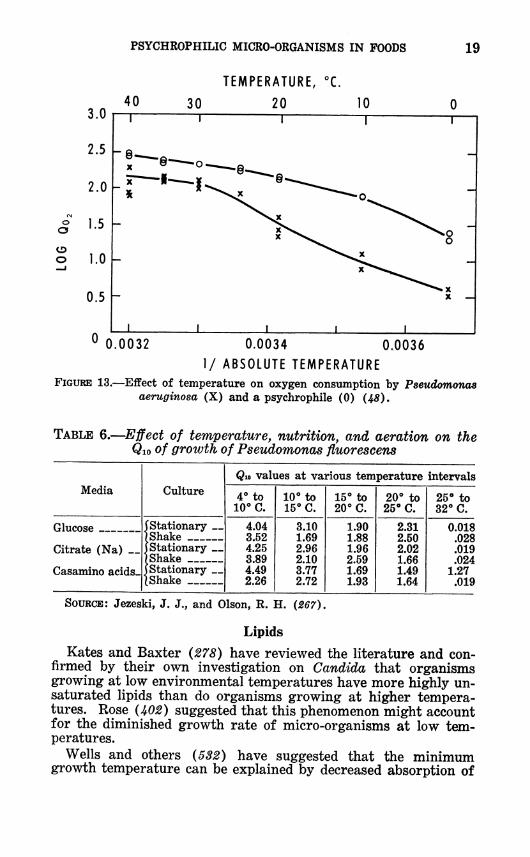
The ft values for oxygen consumption (fig. 13) and for growth (fig. 2) are higher for the mesophiles than for the psychrophiles U8, 229, J¡>77) .2 Table 5 shows that sonication to break up cells brought the Qio of glucose oxidation of a mesophile closer to that of a psychrophile. From this it was concluded that the higher
^ SULTZER, B. M. A COMPARISON OF THE ACTIVITY OF PSYCHROPHILIC AND MESOPHILIC BACTERIA ON SATURATED FATTY ACIDS. Thesis, Ph. D., Mich. State Univ., 104 pp. 1958. (Abstract in Diss. Abs. 21: 26 1961.)
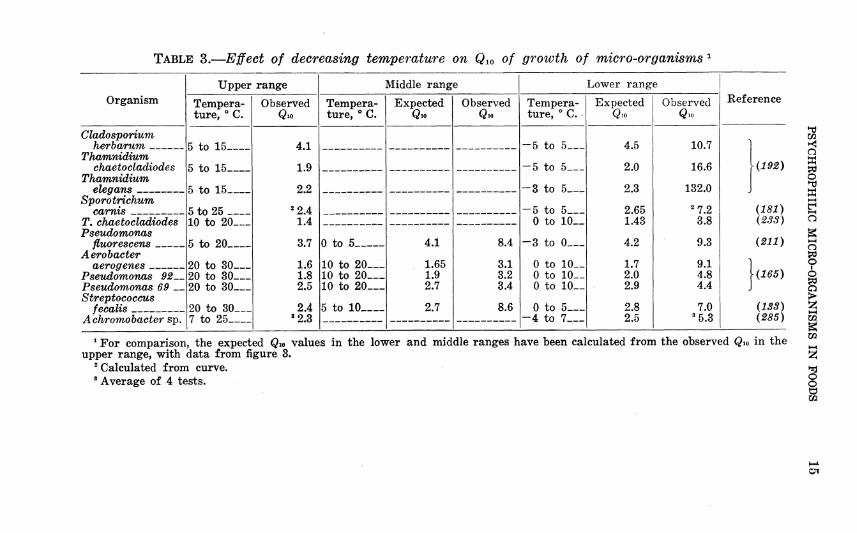
TABLE 3.—Effect of decreasing temperature on Qio of growth of micro-organisms
Upper range Middle range Lower range Organism Tempera- Observed Tempera- Expected Observed Tempera- Expected Observed Reference
ture, ° C. Q.0 ture, ° C. Qio Qio ture, ° C. Qio Qio
Cladosporium herbarum 5 to 15 4.1 -5 to 5 4.5 10.7
Thamnidium chaetocladiodes 5 to 15 1.9 -5 to 5 2.0 16.6 H^92)
Thamnidium elegans
Sporotrichum 5 to 15 2.2 -3 to 5 2.3 132.0
camts 5 to 25 10 to 20
^2.4 1.4
-5 to 5 0 to 10-_
2.65 1.43
^7.2 3.8
(181) T. chaetocladiodes {23S) Psevdomonas
fluorescens 5 to 20 3.7 0 to 5 4.1 8.4 -3 to 0 4.2 9.3 {211) Aerobacter
aerogenes 20 to 30 1.6 10 to 20— 1.65 3.1 0 to 10__ 1.7 9.1 L. X Pseudomonas 92-^ 20 to 30 1.8 10 to 20 1.9 3.2 0 to 10__ 2.0 4.8 \{165) Pseudomonas 69 __ 20 to 30 2.5 10 to 20 2.7 3.4 0 to 10__ 2.9 4.4 J Streptococcus
fecalis « _ 20 to 30 7 to 25
2.4 »2.3
5 to 10 2.7 8.6 0 to 5 -4 to 7
2.8 2.5
7.0 ^5.3
(133) Achromobacter sp. (285)
^ For comparison, the expected Qw values in the lower and middle ranges have been calculated from the observed Qw in the upper range, with data from figure 3.
^ Calculated from curve. * Average of 4 tests.
>< O tu w o
o
o to o É
t—I
Z/2
U2
O O ü U2
z^
16 TECHNICAL BULLETIN 1320, U.S. DEPT. OP AGRICULTURE
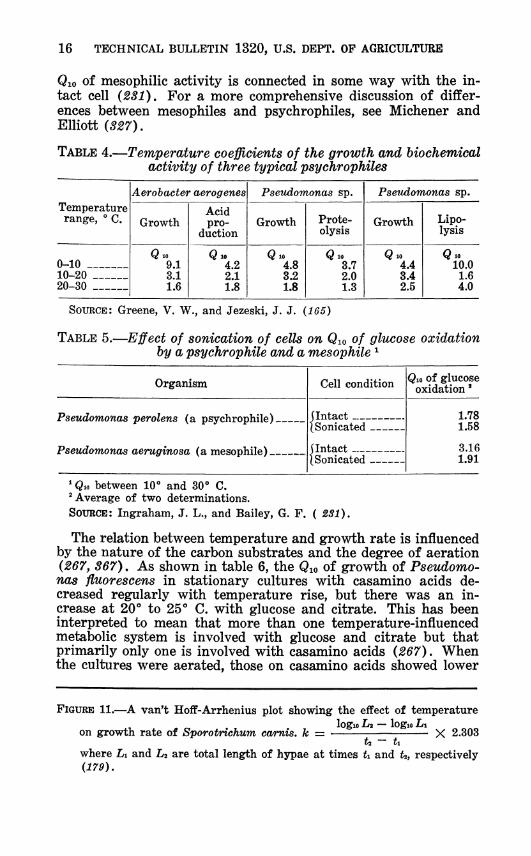
Qio of mesophilic activity is connected in some way with the in- tact cell (231). For a more comprehensive discussion of differ- ences between mesophiles and psychrophiles, see Michener and Elliott (827).
TABLE 4.—Temperature coefficients of the growth and biochemical activity of three typical psychrophiles
Aerobacter aerogenes Pseudomonas sp. Pseudomonas sp. Temperature range, ° C. Growth
Acid pro-
duction Growth Prote-
olysis Growth Lipo-
lysis
0-10 10-20 20-30
Qio 9.1 3.1 1.6
Qio 4.2 2.1 1.8
Qio 4.8 3.2 1.8
Qio 3.7 2.0 1.3
Qio 4.4 3.4 2.5
Qio 10.0
1.6 4.0
SOURCE: Greene, V. W., and Jezeski, J. J. (165)
TABLE 5.—Effect of sonication of cells on Q^ of glucose oxidation by a psychrophile and a mesophile ^
Organism Cell condition Qio of glucose oxidation '
Pseudomonas perolens (a psychrophile)
Pseudomonas aeruginosa (a mesophile)
(Intact (Sonicated
(Intact (Sonicated
1.78 1.58
3.16 1.91
' Qio between 10° and 30° C. ^Average of two determinations. SOURCE: Ingraham, J. L., and Bailey, G. F. ( 231).
The relation between temperature and growth rate is influenced by the nature of the carbon substrates and the degree of aeration (267, 867). As shown in table 6, the Qio of growth of Pseudomo- nas fiuorescens in stationary cultures with casamino acids de- creased regularly with temperature rise, but there was an in- crease at 20° to 25° C. with glucose and citrate. This has been interpreted to mean that more than one temperature-influenced metabolic system is involved with glucose and citrate but that primarily only one is involved with casamino acids (267). When the cultures were aerated, those on casamino acids showed lower
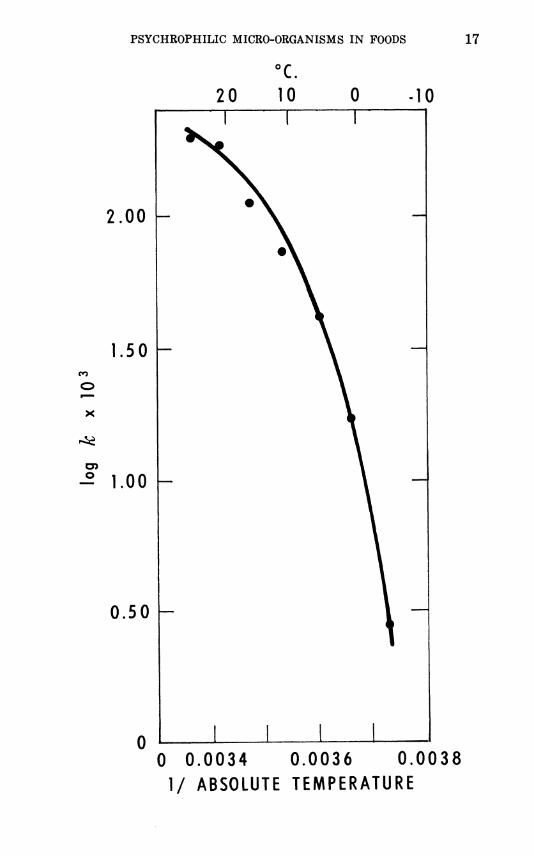
FIGURE 11.—A van't Hoif-Arrhenius plot showing the effect of temperature
on growth rate of Sporotrichum camis. k = logio La — logio Li
X 2.303 U - ti
where Li and L2 are total length of hypae at times U and Uy respectively (179).
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS
20 10
17
0 0.0034 0.0036 0.0038 1/ ABSOLUTE TEMPERATURE
18 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
20 10 1.5 K 1
— *\ -
1.1 \
- //=I6,59o\
-
2o.7 X
LOG
LAM
BE
RTS
o
C
O
-0.1 //=22,44o\
\.
-0.5 1 I 1 J 1 L 0.0033 0.0034 0.0035 0.0036
1/ABSOLUTE TEMPERATURE
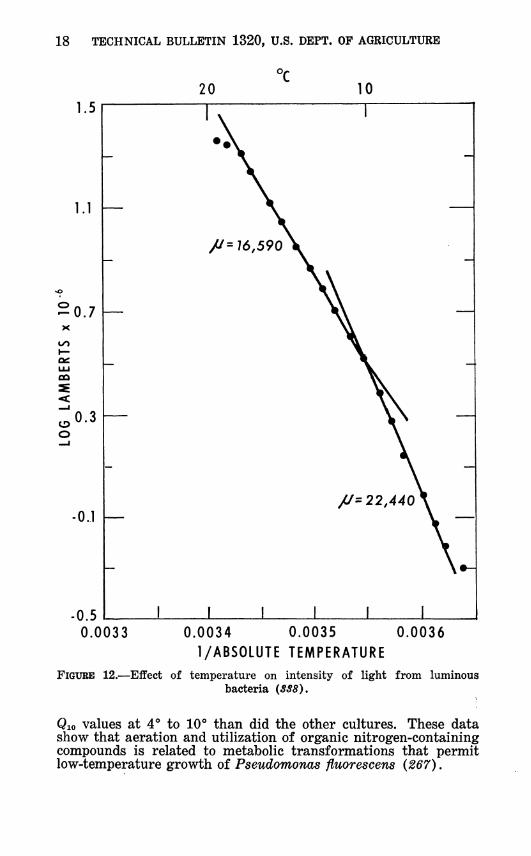
FIGURE 12.—Effect of temperature on intensity of light from luminous bacteria (SS8).
Qio values at 4° to 10"* than did the other cultures. These data show that aeration and utilization of organic nitrogen-containing compounds is related to metabolic transformations that permit low-temperature growth of Pseudomonas fluorescens (267).
PSYCHROPHILIC MICRO-ORGANISMS IN POODS
TEMPERATURE, °C.
19
O O
O o
o 0.0032 0.0034 0.0036
1/ ABSOLUTE TEMPERATURE FIGURE 13.—Effect of temperature on oxygen consumption by Pseudomonas
aeruginosa (X) and a psychrophile (0) (48).
TABLE 6.—Effect of temperature, nutrition, and aeration on the Qio of growth of Pseudomonas fluorescens
Culture Qio values at various temperature intervals
Media 4^ to 10** to 15*» to 20*» to 25*» to 10° C. 15** C. 20*^0. 25*» C. 32*» C.
Glucose (Stationary 4.04 3.10 1.90 2.31 0.018 /Shake » «- 3.52 1.69 1.88 2.50 .028
Citrate (Na) -_ (stationary 4.25 2.96 1.96 2.02 .019 /Shake 3.89 2.10 2.59 1.66 .024
Casamino acids- (Stationary 4.49 3.77 1.69 1.49 1.27 (Shake 2.26 2.72 1.93 1.64 .019
SOURCE: Jezeski, J. J., and Olson, R. H. (267).
Lipids
Kates and Baxter (278) have reviewed the literature and con- firmed by their own investigation on Candida that organisms growing at low environmental temperatures have more highly un- saturated lipids than do organisms growing at higher tempera- tures. Rose (Í02) suggested that this phenomenon might account for the diminished growth rate of micro-organisms at low tem- peratures.
Wells and others (532) have suggested that the minimum grovrth temperature can be explained by decreased absorption of

20 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
nutrients caused by an increase in the amount of fat in the mem- branes. They described one strain of Brevibacterium linens that produced 7.2 percent of fat at 25""C, where it grew well; but it produced 16.7 percent at 4"*, where it grew poorly. Two typical psychrophilic bacteria displayed no such temperature-induced differences. Eklund ^ confirmed that bacteria that grew poorly at 4° produced more fat there than at 9.4° or at 22°. However, one can do no more than speculate that higher fat production or greater unsaturation may explain minimum growth temperature, for much more convincing data are needed.
Enzyme production and activity
Enzymes reduce the energy of activation required to cause chemical reactions under physiological conditions at rates useful to the cell. Although some strains of bacteria cannot ferment carbohydrates near their minimum growth temperatures (20) ^ more commonly micro-organisms produce a greater quantity of enzyme at a low than at a high temperature (table 7 and fig. 14). On the other hand, the activities of the isolated enzymes them- selves often have high temperature maxima—indeed, even above the optimum growth temperature of the organism (fig. 15). The data in figures 14 and 15 are combined in table 8 with those of other workers to show that this relation is commonly found. Precht and others (388) have summarized similar results of German investigations on Streptococcus lactis and a parasitic mold. A psychrophilic bacterium failed to produce formic hydrogenlyase at 35 °C. but produced this enzyme at temperatures between 0° and 20° (511).
TABLE 7.—Catatase activity in psychrophilic bacteria harvested at various ages during incubation at 2"" and at SO"" C.
Incubation temperature,
° C. Harvest
age Specific catalase activity (XlO*)*
Pseudomonas strain 92 Pseudomonas strain 95
30 - _
Days
1 2 3 7
14 5 7
11 14 20
256 198 153
1 1
877 536 382 433 329
427 274 198
2_
35 26
1,137 627 652 595 580
^ Catalatic reaction rate constant at 10° C. per minute per 10* cells. SOURCE: Frank, H. A., Ishabishi, S. T., Reid, A., and Ito, J. S. {134').
'EKLUND, M. W. BIOSYNTHETIC RESPONSES OF POULTRY MEAT ORGANISMS UNDER STRESS. Thesis. Ph. D., Purdue Univ., 128 pp. 1962. (Abstract in Diss. Abs. 23: 2660-1 1963.)
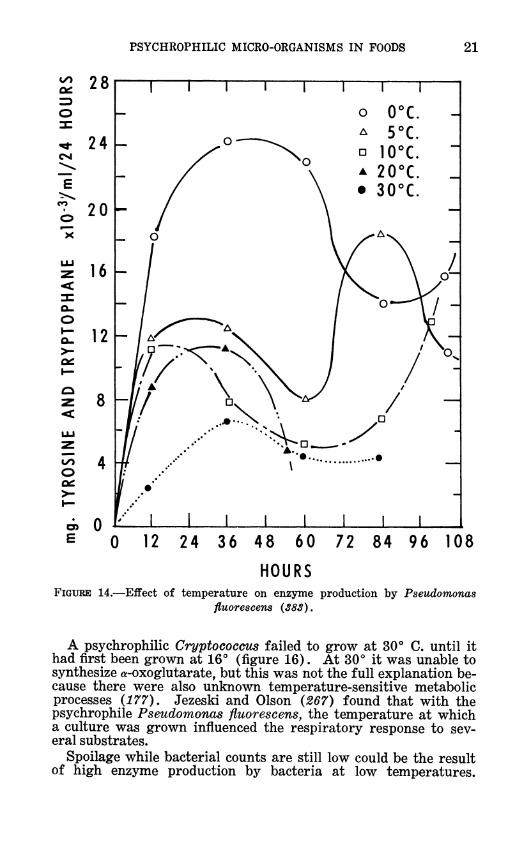
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 21
o, 0 1^ \ \ L S 0 12 24 36 48 60 72 84 96 108
HOURS FIGURE 14.—Effect of temperature on enzyme production by Pseudomonas
fluorescens (383),
A psychrophilic Cryptococcus failed to grow at 30° C. until it had first been grown at 16° (figure 16). At 30° it was unable to synthesize a-oxoglutarate, but this was not the full explanation be- cause there were also unknown temperature-sensitive metabolic processes {177). Jezeski and Olson {267) found that with the psychrophile Pseudomonas fluorescens, the temperature at which a culture was grown influenced the respiratory response to sev- eral substrates.
Spoilage while bacterial counts are still low could be the result of high enzyme production by bacteria at low temperatures.
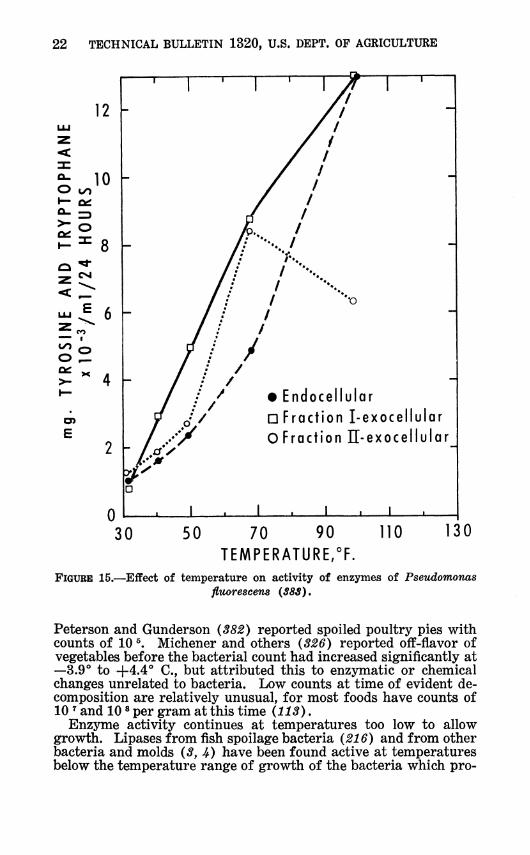
22 TECHNICAL BULLETIN 1320, U.S. DEPT. OP AGRICULTURE
12
<
O oo
>- o
10
8
CSI
S2 u¿ X
E
• Endocelluiar
D Fraction I-exocellular
OFraction IT-exocellular
_L
30 50 70 90 110
TEMPERATURE,°F.
30
FIGURE 15.—Effect of temperature on activity of enzymes of Pseudomonas fluorescens {383).
Peterson and Gunderson {382) reported spoiled poultry pies with counts of 10 ^. Michener and others {826) reported off-flavor of vegetables before the bacterial count had increased significantly at —3.9° to +4.4° C, but attributed this to enzymatic or chemical changes unrelated to bacteria. Low counts at time of evident de- composition are relatively unusual, for most foods have counts of 10 ^ and 10 * per gram at this time (113).
Enzyme activity continues at temperatures too low to allow growth. Lipases from fish spoilage bacteria {216) and from other bacteria and molds (3, Jf) have been found active at temperatures below the temperature range of growth of the bacteria which pro-
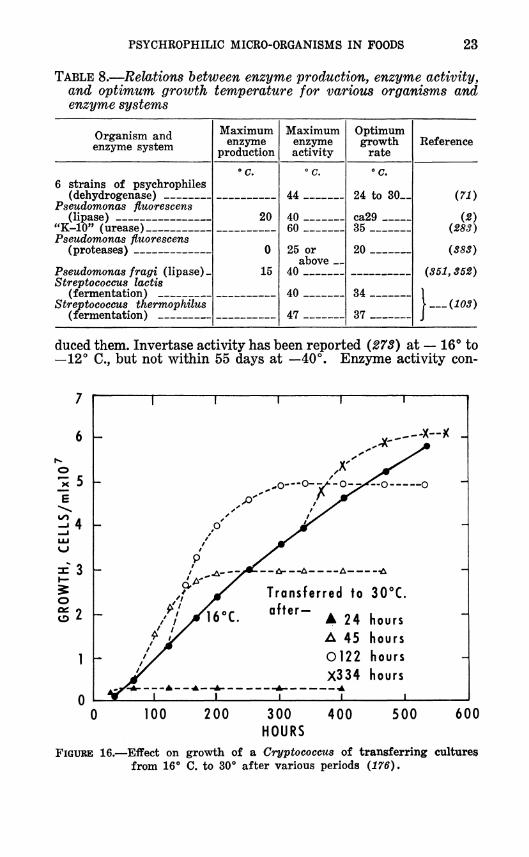
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 23
TABLE 8.—Relations between enzyme production, enzyme activity, and optimum growth temperature for various organisms and enzyme systems
Organism and enzyme system
Maximum enzyme
production
Maximum enzyme activity
Optimum growth
rate Reference
'C. 6 strains of psychrophiles
(dehydrogenase) Pseudomonas fluorescens
(lipase) "K-10" (urease) Pseudomonas fluorescens
(proteases)
20
44
40 60
24 to 30_
ca29 35--
(71)
Pseudomonas fragi (lipase)- Streptococcus lactis
(fermentation) ___. Streptococcus thermophilus
(fermentation)
0
15
25 or above .
40
20
40 ,
47,
34,
37
(383)
{351, 352)
(103)
duced them. Invertase activity has been reported (273) at — 16"* to —12° C, but not within 55 days at —40°. Enzyme activity con-
7
6
o
E CO
4 -
3 -
o CD ^
-
1 1 1 1 1
-X-X -
^'°' .^''
-0
-
/ X / >^
-
— / ..—ytC—^. .-A A -A -
,f' y Transferred to 30°C.
- /'' ^^»f after- ^ ^. , / / ^ 10 t. A 24 hours -
' / A 45 hours
- // 0122 hours - J^ X334 hours
^ •f-*^--.-^--*'--^ |- ^ i i
100 200 300 400 HOURS
500 600
FiGXJRE 16.—Effect on growth of a Cryptococcus of transferring cultures from 16° C. to 30° after various periods (176).

24 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
tinues in soils after bacterial growth has been inhibited by cold {5U).
Activity rates of enzymes at low temperatures above freezing can be predicted by extrapolation from their activity rates at higher temperatures {SO5). The activities of the enzymes with low /A become increasingly important as temperature is lowered. However, when the substrate freezes, the temperature character- istics of enzyme activities usually change abruptly, and thus it is impossible to predict enzymatic reaction rates below 0"* C. from studies made above that temperature (305, JiUe). A high /x in the frozen material shows the efficacy of lowering freezer tempera- tures to slow enzymatic action {JfU6).
Many investigations have shown that enz3mies are little injured by freezing. Indeed some (23, 95, 96, 280) have claimed they were more active for having been frozen and thawed. Young (55^) used 18 cycles of freezing and thawing to prepare cell- free extracts of Escherichia coli for enzyme studies, Luyet and Gehenio (311) reviewed the early literature, which showed that many enzymes can withstand freezing and storage at very low temperatures.
In practical storage tests, various foods have been adversely affected by microbial enzymes at temperatures below the growth range (table 9). On frozen vegetables, however, quality was un- related to the bacterial count (223), It is common knowledge that naturally occurring plant enzymes must be inactivated by blanching before vegetables can be stored in the freezer. Al- though relatively few studies have been made of this subject, it is clear that enzymes of either microbial or nonmicrobial origin may cause food deterioration at temperatures too low for microbial growth.
TABLE 9.—Effect of microbial enzymes on foods stored below the temperature range of growth
Food product Storage temperature
Result of high microbial level Reference
Bacon - -17.8 -10 -18
"Frozen" -12.2
-7 to -29
Rancidity (26 J^) (353) (317) (209) (382) (SA)
Butter - > Lipolysis _ Do _ Deleterious odor
Rancidity - Pork sausage - Chicken pies Starch degradation
Lipolysis Fats - - _ _
Cell size and cell crop
Bacterial cell size is larger at low temperatures than at high temperatures, but the converse is true with yeasts {213, 388). In addition, cell size changes during the various phases of growth. The largest cells occur in late lag phase and during the logarith- mic phase.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 25
Cell crop often has been described as largest below the tempera- ture where growth is most rapid. Hess (211) found total crops of marine bacteria at —3° and 0° C. to be larger than those at 20 ° or 37°, and he recommended maximum cell crop as a criterion of optimum growth temperature. Dorn and Rahn (103) reported that the streptococci do not always produce their largest number of cells at the temperature of most rapid growth, as follows :
Fastest Largest rate of number growth of cells
at— at—
25-30 37
Streptococcus lactis 34 Streptococcus thermophilus 37
Higher cell crops at low temperatures were reported in two investigations on milk (fig. 17). However, data in figure 18 indi- cate the opposite effect. Also, in the genus Arthrobacter, the number of generations produced (equivalent to cell crop) is not greatly affected by incubation temperature within the tempera- ture range of growth of the organism (407).
Most of the foregoing authors failed to consider the oxygen re- lations of their cultures. Sinclair and Stokes (U5) showed that higher cell crops at low temperatures can be explained by the greater solubility of oxygen. With continuous aeration of cultures at both high and low temperatures the difference is erased (table 10). Upadhyay and Stokes (510) found that a facultatively anaerobic psychrophilic rod produced maximal cell crop aerobi- cally at 5° C. but anaerobically at 25°.
TABLE 10.—Effect of temperature and aeration on maximal cell yields of Pseudomonas aeruginosa^
Stationary Aerated Temperature,
° C. Maximal count per milliliter
Time to reach maximum
Maximal count per milliliter
Time to reach maximum
30 -_ . . 20 _>
Billions
6 11
Hours 25 75
Billions 17 19
Hours
25 53
^ In trypticase soy broth containing 1 percent glucose in 1.4 percent phosphate buffer.
SOURCE: Sinclair, N. A., and Stokes, J. L. (445).
Adaptation
Reports of adaptation of micro-organisms to low temperature growth are few. Hess (210, 2H) found that by cultivating bac- terial psychrophiles at 5** C. he was able to produce "adapted" strains that were more active at 0° and at —3° than were strains cultured at 20°. Also one strain of Pseudomonas fluorescens pro-
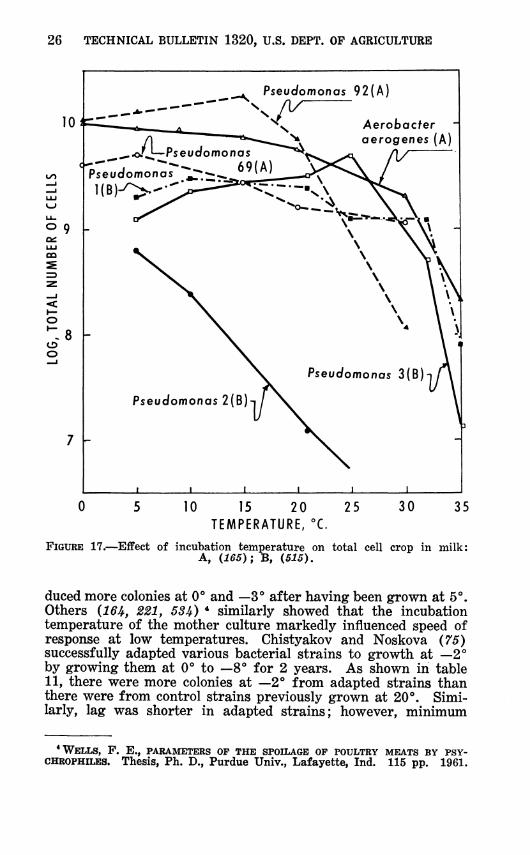
26 TECHNICAL BULLETIN 1320, U.S. DEPT. OP AGRICULTURE
10
O 9 0¿L
< H- O
o" O
Pseudomonas 92(A)
^^*,^Pseüc/omonas "^
Pseudomonas '^'^-^^ _ -
Aerobacfer aerogenes (A)
l(B)-^--
Pseudomonas 2(B)
10 15 20 25 TEMPERATURE, °C.
30 35
FIGURE 17.—Effect of incubation temperature on total cell crop in milk: A, {U5)\ B, (515).
duced more colonies at 0° and —3° after having been grown at 5°. Others {16U, 221, 53i) * similarly showed that the incubation temperature of the mother culture markedly influenced speed of response at low temperatures. Chistyakov and Noskova (75) successfully adapted various bacterial strains to growth at —2° by growing them at 0° to —8'' for 2 years. As shown in table 11, there were more colonies at —2° from adapted strains than there were from control strains previously grown at 20°. Simi- larly, lag was shorter in adapted strains; however, minimum
* WELLS, F. E., PARAMETERS OP THE SPOILAGE OP POULTRY MEATS BY PSY- CHROPHILES. Thesis, Ph. D., Purdue Univ., Lafayette, Ind. 115 pp. 1961.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 27
generation time was very little different (23 to 24 hours and 25 to 26 hours, respectively).
TABLE 11.—Effect of adaptation by growth at 0° to —8 years on growth of bacteria at — j^°
° C. for 2
Species and strain No. Colonies at -2** C
Controls Adapted
Achromobacter sp.: 1 _
Percent
80 59 91 78 84
55 83 97 82
89 85 86
Percent
85 2 _ 98 3 _ _ 132 5 __ 93 6 _ 350
Flavobacterium sp.: 3 108 4 132 7 193
Pseudomonas fluorescens 105 Pseudomonas herbicola:
13 109 14 105
Serratia rubefaciens 111
^ As percentage of those developing at 20° C. SOURCE: Chistyakov, F. M., and Noskova, G. (75).
They obtained similar results with several molds. Adapted strains of Pénicillium glaucum showed a more rapid growth at —5° to 2° C. (table 12). A relatively quick adaptation to rapid growth at 0° was shown by Monilia nigra (fig. 19).
Certainly in geologic time, mutants capable of growth in con- tinuous cold must occur, and these would become predominant in such an environment by selectivity (7, 101). But laboratory studies always cover too brief a period to give data showing an im- pressive degree of adaptation, and artificially induced mutations of this type have not been reported. A review of adaptation was made by Precht and others {888).
TABLE 12.—Effect of adaptation by growth at —5^ C on Pénicillium glaucum at —5° to 2°
growth of
Strain Time required for visible growth at—
-5° C. -2" C. 0° C. 2° C.
Unadapted Days
96 29
Days 19 12
Days 19 12
Days 16
Adapted 9
SOURCE: Chistyakov, F. M., and Noskova, G. (75),
We have found no report that a mesophile has ever been adapted to grow below its normal minimum growth temperature. On

28 TECHNICAL BULLETIN 1320, U.S. DEPT. OP AGRICULTURE
10
i>0
2 7
Streptococcus fecal'is
Streptococcus lactis
1 0 10 20 30 40
TEMPERATURE, °C FIGURE 18.—Effect of incubation temperature on total cell crop in milk (133).
a rich medium, however, a thermophile was made to grow at 37° C. (307).
EFFECT OF WATER AVAILABILITY ON MICROBIAL ACTIVITIES
In addition to being dependent on temperature, microbial activi- ties are also highly dependent on availability of water, and their water and temperature requirements are interdependent. The term usually used to designate availability of water is "water activity" (aw), defined as P/Po, where P and Po are vapor pres- sures of the solution or system under consideration and of pure water, respectively. P/PoXlOO is also the relative humidity (RH) of the atmosphere in equilibrium with the solution. Thus, pure water will have an a^ of 1.00 and be in equilibrium with an RH of 100 percent; a molar concentration of an ideal nonionized solute will have an a^ of 0.9823 U26).
If the solution begins to freeze, pure water freezes out and the solution becomes more concentrated. This process continues as the temperature is lowered. Under these conditions, the solute
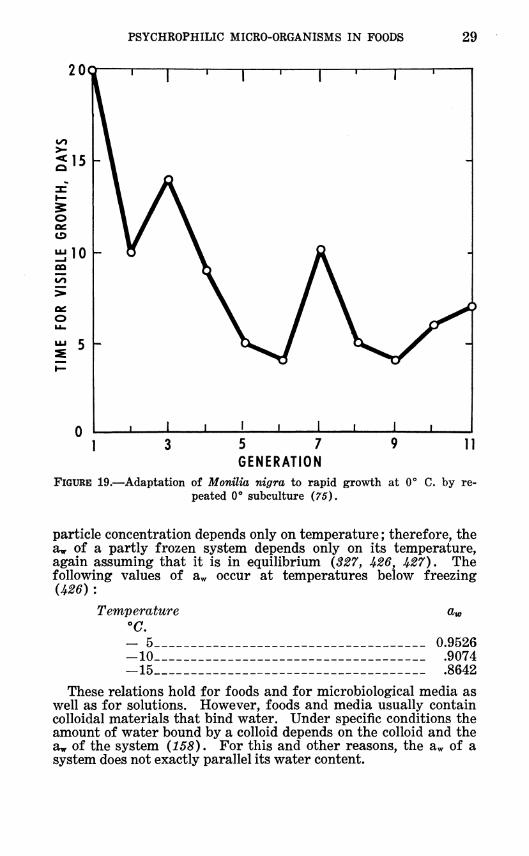
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 29
13 5 7 GENERATION
FIGURE 19.—Adaptation of Monilia nigra to rapid growth at 0° peated 0° subculture (75).
11
C. by re-
particle concentration depends only on temperature ; therefore, the aw of a partly frozen system depends only on its temperature, again assuming that it is in equilibrium (327, J^26, JÍ27) . The following values of aw occur at temperatures below freezing {JÍ26) :
Temperature a«,
- 5 0.9526 -10 .9074 -15 .8642
These relations hold for foods and for microbiological media as well as for solutions. However, foods and media usually contain colloidal materials that bind water. Under specific conditions the amount of water bound by a colloid depends on the colloid and the aw of the system (158). For this and other reasons, the aw of a system does not exactly parallel its water content.
30 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
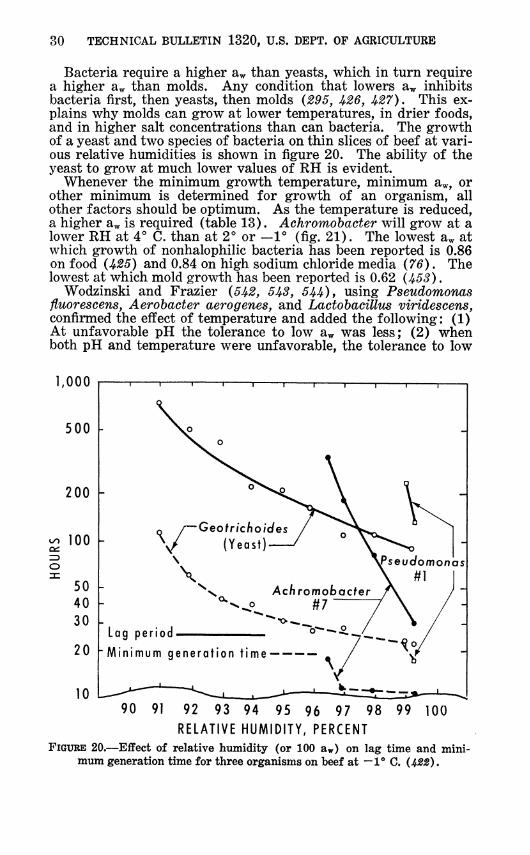
Bacteria require a higher aw than yeasts, which in turn require a higher aw than molds. Any condition that lowers aw inhibits bacteria first, then yeasts, then molds (295, ^26, ^27). This ex- plains why molds can grow at lower temperatures, in drier foods, and in higher salt concentrations than can bacteria. The growth of a yeast and two species of bacteria on thin slices of beef at vari- ous relative humidities is shown in figure 20. The ability of the yeast to grow at much lower values of RH is evident.
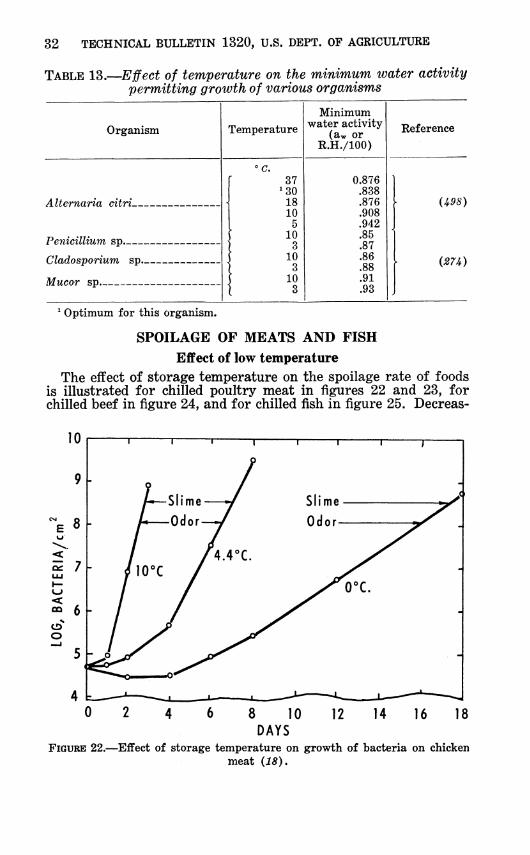
Whenever the minimum growth temperature, minimum aw, or other minimum is determined for growth of an organism, all other factors should be optimum. As the temperature is reduced, a higher aw is required (table 13). Achromobacter will grow at a lower RH at 4° C. than at 2° or —1° (fig. 21). The lowest aw at which growth of nonhalophilic bacteria has been reported is 0.86 on food (Í25) and 0.84 on high sodium chloride media (76). The lowest at which mold growth has been reported is 0.62 (Í53).
Wodzinski and Frazier (54'2, 5^3, 5H), using Pseudomonas fluor escens, Aerobacter aero g enes, and Lactobacillus viridescens, confirmed the effect of temperature and added the following: (1) At unfavorable pH the tolerance to low aw was less; (2) when both pH and temperature were unfavorable, the tolerance to low
1,000
500
200
^ 100
50 40 30
Lag period
'Geotrichoides (Yeast)
seudomonas #1
20 - Minimum generation time
10
Ac/iromobacfer #7
90 91 92 93 94 95 96 97 98 99 100 RELATIVE HUMIDITY, PERCENT
FIGURE 20.—Effect of relative humidity (or 100 aw) on lag time and mini- mum generation time for three organisms on beef at —1° C.
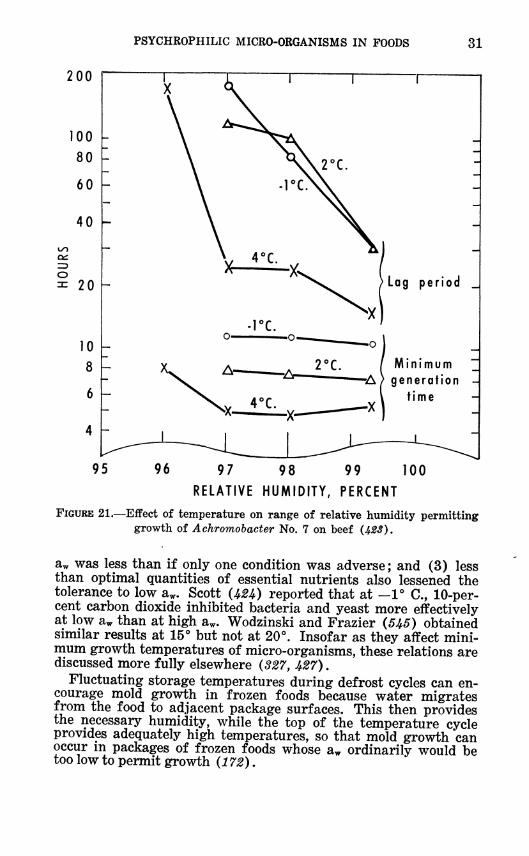
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 31
200
Lag period _
Minimum 'A ^ generation -
time •X
RELATIVE HUMIDITY, PERCENT
FIGURE 21.—Eifect of temperature on range of relative humidity permitting growth of Achromobacter No. 7 on beef (423).
aw was less than if only one condition was adverse; and (3) less than optimal quantities of essential nutrients also lessened the tolerance to low a^. Scott U2i) reported that at —1° C, 10-per- cent carbon dioxide inhibited bacteria and yeast more effectively at low aw than at high a^ Wodzinski and Frazier (54.5) obtained similar results at 15° but not at 20°. Insofar as they affect mini- mum growth temperatures of micro-organisms, these relations are discussed more fully elsewhere (827, 427).
Fluctuating storage temperatures during defrost cycles can en- courage mold growth in frozen foods because water migrates from the food to adjacent package surfaces. This then provides the necessary humidity, while the top of the temperature cycle provides adequately high temperatures, so that mold growth can occur m packages of frozen foods whose aw ordinarily would be too low to permit growth {172).
32 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
TABLE 13.—Ejfect of temperature on the minimum water activity permitting growth of various organisms
Organism
Alternaria citri-
Penicillium sp.
Cladosporium sp..
Mucor sp
Temperature
•c. 37
^30 18 10
5 10
3 10
3 10
3
Minimum water activity
(aw or R.H./IOO)
0.876 .838 .876 .908 .942 .85 .87 .86 .88 .91 .93
Reference
U98)
(27 J^)
^ Optimum for this organism.
SPOILAGE OF MEATS AND FISH Effect of low temperature
The effect of storage temperature on the spoilage rate of foods is illustrated for chilled poultry meat in figures 22 and 23, for chilled beef in figure 24, and for chilled fish in figure 25. Decreas-
8 10 12 14 16 18 DAYS
FIGURE 22.—Eifect of storage temperature on growth of bacteria on chicken meat (18).
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 33
5 10 TEMPERATURE,
15 20
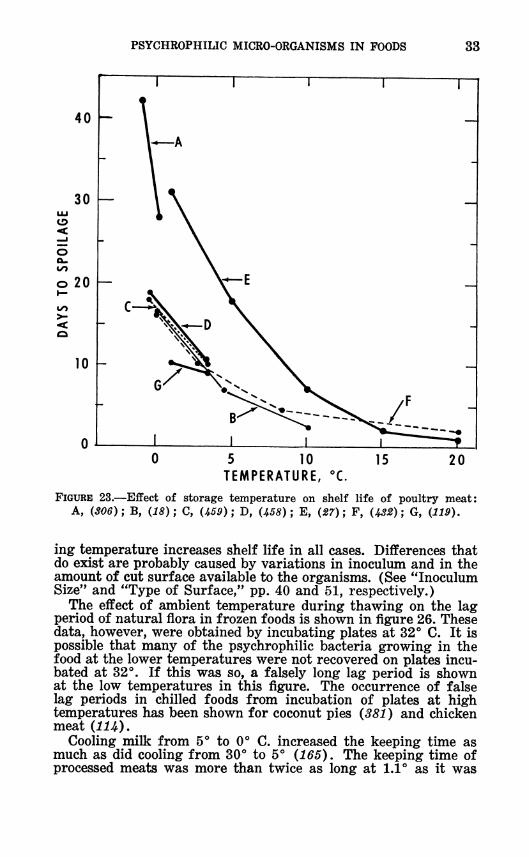
FIGURE 23.—Eifect of storage temperature on shelf life of poultry meat: A, (306); B, {18); C, {1,59) ; D, {JÍ58) ; E, {27); F, {J^2); G, {119).
ing temperature increases shelf life in all cases. Differences that do exist are probably caused by variations in inoculum and in the amount of cut surface available to the organisms. (See "Inoculum Size" and "Type of Surface," pp. 40 and 51, respectively.)
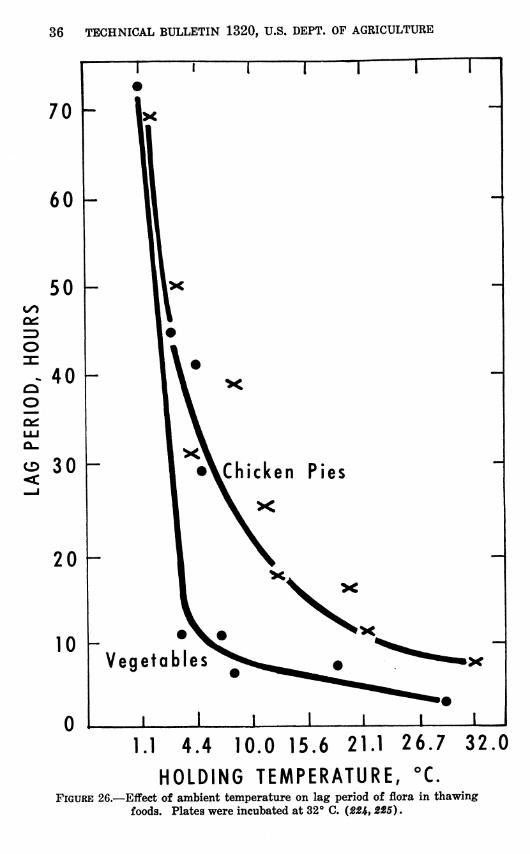
The effect of ambient temperature during thawing on the lag period of natural flora in frozen foods is shown in figure 26. These data, however, were obtained by incubating plates at 32° C. It is possible that many of the psychrophilic bacteria growing in the food at the lower temperatures were not recovered on plates incu- bated at 32°. If this was so, a falsely long lag period is shown at the low temperatures in this figure. The occurrence of false lag periods in chilled foods from incubation of plates at high temperatures has been shown for coconut pies {381 ) and chicken meat {IH).
Cooling milk from 5° to 0° C. increased the keeping time as much as did cooling from 30° to 5° {165). The keeping time of processed meats was more than twice as long at 1.1° as it was
34 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
O 5 10 TEMPERATURE, °C.
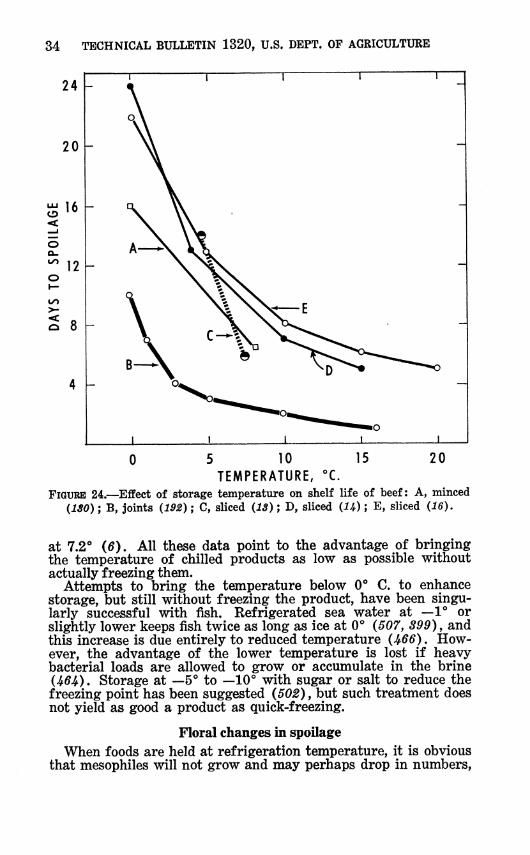
FIGURE 24.—Effect of storage temperature on shelf life of beef: A, minced {ISO) ; B, joints (192) ; C, sliced (13) ; D, sliced (i^) ; E, sliced (16).
Sit 7.2° (6). All these data point to the advantage of bringing the temperature of chilled products as low as possible without actually freezing them.
Attempts to bring the temperature below 0"" C. to enhance storage, but still without freezing the product, have been singu- larly successful with fish. Refrigerated sea water at —V or slightly lower keeps fish twice as long as ice at 0° (507, 399), and this increase is due entirely to reduced temperature (^66). How- ever, the advantage of the lower temperature is lost if heavy bacterial loads are allowed to grow or accumulate in the brine U6^). Storage at —5° to —10° with sugar or salt to reduce the freezing point has been suggested (502), but such treatment does not yield as good a product as quick-freezing.
Floral changes in spoilage When foods are held at refrigeration temperature, it is obvious
that mesophiles will not grow and may perhaps drop in numbers.
PSYCHROPHILIC MICRO-ORGANISMS IN POODS 35
30
20
CD <I5 o a.
O
>- 10 -
5 -
Z \ Z N
Ô \ «^ \
- ü> rE\ \- \
C-
1 1 1 1
—A
1 1 "i"""-....,»^
10 20 TEMPERATURE,
30 40
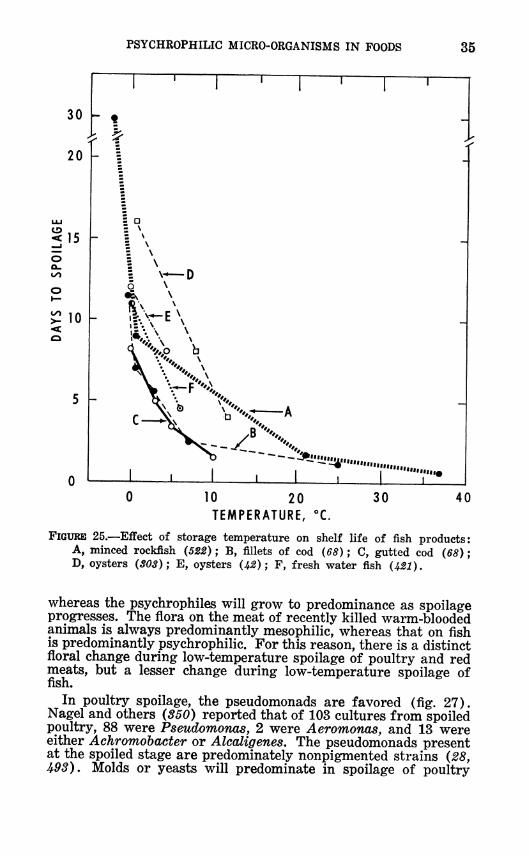
FIGURE 25.—Effect of storage temperature on shelf life of fish products: A, minced rockfish (522) ; B, fillets of cod (68) ; C, gutted cod (68) ; D, oysters (SOS); E, oysters (^) ; F, fresh water fish (421).
whereas the psychrophiles will grow to predominance as spoilage progresses. The flora on the meat of recently killed warm-blooded animals is always predominantly mesophilic, whereas that on ñsh is predominantly psychrophilic. For this reason, there is a distinct floral change during low-temperature spoilage of poultry and red meats, but a lesser change during low-temperature spoilage of fish.
In poultry spoilage, the pseudomonads are favored (fig. 27). Nagel and others (850) reported that of 103 cultures from spoiled poultry, 88 were Pseudomonas, 2 were Aeromonas, and 13 were either Achromobacter or Alcaligenes. The pseudomonads present at the spoiled stage are predominately nonpigmented strains (28, 493). Molds or yeasts will predominate in spoilage of poultry
36 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
—1 r
70
60 _
50
^ 40 o o LU
o 30 <
20 -
10
T 1 r
Vegetables^
1 1 1 1.1 4.4 lO.O 15.6 21.1 26.7 32.0
HOLDING TEMPERATURE, °C. FIGURE 26.—Effect of ambient temperature on lag period of flora in thawing
foods. Plates were incubated at 32° C. (224, 225).
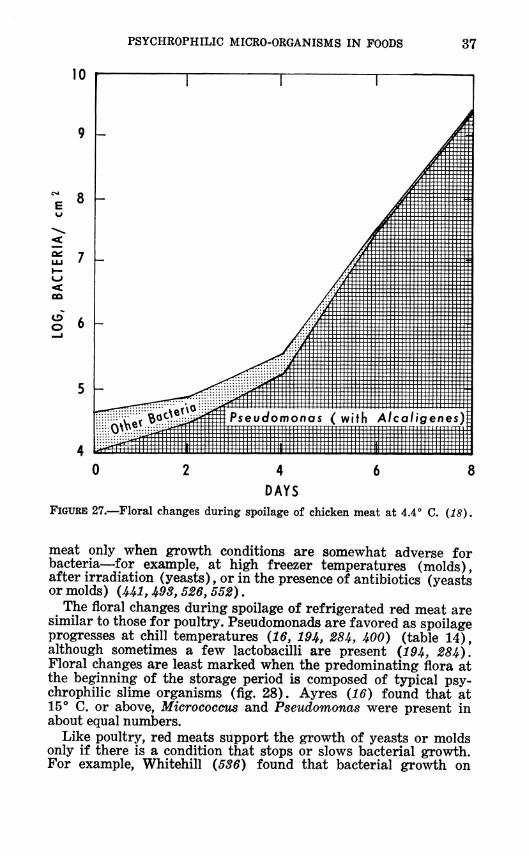
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 37
0 2 4 6 f
DAYS FIGURE 27.--Floral changes during spoilage of chicken meat at 4.4° C. (18),
meat only when growth conditions are somewhat adverse for bacteria—^for example, at high freezer temperatures (molds), after irradiation (yeasts), or in the presence of antibiotics (yeasts or molds) (Ul, 493, 526, 552).
The floral changes during spoilage of refrigerated red meat are similar to those for poultry. Pseudomonads are favored as spoilage progresses at chill temperatures (16, 194, 284, 400) (table 14), although sometimes a few lactobacilli are present (194, 284). Floral changes are least marked when the predominating flora at the beginning of the storage period is composed of typical psy- chrophilic slime organisms (fig. 28). Ayres (16) found that at IB"" C. or above, Micrococcus and Pseudomonas were present in about equal numbers.
Like poultry, red meats support the growth of yeasts or molds only if there is a condition that stops or slows bacterial growth. For example, Whitehill (536) found that bacterial growth on
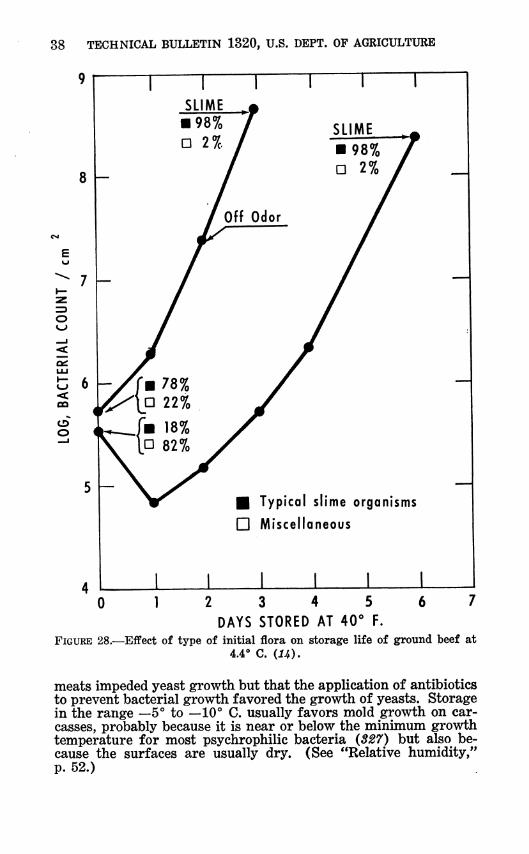
38 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
—\ \ 1 \ 1 r— SLIME ,j
■ Typical slime organisms
D Miscellaneous
1 O 12 3 4 5 6 7
DAYS STORED AT 40° F. FIGURE 28.—Effect of type of initial flora on storage life of ground beef at
4.4° C. iU).
meats impeded yeast growth but that the application of antibiotics to prevent bacterial growth favored the growth of yeasts. Storage in the range —5° to —10° C. usually favors mold growth on car- casses, probably because it is near or below the minimum growth temperature for most psychrophilic bacteria (S27) but also be- cause the surfaces are usually dry. (See "Relative humidity," p. 52.)

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 39
TABLE 14.—Changes in bacterial flora during spoilage of ground beef at 7" C.
Micro-organism
Proportion of total isolates* after—
0 days 14 days
Pseudomonas or Achromobacter, or both Bacillus
Percent
4 28
0 26 42
Percent
84 6 5 5 0
Microbacterium Micrococcus ^ Yeasts _
Total ._ . 100 100
* From plates at 7° C. SOURCE: Rogers, R. E., and McCleskey, C. S. UOO).
Spoilage of sausages falls into a special category because they are usually heated in impervious casings where there is low oxy- gen tension and fairly high salt concentration, and are then re- frigerated. Few heat-resistant organisms can grow under these conditions. In one investigation (Í65), the predominant organ- isms in freshly made liver sausage were aerobic micrococci and sporulating rods, neither of which grew at 5° C. An organism similar to a Lactohacillus and a facultative anaerobic yeast finally spoiled the product after several weeks. Processed vacuum-packed meats have been spoiled by lactobacilli {6). On the other hand, the slime that commonly forms on the surface and between the layers of sausage casings is due to the growth of yeasts, micro- cocci, and aerobic gram-negative rods (lOUy 4^65). A review of sausage spoilage has been made by Jensen (263).
Floral changes during decomposition of fish caught in the tem- perate or arctic zones are not so marked as they are in meat spoilage because the organisms predominating naturally on the surface of the live or recently killed fish, particularly after it contacts ice and deck surfaces, are the same psychrophiles as the ones that cause decomposition (11, 65, 228, US7, US2). However, the PseudomonaS'AchroTnohacter ^ group is favored during spoil- age in ice (69,162, 202, JÍS9, UO, 5JÍ9). In tropic zones the floral change in favor of the Pseudomonas group during low-tempera- ture spoilage is more marked because of the lower percentage of psychrophiles on the recently killed fish (^37). The nonpigmented pseudomonads contribute most of the tissue breakdown and odors of decomposition (HO). FZavobaciermm plays a minor role (^35, 439).
'Many organisms that in earlier work were identified as Achromobacter because they lacked pigments would now, according to the 5th, 6th, and 7th editions of Bergey's Manual (33, U, -45) be classed as Pseudomonas on the basis of flagellation (50, 2J^9).

40 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
Floral changes in favor of the PseudomoncLS-Achromohacter group also occur in crabmeat and in shrimp during spoilage (tables 15 and 16) {5, 60, U95).
TABLE 15.—Changes in flora of Louisiana crabmeat stored at S"" to 5° C.
Colonies picked
Proportion of total isolates
Days in storage Cocci
Pseudomonas- Archomo'
bacter Other
bacteria
1 _ - Number
525 420 598
Percent 60.4 30.9
3.0
Percent 23.4 60.0 96.3
Percent 16.2
5-8 9.1 11-15 .7
SOURCE: Alford, J. A., Tobin, L., and McCleskey, C. S. (5).
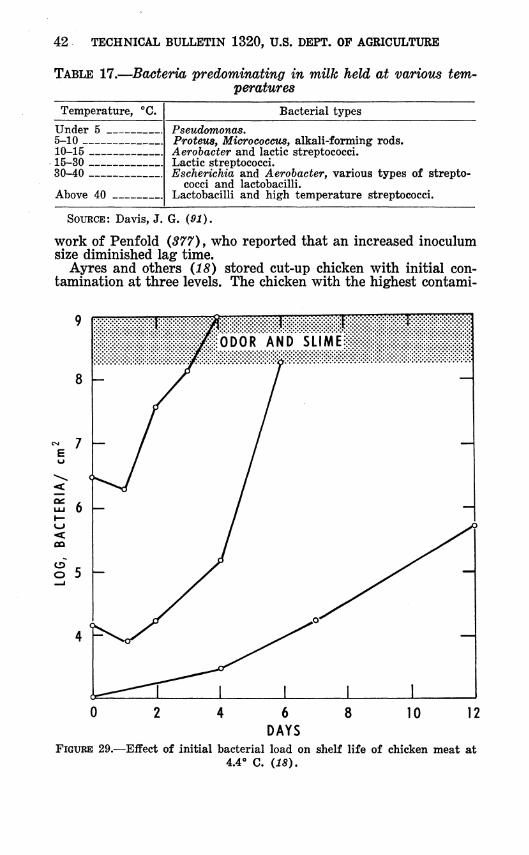
The type of organism growing in milk during storage depends on the storage temperature, as shown in table 17. Pseudomonas predominates at refrigeration temperatures. However, some of the conforms also grow readily in milk at 3*^ to 5° C. (371). On the basis of 586 isolations, Schultze ® reported the bacterial content of pasteurized milk stored at 4° for 1 to 2 weeks as follows:
Percent
7.9 9.2
.7 10.8
.8
100.0
Pseudomonas Alcaligenes Achromobacter Flavobacterium Conforms Yeasts
Total *65.2 percent of these were fluorescent.
Inoculum size It is well established that high initial contamination results in
more rapid spoilage of chilled foods than does low initial con- tamination. Duncan and Nickerson (105), however, showed that inoculum size did not aifect growth rate in the logarithmic phase of a pure culture of Pseudomonas fragi. Essentially the same observation can be made with the natural inoculum on beef and on chicken meat (figs. 28 and 29) with the exception of the lowest inoculum in figure 29, which may not have been followed long enough to reach logarithmic growth. In the experiments repre- sented by these figures, the cultures grown from the smaller inoc- ula had longer lag periods. These observations confirm the early
• SCHULTZE, W. D. AN INVESTIGATION OF PSYCHROPHUJC BACTERIA, I. THE NATURE AND DISTRIBUTION OF PSYCHROPHILIC BACTERIA IN COMMERCIALLY PASTEURIZED DAIRY PRODUCTS. II. STUDIES OP THE INFLUENCE OP GROWTH AND REACTION TEMPERATURE OF THE METABOLISM OF A TYPICAL PSYCHROPHILE, Thesis, Ph. D., Univ. Minn., 190 pp. 1958. (Abstract in. Diss. Abs. 19 (10) : 2434. 1959.)
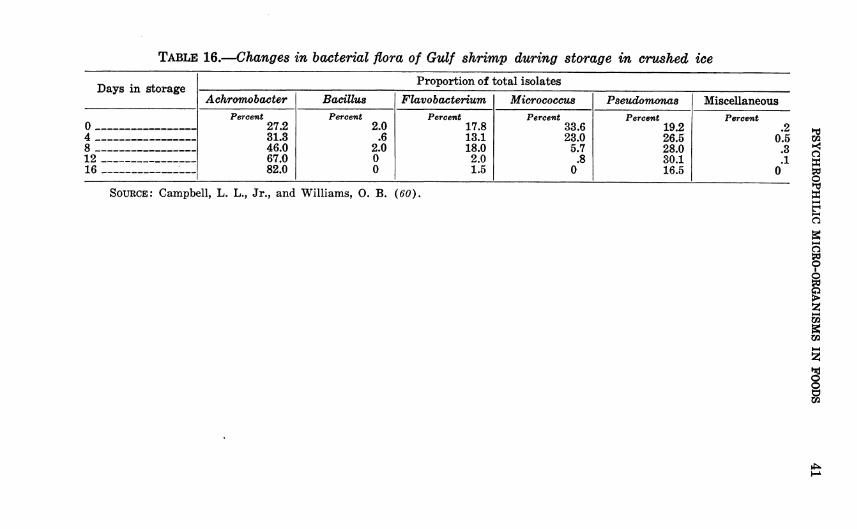
TABLE 16.—Changes in bacterial flora of Gulf shrimp during storage in crushed ice
Days in storage Proportion of total isolates
Achromobacter Bacillus Flavo bacterium Micrococcua Pseudomonas Miscellaneous
0 Percent
27.2 31.3 46.0 67.0 82.0
Percent 2.0
.6 2.0 0 0
Percent 17.8 13.1 18.0 2.0 1.5
Percent 33.6 23.0
5.7 .8
0
Percent 19.2 26.5 28.0 30.1 16.5
Percent 2
4 0 5 3
12 — - — 1 16 - - - —: 0
SOURCE: Campbell, L. L., Jr., and Williams, O. B. (60).
K¡ o to o »Tí
O
É
g
i
42 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
TABLE 17.—Bacteria predominating in milk held at various tem- peratures
Temperature, **€. Bacterial types
Under 5 5-10 10-15 _
Pseudomonas, Proteus, MicrococctLS, alkali-forming rods. Aerobacter and lactic streptococci.
15-30 30-40
Lactic streptococci. Escherichia and Aerobacter, various types of strepto-
cocci and lactobacilli. Lactobacilli and high temperature streptococci. Above 40
SOURCE: Davis, J. G. (91).
work of Penfold (377), who reported that an increased inoculum size diminished lag time.
Ayres and others (18) stored cut-up chicken with initial con- tamination at three levels. The chicken with the highest contami-
FSUMEí^^^^^^
:':':':':':':':':':^':':
10 12 6 DAYS
FIGURE 29.—Effect of initial bacterial load on shelf life of chicken meat at 4.4" C, (18).
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 43
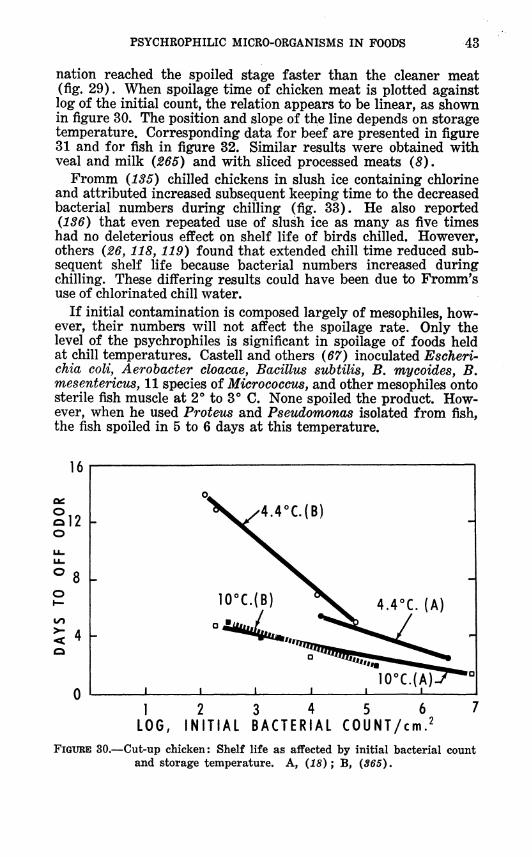
nation reached the spoiled stage faster than the cleaner meat (fig. 29). When spoilage time of chicken meat is plotted against log of the initial count, the relation appears to be linear, as shown in figure 30. The position and slope of the line depends on storage temperature. Corresponding data for beef are presented in figure 31 and for fish in figure 32. Similar results were obtained with veal and milk (265) and with sliced processed meats (8).
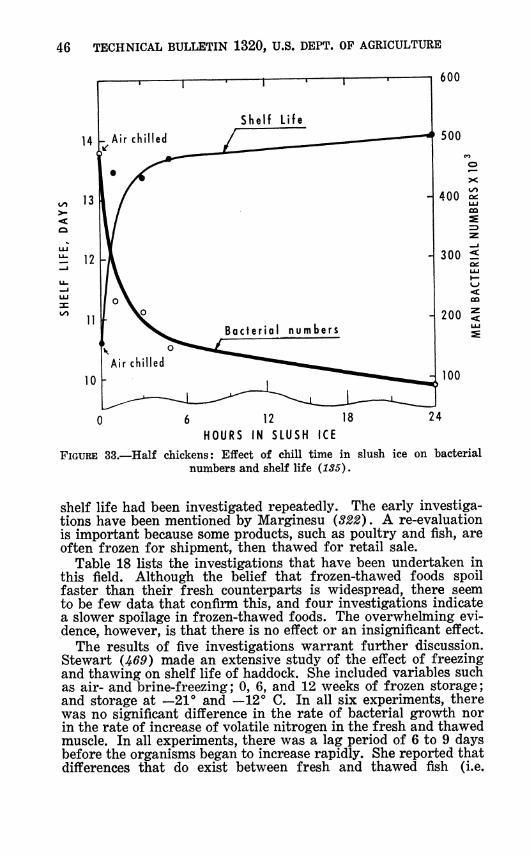
Fromm (135) chilled chickens in slush ice containing chlorine and attributed increased subsequent keeping time to the decreased bacterial numbers during chilling (fig. 33). He also reported (136) that even repeated use of slush ice as many as five times had no deleterious effect on shelf life of birds chilled. However, others (26, 118,119) found that extended chill time reduced sub- sequent shelf life because bacterial numbers increased during chilling. These differing results could have been due to Fromm's use of chlorinated chill water.
If initial contamination is composed largely of mesophiles, how- ever, their numbers will not affect the spoilage rate. Only the level of the psychrophiles is significant in spoilage of foods held at chill temperatures. Castell and others (67) inoculated Escheri- chia coll, Aerobacter cloacae, Bacillus subtilis, B. mycoides, B. mesenterictis, 11 species of Micrococcus, and other mesophiles onto sterile fish muscle at 2"* to 3° C. None spoiled the product. How- ever, when he used Proteits and Pseudomonas isolated from fish, the fish spoiled in 5 to 6 days at this temperature.
16
o
°8 o
< ^ o
'4.4°C.(B)
lOoC.ÍB) A^ 4.40c. (A)
10°C.(A).
12 3 4 5 6 7 LOG, INITIAL BACTERIAL COUNT/cm.^
FiGxniE 30.—Cut-up chicken: Shelf life as aifected by initial bacterial count and storage temperature. A, (18); B, (365).
44 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
24
LOG, INITIAL BACTERIAL COUNT/c m
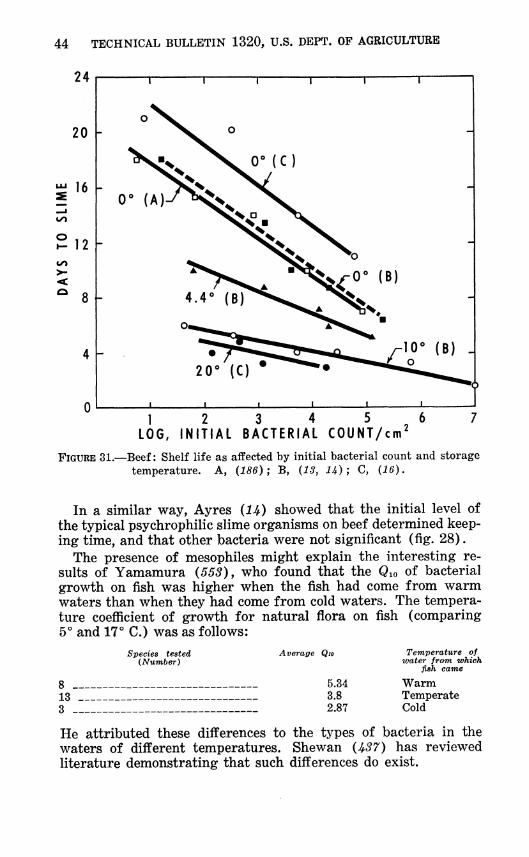
FIGURE 31.—Beef: Shelf life as affected by initial bacterial count and storage temperature. A, (186); B, (13, 1J^) ; C, (16).
In a similar way, Ay res (H) showed that the initial level of the typical psychrophilic slime organisms on beef determined keep- ing time, and that other bacteria were not significant (fig. 28).
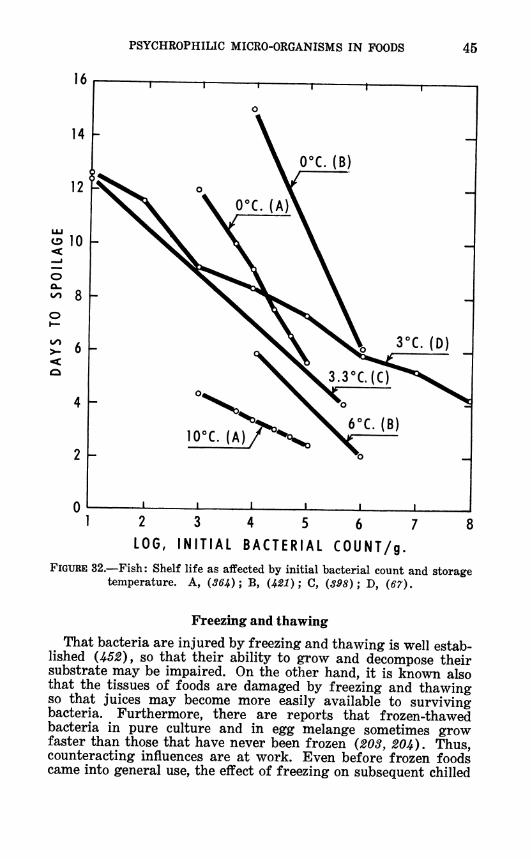
The presence of mesophiles might explain the interesting re- sults of Yamamura (553), who found that the Qio of bacterial growth on fish was higher when the fish had come from warm waters than when they had come from cold waters. The tempera- ture coefficient of growth for natural ñora on fish (comparing 5° and 17° C.) was as follows:
Species tested Average Qio Temperature of (Number) water from which
fish came
8 5.34 Warm 13 3.8 Temperate 3 2.87 Cold
He attributed these differences to the types of bacteria in the waters of different temperatures. Shewan {J^37) has reviewed literature demonstrating that such differences do exist.
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 45
2 3 4 5 6 7 8
LOG, INITIAL BACTERIAL COUNT/g.
FIGURE 32.~Fish: Shelf life as affected by initial bacterial count and storage temperature. A, (36^^); B, (421); C, (398); D, (67).
Freezing and thawing
That bacteria are injured by freezing and thawing is well estab- lished U52), so that their ability to grow and decompose their substrate may be impaired. On the other hand, it is known also that the tissues of foods are damaged by freezing and thawing so that juices may become more easily available to surviving bacteria. ^ Furthermore, there are reports that frozen-thawed bacteria in pure culture and in egg melange sometimes grow faster than those that have never been frozen (208, 20A). Thus, counteracting influences are at work. Even before frozen foods came into general use, the effect of freezing on subsequent chilled
46 TECHNICAL BULLETIN 1320, U.S. DBPT. OP AGRICULTURE
600
500
>- <
± 12
3:
300 5
< CÛ
FIGURE 33.-
6 12 18 HOURS IN SLUSH ICE
-Half chickens: Effect of chill time in slush ice on bacterial numbers and shelf life (ISö),
shelf life had been investigated repeatedly. The early investiga- tions have been mentioned by Marginesu (322). A re-evaluation is important because some products, such as poultry and fish, are often frozen for shipment, then thawed for retail sale.
Table 18 lists the investigations that have been undertaken in this field. Although the belief that frozen-thawed foods spoil faster than their fresh counterparts is widespread, there seem to be few data that confirm this, and four investigations indicate a slower spoilage in frozen-thawed foods. The overwhelming evi- dence, however, is that there is no effect or an insignificant effect.
The results of five investigations warrant further discussion. Stewart (4^69) made an extensive study of the effect of freezing and thawing on shelf life of haddock. She included variables such as air- and brine-freezing; 0, 6, and 12 weeks of frozen storage; and storage at —2V and —12° C. In all six experiments, there was no significant difference in the rate of bacterial growth nor in the rate of increase of volatile nitrogen in the fresh and thawed muscle. In all experiments, there was a lag period of 6 to 9 days before the organisms began to increase rapidly. She reported that differences that do exist between fresh and thawed fish (i.e.
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 47
TABLE 18.—Review of investigations showing the effect of freez- ing and thawing on the subsequent spoilage rate of foods
Effect of freeze-thaw
(investigators' conclusions)
Product Reviewers' comments Pveferences
Fats, proteins—
Meat
An enzyme study; no bacteriological data.
No data presented Claim of faster spoil-
age based on inade- quate data.
No data presented Thawed and nonfroz-
en chickens were separate lots; di-
rect comparison not warranted.
E, coli growth rate Longer lag, slower
growth Longer lag after 3 to
6 months storage. Slightly longer lag period _
(23)
{ISO)
Faster spoilage.
do
Chickens ' do
Eggs _
{167)
{37A) {5U^ 525)
{203, 20U) WS) [Beef, pork
Slower Fish _
{310) spoilage.
do
'Meat _ Investigator's conclu- sion that rate of thaw or length of storage influenced spoilage seems based on inade- quate data.
Growth slightly fast- er on frozen- thawed, but author considered the dif- ference insignifi- cant.
See table 17
{322)
Insignificant effect or none.
do
Horse, pork Beef _
{277)
{286, 287) {U72)
{lU, 355, U56, U57) {115, U71)
{527) {86, j^69, 470)
{m) {535)
Chickens Fish Herring Haddock Halibut Peas
See figure 34 Slightly longer lag __.
* PiVNICK, H. BACTERIOLOGICAL AND BIOCHEMICAL STUDIES OF THE RATE OF DECOMPOSITION OF UNFROZEN AND DEFROSTED COD MUSCLE. Thesis, M. S. Dalhousie Univ., Halifax, N. S., 81 pp. 1949.
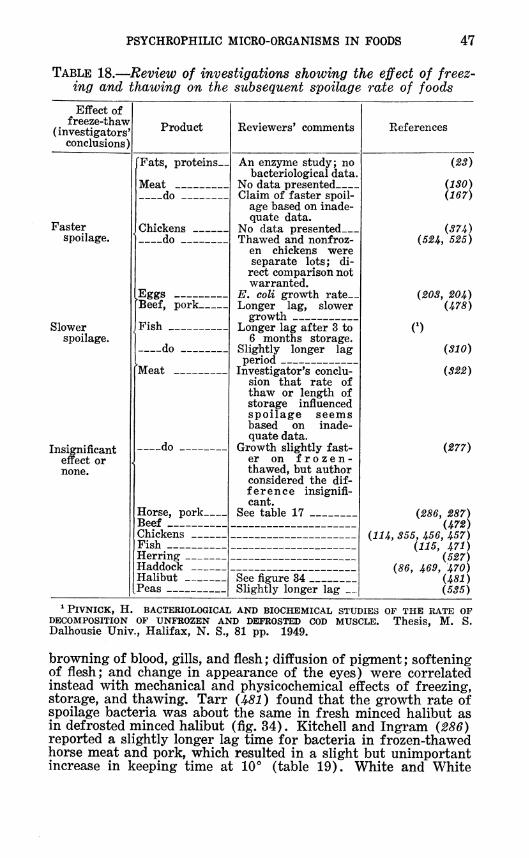
browning of blood, gills, and flesh ; diffusion of pigment ; softening of flesh; and change in appearance of the eyes) were correlated instead with mechanical and physicochemical effects of freezing, storage, and thawing. Tarr (JÍ81 ) found that the growth rate of spoilage bacteria was about the same in fresh minced halibut as in defrosted minced halibut (fig. 34). Kitchell and Ingram {286) reported a slightly longer lag time for bacteria in frozen-thawed horse meat and pork, which resulted in a slight but unimportant increase in keeping time at lO"" (table 19). White and White
48 TECHNICAL BULLETIN 1320, U.S. DEPT. OP AGRICULTURE
O
Oí
< ÛÛ
o" O
11
10
•Viable count, fresh Halibut
-x—x—x-Direct count; fresh Halibut
-a—a—-D-Viable count, defrosted Halibut
..A....A...A.. Direct count, defrosted Halibut
10 20 30 40 50 60
DAYS STORED AT 1.5° C.
70 80 90
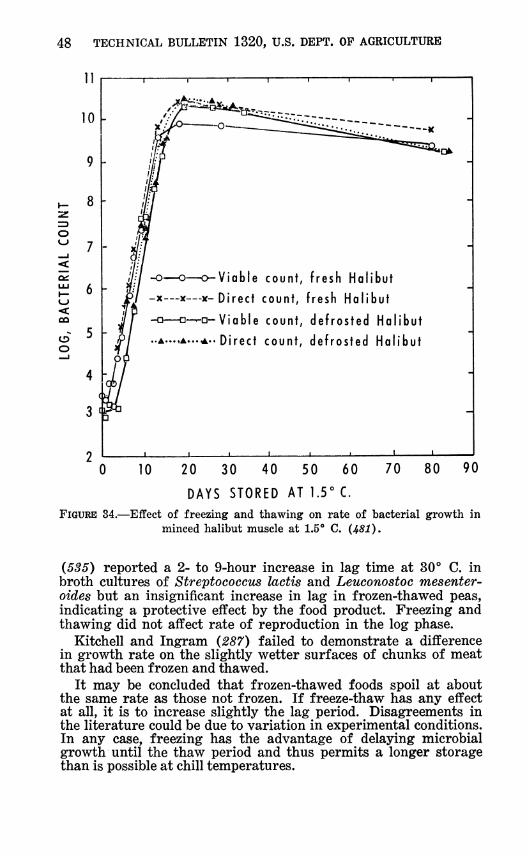
FIGURE 34.—Effect of freezing and thawing on rate of bacterial growth in minced halibut muscle at Lö** C. (481).
(5S5) reported a 2- to 9-hour increase in lag time at 30° C. in broth cultures of Streptococcus lactis and Leuconostoc mesenter- oides but an insignificant increase in lag in frozen-thawed peas, indicating a protective effect by the food product. Freezing and thawing did not affect rate of reproduction in the log phase.
Kitchell and Ingram (287) failed to demonstrate a difference in growth rate on the slightly wetter surfaces of chunks of meat that had been frozen and thawed.
It may be concluded that frozen-thawed foods spoil at about the same rate as those not frozen. If freeze-thaw has any effect at all, it is to increase slightly the lag period. Disagreements in the literature could be due to variation in experimental conditions. In any case, freezing has the advantage of delaying microbial growth until the thaw period and thus permits a longer storage than is possible at chill temperatures.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 49
TABLE 19.—Effect of freezing on subsequent bacterial spoilage of meats at lO"" C.
Product Treatment
Time to reach 30 XlO* bacteria/cm. ^
Frozen- thawed
Not frozen
Pork Minced Hours
20 40 41 50 40 32 35 38 37
Hours 18 33 43 51 38 29 29 29 33.8
Horse do Do . _ Sliced, post rigor Do Sliced, nre-rieror Do Natural surfaces
Stored 7 days at -20** C. Stored 28 days at -20° C. Stored 56 days at -20° C.
Do - Do - Do _
Average _
SOURCE: Kitchell, A. G., and Ingram, M. (286),
Semipreserved canned hams and luncheon meats will keep well, even though they have received only a pasteurization treatment. However, after they have been chilled or frozen and then held above a chill temperature, their keeping time is relatively short. Halvorson and others (197) have reviewed the literature and offer three possible explanations for this phenomenon:
1. Low-temperature storage alters the food to make it a more favorable medium for spore germination and growth.
2. Low-temperature storage may alter spores so that they germinate more rapidly when brought to a higher temper- ature.
3. The spores may actually germinate at low temperatures.
Heat
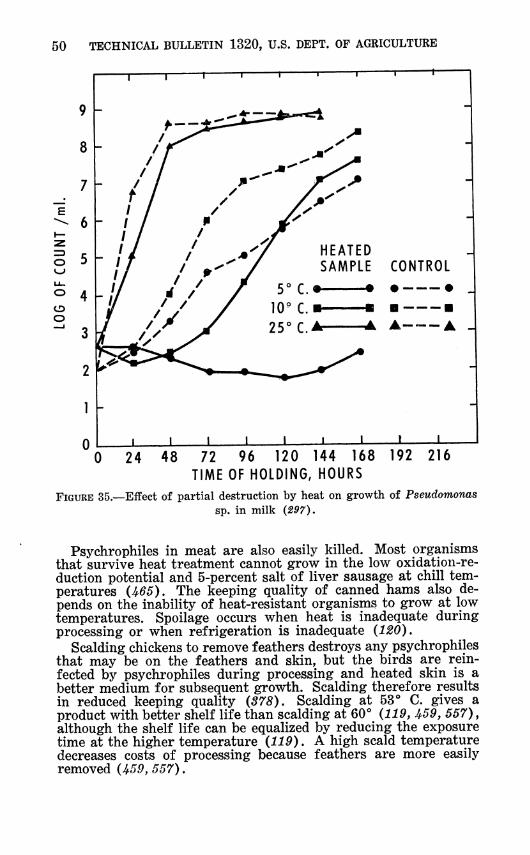
Experience in marine microbiology (561) and in milk products (see p. 79) has estabhshed the fact that most psychrophiles are easily destroyed by relatively mild heat. Bacteria surviving pas- teurization cannot grow in milk at refrigerator temperatures (91, 491, 541). Lawton and Nelson (297\ found that when part of a population of a psychrophilic bacterium was killed by heat, the survivors had a slightly extended lag period at 25 ^^ C, a greatly extended lag at 10°, and there was no growth at 5° in a 7-day period (fig. 35). By dilution techniques it was determined that the absolute level of viable organisms at the beginning of the ex- periment did not affect the lag period in the range of numbers used. Also, chlorine did not result in the same prolonged lag at S**. Sublethal heat also narrows the pH range of growth of Pseudomonas and makes them more fastidious in their require- ments for nutrients (207, 208).
The presence of psychrophiles in pasteurized milk products is due to postpasteurization contamination, and scrupulous sanita- tion after pasteurization is necessary to ensure good keeping time (870).
50
o
o o
TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
-1 1 1 r- T T -r -r
HEATED SAMPLE CONTROL
# # •
A A-'—A -
±. ± X J_ J_ J- 0 24 48 72 96 120 144 168 192 216
TIME OF HOLDING, HOURS
FIGURE 35.—Eifect of partial destruction by heat on growth of Pseudomonas sp. in milk (297),
Psychrophiles in meat are also easily killed. Most organisms that survive heat treatment cannot grow in the low oxidation-re- duction potential and 5-percent salt of liver sausage at chill tem- peratures U65). The keeping quality of canned hams also de- pends on the inability of heat-resistant organisms to grow at low temperatures. Spoilage occurs when heat is inadequate during processing or when refrigeration is inadequate {120).
Scalding chickens to remove feathers destroys any psychrophiles that may be on the feathers and skin, but the birds are rein- fected by psychrophiles during processing and heated skin is a better medium for subsequent growth. Scalding therefore results in reduced keeping quality {S78). Scalding at 53° C. gives a product with better shelf life than scalding at 60° {119,1,59, 557), although the shelf life can be equalized by reducing the exposure time at the higher temperature {119). A high scald temperature decreases costs of processing because feathers are more easily removed {1,59, 557).

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 51
Type of surface
Because micro-organisms need moisture and nutrients, they are inhibited by any film or membrane that keeps these things away from them. Thus, the unbroken skin of a flesh food, a dried sur- face, or a layer of fat reduces the rate and extent of microbial growth.
Spoilage of fresh beef is primarily a surface phenomenon (219). The growth in deep portions, if any occurs, is very slow (335). Bone taint is a deep spoilage due to clostridia (191) but this does not occur at refrigeration temperatures (58,2^0). Haines (180) reported growth on lean meat at —5'' C. but not on connective tissue covering the surface fat. Uncut meat lasts longer than cut meat (192) ; thus growth of bacteria in ground meat is rapid, and very high counts may be expected (284', 530). Fat in ground meat enhances growth (196).
The primary spoilage in poultry, as in red meat, is a surface growth of bacteria whose colonies become visible and coalesce to form slime when spoilage is advanced (18, 306). Bacteria are not carried through the skin to muscle nor to the cavity unless the skin is broken (346). Once bacteria have access to a cut sur- face or the body cavity they multiply more rapidly than on the skin (92, 517), although one group of authors (325) reported a more rapid growth on excised skin than on any other excised tissue.
Bacterial decomposition is slower in undrawn (New York dressed) poultry than in eviscerated poultry because the tissues in the former are protected by uncut skin surfaces (355, 379). In earlier years, carcasses were shipped undrawn and evisceration occurred at the retail level. However, it has been found that dif- fusion of decomposition products from the gut causes an off-flavor in the meat of New York dressed birds (468). Thus when the criterion is flavor, the New York dressed bird has a shorter keep- ing time, even though bacterial growth is less (22) :
Keeping time (days)
New York dressed In ice 1.7° C. 7.2° C.
6 4 4 Eviscerated, ready-to-cook 11 11 9
However, the British still prefer the stronger flavor of meat from uneviscerated chickens (442).
Fish spoilage is likewise primarily due to growth of micro- organisms on the surface, for the flesh of sound living fish is sterile (54, 227). Unopened fish generally keep better than eviscerated fish (527) unless they have been feeding heavily, in which case the opposite is true because of the high level of bacteria and enzymes in the gut (254, 463). Whole fish keep longer than fillets, which in turn keep longer than minced fish (481). Thus to hold fish for the longest time, they should be left in the round or evis- cerated, and fillets should be cut late in the storage period (table 20).

52 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
TABLE 20.—Total storage time required for iced fish and fillets to reach a trimethylamine (TMA) value of 15
Gutted fish stored on ice
Time for fillet stored at 3^ C. to reach TMA of 15
Total storage time to reach TMA of 15
Days 1
Days 4.0 4.0 4.0 4.0 3.5 3.5 1.0
Days 5.0
2 _ - 6.0 3 7.0 6 _ 10.0 8 11.5 10 13.5 13 14.0
SOURCE: Dyer, W. J. and Dyer, F. E. (106).
Although chemical products of decomposition diffuse to the in- side (107) y penetration of organisms into fish ñesh itself is slow. Even fish that are decomposed with heavy slime and odor may still be sterile within the flesh (106, 279, 5U8). On this basis, chemical and bacteriological tests for fish spoilage are most sensi- tive if sampling is limited to the surfaces {5^8).
Niven has described three types of sausage decomposition {359, 860, 861): (1) surface slime, (2) green ring, and (3) green core. Surface slime forms only when insanitation is combined with excess moisture. Green ring and green core are caused by lactobacilli growing before and after the cook, respectively. The color forms only after the sausage is exposed to the oxygen of the air. Adequate sanitation, heat processing, dry storage condi- tions, and refrigeration control all three types of decomposition. Preslicing increases the contaminated surfaces in packaged pre- cooked meats (804^).
Relative humidity
As explained previously, relative humidity (RH) and water activity (aw) are equivalent when the system is in equilibrium. Scott {í26) described the a^ of most fresh foods as 0.98 to above 0.99, so that they offer ideal growth conditions to micro-organ- isms. Meat, however, is frequently stored at a relatively low RH to suppress surface growth of micro-organisms. Under these con- ditions, although the meat surface presumably dries slightly, the aw within the meat is never in equilibrium with the RH of the surrounding air.
Schmidt (416) investigated bacterial growth on large pieces of meat at humidities down to 73 percent. Of course, moisture mov- ing from the interior to the surface of the meat by capillarity and by diffusion presented to the organisms a higher aw than would have occurred in equilibrium conditions. Scott (428), however, studied spoilage of thin slices whose aw came promptly into equi- librium with the RH of the surrounding atmosphere. Despite this difference their results are similar and can be presented together to show the effect of RH and temperature on storage life of beef (fig. 36).
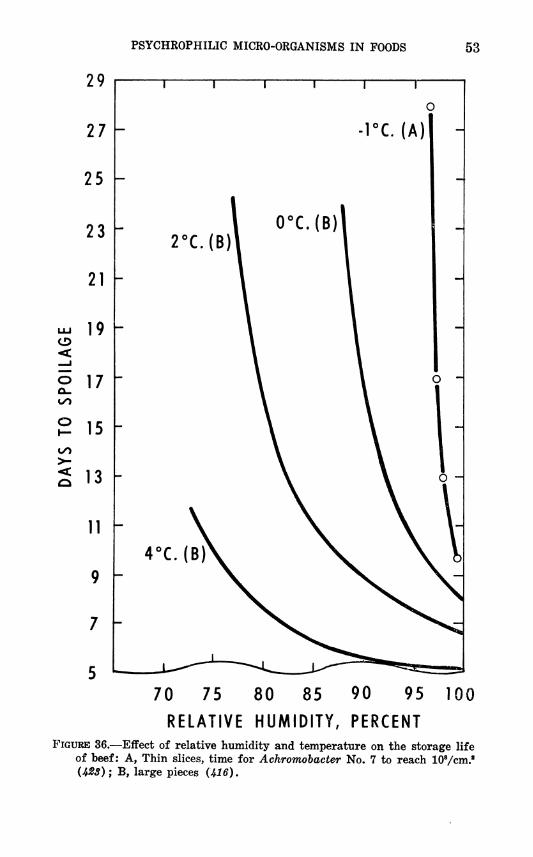
PSYCHKOPHILIC MICRO-ORGANISMS IN POODS 53
70 75 80 85 90 95 100
RELATIVE HUMIDITY, PERCENT FIGURE 36.—Effect of relative humidity and temperature on the storage life
of beef: A, Thin slices, time for Achromobacter No. 7 to reach lOVcm.' (4^3); B, large pieces W6).

54 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
Jepsen (265) stored sides of pork for 8 days at 4° C. at two levels of humidity and found the following :
Bacteria/cm'
Left sides stored at Right sides, stored at Carcass 75 to 80 percent RH 100 percent RH
Thousands Thousands 1 8 490 2 31 130 3 24 110 4 17 58 5 12 190
Allowing cold meat to "sweat'' in a warm room enhances bacterial growth on the surface after it is returned to the cold (192).
Even though surface drying is a major factor in controlling bacterial development in meats, holding them below 85 percent RH causes serious weight loss by evaporation (il7). As a com- promise, meat is generally stored commercially in atmospheres between 85 and 90 percent RH (29, 558).
In meat-storage rooms with this range of humidity, mold growth becomes the major problem. Kaess (27Í) related growth of Mucor, Pénicillium, and Cladosporium on meats to temperature and RH. Rate of growth increased directly as temperature and RH increased, but maximum growth occurred at 95 percent RH, rather than at 100 percent RH, because bacterial growth at 100 percent RH interfered with growth of the molds.
The reported investigations on the effect of RH or aw on poultry spoilage are less comprehensive and less conclusive. When com- pared with ice-chilled poultry, air-chilled poultry has been re- ported to have a longer {289, 290, 878), an equal {21), and a shorter {813) storage life. However, in the tests that showed a shorter storage life, the air-chilled poultry was held at a slightly higher temperature, which may have speeded spoilage somewhat. Ice chilling has been recommended because of the more rapid heat transfer {858), and several investigators have mentioned weight loss by evaporation from air-chilled chickens, which gives the ice chilling a competitive advantage from a profit standpoint. How- ever, prolonged storage in slush ice causes loss of flavor {88Jp).
There is a belief in the fishing industry that water from melting ice has a washing action that removes bacteria from the surfaces, thus enhancing keeping time {JÍ8JÍ). The Department of Scientific and Industrial Research of Great Britain {159,160) has reported that well-iced fish last longer at a high ambient temperature than at a low ambient temperature. They attributed this phenomenon to the greater washing action at the higher temperature. How- ever, no data are presented and no other reports have been found that would prove or disprove this generally held belief. Green {168) reported higher counts on shrimp held in contact with ice than on controls held at the same temperature without contact with ice. She ascribed this difference to the availability of water on the surfaces of the iced shrimp.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 55
Air movement
At relatively low humidities, movement of air over the surface of the meat increases the moisture gradient from the interior of the meat to the air surrounding it. As a result, the aw of the meat surface may be low enough to retard or prevent microbial growth (275). The moving air must be dry (30, 275) ; otherwise the mois- ture gradient will not be steep enough and the meat surface will not be dry. If the air is above 90 or 96 percent EH, the rate of mold or bacterial growth, respectively, is not materially altered by air movement. Scott and Vickery (í28) have pointed out that air movement speeds the chilling of meat and thereby slows bac- terial growth during the initial stages of storage. But they indi- cated that in the absence of surface desiccation, even rapid cooling is not an effective means of bacterial control.
Oxygen
The psychrophilic bacteria that spoil flesh foods grow best aero- bically. However, because some are facultative anaerobes, they can grow on foods held in the absence of air, but more slowly (JÍ5,6í,299,510).
Molds are generally strict aerobes, and in food spoilage their growth is usually limited to surfaces exposed to air. In fact, some film laminates can prevent mold growth on tightly wrapped prod- ucts by preventing the passage of oxygen (H6). However, some strains of molds have been reported that can grow with very small amounts of oxygen ; indeed a strain of Oospora lactis grew in the total absence of measurable amounts of oxygen (328).
Gas storage
Either hydrogen or nitrogen appears valueless for preserving flesh foods at low temperatures, but carbon dioxide increases keep- ing time markedly (57, 83). The mechanism of the effect of car- bon dioxide is not known. Although the pH change that it induces has been suggested (513), others (83, 183) have argued against this theory, and one author (183) suggested that the effect may be due instead to interference with a hydrogenating enzyme.
Susceptibility varies with the organism. Psychrophilic micro- organisms causing spoilage of flesh foods at low temperatures are particularly sensitive to CO2, whereas some of the mesophiles such as Proteus and Aerohacter are less so (83, 183).
It has been suggested that inhibitors are most effective when growth is slow (^26, ^98). It can thus be anticipated that CO2 will be more inhibitory near the minimum temperatures for spoil- age organisms than near the optimum. This was shown for meat spoilage molds (4-99), for Achromobacter (figs. 37 and 38), and for the flora on chicken meat (fig. 39).
Carbon dioxide is more effective in slowing bacterial growth on meats when aw is low than when it is optimal (4'24'). However, in one investigation (5i5) at temperatures of 20'' C, or above, 5 or
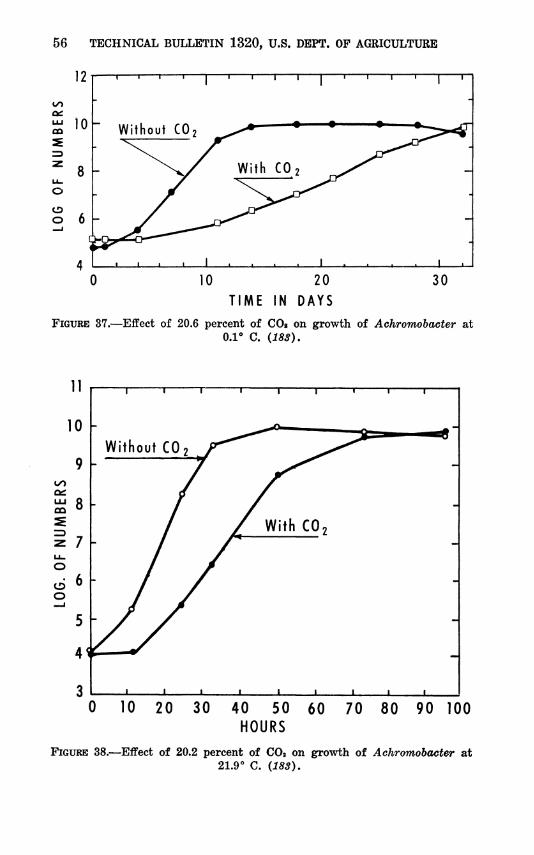
56 TECHNICAL BULLETIN 1320, U.S. DBPT. OP AGRICULTURE
10 20
TIME IN DAYS
FIGURE 37.—Elïect of 20.6 percent of CO» on growth of Achromobacter at 0.1° C. (18S).
U
10 -
CÛ
o o
T r
Without CO
■T r
-I L J L- J L.
0 10 20 30 40 50 60 70 80 90 100 HOURS
FIGURE 38.—Effect of 20.2 percent of CO2 on growth of Achromobacter at 21.9° C. (183).
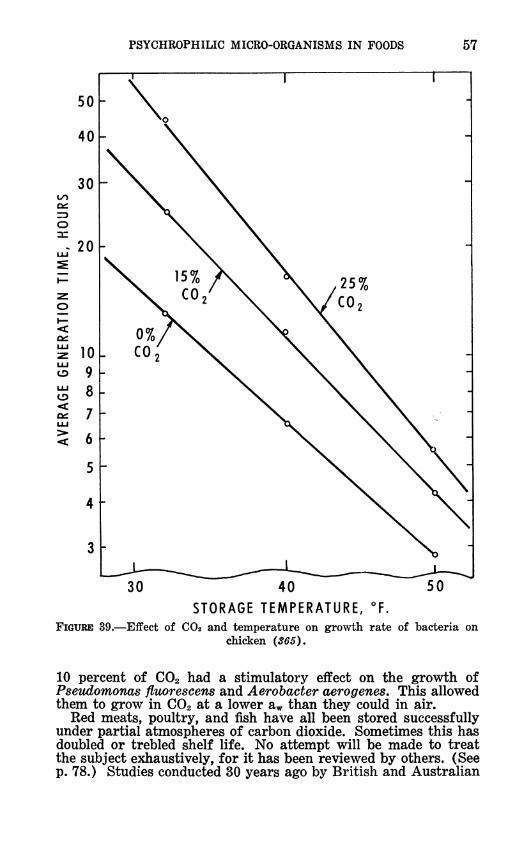
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 57
ID O re
r 20 -
<
O LU O <
STORAGE TEMPERATURE, °F. FIGURE 39.—Effect of CO2 and temperature on growth rate of bacteria on
chicken (S65).
10 percent of CO2 had a stimulatory effect on the growth of Pseudomonas fluorescens and Aerobacter aerogenes. This allowed them to grow in CO2 at a lower aw than they could in air.
Red meats, poultry, and fish have all been stored successfully under partial atmospheres of carbon dioxide. Sometimes this has doubled or trebled shelf life. No attempt will be made to treat the subject exhaustively, for it has been reviewed by others. (See p. 78.) Studies conducted 30 years ago by British and Australian
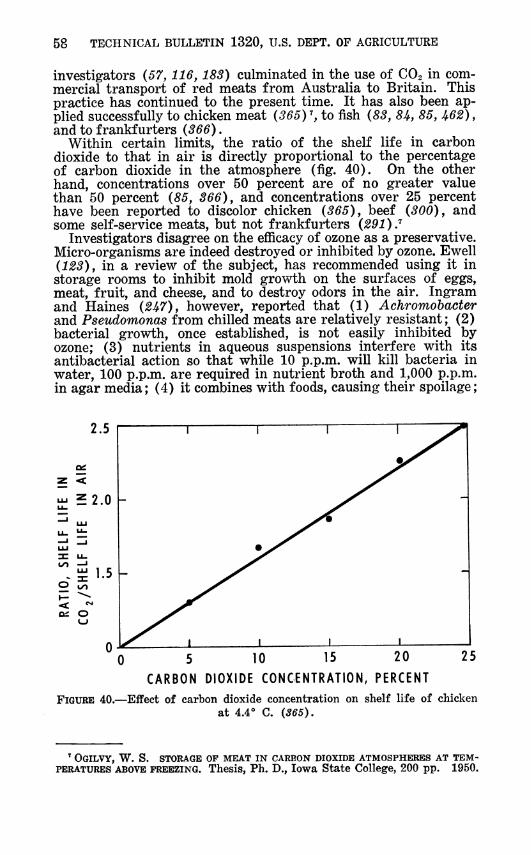
58 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
investigators (57,116,183) culminated in the use of CO2 in com- mercial transport of red meats from Australia to Britain. This practice has continued to the present time. It has also been ap- plied successfully to chicken meat (365) \ to fish (83, 8JÍ, 85, h62), and to frankfurters (366).
Within certain limits, the ratio of the shelf life in carbon dioxide to that in air is directly proportional to the percentage of carbon dioxide in the atmosphere (fig. 40). On the other hand, concentrations over 50 percent are of no greater value than 50 percent (85, 366), and concentrations over 25 percent have been reported to discolor chicken (365), beef (300), and some self-service meats, but not frankfurters (291 ) J
Investigators disagree on the efficacy of ozone as a preservative. Micro-organisms are indeed destroyed or inhibited by ozone. Ewell (123), in a review of the subject, has recommended using it in storage rooms to inhibit mold growth on the surfaces of eggs, meat, fruit, and cheese, and to destroy odors in the air. Ingram and Haines (2A7), however, reported that (1) Achromobacter and Pseudomonas from chilled meats are relatively resistant; (2) bacterial growth, once established, is not easily inhibited by ozone; (3) nutrients in aqueous suspensions interfere with its antibacterial action so that while 10 p.p.m. will kill bacteria in water, 100 p.p.m. are required in nutrient broth and 1,000 p.p.m. in agar media; (4) it combines with foods, causing their spoilage;
FIGURE 40.-
I 5 10 15 20 25 CARBON DIOXIDE CONCENTRATION, PERCENT
-Effect of carbon dioxide concentration on shelf life of chicken at 4.4^ C. (365).
' OGILVY, W. S. STORAGE OF MEAT IN CARBON DIOXIDE ATMOSPHERES AT TEM- PERATURES ABOVE FREEZING. Thesis, Ph. D., Iowa state College, 200 pp. 1950.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 59
and (5) inhibitory concentrations are higher than human beings can tolerate. Ingram and Barnes (2JÍ3) reviewed the use of ozone more critically and reported that it deteriorated rapidly in storage rooms, especially when foods were present; that low RH encour- aged such deterioration ; and that it combined with fats, causing rancidity. They concluded that the effects of relative humidity and temperature on the efficacy of ozone are unclear and require more investigation, although there is some reason to believe that low temperature increases its inhibitory action.
Thus ozone appears to have limited value in inhibiting surface microbial growth on stored foods primarily because of its toxicity to human beings and because of its enhancement of rancidity. However, storage-room air might be treated to remove odors by circulating it through a separate chamber containing ozone and then through charcoal or metal oxides to remove the ozone before it is returned to the storage area (2^3).
Packaging
The effects of humidity, air movement, oxygen, and carbon dioxide on bacterial growth are all involved in the effects of pack- aging. Halleck and others (195) found that when impermeable films were used for packaging meat, the atmosphere within the package inñuenced the growth of bacteria. The RH was high in this entrapped atmosphere, and therefore surface drying was minimized. Despite this they found that growth was marked by a longer lag phase under these films than under permeable films. Under permeable films, growth of bacteria occurred at maximum rates until dehydration or mold growth interfered. Evacuation of the atmosphere from the package wrapped in impermeable film enhanced shelf life. Many others have also found increased shelf life of meats, vegetables, poultry, and crabmeat packaged in im- permeable films or cans, especially when evacuated (8, 18, 6^, 77, 260, 291, 385, 1^55, 533).^ This effect has been attributed to re- tention of CO2 formed by respiration of either the food or the micro-organisms or both (291), or to exclusion of oxygen (77, H3,533), as shown in table 21.
Only a few authors have reported impervious packaging to be of little value for fresh meats. Results of this kind were obtained on sliced meats {30JIP) and on chicken meat {252, 315). However, better color was maintained in the sliced meats in the impervious packages. In one of the chicken meat investigations, (315) air was not evacuated from the packages and they were held at a slightly higher temperature than the unpackaged controls. In the other (252), the meat was treated with an antibiotic, which may have introduced a special factor. The organisms that grow under impermeable films are facultative anaerobes (385). In some instances their growth has been reported only slightly inhibited by low oxygen tension (299, 300).
' See footnote 7, p. 58.
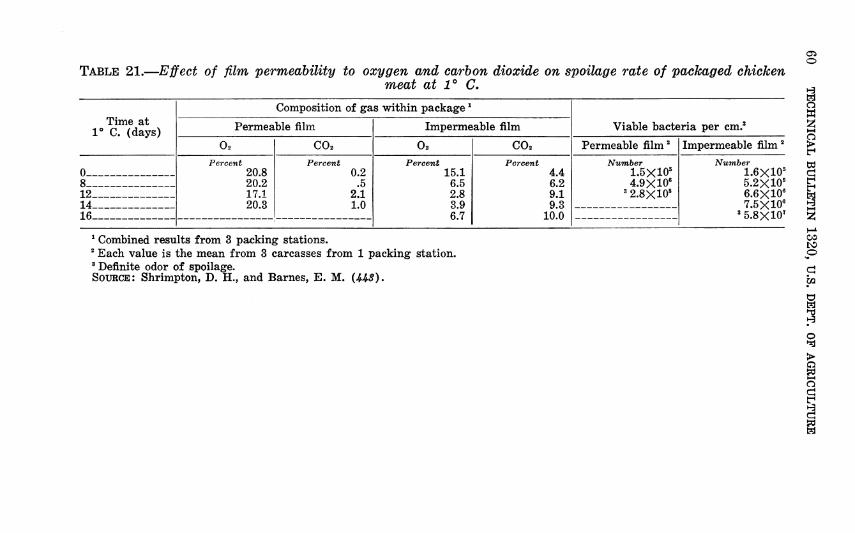
TABLE 21.—Effect of film permeability to oxygen and carbon dioxide on spoilage rate of packaged chicken meat at l"" C.
Composition of gas within package ^ Time at
1° C. (days) Permeable film Impermeable film Viable bacteria per cm.*
0. CO2 0. CO2 Permeable film ^ Impermeable film *
0 Percent
20.8 20.2 17.1 20.3
Percent 0.2
.5 2.1 1.0
Percent 15.1
6.5 2.8 8.9 6.7
Percent 4.4 6.2 9.1 9.3
10.0
Number 1.5X10^^ 4.9X10'
'2.8Xiœ
Number 1.6X10'
8 5.2X10' 12 6.6X10' 14 7.5X10' 16 ' 5.8X10'
* Combined results from 3 packing stations. ^ Each value is the mean from 3 carcasses from 1 packing station. ' Definite odor of spoilage. SOURCE: Shrimpton, D. H., and Barnes, E. M. (-4^5).
O
O a o > r« ttí
i 00 to o d
Ü
o
o o

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 61
Shelf life of cured meat is generally greatly increased by proper packaging. Prepackaged meats such as liver sausage are cooked in the skin, and the skin then protects against further contamina- tion. Films that have low permeability to water and oxygen keep sausages many weeks, whereas highly permeable films permit growth as shown in the following data obtained with liver sausage (182) :
Casing
High permeability. Low permeability
Bacteria per gram
Before storage Number
5,200 5,200
After storage at 5° C. for 78 days
Number
2,400,000 1,400
Various films or laminates have been recommended for cured meat because they inhibit growth by retaining carbon dioxide {291) or by preventing entry of oxygen {UG, 193). Vacuum- packaged cured meats finally spoil from lactic acid bacteria, whereas those permitted access to oxygen also support the growth of yeasts and molds {iSl).
Impervious films have likewise extended the shelf life of smoked fish at chill temperatures. Unfortunately, absence of the normal psychrophilic spoilage flora has permitted the growth of Clos- iridium botulinum type E in some commercial shipments of smoked fish from the Great Lakes Area, and food poisonings have occurred (509). Type E has grown at temperatures as low as 3.3° C. in laboratory studies (reviewed by Michener and Elliott (327).
Stewart (^67) and Ayres (15) have reviewed poultry pack- aging. Gross (169) has reviewed shelf-life problems of packaged foods in general, and Hannan (198) and Ingram (2i2) have re- viewed microbiological aspects of packaged foods.
Acid
Psychrophilic food spoilage bacteria do not grow readily at low pH. Fish spoilage has been inhibited by using acetic or tartaric acids to obtain pH 6 (4^20), or hydrochloric acid to obtain pH 5 (3i9). Spoilage of crabmeat has been inhibited by using phos- phoric acid to obtain pH 5 to 6 (^96), and growth of bacteria in minced beef was slowed by using hydrochloric acid to obtain pH 4J (i9). In two reviews, Ingram (23i, 2A1) has described the eftect of pH on bacteria, particularly as applied to meat spoilage. Witter (5Í1) has reviewed the effect of low pH in inhibiting psychrophilic spoilage of dairy products. The effect of acids in preserving various cured and fermented foods is well known but is outside the scope of this review.
Antemortem treatment
Meat from a hog properly rested and fed before slaughter keeps well, whereas that from a febrile or overfatigued hog de- composes rapidly. The tired animal bleeds less well, and in addi-

62 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
tion contains less muscle glycogen and goes into and out of rigor more rapidly. The pH of muscles containing high quantities of glycogen falls farther to the acid side than does that of muscles containing less glycogen. These factors have been discussed in more detail by various authors {155,186, 23i, 2U, 318).
The situation is similar in other mammals; however, beef is somewhat less susceptible to such differences than is pork (23i). Brown and others (^P) found that when rigor was slower in onset, beef kept better, but the effect was separate from that of pH.
On the other hand, Koonz {289) reported that withholding feed before slaughter of poultry had no effect on subsequent spoilage rate. Thus, there is no advantage in feeding birds before slaughter. In fact, an empty intestinal tract makes for better sanitation during poultry evisceration.
Fish that have been feeding heavily before they are removed from the water will keep less well than those that have empty stomachs. This is because the intestines of a fish full of feed contain many bacteria and in addition the digestive enzymes of the fish are present in high concentration. On the other hand, a fish that has been fasting has few if any bacteria in its intestines {323). For this reason, ''feedy" fish are often kept alive in captivity until their stomachs are empty {216).
Fish that are physically exhausted when they die will go into and out of rigor more rapidly than otherwise, and the pH will re- main low a shorter time. Thus, bacterial decomposition is most rapid in gill-net caught fish, somewhat less so in line-caught fish, and least in seined or trapped fish {90,216).
Autolysis
Before bacteria begin to grow on fish or beef muscle, an auto- lytic breakdown to peptides and amino acids occurs. These simple compounds are not only the first products used by bacteria grow- ing on fish {15i)y but they actually are required before bacteria can build enzymes to attack pure proteins {178). However, the amount of autolytic material present does not affect growth rate on fish tissue because once the proteins are attacked, a sufficient amount of simple nitrogenous material is immediately available {282).
Antibiotics
This review will be limited to a few interesting developments in the field of antibiotic preservation of food. More extensive re- views are listed on page 78.
Some of the antibiotics, primarily chlortetracycline (CTC), en- hance the shelf life of chilled meats, poultry, and fish to a re- markable degree. The effectiveness of antibiotics is greatly en- hanced at low temperatures—^more so than the additive effects of the antibiotic and low temperature would be separately (fig. 41).
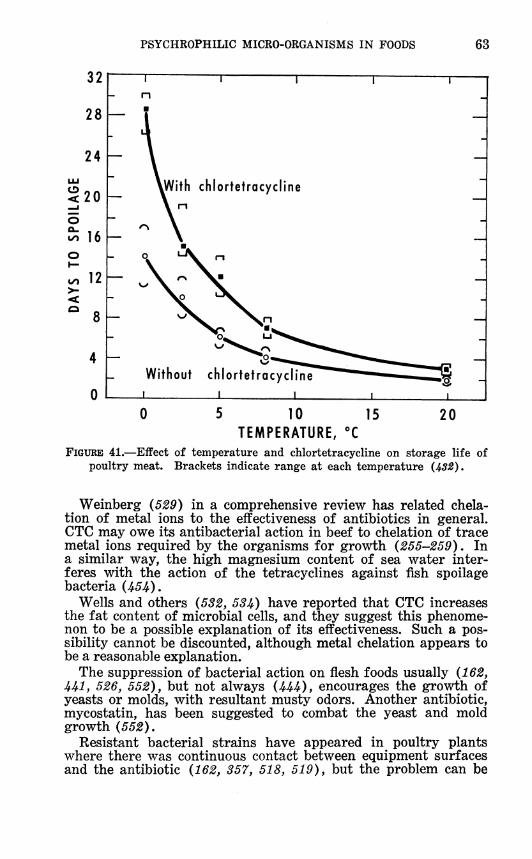
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 63
32
28
24
ÍS20
o ^ 16 o
!l 12 >- < " 8
With chlortetracycline
Without chlortetracy
J I
0 5 10 15 20 TEMPERATURE, "C
FIGURE 41.—Effect of temperature and chlortetracycline on storage life of poultry meat. Brackets indicate range at each temperature (4S2),
Weinberg (529) in a comprehensive review has related chela- tion of metal ions to the effectiveness of antibiotics in general. CTC may owe its antibacterial action in beef to chelation of trace metal ions required by the organisms for growth (255-259). In a similar way, the high magnesium content of sea water inter- feres with the action of the tetracyclines against fish spoilage bacteria (^5^).
Wells and others (532, 5SJf) have reported that CTC increases the fat content of microbial cells, and they suggest this phenome- non to be a possible explanation of its effectiveness. Such a pos- sibility cannot be discounted, although metal chelation appears to be a reasonable explanation.
The suppression of bacterial action on ñesh foods usually (162, hhl, 526, 552), but not always (iM), encourages the growth of yeasts or molds, with resultant musty odors. Another antibiotic, mycostatin, has been suggested to combat the yeast and mold growth (552).
Resistant bacterial strains have appeared in poultry plants where there was continuous contact between equipment surfaces and the antibiotic (162, 257, 518, 519), but the problem can be

64 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
controlled by chlorînation » and other stringent hygienic precau- tions nos). On the other hand, chlorine will destroy CTC if it is applied simultaneously {66). Highly resistant strains develop by a process of selection of the more resistant individuals {1, 489). These are primarily Achromobacter and the less sensitive pig- mented pseudomonads H93).
CTC has been reported to retard the growth of Staphylococcus aureus, Clostridium botuUnum, and spoilage organisms in fish at 10° C. (39). However, one investigation (363) revealed large numbers of the pathogenic yeast Candida parapsilosis in CTC- treated chicken meat; other reports (220, 316) described anti- biotic-resistant strains of Salmonella typhimurium, one of which grew on carcasses of chickens in the absence of the normal spoil- age that would have rendered them inedible (220). Another in- vestigation (i89) failed to confirm that CTC-treated poultry meat represented a health hazard from yeasts or bacteria; however, failure to isolate a given organism does not constitute proof of safety.
The commercial use of antibiotics to preserve fish and poultry has decreased in recent years. One reason is that some producers and handlers have tried to use the antibiotics as a substitute for good sanitation, refrigeration, and rapid handling. If antibiotics are to prove of value, all of these other practices should be con- tinued at the same high level in order to place a product of high quality in the hands of the consumer. In addition, some con- sumers have not been convinced of the safety and usefulness of such additives despite U.S. Food and Drug Administration ap- proval (508).
Salt Psychrophilic bacteria are sensitive to sodium chloride. The
organisms capable of growth in high salt concentrations are rarely able to grow below 5** C. (215, 239, 327, U83, 503). Salted foods are outside the scope of this review. However, small amounts of salt on the surfaces of flesh foods can inhibit bacterial growth by creating a microclimate of moderately high salt content without changing the nature of the foods or contributing enough saltiness to make them unpalatable.
Salt brine dips with acid (153) or without acid (31) have been suggested for fish. Shrimp at —1.1° C. in 6-percent brine re- mained in good condition longer than that in 3-percent brine {U29). Salt brine dips are used regularly for crabmeat and re- duce bacterial loads (63).
Trachéal injection of salt brine was suggested to preserve chickens in areas of the U.S.S.R. where refrigeration was unavail- able (512), and a carageenin gel coating containing 6 percent of salt enhanced the keeping time of frozen-thawed chicken (37U).
• VAUGHN, R. H. Private communication.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 65
However, salt has been only partially successful in enhancing preservation of dairy foods, for many psychrophiles resistant to 4 to 6 percent of salt have been found in such products (5^1 ) •
SPOILAGE OF EGGS
Penetration of shell eggs Only part of the available information on bacterial decomposi-
tion of eggs in the shell will be presented. We refer to the im- portant reviews listed on page 78.
Lorenz and others (308) have quoted nine authors who agree that spoilage organisms are very rarely inside the egg before it is laid. Even the outside of the egg shell is free of bacteria as it leaves the oviduct, but bacterial contamination occurs after the egg is laid U75). Eggs that are obviously dirty, usually with fecal material from the nest, have been found to contain more bacteria after a short period than have obviously clean eggs (139) and in addition they spoil more readily in storage (4'06, 506).
To cause spoilage of the interior, micro-organisms must pene- trate the shell and membranes and overcome the natural inhibi- tory materials within the albumen. The shell contains several hun- dred small holes filled with mucin, which effectively prevents bacterial entry when it is clean and dry (JfOl ). However, abrasion or moisture may remove this mucin, whereupon the shell offers little resistance to the passage of bacteria (137, 138, 151, 156). There is still a question whether shell porosity, which varies with the hen, does (293) or does not (3i2) affect the penetration rate. There is no correlation between shell thickness and permeability to bacteria (138).
The outer shell membrane likewise offers little resistance to bac- terial penetration, but the intact inner membrane is an effective physical barrier to bacteria (110, 151, 292, 321, U75, 523) so that penetration to the albumen is delayed hours (185, 523) or even days (U7, 110, 151, 333). Colonies may grow on the surface of this membrane before the bacteria break through to the albumen (110,156). Florian and Trussell (129) have listed organisms ac- cording to their capabilities of penetration of the inner membrane as "primary" or "secondary" invaders.
Although some investigators (i75) have reported that the mem- branes have an antibacterial activity, subsequent work has shown this to be incorrect (^74). In fact, membranes may actually sup- port bacterial growth (i7,108,151,161).
Wet eggs are much more susceptible to bacterial invasion than are dry eggs (185, 190, 321). Thus, wetwashing enhances spoil- age more than does dry abrasion (505). Similarly, removing cold eggs to a warm, moist atmosphere may result in condensation (sweating) on the shells, which encourages surface growth and entry of bacteria, especially if the eggs are again chilled (131,

66 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
187, 139, 537). High humidity in storage rooms also encourages growth of molds on the shell ll88, i33) (table 22) and enhances penetration of bacteria (3^2). If eggs must be wetwashed, they should then be dried thoroughly {il, 3H).
TABLE 22.—Effect of temperature and humidity on the growth of mold on eggs as judged by eye
Humidity (percent) 0° C. 10° C.
DAYS FOR VERY SLIGHT GROWTH
90 78 24 18 14
17 95 11 98_ 8 100 5
DAYS FOR DISTINCT GROWTH
95 41 29 23
16 98 13 100 9
DAYS FOR HEAVY GROWTH
98 41 35
17 100 - _ 11
SOURCE: Haines, R. B. (188).
A temperature drop, particularly when the shell is wet, may en- courage penetration by pulling bacteria through the shell. This may occur when the egg drops into a dirty or wet nest, when the warm egg is washed in cool water (table 23), or when storage temperature changes during handling. As a result of work such as this, the use of wash water warmer than the egg has been recommended by many investigators and is now commercial prac- tice Ul, 131, Jfl8). Eggs spoil faster when stored at high tem- peratures than when stored at low temperatures {188, 5^7) (table 22).
TABLE 23.—Effect of relative temperature of egg and wash water on rotting during subsequent storage
Treatment ^ Rots
Water and eggs same temperature Eggs, 10^ C; water, 25° C Eggs, 37° C; water, room temperature Immersed 1 hour immediately after being laid Cooled 1 hour, then immersed 1 hour Cooled 24 hours, then immersed 1 hour
* Between 27 and 119 eggs were used for each test. ~ Average of 3 tests.
SOURCE: Haines, R. B. (189).
Percent 13
7 100
'100 *79 '19

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 67
Despite the dangers of washing, most eggs are now wetwashed because the trade prefers a clean product. In many operations all eggs are washed to avoid the need for sorting {Jfl). Washing of dirty eggs is advisable, provided they are washed before bacteria have had time to enter and provided care is taken in the washing procedure {ISl). Funk (i^^) and Winter and others {5S9) have reviewed the use of detergents and detergent-sanitizers and have concluded that they have little value in preventing subsequent spoilage. However, detergent-sanitizers containing a quaternary ammonium compound give Vs to i^ the amount of spoilage as do those without (¿^, S29, SSI, SS2). Iodine in concentrations of 50 to 100 p.p.m. is an effective sanitizer when used following a non- sanitizer detergent (ilS). Antibiotics are valueless in preventing spoilage (82, SSO), and may even increase spoilage {U18). (See also **EfFect of metals on penetration and growth.'')
Shell coatings
Oiling retards evaporation of eg^ contents, but there is still a question as to whether it retards (US), has no effect on (i06), or enhances (52, 281) spoilage of shell eggs.
Thermostabilization
Thermostabilization—a dip in hot oil or hot water long enough to coagulate a thin layer of albumen — retards bacterial entry {125,126, lUy US, U5, HO). This subject has been reviewed by Funk (lU).
Gas storage
Carbon dioxide storage inhibits spoilage organisms on shell eggs as it does on meats and fish (187, 190, SS6) and prevents mold growth as shown in figure 42.
Ozone, at a concentration of 1.5 to 3 p.p.m., successfully pre- vents mold growth on the exterior of the shells of eggs but is in- effective in preventing internal spoilage (122,186, S19).
Internal defenses
Once penetration is achieved, the egg still has natural defenses in the albumen. Garibaldi (lí7) has listed and discussed the factors in albumen that have been reported to inhibit growth: High pH, lysozyme, avidin, ovomucoid, conalbumin, and the pres- ence of only native proteins. Gram-positive bacteria are sensitive to lysozyme, and bacteria requiring biotin or iron are inhibited by the binding action of avidin and conalbumin, respectively. Ovomucoid is an antitrypsin factor, but it does not affect growth of gram-negative bacteria. Garibaldi (1Jí7) and, later. Brooks (Jf7) questioned the inhibitory effect of high pH, per se, explain- ing that the pH effect may be due instead to the greater power
68 TECHNICAL BULLETIN 1320, U.S. DEPT. OP AGRICULTURE
90
j_
• Just perceptible
D Slight
o Marked
X X
O 20 40 60 80 100 CARBON DIOXIDE IN THE ATMOSPHERE, PERCENT
FIGURE 42.—Effect of carbon dioxide on growth of mold on shell eggs at 10** C. over water {3S6).
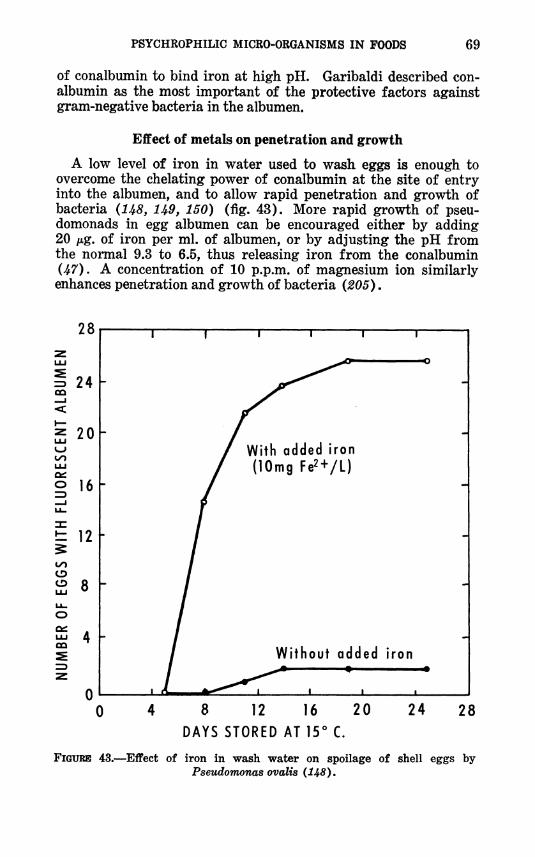
PSYCHROPHILIC MICRO-ORGANISMS IN POODS 69
of conalbumin to bind iron at high pH. Garibaldi described con- albumin as the most important of the protective factors against gram-negative bacteria in the albumen.
EiSiect of metals on penetration and growth
A low level of iron in water used to wash eggs is enough to overcome the chelating power of conalbumin at the site of entry into the albumen, and to allow rapid penetration and growth of bacteria (1^8, HO, 150) (fig. 43). More rapid growth of pseu- domonads in egg albumen can be encouraged either by adding 20 ii,g, of iron per ml. of albumen, or by adjusting the pH from the normal 9,3 to 6.5, thus releasing iron from the conalbumin (^7). A concentration of 10 p.p.m. of magnesium ion similarly enhances penetration and growth of bacteria {205).
28
GQ
<:
24 -
20 -
Oil
? 161-
!= 12
O o 8 - III ^^
o
4 -
With added iron (lOmg Fe2+/L)
Without added iron • 9
0 4 8 12 16 20 24 28
DAYS STORED AT 15° C.
FIGURE 43.—Effect of iron in wash water on spoilage of shell eggs by Pseudomonas ovalis (14S).
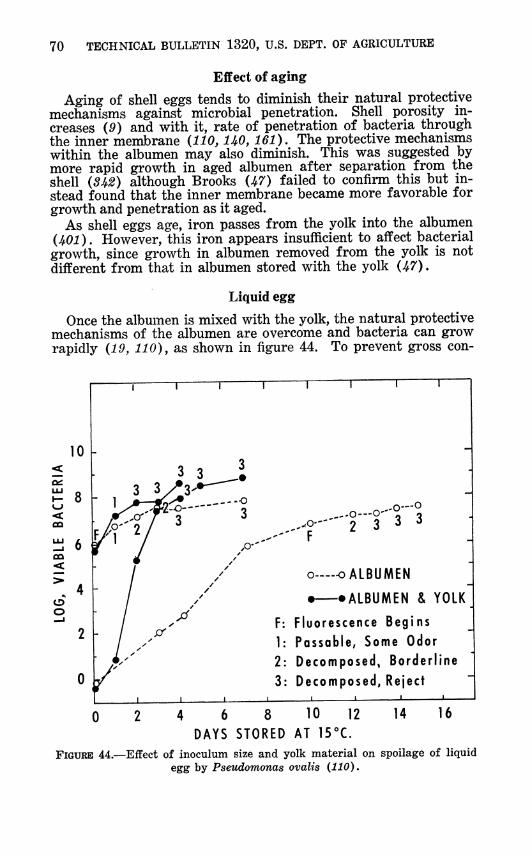
70 TECHNICAL BULLETIN 1320, U.S. DEFT. OF AGRICULTURE
Effect of aging Aging of shell eggs tends to diminish their natural protective
mechanisms against microbial penetration. Shell porosity in- creases (9) and with it, rate of penetration of bacteria through the inner membrane (110, UO, 161). The protective mechanisms within the albumen may also diminish. This was suggested by more rapid growth in aged albumen after separation from the shell {SJÍ2) although Brooks (^7) failed to confirm this but in- stead found that the inner membrane became more favorable for growth and penetration as it aged.
As shell eggs age, iron passes from the yolk into the albumen {JfOl). However, this iron appears insufficient to affect bacterial growth, since growth in albumen removed from the yolk is not different from that in albumen stored with the yolk {A7).
Liquid egg
Once the albumen is mixed with the yolk, the natural protective mechanisms of the albumen are overcome and bacteria can grow rapidly (19, 110), as shown in figure 44. To prevent gross con-
-'-2-r 3 3 -°' 2 3
o—.o ALBUMEN
• »ALBUMEN & YOLK
F: Fluorescence Begins
1: Passable, Some Odor 2: Decomposed, Borderline
3: Decomposed, Reject
0 2 4 6 8 10 12 14 16
DAYS STORED AT IS^C. FIGURE 44.—Effect of inoculum size and yolk material on spoilage of liquid
egg by Pseudomonas ovalis (1X0).

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 71
tamination by micro-organisms, shell eggs should be washed {2Í6, 380, 555) or even pasteurized {HI) before they are broken for liquid Qgg. Also, the magma should be refrigerated quickly {SOI, 802, 555) to prevent growth, particularly if it has been previously subjected to heavy contamination (U9). Even in a well-managed plant, the average total count in liquid egg before freezing or drying often exceeds 10^ per gram, because of the good medium it offers to bacteria {24'6). The liquid egg magma may also be pasteurized to reduce the bacterial count and thereby enhance keeping quality and kill pathogens (2i6, SJÍ7, U79, 5U0).
Similarly, frozen eggs should be defrosted rapidly to prevent bacterial multiplication (53, 276, 390). Liquid egg has a low oxi- dation-reduction potential during spoilage (111) so that only obligate and facultative anaerobes can grow in it, except in sur- face layers.
SPOILAGE OF FRUITS
Spoilage of fresh fruits stored at chill temperatures without other processing is outside the scope of this review and has been reviewed elsewhere (see p.79). Frozen fruits are normally stored at temperatures too low to permit growth, so that microbial spoil- age can occur only before or after storage.
Spoilage of fruits is usually by molds or yeasts, rarely by bac- teria, for the pH of fruits is generally sufRciently low to prevent growth of all bacteria except the lactobacilli. However, the edible organic acids do not owe their antibacterial action solely to their low pH; the anion, too, has antibacterial activity (117). Sugar is usually added before freezing; and the yeasts and molds, which are also more tolerant to high osmotic pressure than are bacteria (239), are again favored. Fermentation by yeasts is a common problem in fruits during delays before freezing.
Before freezing has begun, the added sugar in fruit is a good preservative because of its dehydrating action (37). After freez- ing has started, the sugar may have a protective action but will not affect the a^ of the system (327). The preserving and germi- cidal action of various sugars against bacteria is in the following order of decreasing efficacy: lévulose, dextrose, sucrose, lactose. Yeasts are considerably more tolerant of sugars than are bacteria (117).
Interpretation of the effect of sugar is complicated by the fact that a high sugar content increases the rate of heat transfer, probably because the mass becomes liquid (100, 272). This re- duces the time during which the product is at a temperature favorable for microbial growth.
Early investigators of frozen fruit discovered that molds do not grow if oxygen is excluded (35, 98,100, 820). Therefore, they do not grow in the interior of barrels of frozen berries nor in sealed containers of fruit. Sealed containers of berries did not spoil below —2.2° C. because molds were inhibited by lack of oxygen, bacteria by the acid, and yeasts by the combined effect

72 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
of these with low temperature (35). However, organisms grew in berries at —2° and —4° when the package was not airtight (Si).
Fruits may become contaminated with micro-organisms while still on the plant ; and they may start to break down even before harvest. Breakdown, often due to the same micro-organisms, con- tinues during storage at a rate dependent on temperature (206, JÍ05). If fruits or purees are to be frozen in large containers such as barrels, it is important that the product be cooled to 10° or lower before it is placed in the container, to prevent growth dur- ing the slow freezing period (SOU).
Among the yeasts that grew on grapejuice at low temperatures, Lawrence and others (296) isolated strains that grew well at 0° C, poorly at 17°, and had optima between 7° and 13°. These same strains had maximum growth temperatures below 20° when tested on a temperature gradient incubator (112).
Growth of these yeasts on grapejuice at —2.2° to —5.5° C. can be suppressed by adding sorbic, benzoic, capric, or caprylic acid in concentrations of 0.01 to 0.015 percent, although these levels are too low to prevent growth of these yeasts at room temperature (376).
Microbiological spoilage of citrus juice and juice concentrate has not been reported at subzero temperatures. Berry and others (38) reported no growth of Leuconostoc and Lactobacillus species at 4.4° C. in orange concentrate in the range of 12° to 58° Brix, although these organisms grew in orange concentrate at higher temperatures. These authors observed growth of yeast at 4.4° in 42° Brix concentrate, and Rushing and others (i08) reported growth of yeast at 1.7° in concentrate of 40° to 55° Brix. Micro- bial populations (largely yeasts) in commercial orange concen- trate decreased at 4.4° until the third week of storage, after which they rose rapidly (3i8). Wolford (5JÍ6) found no growth in orange concentrates at 4.4° in 187 days. The microbiology of citrus juices and juice concentrates has been reviewed by Patrick and Hill (373) and that of noncitrus juices by Liithi (309).
Other than juices, few processed fruit products are stored at chill temperatures, but Rushing and Senn (^09) stored citrus salad at temperatures down to -—1.1° C. At this temperature, an initial drop in total count of about 80 percent during the first 25 days was followed by growth. This behavior resembled the be- havior of microbial populations on vegetables at similar tempera- tures (fig. 45). Growth was prevented in the salad by potassium sórbate at 0.066 percent but not at 0.033 percent, while sodium benzoate was ineffective at 0.066 percent. These chemicals were thus less effective than in grapejuice at low temperatures (376).
SPOILAGE OF VEGETABLES
The parasitic micro-organisms that damage fresh vegetables are generally capable of attacking living tissue and are considered plant pathogens. This subject has been reviewed elsewhere (p. 79) and is outside the scope of this review.
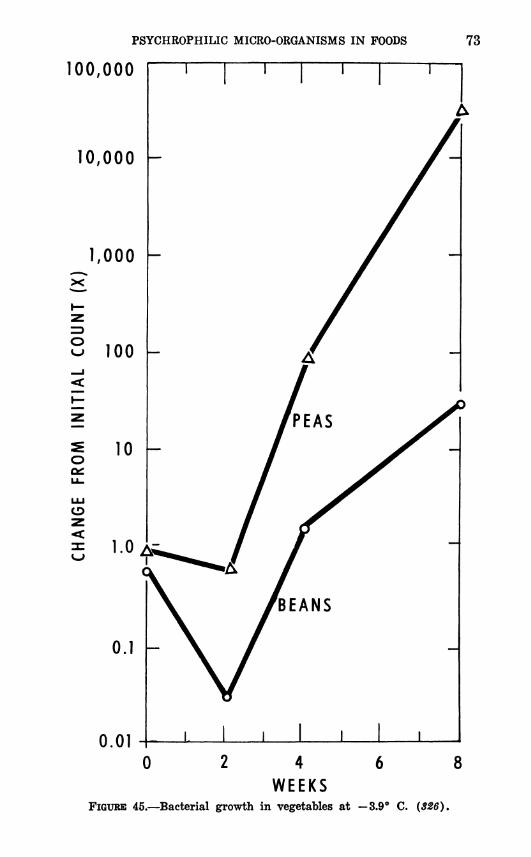
PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 73
100,000
10,000 -
1,000
s 100
o <
0.01 4
WEEKS FIGURE 45.—Bacterial growth in vegetables at —3.9® C.

74 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
Vegetables become contaminated with saprophytic micro-or- ganisms from the soil and from harvesting operations. Some contain small to moderate numbers of bacteria within the tissues {3U5,ill),
However, frozen vegetables differ from most other frozen foods in that they are blanched during processing. Blanching greatly reduces the microbial population {86, 80, 268, 375, UU8, 520, 521). The vegetables before processing have a natural resistance to de- cay (78) which disappears after blanching {36, 99, 268, 375, U9) or after freezing {51 ), presumably as a result of heat killing of the plant tissue with accompanying increase in permeability and leakage of nutrients from the plant cells. Recontamination and growth occur during subsequent processing, and the product may enter the freezer with a large and rapidly growing bacterial popu- lation {268, 375, 4,60, 461, 520, 521 ).
Since blanched vegetables are very susceptible to microbial spoilage, they are always stored at temperatures intended to pre- vent microbial growth. Most of the available information on the microbiology of frozen vegetables, therefore, merely gives the minimum temperature at which microbial growth has been ob- served under particular circumstances {327), or describes the survival of micro-organisms at temperatures too low for growth.
There are a few reports on the microbiology of frozen or thawed vegetables in the bacterial growth range. Growth on vegetables has been reported occasionally down to about —-8** C. and once at —12** {327). Berry {34) found that counts on vegetables rose to over 10^ per gram in 2 months at —4°. Michener and others {326) stored several lots of peas, green beans, cauliflower, and spinach at —3.9°, —1.2°, and +4.5°, and found growth within 8 weeks at the first two temperatures but within 1 week at the third. As shown in figure 45, growth was often preceded by an initial drop in count, presumably caused by death of some species while the others were still in their lag phase. The population of one lot of peas stored at —3.9° reached 10^ per gram (fig. 45). Similarly, Hucker and David {225) incubated frozen-thawed vegetables at +7°, and obtained counts after 70 hours of 10^^ per gram (spin- ach), 10^ per gram (peas), and 10^ to 10^ per gram (corn, lima beans, string beans).
The natural microflora of frozen vegetables grows slowly near the freezing point (figs. 26 and 45). The advantage of permitting frozen vegetables to thaw at low temperatures is evident from the time required for growth to begin. However, these platings were incubated at 30° and 32° C, respectively. Platings at lower temperatures might have shown a somewhat shorter lag time {114,381).
Hucker and David {225) followed the total count of the nat- urally occurring microflora on several frozen vegetables during five successive freeze-thaw cycles but failed to find significant changes. The thaw phase of each cycle lasted 8 hours at 2° or 7° C, which was less than the lag period at these temperatures. While the count varied more at 7° than at 2°, the variations were irregular and presumably were sampling differences.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 75
The process of freezing vegetables may break up clumps of bacteria, thus causing an apparent increase in the number of bacteria {226, SH, 375). In one case these bacterial clumps were observed microscopically before freezing but were not seen in the same sample after freezing {875). When peas or corn were frozen slowly so that they remained for 10 hours or more at their freez- ing point, bacterial counts increased up to threefold after the product reached this temperature {516). Although these authors considered this a result of breaking up of bacterial clumps, Michener and others {326) found that sampling variations of this magnitude are not unusual.
The microbial flora of frozen vegetables is for the most part that which contaminates them after blanching and survives freez- ing. The common types have been listed (^^7-^5i). During spoil- age of previously frozen vegetables at 10 "" to 32° C, initial sour- ing by streptococci was followed by growth of lactobacilli, al- though these groups originally made up only about 5 percent of the flora {U12). At 30**, Streptococcus lactis and Leuconostoc mesenteroides predominated in spoilage of peas {535).
When peas were held at —4° C, they were spoiled by Pseu- domonas fluorescens and a Lactobacilhts, while at —6.7'' they were below the growth range of their bacterial flora and were spoiled by Sporotrichum {37). In the same series of experiments, okra and kale at —6.7° were spoiled by Cladosporium. At 0°, a gram-negative organism resembling Flavobacterium predominated in spoilage of frozen-thawed vegetables {222).
Michener and others {326) failed to find any correlation be- tween microbial growth and development of off-flavors in vege- tables. During storage at —3.9°, —1.2°, and +4.5° C, off-flavors developed before the bacterial count had doubled, and the onset of off-flavor was not correlated with the initial bacterial count. It was concluded that the onset of off-flavor was not due to bacterial growth, although bacterial enzymes could be involved. Further- more, off-flavor developed at a measurable rate even at and below —18° {Jf3i) although bacterial growth is prevented by these tem- peratures.
Prepeeled potatoes are especially subject to microbial deteriora- tion. Although they are not cooked, they are frequently heat- or lye-peeled and treated with sodium bisulfite or other agents to prevent darkening. The cells on the surfaces are killed, and are therefore subject to rapid attack by saprophytic micro-organisms during storage at 0° to 5° C. The methods of processing pre- peeled potatoes have been described {166, 201). The relation be- tween temperature and shelf life of prepeeled potatoes was investi- gated by Ceponis and Friedman (70). They found that the peeled potatoes had to be cooled in water before they were bagged. Other- wise they took 30 to 36 hours to cool to refrigeration temperature. Total counts after processing were 6,000 to 51,000 per gram and included bacteria, yeasts, and sometimes molds. The shelf life at 2° was 8 to 11 days for whole potatoes, and 9 to 13 days for cut strips. Microbial spoilage causing off-odors and flavors was the principal factor limiting shelf life. Bisulfite inhibited microbial

76 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
growth, but its concentrator! was very low near the end of shelf life. Greig and Smith (166) were unable to extend shelf life by- treatments with calcium or sodium propionate or chlortetracy- cline.
Kohman (288) recommended "Frigi-Canning'' of vegetables— a process using only enough heat to kill vegetative cells followed by refrigeration above freezing. Refrigeration would be con- trolled at a temperature low enough to prevent growth of Clos- iridium botuUnum and other food-poisoning organisms. The psychrophiles that could grow are sensitive to heat. This proce- dure, however, has the serious disadvantage that cans accidentally stored at a higher temperature may become hazardous.
SUMMARY AND CONCLUSIONS
An understanding of factors affecting microbial growth at low temperatures will assist the food industry in minimizing spoilage losses in chilled foods. Microbial growth is not a problem in frozen foods unless they are mishandled by being held above -12^ C.
Lag period of microbial growth is shortest at the optimum temperature and becomes progressively longer and approaches infinity as the temperature is lowered. For this reason, growth on frozen foods stored at high freezer temperatures may become evi- dent only after very long periods. Most investigators limit their observations of lag time to a few weeks, but one worker found that Oospora began growth at —8° C. after a 414-day lag. Lag periods vary widely with the species and strain of organism. Psychrophiles have a shorter lag period than mesophiles in the temperature range where both grow.
Growth rate during the logarithmic phase is similarly sensitive to temperature. The growth rate of mesophiles increases faster with temperature rise than does that of psychrophiles.
The temperature coefficient (Qw) and the mathematically re- lated temperature characteristic (ju.) of microbial growth and ac- tivity increase more than one would expect in simple chemical reactions when the temperature approaches the lower limits for growth. For this reason the storage life of chilled foods can be doubled or trebled by keeping them near the freezing point in- stead of at 5^ to 10^ C. where they are often held.
Bacteria often produce extracellular enzymes in greater quantity at temperatures below their optimum than at the optimum or above it. However, the enzymes themselves may alter the sub- strate most rapidly above the optimum or even above the maxi- mum temperature for growth of the organism. A few investiga- tors have reported that some foods held at temperatures near 0** C. spoiled before counts became high. This may have been due to high enzyme production at low temperatures. But usually counts are 10^ or 10^ per gram when spoilage is evident. Microbial enzymes are active below the minimum growth temperature. This is one reason for good sanitation and rapid chilling, which prevent high bacterial levels in frozen foods.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 77
The optimum temperature for cell crop is usually lower than that for growth rate. This can be explained by the greater solu- bility of oxygen at low temperatures. The temperature range of growth is narrow when conditions are adverse but widens when conditions are otherwise satisfactory. Sufficient adaptation to permit rapid growth at low temperatures has been accomplished to only a limited extent. We have found no report of a successful attempt to adapt a mesophile to grow below its normal minimum growth temperature.
A high level of initial contamination results in more rapid spoilage of foods than does a low level, when the foods are held at temperatures suitable for growth. At chill temperatures or below only the level of psychrophiles is significant in this regard.
Frozen-thawed foods spoil at about the same rate as do foods not frozen.
Micro-organisms require high water activity for growth. They must compete with solutes and colloidal water-binding compounds for the water content of the food. Bacteria require more water than do yeasts, which in turn require more than molds.
During spoilage of products of warm-blooded animals at low temperatures, floral changes occur in favor of psychrophiles, which are present originally only as a minority group. A less distinct change occurs in fish products because the initial flora is predominantly psychrophilic, at least in temperate and arctic zones.
Spoilage of meats and fish is primarily a surface phenomenon. Growth is much more rapid on cut surfaces than on skin, fat, or connective tissue.
Psychrophilic spoilage of meats is commonly prevented or re- duced by storage in circulating dry air to dry the surfaces on which spoilage occurs. However, too much surface drying may result in undesirable weight loss.
Psychrophiles are predominantly aerobic, but some are faculta- tive anaerobes, so that exclusion of air slows but does not stop spoilage. Storage in atmospheres containing carbon dioxide is eminently successful in increasing shelf life of chilled flesh foods. Packaging with impermeable films inhibits surface growth of spoilage organisms somewhat by limiting access to oxygen and by allowing an increased carbon dioxide content within the en- trapped atmosphere, but this effect is limited because the humidity is high.
Psychrophilic spoilage bacteria are inhibited by low pH, and thus by the conditions in red meats from animals properly fed and rested before slaughter. These bacteria are sensitive to heat and to moderately high concentrations of salt.
Antibiotics, primarily the tetracyclines, greatly increase keep- ing time of flesh foods, especially at low chill temperatures. This may be the effect of chelation of trace metals essential to bacterial growth. Sometimes the inhibition of bacterial growth allows yeasts or molds to grow, and frequently the use of an antibiotic is followed by appearance of a resistant microbial population.

78 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
Eggs are usually sterile when laid. Dirt, moisture, and drop in temperature encourage microbial penetration. Penetration can be inhibited by good sanitation, by washing with warm water (when washing is necessary), by thermostabilization, or by stor- age in carbon dioxide or ozone. The natural defenses of the albu- men are high pH, lysozyme, avidin, ovomucoid, and conalbumin. Iron in wash water enhances microbial growth in the albumen by saturating the chelating ability of conalbumin at the site of entry. Aging reduces the effectiveness of the natural protective mechanisms of shell eggs. Once the yolk is mixed v/ith the albu- men, as in liquid egg, the protective mechanisms of the albumen are neutralized and growth is rapid; therefore, eggs for breakmg should be clean, and the liquid egg should be refrigerated quickly.
Plant pathogens and saprophytes from the field are responsible for rotting of fresh fruits and vegetables. The microñora of fruits consists primarily of yeasts and molds, because the low pH inhibits bacterial growth. Sugar inhibits growth before freezing. Like- wise anaerobic conditions within impervious packages or in the centers of large containers inhibit mold growth.
Blanching of vegetables greatly reduces their microbial popula- tion, but it also removes their natural resistance to decay so that very rapid bacterial growth occurs on recontamination before freezing. Part of the resulting ñora survives freezing and can grow to the point of spoilage at temperatures at or above —4° C. If spoilage occurs at lower temperatures, it is usually caused by molds. For prepeeled potatoes, which are refrigerated but not frozen, shelf life is about 13 days at 2° C. and is limited by micro- bial spoilage.
REVIEWS AND BIBLIOGRAPHIES ON LOW-TEMPERATURE MICROBIOLOGY
General Reviews.—Psychrophilic bacteria: 1959 (2S2) ; 1960 U7S) ; 1961 (5il) ; 1962 U02). Effect of cold on food micro-^ organisms: 1944 U80) ; 1951 (236); 1955 UO) ; 1962 (61). Food spoilage and the microecology: 1949 (sll) ; 1952 (339) ; 1955 (79, 340); 1959 (il5). Temperature and life: 1935 (32) • 1955 (388).
Bibliographies.—£7#eci of cold: 1937 (253) ; 1946 (528) ; 1947 (17Í); 1948 (10, 173, 175) ; 1961 (476). Fish: 1946 (109) 1949 (56,89).
Reviews of specific ñelds.—Antibiotics : 1953 (482) ; 1954 (531) 1955 (59, 2U) ; 1956 (24, 62, 93, 245) ; 1957 (25, 94, 170 372, 501, 529, 550) ; 1958 (298) ; 1959 (124, 486) ; 1960 (157 862 (pp. 309--327), 487). Baked goods: 1960 (430). Gas storage: 1936 (128) ; 1938 (337) ; 1944 (480) ; 1950 (123) 1951 (365, 366) ; 1954 (243). Chemical preservatives: 1941 (269) ; 1942 (270, 271) ; 1948 (551) ; 1954 (483) ; 1957 (484, 497) ; 1959 (415, 485) ; 1961 (488). Eggs: 1939 (188); 1940 (190) ; 1944 (480, 556) ; 1945 (312) ; 1948 (142) ; 1949 (401) ; 1950 (143); 1952 (246) ; 1955 (144, 539) ; 1960 (17). Fish and shellfish: 1937 (168) ; 1949 (397, 436) ; 1954 (398) ;

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 79
1957 {JÍ8JÍ, A97) ; 1960 {S5A) ; 1961 {US, 127, Í37) ; 1962 US8). Fruits and vegetables, fresh: 1938 (39A); 1941 {893) ; 1943 {iOS) ; 1949 {896) ; 1950 {AOA) ; 1951 {JÍ05, 500) ; 1952 (^P5); 1959 {892); 1960 (:^0^) ; 1961 {250); 1963 (^5i). Fruits and vegetables, processed: 1951 (^i^) ; 1959 {809, 878). Halophilic micro-organisms: 1957 {289). Irradiation: 1957 {889); 1960 (^P^) ; 1961 {97). Marine environment: 1946 {559); 1959 (;^P^); 1962 (5(50). Meats and poultry: 1937 (Í50); 1944 {261); 1945 (;^^5, :g5P) ; 1949 {J,6, 285, 262) ; 1951 {12, 18) ; 1954 {268) ; 1955 (i^, ^^7, 288, 2U8); 1958 (:g^0, je^i, 886) ; 1959 (Í5) ; 1960 {17,121) ; 1962 {199). Milk products: 1943 (5i) ; 1947 {868); 1948 (7^) ; 1951 {91, 102) ; 1953 {217) ; 1954 (^^^) ; 1955 {218, 869) ; 1957 {82Jf) ; 1958 (ÍPO, ^Pi) ; 1960 {A92); 1961 (5^i). Minimum growth temperature: 1962 (^0;^) ; 1964 {827). Packaging: 1953 U(?7) ; 1959 {15,169) ; 1962 {198, 2U2). Soils: 1957 {i9U).
LITERATURE CITED
(1) ABBEY, A., KöHLER, A. R., and HOPPER, P. F. 1960. THE EFFECTIVENESS OF ACRONIZE CHLORTETRACYCLINE IN POUL-
TRY PRESERVATION FOLLOWING LONG TERM COMMERCIAL USE. Food Technol. 14: 609-612.
(2) ALFORD, J. A., and ELLIOTT, L. E. 1960. LIPOLYTIC ACTIVITY OF MICRO-ORGANISMS AT LOW AND INTER-
MEDIATE TEMPERATURES. I. ACTION OF PSEUDOMONAS FLUORES- CENS ON LARD. Food Res. 25: 296-303.
(3) and PIERCE, D. A. 1961. ACTIVITY OF MICROBIAL LIPASES AT TEMPERATURES BELOW 0° C.
(Abstract) Inst. Food Technol., Twenty-first Ann. Mtg., 1961. Program, p. 36.
(4) and PIERCE, D. A. 1961. LIPOLYTIC ACTIVITY OF MICRO-ORGANISMS AT LOW AND INTERMEDI-
ATE TEMPERATURES. III. ACTIVITY OF MICROBIAL LIPASES AT TEMPERATURES BELOW 0° C. Jour. Food Sci. 26: 518-524.
(5) ToBiN, L., and MCCLESKEY, C. S. 1942. BACTERIAL SPOILAGE OF ICED FRESH CRABMEAT. Food Res.
7: 353-359. (6) ALLEN, J. R., and FOSTER, E. M.
1960. SPOILAGE OF VACUUM-PACKED SLICED PROCESSED MEATS DURING REFRIGERATED STORAGE. Food Res. 25: 19-25.
(7) ALLEN, M. B. 1960. UTILIZATION OF THERMAL ENERGY BY LIVING ORGANISMS. IN
Florkin, M., and Mason, H. S., eds., Sources of Free Energy Their Comparative Biochemistry v. 1, pp. 487-514.
(8) ALM, F., ERICHSEN, I., and MOLIN, N. 1961. THE EFFECT OF VACUUM PACKAGING ON SOME SLICED PROCESSED
MEAT PRODUCTS AS JUDGED BY ORGANOLEPTIC AND BACTERIOLOGIC ANALYSIS. Food Technol. 15: 199-203.
(9) ALMQUIST, H. J., and HOLST, W. F. 1931. VARIABILITY OF SHELL POROSITY IN THE HEN EGG. Hilgardia
6: 61-72. (10) ANDERSON, B., and WEIL, B. H.
1948. LITERATURE SEARCH ON THE PRESERVATION OF FOODS BY FREEZING. Ga. State Engin. Expt. Sta. Spec. Rpt. 25, pp. 407-670.
(11) AsCHEHOUG, v., and VESTERHUS, R. 1947. BACTERIOLOGICAL INVESTIGATION ON SPOILAGE OF WINTER HER-
RING DURING STORAGE. Food Res. 12: 55-76.

80 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(12) AYRES, J. C. 1951. SOME BACTERIOLOGICAL ASPECTS OF SPOILAGE OF SELF-SERVICE
MEATS. Amer. Meat Inst., Council on Res., Res. Conf. Proc. 3: 39-53.
SOME BACTERIOLOGICAL ASPECTS OF SPOILAGE OF SELF-SERVICE MEATS. Iowa State Col. Jour. Sei. 26: 31-48.
MICROBIOLOGICAL IMPLICATIONS IN THE HANDLING, SLAUGHTER- ING, AND DRESSING OF MEAT ANIMALS. In Mrak, E. M., ed., Advances in Food Res. 6:109-161.
EFFECT OF SANITATION, PACKAGING AND ANTIBIOTICS ON THE MICROBIAL SPOILAGE OF COMMERCIALLY PROCESSED POULTRY. Iowa State Col. Jour. Sei. 34: 27-46.
TEMPERATURE RELATIONSHIPS AND OTHER CHARACTERISTICS OF THE MICROBIAL FLORA DEVELOPING ON REFRIGERATED BEEF. Food Res. 25: 1-18.
THE RELATIONSHIP OF ORGANISMS OF THE GENUS PSEUDOMONAS TO THE SPOILAGE OF MEAT, POULTRY AND EGGS. Jour. Appl. Bact. 23:471-486. OGILVY, W. S., and STEWART, G. F. POST MORTEM CHANGES IN STORED MEATS. I. MICRO-ORGANISMS ASSOCIATED WITH DEVELOPMENT OF SLIME ON EVISCERATED CUT- UP POULTRY. Food Technol. 4: 199-205. and TAYLOR, B.
1956. EFFECT OF TEMPERATURE ON MICROBIAL PROLIFERATION IN SHELL EGGS. Appl. Microbiol. 4: 355-359.
(20) AZUMA, Y., and CLEGG, L. F. L. 1962. SOME OBSERVATIONS ON CARBOHYDRATE UTILIZATION BY DIFFER-
ENT STRAINS OF PSYCHROPHILES ISOLATED FROM RAW MILK. Eighth Internatl. Cong. Microbiol. Abs. No. B 11.5, p. 47. (Abstract in Biol. Abs. 41: 11769. 1963)
(21) BAKER, R. C. 1957. DETERMINING THE KEEPING QUALITY OF ICE-PACKED AND DRY-
PACKED POULTRY. Poultry Sei. 36: 859-864. (22) NAYLOR, H. B., PFUND, M. C. and OTHERS.
1956. KEEPING QUALITY OF READY-TO-COOK AND DRESSED POULTRY. Poultry Sei. 35: 398-406.
(23) BALLS, A. K., and LINEWEAVER, H. 1938. ACTION OF ENZYMES AT LOW TEMPERATURES. Food Res. 3: 57-
67. (24) BARNES, E. M.
1956. SHOULD ANTIBIOTICS BE USED AS FOOD PRESERVATIVES? AMERI- CAN EXPERIENCE AS A GUIDE. Food Manufacturer [Londonl. 31: 508-510.
(25)
(26)
\j.*j;
1951.
(14) 1955.
(15) 1959.
(16) 1960.
(17) 1960.
(18) 1950.
1957. NEW METHODS IN FOOD PRESERVATION. Roy. Soe. for the Pro- motion Health. Jour. 77: 446-457.
1959. POULTRY PLANT HYGIENE. In [Gt. Brit.] Low Temperature Res. Sta., Cambridge, Ann. Rept. 1959: 24-25.
(27) and SHRIMPTON, D. H. 1957. CAUSES OF GREENING OF UNEVISCERATED POULTRY CARCASSES DUR-
ING STORAGE. Jour. Appl. Bact. 20: 273-285. (28) and SHRIMPTON, D. H.
1958. THE EFFECT OF THE TETRACYLINE COMPOUNDS ON THE STORAGE LIFE AND MICROBIOLOGY OF CHILLED EVISCERATED POULTRY. Jour. Appl. Bact. 21: 313-329.
(29) BARTLEY, J. S., WOLCOTT, K. E., and MCSHANE, J. P. 1949. MEAT PACKING PLANT REFRIGERATION. PART I. CARCASS
COOLERS. Amer. Soe. Refrig. Engin., Refrig. Engin. Appl. Data Sect. 45, 10 pp. In Refrig. Engin, v. 57, No. 3.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 81
(30) BATES, P. K., and HIGHLANDS, M. E. 1934. THE DETERMINATION OF STORAGE CONDITIONS. MEAT REFRIGERA-
TION, ITS UTERATURE AND METHODS OF ANALYSIS. Refrig. Engin. 27: 299-302.
(31) BEDFORD, R. H. 1932. SALT AS A CONTROL OF BACTERIAL DECOMPOSITION OF HALIBUT.
Canada Biol. Bd. Bui. 29: 1-16. (32) BÈLEHRADEK, J.
1935. TEMPERATURE AND LIVING MATTER. 277 pp. Berlin. (Proto- plasma-Monographien, v. 8).
(33) BERGEY, D. H., BREED, R. S., MURRAY, E. G. D., and HITCHENS, A. P. 1939. BERGEY'S MANUAL OF DETERMINATIVE BACTERIOLOGY. Ed. 5,
1032 pp. The Williams and Wilkins Co., Baltimore. (34) BERRY, J. A.
1933. DESTRUCTION AND SURVIVAL OF MICROORGANISMS IN FROZEN PACK FOODS. Jour. Bact. 26: 459-470.
(35)
(36)
1934. COLD TOLERANT MICROORGANISMS AND FROZEN PACK. Canner 78 (11): 13-14.
1937. MICROBIOLOGY OF FROZEN PACK VEGETABLES. West. Canner and Packer. 29 (4): 14-16.
(37) and MAGOON, C. A. 1934. GROWTH OF MICROORGANISMS AT AND BELOW 0° C. Phyto-
pathology 24 : 780-796. (38) BERRY, J. M., WITTER, L. D., and FOLINAZZO, J. F.
1956. GROWTH CHARACTERISTICS OF SPOILAGE ORGANISMS IN ORANGE JUICE AND CONCENTRATE. Food Technol. 10: 553-556.
(39) BLUHM, H. M., and TARR, H. L. A. 1956. EFFECT OF CHLORTETRACYCLINE TREATMENT OF FISH ON DEVEL-
OPMENT OF CERTAIN FOOD-POISONING BACTERIA. Canada Fish- eries Res. Bd. Prog. Rpts. Pacific Coast Stas. 106, pp. 8-10.
(40) BORGSTROM, G. 1955. MICROBIOLOGICAL PROBLEMS OF FROZEN FOOD PRODUCTS. Ju
Mrak, E. M., ed.. Advances in Food Res. 6: 163-230. (41) BOTWRIGHT, W. E.
1953. SOILED EGGS MUST BE CLEANED. Poultry Processing and Mktg. 59 (2) : 18, 35-37.
(42) BoYD, J. W., and TARR, H. L. A. 1956. EFFECT OF CHLORTETRACYCLINE AND STORAGE TEMPERATURE ON
THE QUALITY OF SHUCKED OYSTERS. Canada Fisheries Res. Bd. Prog. Rpts. Pacific Coast Stas. 105, pp. 12-13.
(43) BRAMSTEDT, F., and AUERBACH, M. 1961. THE SPOILAGE OF FRESH-WATER FISH. In Borgstrom, G., ed..
Fish as Food 1: 613-637. (44) BREED, R. S., MURRAY, E. G. D., and HITCHENS, A. P.
1948. BERGEY'S MANUAL OF DETERMINATIVE BACTERIOLOGY. Ed. 6, 1529 pp. The Williams & Wilkins Co., Baltimore.
(45) MURRAY, E. G. D., and SMITH, N. R. 1957. BERGEY'S MANUAL OF DETERMINATIVE BACTERIOLOGY. Ed. 7,
1094 pp. The Williams & Wilkins Co., Baltimore. (46) BROEK, C. J. H., VAN DEN.
1949. HET BEDERF VAN DIERLIJKE VOEDINGSMIDDELEN BEI LAGE TEM- PERATUREN, I. DE ONTWIKKELING VAN DE KENNIS DER BEDERFPROCESSEN. [DETERIORATION OF ANIMAL FOODSTUFFS AT LOW TEMPERATURES. I. DEVELOPMENTS OF OUR KNOWLEDGE OF THE DETERIORATION PROCESS.] Chem. Weekbl. 45: 777-784. (Ab- stract in Chem. Abs. 44: 2659. 1950.)
(47) BROOKS, J. 1960. MECHANISM OF THE MULTIPLICATION OF PSEUDOMONAS IN THE
HEN'S EGG. Jour. Appl. Bact. 23: 499-509.

82 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(48) BROWN, A. D. 1957. SOME GENERAL PROPERTIES OF A PSYCHROPHILIC PSEUDOMONAD :
THE EFFECTS OF TEMPERATURE ON SOME OF THESE PROPERTIES AND THE UTILIZATION OF GLUCOSE BY THIS ORGANISM AND PSEUDOMONAS AERUGINOSA. Jour. Gen. Microbiol. 17: 640- 648.
(49) CooTE, G. G., and MEANEY, M. F. 1957. A PROPERTY OF OX MUSCLE AFFECTING BACTERIAL GROWTH AT 0°.
Jour. Appl. Bact. 20: 75-85. (50) and WEIDEMANN, J. F.
1958. THE TAXONOMY OF THE PSYCHROPHILIC MEAT-SPOILAGE BACTERIA; A REASSESSMENT. Jour. Appl. Bact. 21: 11-17.
(51) BROWN, E. B. 1933. BACTERIAL STUDIES OF DEFROSTED PEAS, SPINACH, AND LIMA
BEANS. Jour. Home Econ. 25: 887-892. (52) BROWN, H. J., and GIBBONS, N. E.
1954. AIR CELL MOLD IN OILED EGGS. Food Technol. 8: 307-311. (53) BROWNLEE, D. S., and JAMES, L. H.
1939. BACTERIAL CONTAMINATION OF FROZEN WHOLE EGGS AND AN IM- PROVED METHOD OF DEFROSTING. World's Poultry Cong. Proc. 7: 488-492.
(54) BRUNS, H. 1909. THE BACTERIOLOGICAL CONDITION OF PREPARED FISH. ARCH. F.
Hyg. 67: 209-236. (Abstract in Chem. Abs. 3: 1293'. 1909.) (55) BUCHANAN, R. E., and FULMER, E. I.
1930. PHYSIOLOGY AND BIOCHEMISTRY OF BACTERIA. VOLUME II. 709 pp. Williams and Wilkins Co., Baltimore.
(56) BucHER, D., HEERDT, M., JR., and OSTERHAUG, K. L. 1949. BIBLIOGRAPHY OF THE PRESERVATION OF FISHERY PRODUCTS BY
FREEZING. I. U.S. Fish and Wildlife Serv., Fishery Leaflet 265, [70] pp.
(57) CALLOW, E. H. 1933. GAS STORAGE. [Gt. Brit.l Dept. Sei. and Indus. Res. Food
Invest. Bd. Rpt. 1932: 112-116. (58) and INGRAM, M.
1955. BONE-TAINT. Food 24: 52-55. (59) CAMPBELL, L. L., JR., and O'BRIEN, R. T.
1955. ANTIBIOTICS IN FOOD PRESERVATION. Food Technol. 9: 461- 465.
(60) and WILLIAMS, 0. B. 1952. THE BACTERIOLOGY OF GULF COAST SHRIMP. IV. BACTERIOLOGI-
CAL, CHEMICAL AND ORGANOLEPTIC CHANGES WITH ICE STORAGE. Food Technol. 6: 125-126.
(61) CAMPBELL SOUP COMPANY. 1962. PROCEEDINGS OF LOW TEMPERATURE MICROBIOLOGY SYMPOSIUM,
1961. 322 pp. Camden, N.J. (62) CAREY, B. W.
1956. THE BIOCHEMISTRY OF ANTIBIOTICS ON THE BACTERIA ASSOCIATED WITH FOOD SPOILAGE. Gior. di Microbiol. 2: 233-257. (Ab- stract in Biol. Abs. 32: 41548. 1958.)
(63) CARLE, L. A., and KYTE, L. 1955. KEEPING QUALITY OF CHILLED DUNGENESS CRABMEAT IN HERME-
TICALLY-SEALED METAL CONTAINERS. Com. Fisheries Rev. 17 (2): 12-14.
(64) CARLSON, C. J. 1954. KEEPING QUALITY OF CHILLED DUNGENESS CRABMEAT PACKED IN
HERMETICALLY-SEALED CONTAINERS. Com. Fisheries Rev. 16 (11): 20-21.
(65) CASTELL, C. H. 1949. REFRIGERATION TEMPERATURES AND THE KEEPING TIME OF FRESH
FISH. Canada Fisheries Res. Bd. Prog. Rpts. Atlantic Coast Stas. 44, pp. 8-12.
(ee) 1960. CHLORINE AFFECTS ANTIBIOTICS. Canad. Fisherman 47 (2) :
37-39. (Abstract in Biol. Abs. 36: 65453. 1961.)

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 83
(67) CASTELL, C. H., ANDERSON, G. W., and PIVNICK, H. 1948. RELATION OF BACTERIAL COUNTS TO QUALITY OF COD FILLETS.
Canada Fisheries Res. Bd. Jour. 7: 378-388. (68) and MACCALLUM, W. A.
1950. THE VALUE OF TEMPERATURES CLOSE TO FREEZING ON THE STOR- AGE OF FISH. Canada Fisheries Res. Bd. Jour. 8: 111-116.
(69) and MAPPLEBECK, E. G. 1952. THE IMPORTANCE OF FLAVOBACTERIUM IN FISH SPOILAGE.
Canada Fisheries Res. Bd, Jour. 9: 148-156. (70) CEPONIS, M. J., and FRIEDMAN, B. A.
1957. TEMPERATURE AND SHELF-LIFE STUDIES WITH PREPEELED PO- TATOES. Amer. Potato Jour. 34: 333-341.
(71) CHANDRA SEKHAR, C. V., and WALKER, N. 1947. PRELIMINARY OBSERVATIONS ON VARIOUS TEMPERATURE CHAR-
ACTERISTICS OF SOME FACULTATIVE PSYCHROPHILIC BACTERIA. Soc. Appl. Bact [Gt. Brit.] Proc. 10: 24-27.
(72) CHEVALIER, R.
1948. LE ROLE BIOLOGIQUE DU FROID EN LAITERIE. Rev. Gen. du Froid 25: 341, 399, 505. (Abstract in Food Sei. Abs. 21: 583. 1949.)
(73) CHISTYAKOV, F. M., and BOCHAROVA, Z. Z. 1937. THE INFLUENCE OF LOW TEMPERATURE ON MICRO-ORGANISMS.
II. INFLUENCE OF LOW TEMPERATURE ON THE DEVELOPMENT OF MOULDS. Mikrobiologiia [U.S.S.R.] 6: 1293-1309.
(74) and BOCHAROVA, Z. Z. 1938. INFLUENCE OF LOW TEMPERATURE ON THE DEVELOPMENT OF
MICROORGANISMS. IV. INFLUENCE OF LOW TEMPERATURE ON THE DEVELOPMENT OF MOLDS. Mikrobiologiia [U.S.S.R.] 7: 838-842.
(75) and NosKovA, G. 1955. THE ADAPTATION OF MICRO-ORGANISMS TO LOW TEMPERATURES.
Ninth Internatl. Cong. Refrig. Proc. 2: 4.230-4.235. (76) CHRISTIAN, J. H. B., and WALTHO, J. A.
1961. THE SODIUM AND POTASSIUM CONTENT OF NONHALOPHILIC BAC- TERIA IN RELATION TO SALT TOLERANCE. Jour. Gen. Microbiol. 25 • 97—102
(77) CLAUSS, W. E., BALL, C. O., and STIER, E. F. 1956. FACTORS AFFECTING QUALITY OF PREPACKAGED MEATS. I. PHYSI-
CAL AND ORGANOLEPTIC TESTS. C. ORGANOLEPTIC AND MIS- CELLANEOUS PHYSICAL CHARACTERISTICS OF PRODUCT. Food Technol. 11: 363-373.
(78) CLAYPOOL, L. L. 1953. PERISHABILITY OF FRESH FRUITS AND VEGETABLES. Pre-pack
Age 6 (11) : 18-20, 22. (79) CLAYSON, D. H. F.
1955. FOOD PERISHABILITY IN TERMS OF MICROECOLOGY. Jour. Sci. Food and Agr. 6: 565-575.
(80) CONE, J. F., DODDS, M. L., GUERRANT, N. B., and OTHERS. 1956. EFFECTS OF THAWING AND REFREEZING ON THE QUALITY OF CER-
TAIN FROZEN FOODS. Pa. Agr. Expt. Sta. Bui. 614, 22 pp. (81) CoRLEY, R. T., LONG, H. F., and HAMMER, B. W.
1943. WATER SUPPLIES OF BUTTER MANUFACTURING PLANTS. lowa Agr. Expt. sta. Res. Bui. 319, pp. 749-794.
(82) CoTTERiLL, 0. J., and HARTMAN, P. 1956. EFFECT OF ANTIBIOTICS ON THE INCIDENCE OF SPOILAGE OF SHELL
EGGS. Poultry Sei. 35: 733-735. (83) COYNE, F. P.
1932. THE EFFECT OF CARBON DIOXIDE ON BACTERIAL GROWTH WITH SPECIAL REFERENCE TO THE PRESERVATION OF FISH. PART I. Trans, and Commun., Soc. Chem. Indus. Jour. [London] 51: 119T-121T.
(84) 1933. THE EFFECT OF CARBON DIOXIDE ON BACTERIAL GROWTH WITH SPE-
CIAL REFERENCE TO THE PRESERVATION OF FISH. [Gt. Brit.] Dept. Sei. and Indus. Res., Food Invest. Bd. Rpt., 1932: 194-196.

84 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(85) COYNE. F. P. 1933. THE EFFECT OF CARBON DIOXIDE ON BACTERIAL GROWTH WITH
SPECIAL REFERENCE TO THE PRESERVATION OF FISH. PART II. GAS STORAGE OF FRESH FISH. Trans. and Comm. Soc. Chem. Indus. Jour. [London] 52: 19T-24T.
(86) CROOKS, G. C, and RITCHIE, W. S. 1938. A STUDY OF RATE OF DECOMPOSITION OF HADDOCK MUSCLE AT
VARIOUS TEMPERATURES AS INDICATED BY AMMONIA CONTENT. Food Res. 3: 589-598.
(87) CROZIER, W. J. 1924. ON THE CRITICAL THERMAL INCREMENT FOR THE LOCOMOTION OF
A DIPLOPOD. Jour. Gen. Physiol. 7: 123-136. (88)
1926. ON CURVES OF GROWTH, ESPECIALLY IN RELATION TO TEMPERA- TURE. Jour. Gen. Physiol. 10: 53-73.
(89) DASSOW, J. [A.] 1949. BIBLIOGRAPHY OF THE PRESERVATION OF FISHERY PRODUCTS BY
FREEZING. II. U.S. Fish and Wildlife Serv. Fishery Leaflet 265 SuppL, [13] pp.
(90) 1956. HANDLING OF FISH ABOARD THE VESSEL. In Handling Fresh
Fish. U.S. Fish and Wildlife Serv. Fishery Leaflet 428, pp. 13—37
(91) DAVIS, J. G. 1951. THE EFFECT OF COLD ON MICRO-ORGANISMS IN RELATION TO DAIRY-
ING. Soc. Appl. Bact. [Gt. Brit.] Proc. 14: 216-242. (92) DAWSON, L. E., and STADELMAN, W. J.
1960. MICRO-ORGANISMS AND THEIR CONTROL ON FRESH POULTRY MEAT (A REVIEW). Mich. Agr. Expt. Sta. Tech. Bui. 278, 39 pp.
(93) DEATHERAGE, F. E. 1956. THE USE OF ANTIBIOTICS IN THE PRESERVATION OF FOODS OTHER
THAN FISH. In Proc. First Internatl. Conf. on the Use of Antibiotics in Agr. Nati. Acad. Sei. Pub. 397, pp. 211-222.
(94) 1957. USE OF ANTIBIOTICS IN THE PRESERVATION OF MEATS AND OTHER
FOOD PRODUCTS. Amer. Jour. Pub. Health 47: 594-600. (95) DEOTTO, E.
1944. L'AZIONE DEL FREDDO SULLA RESPIRAZIONE DELLA CELLULA BAT- TERICA. Arch. f. Expt. Zeilforsch. 25:101-105.
(96) 1946. L^AZIONE DEL FREDDO SULLA RESPIRAZIONE BATTERICA. Rome,
Italy. Accad. Naz. dei LinLcei Atti., Ser 8 (1) : 241-246. (97) DE SERRES, F. J.
1961. SOME ASPECTS OF THE INFLUENCE OF ENVIRONMENT ON THE RADIOSENSITIVITY OF MICRO-ORGANISMS. In Microbial Reaction to Environment. Soc. for Gen. Microbiol. Symposium 11: 196-216.
(98) DiEHL, H. C. 1929. THE COLD PACK METHOD OF PRESERVING BERRIES IN THE PACIFIC
NORTHWEST. Glass Packer 2: 115-120,129. (99) CAMPBELL, H., and BERRY, J. A.
1936. SOME OBSERVATIONS ON THE FREEZING PRESERVATION OF ALDER- MAN PEAS. Food Res. 1: 61-71.
(100) MAGNESS, J. R., GROSS, C. R., and BONNEY, V. B. 1930. THE FROZEN-PACK METHOD OF PRESERVING BERRIES IN THE PA-
CIFIC NORTHWEST. U.S. Dept. Agr. Tech. Bui. 148, 38 pp. (101) Di MENNA, M. E.
1960. YEASTS FROM ANTARCTICA. Jour. Gen. Microbiol. 23: 295-300. (102) DoETSCH, R. N., and SCOTT, W. M.
1951. PSYCHROPHILIC BACTERIA IN MILK AND CREAM. Milk Plant Monthly 40: 30-32 (March).
(103) DORN, F. L., and RAHN, O. 1939. DEFINITION VERSUS MEASUREMENT OF OPTIMAL TEMPERATURE.
Arch. f. Mikrobiol. 10: 6-12.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 85
104) DRAKE, S. D., EVANS, J. B., and NIVEN C. F., JR. 1959. THE IDENTITY OF YEASTS IN THE SURFACE FLORA OF PACKAGED
FRANKFURTERS. Food Res. 24: 243-246. 105) DUNCAN, D. W., JR., and NICKERSON, J. T. R.
1962. EFFECT OF ENVIRONMENTAL AND PHYSIOLOGICAL CONDITIONS ON THE EXPONENTIAL PHASE GROWTH OF PSEUDOMONAS FRAGI (ATCC 4973). In Campbell Soup Co., Camden, N.J. Low Temp. Microbiol. Symposium Proc. 1961: 253-262.
106) DYER, W. J., and DYER, F. E. 1950. AMINES IN FISH MUSCLE. IV. SPOILAGE IN FRESHLY CUT COD
FILLETS. Canada Fisheries Res. Bd. Jour. 7: 580-584. 107) DYER, F. E., and SNOW, J. M.
1947. THE SPOILAGE OF GUTTED COD STORED IN CRUSHED ICE. Canada Fisheries Res. Bd. Prog. Rpts. Atlantic Coast Stas. No. 37, pp. 3-7.
108) ELLIOTT, L. E., and BRANT, A. W. 1957. EFFECT OF SALINE AND EGG SHELL MEMBRANE ON BACTERIAL
GROWTH. Food Res. 22: 241-250. 109) ELLIOTT, R. P.
1946. MICROBIOLOGICAL FLORA IN FISH (A BIBLIOGRAPHY). [Printed on 3x5 inch cards. Availability announced in Com. Fisheries Rev. 8(3): 25.]
SPOILAGE OF SHELL EGGS BY PSEUDOMONADS. Appl. Microbiol. 2: 158-164.
SOME PROPERTIES OF PYOVERDINE, THE WATER-SOLUBLE FLUORES- CENT PIGMENT OF THE PSEUDOMONADS. Appl. Microbiol. 6: 241-246.
TEMPERATURE-GRADIENT INCUBATOR FOR DETERMINING THE TEMPERATURE RANGE OF GROWTH OF MICRO-ORGANISMS. Jour. Bact. 85: 889-894. and MiCHENER, H. D. MICROBIOLOGICAL STANDARDS AND HANDLING CODES FOR CHILLED AND FROZEN FOODS—^A REVIEW. Appl. Microbiol. 9: 452-468. and STRAKA, R. P.
1964. RATE OF MICROBIAL DETERIORATION OF CHICKEN MEAT AT 2" C. AFTER FREEZING AND THAWING. Poultry Sci. 43: 81-86.
115) EMPEY, W. A. 1958. FISH HANDLING AND PRESERVATION, III. CHANGES DURING
FREEZING AND FROZEN STORAGE. Food Preserv. Quart. 18: 76-78.
116) and VICKERY, J. R. 1933. THE USE OF CARBON DIOXIDE IN THE STORAGE OF CHILLED BEEF.
Austral. Council Sei. and Indus. Res. Jour. 6: 233-243. 117) ERICKSON, F. J., and FABIAN, F. W.
1942. PRESERVING AND GERMICIDAL ACTION OF VARIOUS SUGARS AND ORGANIC ACIDS ON YEASTS AND BACTERIA. Food ReS. 7: 68-79.
118) EssARY, E. 0., DAWSON, L. E., and MALLMANN, W. L. 1960. EFFECT OF DIFFERENT LENGTH CHILL PERIODS WITH CHLORTETRA-
CYCLINE AND DIFFERENT HOLDING CONDITIONS ON THE SHELF LIFE OF DRESSED FRYERS. Food Technol. 14: 286-289.
119) MOORE, W. E. C, and KRAMER, C. Y. 1958. INFLUENCE OF SCALD TEMPERATURES, CHILL TIMES, AND HOLDING
TEMPERATURES ON THE BACTERIAL FLORA AND SHELF LIFE OF FRESHLY CHILLED, TRAY-PACK POULTRY. Food Technol. 12: 684-687.
120) EVANS, J. B. 1959. BACTERIAL SPOILAGE OF CANNED HAMS. Amer. Meat Inst.
Found. Cir. 54, 4 pp. 121) and NIVEN, C. F., JR.
1960. MICROBIOLOGY OF MEAT. BACTERIOLOGY. In Amer. Meat Inst. Found., The Science of Meat and Meat Products, 438 pp., (pp. 151-179.) W. H. Freeman & Co., San Francisco and London.
xxv/
1954.
111) 1958.
112) 1963.
113)
^^A\
1961.

86 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(122) EwELL, A. W. . ^ ^ . 1936. THE USE OF OZONE IN COLD STORAGE PLANTS. Ice and Kefrig.
91: 295-296. (123)
1950. OZONE AND ITS APPLICATION IN FOOD PRESERVATION. Amer. Soc. Refrig. Engin., Refrig. Engin. Appl. Data Sect. 50, 4 pp. In Refrig. Engin, v. 58, No. 9.
(124) FäRBER, L. A T> ivyr- T.- i 1959. ANTIBIOTICS IN FOOD PRESERVATION. Ann. Rev. Microbiol.
13: 125-140. (125) FEENEY, R. E., MACDONNELL, L. R., and LORENZ, F. W.
1953. HIGH TEMPERATURE TREATMENT OF SHELL EGGS. (Abstract) Poultry Sei. 32: 899.
(126) MACDONNELL, L. R., and LORENZ, F. W. 1954. HIGH TEMPERATURE TREATMENT OF SHELL EGGS. Food TechnoL
8: 242-245. (127) FiEGER, E. A., and NovAK, A. F.
1961. MICROBIOLOGY OF SHELLFISH DETERIORATION. In Borgstrom, G., ed. Fish as Food 1: 561-611.
(128) FiSK, N. R. 1936. RESEARCHES THAT MADE GAS STORAGE POSSIBLE. PARTS I, II,
and in. Food Indus. 8: 390-391, 446-447, 448, 571-572, 602. (129) FLORIAN, M. L. E., and TRUSSELL, P. G.
1957. BACTERIAL SPOILAGE OF SHELL EGGS. IV. IDENTIFICATION OF SPOILAGE ORGANISMS. Food Technol. 11: 56-60.
(130) FORSTER, J. 1892. UEBER DIE ENTWICKELUNG VON BAKTERIEN BEI NIEDERN TEM-
PERATUREN. Gentbl. f. Bakt. [etc.], Abt. II, 12: 431-436. (131) FoRSYTHE, R. H., AYRES, J. G., and RADLO, J. L.
1953. FACTORS AFFECTING THE MICROBIOLOGICAL POPULATIONS OF SHELL EGGS. Food Technol. 7: 49-56.
(132) FOSTER, E. M. 1959. BACTERIAL PROBLEMS IN PREPACKAGED MEATS. Amer. Meat
Inst. Found. Res. Conf. (Gir. 50) Proc. 11: 61-64. (133) FOTER, M. J., and RAHN, O.
1936. GROWTH AND FERMENTATION OF BACTERIA NEAR THEIR MINI- MUM TEMPERATURE. Jour. Bact. 32: 485-497.
(134) FRANK, H. A., ISHIBASHI, S. T., REíD, A., and ITO, J. S. 1963. CATALASE ACTIVITY OF PSYCHROPHILIC BACTERIA GROWN AT 2
AND 30 C. Appl. Microbiol. 11: 151-153. (135) FROMM, D.
1957. BACTERIAL CONTAMINATION AND SHELF LIFE OF FRESHLY EVIS- CERATED BROILERS AS INFLUENCED BY HOLDING TIME IN SLUSH ICE. Poultry Sei. 36: 1006-1009.
INFLUENCE OF REUSING CHILL TANK SLUSH ICE ON MARKET QUALITY OF EVISCERATED BROILERS. Food Technol. 12: 257- 259.
PERMEABILITY OF THE EGG SHELL AS INFLUENCED BY WASHING, AMBIENT TEMPERATURE CHANGES, AND ENVIRONMENTAL TEM- PERATURE AND HUMIDITY. Poultry Sei. 39: 1490-1495.
LACK OF CORRELATION BETWEEN EGG SHELL THICKNESS AND SHELL PERMEABILITY OF MARKET EGGS. Jour. Food Sci. 27: 111-112. and MARGOLF, P. H. THE INFLUENCE OF SWEATING AND WASHING ON WEIGHT LOSS, BACTERIAL CONTAMINATION AND INTERIOR PHYSICAL QUAUTY OF 12-DAY-OLD SHELL EGGS. Poultry Sei. 37: 1273-1278. and MONROE, R. J.
1960. INTERIOR PHYSICAL QUALITY AND BACTERIAL CONTAMINATION OF MARKET EGGS AS INFLUENCED BY EGG SHELL PERMEABILITY. Food Technol. 14: 401-403.
V-LOU;
1958.
(137) 1960.
(138) 1962.
(139)
í^Á(\\
1958.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 87
141) FUNK, E. M. 1943. PASTEURIZATION OF SHELL EGGS. Mo. AgT. Expt. Sta. Res. Bul.
364, 28 pp. 142)
143)
144)
1948. EXPERIMENTS IN CLEANING SOILED EGGS FOR STORAGE. Mo. Agr. Expt. Sta. Res. Bul. 426, [361 pp.
1950. MAINTENANCE OF QUALITY IN SHELL EGGS BY THERMO-STABILI- ZATION. Mo. Agr. Expt. Sta. Res. Bul. 467, 46 pp.
1955. TREATING SHELL EGGS TO MAINTAIN QUALITY. Mo. Agr. Expt. Sta. Bul. 659, 19 pp.
145) FORWARD, J., and LORAH, M. 1954. MINIMIZING SPOILAGE IN SHELL EGGS BY THERMOSTABILIZATION.
Poultry Sei. 33: 532-538. 146) GALLOWAY, L. D.
1960. PLASTIC FILMS AND MOULD GROWTH. Lab. Pract. 9: 596-597. (Abstract in World Fisheries Abs. 12 (4) : 23. 1961.)
147) GARIBALDI, J. A. 1960. FACTORS IN EGG WHITE WHICH CONTROL GROWTH OF BACTERIA.
Food Res. 25: 337-344. 148) and BAYNE, H. G.
1960. THE EFFECT OF IRON ON THE PSEUDOMONAS SPOILAGE OF EXPERI- MENTALLY INFECTED SHELL EGGS. Poultry Sci. 39: 1517-1520.
149) and BAYNE, H. G. 1962. IRON AND THE BACTERIAL SPOILAGE OF SHELL EGGS. Jour. Food
Sei. 27: 57-59. 150) and BAYNE, H. G.
1962. THE EFFECT OF IRON ON THE PSEUDOMONAS SPOILAGE OF FARM WASHED EGGS. Poultry Sei. 41: 850-853.
151) and STOKES, J. L. 1958. PROTECTIVE ROLE OF SHELL MEMBRANES IN BACTERLVL SPOILAGE
OF EGGS. Food Res. 23. 283-290. 152) GIBBONS, N. E.
1934. A BACTERIOLOGICAL STUDY OF "ICE FILLETS." Ganad. Biol. and Fisheries Contrib. (n.s.) 8: 301-310.
153) 1934. HYDROCHLORIC ACID PRESERVES FISH. Canada Biol. Bd. Prog.
Rpts. Atlantic Coast Stas. 10, p. 7. 154) and REED, G. B.
1930. THE EFFECT OF AUTOLYSIS IN STERILE TISSUES ON SUBSEQUENT BACTERIAL DECOMPOSITION. Jour. Bact. 19: 73-88.
155) and ROSE, D. 1950. EFFECT OF ANTE-MORTEM TREATMENT OF PIGS ON THE QUALITY
OF WILTSHIRE BACON. Canad. Jour. Res. Sect. F, 28: 438-450. 156) GiLLESPiE, J. M., and ScOTT, W. J.
1950. STUDIES IN THE PRESERVATION OF SHELL EGGS. IV. EXPERI- MENTS IN THE MODE OF INFECTION BY BACTERIA. Austral. Jour. Appl. Sei. 1: 514-530.
157) GOLDBERG, H. S. 1960. ANTIBIOTICS. THEIR CHEMISTRY AND NONMEDICAL USES. 608
pp. D. Van Nostrand, Princeton, N.J. 158) GORTNER, R. A., and GORTNER, W. A., editors.
1949. OUTLINES OF BIOCHEMISTRY. Ed. 3, 1078 pp. Wiley, N.Y. 159) [GREAT BRITAIN] DEPARTMENT OF SCIENTIFIC AND INDUSTRIAL RE-
SEARCH 1955. THE EFFECT OF AMBIENT TEMPERATURE ON SPOILAGE. [GT. BRIT.,]
Dept. Sei. and Indus. Res. Food Invest. Bd. Rpt. 1954: 34-35. 160)
161)
1956. THE EFFECT OF AMBIENT TEMPERATURE. [Gt. Brit.] Dept. Sci. and Indus. Res., Food Invest. Bd. Rpt. 1955, 24 pp.
1958. REPORT OF THE SUPERINTENDENT OF THE LOW TEMPERATURE RE- SEARCH STATION FOR 1957. [Gt. Brit.l Dept. Sci. and Indus, Res. Food Invest. Bd. Rpt. 1957: 32-53,

88 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(162) [GREAT BRITAIN] DEPARTMENT OF SCIENTIFIC AND INDUSTRIAL RE- SEARCH.
1960. ON THE HANDLING AND PRESERVATION OF FISH AND FISH PROD- UCTS [Gt. Brit.] Terry Res. Sta., Edinburgh, Rpt. Dir. (1959) 7-8: 20-23.
(163) GREEN, M. 1949. BACTERIOLOGY OF SHRIMP. II. QUANTITATIVE STUDIES ON FRESH-
LY CAUGHT AND ICED SHRIMP. Food Res. 14: 372-383. (164) GREENE, V. W.
1959. INFLUENCE OF TEMPERATURE HISTORY ON THE RESPONSE OF PSYCHROPHILES TO DIFFERENT INCUBATION TEMPERATURES. Jour. Dairy Sei. 42: 1097-1099.
(165) and JEZESKI, J. J. 1954. INFLUENCE OF TEMPERATURE ON THE DEVELOPMENT OF SEVERAL
PSYCHROPHILIC BACTERIA OF DAIRY ORIGIN. Appl. Microbiol. 2: 110-117.
(166) GREIG, W. S., and SMITH, O. 1960. FACTORS AFFECTING QUALITY AND STORAGE LIFE OF PREPEELED
POTATOES AND QUALITY OF FRENCH FRIES. N.Y. (Cornell) Agr. Expt. sta. Mem. 370, 48 pp.
(167) GRESSEL and GRäFE. 1929. ZUR FRAGE DER HALTBARKEIT VON HACKFLEISCH AUS FRISCH-
FLEISCH UND GEFRIERFLEISCH. Berlin Tierärztl. Wochnschr. 45: 430-432.
(168) GRIFFITHS, F. P. 1937. A REVIEW OF THE BACTERIOLOGY OF FRESH MARINE-FISHERY PROD-
UCTS. Food Res. 2: 121-134. (169) GROSS, G. B.
1959. USE OF FLEXIBLE PACKAGING MATERIALS AND INERT GAS TO IN- CREASE THE SHELF LIFE OF POOD PRODUCTS. Food Technol. 13 : 621-623.
(170) GRUBB, T. C. 1957. SYMPOSIUM ON ANTIMICROBIAL PRESERVATIVES. Bact. Rev. 21:
251-254. (171) GUADAGNI, D. G.
1960. CALCULATION OF QUALITY CHANGE FROM TIME-TEMPERATURE RECORDS. In Conference on Frozen Food Quality. U.S. Dept. Agr., Agr. Res. Serv. ARS 74r-21, pp. 13-17.
(172) GUNDERSON, M. F. 1962. MOLD PROBLEM IN FROZEN FOODS. Campbell Soup Co., Camden,
N.J., Low Temp. Microbiol. Symposium Proc. 1961: 299-312. (173) and MCFADDEN, H. W., JR.
1948. BACTERIAL ASPECTS OF PRECOOKED FROZEN FOODS. File MlOO, Contract Wll-183-qm-228. U.S. Quartermaster Food and Container Inst. for the Armed Forces, Rpt. 5, 81 pp.
(174) MCFADDEN, H. W., JR., and MINTHORN, G. 1947. BACTERIAL ASPECTS OF PRECOOKED FROZEN FOODS. File MlOO,
Contract Wll-183-qm-228. U.S. Quartermaster Food and Container Inst. for the Armed Forces, Rpt. 1, 86 pp.
(175) MCFADDEN, H. W., JR., and MINTHORN, G. 1948. BACTERIAL ASPECTS OF PRECOOKED FROZEN FOODS. File MlOO.
Contract Wll~183-qm-228. U.S. Quartermaster Food and Container Inst. for the Armed Forces, Rpt. 2, 96 pp.
(176) HAGEN, P. 0., and ROSE, A. H. 1961. A PSYCHROPHILIC CRYPTOCOCCUS. Canad. Jour. Microbiol. 7:
287-294. (177) and ROSE, A. H.
1962. STUDIES ON THE BIOCHEMICAL BASIS OF THE LOW MAXIMUM TEMPERATURE IN A PSYCHROPHILIC CRYPTOCOCCUS. Jour. Gen. Microbiol. 27: 89-99.
(178) HAINES, R. B. 1930. BIOCHEMICAL STUDIES OP THE ACTION OF BACTERIA ON MEAT AT
DIFFERENT TEMPERATURES. [Gt. Brit.] Dept. Sci. and Indus. Res. Food Invest. Bd. Rpt. 1929: 43.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 89
(179) HAINES, R. B. 1931. THE INFLUENCE OF TEMPERATURE ON THE RATE OF GROWTH OF
SPOROTRICHUM GARNIS, FROM —10° C. TO +30° C. Jour. Expt. BioL8: 379-388.
(180)
(181)
1931. THE GROWTH OF MICRO-ORGANISMS ON CHILLED AND FROZEN MEAT. Trans, and Commun. Soc. Chem. Indus. Jour. [London] 50: 223T-227T.
(182)
(183)
1931. BACTERIOLOGY. [Gt. Brit.] Dept. Sei. and Ind. Res. Food In- vest. Bd. Rpt. 1930: 39-47.
1932. THE INFLUENCE OF TEMPERATURE ON THE RATE OF GROWTH OF SAPROPHYTic ACTiNOMYCES. Jour. Expt. Biol. 9: 45-60.
1933. THE INFLUENCE OF CARBON DIOXIDE ON THE RATE OF MULTIPLICA- TION OF CERTAIN BACTERIA, AS JUDGED BY VIABLE COUNTS. Trans, and Commun., Soc. Chem. Indus. Jour. [London] 52: 13T-17T.
THE MINIMUM TEMPERATURES OF GROWTH OF SOME BACTERIA. Jour. Hyg. 34: 277-282.
BACTERIOLOGY OF THE HEN'S EGG. [Gt. Brit.] Dept. Sci. and Indus. Res., Food Invest. Bd. Ept. 1936: 59-65.
MICROBIOLOGY IN THE PRESERVATION OF ANIMAL TISSUES. [Gt. Brit.] Dept. Sei. and Indus. Res., Food Invest. Bd. Spec. Rpt. 45, 85 pp.
ORSERVATIONS ON THE BACTERIAL FLORA OF THE HEN'S EGG WITH A NEW SPECIES OF PROTEUS AND PSEUDOMONAS CAUSING ROTS IN EGGS. Jour. Hyg. 38: 338-355.
MICROBIOLOGY IN THE PRESERVATION OF THE HEN'S EGG. [Gt. Brit.] Dept. Sei. and Indus. Res. Food Invest. Bd. Spec. Rpt. 47, 65 pp.
THE EFFECT OF TEMPERATURE OF THE WASHING FLUID ON THE SUBSEQUENT BACTERIAL ROTTING OF EGGS. [Gt. Brit.] Dept. Sci. and Indus. Res., Food Invest. Bd. Rpt. 1939: 22-23.
THE PRESERVATION OF EGGS. Chem. and Indus. 18: 391-396.
THE ISOLATION OF AN^ROBES FROM TAINTED MEAT. Chem. and Indus. 19: 413-416.
-and SMITH, E. C. 1933. THE STORAGE OF MEAT IN SMALL REFRIGERATORS. [Gt. Brit.]
Dept. Sci. and Indus. Res., Food Invest. Bd. Spec. Rpt. 43, 30 pp.
(193) HALL, I. C. 1942. ACTION OF VISKING SAUSAGE-CASING MATERIAL ON BACTERIAL
GROWTH. Food Res. 7: 104-110. (194) HALLECK, F. E., BALL, C. 0., and STIER, E. F.
1958. FACTORS AFFECTING QUALITY OF PREPACKAGED MEAT. IV. MICRO- BIOLOGICAL STUDIES., A. CULTURAL STUDIES ON BACTERIAL FLORA OF FRESH MEAT; CLASSIFICATION BY GENERA. Food Technol. 12: 197-203.
(195) BALL, C. 0., and STIER, E. F. 1958. FACTORS AFFECTING QUALITY OF PREPACKAGED MEAT. IV. MICRO-
BIOLOGICAL STUDIES. B. EFFECT OF PACKAGE CHARACTERISTICS, AND OF ATMOSPHERIC PRESSURE IN PACKAGE UPON BACTERIAL FLORA OF MEAT. Food Technol. 12:301-306.
\±0'±J 1934.
(185) 1937.
(186) 1937.
(187) 1938.
(188) 1939.
(189) 1940.
(190) 1940.
(191) 1941.

90 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(196) HALLECK, F. E. BALL, C. 0., and STIER, E. F. 1958. FACTORS AFFECTING QUALITY OF PREPACKAGED MEAT. IV. MICRO-
BIOLOGICAL STUDIES. C. EFFECTS OF INITIAL BACTERIA COUNT, KIND OF MEAT, STORAGE TIME, STORAGE TEMPERATURE, ANTI- OXIDANTS, AND ANTIBIOTICS ON THE RATE OF BACTERIAL GROWTH IN PACKAGED MEAT. Food Technol. 12: 654-659.
(197) HALVORSON, H. 0., WOLF, J., and SRINIVASAN, V. R. 1962. INITIATION OF GROWTH AT LOW TEMPERATURES. Campbell Soup
Co., Camden, N.J., Low Temp. Microbiol. Symposium Proc. 1961: 27-40.
(198) HANNAN, R. S. 1962. SOME PROPERTIES OF FOOD PACKAGING MATERIALS WHICH RELATE
TO THE MICROBIAL FLORA OF THE CONTENTS. Jour. Appl. Bact. 25: 248-258.
(199) HANSEN, N. and RIEMANN, H. 1962. MIKROBIOLOGISCHE BESCHAFFENHEIT VON VORVERPACKTEM
FLEISCH UND VORVERPACKTEN FLEISCHWAREN. Die Fleisch- wirtschaft 9: 861-862, 865-868.
(200) HANSON, H. L., and FLETCHER, L. R. 1958. TIME-TEMPERATURE TOLERANCE OF FROZEN FOODS. XII. TURKEY
DINNERS AND TURKEY PIES. Food Technol. 12: 40-43. (201) HARRINGTON, W. 0., MAYER, P. C, OLSON, R. L., and OTHERS.
1956. OBSERVATIONS ON PREPEELING OF POTATOES. Food Technol. 10: 347-351.
(202) HARRISON, F. C, and SADLER, W. 1929. DISCOLORATION OF HALIBUT. (Abstract) Jour. Bact. 17: 29-30.
(203) HARTSELL, S. E. 1962. THE MICROBIOLOGY OF FROZEN FOODS. Campbell Soup Co., Cam-
den, N.J., LOW Temp. Microbiol. Symposium Proc. 1961: 263- 283.
(204) CHANCE, H. L., LICHSTEIN, H. C, and HALVORSON, H. 0. 1959. SYMPOSIUM ON INITIATION OF BACTERIAL GROWTH. Bact. Rev.
23: 250-272. (205) HäRTUNG, T. E., and STADELMAN, W. J.
1962. THE INFLUENCE OF METALLIC CATIONS ON THE PENETRATION OF THE EGG SHELL MEMBRANES BY PSEUDOMONAS FLUORESCENS. Poultry Sei. 41: 1590-1596.
(206) HARVEY, J. M., and PENTZER, W. T. 1960. MARKET DISEASES OF GRAPES AND OTHER SMALL FRUITS. U.S.
Dept. Agr., Agr. Handb. 189, 37 pp. (207) HEATHER, C. D., and VAN DER ZANT, W. C.
1957. EFFECTS OF TEMPERATURE AND TIME OF INCUBATING AND PH OF PLATING MEDIUM ON ENUMERATING HEAT-TREATED PSYCHRO- PHILIC BACTERIA. Jour. Dairy Sei. 40. 1079-1086.
(208) and VAN DER ZANT, W. C. 1957. EFFECT OF THE PLATING MEDIUM ON THE SURVIVAL OF HEAT-
TREATED CELLS OF PSEUDOMONAS FLUORESCENS. Food ReS. 22: 164-169.
(209) HENDRICKSON, R. L. 1949. BACTERIAL STUDIES RELATED TO THE KEEPING QUALITY OF FROZEN
PORK SAUSAGE. Jour. Anim. Sei. 8: 613. (210) HESS, E.
1933. CULTURAL CHARACTERISTICS OF MARINE BACTERIA IN RELATION TO LOW TEMPERATURES. (Abstract) Nova Scotian Inst. Sei. Proc. and Trans. 18: 292-293.
(211)
(212)
1934. EFFECTS OF LOW TEMPERATURES ON THE GROWTH OF MARINE BACTERIA. Canad. Biol. and Fisheries Contrib. (n.s.) 8: 489- 505.
(213)
1934. TO STOP SPOILAGE KEEP FISH VERY COLD. Canada Biol. Bd. Prog. Rpts., Atlantic Coast Stas. 10, p. 6.
1934. EFFECT OF TEMPERATURE ON THE CELL LENGTH AND SHAPE OF BACILLUS VULGATUS. Canada Biol. Bd. Jour. 1: 109-119.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 91
(214) HESS, E. 1934. CULTURAL CHARACTERISTICS OF MARINE BACTERIA IN RELATION
TO LOW TEMPERATURES AND FREEZING. Canad. Biol. and Fish- eries Contrib. (n.s.) 8: 459-474.
(215)
(216)
1942. STUDIES ON SALT FISH. IX. EFFECT OF ENVIRONMENT UPON GROWTH OF RED HALOPHiLic BACTERIA. Canada Fisheries Res. Bd. Jour. 6: 10-16. (Abstract in Com. Fisheries Abs. 11 (10): 9. 1958.)
1950. BACTERIAL FISH SPOILAGE AND ITS CONTROL. Food Technol. 4: 477-480.
(217) Hiscox, E. R., and BRIGGS, C. A. E. 1953. REVIEWS OF THE PROGRESS OF DAIRY SCIENCE. B. BACTERIOLOGY
AND MYCOLOGY APPLIED TO DAIRYING. Jour. Dairy Res. 20: 381-429. (Abstract in Food Sei. Abs. 26: 746. 1954.)
(218) and BRIGGS, C. A. E. 1955. REVIEWS OF THE PROGRESS OF DAIRY SCIENCE. B. BACTERIOLOGY
AND MYCOLOGY APPLIED TO DAIRYING. Jour. Dairy Res. 22: 391-424. (Abstract in Food Sei. Abs. 28: 1842. 1956.)
(219) HoAGLAND, R., MCBRYDE, C. N., and PowiCK, W. C. 1917. CHANGES IN FRESH BEEF DURING COLD STORAGE AROVE FREEZING.
U.S. Dept. Agr. Dept. Bui. 433, 100 pp. (220) HOBBS, B. C, REEVES, J. C, GARSIDE, J. S., and OTHERS.
1960. ANTIBIOTIC TREATMENT OF POULTRY IN RELATION TO SALMONELLA TYPHIMURIUM. [Gt.Brit.] Min. Health and the Pub. Health Lab. Serv., Monthly Bui. 19: 178-193.
(221) HOROWITZ-WLASSOWA, L. M., and GRINBERG, L. D. 1933. ZUR FRAGE üBER PSYCHROPHILE MIKROBEN. Zentbl. f. Bakt.
[etc.], Abt. 11,89: 54-62. (222) HUCKER, G. J.
1954. LOW TEMPERATURE ORGANISMS IN FROZEN VEGETABLES. Food Technol. 8: 79-82.
(223) BROOKS, R. F., and EMERY, A. J. 1952. THE SOURCE OF BACTERIA IN PROCESSING AND THEIR SIGNIFICANCE
IN FROZEN VEGETABLES. Food Technol. 6: 147-155. (224) and DAVID, E. R.
1957. THE EFFECT OF ALTERNATE FREEZING AND THAWING ON THE TOTAL FLORA OF FROZEN CHICKEN PIES. Food Technol. 11: 354-356.
(225) and DAVID, E. R. 1957. THE EFFECT OF ALTERNATE FREEZING AND THAWING ON THE
TOTAL FLORA OF FROZEN VEGETABLES. Food Technol. 11: 381- 383.
(226) and ROBINSON, W. B. 1950. BACTERIA IN FROZEN VEGETABLES. Food in Canada 11 (3) :
14-15. (227) HUNTER, A. C.
1920. BACTERIAL DECOMPOSITION OF SALMON. Jour. Bact. 5: 353-361. (228)
1920. BACTERIAL GROUPS IN DECOMPOSING SALMON. Jour. Bact. 5: 543-552.
(229) INGRAHAM, J. L. 1958. GROWTH OF PSYCHROPHILIC BACTERIA. Jour. Bact. 76: 75-80.
(230) 1962. NEWER CONCEPTS OF PSYCHROPHILIC BACTERIA. Campbell Soup
Co., Camden, N.J., Low Temp. Microbiol. Symposium 1961: 41-56.
(231) and BAILEY, G. F. 1959. COMPARATIVE STUDY OF EFFECT OF TEMPERATURE ON METABOLISM
OF PSYCHROPHILIC AND MESOPHILIC BACTERIA. Jour. Bact. 77: 609-613.
(232) and STOKES, J. L. 1959. PSYCHROPHILIC BACTERIA. Bact. Rev. 23: 97-108.

92 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(233) INGRAM, M. 1935. THE MICROBIOLOGY OF PORK AND BACON. EFFECT OF TEMPERA-
TURE ON THE FORMATION OF SPORANGIA BY THAMNIDIUM OHAE- TOCLADIODES. [Gr.Brit.] Dept. Sei. and Indus. Res., Food In- vest. Bd. Rpt. 1934: 80-82.
FATIGUE MUSCULAIRE, VHy ET PROLIFÉRATION BACTERIENNE DANS LA VIANDE. Inst. Pasteur [Paris] Ann. 75: 139-146.
BENJAMIN WARD RICHARDSON LECTURE. II. HYGIENE AND STOR- AGE. Roy. Sanit. Inst. Jour. 69: 39-47.
THE EFFECT OF COLD ON MICRO-ORGANISMS IN RELATION TO FOOD. Soc. Appl. Bact. [Gt.Brit.] Proc. 14: 243-260.
MICROBIAL ASSOCIATION OF SEMIPRESERVED MEATS. Inst. Pasteur [Lille] Ann. 7: 32-45.
FACTORS CONTROLLING THE OCCURRENCE OF VARIOUS BACTERIA IN SEMIPRESERVED MEATS. Inst. Pasteur [Lille] Ann. 7: 136-145.
MICRO-ORGANISMS RESISTING HIGH CONCENTRATIONS OF SUGARS OR SALTS. In Microbial Ecology, Soc. for Gen, Microbiol., Symposium 7: 91-133.
L'IMPORTANCE DU POTENTIEL D'OXYDO-RéDUCTION POUR LA MICRO- BIOLOGIE DE LA VIANDE. Rev. des Ferment, et des Indus. Ali- ment., [Brussels] 13: 115-121.
L'IMPORTANCE DU PH POUR LA MICROBIOLOGIE DE LA VIANDE. Rev. des Ferment, et des Indus. Aliment. [Brussels] 13: 139- 146.
MICROBIOLOGICAL PRINCIPLES IN PREPACKAGING MEATS. Jour. Appl. Bact. 25: 259-281. and BARNES, E. M. STERILIZATION BY MEANS OF OZONE. Jour. Appl. Bact. 17: 246-271.
• and BARNES, E. M. PROBLEMS IN THE USE OF ANTIBIOTICS FOR PRESERVING MEAT. Jour. Appl. Bact. 18: 549-564. and BARNES, E. M. PROBLEMS IN THE USE OF ANTIBIOTICS FOR PRESERVING MEAT AND FISH. (Abstract in Food Sei. Abs. 28: 121-136. 1956.) and BROOKS, J. BACTERIOLOGICAL STANDARDS FOR PERISHABLE FOODS. (D) EGGS AND EGG PRODUCTS. Roy. Sanit. Inst. Jour. 72: 411-421. (Ab- stract in Food Sei. Abs. 26: 660. 1954.)
and HAINES, R. B. INHIBITION OP BCTERIAL GROWTH BY PURE OZONE IN THE PRES- ENCE OF NUTRIENTS. Jour. Hyg. 47: 146-158. and KiTCHELL, A. G. THE MICROBIOLOGY OF MEAT SPOILAGE. Inst. Meat Bul. 11, pp. 1-7.
and SHEWAN, J. M. 1960. INTRODUCTORY REFLECTIONS ON THE PSEUDOMONAS-ACHROMO-
BACTER GROUP. Jour. Appl. Bact. 23: 373-378. (250) INSTITUTE FOR RESEARCH ON STORAGE AND PROCESSING OF HORTICUL-
TURAL PRODUCE (Wageningen, The Netherlands). 1961. REFRIGERATED STORAGE OF FRUITS AND VEGETABLES. Inst. Inter-
nal, du Froid Bul., Annexe 1961-1, 374 pp. (251) INTERNATIONAL INSTITUTE OF REFRIGERATION.
1963. RECOMMENDED CONDITIONS FOR LAND TRANSPORT OF PERISHABLE FOODSTUFFS. Ed. 2, 14 pp. Annexe, Internatl. Inst. Refrig. Bul., Commissions 4 and 7.
\iutJ^J
1948.
(235) 1949.
(236) 1951.
(237) 1955.
(238) 1955.
(239) 1957.
(240) 1958.
(241) 1958.
(242) 1962.
(243) 1954.
(244) 1955.
(245) 1956.
(246) 1952.
(247) 1949.
(248) 1955.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 93
(252) JACOBSON, M., SPENCER, J., HARD, M., and WELLER, M. 1959. THE EFFECTS OF ANTIBIOTIC TREATMENTS, PACKAGING FILMS, AND
STORAGE UPON FLAVOR, ODOR, AND APPEARANCE OF CUT-UP FRYERS. Poultry Sei. 38: 40-47.
(253) JAMES, L. H. 1937. ABSTRACTS OF LITERATURE ON REFRIGERATION. Refrig. Engin.
34: 237, 244, 246, 248. (254) JARVIS, N. D.
1943. PRINCIPLES AND METHODS IN THE CANNING OF FISHERY PROD- UCTS. U.S. Fish and Wildlife Serv. Res. Rpt. 7, 366 pp.
(255) JAY, J. M., WEISER, H. H., and DEATHERAGE, F. E. 1956. THE EFFECT OF ANTIBIOTICS ON THE MICROFLORA OF BEEF AND
STUDIES ON THE MODE OF ACTION OF THIS ANTIBIOTIC IN THE PRESERVATION OF MEAT. Antibiotics Ann. 1956-1957: 954-965.
(256) WEISER, H. H., and DEATHERAGE, F. E. 1957. STUDIES ON THE MODE OF ACTION OF CHLORTETRACYCLINE (AURE-
OMYCIN) IN THE PRESERVATION OF BEEF. (Abstract A37) SoC. Amer. Bact., Bact. Proc. for 1957: 19.
(257) WEISER, H. H., and DEATHERAGE, F. E. 1957. STUDIES ON THE MODE OF ACTION OF CHLORTETRACYCLINE IN THE
PRESERVATION OF BEEF. Appl. Microbiol. 5: 400-405. (258) WEISER, H. H., and DEATHERAGE, F. E.
1957. FURTHER STUDIES ON THE PRESERVATION OF BEEF WITH CHLOR- TETRACYCLINE. Food Technol. 11: 563-566.
(259) WEISER, H. H., and DEATHERAGE, F. E. 1958. THE INHIBITION IN BEEF OF CHLORTETRACYCLINE-RESISTANT BAC-
TERIA BY SUB-BACTERIOSTATIC CONCENTRATIONS OF THE ANTI- BIOTIC. Appl. Microbiol. 6: 343-346.
(260) JAYE, M., KITTAKA, R. S., and ORDAL, Z. J. 1962. THE EFFECT OF TEMPERATURE AND PACKAGING MATERIAL ON THE
STORAGE LIFE AND BACTERIAL FLORA OF GROUND BEEF. FOOD Technol. 16 (4) : 95-98.
(261) JENSEN, L. B. 1944. MICROBIOLOGICAL PROBLEMS IN THE PRESERVATION OF MEATS.
Bact. Rev. 8: 161-188. (262) —
1949. MEAT AND MEAT FOODS. 218 pp. The Ronald Press, N.Y. (263)
1954. MICROBIOLOGY OF MEATS. Ed. 3. 422 pp. The Garrard Press, Champaign, 111.
(264) and GRETTIE, D. P. 1937. ACTION OF MICRO-ORGANISMS ON FATS. Food Res. 2: 97-120.
(265) JEPSEN, A. 1945. BIOLOGISKE PROBLEMER I FORBINDELSE MED K0LING OG FRYSNING
AF K0D. [BIOLOGICAL PROBLEMS IN CONNECTION WITH CHILLING AND FREEZING OF MEAT]. I AND II. Dansk Teknisk Tidsskr. 69 (2): 31-34; (3): 47-50.
(266) JEZESKI, J. J. 1954. THE MICROBIOLOGY OF DAIRY PRODUCTS. Ann. Rev. Microbiol.
8: 429-448. (267) and OLSON, R. H.
1962. THE ACTIVITY OF ENZYMES AT LOW TEMPERATURES. Campbell Soup Co., Camden, N.J., Low Temp. Microbiol. Symposium Proc. 1961: 139-155.
(268) JONES, A. H. and FERGUSON, W. E. 1956. THE EVALUATION OF FROZEN VEGETABLES FOR QUALITY. Canad.
Food Indus. 27 (9) : 24, 27, 29, 31. (269) JONES, O.
1941. THE CONTROL OF MEAT SPOILAGE. PART 1. Food 10: 327-328. (270)
1942. THE CONTROL OF MEAT SPOILAGE. PART 2. Food 11: 25-27. (271)
1942. THE CONTROL OF MEAT SPOILAGE. PART 3. Food 11: 36-38.

94 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(272) JoSLYN, M. A., and MARSH, G. L. 1933. CHANGES OCCURRING DURING FREEZING, STORAGE AND THAWING
OF FRUITS AND VEGETABLES. Calif. Agr. Expt. Sta. Bul. 551, 40 pp.
(273) and SHERRILL, M. 1933. INVERSION OF SUCROSE BY INVERTASE AT LOW TEMPERATURES.
Indus, and Engin. Chem. Indus. Ed., 25: 416-417. (274) KAESS, G.
1934. DAS WACHSTUM VON SCHIMMELPILZEN AUF GEKÜHLTEM FLEISCH BEI VERSCHIEDENEN LUFTZUSTÄNDEN^. Ztschr. f. die Gesam. Kälte Indus. 41: 96-102.
(275) 1934. DER EINFLUSS DER LUFTBEWEGUNG AUF DAS WACHSTUM VON
SCHIMMELPILZEN AUF GEKÜHLTEM FLEISCH. Ztschr. f. die Gesam. Kälte Indus. 41: 153-156.
(276) KAHLENBERG, 0. J. ^ ^_ 1949. HOW TO DEFROST FROZEN EGGS. Mfg. Confectioner 29 (6) : 58.
(Abstract in Food Sei. Abs. 23: 1497. 1951.) (277) KALLERT, E.
1928. VERGLEICHENDE UNTERSUCHUNGEN üBER DIE HALTBARKEIT VON HACKFLEISCH AUS FRISCHFLEISCH UND GEFRIERFLEISCH. Berlin Tierärztl. Wochnschr. 44: 547-551.
(278) KATES, M. and BAXTER, R. M. 1962. LIPID COMPOSITION OF MESOPHILIC AND PSYCHROPHILIC YEASTS
(CANDIDA SPECIES) AS INFLUENCED BY ENVIRONMENTAL TEMPER- ATURE. Canad. Jour. Biochem. and Physiol. 40: 1213-1227.
(279) KAYSER, W. 1937. NEUE UNTERSUCHUNGEN ÜBER DIE BAKTERIELLE ZERSETZUNG DER
FISCHE. Berlin. Tierärztl. Wochnschr. 53: 72-76. (280) KlERMEIER, F.
1947. THE INFLUENCE OF FREEZING ON THE ACTIVITY OF ENZYMES. Biochem. Ztschr. 318 (2/3) : 275-296. (Abstract in Refrig. Engin. 57: 657. 1949.)
(281) 1953. [INFLUENCE OF THE RELATIVE HUMIDITY OF THE ATMOSPHERE
AND PRESERVATIVE LIQUID ON EGGS DURING STORAGE] ZtSchr. f. Lebensm. Untersuch, u. Forsch. 97: 11-16 (Abstract in Chem. Abs. 47: 10147a. 1953.)
(282) KiMATA, M. 1938. RELATION BETWEEN AUTOLYTIC AND BACTERIAL AGENCIES IN THE
DECOMPOSITION OF THE MUSCLE OF SOME AQUATIC ANIMALS. Jap. See. Sei. Fisheries Bul. 7: 63. (Abstract 767 in [GtBrit.] Dept. Sei. and Indus. Res., Index to Lit. of Food Invest. 11: 130.)
(283) KiMURA, T. 1958. STUDIES IN THE BACTERIOLOGICAL CHEMISTRY OF SHARK MUSCLE
SPOILAGE. VI. ON THE EFFECTS OF TEMPERATURE UPON THE UREA DECOMPOSING ABILITY OF THE UREA-SPLITTING BACTERIA ISOLATED FROM SHARK MUSCLE. Hokaido Univ. (Japan), Fac- ulty Fisheries Bul. 8: 310-313.
(284) KIRSCH, R. H., BERRY, F. E., BALDWIN, C. L., and FOSTER, E. M. 1952. THE BACTERIOLOGY OF REFRIGERATED GROUND BEEF. Food Res.
17: 495-503. (285) KiSER, J. S.
1944. EFFECTS OF TEMPERATURES APPROXIMATING 0° C. UPON THE GROWTH AND BIOCHEMICAL ACTIVITIES OF BACTERIA ISOLATED FROM MACKEREL. Food Res. 9: 257-267.
(286) KiTCHELL, A. G., and INGRAM, M. 1956. A COMPARISON OF BACTERIAL GROWTH ON FRESH MEAT AND ON
FROZEN MEAT AFTER THAWING. Inst. Pasteur [Lille] Ann. 8: 121-131.
(287) and INGRAM, M. 1959. THE SURVIVAL OF BACTERIA ON FROZEN MEAT AND THEIR GROWTH
AFTER THAWING. Tenth Internatl. Cong. Refrig. Proc. 3: 65-70.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 95
(288) KOHMAN, E. F. 1960. FRIGI-CANNING. Food Technol. 14: 254-256.
(289) KoONZ, C. H. 1945. POST-MORTEM PRESERVATION OF POULTRY QUALITY In Jensen,
L. B., ed., Microbiology of Meats. Ed. 2, pp. 201-231. The Garrard Press, Champaign, 111.
(290) 1946. PROPER CHILLING KEEPS POULTRY SALABLE. U.S. Effff & Poultrv
Mag. 52: 15-16,39-41. ^^ ^ (291) KRAFT, A. A., and AYRES, J. C.
1952. POST-MORTEM CHANGES IN STORED MEATS. IV. EFFECT OF PACKAGING MATERIALS ON KEEPING QUALITY OF SELF-SERVICE MEATS. Food Technol. 6: 8-12.
(292) ELLIOTT, L. E., and BRANT, A. W. 1958. THE SHELL MEMBRANE AS A BARRIER TO BACTERIAL PENETRATION
OF EGGS. Poultry Sei. 37: 238-240. (293) MCNALLY, E. H., and BRANT, A. W.
1958. SHELL QUALITY AND BACTERIAL INFECTION OF SHELL EGGS. Poultry Sei. 37: 638-644.
(294) KRISS, A. E.
1959. MARINE MICROBIOLOGY (DEEP WATER). Acad. Sci., MoSCOW, .c^r^. xr.. U.S.S.R. (Abstract in World Fisheries Abs. 12 (4) : 5. 1961). (295) KUHLWEIN, H.
1952. MIKROBIOLOGISCHE GRUNDLAGEN DER LEBENSMITTELFRISCHHALT- UNG. In Plank, R., ed., Biochemische Grundlagen der Lebens- mittelfrischhaltung, being Handbuch der Kältetechnik, v. 9.
,«^.x -r PP- 167-222. Springer-Verlag. Berlin. (296) LAWRENCE, N. L., WILSON, D. C, and PEDERSON, C. S.
1959. THE GROWTH OF YEASTS IN GRAPEJUICE STORED AT LOW TEMPER- ATURES. II. THE TYPES OF YEAST AND THEIR GROWTH IN PURE CULTURE. Appl. Microbiol. 7: 7-11.
(297) LAWTON, W. C, and NELSON, F. E. 1955. INFLUENCE OF SUBLETHAL TREATMENT WITH HEAT OR CHLORINE
ON THE GROWTH OF PSYCHROPHILIC BACTERIA. Jour. Dairv Sci 38: 380-386.
(298) LEES, K. A. 1958. A SURVEY OF THE ANTIBIOTICS. Pharm. Jour. [London! 181:
163-166. (299) LEISTNER, L.
1956. DIE NEUZEITLICHE VERPACKUNG IN DER FLEISCHWIRTSCHAFT. I. BAKTERIOLOGISCHE PROBLEME BEI DER VORVERPACKUNG VON FLEISCH UND FLEISCHWAREN. Fleischwirtschaft. 8: 422. 425 427-429.
(300) 1957. DIE NEUZEITLICHE VERPACKUNG IN DER FLEISCHWIRTSCHAFT.
II. DIE VAKUUMVERPACKUNG FÜR FLEISCHWAREN, BAKTERIOLOG- ISCH BETRACHTET. Fleischwirtschaft. 9: 262-264.
(301) LEPPER, H. A., BARTRAM, M. T., and HILLIG, F. 1944. DETECTION OF DECOMPOSITION IN LIQUID, FROZEN AND DRIED
EGGS. Assoc. Off. Agr. Chem. Jour. 27: 204-223. (302) BARTRAM, M. T., and HILLIG, F.
1956. AUTHENTIC PACKS OF EDIBLE AND INEDIBLE FROZEN EGGS AND THEIR ORGANOLEPTIC, BACTERIOLOGICAL AND CHEMICAL EXAMINA- TION. Assoc. Off Agr. Chem. Jour. 39: 185-194.
(303) LiEBMAN, H. L., KUDO, G., CHAPEL, J. G., and STERN, J. A. 1957. EFFECTS OF TEMPERATURE UPON THE STORAGE LIFE OF FRESH
SHUCKED PACIFIC OYSTERS (osTREA GiGAs). Com. Fisheries Rev. 19 (12) : 1-4.
(304) LINDERHOLM, K. G.
1960. BAKTRERIOLOGISKA OCH HYGIENISKA UNDERSÖKNINGAR AV VAKU- UMFÖRPACKADE CHARKUTERIVAROR. Nord. Hvff. Tidskr 41 (1-2): 17-38.
(305) LiNEWEAVER, H. 1939. THE ENERGY OF ACTIVATION OF ENZYME REACTIONS AND THEIR
VELOCITY BELOW 0°. Amer. Chem. Soc. Jour. 61: 403-408.

96 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(306) LoCHHEAD, A. G., and LANDERKIN, G. B. 1935. BACTERIOLOGICAL STUDIES OF DRESSED POULTRY. I. PRELIMINARY
INVESTIGATIONS OP BACTERIAL ACTION AT CHILL TEMPERATURES. Sei. Agr. 15: 765-770.
(307) LONG, S. K., and WILLIAMS, O. B. 1959. GROWTH OF OBLIGATE THERMOPHILES AT 37 C. AS A FUNCTION OF
THE CULTURAL CONDITIONS EMPLOYED. Jour. Bact. 77: 545-547. (308) LORENZ, F. W., STARR, P. B., STARR, M. P., and OGASAWARA, F. X.
1952. THE DEVELOPMENT OF PSEUDOMONAS SPOILAGE IN SHELL EGGS. I. PENETRATION THROUGH THE SHELL. Food ReS. 17: 351-360.
(309) LÜTHI, H. 1959. MICROORGANISMS IN NONCITRUS JUICES. In Mrak, E. M., ed..
Advances in Food Res. 9: 221-284. (310) LuiJPEN, A. F. M. G.
1958. OBJECTIVE SPOILAGE TESTS FOR FISH STORED UNDER CONDITIONS OTHER THAN NORMAL CHILLING IN ICE. Jour. Sci. Food and Agr. 9: 410-417.
(311) LUYET, B. J., and GEHENIO, P. M. 1938. THE LOWER LIMIT OF VITAL TEMPERATURES; A REVIEW. Biody-
namica 1: 1-92. (312) MCFARLANE, V. H., WATSON, A. J., and GORESLINE, H. E.
1945. MICROBIOLOGICAL CONTROL IN THE PRODUCTION OF SPRAY DRIED WHITE EGG POWDER. U.S. Egg & Poultry Mag. 51: 250-257, 270-273, 275-277, 279-286.
(313) MCKEE, R. C., CONKEY, J., and CARLSON, J. A. 1959. A STUDY OF THE COMPARATIVE SHELF LIFE OF WET AND DRY-
PACKED POULTRY. Poultry Sei. 38: 260-271. (314) MCNALLY, E. H.
1952. EFFECT OF DRYING AFTER WASHING ON THE INCIDENCE OF EGGS INFECTED BY SPOILAGE MICRO-ORGANISMS. Poultry Sci. 31: 1102-1104.
(315) McViCKER, R. J., DAWSON, L. E., MALLMANN, W. L., and OTHERS. 1959. THE EFFECT OF CHLORTETRACYCLINE ON SHELFLIFE OF FRESH
POULTRY MEAT TREATED IN A COMMERCIAL PROCESSING PLANT. Poultry Sci. 38: 337-343.
(316) MCWHORTER, A. C, MURRELL, M. C. and EDWARDS, P. R. 1963. RESISTANCE OF SALMONELLAE ISOLATED IN 1962 TO CHLORTETRA-
CYCLINE. Appl. Microbiol. 11: 368-370. (317) MACY, H., and STEELE, G. H.
1934. BUTTER AS A SUBSTRATE FOR MOLD GROWTH. Jour. Dairy Sci. 17: 397-407.
(318) MADSEN, J. 1943. UNDERSjeiGELSER OVER SVINEK0DETS HOLDBARHED EFTER SUKKER-
HOLDIGE FODERMIDLER. Nord. Jordbrugsforsk. 5-6: 340-346. (319) MÄDER, K.
1960. OZON IM KÜHLHAUSBETRIEB. Kältetechnik 13 (6) : 284-286. (Abstract Internati. Inst. Refrig. Bul. 41 (3): 824. 1961.)
(320) MAGOON, C. A. 1931. KEEPING QUALITY AS RELATED TO THE DISTRIBUTION PROBLEM.
Ice and Refrig. 80 (1) : 39-41. (321) MALLMANN, W. L., and DAVIDSON, J. A.
1944. OIL-PROTECTED SHELL EGGS. U.S. Egg & Poultry Mag. 50: 113-115, 133, 169-171, 189-191.
(322) MARGINESU, P. 1932. LE CARNI DECONGELATE SI ALTERANO PIU' RáPIDAMENTE DELLE
GARNI PRESCHE? Indus. Ital. delle Conserve Aliment. 7: 232-238.
(323) MARGOLIS, L. 1953. THE EFFECT OF FASTING ON THE BACTERIAL FLORA OF THE IN-
TESTINE OF FISH. Canada Fisheries Res. Bd. Jour. 10: 62-63. (Abstract in Com. Fisheries Abs. 6 (7) : 9-10. 1953.)
(324) MARTH, E. H., and FRAZIER, W. C. 1957. BACTERIOLOGY OF MILK HELD AT FARM BULK COOLING TANK TEM-
PERATURES. HI. PSYCHROPHILES FOUND AND THEIR GROWTH. Jour. Milk and Food Technol. 20: 93-99.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 97
(325) MAY, K. N., IRBY, J. D., and CARMON, J. L. 1962. SHELF LIFE AND BACTERIAL COUNTS OF EXCISED POULTRY TISSUE.
Food Technol. 16 (2) : 66-68. (326) MiCHENER, H. D., DIETRICH, W. C, and THOMPSON, P. A.
1960. TIME-TEMPERATURE TOLERANCE OF FROZEN FOODS. XXII. RELA- TIONSHIP OF BACTERIAL POPULATION TO TEMPERATURE. Food Technol. 14: 290-294.
(327) and ELLIOTT, R. P. 1964. MINIMUM GROWTH TEMPERATURES FOR FOOD POISONING, FECAL
INDICATOR, AND PSYCHROPHILIC MICROORGANISMS. In Chiches- ter, C. 0., ed., Advances in Food Res. 13: 349-396.
(328) MILLER, D. D., and GOLDING, N. S. 1949. THE GAS REQUIREMENTS OF MOLD. V. THE MINIMUM OXYGEN
REQUIREMENTS FOR NORMAL GROWTH AND FOR GERMINATION OF SIX MOLD CULTURES. Jour. Dairy Sci. 32: 101-110.
(329) MILLER, W. A. 1954. THE MICROBIOLOGY OF DIRTY EGGS TREATED IN VARIOUS WAYS AND
STORED AT DIFFERENT TEMPERATURES AND HUMIDITIES. Poultry Sei. 33: 735-742.
(330)
(331)
(332)
1956. THE EFFECT OF COATING THE SHELLS OF WASHED EGGS THAT FORMERLY WERE DIRTY, WITH ANTIBIOTICS, UPON SUBSEQUENT SPOILAGE. Poultry Sei. 35: 241-243.
1957. A COMPARISON OF VARIOUS WASH-WATER ADDITIVES IN PREVENT- ING MICROBIAL DETERIORATION IN WASHED EGGS THAT FORMERLY WERE DIRTY. Poultry Sei. 36: 579-584.
1959. DRY CLEANING SLIGHTLY SOILED EGGS VERSUS WASHING TO PRE- VENT PENETRATION OF SPOILAGE BACTERIA. Poultry Sci. 38: 906-910.
(333) and CRAWFORD, L. B. 1953. SOME FACTORS INFLUENCING BACTERIAL PENETRATION OF EGGS.
Poultry Sci. 32: 303-309. (334) MoNOD, J.
1949. THE GROWTH OF BACTERIAL CULTURES. Ann. Rev. Microbiol. 3 : 371-394.
(335) MORAN, T. 1935. POST-MORTEM AND REFRIGERATION CHANGES IN MEAT. TranS.
and Commun. Soc. Chem. Indus. Jour. [London] 54: 149T- 151T.
(336)
(337)
1937. INHIBITION OF MOULD ON EGGS BY CONTROL OF THE CARBON DIOXIDE AND OXYGEN CONTENT OF THE ATMOSPHERE. [Gt. Brit.] Dept. Sci. and Indus. Res., Food Invest. Bd. Rpt. 1936: 53-55.
1938. GAS STORAGE OF MEAT AND EGGS. Food ReS. 3: 149-154. (338) MORRISON, T. F.
1925. STUDIES ON LUMINOUS BACTERIA. II. THE INFLUENCE OF TEM- PERATURE ON THE INTENSITY OF THE LIGHT OF LUMINOUS BAC- TERIA. Jour. Gen. Physiol. 7: 741-753.
(339) MosSEL, D. A. A. 1952. DE BETEKENIS VAN DE ZGN. BEDERF-ASSOCIATIE VOOR DE LEVENS-
MIDDELEN-MICROBIOLOGIE. Tijdschr. over Plantenziek. 58: 267- 268. (Abstract in Food Sci. Abs. 27: 2522. 1955.)
(340) and INGRAM, M. 1955. THE PHYSIOLOGY OF THE MICROBIAL SPOILAGE OF FOODS. Jour.
Appl. Bact. 18: 232-268. (341) and WESTERDIJK, J.
1949. THE PHYSIOLOGY OF MICROBIAL SPOILAGE IN FOODS. Antonie van Leeuwenhoek Jour. Microbiol. and Serol. 15: 190-202.
(342) MouTNEY, G. J. and VAN DER ZANT, C. 1958. GREEN ROT IN EGGS. Tex. Agr. Expt. Sta. Bui. MP-315, 8 pp.

98 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(343) MÜLLER, M. 1903. ÜBER DAS WACHSTUM UND DIE LEBENSTÄTIGKEIT VON BAKTERIEN,
SOWIE DEN ABLAUF FERMENTATIVER PROZESSE BEI NIEDERER TEM- PERATUR UNTER SPEZIELLER BERÜCKSICHTIGUNG DES FLEISCHES ALS NAHRUNGSMITTEL. Arch. f. Hyg. 47: 127-193.
(344) MuLCOCK, A. P. 1955. SPOILAGE IN FROZEN PEAS. New Zeal. Jour. Sei. and Technol.,
Section B, 37: 15-19. (345) MUNDT, J. 0.
1961. OCCURRENCE OF ENTEROCOCCI ON PLANTS: BUD AND BLOSSOM STUDIES. (Abstract) Soc. Amer. Bact., Bact. Proc. for 1961: A42.
(346) STOKES, R. L. and GOFF, 0. E. 1954. THE SKIN OF BROILERS AS A BARRIER TO BACTERIAL INVASION
DURING PROCESSING. Poultry Sei. 33: 799-802. (347) MURDOCK, C. R., CROSSLEY, E. L., ROBB, J., and OTHERS.
1960. THE PASTEURIZATION OF LIQUID WHOLE EGG. [Gt. Brit.] Med. Res. Council Monthly Bui. 19: 134-152.
(348) MURDOCK, D. I. and BROKAW, C. H. 1961. SOME OBSERVATIONS ON HOLDING 6-OZ. CANS OF FROZEN CONCEN-
TRATED ORANGE JUICE AT ROOM TEMPERATURE AND AT 40** F. A PRELIMINARY REPORT. Fla. State Hort. Soc. Proc. (1961) 74: 271-278.
(349) NADEAU, P. A. 1940. FRESH FISH. II. THE ROLE OF PH IN THE GROWTH OF BACTERIA.
Canada Fisheries Res. Bd. Jour. 5: 121-130. (350) NAGEL, C. W., SIMPSON, K. L., NG, H., and OTHERS.
1960. MICRO-ORGANISMS ASSOCIATED WITH SPOILAGE OF REFRIGERATED POULTRY. Food Technol. 14: 21-23.
(351) NASHIF, S. A., and NELSON, F. E. 1953. THE LIPASE OF PSEUDOMONAS FRAGI. I. CHARACTERIZATION OF
THE ENZYME. Jour. Dairy Sei. 36: 459-470. (352) and NELSON, F. E.
1953. THE LIPASE OF PSUEDOMONAS FRAGI. II. FACTORS AFFECTING LIPASE PRODUCTION. Jour. Dairy Sei. 36: 471-480.
(353) and NELSON, F. E. 1953. THE LIPASE OF PSEUDOMONAS FRAGI. III. ENZYME ACTION IN
CREAM AND BUTTER. Jour. Dairy Sei. 36: 481-488. (354) NETHERLANDS MINISTRY OF AGRICULTURE, FISHERIES, AND FOOD.
1960. CHILLING OF FISH. 276 pp. Ministry of Agriculture, Fisheries, and Food, The Hague, Netherlands. (Abstract in World Fisheries Abs. 12 (1) : 25. 1961.)
(355) NEWELL, G. W., GWIN, J. M., and JULL, M. A. 1948. THE EFFECT OF CERTAIN HOLDING CONDITIONS ON THE QUALITY
OF DRESSED POULTRY. Poultry Sei. 27: 251-256. (356) NG, H., INGRAHAM, J. L., and MARR, A. G.
1962. DAMAGE ANO DEPRESSION IN ESCHERICHIA COLI RESULTING FROM GROWTH AT LOW TEMPERATURES. Jour. Bact. 84: 331- 339.
(357) VAUGHN, R. H., STEWART, G. F., and OTHERS. 1957. ANTIBIOTICS IN POULTRY MEAT PRESERVATION. DEVELOPMENT OF
RESISTANCE AMONG SPOILAGE ORGANISMS. Appl. Microbiol. 5: 331-333.
(358) NiLSSON, T., and REINIUS, L. 1959. EFFEKTEN AV SNABB NEDKYLNING PÂ DEN HYGIENISKA KVALITA-
TEN AV KYCKLING KÖTT. [THE EFFECT OF QUICK COOLING ON THE HYGIENIC QUALITY OF CHICKEN MEAT.] Kyltekn. Tidskr. 18: 103-104. [In Swedish.] (Abstract in Internatl. Inst. Refrig. Bui. 40: 192. 1960.)
(359) NiVEN, C. F., JR. 1948. CAUSES OF GREEN RING SPOILAGE IN SAUSAGE PRODUCTS AND HOW
TO AVOID SUCH TROUBLE. Nati. Provisioner 119 (14) : 112-113. (360)
1951. INFLUENCE OF MICROBES UPON THE COLOR OF MEATS. Amer. Meat Inst. Found. Cir. 2, 11 pp.

PSYCHROPHILIC MICRO-ORGANISMS IN POODS 99
(361) NiVEN, C. F., JR.
1951. SAUSAGE DISCOLORATIONS OF BACTERIAL ORIGIN. Amer. Meat Inst. Found. Bui 13, 24 pp.
(362) and CHESBRO, W. R. 1960. MEAT PRESERVATION. CHEMICALS AND ANTIBIOTICS. In Amer.
Meat Inst. Found., The Science of Meat and Meat Products. 438 pp., pp. 309-327. W. H. Freeman and Co., San Francisco and London.
(363) NJOKU-OBI, A. N., SPENCER, J. V., SAUTER, E. A., and EKLUND, M. W. 1957. A STUDY OF THE FUNGAL FLORA OF SPOILED CHLORTETRACYCLINE
TREATED CHICKEN MEAT. Appl. Microbiol. 5: 319-321. (364) NoTEVARP, 0., and HJORTH-HANSEN, S.
1943. RENHOLDETS BETYDNING FOR FISK OG FISKEVARERS KVALITET. [SIGNIFICANCE OF SANITATION IN THE QUALITY OF FISH AND
FISH PRODUCTS.] Fiskeridir. Smâskr. 1: 1-15. (365) OGILVY, W. S. and AYRES, J. C.
1951. POST-MORTEM CHANGES IN STORED MEATS. II. THE EFFECT OF ATMOSPHERES CONTAINING CARBON DIOXIDE IN PROLONGING THE STORAGE LIFE OF CUT-UP CKHCKEN. Food Technol. 5: 97-102.
(366) and AYRES, J. C. 1951. POST-MORTEM CHANGES IN STORED MEATS. III. THE EFFECT OF
ATMOSPHERES CONTAINING CARBON DIOXIDE IN PROLONGING THE STORAGE LIFE OF FRANKFURTERS. Food Technol. 5: 300-303.
(367) OLSEN, R. H., and JEZESKI, J. J. 1963. SOME EFFECTS OF CARBON SOURCE, AERATION, AND TEMPERATURE
ON GROWTH OF A PSYCHROPHILIC STRAIN OF PSEUDOMONAS FLOURESCENS. Jour. Bact. 86: 429-433.
(368) OLSON, J. C, JR.
1947. BACTERIA IN DAIRY PRODUCTS. PSYCHROPHILES, MESOPHILES, THERMOPHILES AND THERMODURICS. WHAT ARE WE TALKING ABOUT? Milk Plant Monthly. 36 (11) : 32-36.
(369) PARKER, R. B., and MUELLER, W. S. 1955. THE NATURE, SIGNIFICANCE AND CONTROL OF PSYCHROPHILIC
BACTERIA IN DAIRY PRODUCTS. Jour. Milk and Food Technol. 18: 200-203.
(370) WiLLouGHBY, D. S., THOMAS, E. L., and MORRIS, H. A. 1953. THE KEEPING QUALITY OF PASTEURIZED MILK AS INFLUENCED BY
THE GROWTH OF PSYCHROPHILIC BACTERIA AND THE ADDITION OF AUREOMYCIN. Jour. Milk and Food Technol. 16: 213-219.
(371) PANES, J. J., and THOMAS, S. B. 1959. THE MULTIPLICATION OF COLI-AEROGENES BACTERIA IN MILK
STORED AT 3-5°. Jour. Appl. Bact. 22: 272-277. (372) PARTMANN, W.
1957. ANTIBIOTICS IN FOOD PRESERVATION. Ztschr. f. Lebensmtl.- Untersuch. u.-Forsch. 106: 210-227. (Abstract in Chem. Abs. 52: 595. 1958.)
(373) PATRICK, R. and HILL, E. C.
1959. MICROBIOLOGY OF CITRUS FRUIT PROCESSING. Fla. Aer. Exnt. Sta. Bui. 618, 62 pp.
(374) PEARCE, J. A., and LAYERS, C. G. 1949. FROZEN STORAGE OF POULTRY, V. EFFECTS OF SOME PROCESSING
FACTORS ON QUALITY. Canad. Jour. Res., Sect. F., 27: 253-265. (375) PEDERSON, C. S.
1947. SIGNIFICANCE OF BACTERIA IN FROZEN VEGETABLES. Food ReS. 12: 429-438.
(376) ALBURY, M. N., and CHRISTENSEN, M. D. 1961. THE GROWTH OF YEASTS IN GRAPE JUICE STORED AT LOW TEMPER-
ATURE. IV. FUNGISTATIC ACTION OF ORGANIC ACIDS. Appl. Microbiol. 9: 162-167.
(377) PENFOLD, W. J. 1914. ON THE NATURE OF BACTERIAL LAG. Jour. Hvff. 14: 215-241.
(378) PENNINGTON, M. E. 1910. STUDIES OF POULTRY FROM THE FARM TO THE CONSUMER. U.S.
Dept. Agr., Bur. Chem. Cir. 64, 42 pp.

100 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(3791 PENNINGTON, M. E., WITMER, E., and PIERCE, H. C. 1911. THE COMPARATIVE RATE OF DECOMPOSITION IN DRAWN AND UN-
DRAWN MARKET POULTRY. U.S. Dept. AgT., Bur. Chem. Cir. 70, 22 pp.
(380) PENNISTON, V. A., and HEDRICK, L. R. 1947. THE REDUCTION OF BACTERIAL COUNT IN EGG PULP BY USE OF
GERMICIDES IN WASHING DIRTY EGGS. Food Technol. 1: 240- 244.
(381) PETERSON, A. C. T ^ i. n O n 1962. AN ECOLOGICAL STUDY OF FROZEN FOODS. In Campbell bOUp CO.,
Camden, N.J. Low Temp. Microbiol. Symposium 1961: 157- 195.
(382) and GUNDERSON, M. F. 1960. ROLE OF PSYCHROPHILIC BACTERIA IN FROZEN FOOD SPOILAGE.
Food Technol. 14: 413-417. (383) and GUNDERSON, M. F.
1960. SOME CHARACTERISTICS OF PROTEOLYTIC ENZYMES FROM PSEUDOM- ONAS FLUORESCENS. Appl. MicrobioL 8: 98-104.
(384) PiPPEN, E. L., and KLOSE, A. A. 1955 EFFECTS OF ICE WATER CHILLING ON FLAVOR OF CHICKEN. Poul-
try Sei. 34: 1139-1146. (385) PLAZIKOWSKI, U.
1958. HÂLLBARHETEN HOS FÖRPAKADE KÓTT-OCH CHARKUTERIVAROR. Nord. Hyg. Tidskr. 39 (3-4) : 61-71.
(386) PLISZKOWA, A. 1958. KILKA UWAG W SPRAWIE MIKROFLORY SZYNEK PASTERYZOW-
ANYCH. [SOME REMARKS ON THE MICROFLORA OF PASTEURIZED HAMS]. Przem. Spozywczy 12 (4) : 12-15. (English translation in OTS 60: 21511. 1960.)
(387) PORTER, J. R. ^^^^ _ , „_.. 1946. BACTERIAL CHEMISTRY AND PHYSIOLOGY. 1073 pp. John Wiley
and Sons, Inc., New York. (388) PRECHT, H., CHRISTOPHERSON, J., and HENSEL, H.
1955. TEMPERATURE UND LEBEN. 514 pp. Springer-Verlag, Berhn, Göttingen, Heidelberg.
(389) PROCTOR, B. E., and GOLDBLITH, S. A. ^ ^ ^ 1957. PRESERVATION OF FOODS BY IRRADIATION. Amer. Jour. Pub.
Health 47: 439-445. (390) QuiNN, H., and GARNATZ, G. G.
1943. A STUDY OF THE INCREASE OF BACTERIAL COUNTS OF FROZEN WHOLE EGGS, USING THREE METHODS OF THAWING. (Abstract) Jour. Bact. 45: 49-50.
(391) RABOTNOVA, I. L., and MINEEVA, L. A. 1959. A STUDY OF THE LAG PHASE IN MICRO-ORGANISMS. I. EFFECT
OF EXTERNAL CONDITIONS UPON LENGTH OF LAG PHASE IN TORULOPSIS UTILIS AND PS. FLUORESCENS. Mikrobiologiia [USSR] 28: 331^335.
(392) RAMSEY, G. B., FRIEDMAN, B. A., and SMITH, M. A. 1959. MARKET DISEASES OF BEETS, CHICORY, ENDIVE, ESCAROLE, GLOBE
ARTICHOKES, LETTUCE, RHUBARB, SPINACH, AND SWEET POTATOES. U.S. Dept. Agr., Agr. Handb. 155, 42 pp.
(393) and WIANT, J. S. 1941. MARKET DISEASES OF FRUITS AND VEGETABLES: ASPARAGUS,
ONIONS, BEANS, PEAS, CARROTS. CELERY, AND RELATED VEGETA- BLES. U.S. Dept. Agr. Misc. Pub. 440, 70 pp.
(394) WiANT, J. S., and LINK, G. K. K. 1938. MARKET DISEASES OF FRUITS AND VEGETABLES: CRUCIFERS AND
CUCURBITS. U.S. Dept. Agr., Misc. Pub. 292, 74 pp. (395) WiANT, J. S., and MCCOLLOCH, L. P.
1952. MARKET DISEASES OF TOMATOES, PEPPERS, AND EGGPLANTS. U.S. Dept. Agr., Agr. Handb. 28, 54 pp.
(396) WIANT, J. S. and SMITH, M. A. 1949. MARKET DISEASES OF FRUITS AND VEGETABLES: POTATOES. U.S.
Dept. Agr., Misc. Pub. 98, 60 pp.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 101
(397) REAY, G. A., and SHEWAN, J. M. 1949. THE SPOILAGE OF FISH AND ITS PRESERVATION BY CHILLING. In
Mrak, E. M., ed., Advances in Food Res. 2: 343-398. (398) RiEMANN, H., and BRAMSNAES, F.
1954. THE PRESERVATION OF "WET" FISH. Food and Agr. Organ. U.N., Fisheries Bui. 7 (2) : 49-55. (Abstract in Com. Fish- eries Abs. 7 (12) : 3-4 1954).
(399) ROACH, S. W., HARRISON, J. S. M., TARR, H. L. A., and OTHERS. 1961. STORAGE AND TRANSPORT OF FISH IN REFRIGERATED SEA WATER.
Canada Fisheries Res. Bd. Bui. 126, 61 pp. (400) ROGERS, R. E., and MCCLESKEY, C. S.
1957. BACTERIOLOGICAL QUALITY OF GROUND BEEF IN RETAIL MARKETS. Food Technol. 11: 318-320.
(401) ROMANOFF, A. L., and ROMANOFF, A. J. 1949. THE AVIAN EGG. 918 pp. John Wiley and Sons, Inc., New York.
(402) ROSE, A. H. 1962. TEMPERATURE RELATIONSHIPS AMONG MICRO-ORGANISMS. Wal-
lerstein Labs., Commun. 25 (86) : 5-18. (403) ROSE, D. H., BROOKS, C, BRATLEY, C. O., and WINSTON, J. R.
1943. MARKET DISEASES OF FRUITS AND VEGETABLES : CITRUS AND OTHER SUBTROPICAL FRUITS. U.S. Dept. Agr. Misc. Pub. 498, 57 pp.
(404) FISHER, D. F., BROOKS, C, and BRATLEY, C. O. 1950. MARKET DISEASES OF FRUITS AND VEGETABLES : PEACHES, PLUMS,
CHERRIES, AND OTHER STONE FRUITS. U.S. Dept. Agr. MisC. Pub. 228, 27 pp.
(405) McCoLLOCH, L. P., and FISHER, D. F. 1951. MARKET DISEASES OF FRUITS AND VEGETABLES: APPLES, PEARS,
QUINCES. U.S. Dept. Agr. Misc. Pub. 168, rev., 72 pp. (406) RosSER, F. T.
1942. PRESERVATION OF EGGS. II. SURFACE CONTAMINATION ON EGG- SHELL IN RELATION TO SPOILAGE. Canad. Jour. Res., Sect. D, 20: 291-296.
(407) ROTH, N. G., and WHEATON, R. B. 1962. CONTINUITY OF PSYCHROPHILIC AND MESOPHILIC GROWTH CHAR-
ACTERISTICS IN THE GENUS ARTHROBACTER. Jour. Bact. 83: 551-555.
(408) RUSHING, N. B., PATRICK, R., and VELDHUIS, M. K. 1954. EFFECT OF CONCENTRATION OF ORANGE JUICE AND TEMPERATURE
OF STORAGE ON GROWTH AND SURVIVAL OF MICRO-ORGANISMS. Fla. State Hort. Soc. Proc. (1953) 66: 281-286.
(409) and SENN, V. J. 1962. EFFECT OF PRESERVATIVES AND STORAGE TEMPERATURES ON SHELF
LIFE OF CHILLED CITRUS SALADS. Food Technol. 16: 77-79. (410) SALTóN, M. R. J., SCOTT, W. J., and VICKERY, J. R.
1951. STUDIES IN THE PRESERVATION OF SHELL EGGS. VI. THE EFFECT OF PASTEURIZATION ON BACTERIAL ROTTING. Austral. Jour. Appl. Sei. 2: 205-222.
(411) SAMISH, Z., ETINGER-TULCZYNSKA, R., and BICK, M. 1961. MICROFLORA OF HEALTHY TOMATOES. Appl. Microbiol. 9: 20-25.
(412) SANDERSON, N. H., JR. 1941. THE BACTERIOLOGY AND SANITATION OF QUICK FROZEN FOODS.
Refrig. Engin. 42: 228-232. (413) SAUTER, E. A., PETERSEN, C. F., and LAMPMAN, C. E.
1962. THE EFFECTIVENESS OF VARIOUS SANITIZING AGENTS IN THE RE- DUCTION OF GREEN ROT SPOILAGE IN WASHED EGGS. Poultry Sci. 41: 468^73.
(414) SCHELHORN, M., VON 1951. CONTROL OF MICRO-ORGANISMS CAUSING SPOILAGE IN FRUIT AND
VEGETABLE PRODUCTS. In Mrak, E. M., ed.. Advances in Food Res. 3: 429-482.
(415) 1959. KOMBINATION VON METHODEN ZUR VERHINDERUNG DES MIKRO-
BIOLOGISCH BEDINGTEN VERDERBS VON LEBENSMITTELN. Ztschr. f. Lebensm-Untersuch. u.-Forsch. 109: 400-411.

102 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(416) SCHMID, W. 1931. EINFLUSS VON TEMPERATUR UND FEUCHTIGKEIT AUF DAS BAK-
TERIENWACHSTUM AUF GEKÜHLTEM FLEISCH. Ztschr. f. die Gesam. Kälte Indus. 38: 1-5.
(417) SCHMIDT. 1930. ÜBER DEN EINFLUSZ VON TEMPERATUR UND FEUCHTIGKEIT AUF
DAS BAKTERIENWASCHSTUM IN GEKÜHLTEM FLEISCH. Ztschr. f. Angew. Chem. 43: 657-658.
(418) SCHMIDT, F. J., and STADELMAN, W. J. 1957. EFFECTS OF ANTIBIOTICS AND HEAT TREATMENT OF SHELL EGGS
ON QUALITY AFTER STORAGE. Poultry Sci. 36: 1023-1026. (419) ScHNEiTER, R., BARTRAM, M. T., and LEPPER, H. A.
1943. BACTERIOLOGICAL AND PHYSICAL CHANGES OCCURRING IN FROZEN EGGS. Assoc. Off. Agr. Chem. Jour. 26:172-182.
(420) SCHöNBERG, F., and DEBELIC, S. 1933. UEBER FISCHFÄULNISBAKTERIEN UND VERSUCHE ZU IHRER BE-
KÄMPFUNG. Berlin. Tierärztl. Wochnschr. 49: 396-402. (421) SCHWARTZ, W., and ZEISER, TH.
1939. MIKROBIOLOGISCHE UNTERSUCHUNGEN AN SEE-UND SUSZWASSER- FISCHEN. II. DAS BAKTERIEN WACHSTUM AUF GEKÜHLTEN FI- SCHEN IN ABHÄNGIGKEIT VON DER LAGERTEMPERATUR UND DER CO2-BEGASUNG UND DIE WIRKUNG DES ABSCHLEIMENS AUF DIE HALTBARKEIT. Arch. f. Mikrobiol. 10: 322-342. Biol. Abs. 14: 9031.
(422) SCOTT, W. J. 1936. THE GROWTH OF MICRO-ORGANISMS ON OX MUSCLE. I. THE IN-
FLUENCE OF WATER CONTENT OF SUBSTRATE ON RATE OF GROWTH AT —1° C. Austral. Council Sei. and Indus. Res. Jour. 9: 177-190.
(423)
(424)
1937. THE GROWTH OF MICRO-ORGANISMS ON OX MUSCLE. II. THE INFLUENCE OF TEMPERATURE. Austral. Council Sci. and Indus. Res. Jour. 10: 338-350.
1938. THE GROWTH OF MICRO-ORGANISMS ON OX MUSCLE. III. THE INFLUENCE OF 10 PER CENT CARBON DIOXIDE ON RATES OF GROWTH AT "1** C. Austral. Council Sci. and Indus Res. Jour. 11: 266-277.
(425)
(426)
(427)
1953. WATER RELATIONS OF STAPYLOCOCCUS AUREUS AT 30° C. Austral. Jour. Biol. Sci. 6: 549-564.
1957. WATER RELATIONS OF FOOD SPOILAGE MICRO-ORGANISMS. In Mrak, E. M., ed., Advances in Food Res. 7: 83-127.
1962. AVAILABLE WATER AND MICROBIAL GROWTH. Campbell Soup Co., Camden, N.J. Low Temp. Microbiol. Symposium 1961; 89-105.
(428) and VICKEREY, J. R. 1939. INVESTIGATIONS ON CHILLED BEEF. II. COOLING AND STORAGE
IN THE MEATWORKS. Austral. Council Sci. and Indus. Res., Bui. 129, 68 pp.
(429) SEAGRAN, H., COLLINS, J., and IVERSON, J. 1960. PROCESSING AND QUALITY STUDIES OF SHRIMP HELD IN REFRIG-
ERATED SEA WATER AND ICE. Com. Fisheries Rev. 22 (5) : 1-5. (430) SELIBER, G. L. and PUMPYANSKII, A. YA.
1960. THE HISTORY OF THE MICROBIOLOGY OF BAKING. Mikrobiologiia [USSR] 28: 269-281.
(431) SHANK, J. L., and LUNDQUIST, B. R. 1963. THE EFFECT OF PACKAGING CONDITIONS ON THE BACTERIOLOGY,
COLOR, AND FLAVOR OF TABLE-READY MEATS. Food Technol. 17: 1163-1166.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 103
(432) SHANNON, W. G., and STADELMAN, W. J. 1957. THE EFFICACY OF CHLORTETRACYCLINE AT SEVERAL TEMPERA-
TURES IN CONTROLLING SPOILAGE OF POULTRY MEAT. Poultry Sei. 36: 121-123.
(433) SHARP, P. F., and STEWART, G. F. 1936. EFFECT OF RELATIVE HUMIDITY ON THE GROWTH OF MOLD ON
EGGS IN STORAGE. N.Y. (Cornell) Agr. Expt. Sta. Mem. 191, 11 pp.
(434) SHEPHERD, A. D. 1960. RELATIVE STABILITIES OF VARIOUS FROZEN FOODS. In Conference
on Frozen Food Quality. U.S. Dept. Agr., Agr. Res. Serv. ARS 74-21, pp. 18-23.
(435) SHEWAN, J. M. 1944. THE BACTERIAL FLORA OF SOME SPECIES OF MARINE FISH AND ITS
RELATION TO SPOILAGE. Soc. Agr. Bact. [Gt. Brit.] Proc. 15: 56-60. (Abstract in Biol. Abs. 20: 1233. 1946.)
(436)
(437)
(438)
1949. SOME BACTERIOLOGICAL ASPECTS OF HANDLING, PROCESSING AND DISTRIBUTION OF FISH. Roy. Sanit. Inst. Jour. 69: 394-421.
1961. THE MICROBIOLOGY OF SEA-WATER FISH. In Borgstrom, G., ed., Fish as Food. 1: 487-560. Acad. Press. New York, 726 pp.
1962. THE BACTERIOLOGY OF FRESH AND SPOILING FISH AND SOME RE- LATED CHEMICAL CHANGES. Recent Advances in Food Science 1: 167-193.
(439) and GEORGALA, D. L. 1957. THE EFFECT OF SPOILAGE AND HANDLING ON THE BACTERIAL
FLORA OF FISH. Nutr. Soc. [London] Proc. 16: 161-162. (440) HoBBS, G., and HODGKISS, W.
1960. THE PSEUDOMONAS AND ACHROMOBACTER GROUPS OF BACTERIA IN THE SPOILAGE OF MARINE WHITE FISH. Jour. Appl. Bact. 23: 463-468.
(441) SHRIMPTON, D. H. 1957. THE USE OF CHLORTETRACYCLINE (AUREOMYCIN) TO RETARD THE
SPOILAGE OF POULTRY CARCASSES. Jour. Sci. Food and Agr. 8: 485-489.
(442) 1959. QUALITY CONTROL IN MARKETING FRESH POULTRY. [Gt. Brit.]
Dept. Sei. and Indus. Res., Food Invest. Bd. Tech. Paper 7, 25 pp.
(443) and BARNES, E. M. 1960. A COMPARISON OF OXYGEN PERMEABLE AND IMPERMEABLE WRAP-
PING MATERIALS FOR THE PURPOSE OF CHILLED EVISCERATED POULTRY. Chem. and Indus. 49: 1492-1493.
(444) SIMPSON, K. L., NAGEL, C. W., NG, H., and OTHERS. 1959. INFLUENCE OF ACRONIZING ON ORGANISMS CAUSING SPOILAGE IN
COMMERCIALLY PROCESSED FRYERS. Food Technol. 13: 153-154. (445) SINCLAIR, N. A., and STOKES, J. L.
1963. ROLE OF OXYGEN IN THE HIGH CELL YIELDS OF PSYCHROPHILES AND MESOPHILES AT LOW TEMPERATURES. Jour. Bact. 85: 164-167.
(446) SiZER, I. W., and JOSEPHSON, E. S. 1942. KINETICS AS A FUNCTION OF TEMPERATURE OF LIPASE, TRYPSIN,
AND INVERTASE ACTIVITY FROM "70 TO 50° C. ("94 TO 122° F.) Food Res 7: 201-209.
(447) SMART, H. F. 1934. MICRO-ORGANISMS SURVIVING THE STORAGE PERIOD OF FROZEN-
PACK FRUITS AND VEGETABLES. Phytopathology 24: 1319-1331. (448)
1937. TYPES AND SURVIVAL OF SOME MICRO-ORGANISMS IN FROZEN-PACK PEAS, BEANS, AND SWEET CORN GROWN IN THE EAST. Food ReS. 2: 515-528.

104 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(449) SMART, H. F. 1939. MICROBIOLOGICAL STUDIES ON COMMERCIAL PACKS OF FROZEN
FRUITS AND VEGETABLES. Food Res. 4: 293-298. (450) and BRUNSTETTER, B. C.
1936. LIMA BEANS IN FROZEN PACK. I. BLANCHING TESTS. II. MICRO- BIOLOGICAL STUDIES. Canner 83 (10) : 14-16.
(451) and BRUNSTETTER, B. C. 1937. SPINACH AND KALE IN FROZEN PACK. I. SCALDING TESTS. II.
MICROBIOLOGICAL STUDIES. Food Res. 2: 151-163. (452) SMITH, A. U.
1961. BIOLOGICAL EFFECTS OF FREEZING AND SUPERCOOLING. 462 pp. Williams and Wilkins Co., Baltimore, Md.
(453) SNOW, D. 1949. THE GERMINATION OF MOULD SPORES AT CONTROLLED HUMIDITIES.
Ann. Appl. Biol. 36:1-13. (454) SouTHCOTT, B. A., and TARR, H. L. A.
1961. MAGNESIUM AS A FACTOR IN DETERMINING THE COMPARATIVE SENSITIVITY OF MARINE BACTERIA TO FOUR TETRACYCLINE ANTI- BIOTICS. Canad. Jour. Microbiol. 7: 284-286.
(455) SPENCER, J. V., EKLUND, M. W., SAUTER, E. A., and HARD, M. M. 1956. THE EFFECT OF DIFFERENT PACKAGING MATERIALS ON THE SHELF
LIFE OF ANTIBIOTIC TREATED CHICKEN FRYERS. (Abstract) Poultry Sei. 35: 1173.
(456) SAUTER, E. A., and STADELMAN, W. J. 1955. SHELF LIFE OF FROZEN POULTRY MEAT AFTER THAWING. (Ab-
stract) Poultry Sei. 34: 1222. (457) SAUTER, E. A., and STADELMAN, W. J.
1961. EFFECT OF FREEZING, THAW^ING AND STORING BROILERS ON SPOIL- AGE, FLAVOR AND BONE DARKENING. Poulty Sci. 40: 918-920.
(458) and STADELMAN, W. J. 1955. EFFECT OF CERTAIN HOLDING CONDITIONS ON SHELF LIFE OF
FRESH POULTRY MEAT. Food Technol. 9: 358-360. (459) ZiEGLER, F., and STADELMAN, W. J.
1954. RECENT STUDIES OF FACTORS AFFECTING THE SHELF LIFE OF CHICKEN MEAT. Wash. Agr. Expt. Sta. Cir. 254, 5 pp.
(460) SPLITTSTOESSER, D. F., WETTERGREEN, W. P., and PEDERSON, C. S. 1961. CONTROL OF MICRO-ORGANISMS DURING PREPARATION OF VEGETA-
BLES FOR FREEZING. I. GREEN BEANS. Food Technol. 15: 329-331.
(461) WETTERGREEN, W. P. and PEDERSON, C. S. 1961. CONTROL OF MICRO-ORGANISMS DURING PREPARATION OF VEGETA-
BLES FOR FREEZING. II. PEAS AND CORN. Food Technol. 15: 332-334.
(462) STANSBY, M. E., and GRIFFITHS, F. P. 1935. CARBON DIOXIDE IN HANDLING FRESH FISH. HADDOCK. Indus,
and Engin. Chem., Indus. Ed., 27: 1452-1458. (463) and LEMON, J. M.
1941. STUDIES ON THE HANDLING OF FRESH MACKEREL (SCOMBER SCOMBRUS). U.S. Fish and Wildlife Serv. Res. Rpt. 1, 46 pp.
(464) STEINER, G., and TARR, H. L. A. 1955. TRANSPORT AND STORAGE OF FISH IN REFRIGERATED SEA WATER.
II. BACTERIAL SPOILAGE OF BLUEBACK SALMON IN REFRIGERATED SEA WATER AND IN ICE WITH AND WITHOUT ADDED CHLORTETRA- CYCLINE. Canada Fisheries Res. Bd. Prog. Rpts. Pacific Coast Stas. 104, pp. 7-8.
(465) STEINKE, P. K. W., and FOSTER, E. M. 1951. MICROBIAL CHANGES IN REFRIGERATED LIVER SAUSAGE. Food
Res. 16: 245-251. (466) STERN, J., and DASSOW, J. A.
1958. CONSIDERATIONS ON THE USE OF REFRIGERATED BRINE FOR CHILL- ING AND STORING FRESH FISH. Com. Fisheries Rev. 20 (2) : 17-20.
(467) STEWART, G. F. 1953. PACKAGES ARE DIFFERENT. Poultry Processing and Mktg. 59
(7) : 13-15, 34.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 105
(468) STEWART, G. F., LOWE, B., and MORR, M. 1941. POST MORTEM CHANGES IN NEW YORK DRESSED POULTRY AT 35° F.
U.S. Egg & Poultry Mag. 47: 542-544, 571-572. (469) STEWART, M. M.
1934. THE KEEPING QUALITY OF THE THAWED HADDOCK. [Gt. Brit.], Dept. Sei. and Indus. Res., Food Invest. Bd. Rpt. 1933: 173- 178.
(470) 1935. THE KEEPING QUALITY OF HADDOCK FROM COLD STORAGE. Soc.
Chem. Indus. Jour. [London] 54: 92T-96T. (471) STILLE, B.
1941. ÜBER DIE HALTBARKEIT VON GEFRIERFLEISCH NACH DEM AUF- TAUEN. Ztschr. f. die Gesam. Kälte Indus. 48: 176-177.
(472) 1942. ÜBER DIE HALTBARKEIT VON GEFRIERFLEISCH NACH DEM AUF-
TAUEN. Ztschr. f. die Gesam. Kälte Indus. 49: 36-37. (473) STOKES, J. L.
1960. GROWTH OF MICRO-ORGANISMS AT LOW TEMPERATURES. Amer. Meat Inst. Found. (Cir. 61), Res. Conf. Proc. 12: 9-15.
(474) and OSBORNE, W. W. 1956. EFFECT OF THE EGG SHELL MEMBRANE ON BACTERIA. Food Res.
21: 264-269. (475) STUART, L. S., and MCNALLY, E. H.
1943. BACTERIOLOGICAL STUDIES ON THE EGG SHELL. U.S. Egg & Poultry Mag. 49: 28-31, 45-47.
(476) STUTTS, H. P. 1961. BIBLIOGRAPHY OF FROZEN FOODS. U.S. Dept. Agr. Misc. Pub.
868, 206 pp. (477) SULTZER, B. M.
1961. OXIDATIVE ACTIVITY OF PSYCHROPHILIC AND MESOPHILIC BAC- TERIA ON SATURATED FATTY ACIDS. Jour. Bact. 82: 492-497.
(478) SULZBACHER, W. L. 1952. EFFECT OF FREEZING AND THAWING ON THE GROWTH RATE OF
BACTERIA IN GROUND MEAT. Food Technol. 6: 341-343. (479) SzczEPULA, W., and SZCZEPULA, J.
1958. EFFECT OF PASTEURIZATION ON QUANTITY AND MICROFLORA OF CANNED FROZEN WHOLE EGGS. Przem. Spozywczy 12: 21-27. (Abstract in Chem. Abs. 52: 18947a. 1958.)
(480) TANNER, F. W. 1944. THE MICROBIOLOGY OF FOODS. Ed. 2, 1196 pp. Garrard Press,
Champaign, 111. (481) TARR, H. L. A.
1943. RISE AND FALL OF BACTERIAL POPULATIONS IN FISH MUSCLE. Canada Fisheries Res. Bd. Jour. 6: 119-128.
(482) 1953. THE BACTERIAL FLORA OF FRESH FISH AND ITS TECHNOLOGICAL
IMPLICATIONS. In Swedish Inst. for Food Preserv. Res. Göte- borg, Symposium on Cured and Frozen Fish Technology. Proc. Its SIK-Pub. 100, ch. 5, 7 pp.
MICROBIOLOGICAL DETERIORATION OF FISH POST MORTEM. ITS DE- TECTION AND CONTROL. Bact. Rev. 18: 1-15.
PRESERVATION OF FRESH FISH. Arch. f. Fishereiwiss. 8: 9-21.
NEW HORIZONS IN FOOD PRESERVATION. PART 2. Canad. Food Indus. 30 (6) : 25-27.
ANTIBIOTICS AS A PRESERVATIVE MEASURE. Canad. Inst. Food Technol. Nati. Conf., Toronto. 23 pp.
1960. ANTIBIOTICS IN FISH PRESERVATION. Canada Fisheries Res. Bd. Bui. 124, 24 pp. (Abstract in Internatl. Inst. Refrig. Bui. 41 (3): 868. 1961.)
^±00/
1954.
(484) 1957.
(485) 1959.
(486) 1959.
(487)

106 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(488) TARR, H. L. A. 1961. CHEMICAL CONTROL OF MICROBIOLOGICAL DETERIORATION. In
Borgstrom, G., ed.. Fish as Food 1: 639-680. Acad. Press, New York.
(489) THATCHER, F. S., and LOIT, A. 1961. COMPARATIVE MICROFLORA OF CHLORTETRACYCLINE-TREATED AND
NONTREATED POULTRY WITH SPECIAL REFERENCE TO PUBLIC HEALTH ASPECTS. Appl. Microbiol. 9: 39-45.
(490) THOMAS, S. B. 1958. PSYCHROPHILIC MICRO-ORGANISMS IN MILK AND DAIRY PRODUCTS
—PART I. (Abstract in Dairy Sei. Abs. v. 20: col. 366-369). (491)
(492)
1958. PSYCHROPHILIC MICRO-ORGANISMS IN MILK AND DAIRY PRODUCTS —PART II. (Abstract in Dairy Sei. Abs. v. 20, col. 447-468.)
1960. PSYCHROPHILIC BACTERIA IN REFRIGERATED RAW MILK. Dairy Engin. 77 (1): 5-9, 24.
(493) THORNLEY, M. J., INGRAM, M., and BARNES, E. M. 1960. THE EFFECTS OF ANTIBIOTICS AND IRRADIATION ON THE PSEUDO-
MONAS-ACHROMOBACTER FLORA OF CHILLED POULTRY. Jour. Appl. Bact. 23: 487-498.
(494) THORNTON, H. G., and MEIKLEJOHN, J. 1957. SOIL MICROBIOLOGY. Ann. Rev. Microbiol. 11: 123-148.
(495) TOBIN, L., ALFORD, J. A., and MCCLESKEY, C. S. 1941. THE BACTERIAL FLORA OF ICED, FRESH CRABMEAT. (Abstract)
Jour. Bact. 41: 96. (496) and MCCLESKEY, C. S.
1941. BACTERIOLOGICAL STUDIES OF FRESH CRABMEAT. Food Res. 6: 157-167.
(497) ToMiYAsu, Y., and ZENITANI, B. 1957. SPOILAGE OF FISH AND ITS PRESERVATION BY CHEMICAL AGENTS.
In Mrak, E. M., ed.. Advances in Food Res. 7: 41-82. (498) ToMKiNS, R. G.
1929. STUDIES OF THE GROWTH OF MOULDS. I. Roy. Soc. London, Proc, Ser. B, 105: 375-401.
(499)
(500)
1932. THE INHIBITION OF THE GROWTH OF MEAT-ATTACKING FUNGI BY CARBON DIOXIDE. Trans. and Commun., Soc. Chem. Indus. Jour. 51: 261T-264T. (Abstract in Biol. Abs. 7: 11344.1933.)
1951. THE MICROBIOLOGICAL PROBLEMS IN THE PRESERVATION OF FRESH FRUITS AND VEGETABLES. Jour. Sci. Food and Agr. 2: 381-386.
(501) TOMLINSON, N. 1957. ANTIBIOTICS IN FOOD PRESERVATION. Sanitarian 1 (9) : 15-22.
(502) TÖRÖK, G., and ALMáSI, E. 1954. ÉLELMISZEREK TARTÓSÍTÁSA FAGYPONTJUK ALATTI HÔMÉR-
SÉKLETEN, JEGKRISTÁLY KÉPZODÉS NELKÜL. [PRESERVATION OF FOOD-STUFFS AT TEMPERATURES BELOW THEIR FREEZING POINT WITHOUT THE FORMATION OF ICE CRYSTALS]. Élelmezési Ipar [Budapest] 8: 364-369.
(503) TORRES BOTELHO, A. 1958. PROFILAXIA DE ALTERA CAO VERMELHA DE PEIXES SALGADOS. Bol.
da Pesca 11 (58) : 11-23. (Abstract in World Fisheries Abs. 10 (5): 31-32. 1959).
(504) TRESSLER, D. K., and EVERS, C. F. 1957. MICROBIOLOGY OF FROZEN FOODS. In The Freezing Preservation
of Foods. Volume 1. Freezing of Fresh Foods, pp. 1067- 1089 (Ch. 29). Ed. 3, 1214 pp. Avi Pub. Co., Inc., Westport, Conn.
(505) TRUSSELL, P. C, FULTON, C. 0., and CAMERON, C. J. 1955. BACTERIAL SPOILAGE OF SHELL EGGS. II. INCIDENCE OP SPOIL-
AGE IN EGGS FROM NINETY-FOUR FARMS. Food Technol. 9: 130- 134.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 107
(506) TRUSSELL, P. C, TRIGGS, R. E., and GREER, B. A. 1955. BACTERIAL SPOILAGE OF SHELL EGGS. III. FARM PRACTICES PRO-
MOTING SPOILAGE. Food Technol. 9: 134-137. (507) UNITED STATES FISH AND WILDLIFE SERVICE.
1960. REFRIGERATED SEA WATER LENGTHENS STORAGE LIFE OF WHITING. Com. Fisheries Rev. 22 (3) : 15.
(508) UNITED STATES FOOD AND DRUG ADMINISTRATION. 1955-59. TOLERANCES AND EXEMPTIONS FROM TOLERANCES FOR PESTICIDE
CHEMICALS IN OR ON RAW AGRICULTURAL COMMODITIES. TOL- ERANCE FOR RESIDUES OF CHLORTETRACYCLINE. [U.S.] NATI. Arch., Fed. Register 20 (232) : 8776 (1955) ; 24 (77) : 3054 (1959).
(509) UNITED STATES PUBLIC HEALTH SERVICE. 1963. BOTULISM. Morbidity and Mortality Weekly Rpt. 12: 329-330,
337-339, 345, 358, 394, 400. (510) UPADHYAY, J., and STOKES, J. L.
1962. ANAEROBIC GROWTH OF PSYCHROPHILIC BACTERIA. Jour. Bact. 83: 270-275.
(511) and STOKES, J. L. 1962. A TEMPERATURE SENSITIVE FORMIC HYDROGENYLASE IN A PSY-
CHROPHILIC BACTERIUM. (Abstract) Soc. Amer. Bact., Bact. Proc. 62: 124.
(512) USPENSKY, A. A. 1944. TRACHEAL INJECTION METHOD FOR SHORT-TIME PRESERVATION OF
DRESSED POULTRY U.S. Egg & Poultry Mag. 50: 342-345, 373- 374, 412-414, 464-466.
(513) VALLEY, G., and RETTGER, L. F. 1927. THE INFLUENCE OF CARBON DIOXIDE ON BACTERIA. Jour. Bact.
14: 101-137. (514) VANDERLECK, J.
1918. BACTERIA OF FROZEN SOILS IN QUEBEC. II. Roy. Soc. Canada, Trans, (ser. 3), Sect. IV, 12: 1-21.
(515) VAN DER ZANT, W. C, and MOORE, A. V. 1955. THE INFLUENCE OF CERTAIN FACTORS ON THE BACTERIAL COUNTS
AND PROTEOLYTIC ACTIVITIES OF SEVERAL PSYCHROPHILIC OR- GANISMS. Jour. Dairy Sei. 38: 743-750.
(516) VAN ESELTINE, W. P., NELLIS, L. F., LEE, F. A., and HUCKER, G. J. 1948. EFFECT OF RATE OF FREEZING ON BACTERIAL CONTENT OF FROZEN
VEGETABLES. Food Res. 13: 271-280. (517) VAUGHN, R. H., NAGEL, C. W., SAWYER, F. M., and STEWART, G. F.
1957. ANTIBIOTICS IN POULTRY MEAT PRESERVATION. A COMPARISON OF THE TETRACYCLINES. Food Technol. 11: 426-429.
(518) NG, H., STEWART, G. F., and OTHERS. 1959. ANTIBIOTICS IN POULTRY MEAT PRESERVATION: IN VITRO DEVEL-
OPMENT OF BACTERIAL RESISTANCE TO CHLORTETRACYCLINE. (Abstract) Food Technol. 13(4): 23.
(519) NG, H., STEWART, G. F., and OTHERS. 1960. ANTIBIOTICS IN POULTRY MEAT PRESERVATION. DEVELOPMENT IN
VITRO OF BACTERIAL RESISTANCE TO CHLORTETRACYCLINE. Appl. Microbiol. 8: 27-30.
(520) and STADTMAN, T. C. 1946. SANITATION IN THE PROCESSING PLANT AND ITS RELATION TO THE
MICROBIAL QUALITY OF THE FINISHED PRODUCTS. Food Freezing 1: 334-336, 364.
(521) STADTMAN, T. C., and KUENEMAN, R. W. 1946. CONTROL OF MICRO-ORGANISMS IN FROZEN FOOD PLANTS. Quick
Frozen Foods 9(2): 76-78, 114. (522) VENKATARAMAN, R.
1949. EFFECT OF TEMPERATURE ON THE GROWTH OF FISH SPOILAGE BACTERIA. Indian Jour. Med. Res. 37 (1) : 91-99.
(523) WALDEN, C. C., ALLEN I. V. F., and TRUSSELL, P. C. 1956. THE ROLE OF THE EGG SHELL AND SHELL MEMBRANES IN RE-
STRAINING THE ENTRY OF MICRO-ORGANISMS. Poultry Sci. 35: 1190-1196.

108 TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(524) WALKER, E. A., and TAYLOR, J. H. 1960. THE EFFECT OF CHLORTETRACYCLINE ON EVISCERATED FROZEN
CHICKENS COMMERCIALLY PROCESSED IN GREAT BRITAIN. Jour. Appl. Bact. 23: 155-157.
(525) TAYLOR, J. H., and DAVIS, J. G. 1960. THE EFFECT OF CHLORTETRACYCLINE ON EVISCERATED CHICKENS
COMMERCIALLY PROCESSED IN GREAT BRITAIN. Jour. Appl. Bact. 23: 145-154.
(526) WALKER, H. W., and AYRES, J. C. 1959. CHARACTERISTICS OF YEASTS ISOLATED FROM PROCESSED POULTRY
AND THE INFLUENCE OF TETRACYCLINES ON THEIR GROWTH. Appl. Microbiol. 7: 251-255.
(527) WALL, S. 1912. THE PRESERVATION OF FRESH FISH KEPT ON^ICE OR AFTER FREEZ-
ING. Handlingar till Landtbruksveekan âr 1912, 264. centbl. Bakt. [etc.], Abt., Referate 56: 701-702. (Abstract in Chem. Abs. 7: 2633^ 1913.)
(528) WEIL, B. H., and STERNE, F. 1946. LITERATURE SEARCH ON THE PRESERVATION OF FOOD BY FREEZING.
Ga. state Engin. Expt. Sta. Spec. Rpt. 23, 409 pp. (529) WEINBERG, E. D.
1957. THE MUTUAL EFFECTS OF ANTIMICROBIAL COMPOUNDS AND ME- TALLIC CATIONS. Bact. Rev. 21: 46-68.
(530) WEINZIRL, J., and NEWTON, E. B. 1914. BACTERIOLOGICAL ANALYSES OF HAMBURGER STEAK WITH REF-
ERENCE TO SANITARY STANDARDS. Amer. Jour. Pub. Health 4: 413.
(531) WEISER, H. H., KUNKLE, L. E., and DEATHERAGE, F. E. 1954. THE USE OF ANTIBIOTICS IN MEAT PROCESSING. Appl. Microbiol.
2: 88-94. (532) WELLS, F. E., HARTSELL, S. E., and STADELMAN, W. J.
1963. GROWTH OF PSYCHROPHILES. I. LIPID CHANGES IN RELATION TO GROWTH-TEMPERATURE REDUCTIONS. Jour. Food Sci. 28: 140-144.
(533) SPENCER, J. V., and STADELMAN, W. J. 1958. EFFECT OF PACKAGING MATERIALS AND TECHNIQUES ON SHELF
LIFE OF FRESH POULTRY MEAT. Food Technol. 12: 425-427. (534) STADELMAN, W. J., and HARTSELL, S. E.
1963. GROWTH OF PSYCHROPHILES. II. GROWTH OF POULTRY MEAT SPOILAGE BACTERIA AND SOME EFFECTS OF CHLORTETRACYCLINE. Jour. Food Sei. 28: 254-258.
(535) WHITE, A., and WHITE, H. R. 1962. SOME ASPECTS OF THE MICROBIOLOGY OF FROZEN PEAS. Jour.
Appl. Bact. 25: 62-71. (536) WHITEHILL, A. R.
1957. INHIBITION OF YEASTS BY FOOD SPOILAGE ORGANISMS. (Ab- stract) See. Amer. Bact., Bact. Proc. 57: 20.
(537) WILLIAMS, I. L., and YUNG, F. D. 1955. THE EFFECT OF SWEATING ON KEEPING QUALITY OF SHELL EGGS.
Food Res. 20: 221-225. (538) WiNSLOW, C.-E. A., and WALKER, H. H.
1939. THE EARLIER PHASES OF THE BACTERIAL CULTURE CYCLE. Bact. Rev. 3: 147-186.
(539) WINTER, A. R., BURKART, B., CLEMENTS, P., and MACDONALD, L. 1955. CLEANING EGGS WITH DETERGENTS AND DETERGENT-SANITIZERS.
Ohio Agr. Expt. Sta. Res. Bui. 762, 42 pp. (540) GRECO, P. A., and STEWART, G. F.
1946. PASTEURIZATION OF LIQUID-EGG PRODUCTS, I. BACTERIA RE- DUCTION IN LIQUID WHOLE EGG AND IMPROVEMENT IN KEEPING QUALITY. Food Res. 11: 229-245.

PSYCHROPHILIC MICRO-ORGANISMS IN FOODS 109
(541) WITTER, L. D. 1961. PSYCHROPHILIC BACTERIA—^A REVIEW. Jour. Dairy Sei. 44:
983-1015 (542) WoDZiNSKi, R. J., and FRAZIER, W. C.
1960. MOISTURE REQUIREMENTS OF BACTERIA. I. INFLUENCE OF TEM- PERATURE AND PH ON REQUIREMENTS OF PSEUDOMONAS FLUO- RESCENS. Jour. Bact. 79: 572-578.
(543) and FRAZIER, W. C. 1961. MOISTURE REQUIREMENTS OF BACTERIA. II. INFLUENCE OF TEM-
PERATURE, PH, AND MALATE CONCENTRATION ON REQUIREMENTS OF AEROBACTER AER0GENES. Jour. Bact.. 81: 353-358.
(544) and FRAZIER, W. C. 1961. MOISTURE REQUIREMENTS OF BACTERIA. III. INFLUENCE OF TEM-
PERATURE, PH, AND MALATE AND THIAMINE CONCENTRATION ON REQUIREMENTS OF LACTOBACILLUS VIRIDESCENS. Jour. Bact. 81 : 359-365.
(545) and FRAZIER, W. C. 1961. MOISTURE REQUIREMENTS OF BACTERIA. IV. INFLUENCE OF TEM-
PERATURE AND INCREASED PARTIAL PRESSURE OF CARBON DIOXIDE ON REQUIREMENTS OF THREE SPECIES OF BACTERIA. Jour. Bact. 81: 401-408.
(546) WOLFORD, E. R. 1958. EFFECT OF VARIED STORAGE TEMPERATURES ON MICRO-ORGANISMS
IN FROZEN CONCENTRATED ORANGE JUICE. Appl. Microbiol. 6: 139-141.
(547) WOLK, J., MCNALLY, E. H., and SPICKNALL, N. H. 1950. THE EFFECT OF TEMPERATURE ON THE BACTERIAL INFECTION OF
SHELL EGGS. Food Technol. 4: 316-318. (548) WOOD, A. J., SIGURDSSON, G. J., and DYER, W. J.
1942. THE SURFACE CONCEPT IN MEASUREMENT OF FISH SPOILAGE. Canada Fisheries Res. Bd. Jour. 6: 53-62.
(549) WOOD, E. J. F. 1940. STUDIES ON THE MARKETING OF FRESH FISH IN EASTERN AUS-
TRALIA. PART 2. THE BACTERIOLOGY OF SPOILAGE OF MARINE FISH. Austral. Council Sei. and Indus. Res. Pam. 100, 92 pp.
(550) WRENSHALL, C. L. 1957. ADVANCES IN FOOD TECHNOLOGY MADE POSSIBLE THROUGH THE
USE OF ANTIBIOTICS. Antibiotics Ann. 1956-57: 809-815. (551) WYSS, 0.
1948. MICROBIAL INHIBITION BY FOOD PRESERVATIVES. In Mrak, E. M., ed., Advances in Food Research. 1: 373-393.
(552) YACOWITZ, H., PANSY, F., WIND, S., and OTHERS. 1957. USE OF NYSTATIN (MYCOSTATIN) TO RETARD YEAST GR0V7TH ON
CHLORTETRACYCLINE-TREATED CHICKEN MEAT. Poultry Sci. 36: 843-849.
(553) YAMAMURA, Y. 1937. THE RELATION BETWEEN TEMPERATURE AND THE RATE OF DE-
COMPOSITION OF FISH MUSCLE. Jap. Soc. Sci. Fisheries Bui. 6: 141-144.
(554) YOUNG, E. G. 1929. ENDOCELLULAR ENZYMES OF BACILLUS COLI COMMUNIS. BIO-
chem. Jour. 23: 831-839. (555) ZAGAEVSKY, J. S., and LUTIKOVA, P. 0.
1944. SANITARY MEASURES IN THE EGG BREAKING PLANT. U.S. Egg & Poulty Mag. 50: 17-20, 43-46.
(556) and LUTIKOVA, P. 0. 1944. BACTERIAL CONTAMINATION IN EGGS. U.S. Egg & Poultry Mag.
50: 75-77, 88-90. (557) ZiEGLER, F., and STADELMAN, W. J.
1955. THE EFFECT OF DIFFERENT SCALD WATER TEMPERATURES ON THE SHELF LIFE OF FRESH, NONFROZEN FRYERS. Poultry Sci. 34: 237-238.

lio TECHNICAL BULLETIN 1320, U.S. DEPT. OF AGRICULTURE
(558) ZIEGLER, P. T. 1943. THE MEAT WE EAT. 876 pp. Interstate Printers and Pub!.,
Danville, 111. (559) Zo BELL, C. E.
1946. MARINE MICROBIOLOGY, A MONOGRAPH ON HYDRO BACTERIOLOGY. 240 pp. Chronica Botánica Co., Waltham, Mass.
(560) 1962. IMPORTANCE OF MICRO-ORGANISMS IN THE SEA. Campbell Soup
Co., Camden, N.J., Low Temp. Microbiol. Symposium 1961: 107-132.
(561) and CONN, J. E. 1940. STUDIES ON THE THERMAL SENSITIVITY OF MARINE BACTERIA.
Jour. Bact. 40: 223-238.
^ U. S. GOVERNMENT PRINTING OFFICE: 1965—743-027





























