Embed Size (px)
Citation preview

VIROLOGY 64, 330-346 (1973)
Properties of Mouse Leukemia Viruses
IV. Hemagglutination Assay and Characterization of
Hemagglutinating Surface Components
RICHARD WITTER,’ HERMANN FRANK, VOLKER MOENNIG, GERHARD HUNSMANN, JENS LANGE, AND WERNER SCH;niFER
Max-Plan&-Institut fiir virusforschung, Tiibingen, Germany
Accepted April 2, 1973
A hemagglutination (HA) assay for mouse leukemia virus (MuLV) concentrates is described. The assay gives linear dose-response data between concentrations of approximately 10” and 8 X 1Ol2 particles per milliliter. One HA unit was equivalent to approximately 5 X lo7 virus particles in those preparations where surface projec- tions (knobs) were readily detectable on the virions. When knobs were removed during purification or by bromelain treatment, the HA activity was reduced or eliminated. Bromelain-treated virus also lacked glycoprotein component(s), did not absorb neutralizing MuLV antibody, and was noninfectious. Thus, the HA substructure is clearly a surface component of the virion and has an apparent association with the surface knobs, glycoprotein(s), and type-specific antigens. Treatment with neur- aminidase and phoapholipase C, aa required for demonstration of HA activity, damaged the envelope of the virion but did not release hemagglutinating subunits or surface knobs. Active hemagglutinin recovered from enzyme-treated virus by degra- dation with Tween 80 and ether and density gradient centrifugation was stable under various physical treatments but was heat sensitive. Active hemagglutinin could also be recovered from non-enzyme-treated virus by similar procedures. Concentrated preparations of avian myeloblastosis virus and feline leukemia virus agglutinated chicken and sheep erythrocytes, respectively, but in both cases the specific HA activities were low and the reactions could not be inhibited by specific antisera.
INTRODUCTION
The property of hemagglutination (HA) was first associated with RNA tumor viruses by Gross (1959) and Rapp et al. (1962). Crude tissue extracts from mice infected with murine leukemia virus (MuLV) were found by these workers to contain a hemagglutinat- ing activity that was enhanced by or even dependent upon heat treatment (45-55”). The specificity of the HA was not estab- lished, and in one case (Gross, 1959), it was clear that the HA activity was not directly virion-associated.
1 Permanent address : Regional Poultry Re- search Laboratory, Agricultural Research Service, United States Department of Agriculture, East Lansing, Michigan 48823.
Schafer and Szgnt6 (1969) described a virus-specific HA activity associated with concentrates of purified MuLV which had first been treated with neuraminidase and phospholipase C. This HA activity differed from that previously reported because it was dependent on enzyme pretreatment, was markedly heat sensitive and, furthermore, was specifically associated with the MuLV virus particle (Schgfer and Sz&6, 1969). The hemagglutinating subunit subsequently was isolated from enzyme-treated, Tween 80, and ether (Tween-ether) degraded MuLV by Sephadex chromatography and density gradient centrifugation and possessed both type-specific and group-specific (gs) antigens (Schgfer et al., 1972b). SzBnt6 (1971) also
330 Copyright @ 1973 by Academic Press, Inc. All rights of reproduction in any form reserved.

HEMAGGLUTINATION ASSAY 331
reported HA activity with enzyme-treated avian myeloblastosis virus.
Widespread application of these HA pro- cedures, however, was hindered by technical difficulties, particularly autohemagglutina- tion by enzymes alone and nonspecific serum inhibitors which interfered with hemaggluti- nation-inhibition (HI) tests. Nonetheless, the potential value of an HA system seemed to justify further efforts to resolve these difficulties and to determine whether such t,echniques would have value in research with mammalian and avian RNA tumor viruses.
In this report we describe the development of a simple HA assay for MuLV concentrates which can be adequately controlled for spec- ificity. Data on the physicochemical char- acteristics and structure of MuLV hemag- glutinin as well as attempts to identify similar hemagglutinins in avian myeloblas- tosis virus and feline leukemia virus are also presented. Serologic studies including de- scription of HI and indirect HA tests for MuLV are discussed in an accompanying communication (Witter et al., 1973).
MATERIALS AND METHODS
Viruses. Friend leukemia virus (FLV) pro- duced in STU-mouse embryo cells (Schafer and Seifert, 1968), Rauscher leukemia virus (RLV) produced in JLSV, cells (Schafer et al., 1970) and Gross leukemia virus (GLV) produced in permanent rat thymus cell cul- tures originally obtained from Dr. Ioachim, New York (Ioachim, 1969; Schafer et al., 1970) or in mouse 3T3 fibroblasts were con- centrated by 2 cycles of ultracentrifugation at. a performance index (pi) of 10 (Giebler, 1958). The viruses were then banded in sucrose gradients and the material with a density of about 1.16 g/ml, was dialyzed against phosphate-buffered saline (PBS), pH 7.2 and further concentrated, if neces- sary, by centrifugation at pi 10. Alterna- tively, some preparations were used without density gradient centrifugation and some were concentrated by precipitation with ammonium sulfate (Horzinek et al., 1971) or polyethylene glycol (Horzinek, 1969).
Avian myeloblastosis virus (AMV) was recovered from the plasma of AMV- inoculated, leukemic chickens or from media of infected chicken embryo fibroblast cul-
tures. These materials were clarified by low speed centrifugation (3000 g, 15 min) and the virus-containing supernatant was centri- fuged at pi 10. The pellets were further purified by centrifugation on discontinuous and continuous sucrose gradients (Bauer, 1970). The virus band (1.16 g/ml) was dialyzed against PBS, pH 7.2, and further concentrated by centrifugation at pi 10. Plasma virus concentrates were evaluated by the ATPase test (Mommaerts et al., 1954).
Feline leukemia virus (FeLV), Theilen strain, was supplied by Pfizer Research Laboratories, Maywood, New Jersey, as a zonal centrifuged concentrate containing 1.15 X 1012 particles per ml and was further concentrated by pressure dialysis.
Except, where noted, virus concentrates were stored at -70” prior to use.
HA test. The procedures were similar to those of Schafer and SzBnt6 (1969). MuLV and FeLV concentrates were dialyzed against pH 6.4 buffer (prepared from equal volumes of solution A: NaCl, 8.0 g/l; KCl, 0.4 g/l; CaCl2 , 0.14 g/l and MgS04.7 HzO, 0.2 g/l and solution B: 0.15 M, pH 6.4 phosphate buffer). One volume of virus was mixed with 2 volumes of neuraminidase from Vibrio chobrae, 500 units/ml (Behringwerke, Mar- burg) and one volume of phospholipase C from Clostridium welchii, 4 units/mg (Sigma Chemical, St. Louis, Missouri). The phos- pholipase C was prepared as a stock solution in PBS, pH 7.2 at 1 mg/ml and diluted 1: 5 or occasionally 1: 10 prior to use with pH 6.4 buffer. Following incubation of the mixture in tubes at 37” for 30 min with occasional shaking, 0.1 ml was placed in Microtiter “V” plates (Cooke Engineering Co. Alexa,ndria, Virginia) and diluted in pH 6.4 buffer, 0.05 ml per cup. To each cup was added 0.05 ml of a 0.5 % suspension of washed sheep (wether) erythrocytes which were prepared from a 5% stock erythrocyte suspension in PBS, pH 7.2 by dilution with pH 6.4 buffer. The plates were shaken, held at 4” for 2 hr and scored for HA by the pattern method. Titers were recorded as the reciprocal of the highest antigen dilution (in 0.05 ml) causing detectable HA. Similar procedures were used for AMV except that chicken (rooster) erythrocyt#es were used (Szanti, 1971). Con- t’rols included pH 6.4 buffer plus mm-

332 WITTER ET AL.
aminidase, pH 6.4 buffer plus both enzymes and heat-inactivated virus (56’ for 15 min) plus both enzymes.
Isolation of hemagglutinin. Hemagglutinat- ing viral substructures were isolated from MuLV following treatment with neuramini- dase and phospholipase C and degradation with Tween-ether by Sephadex G-150 chromatography (sometimes omitted) and centrifugation in potassium citrate density gradients as described by Schafer et al. (197213). Individual fractions from density gradients were dialyzed against pH 7.2 PBS prior to evaluation for HA. Fractions with peak HA activity were pooled and stored at -20”.
Electron microscopy. The enumeration of particles in purified virus suspensions was conducted by a modification (Frank and Pietrzik, in preparation) of the agar filtration technique described by Kellenberger and Arber (1957). A given volume (0.05 ml) of virus suspension without latex spheres was carefully distributed over the collodion film overlaying the agar matrix in an 8.4 cm diameter petri dish. After filtration, agar blocks were taken from 4 different locations. The collodion films were floated from the agar onto distilled water and 2 grids were prepared from each film. After shadowing with platinum, 3 random micrographs were made from each grid at a magnification of 10,000 X. The particles on each of the 24 micrographs were enumerated and the par- ticle number per ml was calculated by the following formula :
area of petri dish area photographed
X mean No. particles/micrograph X 20
Negative staining was done by first placing a drop of the viral suspension on the grid for adsorption and then applying 1% uranyl acetate (UA) pH 4.4. For the demonstration of knobs, UA was removed within 10 set by drawing off with filter paper. Longer contact between virus particles and UA-solution re- sulted in deeper penetration of the stain and thus in difficulty in visualizing knobs.
Freeze-drying was performed as described by Nermut et al. (1972).
Polyacrylamide gel electrophoresis (PAGE). Virus preparations were treated with dithio-
threitol (DTT) and sodium dodecyl sulfate (SDS) and electrophoresed in gels containing 10% acrylamide plus SDS and DTT as described by Bolognesi and Bauer (1970). The gels were stained for protein with Coomassie blue and for polysaccharides by the periodic acid8chiff (PAS) reaction (Bolognesi and Bauer, 1970).
Complement jixation (CF) test. Dilutions of appropriate antigens were reacted with 2 units of complement in the presence of 4 units of serum. R-gs serum, obtained from a rabbit immunized with purified RLV gs pro- tein and which reacts principally with MuLV gs antigens IV and V (Schafer et al., 1972a), and rabbit anti-AMV gs serum (Bauer, 1970) were used at dilutions of 1: 50 and 1: 32, respectively, for the respective de- tection of MuLV and AMV gs antigens. Titers were the reciprocal of minimal antigen dilution (in 0.025 ml) that fixed 50% of the complement. When not otherwise degraded, virus suspensions were frozen and thawed 3 times prior to testing.
Absorption of neutralizing antibody. Ab- sorption tests were conducted as previously described (Schafer et al., 1972b) except that serum FTE/H, a potent neutralizing serum (titer 1:400) prepared in rabbits against Tween-ether degraded FLV, was used, and that the residual infectivity of virus-serum mixtures was assayed by the XC cell plaque assay for MuLV (Rowe et al., 1970). The FLV was used in concentrations that de- livered about 100 plaques per tissue culture plate. Absorbing antigens were routinely re- acted with FLV in the absence of serum to determine the autoinhibitory activity. How- ever, such inhibitory activity was never sufficient to account for observed reductions of MuLV infectivity by treated sera.
Ouchterlony immunodiflusion tests. Tests employing the complex anti-Rauscher virus RSDS/B serum were conducted as previously described (Schafer et al., 1972a).
Protein. Protein determinations were per- formed by the method of Lowry et al. (1951).
ICESULTS
Speci$city of MuLV Hemagglutination
Demonstration of HA with int,act MuLV is dependent on pretreatment of the virus suspension with neuraminidase and phos-
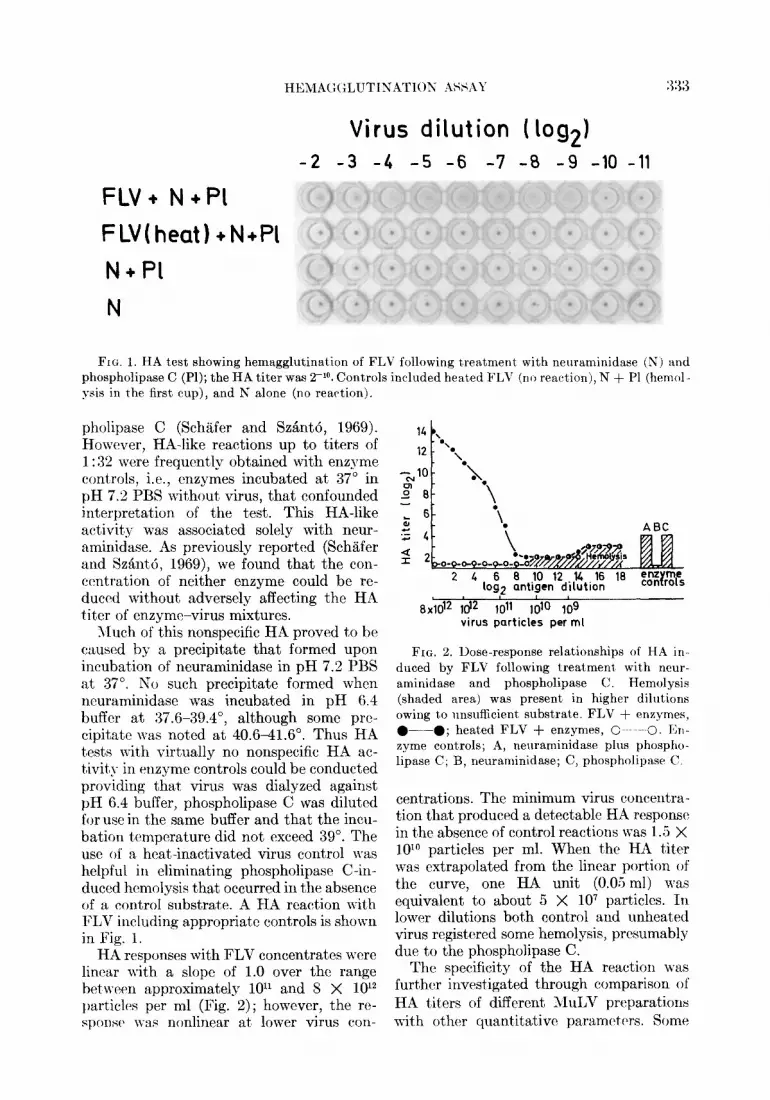
HEMA(:(;LUTINATION ASSAY
Virus dilution (log$ -2 -3 -4 -5 -6 -7 -0 -9 -10 -11
:n3
FLV+ N+PI
FLV( heat) +N+R N+Pl
N
Fro. 1. HA test showing hemagglutination of FLV following treatment with neuraminidase (N) and phospholipase C (Pl); the HA titer was 2- lo. Controls included heated FLV ( no reaction), N + Pl (hemol- ysis in the first cup), and N alone (no reaction).
pholipase C (SchSifer and SzBnt6, 1969). However, HA-like reactions up to titers of 1: 32 were frequently obtained with enzyme cont,rols, i.e., enzymes incubated at 37” in pH 7.2 PBS without’ virus, that confounded interpretation of the test. This HA-like activit,y was associated solely with neur- aminidase. As previously reported (Schgfer and Sz&ntb, 1969), we found that Dhe con- centration of neither enzyme could be re- duced without adversely affecting the HA ti t#er of enzyme-virus mixtures.
Much of this nonspecific HA proved to he caused by a precipitate that formed upon incubat,ion of neuraminidase in pH 7.2 PBS at 37”. No such precipitate formed when ncuraminidase was incubated in pH 6.4 buffer at 37.6-39.4”, although some prc- cipitatc was not’ed at 40.641.6”. Thus HA t,ests with virtually no nonspecific HA ac- tivit#\- in enzyme controls could be conducted providing that virus was dialyzed against pH 6.4 buffer, phospholipase C was diluted for use in the same buffer and that the incu- bation temperature did not exceed 39”. The use of a heat-inactivated virus control was helpful in eliminating phospholipase C-in- duced hcmolysis that occurred in the absence of a control substrate. A HA reaction with FLV including appropriate controls is shown in Fig. 1.
HA responses with FLV concentrates were linear with a slope of 1.0 over the range between approximately 10” and S X 1012 particlrs per ml (Fig. 2) ; however, the re- sponsc~ was nonlinear at’ lower virus con-
.- : 4 P 2
2 4 6 log, 8 ,lO 12 l4 16 18 antlgcn dllutlon
8x1012 d2 i-11 10’0 109 virus particles per ml
FIG. 2. Dose-response relationships of HA in- duced by FLV following treatment with neur- aminidase and phospholipase C. Hemolysis (shaded area) was present in higher dilutions owing t,o unsufficient substrate. FLV + enzymes, 0-0; heated FLV + enzymes, 0 -~ 0. I+:n- zyme controls; A, neuraminidase plus phospho- lipase C; B, neuraminidase; C, phospholipase C.
cent,rations. The minimum virus concentra- tion that produced a detectable HA response in t’he absence of control reactions was 1.5 X lOlo particles per ml. When the HA titer was extrapolated from the linear portion of the curve, one HA unit (0.05 ml) was equivalent to about 5 X lo7 particles. In lower dilutions both control and unheated virus registclred some hemolysis, presumably due to the phospholipase C.
The specificity of the HA reaction was further investigated through comparison of HA titers of different ?IzuLV preparations with other quantitative paramclttrs. Somt:
334 WITTER ET AL.
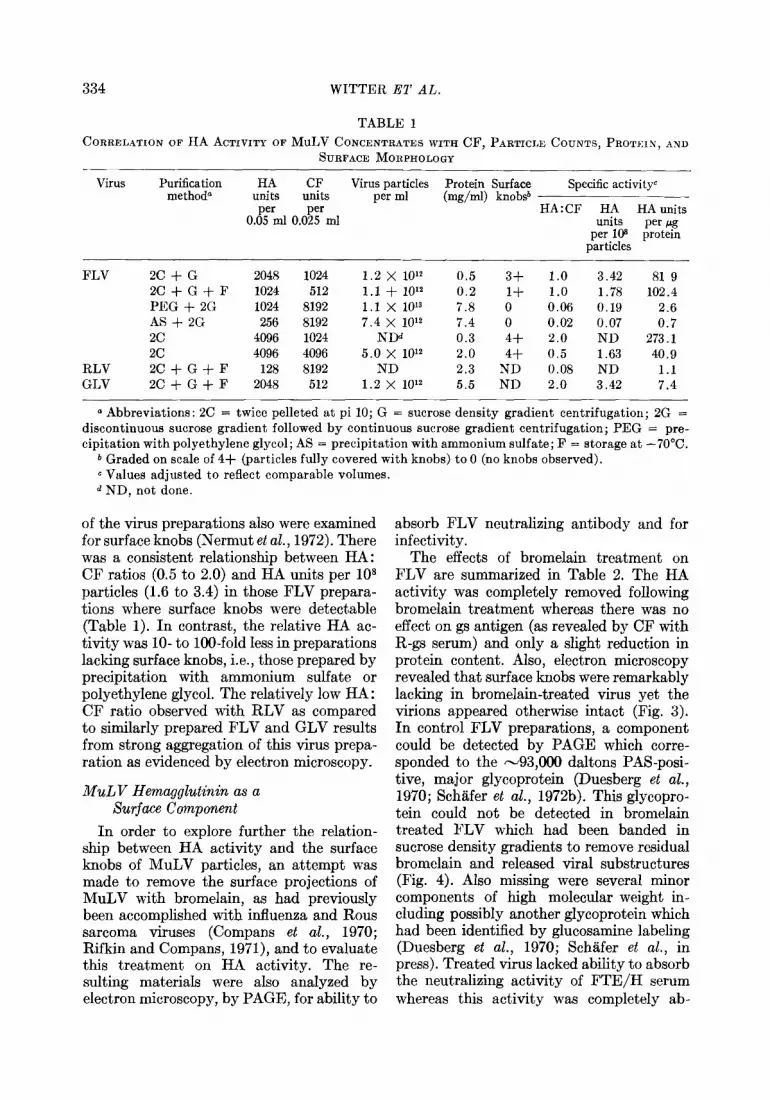
TABLE 1 CORRELATION OF HA ACTIVITY OF MuLV CONCENTRATES WITH CF, PARTICLE COUNTS, PROTEIN, AND
SURFACE MORPHOLOGY
Virus Purification HA CF Virus particles Protein Surface methoda units units per ml (mgjml) knob@
Specific activity”
per per HA:CF HA HA units 0.05 ml 0.025 ml units per erg
per 108 particles
protein
FLV 2C + G 2048 1024 1.2 x 10’2 0.5 3f 1.0 3.42 81 9 2C+G+F 1024 512 1.1 + 10’2 0.2 1+ 1.0 1.78 102.4 PEG + 2G 1024 8192 1.1 x 10’3 7.8 0 0.06 0.19 2.6 AS + 2G 256 8192 7.4 x 10’2 7.4 0 0.02 0.07 0.7 2c 4096 1024 NDd 0.3 4+ 2.0 ND 273.1 2c 4096 4096 5.0 x 10’2 2.0 4+ 0.5 1.63 40.9
RLV 2C+G+F 128 8192 ND 2.3 ND 0.08 ND 1.1 GLV 2C+G+F 2048 512 1.2 x 10’2 5.5 ND 2.0 3.42 7.4
0 Abbreviations: 2C = twice pelleted at pi 10; G = sucrose density gradient centrifugation; 2G = discontinuous sucrose gradient followed by continuous sucrose gradient centrifugation; PEG = pre- cipitation with polyethylene glycol; AS = precipitation with ammonium sulfate; F = storage at -70°C.
b Graded on scale of 4f (particles fully covered with knobs) to 0 (no knobs observed). c Values adjusted to reflect comparable volumes. d ND, not done.
of the virus preparations also were examined for surface knobs (Nermut et al., 1972). There was a consistent relationship between HA: CF ratios (0.5 to 2.0) and HA units per lo* particles (1.6 to 3.4) in those FLV prepara- tions where surface knobs were detectable (Table 1). In contrast, the relative HA ac- tivity was lo- to lOO-fold less in preparations lacking surface knobs, i.e., those prepared by precipitation with ammonium sulfate or polyethylene glycol. The relatively low HA : CF ratio observed with RLV as compared to similarly prepared FLV and GLV results from strong aggregation of this virus prepa- ration as evidenced by electron microscopy.
MuLV Hemagglutinin as a Surface Component
In order to explore further the relation- ship between HA activity and the surface knobs of MuLV particles, an attempt was made to remove the surface projections of MuLV with bromelain, as had previously been accomplished with influenza and Rous sarcoma viruses (Compans et al., 1970; Rifkin and Compans, 1971), and to evaluate this treatment on HA activity. The re- sulting mat,erials were also analyzed by electron microscopy, by PAGE, for ability to
absorb FLV neutralizing antibody and for infectivity.
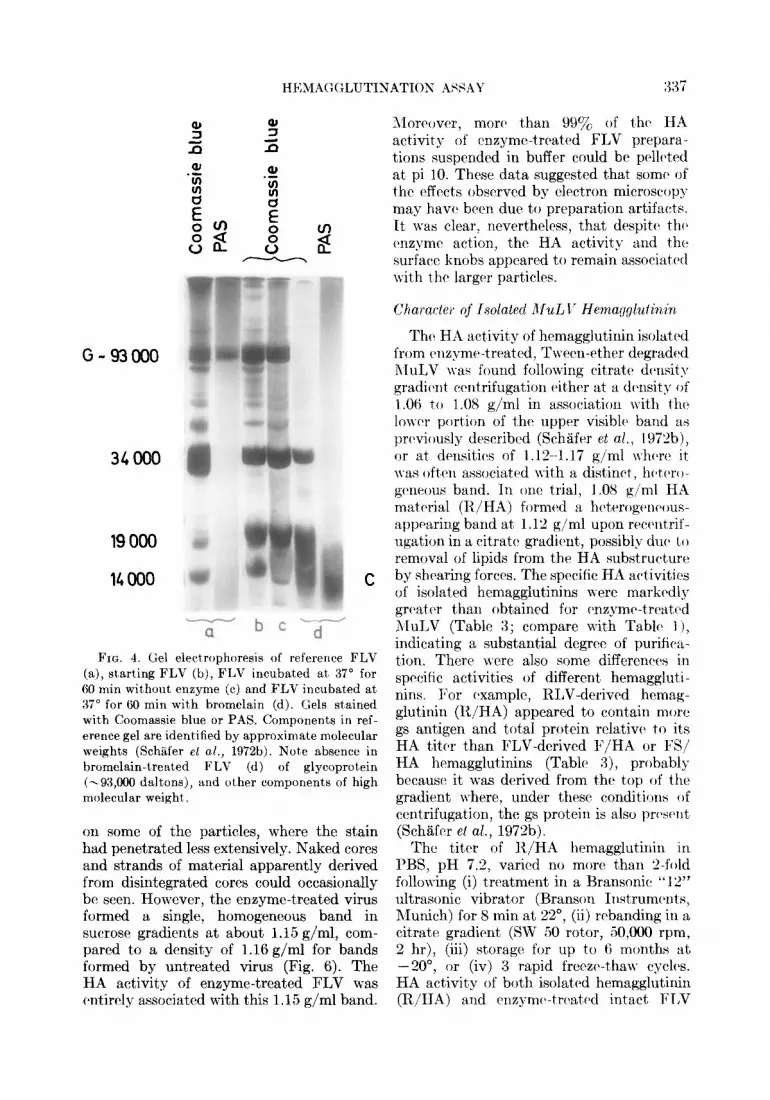
The effects of bromelain treatment on FLV are summarized in Table 2. The HA activity was completely removed following bromelain treatment whereas there was no effect on gs antigen (as revealed by CF with R-gs serum) and only a slight reduction in protein content. Also, electron microscopy revealed that surface knobs were remarkably lacking in bromelain-treated virus yet the virions appeared otherwise intact (Fig. 3). In control FLV preparations, a component could be detected by PAGE which corre- sponded to the -93,000 daltons PAS-posi- tive, major glycoprotein (Duesberg et al., 1970; SchB;fer et al., 1972b). This glycopro- tein could not be detected in bromelain treated FLV which had been banded in sucrose density gradients to remove residual bromelain and released viral substructures (Fig. 4). Also missing were several minor components of high molecular weight in- cluding possibly another glycoprotein which had been identified by glucosamine labeling (Duesberg et al., 1970; Schiifer et al., in press). Treated virus lacked ability to absorb the neutralizing activity of FTE/H serum whereas this activity was completely ab-

HEMAGGLUTINATION ASSAY 33.5
TABLE 2
EFFIXT OF BROMELAIN ON HA ACTIVITY, SURFME PROJECTIONS, AND TYPGSPECIFIC ANTIGENWITY OF FLV
- _--___-~.- -.-~.-.-____ Treatmenta HA titer CF titer Protein Surface
h/ml) knobs” Glycoprotein Absorbing Infectivity*
(PAGE)C activity for (PFU,‘ml) neutralizing antibody”
----.- ~-~ ~ Bromelain <4 2048 0.66 0 - 0.11 <IO Control 4096 2048 0 .70 4+ + 1.07 3570
- CL Concent,rates of FLV were carefully prepared by 2 ultracentrifugations at pi 10 but without banding
on sucrose gradients. To 9 parts of fresh virus in pH 7.2 PBS was added 1 part bromelain solution (30 mg bromelain, 2 m Anson units/mg, in PBS, pH 7.2, containing 0.05 M DTT), and the mixture was incubated 60 min at 37°C. Control virus preparations were diluted 9: 1 with PBS, pH 7.2, and held at 4”. The cont,rol and treated materials were examined for surface knobs and centrifuged at pi 10 to remove bromelain and subviral structures. The resulting pellets were resuspended in pH 6.4 buffer and tested for HA, CF, protein, and absorption of neutralizing antibody. Material for PAGE, antibody absorption and infec- tivity tests was obtained from corresponding experiments which were identical to that described above except that control virus was incubated at 37” and the suspensions were freed of bromelain and subviral components by centrifugation on sucrose density gradients.
h See Fig. 3. c See Fig. 4; component corresponds to the -93,000 dalton glycoprotein described by Dnesberg eh (11.
(1970) and SchLfer et al. (197213). d Expressed as the percent of virus plaques (XC test) neutralized by unabsorbed FLV control serum
less the percent of virus plaques neutralized by absorbed serum divided by the percent of virus plaques neutralized by the unabsorbed serum. Each absorbing antigen was tested at a dilution of l:lO, i.e., pro- tein concentration of 0.066 to 0.070 mg/ml. Values of 1.0 indicate complete absorption; values of 0 indi- rate no absorption.
e XC test (Rowe et al., 1970).
sorbed by control FLV. Finally, as also re- ported for Rous sarcoma virus by Rifkin and Compans (1971), bromelain-treated FLV was noninfectious. These results were wholly reproducible in a series of cor- responding replicate experiments.
Because of the apparent association be- tween the loss of HA activity and the loss of surface knobs, glycoprotein(s) and type- specific antigenicity in bromelain-treated FLV, it seemed likely that all these ac- tivities or substances were related and were simultaneously removed by the bromelain treatment. However, the supernatant frac- tion of bromelain-treated FLV after pelleting at pi 10 lacked HA activity, and in parallel studies, isolated RLV hemagglutinin (R/ HA) was also completely inactivated by bromelain treatment. Thus, because of the inactivating action of bromelain on HA, the possibility that the HA subunit remained with the virion in an inactive form could not’ be completely excluded.
In other studies, we found no HA activity
associated with highly purified FLV cores (CF titer L 2048) isolated from ether- treated FLV by differential centrifugation. Furthermore, as we describe in the ac- companying report (Witter et al., 1973), the virion-associated HA activity of MuLV is accessible to inactivation by antibody. Thus there seems little doubt from these data and those of the previous section taken together that the hemagglutinin of MuLV is a surface component, possibly associated with the surface knobs.
Neuraminidase and Phospholipase C Treatment of MULTI
Treatment with neuraminidase and phos- pholipase C, as required for demonstration of HA, damaged to some extent the envelope of FLV particles (Fig. 5). Effects observed by electron microscopy ranged from com- plete or partial removal of the envelope to increased permeability as evidenced by in- creased penetration of uranyl acetat’e. Never- theless, surface knobs could be demonstrat,ed
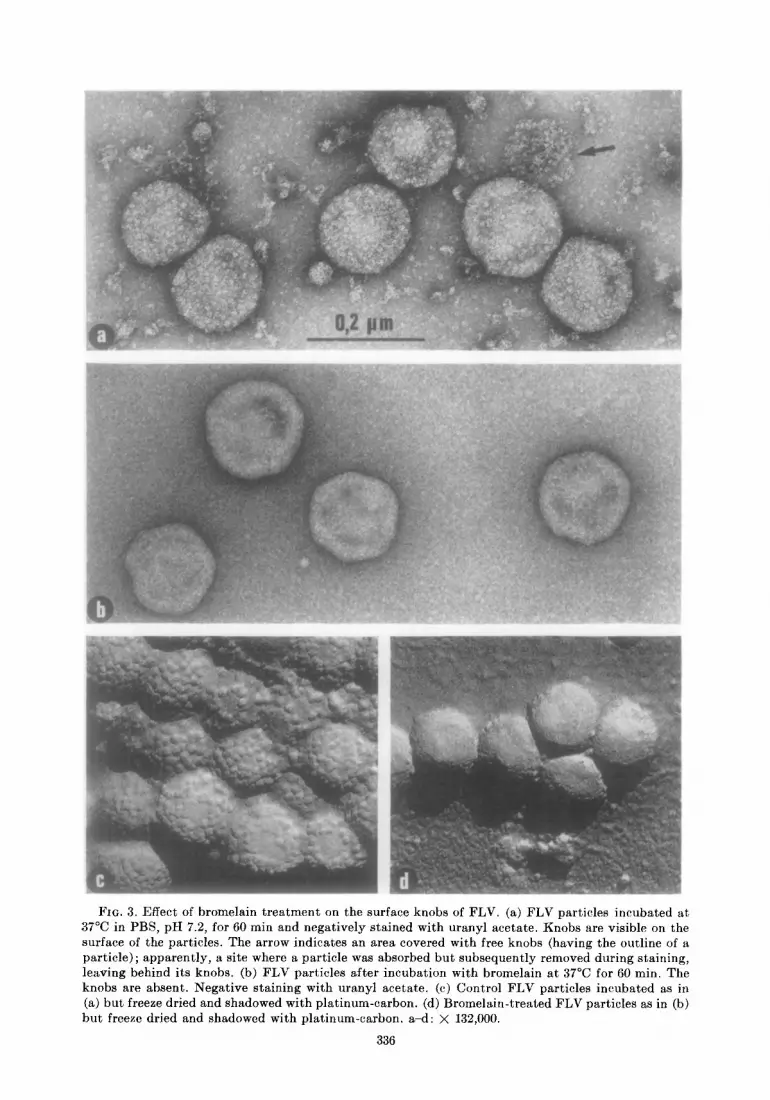
FIG. 3. Effect of bromelain treatment on the surface knobs of FLV. (a) FLV particles incubated at 37°C in PBS, pH 7.2, for 60 min and negatively stained with uranyl acetate. Knobs are visible on the surface of the particles. The arrow indicates an area covered with free knobs (having the outline of a particle); apparently, a site where a particle was absorbed but subsequently removed during staining, leaving behind its knobs. (b) FLV particles after incubation with bromelain at 37°C for 60 min. The knobs are absent. Negative staining with uranyl acetate. (c) Control FLV particles incubated as in (a) but freeze dried and shadowed with platinum-carbon. (d) Bromelain-treated FLV particles as in (b) but freeze dried and shadowed with platinum-carbon. a-d: X 132,000.
336
HRMAGGLUTINATION ASS.4Y 337
G-93000
34000
19000
14000 C
-bee a FIG. 4. Gel electrophoresis of reference FLV
(a), starting FLV (b), FLV incubated at 37” for 60 min without enzyme (c) and FLV incubated at 37” for 60 min with bromelain (d). Gels stained with Coomassie blue or PAS. Components in ref- erence gel are identified by approximate molecular weights (Schilfer et al., 197213). Note absence in bromelain-treated FLV (d) of glycoprotein (-93,000 daltons), and other components of high molecular weight.
on some of the particles, where the stain had penetrated less extensively. Naked cores and strands of material apparently derived from disintegrated cores could occasionally be seen. However, the enzyme-treated virus formed a single, homogeneous band in sucrose gradients at about 1.15 g/ml, com- pared to a density of 1.16 g/ml for bands formed by untreated virus (Fig. 6). The HA activity of enzyme-treated FLV was cbntirely associated with this 1.15 g/ml band.
Moreover, mor(l than 99% of th(l HA activity of enzyme-treated FLV prepara- tions suspended in buffer could be pcll(%ed at pi 10. These data suggested that some’ of the effects observed by electron microscop) may havcx been due t’o preparation artifacts. It was clear, nevertheless, that despitck tht> c’nzymc action? the HA activit’y and the surface knobs appeared to remain associated wit,h the larger particles.
Character of Isolatecl AIuLI’ Hema~gluhin~
Thtb HA activity of hemagglutinin isolated from rnzymr-treated, Twecn-ether drgradcd AIuLV I\-as found following citrate dcnsit> gradiclnt ccntrifugation t4htr at a d(>nsitg of 1.06 to 1.08 g/ml in association with the lower portion of thr upper visibhb band as previously described (Schiifer et al., 1 Y72b), or at dtinsitios of 1.12-l .17 g/ml whtxn> it was oftctn associated wi-ith a distinct, h(+ero- gc~neous band. In one trial, 1.08 g/ml HA matrlrial (R/HA) formed a hcterogc~ncous- appearing band at 1.12 g/ml upon reccnt,rif- ugation in a citratch gradient., possibly dut> t,o removal of lipids from the HA substruct,urc by shearing forces. The sprcific HA activities of isolated hemagglutinins were markcdl3 grcxatc>r t,han obtained for cxnzymc-treat,cd hluLV (Table 3; compare v&h Table 1 1, indicating a substantial degree of purifica- t’ion. There were also some differenccbs in specific activities of different hemaggluti - nins. For Cxample, RLV-derived hernag- glutinin (R/HA) appeared to contain more gs antigen and Ma1 protein relativr to its HA titer than FLV-derived F/HA or FS/ HA hemagglutinins (Table 8), probably because> it was derived from the top of the gradient where, under these conditions of centrifugation, the gs protein is also prc>sclnt (Sch5ifer et al., 1972b).
The t,itcr of R/HA hemagglutinin in PBS, pH 7.2, varied no more t,han Z-fold following (i) t,rcat,ment in a Bransonic “ 12” ultrasonic vibrator (Branson Instrumcbnts, Munich) for 8 min at 22”, (ii) rebanding in a citrate gradient (SW 50 rotor, 50,000 rpm, 2 hr), (iii) storage for up to 6 mont,hs at -2O”, or (iv) 3 rapid frecze-t8ha\v cycles. HA activit,y of both isolat,cd hemagglutinin (R/HA) and rnzym+trc,atcd intact FT,V
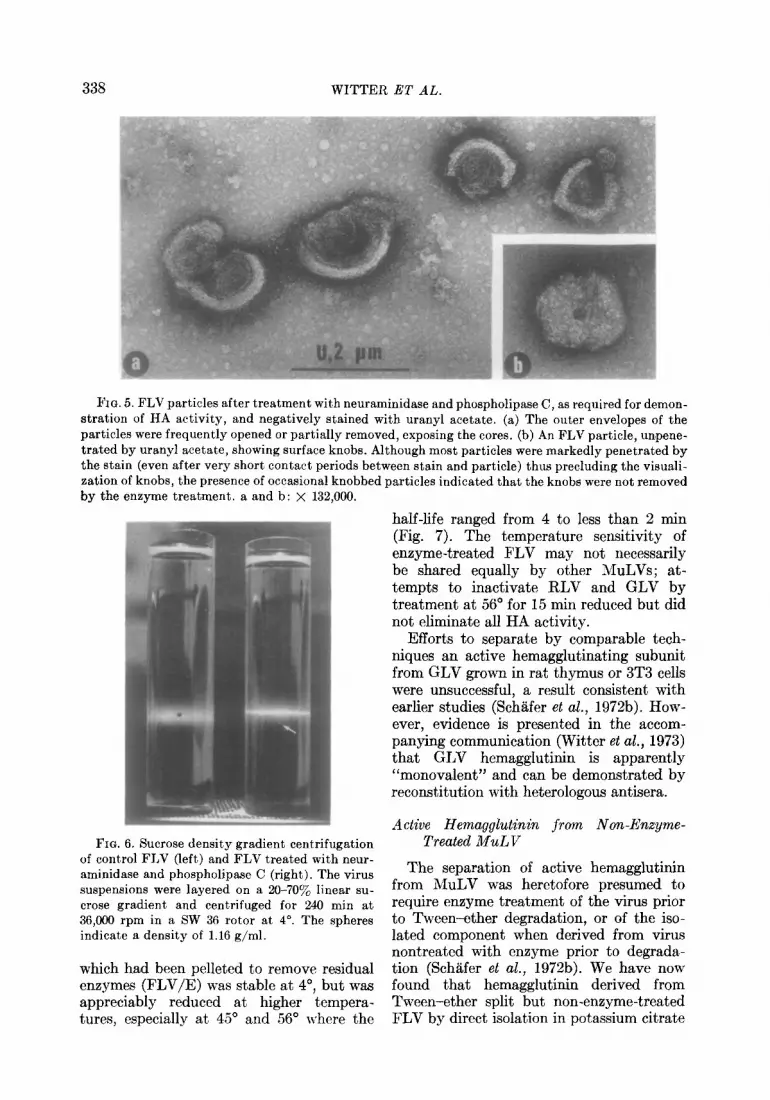
338 WITTER ET AL.
FIG. 5. FLV particles after treatment with neuraminidase and phospholipase C, as required for demon- stration of HA activity, and negatively stained with uranyl acetate. (a) The outer envelopes of the particles were frequently opened or partially removed, exposing the cores. (b) An FLV particle, unpene- trated by uranyl acetate, showing surface knobs. Although most particles were markedly penetrated by the stain (even after very short contact periods between stain and particle) thus precluding the visuali- zation of knobs, the presence of occasional knobbed particles indicated that the knobs were not removed by the enzyme treatment. a and b: X 132,000.
half-life ranged from 4 to less than 2 min (Fig. 7). The temperature sensitivity of enzyme-treated FLV may not necessarily be shared equally by other MuLVs; at- tempts to inactivate RLV and GLV by treatment at 56” for 15 min reduced but did not eliminate all HA activity.
Efforts to separate by comparable tech- niques an active hemagglutinating subunit from GLV grown in rat thymus or 3T3 cells were unsuccessful, a result consistent with earlier studies (Schiifer et al., 197213). How- ever, evidence is presented in the accom- panying communication (Witter et al., 1973) that GLV hemagglutinin is apparently “monovalent” and can be demonstrated by reconstitution with heterologous antisera.
FIG. 6. Sucrose density gradient centrifugation of control FLV (left) and FLV treated with neur- aminidase and phospholipase C (right). The virus suspensions were layered on a 20-707, linear su- crose gradient and centrifuged for 240 min at 36,000 rpm in a SW 36 rotor at 4”. The spheres indicate a density of 1.16 g/ml.
which had been pelleted to remove residual enzymes (FLV/E) was stable at 4’, but was appreciably reduced at higher tempera- tures, especially at 45’ and 56” where the
Active Hemagglutinin from Non-Enzyme- Treated MuLV
The separation of active hemagglutinin from MuLV was heretofore presumed to require enzyme treatment of the virus prior to Tween-ether degradation, or of the iso- lated component when derived from virus nontreated with enzyme prior to degrada- tion (Sch8fer et al., 1972b). We have now found that hemagglutinin derived from Tween-ether split but non-enzyme-treated FLV by direct isolation in potassium citrate

HEMAGGLUTINATIOK ASSAY
TABLE 3
Hemagglutinin
PROPERTIES OF ISOLATED MuLV HI~:M.~GGIXTINISS __- ~_ ~-~~.- ~~ ~~~- -. ~~ ~ -- ~~~
Source virus Density of peak H.\ titer Specific activity fraction” (mg/ml)
HA:CF H.1 units per gg protein
R/HAh RLV 1.08 1024 4 2tiKO F/HA6 FLV 1.17 256 8 x540 N/H& FLV 1.12 12x 16 7320
‘I From potassium citrate density gradients. b Isolated from neuraminidase and phospholipase C-treated, Tween 80 and ether-degraded MuLV by
centrifugation on potassium citrate gradients. c Isolated as above except that the Tweet)-ether concnentrate was c~hromatographed on Sephadex
G-150 prior to density gradient centrifugation.
FLVIE
2 4 , I I I 1 I I I I 1 I 0 2 4 8 16 32 64 128 0 2 4 8 16 32 64 128
minutes
FIG. 7. Temperature sensitivity of the HA activity of RLV isolated hemagglutinin (R/HA) and en- zyme-treated FLV (FLV/E). Temperature: 4”C, 60; 37”C,O---0; 45”C, H; 56”C, O-.-II.
gradients possesses HA activity that is not dependent on or enhanced by subsequent enzyme treatment. This material was re- covered from a broad region (peak at 1.14 g/ml) of the gradient (Fig. 8). The HA activity of this material could be strongly inhibited by immunoglobulin G (IgG) ob- tained from the serum (FV525/1) of a rabbit immunized with SDS-degraded FLV, but not by IgG from normal rabbit sera. Proto- cols for HI tests are given in the accom- panying communication (Witter et al., 1973).
Further analysis of this HA material re- vealed no important differences compared to chemical and serological properties pre- viously reported for MuLV hemagglutinin (Schiifer et al., 197213). The material as studied by PAGE (Fig. 9) apparently con- tained the -93,000 daltons glycoprotein although curiously as 2 PAS-positive com- ponents (possibly aggregation products) in- stclad of the usual single band (Schiifer et al.,
1972b). As another major component a protein appeared in the 19,000 daltons position.
The material absorbed neutralizing anti- body when tested against FTE/H serum (absorbing activities of 0.49 and 0.15 at 1:25 and 1:50 dilutions of antigen; see legend Table 2). This was in agreement with SchHfer et al. (1972b), who reported that hemagglutinating substructures isolated from Twcen-ether degraded FLV at a den- sity of 1.06 g/ml absorbed homologous FLV but not heterologous GLV neutralizing anti- body. When analyzed in Ouchterlony tests against RSDS/2 serum, the HA material contained principally antigens Igs and IIv- gs, with only a minor amount of antigen IVgs (Schgfer et al., 1972a) (Fig. 10). Thus, these data provide direct evidence that the hemagglutinin exists in situ in the MuLV particle and is revealed or uncovered rather
340 WITTER ET AL.
HA OD280 I
2
1
3.0
1 ZO-
g/ml
LO-
5a- \
LO-
3.0 -
2D-
IO- ‘\,/’
I *%./ •/--~
HA / 1-0
--- 5 10 15 fraction number
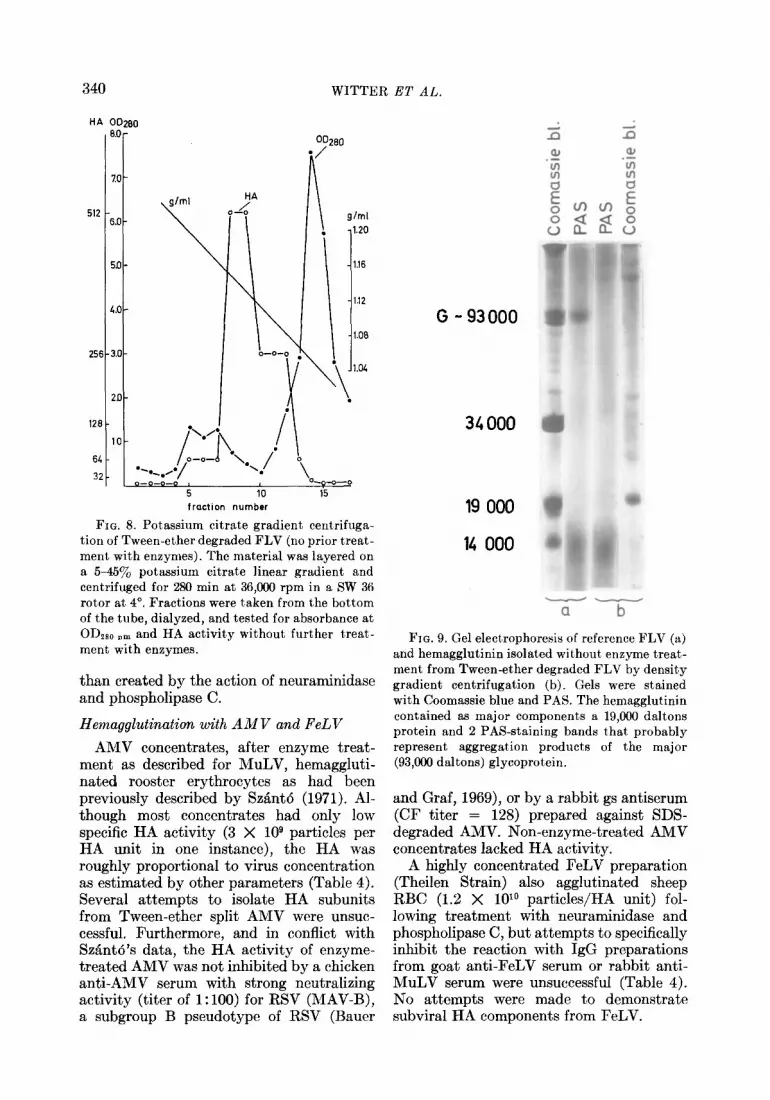
FIG. 8. Potassium citrate gradient centrifuga- tion of Tween-ether degraded FLV (no prior treat- ment with enzymes). The material was layered on a 55457& potassium citrate linear gradient and centrifuged for 280 min at 36,000 rpm in a SW 36 rotor at 4”. Fractions were taken from the bottom of the tube, dialyzed, and tested for absorbance at ODzso nm and HA activity without further treat- ment with enzymes.
than created by the action of neuraminidase and phospholipase C.
Hemagglutination with AMV and FeLV
AMV concentrates, after enzyme treat- ment as described for MuLV, hemaggluti- nated rooster erythrocytes as had been previously described by Szant6 (1971). Al- though most concentrates had only low specific HA activity (3 X log particles per HA unit in one instance), the HA was roughly proportional to virus concentration as estimated by other parameters (Table 4). Several attempts to isolate HA subunits from Tween-ether split AMV were unsuc- cessful. Furthermore, and in conflict with Szant6’s data, the HA activity of enzyme- treated AMV was not inhibited by a chicken anti-AMV serum with strong neutralizing activity (titer of 1: 100) for RSV (MAV-B), a subgroup B pseudotype of RSV (Bauer
-- a b
FIG. 9. Gel electrophoresis of reference FLV (a) and hemagglutinin isolated without enzyme treat- ment from Tween-ether degraded FLV by density gradient centrifugation (b). Gels were stained with Coomassie blue and PAS. The hemagglutinin contained as major components a 19,000 daltons protein and 2 PAS-staining bands that probably represent aggregation products of the major (93,000 daltons) glycoprotein.
and Graf, 1969), or by a rabbit gs antiserum (CF titer = 128) prepared against SDS- degraded AMV. Non-enzyme-treated AMV concentrates lacked HA activity.
A highly concentrated FeLV preparation (Theilen Strain) also agglutinated sheep RBC (1.2 X lOlo particles/HA unit) fol- lowing treatment with neuraminidase and phospholipase C, but attempts to specifically inhibit the reaction with IgG preparations from goat anti-FeLV serum or rabbit anti- MuLV serum were unsuccessful (Table 4). No attempts were made to demonstrate subviral HA components from FeLV.

HEMAGGLUTINATION ASSAY 341
Evidence for the specificity of the HA reaction with XluLV has been given prc- viously (Schgfer and S&&6, 1969; Sch%fer et al., 1972b). However, HA-like reactions occasionally wcr(’ obtained with enzyme
FIG. 10. Ouchterlony tests with the hemsgglu- tinin (HA) isolated from Tween-ether degraded FLV without enzymes (see Fig. 8) tested against RSl)S/S serum (center well). Tween-ether de- graded FLV serves as a reference antigen. Dem- onstrated in both reference FLV and HA are anti- gens Igs and II T+a< ilntigen IVgs (gs-1 and gs-2; gs-3 not demonstrated by serum) is present, in HA in very low amounts.
(neuraminidase) controls which proved to be associated with the use of PBS, pH 7.2. This problem was successfully resolved bq the use of pH 6.4 buffer as previously rccom- mended by SchBfer and SzW6 (1969) and by careful control of the incubation tempera- ture. Under these conditions, background HA-like reactions in neuraminidase controls were eliminated. Hemolysis caused by phos- pholipase C could be controlled by inclusion of a control substrate such as heat-ina&- vated l\luLV.
Other cvidencc for specificity oht,ained in these experiments included linear dose de- pendency (over at least a limited range) and good correlation of HA titers of intact, knobbcd virions with particle count,s, CF titers, and protein. Failure to obtain linear dose response relationships with lower doses of virus w-as possibly due to oxcessivcb M- zyme concentrations, which were maintained at a constant level over all virus dilutions. Variation of enzyme concentrations with the amount, of protein in the sample migth be investigated as a means of improving t,he
TABLE 4
INDU(~TION OF HA IIY I<NZYME-TRK’:BTED AMV IND FeLV ASD COMPA~~ISON WITH OTHI:R PLRAMI<:TEIM -__._
Virus Material Purification HA titer CF titer ATPase method*
Virus particles Inhibition activity& per ml of HA I,>
specific antiserum __.~
AMV Plasma C 12x 4096 13.3 No, Plasma c + 2G 32 1024 x .o CEF fltlid C 16 1024 CEF fluid c i- 2G 4 256 Plasma C + 2G 32 x192 4.0 2 x 10’2
FeLl Zonal + PD 32 x x IO’% NW ~-.--.--~~
(L Abbreviations: CEF = chick embryo fibroblast culture, C = ultracentrifugation at pi 10, 2G = dis- caontinuous sucrose density gradient followed by continuous surrose density gradient centrifugation, Zonal = zonal centrifugation, PI) = pressure dialysis,
h Expressed as units/ml, each unit consisting of that amount. of ATPase activity capable of reacsting with the test reagent in 2 min (Mommaerts et al., 1954).
( Twofold dilutions of heated anti-AMV chicken serum and anti-AM\’ gs rabbit, serum were re- acted with 8 HA units of AMV plasma pellet antigen which cont,ained neuraminidase and phospholi- pase C. After incubat,ion of serum-virus mixtures for 40 min at 4”, washed chicken erythrocaytes were added and the test was held 2 hr at 4’ prior to reading. HI t,iters of both sera were <4.
d Particle count, courtesy of Pfizer Research Laboratories, Maywood, New Jersey. p IgG preparations of anti-FeSV goat serum FeSV/22 and anti-FeLV goat serum FeLV/2 were serially
dilut)ed and react.ed with 8 HA units of FeLV antigen prepared by treatment with neuraminidase and phospholipase C but, centrifuged at pi 10 to remove residual enzymes. After incubation for 40 min at, 22”, washed sheep erythrocytes were added. The test was read after 2 hr at 4’. Further details on sera and HI procedures are in the accompanying report (Wit,ter et a/., 1973). All sera lacked HI activity at a 1:” dilution.

342 WITTER ET AL.
sensitivity with which low concentrations of MuLV may be detected. Extensive trials involving the inhibition of HA by IgG preparations of specific MuLV antisera are presented in the accompanying report (Witter et al., 1973) and provide con- firmatory evidence that the HA induced by neuraminidase and phospholipase C- treated MuLV is associated with a virus- specific substructure.
In these trials, approximately 5 X 10’ enzyme-treated, knob-containing FLV par- ticles (values for 4 preparations were 2.9 X lo’, 5.0 X lo’, 5.6 X lo’, and 6.1 X 10’) were equivalent to one HA unit. Cor- responding values reported for influenza virus averaged about 1.6 X 10’ particles per HA unit (Donald and Isaacs, 1954) ; how- ever, the data are not strictly comparable because of differences in volume used in defining HA units.
The recent identification of surface pro- jections on MuLV particles by Nermut et ab. (1972) raised the intriguing possibility that these projections were physical sites of HA activity, as is the case with myxoviruses. Indeed, a positive correlation between HA titer and surface knobs was obtained on a limited number of samples. Furthermore, bromelain treatment of MuLV simultane- ously removed the projections and abolished all HA activity. Bromelain-treated virus also lacked glycoprotein(s) and type-specific antigens as revealed by its inability to absorb neutralizing antibody. However, failure to purify and isolate the projections as distinct subunits with directly demonstrable HA activity has thus far prevented confirmation of this possibility. It has been observed that during normal centrifugation of MuLV in sucrose gradients, some HA-active material is present near the top of the gradient, clearly distinct from the 1.16 g/ml virus band (Schafer, unpublished data). This material could represent surface projections since density gradient centrifugation tends to physically remove such projections (Frank, unpublished data). In any case it can be concluded that HA activity resides at the surface of the particles. Further support for this is provided by the absence of HA ac- tivity in purified MuLV cores and by the accessibility of the virus-associated hemag-
glutinin to inhibition by antiserum (Witt’er et al., 1973).
In different experiments, isolated hemag- glutinin banded at varying densities in citrate gradients Moreover, HA material isolated at higher densities (1.12 to 1.17 g/ml) was usually associated with visible, heterogeneous bands whereas hemagglutinins from the top of the gradients (1.06 to 1.08 g/ml) formed no distinct bands, possibly due to interference by lipids. Since the HA sub- unit itself is probably associated with some lipid (Schafer e2 al., 1972b), the material banding at higher densities may represent hemagglutinin from which some lipid has been removed by shearing forces during density gradient centrifugation.
The HA activity of MuLV hemagglutinin preparations was reasonably stable following various physical treatments such as freeze- thaw, ultrasonic vibration and recentrifuga- tion on citrate gradients. However, the HA activity of isolated RLV hemagglutinin and enzyme-treated FLV were both heat sensi- tive with a half-life of 4 min or less at 45” and higher. The HA activities of enzyme- treated RLV and GLV seemed less sensitive to inactivation at 56” than that of FLV; however, this also could be related to the different host cells in which the viruses were, respectively, propagated.
The chemical composition of the MuLV hemagglutinating substructure has not been fully elucidated; however, our data in com- bination with those reported earlier (Schafer et al., 1972b) provide some insight into this question. It seems virtually certain that glycoprotein is involved. A major glycopro- tein was contained in hemagglutinin isolated from non-enzyme-treated FLV, and bro- melain-treated FLV lacked this glycoprotein as well as HA activity. It may well be that, as in the case of AMV (Bolognesi et al., 1972), a minor glycoprotein (Duesberg et al., 1970; Schafer et al., in press), which in our system escaped detection by the methods used, exists in association with the former. By comparison it is interesting that the hemagglutinin of influenza virus is a surface complex of two glycoproteins (Schultze, 1972) which, when released from the virion, aggregate in association with lipid to form HA-active, multimeric structures (Laver and

HEMAGGLUTINATION ASSAY 3 43
Valentine, 1969). Type-specific and certain more broadly reacting antigens have also been related to surface glycoproteins of AMV (Bolognesi et al., 1972), and it may be pertinent that such antigens are also as- sociated with isolated MuLV hemagglutinin. Antigen Igs which is detectable in Ouchterlony tests with RSDS/2 serum is probably not a component of the hemag- glutinin since it is removed by pretreatment of virus with neuraminidase and phospho- lipase C (Schtifer et al., 197213). The variable sedimentation in density gradients suggests that a lipid moiety may also be involved. The finding that hemagglutinin can be isolated without enzyme treatment indicates that the hemagglutinating substructure exists on or within the virus particle and is not an artificial product of enzyme action.
Although both AMV and FeLV produced some HA with chicken and sheep erythro- cytes, respectively, the specificity of these reactions remained equivocal. Compared to MuLV, the HA activity of AMV was about 60-fold less per virus particle and was not inhibited by gs or type-specific antisera. Furthermore, no HA subunits could be re- covered by methods applicable to the isola- tion of MuLV hemagglutinin. These results are consistent wit’h data of Bauer and Bolognesi (personal communication) but conflict, at least in part, with that of SzBnt6 (1971), who reported inhibition of AMV- induced HA by specific antiserum. In our trials, whole serum was used for HI tests since such sera lacked nonspecific inhibitory activity against AMV HA antigen. Similarly, t,hc HA activity of FeLV was 250-fold less per particle than for MuLV and also could not be inhibited by specific FeLV or MuLV antiserum (IgG). However, the existence of hemagglutinins in these viruses should prob- ably not be excluded. Variations in type of erythrocytes or conditions of hemagglutina- tion were not studied. Furthermore hemag- glutinins are normally complex structures which can be rendered monovalent during isolation procedures and, therefore, may require special techniques for their demon- stration. Such appears to be the case for GLV, since the isolat’ed hemagglutinating substructure lacks HA activity unless re- const,ituted with het’erotypic antiserum
(Witt.er el al., 1973). Thus, it is possible that further efforts to demonstrate specific HA substructures in AMV and FeLV and other RNA tumor viruses might yet be successful.
The HA reaction has several possible ap- plications t,o studies on MuLV and mouse leukemia. When used with another param- eter such as CF, the relative (specific) HA activity of a virus concentrat’e should indi- cate whether surface knobs are present and thus provide a measure of virion intactness without involving electron microscopy. The usefulness of HA for quantifying virus suspensions is probably restricted to viral concentrates but could, in such cases, have advantages over other methods. Once an HA:particle ratio is established for a given virus and purification system, it should bc possible to use HA to directly quant’itate viral preparations. Indeed, such methods are in routine use at this laboratory. HA act,ivity was not’ directly correlated with infectivity in these studies; however, FLV particles from which surface knobs had been removed by bromelain treat,ment lacked infectivity as well as HA activity. A further application of HA procedures is in the pro- duction of antigens with HA activity for use in serologic tests as described in t#hc accompanying paper (Witter et al., 1973). Finally, hemagglutinating surface compo- nents of MuLV may prove to have dir& biological significance. The so-called G sol- uble antigen (GSA) present’ in the plasma of GLV-infect,ed mice is an indic,ator of the probability of leukemia development8 (Ploki et al., 1968) and, furthermore. immune complexes of GSA and its antibody have been ctiological!? linked to an autoimmune glomerulonephrltls in XZB mice (Mellors et al., 1971). Because of it,s size (passes 10 nm filters) and type-specific antigenicity (Aoki et al., 1968), GSA bears a striking re- semblance to the surface knobs of MuLV. Since the surface knobs are loosely att8achcld to the virion (Nermut, et al., 1972; SchSifer et al., in press), it is conceivable that defolia- tion of surface knobs may also occur in ~:i~o. It would seem highly interesting to examine MuLV-infected mice for hemagglutinating soluble ant,igens because, if suc:h were dem- onstrated, the HA assay might prove to be a powerful tool in the study of biologi&

344 WITTER ET AL.
interactions between MuLV’s and their hosts.
ACKNOWLEDGMENTS
The authors thank L. Pister, E. Seifert, and D. Willutzki for excellent technical assistance.
REFERENCES
AOKI, T., BOYSE, E. A., and OLD, L. J. (1968). Wild-type gross leukemia virus. I. Soluble anti- gen (GSA) in the plasma and tissues of infected mice. J. Nat. Cancer Inst. 41, 89-96.
BAUER, H. (1970). Untersuchungen iiber Struktur, Vermehrung und onkogene Wirkung der Hiihner- Leukose-Sarkomatose-Viren. Zentralbl. Veteri- naermed. Reihe B 17,582430.
Baus~, H., and GRAF, T. (1969). Evidence for the possible existence of two envelope antigenic de- terminants and corresponding cell receptors for avian tumor viruses. Virology 37, 157-161.
BOLOGNESI, D. P., and BAUER, H. (1970). Polypep- tides of avian RNA tumor viruses. I. Isolation and physical and chemical analysis. Virology 42, 1097-1112.
BOLOGNESI, D. P., BAUER, H., GELDERBLOM, H., and H~~PF,R, G. (1972). Polypeptides of avian RNA tumor viruses. IV. Components of the viral envelope. Virology 47, 551-566.
COMPANS, R. W., KLENK, H. D., CALIGUIRI, L. A., and CHOPPIN, P. W. (1970). Influenza vi- rus proteins. I. Analysis of polypeptides of the virion and identification of spike glycoproteins. Virology 47, 880-889.
DONALD, H. B., and ISA.%CS, A. (1954). Counts of influenza virus particles. J. Gen. Microbial. 10, 457-464.
DUESBERG, P. H., MARTIN, G. S., and VOGT, P. K. (1970). Glycoprotein components of avian and murine RNA tumor viruses. Vz’rology 41, 631-646.
GIEBLER, P. (1958). Definierung und Auswertung priparativer Ultrazentrifugierungen mit Hilfe des Performance Index (pi). Z. Naturforsch. B 13, 238-241.
GROSS, L. (1959). Agglutinating action of heat- inactivated passage A mouse leukemia filtrates on mouse red blood cells. Proc. Sot. Exp. BioZ. Med. 101, 113-117.
HORZINEK, M. (1969). A simple method for con- centration of arboviruses propagated in tissue culture. Amer. J. Trop. Med. Hyg. 18, 588-591.
HORZINEK, M., MA~SS, J., and LAUFS, R. (1971). Studies on the substructure of Togaviruses. IT. Analysis of equine arteritis, rubella, bovine vi- ral diarrhea, and hog cholera viruses. Arch. Ges. Virusforsch. 33, 306&318.
IOliCaIM, H. L. (1969). Divergence in tumor induc-
tion of thymus cells transformed in vitro by Gross leukemia virus. J. Nat. Cancer Inst. 42, 101-113.
KELLENBERGER, E., and ARBER, W. (1957). Elec- tron microscopical studies of phage multiplica- tion. I. Method for quantitative analysis of par- ticle suspensions. Virology 3, 245-255.
LAVER, W. G., and VALENTINE:, R. C. (1969). Mor- phology of the isolated hemagglutinin and neur- aminidase subunits of influenza virus. Virology 38, 105-119.
LOWRY, 0. H., ROSEBROUGH, H. J., FARR, L., and RANDALL, R. J. (1951). Protein measurement with Folin phenol reagenti J. Biol. Chem. 193, 265-275.
MELLORS, R. C., SHIRAI, T., AOKI, T., HUEBNER, R. J., and KRAWCZYNSKI, K. (1971). Wild-type Gross leukemia virus and the pathogenesis of the glomerulonephritis of New Zealand mice. J. Exp. Med. 133, 113-132.
MOMMAERTS, E. B., SHARP, 1). G., ECKERT, E. A., BEARD, D., and BEARD, J. W. (1954). Virus of avian erythromyeloblastic leukosis. I. Relation of specific plasma particles to the dephosphoryl- ation of adenosine triphosphate. J. Nat. Cancer Inst. 14, 1011-1025.
NERMUT, M. V., FRANK, H., and SCHAFER, W. (1972). Properties of mouse leukemia viruses. III. Electron microscopic appearance as re- vealed after conventional preparation tech- niques as well as freeze-drying and freeze- etching. Virology 49, 345-358.
RAPP, F., STEINGLASS, M., and FRIEND, C. (1962). Hemagglutination by Swiss mouse leukemia vi- rus. (Abstract.) Bacterial. Proc. 21, 134.
RIFKIN, D. B., and COMPANS, R. W. (1971). Iden- tification of the spike proteins of Rous sarcoma virus. Virology 46, 485-489.
ROWE, W. P., PUGH, W. E., and HARTLE:Y, J. W. (1970). Plaque assay techniques for murine leu- kemia viruses. Virology 42, 1136-1137.
SCHAFER, W., and SEIFERT, E. (1968). Production of a potent complement-fixing murine leukemia virus antiserum from the rabbit and its reactions with various types of tissue culture cells. Viral- ogy 35, 323-328.
SCH;~FER, W., and SZANT~, J. (1969). Studies on mouse leukemia viruses. II. Nachweis eines virus-spezifischen Hlmagglutinins. Z. Natur- forsch. B 24, 1324-1331.
SCHAFER, W., LANGE, J., PISTER, L., SEIFERT, E., DF,NORONHA, F., and SCHMIDT, F. W. (1970). Vergleichende serologische Untersuchungen iiber LeukCmieviren. Eine Komplementbindungsre- aktion zum Nachweis der bei Leukilmieviren verschiedener SLuger vorkommenden gemein- samen antigenen Komponente. Z. Naturforsch. B 25, 1029-1036.

HEMAGGLUTINATION ASSAY 345
SCHUER, W., FISCHINGKH, J., LANGE, J., and POSTER, L. (1972a). Properties of mouse Leuke- mia viruses. I. Characterization of various anti- sera and serological identification of viral com- ponents. Virology 47, 197-209.
SCH~~FI.:R, W., L.IN(;Ic, J., FISCHINGER, P. J., FR.ZNK, H., BOLOGNESI, D. P., and PISTER, L. (1972b). Properties of mouse Leukemia viruses. II. Isolation of viral components. virology 47, 210-228.
SCH~FIX, W., B.\w:a, H., BOLOGNESI, D. P., FISCHINC;I:I~, P., FRANK, H., GELDERBLOM, H., L~NGK, J., and NERMUT, M. V. (in press). Stud- ies on strrlct,ural and antigenic properties of C-type viruses. Proc. 25th Symp. Fundament.
Cancer Res., M. D. Anderson Hospital and Tu- mor Inst,itute, Houston, Texas, 1972.
SCHULTZL, I. T. (1972). The st,ructure of infIuenza virus. II. A model based on the morphology and composition of subviral particles. T’irology 47, 181-196.
SHANTY, J. (1971). Hemagglutination reaction with avian Myeloblastosis virus. Acta Viral. (Pragtrr I 15, 245-218.
WITTER, 1~. L., HUNSMANN, G., L.IN(;I,;, J., and SCH;~FER, W. (1973). Properties of 1110118e leu- kemia viruses. V. Hemagglutination-inhibition and indirect hemagglutination test)s. Virolog?l 54, 346-358.

![Analysis of proteins of mouse sarcoma pseudotype viruses: type-specific radioimmunoassays for ecotropic virus p30's. [¹²âµI tracer technique]](https://img.dokumen.tips/doc/110x75/635d7238a0f1eac29f0c3205/analysis-of-proteins-of-mouse-sarcoma-pseudotype-viruses-type-specific-radioimmunoassays.jpg)



























