Embed Size (px)
Citation preview

MI67CH12-Cowman ARI 5 August 2013 16:34
Plasmodium Nesting: Remakingthe Erythrocyte from theInside OutJustin A. Boddey and Alan F. CowmanDivision of Infection and Immunity, The Walter and Eliza Hall Institute of Medical Research,Parkville, Victoria 3052, Australia; email: [email protected], [email protected]
Department of Medical Biology, University of Melbourne, Parkville, Victoria 3010, Australia
Annu. Rev. Microbiol. 2013. 67:243–69
First published online as a Review in Advance onJune 26, 2013
The Annual Review of Microbiology is online atmicro.annualreviews.org
This article’s doi:10.1146/annurev-micro-092412-155730
Copyright c© 2013 by Annual Reviews.All rights reserved
Keywords
malaria, trafficking, export, PEXEL, translocon
Abstract
One of the most fascinating and remarkable features of Plasmodium parasites,which cause malaria, is their choice of erythrocytes as the principal host cellsin which to reside during infection of a vertebrate host. Parasites completelyrenovate the terminally differentiated cells, which lack most of the normalorganelles and functions of other cells, such as a nucleus and the machin-ery to express and transport proteins to subcellular locations. Erythrocyteremodeling begins immediately after invasion by the Plasmodium parasite,by expression and export of many hundreds of proteins that assemble intomolecular machinery in the host cell that permit protein trafficking, har-vesting of nutrients, and mechanisms to evade host immune responses. Inthis review, we discuss recent studies on erythrocyte remodeling, includingmechanisms of protein export as well as the identity, functions, and subcel-lular locations of key exported proteins.
243
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
Protein export:transport of malarialproteins from withinthe parasite into theinfected erythrocyte
Contents
INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 244MALARIA THE DISEASE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 244ERYTHROCYTE REMODELING . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 246MECHANISMS OF PROTEIN EXPORT . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 248
Role of the PEXEL/VTS in Export. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 248Plasmepsin V and the Matured N Terminus of PEXEL Proteins . . . . . . . . . . . . . . . . . . 249How Are PNEPs Exported? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 249How Is PfEMP1 Exported? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 251Role of Phosphatidylinositol-3-Phosphate . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 257
PROTEIN EXPORT MACHINERY . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 258EXPORTED PROTEINS AND THEIR FUNCTIONS . . . . . . . . . . . . . . . . . . . . . . . . . . . 259
The Exportome of Plasmodium Spp. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 259Cytoadherence Complex . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 260PfEMP1 Trafficking and Display on the Erythrocyte Surface . . . . . . . . . . . . . . . . . . . . . 261Enhancing Erythrocyte Rigidity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 261New Permeability Pathways . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 261
TIMING OF EVENTS IN EXPORT . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 262CONCLUSIONS AND OUTLOOK. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 262
INTRODUCTION
Malaria is a mosquito-borne disease caused by infection with Plasmodium parasites. The parasitesare transmitted by an arthropod vector (Anopheles mosquitoes) to vertebrate hosts, which includebirds, reptiles, rodents, and primates (including humans). Five Plasmodium species infect humans,namely P. falciparum, P. vivax, P. ovale, P. malariae, and P. knowlesi; however, infection withP. falciparum causes the most severe disease. Establishment of infection is dependent on the par-asite’s ability to profoundly remodel erythrocytes. Cellular remodeling engineers an intracellularniche that fosters evasion of host immune responses and scavenging of host nutrients, permittingthe parasite to replicate and thrive. This remarkable remodeling process occurs through the ex-pression of a large repertoire of specialized proteins that are trafficked through a sophisticatedexport pathway into the host cell (38, 56, 78, 94). The renovation of host erythrocytes by exportedparasite proteins ensures the parasite’s survival but also contributes to virulence and pathogenesisof malaria (reviewed in 55).
In this chapter, we discuss the current state of knowledge of protein export and host cell remod-eling by Plasmodium, with an emphasis on P. falciparum, for which most information is known. Wediscuss the key exported proteins deployed by parasites, their functions in virulence and remod-eling, and the mechanisms that deliver them to their subcellular locations within erythrocytes.
MALARIA THE DISEASE
In 2010, malaria caused 216 million infections and 655,000 deaths; 91% of cases occurred inAfrica and 86% of all deaths occurred in children less than five years old (103). Almost half ofthe world’s population is at risk of malaria infection and over 100 countries are endemic for thedisease, primarily in sub-Saharan Africa, Southeast Asia, the Indian subcontinent, South America,
244 Boddey · Cowman
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.
MI67CH12-Cowman ARI 5 August 2013 16:34
Sporozoite invasion
Macrogametocyte
Exflagellatedmicrogametocyte
Zygote
Ookinete
Oocyst
Sporozites
Salivaryglands
Mosquito
Liver stage
Ring
Sporozites
Trophozoite
Schizont
Egress
Bloodstage
Sexualstage
Merozoite invasion
Gametocytes
Mosquitoblood meal
Mosquitobloodmeal
Oocyst rupture
Temperatureshift
Midgut
Basallamina
Hepatocyteegress
Hepatocyte
Human host
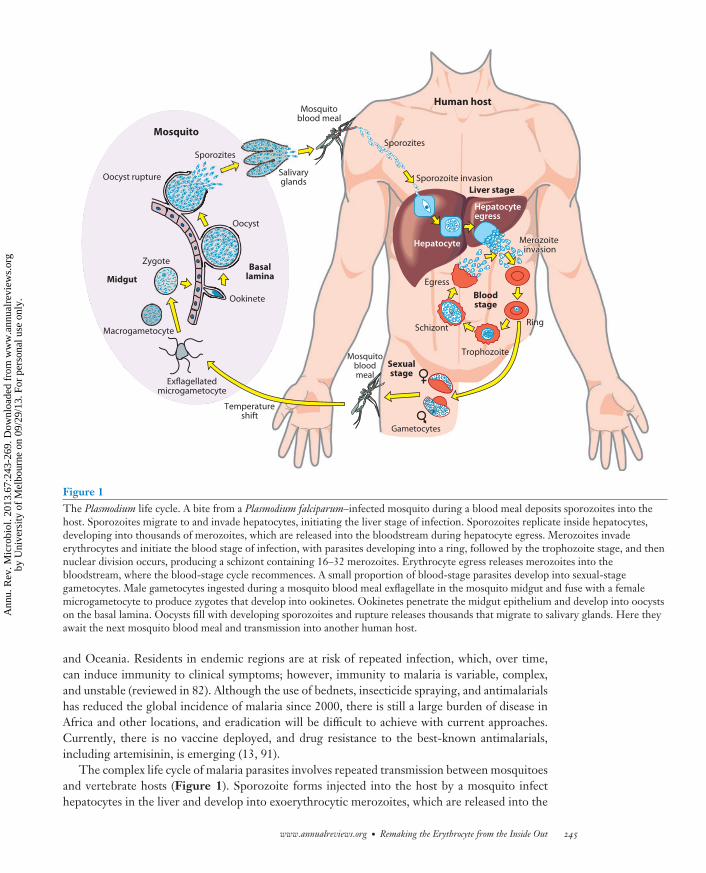
Figure 1The Plasmodium life cycle. A bite from a Plasmodium falciparum–infected mosquito during a blood meal deposits sporozoites into thehost. Sporozoites migrate to and invade hepatocytes, initiating the liver stage of infection. Sporozoites replicate inside hepatocytes,developing into thousands of merozoites, which are released into the bloodstream during hepatocyte egress. Merozoites invadeerythrocytes and initiate the blood stage of infection, with parasites developing into a ring, followed by the trophozoite stage, and thennuclear division occurs, producing a schizont containing 16–32 merozoites. Erythrocyte egress releases merozoites into thebloodstream, where the blood-stage cycle recommences. A small proportion of blood-stage parasites develop into sexual-stagegametocytes. Male gametocytes ingested during a mosquito blood meal exflagellate in the mosquito midgut and fuse with a femalemicrogametocyte to produce zygotes that develop into ookinetes. Ookinetes penetrate the midgut epithelium and develop into oocystson the basal lamina. Oocysts fill with developing sporozoites and rupture releases thousands that migrate to salivary glands. Here theyawait the next mosquito blood meal and transmission into another human host.
and Oceania. Residents in endemic regions are at risk of repeated infection, which, over time,can induce immunity to clinical symptoms; however, immunity to malaria is variable, complex,and unstable (reviewed in 82). Although the use of bednets, insecticide spraying, and antimalarialshas reduced the global incidence of malaria since 2000, there is still a large burden of disease inAfrica and other locations, and eradication will be difficult to achieve with current approaches.Currently, there is no vaccine deployed, and drug resistance to the best-known antimalarials,including artemisinin, is emerging (13, 91).
The complex life cycle of malaria parasites involves repeated transmission between mosquitoesand vertebrate hosts (Figure 1). Sporozoite forms injected into the host by a mosquito infecthepatocytes in the liver and develop into exoerythrocytic merozoites, which are released into the
www.annualreviews.org • Remaking the Erythrocyte from the Inside Out 245
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
J-dot: vesicle in theerythrocyte cytoplasmcontaining exportedparasite chaperones
Maurer’s cleft:flattened membranousstructures produced byparasites in theinfected erythrocytefor protein transport
PVM:parasitophorousvacuole membrane
KAHRP:knob-associatedhistidine-rich protein
PfEMP1: Plasmodiumfalciparum erythrocytemembrane protein 1
NPP: newpermeability pathway
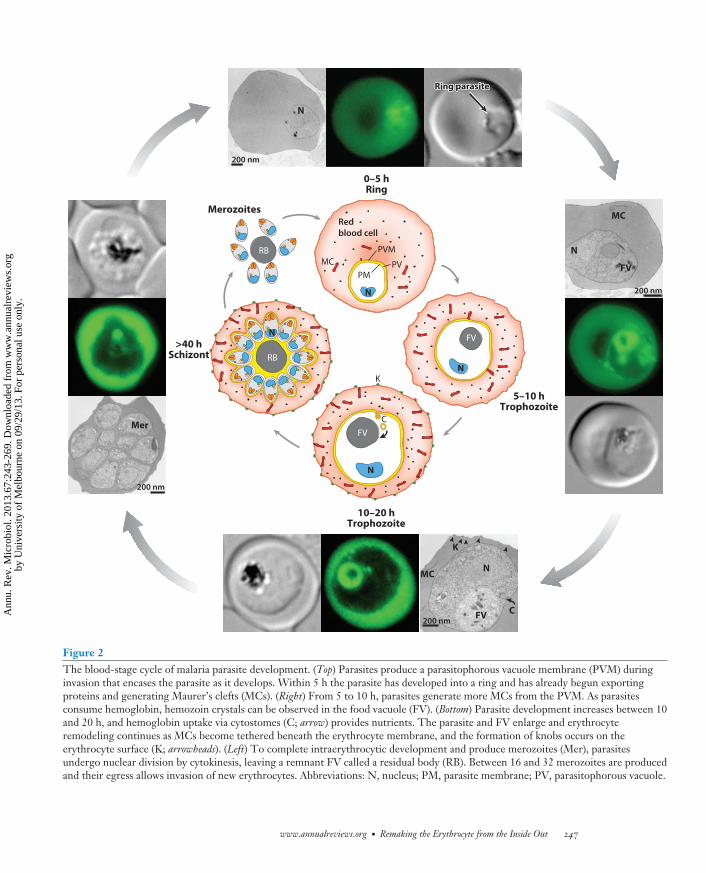
bloodstream, where they invade circulating erythrocytes. The asexual bloodstage of P. falciparumis cyclic and involves differentiation from invading merozoite to ring (0–5 h), to trophozoite (10–40 h), and finally schizont (40–48 h) stages (Figures 1 and 2). During intracellular development,deformability of P. falciparum–infected erythrocytes is dramatically reduced and the cell is endowedwith the ability to adhere to host endothelial cells and other erythrocytes (Figure 3) (reviewedin 2). Cytoadherence confers on P. falciparum–infected erythrocytes the ability to sequester inmajor organs of the host, preventing circulation through the spleen and destruction by splenicmacrophages. Sequestration of P. falciparum–infected erythrocytes can lead to occlusion of bloodflow and infiltration of immune effector cells, which is linked to severe pathogenesis and death(reviewed in 2, 60).
ERYTHROCYTE REMODELING
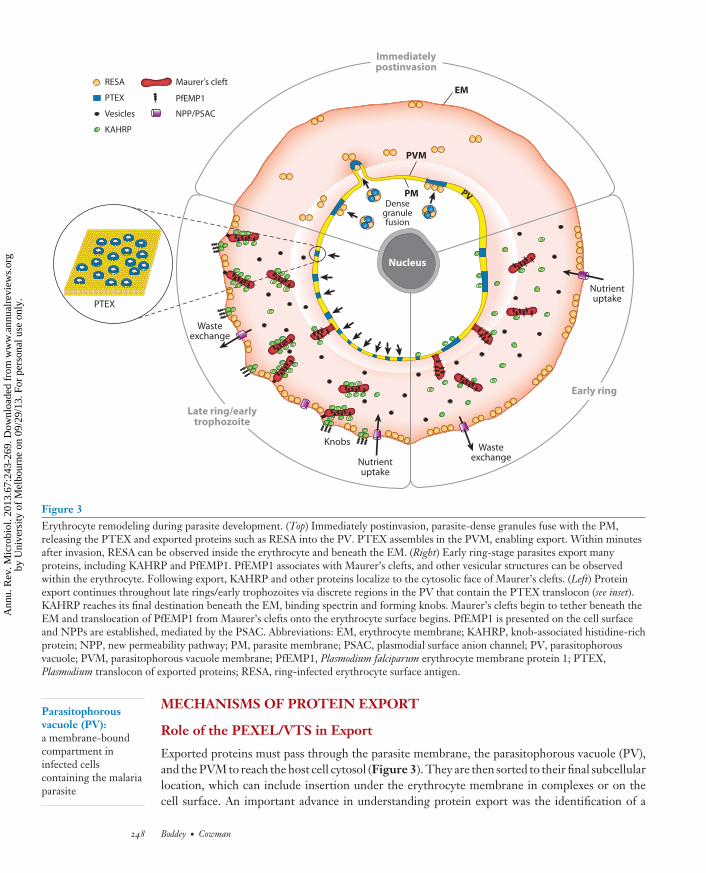
The extraordinary process of erythrocyte remodeling is achieved by large-scale export of pro-teins beyond the confines of the parasite and the encasing parasitophorous vacuole membrane(PVM) (Figure 3) (reviewed in 52). Remodeling is dependent on construction of an extraparasitictrafficking network in the host cell that distributes and sorts exported proteins to their subcel-lular locations. A key feature of the extraparasite protein trafficking network is exomembranousstructures of various sizes (93), including Maurer’s clefts (57), electron-dense vesicles (∼80 nmdiameter) (33, 35, 36), and J-dots (49) (Figures 2 and 3). Maurer’s clefts and J-dots are mobilestructures in the erythrocyte cytoplasm during early stages of parasite development (31). Mau-rer’s clefts are large, flattened membranous structures that bud from the PVM in early ring-stagedevelopment and they migrate toward the erythrocyte membrane, where they become physicallytethered as the parasite differentiates into a trophozoite (34, 86, 98). Maurer’s cleft tethering isimportant for trafficking of protein complexes from within them to the underside and surfaceof the host cell membrane (34, 98, 105). The function of electron-dense vesicles and J-dots isunknown, although they may facilitate trafficking of adherence proteins from Maurer’s clefts ontothe erythrocyte membrane (48, 49).
Cytoadherence of P. falciparum–infected erythrocytes is conferred by expression of a high-molecular-weight protein, P. falciparum erythrocyte membrane protein 1 (PfEMP1) (3, 81, 89).The PfEMP1 protein family displays enormous sequence diversity and host-cell-binding selec-tivity, and these features contribute to the spectrum of disease severity and pathology (reviewedin 2). Sequestration in the capillaries of the brain and placenta during pregnancy contributes sig-nificantly to the morbidity and mortality rates from malaria. PfEMP1 is assembled into electron-dense knobs, consisting primarily of the knob-associated histidine-rich protein (KAHRP), andthese structures are inserted under the erythrocyte membrane (67, 95, 105). The PfEMP1 familyof proteins are encoded by approximately 60 var (variable) genes per haploid genome (25) and theyshow monoallelic expression (79). The ability to switch var gene expression provides a mechanismof antigenic variation important for evasion of host immune responses. The PfEMP1 family pro-teins are targets of acquired immunity during infection and may be candidates for development ofinterventions against malaria. The significant diversity of this protein family in the P. falciparumpopulation is a possible impediment to this goal.
Another key feature of erythrocyte remodeling is the establishment of a new permeability path-way (NPP) that confers a profound increase in permeability of the infected erythrocyte membrane(1, 27, 88). The NPP acts as a conduit for the import of nutrients and exchange of waste products,allowing parasites to condition the intracellular ionic environment. The NPP has also been namedthe plasmodial surface anion channel (65). The proteins involved in each of these processes arefurther described below.
246 Boddey · Cowman
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.
MI67CH12-Cowman ARI 5 August 2013 16:34
PV
PVM
N
Red
blood cell
FV
N
N
FV
RB
PM
K
Merozoites
MC
N
RB
CMer
N
MC
N
FV
0–5 hRing
5–10 hTrophozoite
10–20 hTrophozoite
>40 hSchizont
MC
FV
K
N
C
200 nm
200 nm
200 nm
200 nm
Ring parasiteRing parasiteRing parasite
Figure 2The blood-stage cycle of malaria parasite development. (Top) Parasites produce a parasitophorous vacuole membrane (PVM) duringinvasion that encases the parasite as it develops. Within 5 h the parasite has developed into a ring and has already begun exportingproteins and generating Maurer’s clefts (MCs). (Right) From 5 to 10 h, parasites generate more MCs from the PVM. As parasitesconsume hemoglobin, hemozoin crystals can be observed in the food vacuole (FV). (Bottom) Parasite development increases between 10and 20 h, and hemoglobin uptake via cytostomes (C; arrow) provides nutrients. The parasite and FV enlarge and erythrocyteremodeling continues as MCs become tethered beneath the erythrocyte membrane, and the formation of knobs occurs on theerythrocyte surface (K; arrowheads). (Left) To complete intraerythrocytic development and produce merozoites (Mer), parasitesundergo nuclear division by cytokinesis, leaving a remnant FV called a residual body (RB). Between 16 and 32 merozoites are producedand their egress allows invasion of new erythrocytes. Abbreviations: N, nucleus; PM, parasite membrane; PV, parasitophorous vacuole.
www.annualreviews.org • Remaking the Erythrocyte from the Inside Out 247
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.
MI67CH12-Cowman ARI 5 August 2013 16:34
Immediatelypostinvasion
Late ring/earlytrophozoite
Densegranulefusion
Early ring
RESA
PTEX
Vesicles
KAHRP
Maurer’s cleft
PfEMP1
Wasteexchange
Nutrientuptake
NPP/PSAC
Nucleus
Knobs
PM
PVM
EM
PV
PTEX
Nutrientuptake
Wasteexchange
Figure 3Erythrocyte remodeling during parasite development. (Top) Immediately postinvasion, parasite-dense granules fuse with the PM,releasing the PTEX and exported proteins such as RESA into the PV. PTEX assembles in the PVM, enabling export. Within minutesafter invasion, RESA can be observed inside the erythrocyte and beneath the EM. (Right) Early ring-stage parasites export manyproteins, including KAHRP and PfEMP1. PfEMP1 associates with Maurer’s clefts, and other vesicular structures can be observedwithin the erythrocyte. Following export, KAHRP and other proteins localize to the cytosolic face of Maurer’s clefts. (Left) Proteinexport continues throughout late rings/early trophozoites via discrete regions in the PV that contain the PTEX translocon (see inset).KAHRP reaches its final destination beneath the EM, binding spectrin and forming knobs. Maurer’s clefts begin to tether beneath theEM and translocation of PfEMP1 from Maurer’s clefts onto the erythrocyte surface begins. PfEMP1 is presented on the cell surfaceand NPPs are established, mediated by the PSAC. Abbreviations: EM, erythrocyte membrane; KAHRP, knob-associated histidine-richprotein; NPP, new permeability pathway; PM, parasite membrane; PSAC, plasmodial surface anion channel; PV, parasitophorousvacuole; PVM, parasitophorous vacuole membrane; PfEMP1, Plasmodium falciparum erythrocyte membrane protein 1; PTEX,Plasmodium translocon of exported proteins; RESA, ring-infected erythrocyte surface antigen.
Parasitophorousvacuole (PV):a membrane-boundcompartment ininfected cellscontaining the malariaparasite
MECHANISMS OF PROTEIN EXPORT
Role of the PEXEL/VTS in Export
Exported proteins must pass through the parasite membrane, the parasitophorous vacuole (PV),and the PVM to reach the host cell cytosol (Figure 3). They are then sorted to their final subcellularlocation, which can include insertion under the erythrocyte membrane in complexes or on thecell surface. An important advance in understanding protein export was the identification of a
248 Boddey · Cowman
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
Plasmodium exportelement (PEXEL):an amino acidsignature in the Nterminus of proteinsthat is necessary forexport
Endoplasmicreticulum (ER):a secretory organelle;the first step in thesecretory and exportpathways in malariaparasites
Plasmepsin V:a parasite protease;cleaves the PEXELmotif in proteinsdestined for export
PEXEL-negativeexported protein(PNEP): an exportedmalarial proteinlacking a PEXELmotif
pentameric amino acid sequence (RxLxE/D/Q) required for export, named the Plasmodium exportelement (PEXEL) (56) or vacuolar targeting sequence (VTS) (38), located 20–30 amino acids C-terminal from the signal sequence in proteins destined for export. A similar export motif (RxLR)was identified in the N terminus of effector proteins from oomycete pathogens (102), and despitesuggestions that the mechanism of effector trafficking for Plasmodium spp. and oomycetes wassimilar, subsequent work has shown substantial differences, and it is unlikely they are functionallyequivalent (23, 42, 100, 107).
Plasmepsin V and the Matured N Terminus of PEXEL Proteins
Although it was clear that the PEXEL is required for the export of proteins, its role was notunderstood (38, 56). It has been demonstrated that exported proteins are proteolytically cleavedon the C-terminal side of the conserved leucine (i.e., RxL↓ xE/D/Q) and N-acetylated (i.e.,Ac-xE/Q/D), which showed that the PEXEL motif is a recognition site for a protease that actsin the endoplasmic reticulum (ER), as cleavage was sensitive to brefeldin A (12). Subsequently,the ER resident protease, plasmepsin V, was shown to be responsible for PEXEL cleavage, andthis is the first step in a pathway for export of proteins (7, 76). The PEXEL arginine and leucineresidues are required for recognition and cleavage by plasmepsin V, whereas the fifth amino acidis important in export postcleavage (7, 8, 38, 56). Consequently, the function of the PEXELmotif is twofold: to enable identification of PEXEL proteins for export and cleavage to releasethe N terminus from the ER membrane, and to uncover the export signal Ac-xE/Q/D at the newN terminus, which directs the mature protein to the host cell (7, 76).
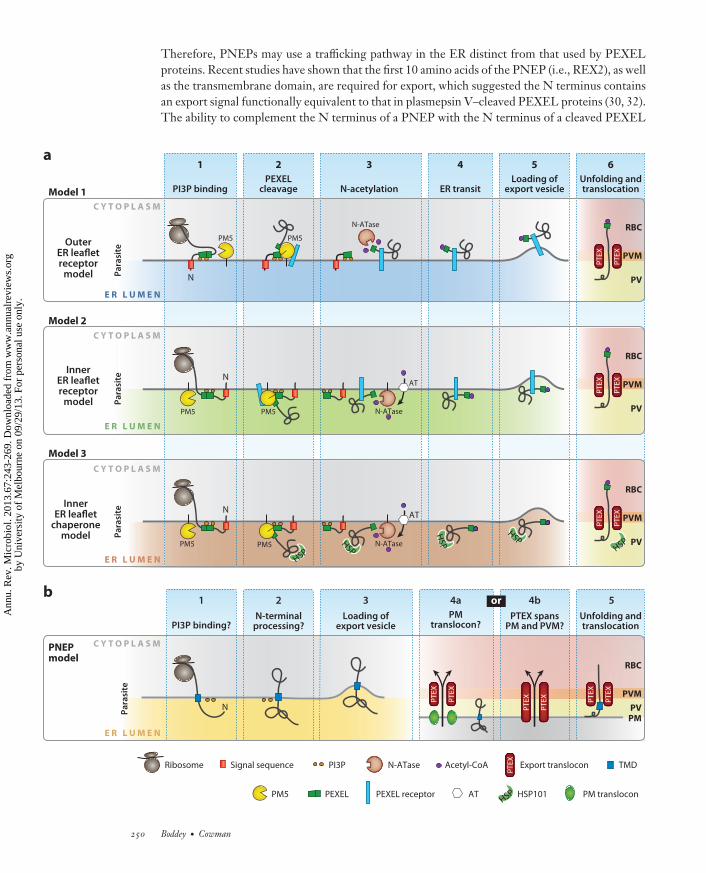
The importance of the processed Ac-xE/Q/D N terminus in trafficking events after plasmepsinV cleavage has been demonstrated by engineering protein reporters cleaved in the ER either bysignal peptidase or by plasmepsin V to produce identical proteins. The reporter protein cleavedby signal peptidase to produce Ac-xQ at the N terminus was secreted to the PV, not exported(7). In contrast, the reporter protein processed by plasmepsin V to produce the same N terminus(Ac-xQ) was efficiently exported. Therefore, plasmepsin V recognition and cleavage reveals asignal that is essential for progression into the export pathway. This finding has been confirmedby a recent study fusing the protein sequence from a mature PEXEL protein (i.e., starting xE)onto a domain from a viral capsid protease that self-cleaves upon folding in the ER to reveal themature PEXEL protein, and this processed protein was also efficiently exported (92). This showedthat plasmepsin V function could be replaced, analogous to PEXEL-negative protein export (seebelow). Nevertheless, it was clear that processing of PEXEL proteins reveals the export signal andthis is essential. The precise mechanism(s) after cleavage that directs mature PEXEL proteins tothe PVM remains unknown; however, like most mechanisms of cargo selection in the ER, it mayinvolve specific proteins interacting directly or indirectly with plasmepsin V, be it a chaperone orcargo receptor, that escort the protein after PEXEL processing (Figure 4).
How Are PNEPs Exported?
Plasmodium spp. express a second class of exported proteins, called PEXEL-negative exported pro-teins (PNEPs) (reviewed in 83), that do not contain an N-terminal hydrophobic signal sequence,a PEXEL motif, or other conserved export sequences. PNEPs are a relatively small group inP. falciparum and include proteins such as ring-exported proteins 1 and 2 (REX1, REX2) (21, 32)and skeleton binding protein 1 (SBP1) (see Table 1). The major virulence protein PfEMP1 isalso defined as a PNEP as this family has no hydrophobic signal sequence and does not possessa PEXEL at the N terminus; PfEMP1 and other PNEPs cannot be cleaved by plasmepsin V (6).
www.annualreviews.org • Remaking the Erythrocyte from the Inside Out 249
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.
MI67CH12-Cowman ARI 5 August 2013 16:34
Therefore, PNEPs may use a trafficking pathway in the ER distinct from that used by PEXELproteins. Recent studies have shown that the first 10 amino acids of the PNEP (i.e., REX2), as wellas the transmembrane domain, are required for export, which suggested the N terminus containsan export signal functionally equivalent to that in plasmepsin V–cleaved PEXEL proteins (30, 32).The ability to complement the N terminus of a PNEP with the N terminus of a cleaved PEXEL
C Y T O P L A S M
E R L U M E N
C Y T O P L A S M
E R L U M E N
C Y T O P L A S M
E R L U M E N
C Y T O P L A S M
E R L U M E N
PT
EX
PT
EX
PT
EX
PT
EX
PT
EX
PT
EX
PT
EX
PT
EX
PT
EX
PT
EX
PT
EX
PT
EX
PT
EX
6
Unfolding andtranslocation
N
1 2 3 4 5
Model 1
Model 2
N
Model 3
N
RBC
PV
PVMH
SP
HSP
N-ATase
PM5
PM5 N-ATase
RBC
PV
PVM
RBC
PV
PVM
HSPPM5
AT
Pa
rasi
teP
ara
site
Pa
rasi
te
PM5
PM5
1 2 3
PNEPmodel
N
N-terminalprocessing?
Loading ofexport vesicle
RBC
PV
PVM
5
Unfolding andtranslocationPI3P binding?
PTEX spansPM and PVM?
4b4a
PMtranslocon?
PM
Ribosome Signal sequence
PEXEL
PI3P
PEXEL receptor
Acetyl-CoA
HSP HSP101
Export translocon
PM5 AT
N-ATase TMD
PM translocon
PM5
HSP
HSP
OuterER leafletreceptor
model
InnerER leafletreceptor
model
PI3P bindingPEXEL
cleavage N-acetylation ER transitLoading of
export vesicle
InnerER leaflet
chaperonemodel
Pa
rasi
te
N-ATase
AT
a
bor
250 Boddey · Cowman
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
protein strongly supports this hypothesis (30). Trafficking of PNEPs is also brefeldin A sensitive,suggesting that it involves vesicular trafficking from the ER to the parasite membrane (30). Thisindicates that although there are distinct differences between trafficking of PNEPs and traffickingof PEXEL proteins, there may be some overlap in their export mechanism. While we do notunderstand the molecular basis for PEXEL and PNEP export, it is likely that the differences be-tween them may be important with respect to timing of expression, subcellular localization, andtopology of the proteins in the erythrocyte.
Another difference between PEXEL proteins and PNEPs is acetylation of the N terminus;however, the importance of this feature in export is uncertain. Whereas virtually all exportedPEXEL proteins are N-acetylated after cleavage (8, 12), the PNEP REX2 is not N-acetylated afterN-terminal processing in parasites (32). N-acetylation is common in eukaryotes and generallyoccurs in the cytoplasm. In Plasmodium spp. this would require PEXEL proteins to face theoutside of the ER membrane rather than the ER lumen during transport, and this has beensuggested as a potential route for PEXEL protein export (51, 75) (Figure 4). However, eukaryoticcells possess ER acetylation machinery (41, 45, 104). BLAST searches of Plasmodium genomesequences (25) using this human machinery readily identified homologues of these components(PF3D7_1437000, an N-acetyltransferase homologue, and PF3D7_1036800, a putative acetyl-CoA transporter) in all Plasmodium species. Although the role of N-acetylation in export is unclear,it may enhance the avidity of protein-protein interactions between the plasmepsin V–processedN terminus of exported proteins and potential PEXEL receptors or chaperones (80) (Figure 4).
How Is PfEMP1 Exported?
Export of the PNEP protein PfEMP1 is a complex and multistep process as it is large with avariable number of folded domains (Figure 5) and must traverse the parasite membrane, PVM,Maurer’s cleft membrane, and erythrocyte membrane to display the adhesive ectodomain on theoutside surface. The first step in export requires the transmembrane domain for ER entry, as is the
←−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−Figure 4Models of PEXEL and PNEP export. (a) Three models of PEXEL protein export. In Model 1, PEXEL proteins are targeted to the ERby their signal sequence anchored in the ER membrane. The PEXEL binds PI3P present on the cytoplasmic ER leaflet ( gray), possiblytargeting the protein toward PM5 (Step 1). In Step 2, PEXEL processing occurs and the matured protein is selected by an ER receptor.The new N terminus of the PEXEL protein is acetylated by a cytoplasmic N-ATase using cytoplasmic acetyl-CoA (Step 3). Cargoproteins transit the ER membrane (Step 4) and are loaded onto vesicle membranes via an ER receptor (Step 5). Vesicles dock at regionsof the PV containing the PTEX translocon in the PVM. Cargo proteins are released, unfolded, and translocated across the PVM intothe RBC (Step 6). In Model 2, PEXEL proteins are targeted to the ER, where PI3P binding, PEXEL cleavage, and cargo recruitmentoccur inside the ER lumen ( green) (Steps 1 and 2). However, Step 3 involves N-acetylation of the new N terminus in the ER by anER-resident N-ATase that utilizes acetyl-CoA imported into the ER by an AT. Cargo proteins transit the ER and traffic to the PV viavesicles (Steps 4 and 5). Step 6 occurs as in Model 1. In Model 3, Steps 1 to 6 occur as in Model 2, except PEXEL-cleaved cargo arerecruited by Hsp101 (Hsp) after cleavage by PM5 (Step 2) and escort cargo to PTEX (Step 6). (b) Model of PNEP export. In Step 1,proteins are targeted to the ER by a TMD. PNEP-PI3P binding at the parasite ER is unknown. In Step 2, the N terminus of PNEPsmay be processed (e.g., REX2 is cleaved by an unknown protease, not PM5). Also of note, REX2 was not N-acetylated after processing(32). In Step 3, proteins transit the ER and are loaded into export vesicles for transport to the PM. In Step 4, protein unfolding isrequired at the PM, suggesting a translocon mechanism (30). This could be a unique PM translocon (Step 4a) or indicate that PTEXspans both the PM and the PVM (Step 4b). The protein is translocated into the RBC (Step 5) and associates with Maurer’s clefts.Abbreviations: AT, acetyl-CoA transporter; ER, endoplasmic reticulum; N-ATase, N-acetyltransferase; PEXEL, Plasmodium exportelement; PI3P, phosphatidylinositol-3-phosphate; PM, parasite membrane; PM5, plasmepsin V; PNEP, PEXEL-negative exportedprotein; PTEX, Plasmodium translocon of exported proteins; PV, parasitophorous vacuole; PVM, parasitophorous vacuole membrane;REX, ring-exported protein 2; RBC, red blood cell; TMD, transmembrane domain.
www.annualreviews.org • Remaking the Erythrocyte from the Inside Out 251
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.
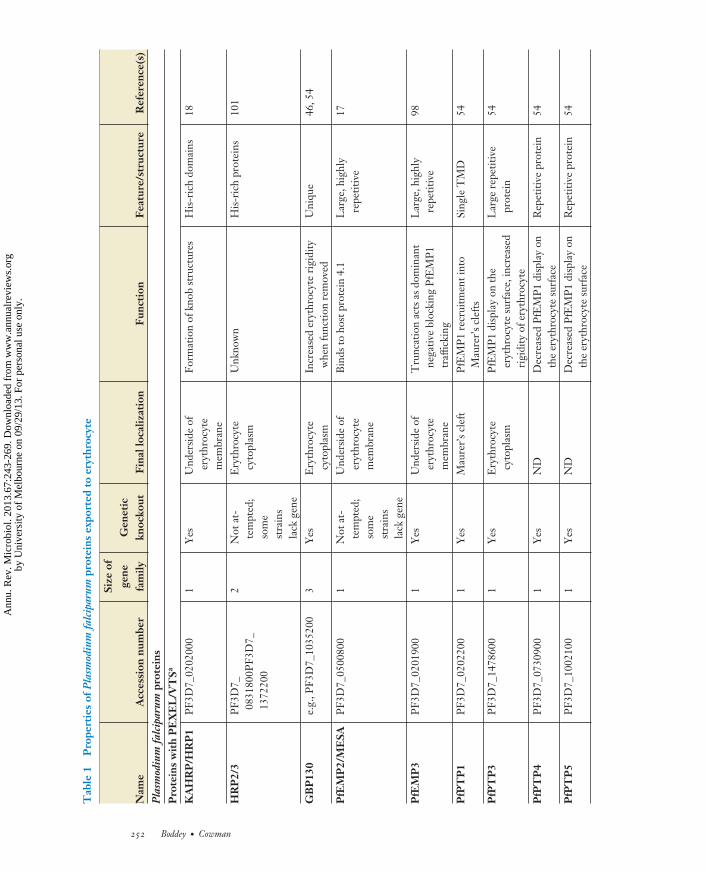
MI67CH12-Cowman ARI 5 August 2013 16:34
Tab
le1
Pro
pert
ies
ofPl
asm
odiu
mfa
lcip
arum
prot
eins
expo
rted
toer
ythr
ocyt
e
Nam
eA
cces
sion
num
ber
Size
ofge
nefa
mily
Gen
etic
knoc
kout
Fina
lloc
aliz
atio
nFu
ncti
onFe
atur
e/st
ruct
ure
Ref
eren
ce(s
)Pl
asm
odiu
mfa
lcip
arum
prot
eins
Pro
tein
sw
ith
PE
XE
L/V
TSa
KA
HR
P/H
RP
1P
F3D
7_02
0200
01
Yes
Und
ersi
deof
eryt
hroc
yte
mem
bran
e
Form
atio
nof
knob
stru
ctur
esH
is-r
ich
dom
ains
18
HR
P2/
3P
F3D
7_08
3180
0PF3
D7_
1372
200
2N
otat
-te
mpt
ed;
som
est
rain
sla
ckge
ne
Ery
thro
cyte
cyto
plas
mU
nkno
wn
His
-ric
hpr
otei
ns10
1
GB
P13
0e.
g.,P
F3D
7_10
3520
03
Yes
Ery
thro
cyte
cyto
plas
mIn
crea
sed
eryt
hroc
yte
rigi
dity
whe
nfu
nctio
nre
mov
edU
niqu
e46
,54
PfE
MP
2/M
ESA
PF3
D7_
0500
800
1N
otat
-te
mpt
ed;
som
est
rain
sla
ckge
ne
Und
ersi
deof
eryt
hroc
yte
mem
bran
e
Bin
dsto
host
prot
ein
4.1
Lar
ge,h
ighl
yre
petit
ive
17
PfE
MP
3P
F3D
7_02
0190
01
Yes
Und
ersi
deof
eryt
hroc
yte
mem
bran
e
Tru
ncat
ion
acts
asdo
min
ant
nega
tive
bloc
king
PfE
MP
1tr
affic
king
Lar
ge,h
ighl
yre
petit
ive
98
PfP
TP
1P
F3D
7_02
0220
01
Yes
Mau
rer’
scl
eft
PfE
MP
1re
crui
tmen
tint
oM
aure
r’s
clef
tsSi
ngle
TM
D54
PfP
TP
3P
F3D
7_14
7860
01
Yes
Ery
thro
cyte
cyto
plas
mP
fEM
P1
disp
lay
onth
eer
ythr
ocyt
esu
rfac
e,in
crea
sed
rigi
dity
ofer
ythr
ocyt
e
Lar
gere
petit
ive
prot
ein
54
PfP
TP
4P
F3D
7_07
3090
01
Yes
ND
Dec
reas
edP
fEM
P1
disp
lay
onth
eer
ythr
ocyt
esu
rfac
eR
epet
itive
prot
ein
54
PfP
TP
5P
F3D
7_10
0210
01
Yes
ND
Dec
reas
edP
fEM
P1
disp
lay
onth
eer
ythr
ocyt
esu
rfac
eR
epet
itive
prot
ein
54
252 Boddey · Cowman
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.
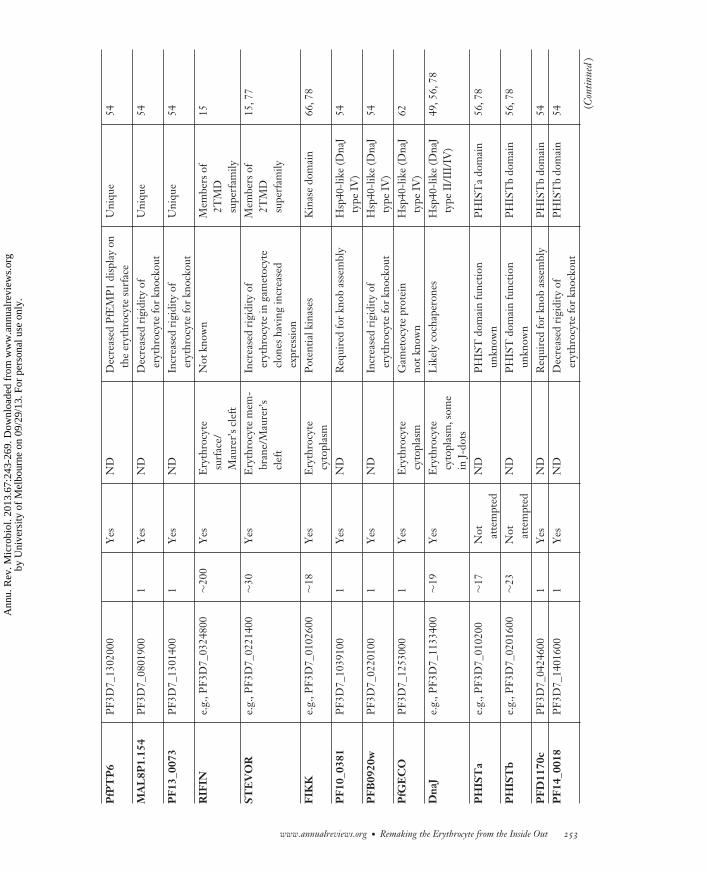
MI67CH12-Cowman ARI 5 August 2013 16:34
PfP
TP
6P
F3D
7_13
0200
0Y
esN
DD
ecre
ased
PfE
MP
1di
spla
yon
the
eryt
hroc
yte
surf
ace
Uni
que
54
MA
L8P
1.15
4P
F3D
7_08
0190
01
Yes
ND
Dec
reas
edri
gidi
tyof
eryt
hroc
yte
for
knoc
kout
Uni
que
54
PF1
3_00
73P
F3D
7_13
0140
01
Yes
ND
Incr
ease
dri
gidi
tyof
eryt
hroc
yte
for
knoc
kout
Uni
que
54
RIF
INe.
g.,P
F3D
7_03
2480
0∼2
00Y
esE
ryth
rocy
tesu
rfac
e/M
aure
r’s
clef
t
Not
know
nM
embe
rsof
2TM
Dsu
perf
amily
15
STE
VO
Re.
g.,P
F3D
7_02
2140
0∼3
0Y
esE
ryth
rocy
tem
em-
bran
e/M
aure
r’s
clef
t
Incr
ease
dri
gidi
tyof
eryt
hroc
yte
inga
met
ocyt
ecl
ones
havi
ngin
crea
sed
expr
essi
on
Mem
bers
of2T
MD
supe
rfam
ily
15,7
7
FIK
Ke.
g.,P
F3D
7_01
0260
0∼1
8Y
esE
ryth
rocy
tecy
topl
asm
Pot
entia
lkin
ases
Kin
ase
dom
ain
66,7
8
PF1
0_03
81P
F3D
7_10
3910
01
Yes
ND
Req
uire
dfo
rkn
obas
sem
bly
Hsp
40-l
ike
(Dna
Jty
peIV
)54
PFB
0920
wP
F3D
7_02
2010
01
Yes
ND
Incr
ease
dri
gidi
tyof
eryt
hroc
yte
for
knoc
kout
Hsp
40-l
ike
(Dna
Jty
peIV
)54
PfG
EC
OP
F3D
7_12
5300
01
Yes
Ery
thro
cyte
cyto
plas
mG
amet
ocyt
epr
otei
nno
tkno
wn
Hsp
40-l
ike
(Dna
Jty
peIV
)62
Dna
Je.
g.,P
F3D
7_11
3340
0∼1
9Y
esE
ryth
rocy
tecy
topl
asm
,som
ein
J-do
ts
Lik
ely
coch
aper
ones
Hsp
40-l
ike
(Dna
Jty
peII
/III
/IV
)49
,56,
78
PH
IST
ae.
g.,P
F3D
7_01
0200
∼17
Not atte
mpt
edN
DP
HIS
Tdo
mai
nfu
nctio
nun
know
nP
HIS
Ta
dom
ain
56,7
8
PH
IST
be.
g.,P
F3D
7_02
0160
0∼2
3N
ot atte
mpt
edN
DP
HIS
Tdo
mai
nfu
nctio
nun
know
nP
HIS
Tb
dom
ain
56,7
8
PFD
1170
cP
F3D
7_04
2460
01
Yes
ND
Req
uire
dfo
rkn
obas
sem
bly
PH
IST
bdo
mai
n54
PF1
4_00
18P
F3D
7_14
0160
01
Yes
ND
Dec
reas
edri
gidi
tyof
eryt
hroc
yte
for
knoc
kout
PH
IST
bdo
mai
n54
(Con
tinue
d)
www.annualreviews.org • Remaking the Erythrocyte from the Inside Out 253
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.
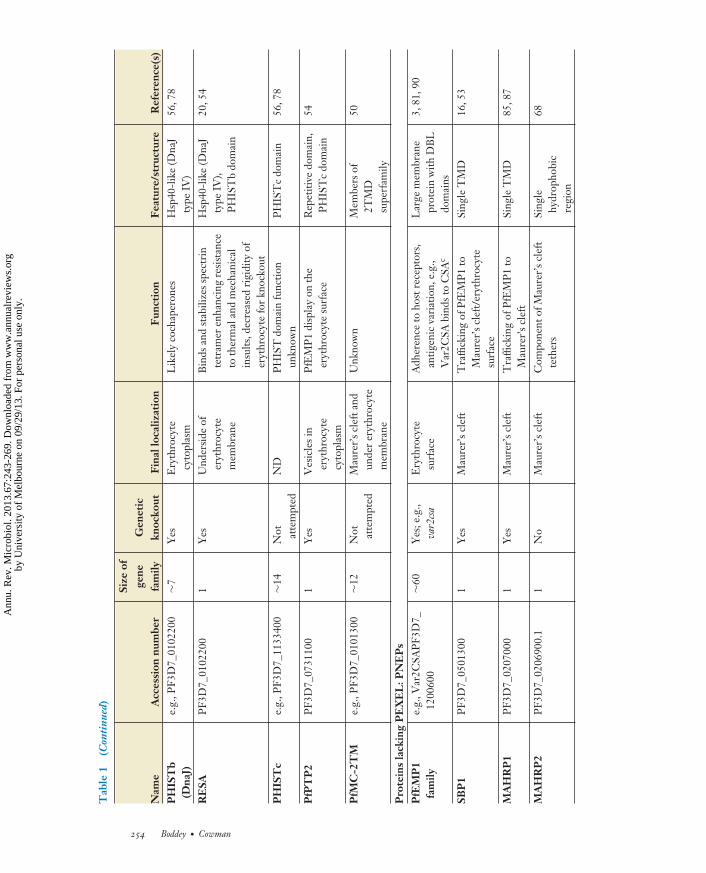
MI67CH12-Cowman ARI 5 August 2013 16:34
Tab
le1
(Con
tinu
ed)
Nam
eA
cces
sion
num
ber
Size
ofge
nefa
mily
Gen
etic
knoc
kout
Fina
lloc
aliz
atio
nFu
ncti
onFe
atur
e/st
ruct
ure
Ref
eren
ce(s
)P
HIS
Tb
(Dna
J)e.
g.,P
F3D
7_01
0220
0∼7
Yes
Ery
thro
cyte
cyto
plas
mL
ikel
yco
chap
eron
esH
sp40
-lik
e(D
naJ
type
IV)
56,7
8
RE
SAP
F3D
7_01
0220
01
Yes
Und
ersi
deof
eryt
hroc
yte
mem
bran
e
Bin
dsan
dst
abili
zes
spec
trin
tetr
amer
enha
ncin
gre
sist
ance
toth
erm
alan
dm
echa
nica
lin
sults
,dec
reas
edri
gidi
tyof
eryt
hroc
yte
for
knoc
kout
Hsp
40-l
ike
(Dna
Jty
peIV
),P
HIS
Tb
dom
ain
20,5
4
PH
IST
ce.
g.,P
F3D
7_11
3340
0∼1
4N
ot atte
mpt
edN
DP
HIS
Tdo
mai
nfu
nctio
nun
know
nP
HIS
Tc
dom
ain
56,7
8
PfP
TP
2P
F3D
7_07
3110
01
Yes
Ves
icle
sin
eryt
hroc
yte
cyto
plas
m
PfE
MP
1di
spla
yon
the
eryt
hroc
yte
surf
ace
Rep
etiti
vedo
mai
n,P
HIS
Tc
dom
ain
54
PfM
C-2
TM
e.g.
,PF3
D7_
0101
300
∼12
Not atte
mpt
edM
aure
r’s
clef
tand
unde
rer
ythr
ocyt
em
embr
ane
Unk
now
nM
embe
rsof
2TM
Dsu
perf
amily
50
Pro
tein
sla
ckin
gP
EX
EL
:PN
EP
sP
fEM
P1
fam
ilye.
g.,V
ar2C
SAP
F3D
7_12
0060
0∼6
0Y
es;e
.g.,
var2
csaE
ryth
rocy
tesu
rfac
eA
dher
ence
toho
stre
cept
ors,
antig
enic
vari
atio
n,e.
g.,
Var
2CSA
bind
sto
CSA
c
Lar
gem
embr
ane
prot
ein
with
DB
Ldo
mai
ns
3,81
,90
SBP
1P
F3D
7_05
0130
01
Yes
Mau
rer’
scl
eft
Tra
ffick
ing
ofP
fEM
P1
toM
aure
r’s
clef
t/er
ythr
ocyt
esu
rfac
e
Sing
leT
MD
16,5
3
MA
HR
P1
PF3
D7_
0207
000
1Y
esM
aure
r’s
clef
tT
raffi
ckin
gof
PfE
MP
1to
Mau
rer’
scl
eft
Sing
leT
MD
85,8
7
MA
HR
P2
PF3
D7_
0206
900.
11
No
Mau
rer’
scl
eft
Com
pone
ntof
Mau
rer’
scl
eft
teth
ers
Sing
lehy
drop
hobi
cre
gion
68
254 Boddey · Cowman
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.
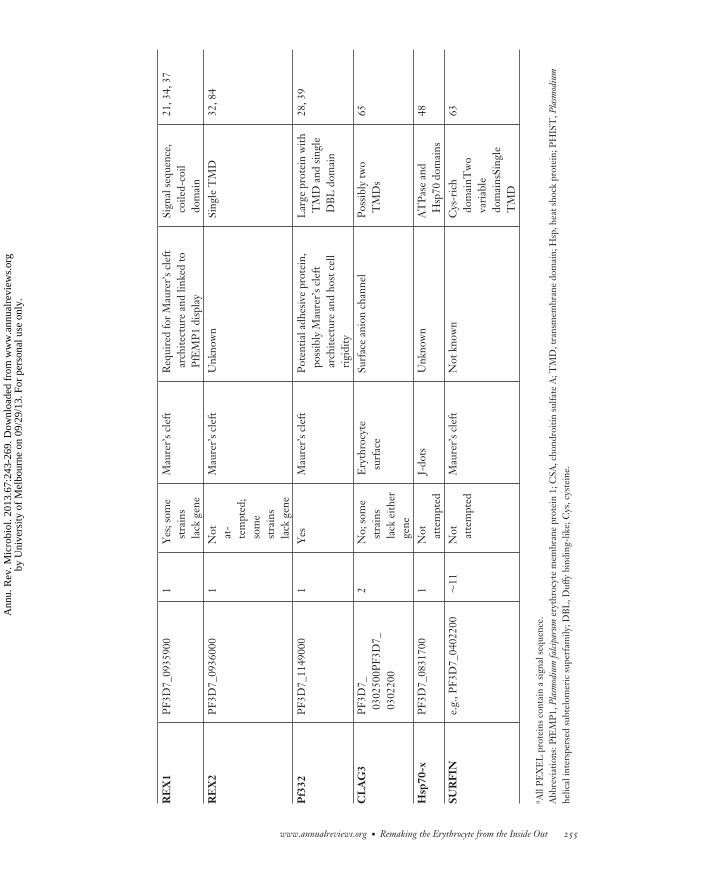
MI67CH12-Cowman ARI 5 August 2013 16:34
RE
X1
PF3
D7_
0935
900
1Y
es;s
ome
stra
ins
lack
gene
Mau
rer’
scl
eft
Req
uire
dfo
rM
aure
r’s
clef
tar
chite
ctur
ean
dlin
ked
toP
fEM
P1
disp
lay
Sign
alse
quen
ce,
coile
d-co
ildo
mai
n
21,3
4,37
RE
X2
PF3
D7_
0936
000
1N
ot at-
tem
pted
;so
me
stra
ins
lack
gene
Mau
rer’
scl
eft
Unk
now
nSi
ngle
TM
D32
,84
Pf3
32P
F3D
7_11
4900
01
Yes
Mau
rer’
scl
eft
Pot
entia
ladh
esiv
epr
otei
n,po
ssib
lyM
aure
r’s
clef
tar
chite
ctur
ean
dho
stce
llri
gidi
ty
Lar
gepr
otei
nw
ithT
MD
and
sing
leD
BL
dom
ain
28,3
9
CL
AG
3P
F3D
7_03
0250
0PF3
D7_
0302
200
2N
o;so
me
stra
ins
lack
eith
erge
ne
Ery
thro
cyte
surf
ace
Surf
ace
anio
nch
anne
lP
ossi
bly
two
TM
Ds
65
Hsp
70-x
PF3
D7_
0831
700
1N
ot atte
mpt
edJ-
dots
Unk
now
nA
TP
ase
and
Hsp
70do
mai
ns48
SUR
FIN
e.g.
,PF3
D7_
0402
200
∼11
Not atte
mpt
edM
aure
r’s
clef
tN
otkn
own
Cys
-ric
hdo
mai
nTw
ova
riab
ledo
mai
nsSi
ngle
TM
D
63
a All
PE
XE
Lpr
otei
nsco
ntai
na
sign
alse
quen
ce.
Abb
revi
atio
ns:P
fEM
P1,
Plas
mod
ium
falci
paru
mer
ythr
ocyt
em
embr
ane
prot
ein
1;C
SA,c
hond
roiti
nsu
lfate
A;T
MD
,tra
nsm
embr
ane
dom
ain;
Hsp
,hea
tsho
ckpr
otei
n;P
HIS
T,P
lasm
odiu
mhe
lical
inte
rspe
rsed
subt
elom
eric
supe
rfam
ily;D
BL
,Duf
fybi
ndin
g-lik
e;C
ys,c
yste
ine.
www.annualreviews.org • Remaking the Erythrocyte from the Inside Out 255
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.
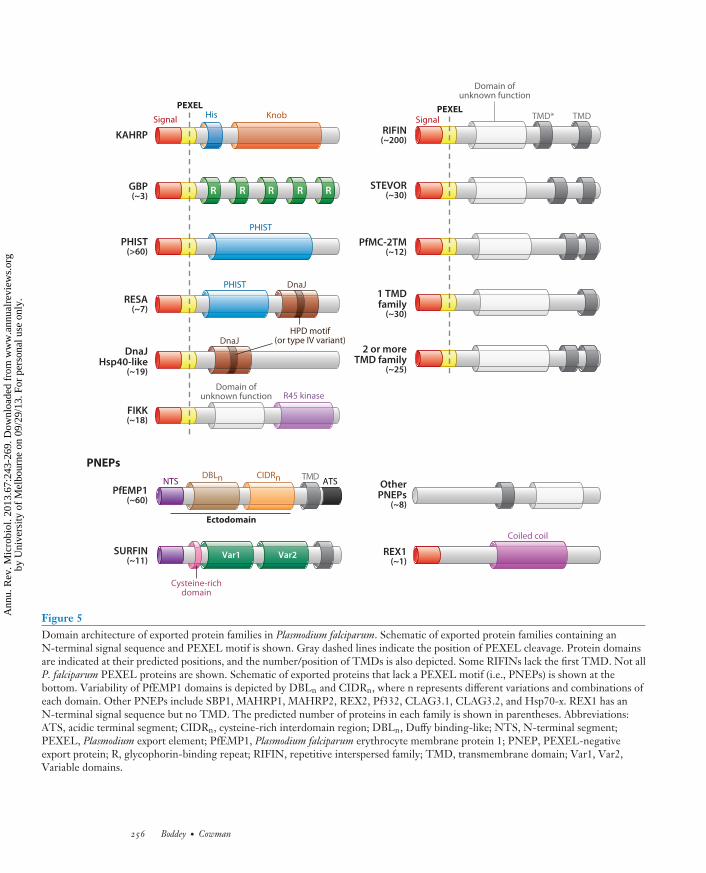
MI67CH12-Cowman ARI 5 August 2013 16:34
PHIST(>60)
GBP(~3)
R R R R R
KAHRP RIFIN(~200)
STEVOR(~30)
PfMC-2TM(~12)
PEXEL PEXEL
FIKK(~18)
PfEMP1(~60)
Ectodomain
SURFIN(~11)
PNEPs
RESA(~7)
DnaJHsp40-like
(~19)
1 TMDfamily
(~30)
2 or moreTMD family
(~25)
OtherPNEPs
(~8)
REX1(~1)
HPD motif(or type IV variant)
SignalSignalHis Knob
PHIST
PHIST DnaJ
DnaJ
Domain ofunknown function R45 kinase
TMD
TMD* TMD
NTSDBLn CIDRn ATS
Cysteine-richdomain
Var1 Var2
Coiled coil
Domain ofunknown function
Figure 5Domain architecture of exported protein families in Plasmodium falciparum. Schematic of exported protein families containing anN-terminal signal sequence and PEXEL motif is shown. Gray dashed lines indicate the position of PEXEL cleavage. Protein domainsare indicated at their predicted positions, and the number/position of TMDs is also depicted. Some RIFINs lack the first TMD. Not allP. falciparum PEXEL proteins are shown. Schematic of exported proteins that lack a PEXEL motif (i.e., PNEPs) is shown at thebottom. Variability of PfEMP1 domains is depicted by DBLn and CIDRn, where n represents different variations and combinations ofeach domain. Other PNEPs include SBP1, MAHRP1, MAHRP2, REX2, Pf332, CLAG3.1, CLAG3.2, and Hsp70-x. REX1 has anN-terminal signal sequence but no TMD. The predicted number of proteins in each family is shown in parentheses. Abbreviations:ATS, acidic terminal segment; CIDRn, cysteine-rich interdomain region; DBLn, Duffy binding-like; NTS, N-terminal segment;PEXEL, Plasmodium export element; PfEMP1, Plasmodium falciparum erythrocyte membrane protein 1; PNEP, PEXEL-negativeexport protein; R, glycophorin-binding repeat; RIFIN, repetitive interspersed family; TMD, transmembrane domain; Var1, Var2,Variable domains.
256 Boddey · Cowman
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
case for other PNEPs, confirming important trafficking information is encoded in this region (44).For export and display of PfEMP1 on the host cell surface, the semiconserved head region (NTS,DBL1, and CIDR domains) and the transmembrane domain and cytoplasmic tail are required (59).The N terminus of PfEMP1 is also predicted to be myristoylated in the parasite cytoplasm, whichmay facilitate protein trafficking as well as anchoring the protein in the erythrocyte membrane(72).
PfEMP1 expression starts 4 h postinvasion and does not appear on the surface of theP. falciparum–infected erythrocyte until 16 h; however, it is trafficked to and stored in Maurer’sclefts (47, 69, 105). Whether PfEMP1 is translocated across the PVM using the PTEX translocon(see below) is unknown (19). It has been proposed that PfEMP1 is incorporated into Maurer’sclefts as they form at the PVM; however, it was recently shown that Maurer’s clefts are gener-ated early after invasion, before PfEMP1 would reach the PVM, and that the number of cleftsdoes not increase over time (see Timing of Events in Export, below) (31). This finding suggestsPfEMP1 may be trafficked to Maurer’s clefts after they have formed, involving transit as a solublecomplex (44, 69). The next step in transport of PfEMP1 is transfer from Maurer’s clefts onto theerythrocyte surface, and this process is poorly defined (44, 59).
Role of Phosphatidylinositol-3-Phosphate
Another mechanism for export has been proposed, involving binding of the PEXEL tophosphatidylinositol-3-phosphate (PI3P) on the lumenal side of the ER membrane, prior to plas-mepsin V cleavage (5) (Figure 4). PI3P binding of PEXEL proteins upon ER targeting couldrecruit cargo to export competent or plasmepsin V–rich zones in the ER, but binding wouldattach the PEXEL to the membrane, away from the plasmepsin V active site. The PI3P hy-pothesis originated from studies on Phytophthora infestans, a pathogenic oomycete that expressesvirulence effectors with an N-terminal RxLR motif required for host cell entry (42). However,there is considerable uncertainty of a role for PI3P in the export of effector proteins in oomycetes(99, 107). The PEXEL motif of Plasmodium spp. may be functionally equivalent to the RxLRmotif in oomycetes, and this was shown by complementation of each signal for export (4, 29).Although this suggested complementarity between the RxLR from oomycetes and PEXEL fromPlasmodium spp., several other laboratories have so far not been able to demonstrate export ofoomycete proteins in P. falciparum (30).
In vitro experiments have shown that PI3P binds to the PEXEL and the key amino acidinvolved was the conserved arginine, providing evidence that PI3P may be involved in export(5). This hypothesis suggests that proteins destined for export bind to PI3P in the ER lumenand are trafficked via a vesicular pathway and released from the membrane by plasmepsin Vcleavage (Figure 4). It is unclear how PI3P binding contributes to export as plasmepsin V veryrapidly cleaves the PEXEL motif in the parasite ER, which would release the proteins from PI3Pbinding. In addition, efficient export was observed when the normal N-terminal export signal(i.e., xE) was revealed by cleavage of a viral capsid protease (92). Because these constructs lack thePEXEL arginine and leucine (required for plasmepsin V cleavage), they would not bind PI3P, andthese results are therefore inconsistent with ER lipid binding as the export mechanism. A potentialrole for PI3P binding in the export of PfEMP1 and other PNEPs has also been suggested and arecombinant PfEMP1 N terminus reportedly binds this lipid via a conserved lysine residue (i.e.,KxLxD). The role of lysine binding to PI3P in export is unclear as substitution of the PEXELarginine for lysine in KAHRP (i.e., KxLxQ) completely blocked export; the chimeric motif couldnot be cleaved by plasmepsin V and the protein accumulated in the ER (6). Other PNEPs have
www.annualreviews.org • Remaking the Erythrocyte from the Inside Out 257
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
Plasmodiumtranslocon ofexported proteins(PTEX translocon):a parasite-derivedcomplex that enablesexport across the PVM
not been shown to bind PI3P, and at this stage there is no convincing evidence that PI3P plays arole in the export of PfEMP1, or other PNEPs, in infected erythrocytes.
In P. falciparum PI3P was localized to the lumenal side of the ER membrane by transgenicexpression of FYVE domains that bind specifically to this lipid, which was important for proteinexport (5). The presence of PI3P in this compartment of the ER is unusual in eukaryotic cellsand only occurs under certain circumstances, such as transient recruitment of PI3-kinase to thelumenal side of the ER for autophagosome formation, where PI3P is absent. There is no obviousPI3-kinase residing within the ER or secretory pathway, and autophagosomal-like structures havenot been observed in P. falciparum blood stages (22, 43). Moreover, ER recruitment of PI3-kinase requires a three-member complex, and orthologues cannot be identified in the genomesof different Plasmodium spp. (43). Thus, the characterized machinery that recruits PI3-kinase tothe ER may not be present in protozoa. If Plasmodium spp. produce ER-resident PI3P in a steadystate, as would be required for continuous protein export, it would be the first eukaryotic organismidentified to do so.
PROTEIN EXPORT MACHINERY
For export, PEXEL proteins and PNEPs must traverse the parasite membrane and PVM, and atranslocation machine is hypothesized to be responsible for this process (56). Proteomic analysis ofdetergent-resistant membranes identified a complex with properties expected of a translocon; it wasnamed Plasmodium translocon of exported proteins (PTEX) (19). PTEX is a >1.2-MDa multimericcomplex (11) composed of at least five members: exported protein 2 (EXP2) (40), thioredoxin 2(Trx2) (10), heat shock protein 101 (Hsp101), PTEX150, and PTEX88. PTEX components areconserved in Plasmodium spp., as expected for a protein export translocon. In addition, they areexpressed in dense granules of merozoites and deposited into the PVM after invasion when proteinexport commences (11). Furthermore, PTEX specifically binds PEXEL proteins, consistent withtheir translocation by this protein machine; however, a direct interaction between PTEX andPNEPs has not been demonstrated (19). The properties of PTEX in Plasmodium spp. is consistentwith a potential role in export across the PVM.
Biochemical analysis and homology with proteins of known function have suggested possi-ble roles for PTEX components. Hsp101 is a ClpA/B-like AAA+ ATPase of a type commonlyassociated with translocons and is expected to function in protein unfolding for translocationthrough the translocon pore (19) (Figure 6). EXP2 forms ∼600-kDa homo-oligomers and ismore tightly membrane associated than other components, consistent with its hypothesized roleas the membrane-spanning pore (11). Solubility studies show the PTEX complex is arranged inthe order EXP2-PTEX150-Hsp101, with EXP2 inserting into the PVM (11). Presumably, Trx2is involved in chemical reduction of cysteine-linked proteins for unfolding by Hsp101 beforethreading through the channel. PTEX150 and PTEX88 have no homology to other proteins andtheir function is unknown.
There is circumstantial evidence that PTEX is the export translocon in Plasmodium spp.; how-ever, lack of a good system for conditional gene expression has made it difficult to provide directevidence. Studies using three-dimensional structured illumination microscopy have provided di-rect evidence that PTEX is indeed the export translocon (74). This finding was based on previousstudies with dihydrofolate reductase fused to a signal sequence and PEXEL, which demonstratedthat unfolding is required for export, as antifolate drugs binding dihydrofolate reductase preventprotein unfolding and block export (26). Blocked translocation intermediates spatially associatewith PTEX clusters, providing direct evidence that export of PEXEL proteins occurs in PTEX-rich domains of the PVM (74).
258 Boddey · Cowman
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
PVMPVM
PMPM
PTEX150
Hsp101PTEX88
Trx2
Protein for exportis unfolded
Parasite cytoplasm
Erythrocytecytoplasm
Exported proteinis refolded
ATP
ADP
Vesicle
PV
PV
PVM
PM
EXP2
Figure 6Model of translocation by the Plasmodium translocon of exported proteins (PTEX). Proteins destined forexport arrive at the parasitophorous vacuole (PV) by fusion of an export vesicle with the parasite membrane(PM). The protein is recognized in the vacuolar space and unfolded perhaps by Hsp101, which contains anAAA+ ATPase domain. The protein is fed through the Hsp101-PTEX150 complex to EXP2, which is theputative pore in the parasitophorous vacuole membrane (PVM). Once in the erythrocyte cytoplasm theprotein is refolded, which may involve parasite and/or host chaperones.
The current translocation model is that proteins for export arrive at the vacuolar space byvesicular trafficking and are unfolded. Hsp101 tightly binds a PTEX150 multimer and they as-sociate with oligomerized EXP2 present in the PVM (Figure 6). Cargo proteins would transitthe translocon pore and be refolded into the erythrocyte by exported parasite proteins such asHSP70-x/HSP40-like (see next section) and possibly by host chaperones.
EXPORTED PROTEINS AND THEIR FUNCTIONS
The Exportome of Plasmodium Spp.
The PEXEL motif is highly conserved and its identification in P. falciparum provided the meansto predict the exportome for Plasmodium spp. (38, 56). This revealed P. falciparum encodes thelargest exportome, with over 6% of its genome devoted to >350 exported proteins (78, 94). Thelarge catalogue of PEXEL proteins unique to P. falciparum is due partly to expansion and radi-ation of genes encoding protein families, including the PHIST (Plasmodium helical interspersedsubtelomeric) superfamily and heat shock protein 40-like (Hsp40-like) families (78) (Table 1). Alarge-scale investigation revealed that some P. falciparum PEXEL proteins are involved in cellularremodeling, cytoadherence, and parasite survival (54) (Table 1). The smaller PEXEL exportomefor parasites such as P. berghei and P. yoelii suggests they require fewer exported proteins to establishtheir intracellular niche, or deploy a larger catalogue of PNEPs yet to be identified.
It has not been possible to predict the full repertoire of exported proteins in Plasmodium spp.using bioinformatics, as PNEPs lack a conserved sequence element. The number of PNEPs inP. falciparum is presently relatively small, with the PfEMP1 and SURFIN families (106), as wellas nine others, currently known (Table 1; Figure 5). Because P. falciparum uniquely expressesthe large adhesive PfEMP1 family (as does P. reichenowi ) and extrudes Maurer’s clefts into theerythrocyte to transport and sort proteins, it might be expected that a proportion of exportedP. falciparum proteins are unique and required for the trafficking and function of PfEMP1. Theexpression of 19 exported Hsp40-like proteins (9, 78) and at least one exported Hsp70 protein
www.annualreviews.org • Remaking the Erythrocyte from the Inside Out 259
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
(48) is consistent with this possibility (see below). Functional analysis of exported P. falciparumproteins has shown that many are involved in PfEMP1 trafficking and surface display, as well asmaintaining Maurer’s clefts architecture (Table 1). Examples are discussed below.
Cytoadherence Complex
PfEMP1-mediated cytoadherence to host receptors requires the cytoadherence complex, which isessential for structurally reinforcing PfEMP1 in the cell membrane and is composed of ancillaryexported proteins (18, 105). The best-studied example is KAHRP, which is exported soon aftermerozoite invasion. After export, KAHRP concentrates around the cytoplasmic face of Maurer’sclefts (105), possibly associating with the acidic terminal sequence (ATS) of PfEMP1 (67, 95,96). Recent in vitro binding studies suggest that KAHRP may not directly interact with the ATSbut that another exported protein, having a PHISTc domain (PF3D7_0936800), may bind (58).After interacting with Maurer’s clefts, KAHRP then traffics to and binds host spectrin in thecytoskeleton (67, 70) to generate one of the most prominent features of the P. falciparum–infectederythrocyte, knobs, which are a principal component of the cytoadherence complex (Figure 3).
Maurer’s clefts migrate to the erythrocyte periphery, transporting PfEMP1 and other pro-teins/complexes (see below) to beneath the erythrocyte membrane. Maurer’s clefts eventu-ally tether to the host cell membrane before another remarkable remodeling step ensues—translocation of PfEMP1 from Maurer’s clefts onto the erythrocyte surface. This process beginsat approximately 16 h post merozoite invasion (Figure 3) by mechanism(s) that are poorly under-stood (see below for a description of exported proteins involved) (47, 69). It may involve vesicularbudding from Maurer’s clefts and fusion with the erythrocyte membrane, presenting PfEMP1on the surface, where it concentrates within knobs, with the adherent ectodomain facing externalmilieu.
KAHRP is the main constituent of knobs, and disruption of its expression causes loss of knobs onthe cell surface (18). However, although this does not reduce PfEMP1 trafficking to the erythrocytesurface, consistent with the suggestion that it does not directly bind the ATS domain, it doesdramatically reduce cytoadherence under physiological flow conditions. This demonstrates thatknobs are important for PfEMP1-mediated adherence. Knob formation does not rely solely onKAHRP. The exported PEXEL proteins, PFD1170c and PF10_0381, play key roles as disruptionof the corresponding genes gave a knobless phenotype (PfEMP1 surface localization was againnot affected), similar to that observed when KAHRP expression was disrupted (54).
PFD1170c has a PHISTb domain that is likely involved in protein-protein interactions; how-ever, its specific function in knob assembly is currently unknown (54). PF10_0381 contains a typeIV DnaJ domain and is classified as Hsp40-like (Table 1; Figure 5). Hsp40 proteins are cochap-erones that regulate Hsp70 function; they are divided into four classes based on the presence orabsence of four domains. These include the DnaJ domain with a conserved His-Pro-Asp (HPD)motif (Figure 5), a Gly/Phe-rich region, a cysteine-rich zinc-binding domain, and a C-terminalsubstrate-binding domain (14). Type I Hsp40 proteins have all four domains; type II proteins lackthe zinc-binding domain; and type III and IV proteins have a signature DnaJ domain, with typeIV exhibiting variations in the HPD motif (9, 14, 97). The HPD motif is essential for stimulatingHsp70 ATPase activity, and its mutation abolishes the Hsp40-Hsp70 interaction. P. falciparum ispredicted to encode 19 exported Hsp40-like proteins (type II/III/IV), with 11 possessing the varianttype IV HPD motif (Table 1; Figure 5). This suggests they perform important cochaperone-likefunctions in erythrocytes, independently of Hsp70, and the role of PF10_0381 in knob assemblyis a good example. Recently, an exported parasite Hsp70 was identified (Hsp70-x) and it may bea target of one or more exported type II/III Hsp40-like proteins (48) (Table 1).
260 Boddey · Cowman
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
PfEMP1 Trafficking and Display on the Erythrocyte Surface
Following formation of the cytoadherence complex, PfEMP1 is exposed on the erythrocyte sur-face, a process requiring the function of a number of PNEPs and PEXEL proteins. Several PNEPscharacterized in P. falciparum are required for trafficking of PfEMP1. Disruption of the genes en-coding membrane-associated histidine-rich protein 1 (MAHRP1) (87) and SBP1 (16, 53) abolisheddisplay of PfEMP1 on the infected erythrocyte surface. MAHRP1 and SBP1 are membrane pro-teins and localize in Maurer’s clefts. They are thought to facilitate PfEMP1 transfer to Maurer’sclefts during, or after, their genesis from the PVM. Loss of MAHRP1 function destabilizes Mau-rer’s clefts and it may play a structural role rather than a direct role in PfEMP1 transfer. The PNEPREX1 is also localized in Maurer’s cleft by a coiled-coil domain (21) (Figure 5). REX1 mutationdecreased PfEMP1 trafficking to the erythrocyte surface; however, these mutations destabilizedMaurer’s clefts architecture, suggesting it may be a structural protein.
Six PEXEL proteins identified in P. falciparum play roles in PfEMP1 trafficking and are namedPTP1–6 (PfEMP1-trafficking proteins 1–6) (54) (Table 1). Disruption of PTP3, PTP4, and PTP6expression in P. falciparum resulted in decreased levels of exported PfEMP1, whereas inactivationof PTP1, PTP2, and PTP3 expression abolished PfEMP1 display on the erythrocyte surface.PTP1 localizes in Maurer’s clefts and may help load or traffic PfEMP1 from the PVM into thesestructures. PTP2 and PTP3 are localized in Maurer’s clefts and also in vesicles in the erythrocytecytosol; their function is essential for the translocation of PfEMP1 from Maurer’s clefts onto theerythrocyte surface.
Enhancing Erythrocyte Rigidity
A key consequence of P. falciparum infection is that erythrocytes lose their deformability, be-coming increasingly spherocytic and rigid. Although this improves the ability to cytoadhere tohost receptors, it can block microcapillaries in the vasculature of major organs, contributing tomalaria pathogenesis (64). Uninfected erythrocytes are highly deformable, due largely to the elas-tic properties of the underlying cytoskeleton; however, exported parasite proteins that bind theerythrocyte cytoskeleton dramatically decrease its elasticity. The ring-infected erythrocyte surfaceantigen (RESA) is one such protein, and it is deployed into the erythrocyte soon after merozoiteinvasion, where it binds spectrin (24). This stabilizes the spectrin tetramer against dissociation,increasing cellular rigidity and sensitivity to thermal degradation (61, 71). RESA also slows thetransit velocity of parasitized erythrocytes, especially at febrile temperature (20). RESA is impor-tant for protecting the integrity of parasitized erythrocytes during febrile attack. Other proteinsassociated with modulation of parasite-infected erythrocyte rigidity include Pf332, a megadaltonprotein (28, 39), two exported FIKK kinases that appear to phosphorylate the cytoskeleton (66),members of the STEVOR family (77), and several PEXEL proteins examined in a large-scalegene knockout screen (54) (Table 1).
New Permeability Pathways
Parasite expansion through intraerythrocytic development requires nutrient uptake and wasteexchange. Parasites rapidly establish a new permeability pathway (NPP) in the infected erythrocytemembrane, conferring significant permeability to the membrane (1, 27, 88). The NPP ensures anoverall shift in the intracellular ionic environment so the cell does not lyse from osmotic stress ascontents change. The NPP is also called the plasmodial surface anion channel and is conferred byexpression of either of two exported PNEPs (CLAG3.1/3.2) that are conserved across Plasmodium
www.annualreviews.org • Remaking the Erythrocyte from the Inside Out 261
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
spp. (65). Either CLAG3.1 or CLAG3.2 is inserted into the membrane of the host cell duringmerozoite invasion or soon after by an unknown mechanism. The two CLAG3 proteins undergoexpression switching, a property that may assist in evading host immune responses, as a portionof these proteins may be exposed on the erythrocyte surface.
TIMING OF EVENTS IN EXPORT
The ability to perform live-cell imaging and super-resolution microscopy has provided new toolsto follow progression of P. falciparum parasites through the life cycle as well as new insights intomerozoite invasion of erythrocytes and cellular remodeling events (31, 74). Merozoite invasion isan ordered process with the final stages involving release of apical organelles for the formationof the PVM and PV from parasite lipid and proteins (73). After invasion is completed, the ringstage is surprisingly dynamic, going through actin-dependent changes in shape that include elon-gated amoeboid-like extensions (31). Visualization of proteins via super-resolution microscopyhas shown that some of the shape changes observed in the newly invaded ring may result fromfusion of dense granules with the parasite membrane and release of contents (74). This importantstep establishes the structure of the PV and PVM by insertion of proteins and complexes such asthe PTEX translocon machinery from dense granules (11). Rapid deployment of the transloconto the PVM would enable export of proteins such as RESA within minutes of merozoite invasion,where these proteins traffic to the underside of erythrocyte membranes associating with spectrin.
The Maurer’s clefts are formed during ring-stage development of parasites by budding fromthe PVM. Previously, Maurer’s clefts were thought to form throughout ring-stage developmentof P. falciparum; however, a recent study has suggested they are synthesized in early ring stages andtheir numbers remain stable throughout blood-stage development (31). This has raised questionsas to how proteins that localize to Maurer’s clefts are trafficked to this organelle when they areexpressed after Maurer’s clefts are released from the PV. It recalled previous models that suggestedmembrane-associated proteins such as PfEMP1 are trafficked to Maurer’s clefts from the PV invesicles or as a soluble complex (69). Certainly, soluble PEXEL-containing proteins, such asKAHRP and PfEMP3, are expressed and exported approximately 4 h postinvasion through theparasite membrane and PVM into the host cell cytosol, where they bind to the outside of Maurer’sclefts, a step that is essential for assembly in complexes under the parasitized erythrocyte (98, 105).Regardless of the timing of Maurer’s cleft formation and release, they play an essential role in hostcell remodeling.
CONCLUSIONS AND OUTLOOK
Many viruses and bacteria commandeer host cell machinery to subvert cellular processes andensure their survival and proliferation. In contrast, the malaria parasite invades and renovatesthe erythrocyte, turning a terminally differentiated cell, lacking most of the machinery and func-tions of a typical cell, into a dynamic and privileged niche in which it can grow, transport manyhundreds of proteins, scavenge nutrients, sequester in the presence of a circulating bloodstream,and facilitate transmission back into mosquitoes. Our understanding of this process has greatlyincreased; however, there is much to understand. Most exported proteins have been identifiedthrough bioinformatic searches of Plasmodium genome sequences using the PEXEL motif—howmany exported proteins remain undiscovered? Of the more than 540 PNEPs and PEXEL proteinsso far identified in P. falciparum, only a fraction have been characterized functionally.
The PTEX complex has been identified at the PVM and it has the features expected of atranslocation machine, but proof of its role in this process is lacking. Although it is membrane
262 Boddey · Cowman
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
associated, it is not clear whether it is associated only with the PVM or whether it spans the PV toconnect directly to the parasite membrane. The latter model would be similar to several bacterialsecretion systems that deploy both soluble and membrane-associated proteins across two or moremembranes in one translocation step.
Plasmodium spp. have developed two modes of protein export—one for PEXEL proteins andanother for PNEPs—and the reasons are not obvious, but different routes may be required forparticular subcellular locations and membrane topologies in the host cell. Plasmepsin V cleavageof the PEXEL motif in the ER is a key event that identifies these proteins for export, but therole of PI3P in exporting either PEXEL proteins or PNEPs is presently not clear. Currentdata suggest that the PTEX translocon is involved in exporting PEXEL-containing proteins;however, it is also likely involved in trafficking of PNEPs. Once these proteins are traffickedacross the parasite membrane and PVM, are they loaded directly into Maurer’s clefts or is therea vesicular system, or do they transfer as chaperoned soluble complexes? It is likely that all thesepossibilities function in Plasmodium spp., and the exact trafficking route they take depends ontheir final localization in the infected erythrocyte.
SUMMARY POINTS
1. A large number of proteins are exported to the P. falciparum–infected erythrocyte andthese proteins play an important role in remodeling the host cell.
2. PfEMP1 is a major virulence protein in P. falciparum responsible for cytoadherence andantigenic variation.
3. The presence of a PEXEL/VTS motif, and its processing by plasmepsin V, is requiredfor the export of several hundred proteins, and this is conserved in Plasmodium spp.
4. There are two classes of exported proteins, PEXEL and PNEP, and they appear to beexported by different trafficking routes, at least through the ER.
5. The PTEX complex located at the PVM has been implicated as a translocation machinefor protein export.
FUTURE ISSUES
1. The location and role of PI3P binding by PEXEL proteins prior to cleavage by plasmepsinV need to be clarified.
2. The mechanism of PEXEL export immediately following plasmepsin V requires exam-ination. Is there a PEXEL receptor or chaperone?
3. The trafficking route for PEXEL proteins and PNEPs from the ER to the parasitemembrane needs to be defined.
4. Functional evidence that the PTEX complex is involved in exporting PEXEL proteinsand PNEPs is required.
5. The mechanism of protein transfer from the PVM to Maurer’s clefts and to the erythro-cyte membrane requires detailed investigation.
www.annualreviews.org • Remaking the Erythrocyte from the Inside Out 263
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings thatmight be perceived as affecting the objectivity of this review.
ACKNOWLEDGMENTS
We thank all past and present members of the laboratory for their contributions. T. Spielmannand J. Przyborski are thanked for advice on the manuscript. Funding by the National Health andMedical Research Council of Australia, the Australian Research Council, and the Howard HughesMedical Institute is acknowledged. We apologize to colleagues whose work we could not cite dueto page limitations.
LITERATURE CITED
1. Alkhalil A, Cohn JV, Wagner MA, Cabrera JS, Rajapandi T, Desai SA. 2004. Plasmodium falciparumlikely encodes the principal anion channel on infected human erythrocytes. Blood 104:4279–86
2. Arnot DE, Jensen AT. 2011. Antigenic variation and the genetics and epigenetics of the PfEMP1 ery-throcyte surface antigens in Plasmodium falciparum malaria. Adv. Appl. Microbiol. 74:77–96
3. Baruch DI, Pasloske BL, Singh HB, Bi X, Ma XC, et al. 1995. Cloning the P. falciparum gene encod-ing PfEMP1, a malarial variant antigen and adherence receptor on the surface of parasitized humanerythrocytes. Cell 82:77–87
4. Bhattacharjee S, Hiller NL, Liolios K, Win J, Kanneganti TD, et al. 2006. The malarial host-targetingsignal is conserved in the Irish potato famine pathogen. PLoS Pathog. 2:e50
5. Bhattacharjee S, Stahelin RV, Speicher KD, Speicher DW, Haldar K. 2012. Endoplasmic reticulumPI(3)P lipid binding targets malaria proteins to the host cell. Cell 148:201–12
6. Boddey JA, Carvalho TG, Hodder AN, Sargeant TJ, Sleebs BE, et al. 2013. Role of Plasmepsin V inexport of diverse protein families from the Plasmodium falciparum exportome. Traffic 14:532–50
7. Boddey JA, Hodder AN, Gunther S, Gilson PR, Patsiouras H, et al. 2010. An aspartyl protease directsmalaria effector proteins to the host cell. Nature 463:627–31
8. Boddey JA, Moritz RL, Simpson RJ, Cowman AF. 2009. Role of the Plasmodium export element intrafficking parasite proteins to the infected erythrocyte. Traffic 10:285–99
9. Botha M, Pesce ER, Blatch GL. 2007. The Hsp40 proteins of Plasmodium falciparum and other Apicom-plexa: regulating chaperone power in the parasite and the host. Int. J. Biochem. Cell Biol. 39:1781–803
10. Boucher IW, McMillan PJ, Gabrielsen M, Akerman SE, Brannigan JA, et al. 2006. Structural and bio-chemical characterization of a mitochondrial peroxiredoxin from Plasmodium falciparum. Mol. Microbiol.61:948–59
11. Bullen HE, Charnaud SC, Kalanon M, Riglar DT, Dekiwadia C, et al. 2012. Biosynthesis, localization,and macromolecular arrangement of the Plasmodium falciparum translocon of exported proteins (PTEX).J. Biol. Chem. 287:7871–84
12. Chang HH, Falick AM, Carlton PM, Sedat JW, DeRisi JL, Marletta MA. 2008. N-terminal processingof proteins exported by malaria parasites. Mol. Biochem. Parasitol. 160:107–15
13. Cheeseman IH, Miller BA, Nair S, Nkhoma S, Tan A, et al. 2012. A major genome region underlyingartemisinin resistance in malaria. Science 336:79–82
14. Cheetham ME, Caplan AJ. 1998. Structure, function and evolution of DnaJ: conservation and adaptationof chaperone function. Cell Stress Chaperones 3:28–36
15. Cheng Q, Cloonan N, Fischer K, Thompson J, Waine G, et al. 1998. stevor and rif are Plasmodiumfalciparum multicopy gene families which potentially encode variant antigens. Mol. Biochem. Parasitol.97:161–76
16. Cooke BM, Buckingham DW, Glenister FK, Fernandez KM, Bannister LH, et al. 2006. A Maurer’scleft-associated protein is essential for expression of the major malaria virulence antigen on the surfaceof infected red blood cells. J. Cell Biol. 172:899–908
264 Boddey · Cowman
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
17. Coppel RL, Culvenor JG, Bianco AE, Crewther PE, Stahl HD, et al. 1986. Variable antigen associatedwith the surface of erythrocytes infected with mature stages of Plasmodium falciparum. Mol. Biochem.Parasitol. 20:265–77
18. Crabb BS, Cooke BM, Reeder JC, Waller RF, Caruana SR, et al. 1997. Targeted gene disruption showsthat knobs enable malaria-infected red cells to cytoadhere under physiological shear stress. Cell 89:287–96
19. de Koning-Ward TF, Gilson PR, Boddey JA, Rug M, Smith BJ, et al. 2009. A newly discovered proteinexport machine in malaria parasites. Nature 459:945–49
20. Diez-Silva M, Park Y, Huang S, Bow H, Mercereau-Puijalon O, et al. 2012. Pf155/RESA proteininfluences the dynamic microcirculatory behavior of ring-stage Plasmodium falciparum infected red bloodcells. Sci. Rep. 2:614
21. Dixon MW, Hawthorne PL, Spielmann T, Anderson KL, Trenholme KR, Gardiner DL. 2008. Tar-geting of the ring exported protein 1 to the Maurer’s clefts is mediated by a two-phase process. Traffic9:1316–26
22. Duszenko M, Ginger ML, Brennand A, Gualdron-Lopez M, Colombo MI, et al. 2011. Autophagy inprotists. Autophagy 7:127–58
23. Ellis JG, Dodds PN. 2011. Showdown at the RXLR motif: serious differences of opinion in how effectorproteins from filamentous eukaryotic pathogens enter plant cells. Proc. Natl. Acad. Sci. USA 108:14381–82
24. Foley M, Tilley L, Sawyer WH, Anders RF. 1991. The ring-infected erythrocyte surface antigen ofPlasmodium falciparum associates with spectrin in the erythrocyte membrane. Mol. Biochem. Parasitol.46:137–47
25. Gardner MJ, Hall N, Fung E, White O, Berriman M, et al. 2002. Genome sequence of the humanmalaria parasite Plasmodium falciparum. Nature 419:498–511
26. Gehde N, Hinrichs C, Montilla I, Charpian S, Lingelbach K, Przyborski JM. 2009. Protein unfolding isan essential requirement for transport across the PVM of Plasmodium falciparum. Mol. Microbiol. 71:613–28
27. Gero AM, Wood AM. 1991. New nucleoside transport pathways induced in the host erythrocyte mem-brane of malaria and Babesia infected cells. Adv. Exp. Med. Biol. 309A:169–72
28. Glenister FK, Fernandez KM, Kats LM, Hanssen E, Mohandas N, et al. 2009. Functional alteration ofred blood cells by a megadalton protein of Plasmodium falciparum. Blood 113:919–28
29. Grouffaud S, van West P, Avrova AO, Birch PR, Whisson SC. 2008. Plasmodium falciparumand Hyaloperonospora parasitica effector translocation motifs are functional in Phytophthora infestans.Microbiology 154:3743–51
30. Gruring C, Heiber A, Kruse F, Flemming S, Franci G, et al. 2012. Uncovering common principles inprotein export of malaria parasites. Cell Host Microbe 12:717–29
31. Gruring C, Heiber A, Kruse F, Ungefehr J, Gilberger TW, Spielmann T. 2011. Development and hostcell modifications of Plasmodium falciparum blood stages in four dimensions. Nat. Commun. 2:165
32. Haase S, Herrmann S, Gruring C, Heiber A, Jansen PW, et al. 2009. Sequence requirements for theexport of the Plasmodium falciparum Maurer’s clefts protein REX2. Mol. Microbiol. 71:1003–17
33. Hanssen E, Carlton P, Deed S, Klonis N, Sedat J, et al. 2010. Whole cell imaging reveals novel modularfeatures of the exomembrane system of the malaria parasite, Plasmodium falciparum. Int. J. Parasitol.40:123–34
34. Hanssen E, Hawthorne P, Dixon MW, Trenholme KR, McMillan PJ, et al. 2008. Targeted mutagen-esis of the ring-exported protein-1 of Plasmodium falciparum disrupts the architecture of Maurer’s cleftorganelles. Mol. Microbiol. 69:938–53
35. Hanssen E, Knoechel C, Klonis N, Abu-Bakar N, Deed S, et al. 2011. Cryo transmission X-ray imagingof the malaria parasite, P. falciparum. J. Struct. Biol. 173:161–68
36. Hanssen E, Sougrat R, Frankland S, Deed S, Klonis N, et al. 2008. Electron tomography of the Maurer’scleft organelles of Plasmodium falciparum-infected erythrocytes reveals novel structural features. Mol.Microbiol. 67:703–18
37. Hawthorne PL, Trenholme KR, Skinner-Adams TS, Spielmann T, Fischer K, et al. 2004. A novelPlasmodium falciparum ring stage protein, REX, is located in Maurer’s clefts. Mol. Biochem. Parasitol.136:181–89
www.annualreviews.org • Remaking the Erythrocyte from the Inside Out 265
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
38. Hiller NL, Bhattacharjee S, van Ooij C, Liolios K, Harrison T, et al. 2004. A host-targeting signal invirulence proteins reveals a secretome in malarial infection. Science 306:1934–37
39. Hodder AN, Maier AG, Rug M, Brown M, Hommel M, et al. 2009. Analysis of structure and functionof the giant protein Pf332 in Plasmodium falciparum. Mol. Microbiol. 71:48–65
40. Johnson D, Gunther K, Ansorge I, Benting J, Kent A, et al. 1994. Characterization of membrane proteinsexported from Plasmodium falciparum into the host erythrocyte. Parasitology 109(Pt. 1):1–9
41. Jonas MC, Pehar M, Puglielli L. 2010. AT-1 is the ER membrane acetyl-CoA transporter and is essentialfor cell viability. J. Cell Sci. 123:3378–88
42. Kale SD, Gu B, Capelluto DG, Dou D, Feldman E, et al. 2010. External lipid PI3P mediates entry ofeukaryotic pathogen effectors into plant and animal host cells. Cell 142:284–95
43. Kitamura K, Kishi-Itakura C, Tsuboi T, Sato S, Kita K, et al. 2012. Autophagy-related Atg8 localizes tothe apicoplast of the human malaria parasite Plasmodium falciparum. PLoS One 7:e42977
44. Knuepfer E, Rug M, Klonis N, Tilley L, Cowman AF. 2005. Trafficking of the major virulence factorto the surface of transfected P. falciparum-infected erythrocytes. Blood 105:4078–87
45. Ko MH, Puglielli L. 2009. Two endoplasmic reticulum (ER)/ER Golgi intermediate compartment-basedlysine acetyltransferases post-translationally regulate BACE1 levels. J. Biol. Chem. 284:2482–92
46. Kochan J, Perkins M, Ravetch JV. 1986. A tandemly repeated sequence determines the binding domainfor an erythrocyte receptor binding protein of P. falciparum. Cell 44:689–96
47. Kriek N, Tilley L, Horrocks P, Pinches R, Elford BC, et al. 2003. Characterization of the pathway fortransport of the cytoadherence-mediating protein, PfEMP1, to the host cell surface in malaria parasite-infected erythrocytes. Mol. Microbiol. 50:1215–27
48. Kulzer S, Charnaud S, Dagan T, Riedel J, Mandal P, et al. 2012. Plasmodium falciparum-encoded exportedhsp70/hsp40 chaperone/co-chaperone complexes within the host erythrocyte. Cell Microbiol. 14:1784–95
49. Kulzer S, Rug M, Brinkmann K, Cannon P, Cowman A, et al. 2010. Parasite-encoded Hsp40 proteinsdefine novel mobile structures in the cytosol of the P. falciparum-infected erythrocyte. Cell Microbiol.12:1398–420
50. Lavazec C, Sanyal S, Templeton TJ. 2007. Expression switching in the stevor and Pfmc-2TM superfam-ilies in Plasmodium falciparum. Mol. Microbiol. 64:1621–34
51. Lingelbach K, Przyborski JM. 2006. The long and winding road: protein trafficking mechanisms in thePlasmodium falciparum infected erythrocyte. Mol. Biochem. Parasitol. 147:1–8
52. Maier AG, Cooke BM, Cowman AF, Tilley L. 2009. Malaria parasite proteins that remodel the hosterythrocyte. Nat. Rev. Microbiol. 7:341–54
53. Maier AG, Rug M, O’Neill MT, Beeson JG, Marti M, et al. 2007. Skeleton-binding protein 1 func-tions at the parasitophorous vacuole membrane to traffic PfEMP1 to the Plasmodium falciparum-infectederythrocyte surface. Blood 109:1289–97
54. Maier AG, Rug M, O’Neill MT, Brown M, Chakravorty S, et al. 2008. Exported proteins required forvirulence and rigidity of Plasmodium falciparum-infected human erythrocytes. Cell 134:48–61
55. Marti M, Baum J, Rug M, Tilley L, Cowman AF. 2005. Signal-mediated export of proteins from themalaria parasite to the host erythrocyte. J. Cell Biol. 171:587–92
56. Marti M, Good RT, Rug M, Knuepfer E, Cowman AF. 2004. Targeting malaria virulence and remodelingproteins to the host erythrocyte. Science 306:1930–33
57. Maurer G. 1902. Die Malaria pernciosa. Zentralbl. Bakteriol. Parasitenkd. 23:695–71958. Mayer C, Slater L, Erat MC, Konrat R, Vakonakis I. 2012. Structural analysis of the Plasmodium falci-
parum erythrocyte membrane protein 1 (PfEMP1) intracellular domain reveals a conserved interactionepitope. J. Biol. Chem. 287:7182–89
59. Melcher M, Muhle RA, Henrich PP, Kraemer SM, Avril M, et al. 2010. Identification of a role for thePfEMP1 semi-conserved head structure in protein trafficking to the surface of Plasmodium falciparuminfected red blood cells. Cell Microbiol. 12:1446–62
60. Miller LH, Greenwood B. 2002. Malaria—a shadow over Africa. Science 298:121–2261. Mills JP, Diez-Silva M, Quinn DJ, Dao M, Lang MJ, et al. 2007. Effect of plasmodial RESA protein
on deformability of human red blood cells harboring Plasmodium falciparum. Proc. Natl. Acad. Sci. USA104:9213–17
266 Boddey · Cowman
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
62. Morahan BJ, Strobel C, Hasan U, Czesny B, Mantel PY, et al. 2011. Functional analysis of the exportedtype IV HSP40 protein PfGECO in Plasmodium falciparum gametocytes. Eukaryot. Cell 10:1492–503
63. Mphande FA, Ribacke U, Kaneko O, Kironde F, Winter G, Wahlgren M. 2008. SURFIN4.1, a schizont-merozoite associated protein in the SURFIN family of Plasmodium falciparum. Malaria J. 7:116
64. Nash GB, O’Brien E, Gordon-Smith EC, Dormandy JA. 1989. Abnormalities in the mechanical prop-erties of red blood cells caused by Plasmodium falciparum. Blood 74:855–61
65. Nguitragool W, Bokhari AA, Pillai AD, Rayavara K, Sharma P, et al. 2011. Malaria parasite clag3 genesdetermine channel-mediated nutrient uptake by infected red blood cells. Cell 145:665–77
66. Nunes MC, Okada M, Scheidig-Benatar C, Cooke BM, Scherf A. 2010. Plasmodium falciparum FIKKkinase members target distinct components of the erythrocyte membrane. PLoS One 5:e11747
67. Oh SS, Voigt S, Fisher D, Yi SJ, LeRoy PJ, et al. 2000. Plasmodium falciparum erythrocyte membraneprotein 1 is anchored to the actin-spectrin junction and knob-associated histidine-rich protein in theerythrocyte skeleton. Mol. Biochem. Parasitol. 108:237–47
68. Pachlatko E, Rusch S, Muller A, Hemphill A, Tilley L, et al. 2010. MAHRP2, an exported protein ofPlasmodium falciparum, is an essential component of Maurer’s cleft tethers. Mol. Microbiol. 77:1136–52
69. Papakrivos J, Newbold CI, Lingelbach K. 2005. A potential novel mechanism for the insertion of a mem-brane protein revealed by a biochemical analysis of the Plasmodium falciparum cytoadherence moleculePfEMP-1. Mol. Microbiol. 55:1272–84
70. Pei X, An X, Guo X, Tarnawski M, Coppel R, Mohandas N. 2005. Structural and functional stud-ies of interaction between Plasmodium falciparum knob-associated histidine-rich protein (KAHRP) anderythrocyte spectrin. J. Biol. Chem. 280:31166–71
71. Pei X, Guo X, Coppel R, Bhattacharjee S, Haldar K, et al. 2007. The ring-infected erythrocyte surfaceantigen (RESA) of Plasmodium falciparum stabilizes spectrin tetramers and suppresses further invasion.Blood 110:1036–42
72. Rask TS, Hansen DA, Theander TG, Gorm Pedersen A, Lavstsen T. 2010. Plasmodium falciparumerythrocyte membrane protein 1 diversity in seven genomes—divide and conquer. PLoS Comput. Biol.6(9):pii:e1000933
73. Riglar DT, Richard D, Wilson DW, Boyle MJ, Dekiwadia C, et al. 2011. Super-resolution dissection ofcoordinated events during malaria parasite invasion of the human erythrocyte. Cell Host Microbe 9:9–20
74. Riglar DT, Rogers KL, Hanssen E, Turnbull L, Bullen HE, et al. 2013. Spatial association with PTEXcomplexes defines regions for effector export into Plasmodium falciparum-infected erythrocytes. Nat.Commun. 4:1415
75. Romisch K. 2012. Diversion at the ER: how Plasmodium falciparum exports proteins into host erythro-cytes. F1000 Res. 1:12
76. Russo I, Babbitt S, Muralidharan V, Butler T, Oksman A, Goldberg DE. 2010. Plasmepsin V licensesPlasmodium proteins for export into the host erythrocyte. Nature 463:632–36
77. Sanyal S, Egee S, Bouyer G, Perrot S, Safeukui I, et al. 2012. Plasmodium falciparum STEVOR proteinsimpact erythrocyte mechanical properties. Blood 119:e1–8
78. Sargeant TJ, Marti M, Caler E, Carlton JM, Simpson K, et al. 2006. Lineage-specific expansion ofproteins exported to erythrocytes in malaria parasites. Genome Biol. 7:R12
79. Scherf A, Hernandez-Rivas R, Buffet P, Bottius E, Benatar C, et al. 1998. Antigenic variation in malaria:in situ switching, relaxed and mutually exclusive transcription of var genes during intra-erythrocyticdevelopment in Plasmodium falciparum. EMBO J. 17:5418–26
80. Scott DC, Monda JK, Bennett EJ, Harper JW, Schulman BA. 2011. N-terminal acetylation acts as anavidity enhancer within an interconnected multiprotein complex. Science 334:674–78
81. Smith JD, Chitnis CE, Craig AG, Roberts DJ, Hudson-Taylor DE, et al. 1995. Switches in expressionof Plasmodium falciparum var genes correlate with changes in antigenic and cytoadherent phenotypes ofinfected erythrocytes. Cell 82:101–10
82. Spence PJ, Langhorne J. 2012. T cell control of malaria pathogenesis. Curr. Opin. Immunol. 24:444–4883. Spielmann T, Gilberger TW. 2010. Protein export in malaria parasites: Do multiple export motifs add
up to multiple export pathways? Trends Parasitol. 26:6–10
www.annualreviews.org • Remaking the Erythrocyte from the Inside Out 267
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
84. Spielmann T, Hawthorne PL, Dixon MW, Hannemann M, Klotz K, et al. 2006. A cluster of ringstage-specific genes linked to a locus implicated in cytoadherence in Plasmodium falciparum codes forPEXEL-negative and PEXEL-positive proteins exported into the host cell. Mol. Biol. Cell 17:3613–24
85. Spycher C, Klonis N, Spielmann T, Kump E, Steiger S, et al. 2003. MAHRP-1, a novel Plasmod-ium falciparum histidine-rich protein, binds ferriprotoporphyrin IX and localizes to the Maurer’s clefts.J. Biol. Chem. 278:35373–83
86. Spycher C, Rug M, Klonis N, Ferguson DJ, Cowman AF, et al. 2006. Genesis of and trafficking to theMaurer’s clefts of Plasmodium falciparum-infected erythrocytes. Mol. Cell. Biol. 26:4074–85
87. Spycher C, Rug M, Pachlatko E, Hanssen E, Ferguson D, et al. 2008. The Maurer’s cleft proteinMAHRP1 is essential for trafficking of PfEMP1 to the surface of Plasmodium falciparum-infected ery-throcytes. Mol. Microbiol. 68:1300–14
88. Staines HM, Rae C, Kirk K. 2000. Increased permeability of the malaria-infected erythrocyte to organiccations. Biochim. Biophys. Acta 1463:88–98
89. Su XZ, Heatwole VM, Wertheimer SP, Guinet F, Herrfeldt JA, et al. 1995. The large diverse gene familyvar encodes proteins involved in cytoadherence and antigenic variation of Plasmodium falciparum-infectederythrocytes. Cell 82:89–100
90. Su XZ, Kirkman LA, Fujioka H, Wellems TE. 1997. Complex polymorphisms in an ∼330 kDa proteinare linked to chloroquine-resistant P. falciparum in Southeast Asia and Africa. Cell 91:593–603
91. Takala-Harrison S, Clark TG, Jacob CG, Cummings MP, Miotto O, et al. 2013. Genetic loci associatedwith delayed clearance of Plasmodium falciparum following artemisinin treatment in Southeast Asia. Proc.Natl. Acad. Sci. USA 110:240–45
92. Tarr SJ, Cryar A, Thalassinos K, Haldar K, Osborne AR. 2013. The C-terminal portion of the cleavedHT motif is necessary and sufficient to mediate export of proteins from the malaria parasite into its hostcell. Mol. Microbiol. 87:835–50
93. Trelka DP, Schneider TG, Reeder JC, Taraschi TF. 2000. Evidence for vesicle-mediated trafficking ofparasite proteins to the host cell cytosol and erythrocyte surface membrane in Plasmodium falciparuminfected erythrocytes. Mol. Biochem. Parasitol. 106:131–45
94. van Ooij C, Tamez P, Bhattacharjee S, Hiller NL, Harrison T, et al. 2008. The malaria secretome: fromalgorithms to essential function in blood stage infection. PLoS Pathog. 4:e1000084
95. Waller KL, Cooke BM, Nunomura W, Mohandas N, Coppel RL. 1999. Mapping the binding do-mains involved in the interaction between the Plasmodium falciparum knob-associated histidine-rich pro-tein (KAHRP) and the cytoadherence ligand P. falciparum erythrocyte membrane protein 1 (PfEMP1).J. Biol. Chem. 274:23808–13
96. Waller KL, Nunomura W, Cooke BM, Mohandas N, Coppel RL. 2002. Mapping the domains of thecytoadherence ligand Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) that bind to theknob-associated histidine-rich protein (KAHRP). Mol. Biochem. Parasitol. 119:125–29
97. Walsh P, Bursac D, Law YC, Cyr D, Lithgow T. 2004. The J-protein family: modulating proteinassembly, disassembly and translocation. EMBO Rep. 5:567–71
98. Waterkeyn JF, Wickham ME, Davern K, Cooke BM, Reeder JC, et al. 2000. Targeted mutagenesis ofPlasmodium falciparum erythrocyte membrane protein 3 (PfEMP3) disrupts cytoadherence of malaria-infected red blood cells. EMBO J. 19:2813–23
99. Wawra S, Agacan M, Boddey JA, Davidson I, Gachon CM, et al. 2012. Avirulence protein 3a (AVR3a)from the potato pathogen Phytophthora infestans forms homodimers through its predicted translocationregion and does not specifically bind phospholipids. J. Biol. Chem. 287:38101–9
100. Wawra S, Bain J, Durward E, de Bruijn I, Minor KL, et al. 2012. Host-targeting protein 1 (SpHtp1)from the oomycete Saprolegnia parasitica translocates specifically into fish cells in a tyrosine-O-sulphate-dependent manner. Proc. Natl. Acad. Sci. USA 109:2096–101
101. Wellems TE, Walliker D, Smith CL, do Rosario VE, Maloy WL, et al. 1987. A histidine-rich proteingene marks a linkage group favored strongly in a genetic cross of Plasmodium falciparum. Cell 49:633–42
102. Whisson SC, Boevink PC, Moleleki L, Avrova AO, Morales JG, et al. 2007. A translocation signal fordelivery of oomycete effector proteins into host plant cells. Nature 450:115–18
103. WHO. 2011. World Malaria Report 2011. Geneva: WHO
268 Boddey · Cowman
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67CH12-Cowman ARI 5 August 2013 16:34
104. Wiame E, Tyteca D, Pierrot N, Collard F, Amyere M, et al. 2010. Molecular identification of aspartateN-acetyltransferase and its mutation in hypoacetylaspartia. Biochem. J. 425:127–36
105. Wickham ME, Rug M, Ralph SA, Klonis N, McFadden GI, et al. 2001. Trafficking and assembly of thecytoadherence complex in Plasmodium falciparum-infected human erythrocytes. EMBO J. 20:1–14
106. Winter G, Kawai S, Haeggstrom M, Kaneko O, von Euler A, et al. 2005. SURFIN is a polymorphicantigen expressed on Plasmodium falciparum merozoites and infected erythrocytes. J. Exp. Med. 201:1853–63
107. Yaeno T, Li H, Chaparro-Garcia A, Schornack S, Koshiba S, et al. 2011. Phosphatidylinositolmonophosphate-binding interface in the oomycete RXLR effector AVR3a is required for its stabilityin host cells to modulate plant immunity. Proc. Natl. Acad. Sci. USA 108:14682–87
www.annualreviews.org • Remaking the Erythrocyte from the Inside Out 269
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67-FrontMatter ARI 12 August 2013 12:26
Annual Review ofMicrobiology
Volume 67, 2013 Contents
Fifty Years Fused to LacJonathan Beckwith � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
3′ Cap-Independent Translation Enhancers of Plant VirusesAnne E. Simon and W. Allen Miller � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �21
Acyl-Homoserine Lactone Quorum Sensing: From Evolutionto ApplicationMartin Schuster, D. Joseph Sexton, Stephen P. Diggle, and E. Peter Greenberg � � � � � � � � �43
Mechanisms of Acid Resistance in Escherichia coliUsheer Kanjee and Walid A. Houry � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �65
The Biology of the PmrA/PmrB Two-Component System: The MajorRegulator of Lipopolysaccharide ModificationsH. Deborah Chen and Eduardo A. Groisman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �83
Transcription Regulation at the Core: Similarities Among Bacterial,Archaeal, and Eukaryotic RNA PolymerasesKimberly B. Decker and Deborah M. Hinton � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 113
Bacterial Responses to Reactive Chlorine SpeciesMichael J. Gray, Wei-Yun Wholey, and Ursula Jakob � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 141
It Takes a Village: Ecological and Fitness Impactsof Multipartite MutualismElizabeth A. Hussa and Heidi Goodrich-Blair � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 161
Electrophysiology of BacteriaAnne H. Delcour � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 179
Microbial Contributions to Phosphorus Cycling in Eutrophic Lakesand WastewaterKatherine D. McMahon and Emily K. Read � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 199
Structure and Operation of Bacterial Tripartite PumpsPhilip Hinchliffe, Martyn F. Symmons, Colin Hughes, and Vassilis Koronakis � � � � � � � � � 221
Plasmodium Nesting: Remaking the Erythrocyte from the Inside OutJustin A. Boddey and Alan F. Cowman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 243
vi
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67-FrontMatter ARI 12 August 2013 12:26
The Algal Past and Parasite Present of the ApicoplastGiel G. van Dooren and Boris Striepen � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 271
Hypoxia and Gene Expression in Eukaryotic MicrobesGeraldine Butler � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 291
Wall Teichoic Acids of Gram-Positive BacteriaStephanie Brown, John P. Santa Maria Jr., and Suzanne Walker � � � � � � � � � � � � � � � � � � � � � � 313
Archaeal Biofilms: The Great UnexploredAlvaro Orell, Sabrina Frols, and Sonja-Verena Albers � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 337
An Inquiry into the Molecular Basis of HSV Latency and ReactivationBernard Roizman and Richard J. Whitley � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 355
Molecular Bacteria-Fungi Interactions: Effects on Environment, Food,and MedicineKirstin Scherlach, Katharina Graupner, and Christian Hertweck � � � � � � � � � � � � � � � � � � � � � � � 375
Fusarium PathogenomicsLi-Jun Ma, David M. Geiser, Robert H. Proctor, Alejandro P. Rooney,
Kerry O’Donnell, Frances Trail, Donald M. Gardiner, John M. Manners,and Kemal Kazan � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 399
Biological Consequences and Advantages of AsymmetricBacterial GrowthDavid T. Kysela, Pamela J.B. Brown, Kerwyn Casey Huang, and Yves V. Brun � � � � � � � 417
Archaea in Biogeochemical CyclesPierre Offre, Anja Spang, and Christa Schleper � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 437
Experimental Approaches for Defining Functional Roles of Microbesin the Human GutGautam Dantas, Morten O.A. Sommer, Patrick H. Degnan,
and Andrew L. Goodman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 459
Plant Cell Wall Deconstruction by Ascomycete FungiN. Louise Glass, Monika Schmoll, Jamie H.D. Cate, and Samuel Coradetti � � � � � � � � � � � � 477
Cnidarian-Microbe Interactions and the Origin of InnateImmunity in MetazoansThomas C.G. Bosch � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 499
On the Biological Success of VirusesBrian R. Wasik and Paul E. Turner � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 519
Prions and the Potential Transmissibility of ProteinMisfolding DiseasesAllison Kraus, Bradley R. Groveman, and Byron Caughey � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 543
Contents vii
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.

MI67-FrontMatter ARI 12 August 2013 12:26
The Wonderful World of Archaeal VirusesDavid Prangishvili � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 565
Tip Growth in Filamentous Fungi: A Road Trip to the ApexMeritxell Riquelme � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 587
A Paradigm for Endosymbiotic Life: Cell Differentiation of RhizobiumBacteria Provoked by Host Plant FactorsEva Kondorosi, Peter Mergaert, and Attila Kereszt � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 611
Neutrophils Versus Staphylococcus aureus: A Biological Tug of WarAndras N. Spaan, Bas G.J. Surewaard, Reindert Nijland,
and Jos A.G. van Strijp � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 629
Index
Cumulative Index of Contributing Authors, Volumes 63–67 � � � � � � � � � � � � � � � � � � � � � � � � � � � 651
Errata
An online log of corrections to Annual Review of Microbiology articles may be found athttp://micro.annualreviews.org/
viii Contents
Ann
u. R
ev. M
icro
biol
. 201
3.67
:243
-269
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f M
elbo
urne
on
09/2
9/13
. For
per
sona
l use
onl
y.





























