Embed Size (px)
Citation preview

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Basic and Applied Ecology 12 (2011) 1–9
Plant invasions in temperate forests: Resistance or ephemeralphenomenon?
Franz Essla,∗, Norbert Milasowszkyb, Thomas Dirnböcka
aEnvironment Agency Austria, Spittelauer Lände 5, A-1090 Vienna, AustriabInstitut für angewandte Biologie und Umweltbildung (IFABU), Argentinierstraße 54/21, A-1040 Vienna, Austria
Received 23 March 2010; accepted 13 October 2010
Abstract
Invasion patterns in Europe are uneven across habitats. In particular, temperate zonal forests are relatively little affected byinvasions. However, long generation periods of forest species and the rarity of disturbance events may lead to long time lags,and so, invasion resistance of temperate forests may have been overestimated.
Based on the inventory of the alien vascular flora of 15 study sites in East Austrian lowland forest patches adjacent to villages,we analysed diversity and temporal trends of their alien flora and the influence of within-habitat diversity, occurrence of forestareas dominated by alien tree species and adjacent land use diversity on the level of invasion.
In total, we recorded 119 alien (neophyte) species. Species introduced by horticulture are of overriding importance (86% ofall aliens). Using the year of first record in Austria as a proxy for local residence time, numbers of aliens have increased linearlyby five species per decade since 1800. A GLM including the study site size and ratio of forest areas dominated by alien vs.native tree species explains 64% of the variance of the number of alien species. Size was more important, but the ratio of forestareas dominated by alien trees (Ailanthus altissima, Robinia pseudacacia) significantly increased alien species number.
Our results show, that invasion of alien trees with strong impacts on ecosystem properties facilitates invasion in CentralEuropean forests, leading to “invasional meltdown”. Further, high levels of propagule pressure, created by adjacent settlementsand gardens, may foster invasions. Currently observed low levels of invasion in Central European forests situated in greaterdistance to settlements may turn out to be an ephemeral phenomenon. Spread of alien forest plants on the landscape level mayultimately, although possibly only over long time periods, lead to increased levels of invasion.
Zusammenfassung
Neophyten besiedeln unterschiedliche Lebensräume Europas in unterschiedlichem Ausmaß. Im Besonderen gelten temper-ate zonale Wälder als nur wenig von biologischen Invasionen betroffen. Allerdings können die lange Generationszeit vonWaldarten und die Seltenheit von Störungserereignissen zu besonders langen Verzögerungen von Invasionen führen, sodass dieInvasionsresistenz temperater Wälder möglicherweise überschätzt wird.
Auf Basis eines Inventars der Neophytenflora von 15 an Siedlungen angrenzenden Waldflächen im östlichen TieflandÖsterreichs untersuchten wir Diversität und zeitliche Trends der Neophytenflora sowie die Bedeutung der Habitatvielfalt,des Vorkommens von durch nichtheimische Baumarten dominierten Waldflächen und angrenzender Landnutzung.
∗Corresponding author. Tel.: +43 1 31303 3323; fax: +43 1 31304 3700.E-mail address: [email protected] (F. Essl).
1439-1791/$ – see front matter © 2010 Gesellschaft für Ökologie. Published by Elsevier GmbH. All rights reserved.doi:10.1016/j.baae.2010.10.003

Author's personal copy
2 F. Essl et al. / Basic and Applied Ecology 12 (2011) 1–9
Insgesamt fanden wir 119 Neophytentaxa. Aus Gartenkultur verwilderte Arten dominierten (86% aller Neophyten). Auf Basisdes Erstnachweises der Arten in Österreich als Maßzahl für lokale Anwesenheitszeit wuchs die Neophytenflora mit einer linearenZunahme von 5 Arten pro Dekade seit 1800 an. In den letzten Jahrzehnten haben Sträucher, aus Asien stammende Arten, inGartenkultur entstandene Sippen (Anökophyten) und als Gartenpflanzen eingeführte Arten überproportional zugenommen.
Ein GLM mit Flächengröße und dem Anteil von durch nichtheimische Baumarten dominierten Waldflächen erklärt 64%der Varianz der Neophytenzahl. Flächengröße war wichtiger, aber der Anteil von durch nichtheimische Baumarten (Ailanthusaltissima, Robinia pseudacacia) dominierten Waldflächen erhöhte ebenfalls die Neophytenzahl signifikant.
Unsere Ergebnisse zeigen, dass das Eindringen nichtheimischer Baumarten mit starken ökosystemaren Auswirkungen dieAusbreitung anderer Neophyten in mitteleuropäischen Wäldern begünstigt und zu einem “invasional meltdown” führt. Angren-zende Siedlungen und Gärten scheinen wegen des hohen Samendrucks Invasionen zu begünstigen. Die derzeit beobachtetegeringe Ausbreitung von Neophyten in mitteleuropäischen Wäldern in größerem Abstand zu Siedlungen mag sich daher zukün-ftig als ein temporäres Phänomen herausstellen. Die Ausbreitung von Neophyten auf der Landschaftsebene mag letztlich,wenngleich vermutlich über längere Zeiträume, zu einem höheren Invasionsniveau führen.© 2010 Gesellschaft für Ökologie. Published by Elsevier GmbH. All rights reserved.
Keywords: Central Europe; Habitat diversity; Horticulture; Invasional meltdown; Life form; Pathway; Propagule pressure; Residence time;Temporal change; Vascular plants
Introduction
The rate of biological invasions is rapidly increasingworldwide (Lambdon et al. 2008; Hulme, Pysek, Nentwig,& Vilá 2009). Invasive species (transformer species sensuRichardson et al. 2000) are causing large detrimental effectson both biodiversity and human well-being (Vilà et al. 2009).Recent research has demonstrated that, besides climatematching, human activities are an important determinantof the biogeography of invasions (Taylor & Irwin 2004;Hulme 2009). In particular, human population size andvarious indicators of socio-economic behaviour have beenshown to be linked to patterns of invasion (McKinney2001; Pysek, Kucera, & Jarosik 2002; Taylor & Irwin 2004;Hulme 2009), because they are surrogates of propagule pres-sure and changed disturbance regimes of natural systems(McKinney 2001; Lockwood, Cassey, & Blackburn 2005;Colautti, Grigorovich, & MacIsaac 2006; Krivánek, Pysek,& Jarosik 2006).
Biological invasions are often characterized by time lagsbetween first introduction of a species in a new territory andthe start of invasive spread (Kowarik 1995; Richardson et al.2000). The duration of such time lag phenomena largelydepends on species traits (e.g. life cycle), introduction his-tory (e.g. amount of propagules or individuals introduced)and matching of the alien species autecology with the habitatconditions in the new territory (Pysek & Richardson 2007;Bucharova & van Kleunen 2009; Pysek, Krivánek, & Jarosik2009) or the time necessary for phenological or genetic adap-tations to new abiotic and biotic environments. For long-livedspecies like shrubs or trees such time lags may be substantial,reaching decades or centuries (Kowarik 1992, 1995; Pyseket al. 2009, but see Daehler 2009).
Invasion patterns in Europe are inconsistent across habi-tats (Chytry et al. 2008). So far, temperate zonal forests arerelatively little affected by invasions (Chytry, Pysek, Tichy,Knollova, & Danihelka 2005; Chytry et al. 2008; Walter, Essl,
Englisch, & Kiehn 2005). However, taking into account thatforests are dominated by woody species with long generationperiods and invasion into forests predominantly occurs whendisturbances create gaps or clearings (Chytry et al. 2005,2008) leading to temporarily increased resource availability,which can foster plant invasions (Davis, Grime, & Thompson2000), invasions in temperate forests may exhibit particu-larly long time lags. Hence, invasion resistance of temperateforests might have been overestimated.
Based on the inventory of the alien vascular flora of 15study sites in East Austrian lowland forest patches, we askthe following questions: (1) What is the diversity, structureand composition of the alien flora? (2) What are the tempo-ral trends (time elapsed since the invasion of the species inAustria) in alien species composition and is there a changeover time in the importance of pathways, regions of originand life forms? (3) What are the effects of within-habitatdiversity, occurrence of forest areas dominated by alien treespecies and adjacent land use type diversity on the level ofinvasion?
Data
Study area
The study area is situated in the Marchfeld region inthe lowlands of Eastern Austria, 30 km east of Vienna(48◦17′–48◦20′N, 16◦35′–16◦43′E, Fig. 1). The flat area ispart of the glacial Danube valley (165 m a.s.l.). The bedrockconsists of gravel deposited by the Danube during the IceAges, which is covered by sandy alluvial soils. The climateis temperate pannonic, with cool winters and warm summers,average annual temperature being 10 ◦C and annual precipita-tion 550–600 mm (1961–1990, Wiesbauer & Mazucco 1997).The Marchfeld is dominated by agriculture, so only few sig-nificant forests remain. However, in the central part of the

Author's personal copy
F. Essl et al. / Basic and Applied Ecology 12 (2011) 1–9 3
Fig. 1. Location of the study area in East Austria and of the 15forest patches within the central Marchfeld region. ©BEV 2010(T2010/70002).
Marchfeld, several extensive forests of approximately 10 km2
exist in the environs of the villages of Deutsch Wagram,Straßhof and Gänserndorf. These forests have been replantedon formerly extensively grazed dry grassland in the 18th and19th century (Wiesbauer & Mazucco 1997). Due to theirpoor growth, they are currently not or only extensively usedsilviculturally.
Species data
In the years 2008 and 2009, the alien vascular flora of 15forest patches of different sizes (ranging from 2.4 ha to 47 ha)of the central Marchfeld was recorded (Fig. 1). The study sitesare separated from each other by settlements and agriculturalfields, and the minimum distance between individual studysites is 150 m. All sites are dominated by old-growth forest(stand age > 60 years), only small fractions of the sites areoccupied by medium-aged forest (minimum stand age: 30years), whereas young-growth areas are of negligible size.Due to their uniform abiotic characteristics (flat topography,identical soil types and climate) and history, these forest sitesoffer a unique possibility for a comparative assessment ofinvasions.
For each forest patch, a set of variables was collected (seeTable S1). Based on the digital AustrianMap (www.amap.at),we calculated the size of each site. Based on field data andaerial photos, we mapped and calculated the percentage ofthree different forest types to account for habitat diversityand forestry intensity within the sites: native broadleavedforests (for.nb) dominated by native oaks (Quercus cerris, Q.petraea, Q. robur), native conifer plantations (for.ncp) dom-inated by pine species native to eastern Austria (Pinus nigra,P. sylvestris), and forests dominated by alien broadleavedtrees (Ailanthus altissima, Robinia pseudacacia; for.ab). Toaccount for the influence of adjacent habitat diversity, thepresence/absence of the following four habitats was recorded:large (two lane) roads (l.s.), railway (r.w.), crop fields (fi.),built-up areas (b.u.).
We collated a complete list of alien vascular plants, the fre-quency was assessed in five frequency classes (see Table S2).
For analyses, we used presence/absence data only. We con-sidered only alien species recorded after 1500 in Austria(neophytes), while old aliens (archaeophytes) were excluded(Richardson et al. 2000). To sample a species list as completeas possible, each site was visited at least four times in differentmonths of the growing season to cover different phenolog-ical periods. To avoid size-related sampling biases, surveysin larger sites lasted longer, with total sampling effort beingapproximately 1 h per 4 ha. Data collection was done by sys-tematically surveying all parts of the patches, while excludingforest edges. Taxonomically critical species were collectedand verified or determined by specialists (see “Acknowledge-ments”). A few species could only be identified to genus level,but they were treated as species in the analyses.
For each species, we collated a set of additional vari-ables: first record in Austria (as a proxy for the localresidence time), region of origin (continents), and dom-inant pathways (intentional introduction by horticulture[including ornamental trees] or forestry; unintentional intro-duction) based on the information provided in AliensAustria(2009). All species were assigned to life form categories:macro-phanerophytes (trees), nano-phanerophytes (shrubs),long-lived herbs (hemicryptophytes, geophytes) and shortlived herbs (annuals and biennals). Based on the databaseAliensAustria (2009), each species was classified accord-ing to its habitat affiliation as forest species or as non-forestspecies. The latter mainly consist of ruderal and grasslandspecies which occur in small gaps, along forest edges or tracksinside forests.
Taxonomy and nomenclature follow Fischer, Adler, andOswald (2008), for alien species not covered there, we fol-lowed the database AliensAustria (2009).
Statistical analyses
When testing for the influence of within-habitat and adja-cent land type diversity, we used two approaches in order toaccount for the different sizes of the forest patches. First, logarea was used as a covariate in the analysis. The independentvariables are thus related to the residual variation of the diver-sity by size model. Second, all independent variables werestandardized for the area effect by dividing through size.
Two variables were used, which are assumed to determinethe variation in species richness: Forest habitat diversity ofthe sites was expressed as the number of forest habitat typesdivided by area of the respective site (div.hab). To analysethe effects of adjacent habitat diversity, the presence andabsence of adjacent habitats were summed and then dividedby the size of the respective site (div.a). Since the numberof alien species was the response, we used generalised lin-ear models (GLM) from the Poisson family with a log-linkfunction. Significance was tested by dropping the respectiveindependent variable from the model with the size as a covari-ate using the likelihood ratio statistic and assuming a χ2-testdistribution under the null hypothesis. In addition, quasipois-

Author's personal copy
4 F. Essl et al. / Basic and Applied Ecology 12 (2011) 1–9
Table 1. Numbers of alien species of 15 forest patches as explainedby the ratio of forests areas dominated by alien vs. native treespecies within each forest patch (for.r) using generalized linearmodels (GLM) with Poisson distribution and a log-link function(null deviance: 46.9, residual deviance: 17.2). The log of the sizeof the forest patch is used as a covariate to control for area-relatedspecies richness. Significance is determined by dropping the respec-tive variable from the full model using the likelihood ratio statistic(LR) assuming a χ2 distribution. df, dev: the change in the degrees offreedom and the deviance respectively when dropping the respectivevariable from the full model; coeff: coefficient.
df dev coeff AIC LR p-Value
log (size) 1 40.9 +0.25 127.9 23.8 <0.001for.r 1 20.4 −0.44 107.3 3.2 0.073
son models were used accounting for overdispersion but asthe results were the same, we here show the simpler Pois-son models. Due to the limited number of investigated forestpatches separate models were used for: (1) the ratio (for.r)between area dominated by alien (for.ab) and area dominatedby native (for.ncb + for.nb) tree species within each forestpatch. This ratio ranges between 0 and 0.7, because area offorests dominated by native tree species is higher than forestarea dominated by alien trees in each site; (2) the diversity ofhabitats within (div.hab) and adjacent to (div.a) the respectiveforest patch. As a result of the limited sample size we acceptthe alternative hypotheses at an �-level of 0.1.
We tested the effect of residence time of the alien speciesin Austria on their abundance on the local scale (number ofinvaded forest patches) using a Kruskal–Wallis–H test. Toaccount for differences in invasion success of alien speciesof different habitat affiliations, we did this analysis sepa-rately for forest and non-forest species. We analysed temporalchanges in the composition of the alien flora by testing fordifferences of the residence time in Austria for different path-ways, regions of origin and life forms by means of KW-tests.
Statistical analyses were carried out in SPSS Version11.5.1 (Anonymous 2002) and R (R Development Core Team2009).
Results
Diversity, structure and composition of the alienflora
In total, we recorded 119 alien (neophyte) species in the15 study sites (see Table S2). Six species were recorded inall sites, whereas 51 species were rare (recorded at most intwo sites). Alien species numbers per site ranged from 25to 69 species. Size of the forest patch has a large impacton alien species number (Table 1). The most important lifeform present are long-lived herbs (36% of all alien species),followed by shrubs (27%), short-lived herbs and trees (19%
each). Tree species, however, colonized on average the high-est number of sites (8.5 sites), followed by shrubs (5.7 sites),short-lived herbs (4.0 sites) and long-lived herbs (3.9 sites).
North America, Europe and Asia are of nearly equal impor-tance as regions of origin (29–32 species), and 14 species haveoriginated in cultivation (anecophytes). No species native toother continents were found. Species introduced intentionallyare by far the largest group (88% of all aliens). The pathwayof overriding importance is horticulture (86% of all aliens),whereas only 2% of the species have been introduced forsilviculture.
Temporal change in composition of life forms,pathways and regions of origin
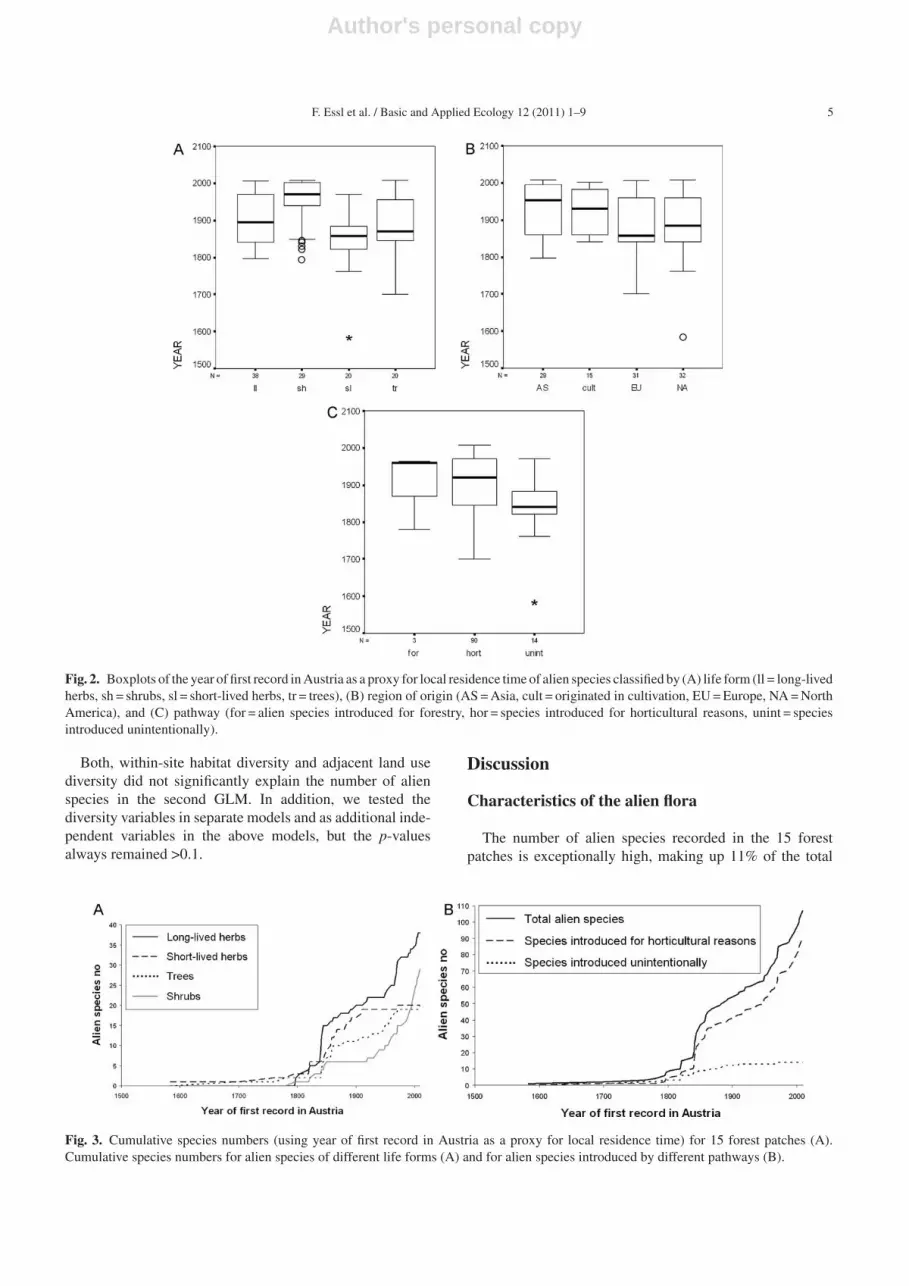
Using the year of first record in Austria as a proxy for localresidence time, invasion rate has been low until 1800. Sincethen, the number of aliens has accumulated much faster witha linear increase of 5 species per decade (Fig. 2A). Speciesintroduced after 1900 into Austria make up 50% of all alienspecies recorded.
Alien species of different life forms significantly differin their residence time (p < 0.001, KW-test). Shrubs are themost recent invaders compared to aliens of other life forms(Figs. 2A and 3A). European and North American specieshave been introduced earlier into Austria, whereas speciesnative to Asia or originated in cultivation are more recentintroductions (Fig. 2B) (p = 0.016, KW-test).
Importance of pathways has changed over time(Figs. 2C and 3B). Species introduced unintentionally arepredominantly old introductions (86% introduced before1900 into Austria), whereas only 46% of the species intro-duced for horticultural purposes have been recorded inAustria before 1900. Temporal differences between pathwaysare highly significant (p = 0.005, KW-test).
Residence time of the species in Austria had a signif-icant positive influence on the number of forest patchesinvaded (Fig. 4); this is true for forest and non-forest species(p < 0.001, KW-test), although for the latter the numbers ofinvaded forest patches are significantly lower at all timeperiods. Forest species were recorded throughout the forestsites, whereas non-forest species tended to invade gaps, foresttracks and young-growth areas.
The effect of within and adjacent habitatdiversity
The GLM including size and ratio of alien vs. native forestsexplains 64% of the variance of the number of alien species.Size was clearly more important, but the ratio significantlycontributes to the explained variance (p < 0.1; Table 1). Anegative coefficient of −0.44 means that with every 10%more of alien forest area per site, approximately 1.6 alienplant species are added to the forest vegetation.
Author's personal copy
F. Essl et al. / Basic and Applied Ecology 12 (2011) 1–9 5
Fig. 2. Boxplots of the year of first record in Austria as a proxy for local residence time of alien species classified by (A) life form (ll = long-livedherbs, sh = shrubs, sl = short-lived herbs, tr = trees), (B) region of origin (AS = Asia, cult = originated in cultivation, EU = Europe, NA = NorthAmerica), and (C) pathway (for = alien species introduced for forestry, hor = species introduced for horticultural reasons, unint = speciesintroduced unintentionally).
Both, within-site habitat diversity and adjacent land usediversity did not significantly explain the number of alienspecies in the second GLM. In addition, we tested thediversity variables in separate models and as additional inde-pendent variables in the above models, but the p-valuesalways remained >0.1.
Discussion
Characteristics of the alien flora
The number of alien species recorded in the 15 forestpatches is exceptionally high, making up 11% of the total
Fig. 3. Cumulative species numbers (using year of first record in Austria as a proxy for local residence time) for 15 forest patches (A).Cumulative species numbers for alien species of different life forms (A) and for alien species introduced by different pathways (B).

Author's personal copy
6 F. Essl et al. / Basic and Applied Ecology 12 (2011) 1–9
Fig. 4. Number of forest patches (total n = 15) invaded by alienforest (grey bars) and non-forest (white bars) species classified bytime of arrival in Austria (year of first record, 5 periods).
alien flora of Austria (1110 alien vascular plants, Rabitsch &Essl, 2006) on a total area of 307.2 ha. In addition, this speciesnumber was recorded in forests only. Besides the proximityof extensive settlements and gardens leading to high propag-ule pressure, another reason for this high level of invasionmay be attributed to the mild climate of the study region, aslevels of invasion of aliens in Central Europe are higher inthe lowlands (Pysek et al. 2002; Chytry et al. 2008).
Although the Austrian alien flora is exceptionally well doc-umented (Essl & Rabitsch 2002; AliensAustria 2009), severalof the alien species recorded have not or only very rarelybeen found before in Austria (e.g. Amelanchier lamarckii,Anemone blanda, Fritillaria imperialis, Kolkwitzia amabilis,Ribes multiflorum) and testify the dynamics of alien speciesspread in Europe (Lambdon et al. 2008; DAISIE 2009).Furthermore, several species recorded only recently for thefirst time in Austria are wide-spread in our forest patches(exclusively shrubs as Cotoneaster divaricatus, C. dielsianus,Kolkwitzia amabilis, Ribes multiflorum, see Table S2). Theincipient invasion of evergreen broadleaved shrubs and treesin the warmest parts of Central Europe due to global warm-ing (Walther 2002; Walther et al. 2009) was confirmed, asseveral species, most of them being rare and evidently onlyrecently escaped, were recorded (Euonymus fortunei, Vibur-num rhytidophyllum). Escaped fruit and nut trees of adjacentgardens (e.g. Juglans regia, Malus domestica, Morus alba,Prunus spp., Pyrus communis) are conspicuously common.
Interestingly, species of those life forms with the longestgeneration periods (trees, shrubs) occurred on average onmore sites than species with shorter generation periods,although they are on average no longer (trees) or even fora shorter time recorded as alien in Austria. We argue thatzoochory (many shrubs and trees belong to species dispersedby birds or mammals, e.g. Cotoneaster spp., Juglans regia,Prunus spp., Ribes spp.), and changes in horticultural fash-
ions (e.g. recent wide spread planting of ornamental shrubs)have contributed to this pattern, as both processes enhanceinvasion velocity at the landscape scale (Kowarik 2003b).
Temporal changes in alien species composition
We used year of first record in Austria as a proxy for localresidence time. This approach has been used widely in assess-ments of the influence of residence time on invasion successof alien species (e.g. Castro, Figueroa, Munoz-Schick, &Jaksic 2005; Pysek & Jarosik 2005; Essl & Dirnböck 2008)and we consider it a robust measure of temporal patterns ofinvasion at the local scale, although it also bears its limita-tions (e.g. local invasion may in fact have started significantlylater than on the national level). Using this proxy we inferthat the temporal changes in composition of the alien floraare substantial with regard to regions of origin, life formsand pathways. In the last decades, shrubs, species native toAsia, originated in cultivation, or introduced by horticulturehave increased disproportionally in the alien flora of the studysites.
The overriding importance of ornamentals in the alien floraof the studied forest patches (86% compared to 57% of thenational alien vascular plant flora, Essl & Rabitsch 2002) maybe attributed to the combined effects of several factors. First,in Europe (Hulme et al. 2008; Lambdon et al. 2008) and Aus-tria as well, horticulture is by far the most important pathwayfor alien vascular plants. Second, they mostly are and wereintroduced repeatedly into new regions, and horticulturalusage of these species in different environments increasespropagule pressure and the likelihood that these species willescape (Kowarik 2003b; Dehnen-Schmutz, Touza, Perrings,& Williamson 2007a, 2007b; Bucharova & van Kleunen2009). All of our study sites are adjacent to settlements withgardens constructed in the 1950s to 1980s, where planting ofornamentals and the deposition of garden wastes in adjacentforests increase local levels of propagule pressure. Third, thenumbers of cultivated ornamentals increased strongly in thelast decades (Kowarik 2003a; Dehnen-Schmutz et al. 2007b).
Predominance of early introductions in European andNorth American species compared to those originatingfrom Asia or from horticulture indicates the importance ofchanges in the structure of international trade (Hulme et al.2008; Hulme 2009). General and horticultural trade betweenEurope and temperate East Asia – the dominant region oforigin for Asian origin – has belatedly gained momentum(Mitchell 2003; Hulme 2009). Species which have originatedin cultivation mostly as results of intensive breeding efforts,have in several cases been created only during the last decades(e.g. Fallopia × bohemica, Galeobdolon argentatum). On theother hand, species native to adjacent regions of Europe or tothe former European colonies in North America were intro-duced earlier, giving them a headstart.
As expected and in concordance with recent studies onplant invasions in Europe (Pysek et al. 2009; Williamson et al.

Author's personal copy
F. Essl et al. / Basic and Applied Ecology 12 (2011) 1–9 7
2009), residence time of the species in Austria had a signifi-cant effect on the number of invaded forest patches. Numbersof invaded forest patches increased stronger with residencetime for forest species, and non-forest species invaded fewerforest patches, which reflects the fact, that the latter find onlyvery localized appropriate habitats in small forest gaps oralong tracks.
The late date of first records of shrubs in Austria comparedto other life forms was unexpected and may partly result fromchanging fashions in horticulture (e.g. recently increasedplantation of ornamental shrub species of Cotoneaster spp.and Ribes spp.).
Role of within-habitat and adjacent land usediversity
As expected by biogeographic theory and empirical evi-dence (e.g. McKinney 2001; Pysek et al. 2002; Essl &Dirnböck 2008), patch size explained a large part of the vari-ance. Second to this variable, only the proportion of alienforests contributed to the number of alien species. That is,the presence and extent of forests dominated by Ailanthusaltissima and Robinia pseudacacia contributed to highernumbers of alien species. Both species form dense stands,and particularly the latter one, which is also more widespreadin the study sites, has severe impacts on ecosystem proper-ties, especially by altering the nutrient cycle (Kowarik 2003b;Kowarik & Säumel 2007; Kleinbauer, Dullinger, Peterseil, &Essl 2009). A recent study for Austria has shown, that Robiniapseudacacia is currently invading a wide range of habitattypes (including oak forests, the dominant habitat type in ourstudy region) and that climate change will increase invasionrisks substantially across all habitat types (Kleinbauer et al.2009). Hence, both climate change and secondary releasesby planting for forestry or landscaping (Kowarik 2003b) willcontribute to future spread of Robinia pseudacacia in theforest patches.
Our results show, that invasion of alien trees with strongimpacts on ecosystem properties, facilitates invasion in Cen-tral European forests – a feature, which has been aptly termed“invasional meltdown” (Simberloff & Von Holle 1999). Sur-prisingly, within and adjacent land use did not influencethe numbers of alien species. We argue that this is due tothe fact that all sites were adjacent to gardens and built-upareas which led to high levels of local propagule pressureand which masked possible influences of other land usetypes.
Resistance of temperate forests to invasion
Open habitats (e.g. industrial, man-made habitats, grass-lands) are more receptive to invasions than closed habitats(i.e. forests; Chytry et al. 2008). In Europe, temperate forests(except for forests shaped by natural disturbances, i.e. flood-plain forests) are considered to be rather resistant to plant
invasions (Lambdon et al. 2008; Chytry et al. 2008), andthere are only few substantial and wide spread invasions(Chytry et al. 2005; Walter et al. 2005). The cause of thispaucity of invasions is primarily seen as a lack of avail-able resources and strong interspecific competition (Daviset al. 2000), which limit recruitment and the establishmentof alien species (Chytry et al. 2008). However, as forestsare dominated by long-lived woody species, time lag phe-nomena stemming from long intervals between disturbances(e.g. gaps created by windfalls or tree harvesting), and disper-sal limitation (e.g. aggravated by isolation of forest patches)may be of particular relevance (Svenning & Skov 2004,2007), as they may delay alien species spread and hencemask invasibility of forests. Additional data from transectslaid through the study sites (Essl unpubl.) show that thereare no significant correlations between stand age and num-bers of alien species recorded. So, we conclude, that inforest succession numbers of alien species in the investigatedforests does not decline significantly as forest successionproceeds (e.g. as a consequence of shading and lack of dis-turbance).
Conclusions
Our case study shows, that temperate forests are notresistant to invasions. High levels of propagule pressure,created by adjacent settlements and gardens, may lead tohigh and increasing levels of invasion, which ultimately mayprofoundly change the structure and composition of the veg-etation. Further, invasion of alien species with strong impacton ecosystem properties (transformer species) may enhanceinvasion rates in Central European forests. Most tree specieswith strong impacts on native vegetation in Central Europeare strongly controlled by temperature, so climate warmingwill increase their potential distribution (Chytry et al. 2005;Kleinbauer et al. 2009), and possibly the range of habitatscolonized.
Although the setting of our study sites is context spe-cific (e.g. large adjacent settlements with gardens, lowlandforests), observed low levels of invasion in Central Euro-pean forests situated at a greater distance to settlementsmay turn out to be an ephemeral phenomenon. Spreadof alien forest plants on the landscape level may ulti-mately, although possibly only over long time periods dueto significant lag times, lead to increased levels of inva-sion.
Acknowledgements
We are grateful to Christian Schröck and Oliver Stöhrfor their assistance in determining critical specimens. Weare grateful for constructive comments and suggestions ofthree anonymous reviewers and the managing editor K. Höve-meyer.

Author's personal copy
8 F. Essl et al. / Basic and Applied Ecology 12 (2011) 1–9
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at doi:10.1016/j.baae.2010.10.003.
References
AliensAustria, (2009). Austrian database of alien plant, animal andfungi species. Umweltbundesamt, Vienna (accessed 15.09.09).
Anonymous. (2002). SPSS for Windows Release 11.5.1. Chicago:SPSS Inc.
Bucharova, A., & van Kleunen, M. (2009). Introduction history andspecies characteristics partly explain naturalization success ofNorth American woody species in Europe. Journal of Ecology,97, 230–238.
Castro, S. A., Figueroa, J. A., Munoz-Schick, M., & Jaksic, F. M.(2005). Minimum residence time, biogeographical origin, andlife cycle as determinants of the geographical extent of natural-ized plants in continental Chile. Diversity and Distributions, 11,183–191.
Chytry, M., Pysek, P., Tichy, L., Knollova, I., & Danihelka, J. (2005).Invasions by alien plants in the Czech Republic: A quantitativeassessment across habitats. Preslia, 77, 339–354.
Chytry, M., Maskell, L. C., Pino, J., Pysek, P., Vilá, M., Font, X.,et al. (2008). Habitat invasions by alien plants: A quantitativecomparison among Mediterranean, subcontinental and oceanicregions of Europe. Journal of Applied Ecology, 45, 448–458.
Colautti, R. I., Grigorovich, I. A., & MacIsaac, H. J. (2006). Propag-ule pressure: A null model for biological invasions. BiologicalInvasions, 8, 1023–1037.
Daehler, C. C. (2009). Short lag times for invasive tropical plants:Evidence from experimental plantings in Hawai’i. PLoS ONE,4(2), e4462, doi:10.1371/journal.pone.0004462
DAISIE (Ed.). (2009). The Handbook of Alien Species in Europe.Dordrecht: Springer.
Davis, M. A., Grime, J. P., & Thompson, K. (2000). Fluctuatingresources in plant communities: A general theory of invasibility.Journal of Ecology, 88, 528–534.
Dehnen-Schmutz, K., Touza, J., Perrings, C., & Williamson, M.(2007a). The horticultural trade and ornamental plant invasionsin Britain. Conservation Biology, 21, 224–231.
Dehnen-Schmutz, K., Touza, J., Perrings, C., & Williamson, M.(2007b). A century of the ornamental plant trade and its impacton invasion success. Diversity and Distributions, 13, 527–534.
Essl, F., & Dirnböck, T. (2008). Diversity of native and alien vascularplants of dry grasslands in Central Europe. Applied VegetationScience, 11, 441–450.
Essl, F., & Rabitsch, W. (2002). Neobiota in Österreich. Umwelt-bundesamt: Wien.
Fischer, M. A., Adler, W., & Oswald, K. (2008). Exkursions-flora für Österreich, Liechtenstein und Südtirol. BiologiezentrumOberösterreich: Linz.
Hulme, P. E. (2009). Trade, transport and trouble: Managing invasivespecies pathways in an era of globalization. Journal of AppliedEcology, 46, 4–10.
Hulme, P. E., Bacher, S., Kenis, M., Klotz, S., Kühn, I., Minchin,D., et al. (2008). Grasping at the routes of biological invasions:A framework for integrating pathways into policy. Journal ofApplied Ecology, 45, 403–414.
Hulme, P. E., Pysek, P., Nentwig, W., & Vilá, M. (2009). Will threatof biological invasions unite the European Union? Science, 324,40–41.
Kleinbauer, I., Dullinger, S., Peterseil, J., & Essl, F. (2009). Climatechange might drive the invasive tree Robinia pseudacacia intonature reserves and endangered habitats. Biological Conserva-tion, 143, 382–390.
Kowarik, I. (1992). Einführung und Ausbreitung nichtheimischerGehölzarten in Berlin und Brandenburg und ihre Folgen für Floraund Vegetation. Verhandlungen des botanischen Vereins BerlinBrandenburg, Beiheft, 3, 1–180.
Kowarik, I. (1995). Time lags in biological invasions with regard tothe success and failure of alien species. In P. Pysek, K. Prach, M.Rejmánek, & M. Wade (Eds.), Plant Invasions, general aspectsand special problems (pp. 15–38). Amsterdam: SPB AcademicPublishers.
Kowarik, I. (2003a). Biologische Invasionen: Neophyten und Neo-zoen in Mitteleuropa. Stuttgart: E. Ulmer Verlag.
Kowarik, I. (2003b). Human agency in biological invasions: Sec-ondary releases foster naturalisation and population expansionof alien plant species. Biological Invasions, 5, 281–300.
Kowarik, I., & Säumel, I. (2007). Biological flora of Central Europe:Ailanthus altissima (Mill.) Swingle. Perspectives in Plant Ecol-ogy, Evolution and Systematics, 8, 207–237.
Krivánek, M., Pysek, P., & Jarosik, V. (2006). Planting history andpropagule pressure as predictors of invasion by woody speciesin a temperate region. Conservation Biology, 20, 1487–1498.
Lambdon, P. W., Pysek, P., Basnou, C., Hejda, M., Arianoutsou,M., Essl, F., et al. (2008). Alien flora of Europe: Species diver-sity, temporal trends, geographical patterns and research needs.Preslia, 80, 101–149.
Lockwood, J. L., Cassey, P., & Blackburn, T. (2005). The role ofpropagule pressure in explaining species invasions. Trends inEcology and Evolution, 20, 223–228.
McKinney, M. L. (2001). Effects of human population, area, andtime on non-native plant and fish diversity in the United States.Biological Conservation, 100, 243–252.
Mitchell, B. R. (2003). International historical statistics: Europe,1750–1993. New York: Stockton Press.
Pysek, P., Kucera, T., & Jarosik, V. (2002). Plant species richnessof nature reserves: The interplay of area, climate and habitat in acentral European landscape. Global Ecology and Biogeography,11, 279–289.
Pysek, P., & Jarosik, V. (2005). Residence time determinesthe distribution of alien plants. In I. Inderjit (Ed.), Invasiveplants: Ecological and agricultural aspects (pp. 77–96). Basel:Birkhäuser.
Pysek, P., & Richardson, D. M. (2007). Traits associated with inva-siveness: Where do we stand? In W. Nentwig (Ed.), Biologicalinvasions. Ecological studies (pp. 97–122). Berlin: Springer.
Pysek, P., Krivánek, M., & Jarosik, V. (2009). Planting intensity,residence time, and species traits determine invasion success ofalien woody species. Ecology, 90, 2734–2744.
R Development Core Team. (2009). R: A language and environ-ment for statistical computing. Vienna, Austria: R Foundationfor Statistical Computing. URL: http://www.R-project.org
Rabitsch, W., & Essl, F. (2006). Biological invasions in Aus-tria: Patterns and case studies. Biological Invasions, 8,295–308.
Richardson, D. M., Pysek, P., Rejmánek, M., Barbour, M. G.,Panetta, F. D., & West, C. J. (2000). Naturalisation and inva-

Author's personal copy
F. Essl et al. / Basic and Applied Ecology 12 (2011) 1–9 9
sion of alien plants: Concepts and definitions. Diversity andDistributions, 6, 93–107.
Simberloff, D., & Von Holle, B. (1999). Positive interactions of non-indigenous species: Invasional meltdown? Biological Invasions,1, 21–32.
Svenning, J. -C., & Skov, F. (2004). Limited filling of the potentialrange in European tree species. Ecology Letters, 7, 565–573.
Svenning, J. -C., & Skov, F. (2007). Could the tree diversity pattern inEurope be generated by postglacial dispersal limitation? EcologyLetters, 10, 453–460.
Taylor, B. W., & Irwin, R. E. (2004). Linking economic activities tothe distribution of exotic plants. Proceedings of the NationalAcademy of Sciences of the United States of America, 101,17725–17730.
Vilà, M., Basnou, C., Pysek, P., Josefsson, M., Genovesi, P.,Gollasch, S., et al. (2009). How well do we understand theimpacts of alien species on ecosystem services? A pan-European
cross-taxa assessment. Frontiers in Ecology and the Environ-ment, 8, 135–144.
Walter, J., Essl, F., Englisch, T., & Kiehn, M. (2005). Neophytes inAustria: Habitat preferences and ecological effects. Neobiota, 6,13–25.
Walther, G. -R. (2002). Weakening of climatic constraints withglobal warming and its consequences for evergreen broad-leavedspecies. Folia Geobotanica, 37, 129–139.
Walther, G. -R., Roques, A., Hulme, P. E., Sykes, M. T., Pysek, P.,Kühn, I., et al. (2009). Alien species in a warmer world: Risks andopportunities. Trends in Ecology and Evolution, 24, 686–693.
Wiesbauer, H., & Mazucco, K. (1997). Dünen in Niederösterreich.Fachberichte des NÖ Landschaftsfonds, 6, 91.
Williamson, M., Dehnen-Schmutz, K., Kühn, I., Hill, M., Klotz,S., Milbau, A., et al. (2009). The distribution of range sizes ofnative and alien plants in four European countries and the effectsof residence time. Diversity and Distributions, 15, 158–166.





























