Embed Size (px)
Citation preview

Pinocchio, a Novel Protein Expressed in the Antenna,Contributes to Olfactory Behaviorin Drosophila melanogaster
Stephanie M. Rollmann,1,3 Trudy F. C. Mackay,2,3 Robert R. H. Anholt1,2,3
1 Department of Zoology, North Carolina State University, Campus Box 7617, Raleigh,North Carolina 27695-7617
2 Department of Genetics, North Carolina State University, Campus Box 7617, Raleigh,North Carolina 27695-7617
3 W. M. Keck Center for Behavioral Biology, North Carolina State University,Campus Box 7617, Raleigh, North Carolina 27695-7617
Received 1 September 2004; accepted 3 October 2004
ABSTRACT: Most organisms depend on chemore-
ception for survival and reproduction. In Drosophila mel-anogaster multigene families of chemosensory receptors
and putative odorant binding proteins have been identi-
fied. Here, we introduce an additional distinct protein,
encoded by the CG4710 gene, that contributes to olfac-
tory behavior. Previously, we identified through P[lArB]-
element mutagenesis a smell impaired (smi) mutant,
smi21F, with odorant-specific defects in avoidance
responses. Here, we show that the smi21F mutant also
exhibits reduced attractant responses to some, but not all,
of a select group of odorants. Furthermore, electroanten-
nogram amplitudes are increased in smi21F flies. Charac-
terization of flanking sequences of the P[lArB] insertionsite, complementation mapping, phenotypic reversion
through P-element excision, and expression analysis
implicate a predicted gene, CG4710, as the candidate smi
gene. CG4710 produces two transcripts that encode pro-
teins that contain conserved cysteines and which are
reduced in the smi21F mutant. Furthermore, in situ
hybridization reveals CG4710 expression in the third
antennal segment. We have named this gene of
previously unknown function and its product ‘‘Pinocchio
(Pino)’’. ' 2005 Wiley Periodicals, Inc. J Neurobiol 63: 146–158, 2005
Keywords: olfaction; P-element mutagenesis; behavioral
genetics; chemoreception
INTRODUCTION
Chemoreception is the principle modality by which
many organisms gain information from their external
environment and is critical for an organism’s survival
and for modulating social interactions. In recent years
significant progress has been made in our understand-
ing of the molecular mechanisms underlying odor
recognition, but the link between odor perception and
odor-guided behavior remains poorly understood.
With its sequenced genome and well developed
genetic technology, Drosophila melanogaster pro-
vides a powerful model system to investigate how
ensembles of genes mediate odor-guided behavior.
Insect olfactory systems are remarkably similar in
functional organization to those of vertebrates in that
peripheral chemosensory neurons converge on a few
output neurons in distinct spherical structures of neu-
ropil called ‘‘glomeruli’’ (reviewed by Hildebrand
Correspondence to: R. R. H. Anholt ([email protected]).Contract grant sponsor: National Institutes of Health; contract
grant numbers: GM059469, GM45344, and GM45146.Contract grant sponsor: W. M. Keck Foundation.
# 2005 Wiley Periodicals, Inc.Published online 17 February 2005 in Wiley InterScience (www.interscience.wiley.com).DOI 10.1002/neu.20123
146

and Shepherd, 1997). Insect olfactory systems, how-
ever, are quantitatively less complex than their
vertebrate counterparts. The olfactory system of
D. melanogaster contains only about 1200 chemosen-
sory neurons located in the third antennal segments
and the maxillary palps (reviewed by Stocker, 1994).
Chemosensory neurons from the antennae and maxil-
lary palps project to 43 identified glomeruli in each
antennal lobe (Laissue et al., 1999; Vosshall et al.,
2000; Gao et al., 2000), a small number compared to
the 1800 glomeruli in the mouse olfactory bulb
(Pomeroy et al., 1990).
D. melanogaster was the first insect species in
which families of olfactory receptors (Clyne et al.,
1999; Vosshall et al., 1999; Gao and Chess, 1999)
and gustatory receptors (Scott et al., 2001; Dunipace
et al., 2001) were identified. The genome of D. mela-nogaster contains approximately 60 putative olfac-
tory receptors (Rubin, 2000), similar to the number of
glomeruli in each antennal lobe (Laissue et al., 1999).
Odorant receptors have distinct expression patterns in
the antennae and/or maxillary palps, although one
receptor, Or83b, is expressed ubiquitously (Vosshall
et al., 1999, 2000) and is essential for transporting
other odorant receptors to the dendritic membranes
(Larsson et al., 2004). Electrophysiological studies
demonstrated that olfactory neurons in both the max-
illary palps and the third antennal segments are com-
partmentalized according to strict pairing rules so
that the same pairs of neurons with distinct odorant
specificities are always housed together in the same
sensilla (de Bruyne et al., 1999, 2001). Olfactory neu-
rons expressing the same odorant receptor project
bilaterally to one, or sometimes two, glomeruli in
each antennal lobe (Vosshall et al., 2000; Gao et al.,
2000), displaying a similar convergence to that
observed in the vertebrate olfactory system (Ressler
et al., 1994; Vassar et al., 1994; Mombaerts et al.,
1996). Systematic single cell tracing experiments
have characterized individual projection neurons
from defined glomeruli in the antennal lobe to higher
brain regions, the mushroom body and lateral horn of
the protocerebrum (Wong et al., 2002; Marin et al.,
2002). Elegant electrophysiological studies, in which
odorant receptors were misexpressed in sensilla con-
taining neurons that lacked their endogenous odorant
receptor, have identified the response spectra of a
wide array of Drosophila odorant receptors (Hallem
et al., 2004).
In addition to families of chemosensory receptors,
a large family of odorant binding proteins (OBPs) is
also differentially expressed (Hekmat-Scafe et al.,
1997, 2002; Park et al., 2000; Galindo and Smith,
2001; Shanbhag et al., 2001; Vogt et al., 2002). OBPs
are secreted into the sensillar perilymph and,
although their precise function has not been conclu-
sively established, a prevailing hypothesis is that they
facilitate transport of hydrophobic odorants through
the perilymph to receptors at the chemosensory mem-
brane (Kaissling, 1996; Du and Prestwich, 1995;
Robertson et al., 1999; Vogt et al., 1999). OBPs form
a structurally diverse family of proteins characterized
by six cysteines at conserved positions (Vogt et al.,
1991). Crystallographic studies on the pheromone
binding protein of the silk moth Bombyx mori suggestthat the cysteines stabilize a binding pocket that
undergoes pH-dependent conformational changes for
the binding and release of the pheromone, bombykol
(Leal et al., 1999; Wojtasek and Leal, 1999; Sandler
et al., 2000; Damberger et al., 2000; Leal, 2000;
Horst et al., 2001). In Drosophila, an OBP null
mutant, lush, showed impaired avoidance responses
to ethanol and propanol (Kim et al., 1998; Kim and
Smith, 2001), and the crystal structure of the alcohol
binding site of the Lush protein has been described
(Kruse et al., 2003). The functional relationships
between OBPs and chemosensory receptors and phys-
iological responsiveness of identified olfactory neu-
rons in the antennae and the maxillary palps,
however, remain to be clarified. Here, we introduce
an additional protein, encoded by the CG4710 gene,
which is distinct from the classic OBP family, but
also contributes to olfactory behavior.
MATERIALS AND METHODS
Fly Stocks
The smi21F line has a single P[lArB]-element insertion at
cytological position 21F in the isogenic Samarkand (Sam;ry506) background (Anholt et al., 1996). P-element excision
lines of smi21F were generated in the Samarkand back-
ground, as described below. Deficiency line, Df(2L)ast2/SM1, was obtained from the Bloomington Drosophila StockCenter (Bloomington, IN). All flies were reared on an agar-
yeast-molasses medium in vials maintained at 258C and
under a 12 h light/dark cycle.
Behavioral Assays
Avoidance Assay. Flies, 5–7 days posteclosion, were
removed from their food source 1.5–2 h prior to assay.
Avoidance responses to 0.3% (v/v) benzaldehyde were con-
ducted in an environmental chamber at 258C and 70%
humidity as described by Anholt et al. (1996). Briefly, one
replicate assay consisted of a single sex group of five indi-
viduals. Test vials were divided into two compartments by
placing a mark on the wall 3 cm from the bottom of the
Pinocchio Contributes to Olfaction 147

vial. The odorant was introduced on a cotton swab wedged
between the plug and the wall of the vial and the number of
flies present in the compartment remote from the odor
source was measured at 5 s intervals for 60 s. Measure-
ments began 15 s after the introduction of odorant. The
‘‘avoidance score’’ for the replicate is the average of the 10
counts, with a score of 5 indicating a maximal repellent
response. Replicate assays of each line were conducted.
Statistically significant deviations from wild-type behavior
were evaluated by ANOVA.
Attraction Assay. Olfactory trap assays were conducted
essentially as described by Woodard et al. (1989). Ten flies
of a single sex were placed in a Petri dish (100 � 15 mm)
containing 10 mL of 1% agarose and a trap constructed
from a 1.5 mL centrifuge tube and a 200 �L wide orifice
pipet tip. The tube was cut approximately 3 mm from its
end and the inverted pipette tip inserted into the tube. Three
hundred microliter of odorant in 1% agarose was applied to
the inside of the tube cap. Odorants tested were obtained
from Aldrich Chemical Co. (Milwaukee, WI) and included
acetic acid, 1-pentanol, acetone, helional, citralva, 1-buta-
nol, geraniol, cyclohexanone, amyl acetate, propanol, 3-
octanol, p-cresol, E2-hexenal, propionic acid, acetaldehyde,acetal, and ethyl propionate. After 48 h the number of flies
trapped was recorded. The ‘‘attraction score’’ is the average
number of flies trapped among replicates. Comparisons
between lines were made by three-way factorial fixed
effects ANOVA using SAS Proc GLM (SAS Institute Inc.,
Cary, NC) and posthoc Tukey tests for significant differen-
ces among means.
Electrophysiological Recordings
Electroantennogram (EAG) recordings were obtained from
Sam; ry506 (n ¼ 10) and smi21F (n ¼ 10) Drosophila 3–5
days posteclosion. Individual flies were immobilized in a
pipette tip with the anterior portion of the head protruding
from the tip. Ground and recording glass capillary electro-
des (1.5 mm outer diameter) were filled with Drosophilasaline (188 mM NaCl, 5 mM KCl, 2 mM CaCl2�2H2O). The
ground electrode was subsequently placed into the head
capsule and the recording electrode onto the anterior distal
region of the third antennal segment. A constant air stream
of �1.4 L/min flowed continuously over the antenna.
Pulses of odor consisted of 3 mL of air from a 5 cc syringe
rapidly puffed over a filter disk saturated with the diluted
odorant and then directly introduced into the air stream.
Odorants were diluted in paraffin oil (Fluka, Buchs, Swit-
zerland) and tested at three different concentrations. Each
sample was puffed two times and EAG peak amplitudes
were measured and averaged. Data were acquired using a
Syntech Intelligent Data Acquisition Controller IDAC-4-
USB (Hilversum, The Netherlands) and recording, storing,
and quantifying EAG peak amplitudes was carried out with
EAG2000 version 2.7 software (Syntech). Comparisons of
peak amplitudes between lines were evaluated by three-way
factorial ANOVA.
Quantitative Complementation Test
Deficiency line, Df(2L)ast2/SM1, which contains a deletion
over 21D1-2 to 22B2-3, was crossed to smi21F and Sam;ry506, and olfactory avoidance behavior of the F1 progeny ofeach cross assessed (Anholt et al., 1996). The data were ana-
lyzed by two-way factorial fixed effects ANOVA and quan-
titative failure to complement was inferred if the line (L) or
line by sex (S) interaction terms were significant according
to the model y¼ � þ Lþ Sþ L� Sþ E, where E indicates
error (Pasyukova et al., 2000). Significant differences
among means were determined by a posthoc Tukey test.
Phenotypic Reversion throughP-Element Excision
The P[lArB] construct was mobilized by crossing smi21Ffemales to w; Cy/Sp; SbD2-3/TM6,Tb males. Male offspring
of the genotype Sam; Cy/smiP21F; SbD2-3/ry506 were then
crossed to Sam; Cy/Sp; ry506 females and single male off-
spring in which the P-element was excised, Sam; Cy/smiP�;ry506, were again crossed to Sam; Cy/Sp; ry506 females. Male
and female Sam; Cy/smiP�; ry506 were subsequently mated
to make a homozygous P-element excision line in the iso-
genic Samarkand background. The precise excision of the
construct was verified using Southern blots of HindIII-digested genomic DNA from Sam; ry506, smi21F and P-ele-ment excision lines hybridized to a plasmid-rescued genomic
DNA fragment flanking the P-element insertion site in
smi21F. Avoidance responses to benzaldehyde of P-element
excision lines were assessed and data were analyzed using a
single classification fixed effects ANOVA according to the
model y ¼ � þ L þ E, where L designates line and E error.
mRNA Quantification
Levels of mRNA were quantified in Sam; ry506 and smi21Flines using real-time quantitative PCR. Total RNA was iso-
lated from male and female Drosophila heads using Trizol
(Gibco BRL, Gaithersburg, MD). Three replicate RNA iso-
lations were prepared for each line. The RNA was purified
by DNase treatment followed by a phenol and chloroform
extraction. cDNA was generated from 5 �g of total RNA by
reverse transcription (Stratagene, La Jolla, CA) and tran-
script-specific primers were designed by Primer Express
software (PE Applied Biosystems, Foster City, CA).
Primers and probes are as follows: transcript I, 50-CCA-CCGACAAGCTGAACTTCA-30, 50-GCAGACAGACCG-AGATTGTG-30, 6FAM-TCAGCAGCCAATATGTCG-
ATCGCCA-TAMRA; transcript II, 50-GCAGCACAG-GAATCCCAATT-30, 50-TGGCGATCGACATATTGGC-30, 6FAM-TGCTGTAAATAACCCAACAAATTGCCAT-
CAG-TAMRA. Ribosomal protein 49 (rp49) was used as
an endogenous control. Sequences for rp49 primers and
Taqman probe were: 50-CAAGAAGTTCCTGGTGCA-CAA-30, 50-AGTAAACGCGGTTCTGCATGA-30, and
6FAM-TGCGCGAGCTGGAGGTCCTG-TAMRA. The
mRNA levels were measured using the GeneAmp 5700
148 Rollmann et al.

system (PE Applied Biosystems) and the relative standard
curve method. Standard curves for rp49, CG4552, and
CG4710 transcripts I and II were constructed using dilu-
tions of cDNA from Sam; ry506. Each of the cDNA samples
was amplified in duplicate and expression levels determined
relative to the Sam; ry506 standard curve. Differences in
expression between genes in the vicinity of the P[lArB]insertion site and differences in gene expression between
smi21F and the host strain were determined by two-way
factorial ANOVA and posthoc Tukey tests, where applica-
ble. Expression of both transcripts of CG4710 was further
verified by complete sequencing of full length amplicons
corresponding to their coding regions.
In Situ Hybridization
mRNA expression patterns of the smi21F gene were deter-
mined by in situ hybridization to 10 � formalin-fixed and
paraffin-embedded sections of randomly oriented Sam;ry506 heads (Vosshall et al., 2000, with minor modification).
PCR-amplified DNA fragments corresponding to unique
portions of the coding region for each CG4710 transcript
(Fig. 3) and the neighboring gene, CG4552, were cloned
into the TA dual promotor cloning vector (Invitrogen,
Carlsbad, CA), and, following linearization of the vector,
digoxigenin-labeled antisense and sense riboprobes were
generated using SP6 and T7 RNA polymerases, respec-
tively. After deparaffinization and rehydration through
graded alcohols, slides were placed in 4% paraformalde-
hyde and subsequently washed with phosphate buffered sal-
ine (PBS). Sections were then incubated with 5 �g/mL
proteinase K for 30 min at 378C, followed by treatment
with acetic anhydride. The sections were then hybridized
for 16 h at 558C in hybridization buffer (50% formamide,
5X SSC, 5X Denhardts, 50 �g/mL yeast tRNA, 50 �g/mL
heparin, 500 �g/mL sheared salmon sperm DNA, 2.5 mMEDTA, 0.25% CHAPS, and 0.1% Tween 20) with either a
Figure 1 Behavioral responses of smi21F (white bars) and wild-type flies (Sam; ry506, blackbars) to odorants. Flies of the smi21F line show sex-specific reductions in olfactory responses to
acetic acid and propionic acid. (A) Responses of females to acetic acid, and (B) responses of males
to propionic acid. For each line, 10 (A) or 15 (B) replicates were measured for each of four odorant
concentrations.
Pinocchio Contributes to Olfaction 149

heat denatured antisense or sense probe. After hybridiza-
tion, the slides were washed at 558C with 5X SSC followed
by 20 min sequential washes at 558C with 0.2X SSC and
PBS, 0.1% Triton X-100. Sections were then blocked with
heat inactivated normal goat serum in PBS, 0.1% Triton
X-100. Hybridization products were visualized with an
alkaline phosphatase-conjugated antidigoxigenin antibody
(Roche Molecular Biochemicals, Indianapolis, IN), using
nitro blue tetrazolium chloride and 5-bromo-4-chloro-
3-indolyl-phosphate as substrates. Images were digitally
captured and processed with Adobe Photoshop.
RESULTS
Phenotypic Characterization of thesmi21FMutant
Previously, we identified through P-element inser-
tional mutagenesis a smell impaired (smi) mutant,
smi21F, that shows odorant-specific defects in olfac-
tory avoidance behavior (Anholt et al., 1996). Aber-
rant behavioral responses were observed only in
females to benzaldehyde and 2-isobutylthiazole at
high concentrations, but not to 2-n-propylpyrazine.To further examine the specificity of the smell
impairment in this mutant, we asked whether smi21Fflies also show aberrant attractant responses to odor-
ants. Using an olfactory trap assay, we first examined
responses of the P-element free host strain, Sam;ry506, to a series of odorants, used previously by other
investigators (de Bruyne et al., 2001) and represent-
ing different structural classes, including acetic acid,
1-pentanol, acetone, helional, citralva, 1-butanol,
geraniol, cyclohexanone, amyl acetate, propanol, 3-
octanol, p-cresol, E2-hexenal, propionic acid, acetal-
dehyde, acetal, and ethyl propionate. Under our
experimental conditions [a concentration of 1% (v/v)
of each odorant] Sam; ry506 flies responded to only
five of the odorants tested, namely acetic acid, acetal-
dehyde, acetal, propionic acid, and ethyl propionate.
Although other odorants might have elicited
responses at different odorant concentrations, we
decided to focus our experiments on these five odor-
ants and generated dose-response curves for each of
them. Dose-response curves showed an increase in
attractant behavior at low odorant concentrations and
a decline at higher concentrations. We compared
attractant responses of Sam; ry506 and smi21F flies at
a range of concentrations optimal for resolving differ-
ences in attractant responses between wild-type and
mutant flies (Fig. 1). No statistically significant dif-
ferences between responses of smi21F and their wild-
type controls to ethyl propionate, acetaldehyde, and
acetal were observed (data not shown). However,
smi21F flies showed marked sex-specific reductions
in responsiveness to acetic acid and propionic acid
(Fig. 1; Table 1). The response to acetic acid was
reduced in females [Fig. 1(A); Table 1]. An apparent
shift of the dose-response toward higher stimulus
concentrations of acetic acid in the smi21F mutant is
evident from a significant difference at the intermedi-
ate concentration of 0.3% (v/v) acetic acid (p <0.0001). No statistically significant differences were
resolved at lower concentrations near threshold or at
saturating concentrations.
We also observed impaired olfactory behavior
with propionic acid [Fig. 1(B); Table 1]. In contrast
to the response to acetic acid, males, but not females,
are smell impaired at high concentrations of pro-
pionic acid, reminiscent of sexually dimorphic olfac-
tory defects observed previously for avoidance
responses to benzaldehyde and 2-isobutylthiazole
(Anholt et al., 1996). A nominally significant reduc-
tion in female response to propionic acid was also
observed at intermediate concentrations, but was not
significant after Bonferroni correction.
Table 1 Three-Way Factorial ANOVA of Behavioral Response of smi21F and Wild-Type Files to Acetic Acid and
Propionic Acidy
Acetic Acid Propionic Acid
Mean square F Value p Value Mean square F Value p Value
Line 2.3 9.11 * 5.5 17.2 ****
Sex 1.76 6.99 * 1.83 5.71 *
Treatment 9.21 36.49 *** 10.71 33.53 ****
L � S 0.001 0 ns 0.1 0.31 ns
L � T 0.79 3.12 ns 1.89 5.93 ***
S � T 0.37 1.45 ns 0.16 0.51 ns
L � S � T 0.12 0.46 ns 1.17 3.66 **
yThe model used was y ¼ � þ L þ T þ S þ L � T þ L � S þ L � T � S þ E with the three main fixed effects being line (L), treatment
(T, concentration of odorant), and sex (S), and where E indicates error. p values of <0.05, <0.01, <0.001, and <0.0001 are indicated by *, **,
***, and ****, respectively; ns, not significant.
150 Rollmann et al.
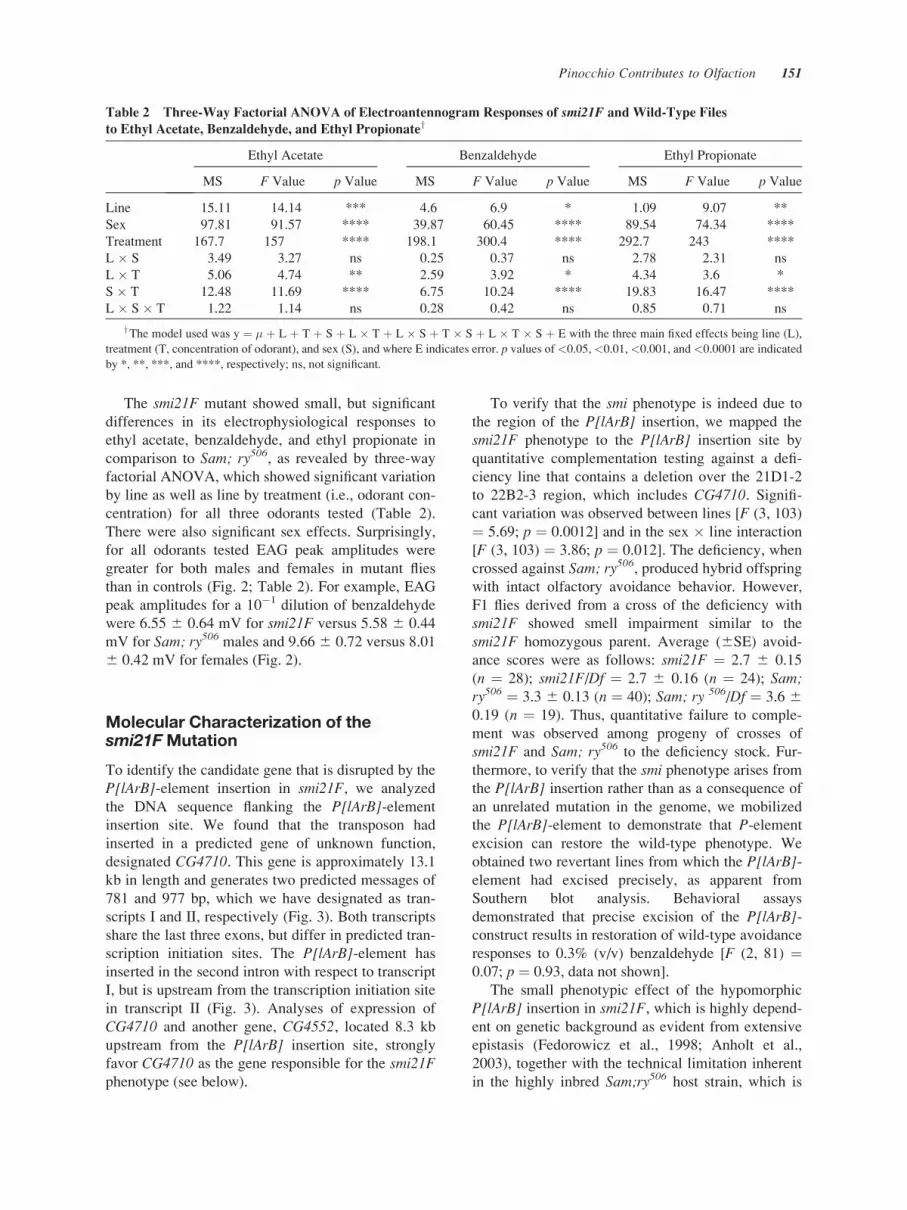
The smi21F mutant showed small, but significant
differences in its electrophysiological responses to
ethyl acetate, benzaldehyde, and ethyl propionate in
comparison to Sam; ry506, as revealed by three-way
factorial ANOVA, which showed significant variation
by line as well as line by treatment (i.e., odorant con-
centration) for all three odorants tested (Table 2).
There were also significant sex effects. Surprisingly,
for all odorants tested EAG peak amplitudes were
greater for both males and females in mutant flies
than in controls (Fig. 2; Table 2). For example, EAG
peak amplitudes for a 10�1 dilution of benzaldehyde
were 6.55 6 0.64 mV for smi21F versus 5.58 6 0.44
mV for Sam; ry506 males and 9.66 6 0.72 versus 8.01
6 0.42 mV for females (Fig. 2).
Molecular Characterization of thesmi21FMutation
To identify the candidate gene that is disrupted by the
P[lArB]-element insertion in smi21F, we analyzed
the DNA sequence flanking the P[lArB]-element
insertion site. We found that the transposon had
inserted in a predicted gene of unknown function,
designated CG4710. This gene is approximately 13.1
kb in length and generates two predicted messages of
781 and 977 bp, which we have designated as tran-
scripts I and II, respectively (Fig. 3). Both transcripts
share the last three exons, but differ in predicted tran-
scription initiation sites. The P[lArB]-element has
inserted in the second intron with respect to transcript
I, but is upstream from the transcription initiation site
in transcript II (Fig. 3). Analyses of expression of
CG4710 and another gene, CG4552, located 8.3 kb
upstream from the P[lArB] insertion site, strongly
favor CG4710 as the gene responsible for the smi21Fphenotype (see below).
To verify that the smi phenotype is indeed due to
the region of the P[lArB] insertion, we mapped the
smi21F phenotype to the P[lArB] insertion site by
quantitative complementation testing against a defi-
ciency line that contains a deletion over the 21D1-2
to 22B2-3 region, which includes CG4710. Signifi-cant variation was observed between lines [F (3, 103)
¼ 5.69; p ¼ 0.0012] and in the sex � line interaction
[F (3, 103) ¼ 3.86; p ¼ 0.012]. The deficiency, when
crossed against Sam; ry506, produced hybrid offspring
with intact olfactory avoidance behavior. However,
F1 flies derived from a cross of the deficiency with
smi21F showed smell impairment similar to the
smi21F homozygous parent. Average (6SE) avoid-
ance scores were as follows: smi21F ¼ 2.7 6 0.15
(n ¼ 28); smi21F/Df ¼ 2.7 6 0.16 (n ¼ 24); Sam;ry506 ¼ 3.3 6 0.13 (n ¼ 40); Sam; ry 506/Df ¼ 3.6 60.19 (n ¼ 19). Thus, quantitative failure to comple-
ment was observed among progeny of crosses of
smi21F and Sam; ry506 to the deficiency stock. Fur-
thermore, to verify that the smi phenotype arises fromthe P[lArB] insertion rather than as a consequence of
an unrelated mutation in the genome, we mobilized
the P[lArB]-element to demonstrate that P-element
excision can restore the wild-type phenotype. We
obtained two revertant lines from which the P[lArB]-element had excised precisely, as apparent from
Southern blot analysis. Behavioral assays
demonstrated that precise excision of the P[lArB]-construct results in restoration of wild-type avoidance
responses to 0.3% (v/v) benzaldehyde [F (2, 81) ¼0.07; p ¼ 0.93, data not shown].
The small phenotypic effect of the hypomorphic
P[lArB] insertion in smi21F, which is highly depend-
ent on genetic background as evident from extensive
epistasis (Fedorowicz et al., 1998; Anholt et al.,
2003), together with the technical limitation inherent
in the highly inbred Sam;ry506 host strain, which is
Table 2 Three-Way Factorial ANOVA of Electroantennogram Responses of smi21F and Wild-Type Files
to Ethyl Acetate, Benzaldehyde, and Ethyl Propionatey
Ethyl Acetate Benzaldehyde Ethyl Propionate
MS F Value p Value MS F Value p Value MS F Value p Value
Line 15.11 14.14 *** 4.6 6.9 * 1.09 9.07 **
Sex 97.81 91.57 **** 39.87 60.45 **** 89.54 74.34 ****
Treatment 167.7 157 **** 198.1 300.4 **** 292.7 243 ****
L � S 3.49 3.27 ns 0.25 0.37 ns 2.78 2.31 ns
L � T 5.06 4.74 ** 2.59 3.92 * 4.34 3.6 *
S � T 12.48 11.69 **** 6.75 10.24 **** 19.83 16.47 ****
L � S � T 1.22 1.14 ns 0.28 0.42 ns 0.85 0.71 ns
yThe model used was y ¼ � þ L þ T þ S þ L � T þ L � S þ T � S þ L � T � S þ E with the three main fixed effects being line (L),
treatment (T, concentration of odorant), and sex (S), and where E indicates error. p values of <0.05, <0.01, <0.001, and <0.0001 are indicated
by *, **, ***, and ****, respectively; ns, not significant.
Pinocchio Contributes to Olfaction 151

not robust enough for transformation, precluded phe-
notypic rescue of the smi21F mutant phenotype by
introduction of a wild-type copy of the CG4710 gene.
Accumulation of deleterious mutations during more
than a decade of inbreeding has rendered the isogenic
Sam;ry506 host strain unsuitable for introduction of
transgenic constructs, because embryos of this strain
do not survive the microinjection procedure. Further-
more, the phenotypic effect of the smi21F mutation is
within the standard error of natural phenotypic varia-
Figure 2 EAG responses in the smi21F mutant and the Sam; ry506 control. (A) Typical EAG
responses of smi21F and Sam; ry506 flies illustrate the increased peak amplitudes in response to the
delivery of a 10�1 dilution of benzaldehyde to the antenna. The scale bars designate 5 mV (ordi-
nate) and 2 s (abscissa). (B,C) EAG amplitudes of male (B) and female (C) Sam; ry506 (gray bars)
and smi21F (black bars) to ethyl acetate, benzaldehyde, and ethyl propionate at dilutions of 10�5,
10�3, and 10�1. Data were analyzed by three-way factorial ANOVA, which revealed significant
variation by line and line � treatment (Table 2).
152 Rollmann et al.

tion (Mackay et al., 1996) and can only be resolved
under controlled genetic background conditions, pre-
cluding the introduction of a transgene by crossing it
from a different genetic background into the mutant
stock. Despite this limitation, the position of the P-element, expression analyses described below, and
reduction of the CG4710 transcript observed in
microarray analyses Anholt et al. (2003) strongly
implicate CG4710 as the candidate gene responsible
for the smi21F mutant phenotype. We have named
this gene of previously unknown function and its
product Pinocchio (Pino).The Pino gene encodes a 249 amino acid polypep-
tide and a 171 amino acid polypeptide corresponding
to transcripts I and II, respectively (Fig. 3). Both pol-
ypeptides are identical, except for the N-terminus,
which is 78 amino acids longer in transcript I (Fig.
4). The Pinocchio proteins contain 12 cysteines, five
of which can be aligned with characteristic cysteines
at conserved positions of OBPs (Robertson et al.,
1999; Galindo and Smith, 2001), although the overall
sequence is vastly divergent from the OBP family.
We identified orthologues of Pinocchio in the
recently sequenced Anopheles gambiae (Holt et al.,
2002) and Apis mellifera genomes (Fig. 4). It is of
interest to note that Anopheles gambiae also
expresses two transcripts corresponding to the Droso-phila Pino transcripts I and II with �40% sequence
identity, including conservation of the cysteines.
Expression of Pino Transcripts
To provide evidence that the P[lArB]-element inser-
tion in Pinosmi21F reduces gene expression we quanti-
fied mRNA levels in heads of wild-type and
Pinosmi21F flies using real-time quantitative PCR.
Transcript I was reduced 287 6 128 (SE)-fold in the
Pinosmi21F mutant (n ¼ 6), whereas transcript II
showed a 2.57 6 0.28 (SE)-fold reduction (n ¼ 6). In
contrast, the CG4552 transcript showed a 0.77 60.06 (SE)-fold difference compared to the wild-type
(n ¼ 6). To assess statistical significance the data
were analyzed by analysis of variance (Table 3).
There was a significant reduction in expression of
Pino transcript I in Pinosmi21F compared to the wild-
Figure 3 Structure of the CG4710 gene. The P[lArB] insertion site and orientations of genes and
the P-element are indicated by arrowheads and arrows, respectively. The horizontal line represents
genomic DNA in the CG4710 region. Regions of the CG4710 gene that give rise to alternative tran-scripts I and II are schematically represented. Black boxes in these regions represent coding
regions. The numerals 1 and 2 inside the squares indicate regions specific for transcript I and II,
respectively, amplified for in situ hybridizations (Fig. 5) and QPCR (Table 3).
Pinocchio Contributes to Olfaction 153

type, while there was no significant difference in
expression of the endogenous control [Table 3(A)].
There were also significant differences in gene
expression levels when Pino transcript II, CG4552,and the endogenous control were analyzed [males,
Table 3(B), and females, Table 3(C), were analyzed
separately]. Posthoc Tukey tests (p < 0.05) indicate
that there were differences in expression in all three
genes among males with the greatest difference in
transcript II; in females, however, only differences in
expression of the Pino transcript II were significant.
These data support the notion that disruption of
Pinocchio accounts for the smell-impaired phenotype
of smi21F mutants.
If the proteins encoded by Pinocchio contribute to
olfaction, we expect Pinocchio to be expressed in
chemosensory tissues. Previously, we observed in the
Pinosmi21F mutant heterogeneous lacZ reporter gene
expression throughout the third antennal segment.
Reporter gene expression was widespread, but espe-
cially intense2 in the proximal region of the third
antennal segment (Anholt et al., 1996). In situ hybrid-
ization in Sam; ry506 flies with a riboprobe corre-
sponding to an exon of Pino transcript I reveals
extensive expression in the third antennal segments
[Fig. 5(A)], recapitulating the previously observed
lacZ reporter gene expression pattern (Anholt et al.,
1996). In contrast, we did not detect expression of
CG4552 in the third antennal segment, further corrob-
orating evidence that Pinocchio rather than the neigh-
boring gene is affected by the P[lArB]-insertion in the
smi21F mutant. We did not observe differences in
expression patterns between males and females. Stain-
ing is not observed with a sense riboprobe [Fig. 5(B)],
which shows that the staining observed with the anti-
sense probe is specific. Examination of sections
Figure 4 Amino acid sequence alignments of Pinocchio gene products with their orthologues in
the honey bee, Apis mellifera, and the malaria mosquito, Anopheles gambiae. Amino acid sequen-
ces of Pinocchio transcripts I and II were aligned with the partial sequence of the Apis melliferahmm485 protein and Anopheles gambiae XP_317770 and XP_317771 transcripts. Sequences were
aligned with the ClustalW program and alignments were refined by eye. Conserved amino acids
between Pinocchio and orthologues from the other two species are shown in red and conserved cys-
teines are shown in blue.
154 Rollmann et al.

through heads cut in various orientations and hybri-
dized to sense and antisense probes did not reveal
specific staining in the brain or in the proboscis.
DISCUSSION
Previous studies showed that the Pinosmi21F mutant
exhibits subtle odorant-specific defects in olfactory
avoidance behavior (Anholt et al., 1996). Subsequent
studies discovered that despite the relatively small
effect on olfactory behavior, the smi21F locus dis-
plays extensive epistasis with other co-isogenic smiloci (Fedorowicz et al., 1998), which led us to
hypothesize that the candidate smi gene at this locus
encodes a protein that may be required in early stages
of the process of odor recognition. Characterization
of flanking sequences of the P[lArB] insertion site,
complementation mapping, and phenotypic reversion
through P-element excision identified the CG4710gene, which we have named Pinocchio, as the candi-
date smi gene at the smi21F locus. Furthermore,
expression analysis using high-density oligonucleo-
tide microarrays showed reduction of CG4710expression (but not CG4552) in the Pinosmi21F geneticbackground (Anholt et al., 2003).
The smi21F flies show aberrant olfactory res-
ponses to benzaldehyde (Anholt et al., 1996), acetic
acid, and propionic acid (Fig. 1). We observed
increased EAG amplitudes in the smi21F mutant
upon application of benzaldehyde, ethyl acetate (used
as standard), and ethyl propionate (Fig. 2). We were
not able to test acids as they give rise to artifacts in
the EAG recordings. The increased EAG amplitude
to ethyl propionate was not correlated with an altera-
tion in the behavioral response to this odorant.
Although a strict correlation between the EAG and
expression of the behavior might not be a prioriexpected, our observations could reflect greater sensi-
tivity of the electrophysiological assay compared to
the olfactory trap assay in assessing the subtle pheno-
typic effects of this hypomorphic mutation.
Throughout our experiments we noticed concen-
tration-dependent sexual dimorphism in olfactory
impairments between Pinosmi21F males and females.
Previous studies demonstrated that the genetic archi-
tecture that underlies olfactory behavior is different
in males and females (Mackay et al., 1996; Anholt,
2004). Such sexual dimorphism can arise from differ-
ences in gene regulation in males and females or from
differences in epistatic interactions in each sex envi-
ronment (Anholt et al., 2003; Anholt, 2004; Anholt
and Mackay, 2004). Because we did not observe
overt differences in gene expression between males
and females and because epistatic interactions in
the olfactory subgenome are extensive and involve
Pinosmi21F, we favor the latter interpretation.Previously, we reported transcriptional profiles of
five coisogenic smi lines, including smi21F, and iden-
tified transregulated genes in each smi background
(Anholt et al., 2003). These studies showed extensive
transregulation of antibacterial peptides, associated
with immune defense, and biotransformation
enzymes uniquely in the Pinosmi21F genetic back-
ground (Anholt et al., 2003). In contrast, only a single
OBP transcript, Obp8a, was up-regulated in the
Pinosmi21F genetic background. These observations
and the ubiquitous expression of Pinocchio through-
out the third antennal segment, in contrast to most
conventional OBPs (Galindo and Smith, 2001), lead
us to speculate that Pinocchio may contribute to the
removal of xenobiotics, including odorants and bacte-
rial toxins, rather than odorant recognition. This
hypothesis would be in line with our observation that
EAG amplitudes are increased in the hypomorphic
Pinosmi21F mutant, possibly as a consequence of
slower odorant removal.
Pinocchio is widely expressed throughout the third
antennal segment. In addition, Pino mRNAs have also
been identified in embryonic cDNA libraries and in
adult testis. The functional significance of transcription
in these tissues remains to be evaluated. Transcrip-
Table 3 Two-Way Factorial ANOVA of pinosmi21F
Expression Levels
Mean square F Value p Value
A. pino transcript I expression levels between
smi21F and wild-type filesy
Line 457375 11.73 **
Sex 180415 4.63 ns
L � S 173794 4.46 ns
B. Male differences in expression among pinotranscript II, CG4552, and rp49{
Gene 2.33 50.77 ***
Isolation 0.03 0.63 ns
G � I 0.05 1.15 ns
C. Female differences in expression among pinotranscript II, CG4552, and rp49{
Gene 1.3 16.08 ***
Isolation 0.22 2.72 ns
G � I 0.02 0.2 ns
yNo significant difference between lines was observed for rp49.
The model used was y ¼ � þ L þ S þ L � S þ E with the two
main fixed effects being line (L) and sex (S), and{y ¼ � þ G þ I þ G � I þ E with two main fixed effects being
gene (G) and RNA isolation (I), where E indicates error. p values of<0.01 and <0.001 are indicated by ** and ***, respectively; ns,
not significant.
Pinocchio Contributes to Olfaction 155

tional profiling studies suggest that Pinocchio may
contribute both to odorant binding and immune
defense (Anholt et al., 2003). Considering the ubiqui-
tous expression of Pino in the antenna, the odorant-
specific effects in the Pinosmi21F mutant are intriguing.
It is tempting to speculate that additional proteins in
the perilymph, possibly OBPs, may functionally inter-
act with Pinocchio. Galindo and Smith (2001) showed
differential expression of GFP-reporter genes driven
by different OBP promoters in both olfactory and
gustatory organs of Drosophila with some OBPs
showing widespread expression and others confined
to only a few cells. It is of interest to note that expres-
sion of Obp8a is up-regulated in the Pinosmi21F
genetic background (Anholt et al., 2003). A detailed
characterization of putative interactions of Pinocchio
with odorants and other components of the chemo-
sensory system in Drosophila awaits further studies.
We thank Paul Gilligan III, Nalini Kulkarni, and Monica
Mattmuller for excellent technical assistance and Yehuda
Ben-Shahar for providing sequence information for design-
ing the rp49 probes. We thank Dr. John Carlson and Rick-
ard Ignell for introducing us to EAG recording procedures
and Dr. Coby Schal for making his electrophysiological
equipment available. S. M. Rollmann is the recipient of a
Keck postdoctoral fellowship and a postdoctoral National
Research Service Award (GM20897).
REFERENCES
Anholt RRH. 2004. Genetic modules and networks for
behavior: Lessons from Drosophila. BioEssays 26:
1299–1306.
Anholt RRH, Dilda CL, Chang S, Fanara JJ, Kulkarni NH,
Ganguly I, Rollmann SM, Kamdar KP, Mackay TFC.
2003. The genetic architecture of odor-guided behavior
in Drosophila: Epistasis and the transcriptome. Nat Genet
35:180–184.
Anholt RRH, Lyman RF, Mackay TFC. 1996. Effects of
single P-element insertions on olfactory behavior in
Drosophila melanogaster. Genetics 143:293–301.
Anholt RRH, Mackay TFC. 2004. The design of genetic
analyses of complex behaviors in Drosophila. Nat Rev
Genet 5:838–849.
Clyne PJ, Warr CG, Freeman MR, Lessing D, Kim JH,
Carlson JR. 1999. A novel family of divergent seven-
transmembrane proteins: Candidate odorant receptors in
Drosophila. Neuron 22:327–338.
Damberger F, Nikonova L, Horst R, Peng G, Leal WS,
Wuthrich K. 2000. NMR characterization of a pH-
dependent equilibrium between two folded solution con-
formations of the pheromone-binding protein from
Bombyx mori. Protein Sci 9:1038–1041.
de Bruyne M, Clyne PJ, Carlson JR. 1999. Odor coding in a
model olfactory organ: The Drosophila maxillary palp.
J Neurosci 19:4520–4532.
de Bruyne M, Foster K, Carlson JR. 2001. Odor coding in
the Drosophila antenna. Neuron 30:537–552.
Figure 5 Expression patterns of Pinocchio. Panel (A) shows expression of Pino transcript I in
cells in the third antennal segment of adult flies hybridized with an antisense riboprobe specific to
transcript I. Panel (B) shows lack of hybridization with a sense riboprobe to transcript I, demon-
strating specificity of the staining in panel (A). The second and third antennal segments are desig-
nated II and III, respectively; ‘‘s’’ designates the sacculus.
156 Rollmann et al.

Du G, Prestwich GD. 1995. Protein structure encodes the
ligand binding specificity in pheromone binding proteins.
Biochemistry 34:8726–8732.
Dunipace L, Meister S, McNealy C, Amrein H. 2001.
Spatially restricted expression of candidate taste recep-
tors in the Drosophila gustatory system. Curr Biol
11:822–835.
Fedorowicz GM, Fry JD, Anholt RRH, Mackay TFC. 1998.
Epistatic interactions between smell impaired loci in
Drosophila melanogaster. Genetics 148:1885–1891.
Galindo K, Smith DP. 2001. A large family of divergent
Drosophila odorant-binding proteins expressed in gusta-
tory and olfactory sensilla. Genetics 159:1059–1072.
Gao Q, Chess A. 1999. Identification of candidate Droso-
phila olfactory receptors from genomic DNA sequence.
Genomics 60:31–39.
Gao Q, Yuan B, Chess A. 2000. Convergent projections of
Drosophila olfactory neurons to specific glomeruli in the
antennal lobe. Nat Neurosci 3:780–785.
Hallem EA, Ho MG, Carlson JR. 2004. The molecular basis of
odor coding in the Drosophila antenna. Cell 117:965–979.
Hekmat-Scafe DS, Scafe CR, McKinney AJ, Tanouye MA.
2002. Genome-wide analysis of the odorant-binding pro-
tein gene family in Drosophila melanogaster. Genome
Res 12:1357–1369.
Hekmat-Scafe DS, Steinbrecht RA, Carlson JR. 1997.
Coexpression of two odorant-binding protein homologs
in Drosophila: Implications for olfactory coding. J Neuro-
science 17:1616–1624.
Hildebrand JG, Shepherd GM. 1997. Mechanisms of olfac-
tory discrimination: Converging evidence for common
principles across phyla. Annu Rev Neurosci 20:595–631.
Holt RA, Subramanian GM, Halpern A, Sutton GG, Charlab
R, Nusskern DR, Wincker P, Clark AG, Ribeiro JM,
Wides R, et al. 2002. The genome sequence of the malaria
mosquito Anopheles gambiae. Science 298:129–149.
Horst R, Damberger F, Luginbuhl P, Guntert P, Peng G,
Nikonova L, Leal WS, Wuthrich K. 2001. NMR structure
reveals intramolecular regulation mechanism for phero-
mone binding and release. Proc Natl Acad Sci USA
98:14374–14379.
Kaissling KE. 1996. Peripheral mechanisms of pheromone
reception in moths. Chem Senses 21:257–268.
Kim MS, Repp A, Smith DP. 1998. LUSH odorant-binding
protein mediates chemosensory responses to alcohols in
Drosophila melanogaster. Genetics 150:711–721.
Kim MS, Smith DP. 2001. The invertebrate odorant-bind-
ing protein LUSH is required for normal olfactory behav-
ior in Drosophila. Chem Senses 26:195–199.
Kruse SW, Zhao R, Smith DP, Jones DN. 2003. Structure
of a specific alcohol-binding site defined by the odorant
binding protein LUSH from Drosophila melanogaster.
Nat Struct Biol 10:694–700.
Laissue PP, Reiter C, Hiesinger PR, Halter S, Fischbach
KF, Stocker RF. 1999. Three-dimensional reconstruction
of the antennal lobe in Drosophila melanogaster. J Comp
Neurol 405:543–552.
Larsson MC, Domingos AI, Jones WD, Chiappe ME,
Amrein H, Vosshall LB. 2004. Or83b encodes a broadly
expressed odorant receptor essential for Drosophila
olfaction. Neuron 43:703–714.
Leal WS. 2000. Duality monomer-dimer of the pheromone-
binding protein from Bombyx mori. Biochem Biophys
Res Commun 268:521–529.
Leal WS, Nikonova L, Peng G. 1999. Disulfide structure of
the pheromone binding protein from the silkworm moth,
Bombyx mori. FEBS Lett 464:85–90.
Mackay TFC, Hackett JB, Lyman RF, Wayne ML, Anholt
RRH. 1996. Quantitative genetic variation of odor-
guided behavior in a natural population of Drosophila
melanogaster. Genetics 144:727–735.
Marin EC, Jefferis GS, Komiyama T, Zhu H, Luo L. 2002.
Representation of the glomerular olfactory map in the
Drosophila brain. Cell 109:243–255.
Mombaerts P, Wang F, Dulac C, Chao SK, Nemes A, Men-
delsohn M, Edmondson J, Axel R. 1996. Visualizing an
olfactory sensory map. Cell 87:675–686.
Park SK, Shanbhag SR, Wang Q, Hasan G, Steinbrecht RA,
Pikielny CW. 2000. Expression patterns of two putative
odorant-binding proteins in the olfactory organs of Dro-
sophila melanogaster have different implications for their
functions. Cell Tissue Res 300:181–192.
Pasyukova EG, Vieira C, Mackay TFC. 2000. Deficiency
mapping of quantitative trait loci affecting longevity in
Drosophila melanogaster. Genetics 156:1129–1146.
Pomeroy SL, LaMantia AS, Purves D. 1990. Postnatal con-
struction of neural circuitry in the mouse olfactory bulb.
J Neurosci 10:1952–1966.
Ressler KJ, Sullivan SL, Buck LB. 1994. Information cod-
ing in the olfactory system: evidence for a stereotyped
and highly organized epitope map in the olfactory bulb.
Cell 79:1245–1255.
Robertson HM, Martos R, Sears CR, Todres EZ, Walden KK,
Nardi JB. 1999. Diversity of odourant binding proteins
revealed by an expressed sequence tag project on male
Manduca sexta moth antennae. Insect Mol Biol 8:501–518.
Rubin GM. 2000. Comparative genomics of the eukaryotes.
Science 287:2204–2215.
Sandler BH, Nikonova L, Leal WS, Clardy J. 2000. Sexual
attraction in the silkworm moth: structure of the phero-
mone-binding-protein-bombykol complex. Chem Biol
7:143–151.
Scott K, Brady R Jr, Cravchik A, Morozov P, Rzhetsky A,
Zuker C, Axel RA. 2001. Chemosensory gene family
encoding candidate gustatory and olfactory receptors in
Drosophila. Cell 104:661–673.
Shanbhag SR, Hekmat-Scafe D, Kim MS, Park SK, Carlson
JR, Pikielny C, Smith DP, Steinbrecht RA. 2001. Expres-
sion mosaic of odorant-binding proteins in Drosophila
olfactory organs. Microscopy Res Technique 55:297–306.
Stocker RF. 1994. The organization of the chemosensory
system in Drosophila melanogaster: a review. Cell Tissue
Res 275:3–26.
Vassar R, Chao SK, Sitcheran R, Nunez JM, Vosshall LB,
Axel R. 1994. Topographic organization of sensory pro-
jections to the olfactory bulb. Cell 79:981–991.
Vogt RG, Callahan FE, Rogers ME, Dickens JC. 1999.
Odorant binding protein diversity and distribution among
Pinocchio Contributes to Olfaction 157

the insect orders, as indicated by LAP, an OBP-related
protein of the true bug Lygus lineolaris (Hemiptera,
Heteroptera). Chem Senses 24:481–495.
Vogt RG, Rogers ME, Franco MD, Sun M. 2002. A compa-
rative study of odorant binding protein genes: differential
expression of the PBP1-GOBP2 gene cluster in Manduca
sexta (Lepidoptera) and the organization of OBP genes
in Drosophila melanogaster (Diptera). J Exp Biol 205:
719–744.
Vogt RG, Rybczynski R, Lerner MR. 1991. Molecular
cloning and sequencing of general odorant-binding
proteins GOBP1 and GOBP2 from the tobacco hawk
moth Manduca sexta: comparisons with other insect
OBPs and their signal peptides. J Neurosci 11:2972–
2984.
Vosshall LB, Amrein H, Morozov PS, Rzhetsky A, Axel R.
1999. A spatial map of olfactory receptor expression in
the Drosophila antenna. Cell 96:725–736.
Vosshall LB, Wong AM, Axel R. 2000. An olfactory sen-
sory map in the fly brain. Cell 102:147–159.
Wojtasek H, Leal WS. 1999. Conformational change in the
pheromone-binding protein from Bombyx mori induced
by pH and by interaction with membranes. J Biol Chem
274:30950–30956.
Wong AM, Wang JW, Axel R. 2002. Spatial representation
of the glomerular map in the Drosophila protocerebrum.
Cell 109:229–241.
Woodard C, Huang Sun H, Helfand SL, Carlson J. 1989.
Genetic analysis of olfactory behavior in Drosophila: a
new screen yields the ota mutants. Genetics 123:315–326.
158 Rollmann et al.





























