Embed Size (px)
Citation preview

This article was downloaded by: [Tamas Komives]On: 30 April 2013, At: 01:57Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
International Journal of PhytoremediationPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/bijp20
Phytoextraction potential of wild type and 35S-gshItransgenic poplar trees (Populus × canescens) forenvironmental pollutants herbicide paraquat, saltsodium, zinc sulfate and nitric oxide in vitroGábor Gyulai a , András Bittsánszky b , Zoltán Szabó c , Luther Waters Jr. d , Gábor Gullner b ,Györgyi Kampfl e , György Heltai e & Tamás Kőmíves b
a Institute of Genetics and Biotechnology , Szent István University , 2103 , Gödöllő , Hungaryb Plant Protection Institute, Agricultural Research Centre, Hungarian Academy of Sciences ,Budapest , 1022 , Hungaryc Agricultural Biotechnology Center , 2100 , Gödöllő , Hungaryd Department of Horticulture, College of Agriculture , Auburn University , Alabama , 36849 ,USAe Department of Chemistry and Biochemistry , Szent István University , 2103 , Gödöllő ,HungaryAccepted author version posted online: 29 Apr 2013.
To cite this article: Gábor Gyulai , András Bittsánszky , Zoltán Szabó , Luther Waters Jr. , Gábor Gullner , Györgyi Kampfl ,György Heltai & Tamás Kőmíves (2013): Phytoextraction potential of wild type and 35S-gshI transgenic poplar trees (Populus ×canescens) for environmental pollutants herbicide paraquat, salt sodium, zinc sulfate and nitric oxide in vitro , InternationalJournal of Phytoremediation, DOI:10.1080/15226514.2013.783553
To link to this article: http://dx.doi.org/10.1080/15226514.2013.783553
Disclaimer: This is a version of an unedited manuscript that has been accepted for publication. As a serviceto authors and researchers we are providing this version of the accepted manuscript (AM). Copyediting,typesetting, and review of the resulting proof will be undertaken on this manuscript before final publication ofthe Version of Record (VoR). During production and pre-press, errors may be discovered which could affect thecontent, and all legal disclaimers that apply to the journal relate to this version also.
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form toanyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses shouldbe independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims,

proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly inconnection with or arising out of the use of this material.
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 1
Phytoextraction potential of wild type and 35S-gshI transgenic poplar trees (Populus ×
canescens) for environmental pollutants herbicide paraquat, salt sodium, zinc sulfate and
nitric oxide in vitro
Gábor Gyulai1*, András Bittsánszky2, Zoltán Szabó3, Luther Waters Jr.4, Gábor Gullner2,
Györgyi Kampfl5, György Heltai5 and Tamás Kőmíves2
1Institute of Genetics and Biotechnology, Szent István University, 2103 Gödöllő, Hungary
2Plant Protection Institute, Agricultural Research Centre, Hungarian Academy of Sciences,
Budapest 1022, Hungary
3Agricultural Biotechnology Center, 2100 Gödöllő, Hungary
4Department of Horticulture, College of Agriculture, Auburn University, Alabama 36849, USA
5Department of Chemistry and Biochemistry, Szent István University, 2103 Gödöllő, Hungary
*Correspondence: [email protected]
E-mail addresses: [email protected]; [email protected];
[email protected]; [email protected]; [email protected];
[email protected]; [email protected]
Abstract
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 2
Phytoextraction potentials of two transgenic (TR) poplar (Populus × canescens) clones TRggs11
and TRlgl6 were compared with that of wild-type (WT) following exposure to paraquat, zinc
sulfate, common salt and nitric oxide (NO), using a leaf-disc system incubated for 21 days on
EDTA-containing nutritive WPM media in vitro. Glutathione (GSH) contents of leaf discs of
TRlgl6 and TRggs11 showed increments to 296 % and 190%, respectively, compared with WT.
NO exposure led to a twofold GSH content in TRlgl6, which was coupled with a significantly
increased sulfate uptake when exposed to 10-3 M ZnSO4. The highest mineral contents of Na, Zn,
Mn, Cu and Mo was observed in the TRggs11 clone. Salt-induced activity of catalase enzyme
increased in both TR clones significantly compared with WT under NaCl (0.75% and 1.5%)
exposure. The in silico sequence analyses of gsh1 genes revealed that P. x canadensis and Salix
sachalinensis show the closeset sequence similarity to that of P. x canescens, which predicted an
active GSH production with high phytoextraction potentials of these species with indication for
their use where P. x canescens can not be grown.
Keywords: Phytoextraction, environmental pollutants, gene sequence analyses
1. Introduction
Poplars are frequently used for treating manure of swine houses in Hungary (Europe), and when
conditions for nitrification and denitrification are suboptimal, volatile nitrogen oxides (NO and
N2O) can be released (Kim et al., 2007). Our study aimed to evaluate the response of poplar to
NO gas. Soils around Gödöllő (Hungary) have naturally high levels of Zn content (53.3±2.3 mg
kg-1 DW soil; Gyulai et al., 2012b), and due to the overuse of paraquat in agriculture
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 3
(Bittsánszky et al. 2009), experiments were designed to study zinc and paraquat tolerance of
poplars. Primary and secondary soil salinity is also a major environmental hazard not only in
Hungary but in the USA and Australia (Csillag et al., 1993; Metternicht and Zinck, 2003), which
is a challenge to study plant salt tolerance (Yamaguchi and Blumwald, 2005; Binh et al., 1992).
Numerous plant species are able to tolerate toxic substances and heavy metals from polluted soils
and air, such as the annual species Thlaspi caerulescens, a known nickel (Ni) and zinc (Zn)
hyperaccumulator plant; Brassica juncea, a Pb accumulator (Gleba et al., 1999); and the arsenic
(As) hyperaccumulator Pteris vittata (Nagarajan and Ebbs, 2010). Metallocrops such as oat
(Avena sativa), barley (Hordeum vulgare) and Indian mustard (B. juncea) also tend to take up
high level of Cu, Cd and Zn in hydroponics (Ebbs and Kochian, 1998). Unlike these annual
plants, given their high growth rate and perennial life history, woody poplars of both wild type
(WT) and transgenic (TR) were found to be optimal for the elimination of environmental
contaminants through phytoextraction (Peuke and Rennenberg, 2005; Bittsánszky et al., 2005;
Gyulai et al., 2012a).
Poplar trees, used in our study, were transformed to overexpress a bacterial gene encoding
gamma-glutamylcysteine synthetase (EC 6.3.2.2), which is the rate-limiting regulatory enzyme
in the biosynthesis of the ubiquitous tripeptide thiol compound glutathione (GSH) (Peuke and
Rennenberg, 2005). The overexpressed protein was directed either in the cytosol (TRggs11 line)
(Arisi et al., 1997) or in the chloroplasts (TRlgl6 line) (Noctor et al.,1998).
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 4
The 35S-gshI TR poplars were previously investigated in vitro (Gullner et al., 2001; Gyulai et
al., 2005; Bittsánszky et al., 2006) and in situ (Peuke and Rennenberg, 2005; Peuke et al., 2012).
The gshI transgene incorporation (Gyulai et al., 2005) and gene expression levels (Bittsánszky et
al., 2006) were characterized with the capability for further gene activation by treatment with the
DNA-demethylating agent 5,6-dihydro-5'-azacytidine hydrochloride (DHAC) (Gyulai et al.,
2012a).
In the study presented GSH and cysteine (Cys) content, mineral uptake capacity, and the activity
of stress indicator enzyme catalaze (CAT) was measured in poplar leaf tissue exposed to
environmental aerial contaminants nitric oxide (NO), zinc sulfate (ZnSO4), common salt (NaCl),
and the herbicide paraquat (PQ) in aseptic leaf disc cultures in vitro. In silico sequence analyses
of plant gsh1 genes were carried out to identify plant species with high sequences similarities to
those of poplar.
2. Methods
2.1. Plant materials
A clone of the untransformed (INRA-717-1-B4) natural hybrid poplar P. × canescens (P.
tremula × P. alba), and two genetically transformed lines were used that overexpress the
bacterial (E. coli) gshI gene (gamma glutamylcysteine synthetase; EC 6.3.2.2) (NCBI: X03954;
Watanabe et al., 1986). Unlike TRggs11 (35s-gshI), transgene cassette of TRlgl6 (35S-gshI-
rbcS) carried an additional targeting sequence (32 to 202 of the 206 bp stretch; NCBI M25614)
of transit peptide (RBCS) gene rbcS (RuBPCase SSU: small subunit of RuBPCase, ribulose-1,5-
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 5
bisphosphate carboxylase, EC 4.1.1.39), which facilitates the translocation of cytosolically
synthesized RBCS-GSH complex into chloroplasts (Leplé et al., 1992; Arisi et al., 1997; Noctor
et al., 1998).
Poplars were micropropagated and maintained in aseptic shoot cultures from nodal segments
following the methods of Gyulai et al. (2005) and Koprivova et al. (2002). Briefly, shoot
segments (0.5 cm) were placed on woody plant medium (WPM) (Lloyd and McCown, 1980)
supplemented with benzyl adenine (BA; 0.5 mg-1 L) and α-naphthaleneacetic acid (NAA; 0.2
mg-1 L), then incubated for 21 days under a 16 h/8 h (light/dark) photoperiod (3000 lux, 4.05
µmol/photons/m2/s). Auxiliary shoots that developed were excised, transferred to hormone-free
WPM media to induce rooting and incubated for an additional 21 days. Leaves of aseptic shoots
were sampled and used for leaf-disc cultures in accordance with the method of Gyulai et al.
(2012a).
The woody plant medium (WPM) (Duchefa Biochemie NL; #M0220) included the following
nutritive elements: NH4NO3 (400.0 mg L-1), H3BO3 (6.2 mg L-1), CaCl2 (72.5 mg L-1), Ca(NO3)2
x 4H2O (386.0 mg L-1), CuSO4 x 5H2O (0.25 mg L-1), EDTA x 2H2O (37.3 mg L-1), FeSO4
x7H2O (27.85 mg L-1), MgSO4 (180.7 mg L-1), MnSO4 x H2O (22.3 mg L-1), Na2MoO4 x 2H2O
(0.25 mg L-1), KH2PO4 (170.0 mg L-1), K2SO4 (990.0 mg L-1) and ZnSO4 x 7H2O (8.6 mg L-1).
2.2. Nitric oxide exposure
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 6
Aseptic WPM medium was supplemented with a concentration series (10-7 to 10-4 M) of NaNP
(Na-nitroferricyanide III; Na2Fe(CN)5NO × 2H2O) to release NO following the method of
Floryszak-Wieczorek et al. (2006). Gas samples (250 µl) were collected with gas-tight Hamilton
syringes from the headspace of each culture vessel (1 L volume each) and analyzed with an
ANTEK 7050 NO-analyser using a chemiluminescent detector. The NO content was calculated
by one-point linear calibration using external NO gas for calibration. The peak areas, which were
proportional to the measured gas concentrations, were calculated with an HP 3396 Integrator and
Microsoft Excel software in accordance with Kampfl et al. (2007).
2.3. Paraquat (PQ) exposure
Aseptic WPM medium, solidified with 0.8 g L-1 agar, was supplemented with a concentration
series (10-8, 5 × 10-8, 10-7 and 5 × 10-7 M) of the herbicide paraquat (methyl viologen; 1,1'-
dimethyl-4.4'-bipyridinium dichloride) combined with and without 10-7 M NaNP. Leaf discs (8
mm diameter; three leaf discs per petri dishes of 6 cm, in triplicates) were laid onto the surface of
media and incubated for 21 days under a 16 h/8 h (light/dark) photoperiod (3000 lux, 4.05 µmol
photons/m2/s).
2.4. ZnSO4 exposure
Leaf discs were excised and placed on aseptic EDTA-containing (37.25 mg-1 L) WPM agar
media supplemented with a concentration series of ZnSO4 (10-2 M to 10-4 M), and incubated for
21 days, according to the standard aseptic culture methods, under a 16 h/8 h (light/dark)
photoperiod (3000 lux, 4.05 µmol photons/m2/s) following the method of Gyulai et al. (2005,
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 7
2012a). Untreated basal WPM medium contained 29.9068 µM Zn2+ (equal to 1.955 mg-1 L Zn2+)
and 14.36 meq-1 L SO42- (George et al., 1987).
2.5. Bioconcentration factor
Bioconcentration factor (BCF) ratios were calculated by dividing the molar concentrations of the
elements measured in leaf discs with the element concentrations supplied in the WPM medium
according to Mackay (1982).
2.6. NaCl exposure
NaCl was added to WPM agar media at the concentrations of 0.75%, 1.5%, 2.25%, and 3.0%
prior to autoclaving the media. Leaf discs were incubated for 21 days under a 16 h/8 h
(light/dark) photoperiod (3000 lux, equal to 4.05 µmol photons/m2/s). Basal WPM medium
contained 4.59 mg-1 L Na supplied as the Fe-chelating agent Na2EDTA × 2H2O (37.25 mg-1 L)
(George et al., 1987).
2.7. Inductively coupled plasma emission spectrometry
The Na, Zn, Mn, Cu and Mo elemental content of poplar leaf discs was determined by ICP
(Inductively Coupled Plasma emission spectrometry) following the methods of Zarcinas et al.
(1987). Leaf discs were oven-dried at 70 ºC overnight, and 0.5 mg leaf tissue of each treatment
was transferred to digestion vessels in triplicates, digested with a mixture of 5 ml HNO3 and 2 ml
H2O2 using a MILESTONE 200 MEGA microwave oven. The microwave digestion program
was as follows: 6 min at 250 W; 6 min at 400 W; 6 min at 650 W; 6 min at 250 W closed by
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 8
cooling for 5 min to room temperature. The digested solution was filtered through Macherey-
Nagel G 1/4 filter-paper, diluted to 25 ml, and was analyzed with Jobin Yvon 24 sequent ICP
Optical Emission Spectrometer. The operation parameters were as follows: incident power, 1
kW; outer argon flow 12 L min-1; intermediate argon flow 0.2 L min-1; inner argon flow 1.1 L
min-1; with nebulizer Meinhardt type nebulizer, and sample uptake rate 1 mL min-1 according to
Heltai et al. (2000).
2.8. HPLC analysis
Total GSH contents were measured using 0.1 g fresh leaf tissue of leaf discs after grinding in
liquid nitrogen. The content of cysteine (Cys), an amino acid precursor of GSH, was also
determined by reverse-phase HPLC with spectrofluorometric detection after derivatisation with
monobromobimane according to the methods of Strohm et al. (1995).
2.9. Catalase (CAT) activity
CAT activity was measured in extracts of 0.2 g fresh leaf tissue of leaf discs after grinding in
liquid nitrogen following the method of Aebi (1984).
2.10. Statistical analysis
At least three independent parallel measurements were carried out in each experiment.
Differences between mean values were assessed with Student's t-test at a significance level of P
= 0.05.
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 9
2.11. Multiple sequence alignments
Nucleotide sequences of gsh1 genes were downloaded from the National Center for
Biotechnology Information (NCBI) databases (Altschul et al., 1997). Multiple sequence
alignments were applied in silico with the software programs BioEdit Sequence Alignment
Editor (North Carolina State University, USA) (Hall, 1999), MULTALIN (Combet et al., 2000),
CLUSTAL W (Thompson et al., 1994), and FastPCR (Kalendar et al., 2009). For BLAST
analysis the NCBI databases were used (Altschul et al., 1997). Hierarchical cluster analysis with
the Maximum Likelihood (ML) method (Hillis et al., 1994) was used to construct dendrograms
from the alignment sequences using computer program MEGA4 (Tamura et al., 2007).
3. Results
3.1. Paraquat exposure
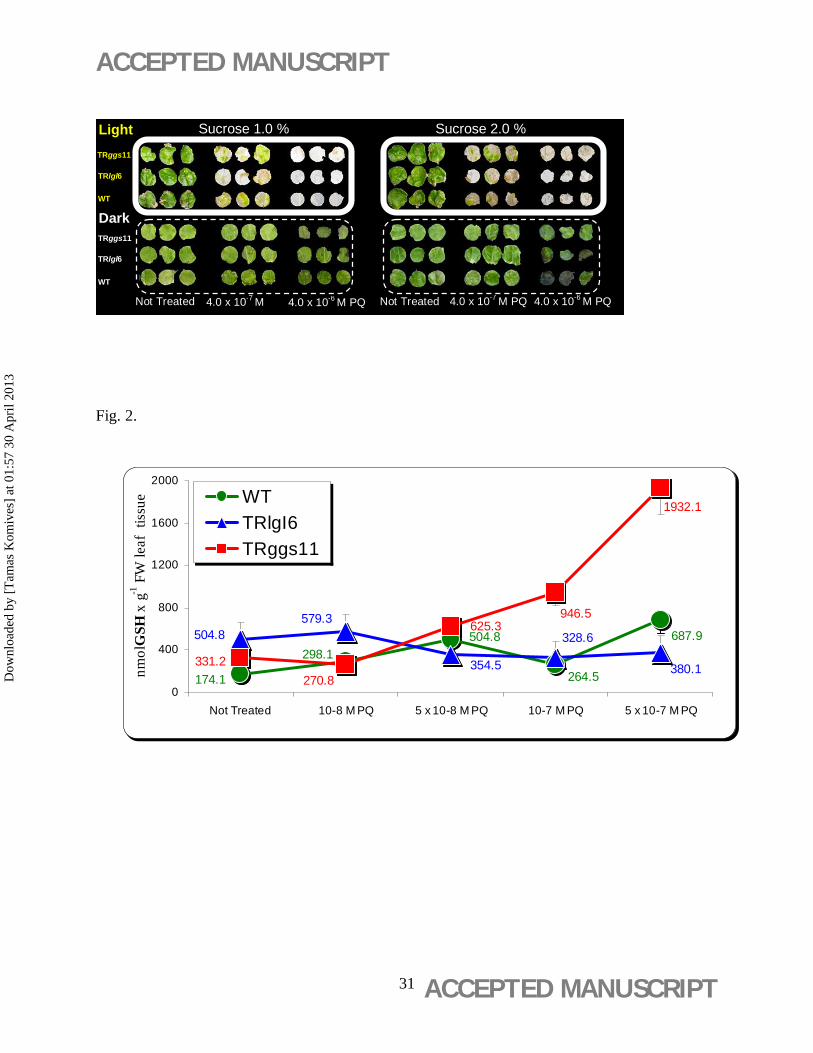
A concentration of 4.0 × 10-6 M paraquat led to bleaching the leaf discs (Figure 1) of each of the
poplar clones. A concentration of 4.0 × 10-7 M paraquat caused chloroplast sublethality with a
mixture of bleached and green spots on the leaf discs (Figure 1). No bleaching effect of paraquat
was observed in the dark-incubated leaf discs (Figure 1).
3.2. GSH contents
The GSH content in both transgenic lines significantly increased by 190% (331.1±13 nmol GSH
g-1 FW for TRggs11) and 296% (504.8±22 nmol GSH g-1 FW for TRlgl6), respectively,
compared with that of WT (174.3±11 nmol GSH g-1 FW) (Figure 2).
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 10
Under paraquat treatment, the GSH content of TRggs11 clone showed a concentration-dependent
increment with increasing concentration of the herbicide and peaked at 5 × 10-7 M paraquat
(1932.1±70 nmol GSH g-1 FW) (Figure 2).
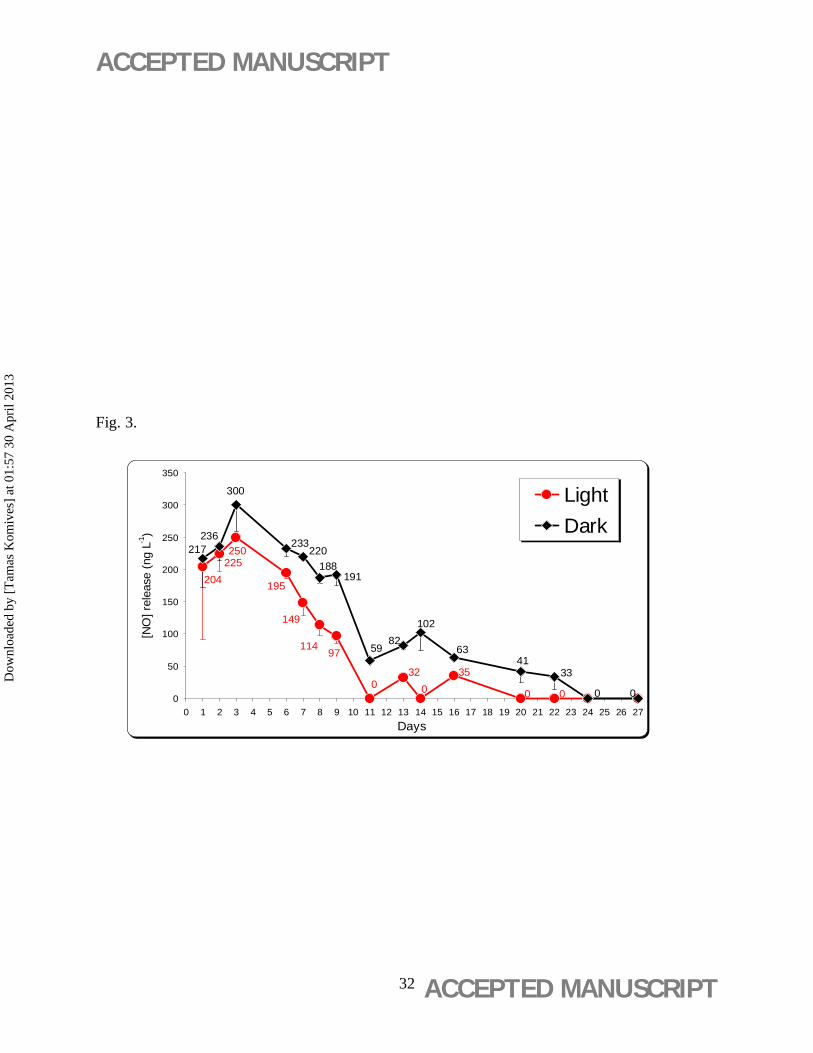
3.3. Nitric oxide exposure
The light/dark-dependent NO release from NaNP did not show significant differences between
light and dark incubation, with the highest release of NO (300±16 ng-1 L) at the 3rd day of the
dark incubated vessels (Figure 3).
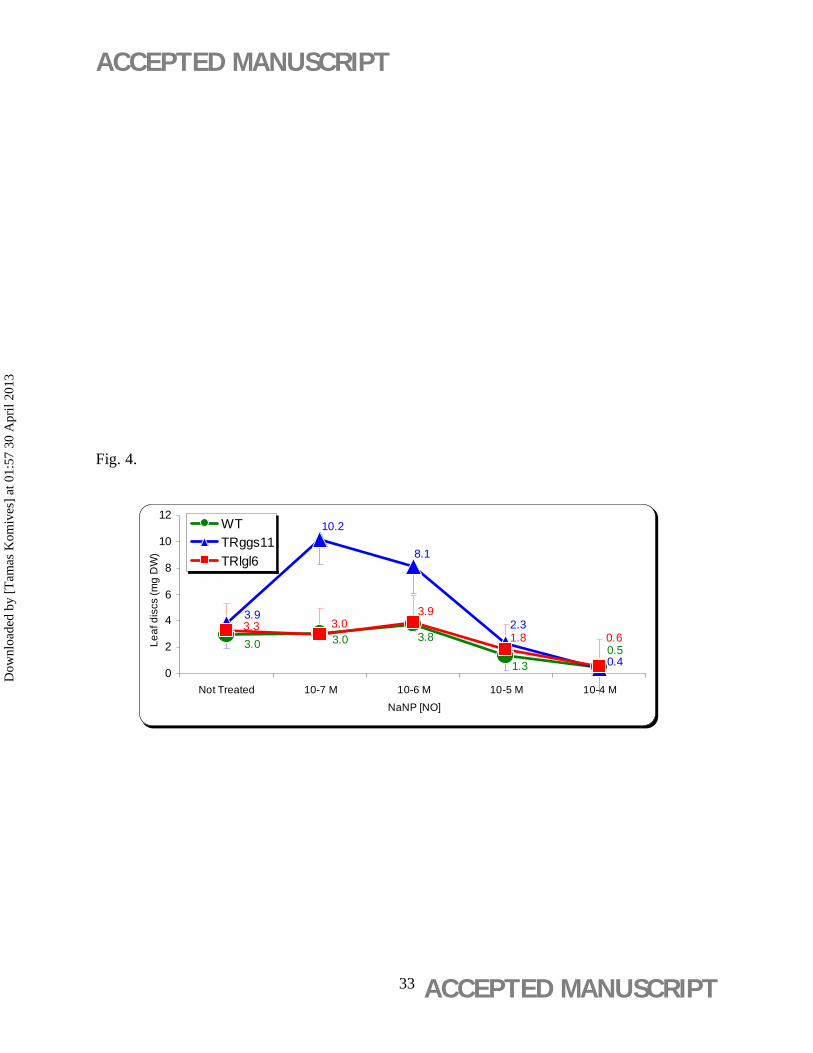
Dry weight (DW) of leaf discs of the TRggs11 clone tripled at a concentration of 10-7 M NaNP,
but NaNP had no effect on the DW of TRlgl6 and WT (Figure 4).
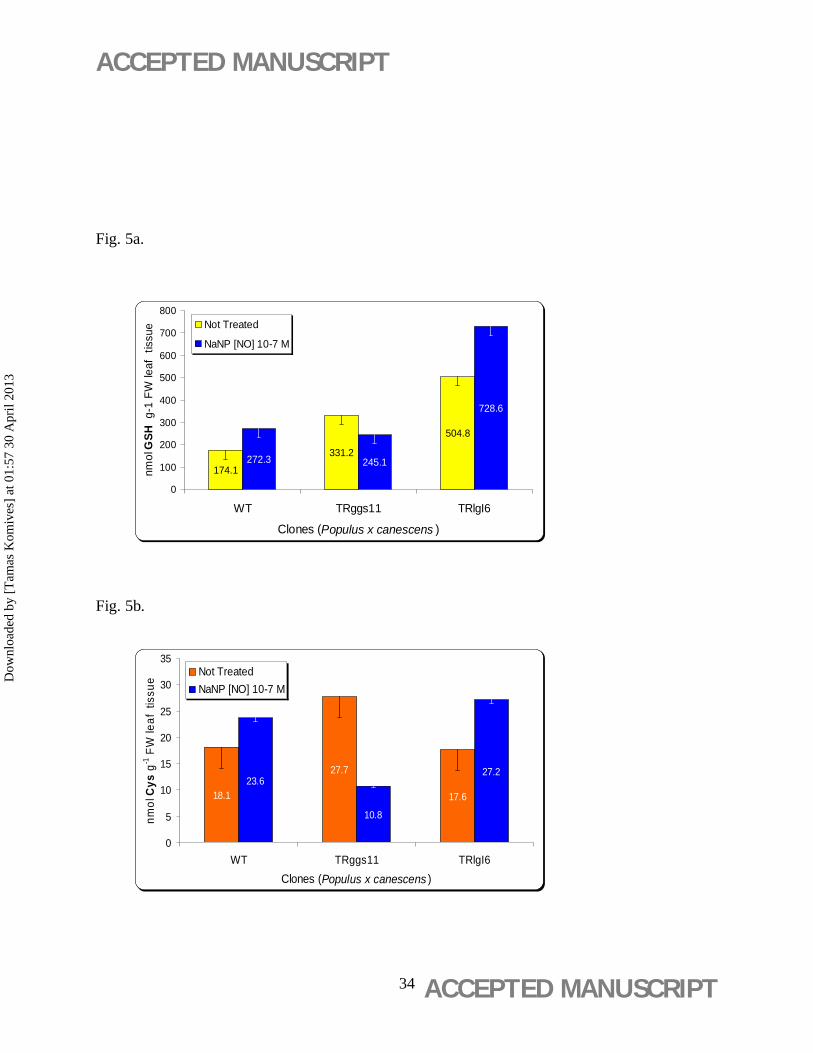
Nitric oxide-induced GSH overproduction at 10-7 M NaNP in the TRlgl6 clone (from 504.8±22
to 728.6±29 nmol GSH g-1 FW) and in the WT (from 174.3±11 to 272.3±29 nmol GSH g-1 FW)
(Figure 5a). The NO had no effect on the GSH content of TRggs11 clone, however the GSH
precursor amino acid Cys showed the highest level (27.7±1.1 nmol GSH g-1 FW) in this clone
without NO treatment (Figure 5b).
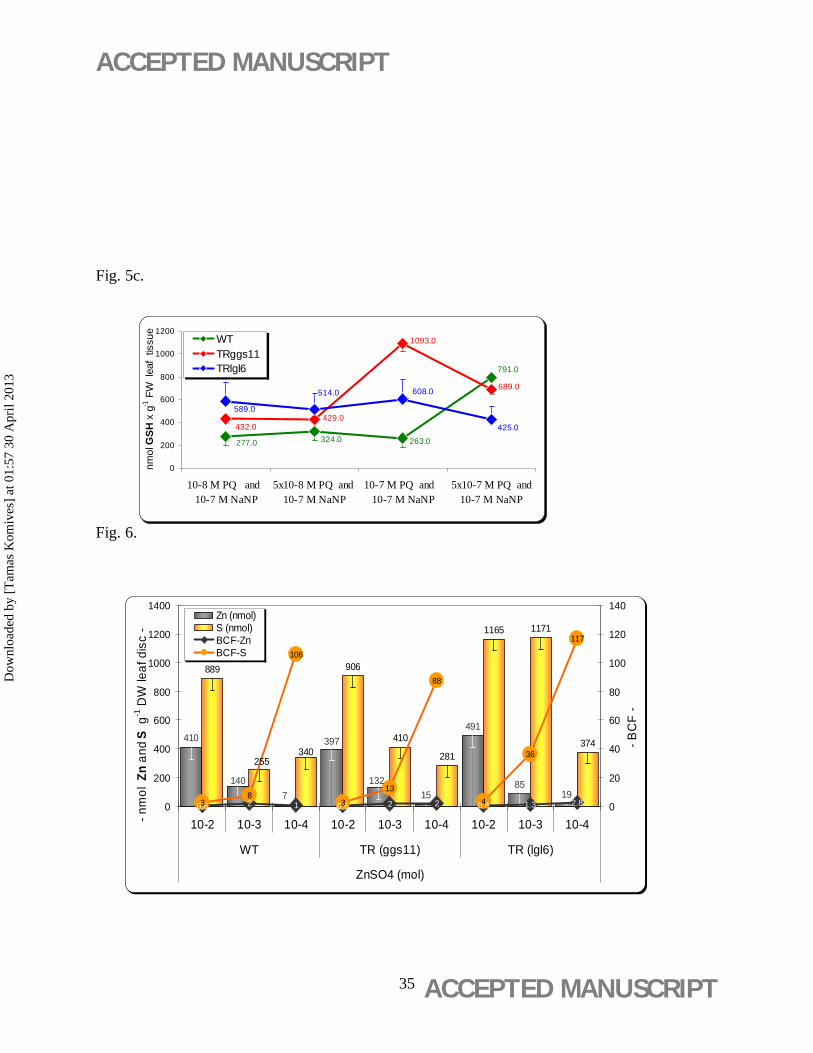
NaNP (10-7 M) combined with PQ (10-7 M) generated the highest level of GSH production
(1093.0±45 nmol GSH g-1 FW) in the TRggs11 clone (Figure 5c). The WT also showed PQ-
stimulated NO-responsiveness (791.0±30 nmol GSH g-1 FW) at a lower (5 × 10-7 M)
concentration of PQ combined with 10-7 M NaNP (Figure 5c).
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 11
3.4. Zn2+ and SO42- uptake of poplars
Significantly higher S uptake capacity was observed by the TRlgl6 clone at 10-3 M ZnSO4
concentration (1171±48 nmol S g-1 DW) compared with that of TRggs11 (906±37 nmol S g-1
DW) and the WT (889±38 nmol S g-1 DW) (Figure 6).
When molar concentrations were calculated, the content of Zn and S in the leaf discs showed not
the expected 1:1 ratio with the highest level of Zn exclusions in Trlgl6 at 10-3 ZnSO4 (Figure 6).
Consequently, BCFZn of leaf discs showed low and concentration-independent values in each
clones, whereas BCFS values peaked at 10-4 M ZnSO4 in all clones with the highest value for
TRlgl6 (Figure 6).
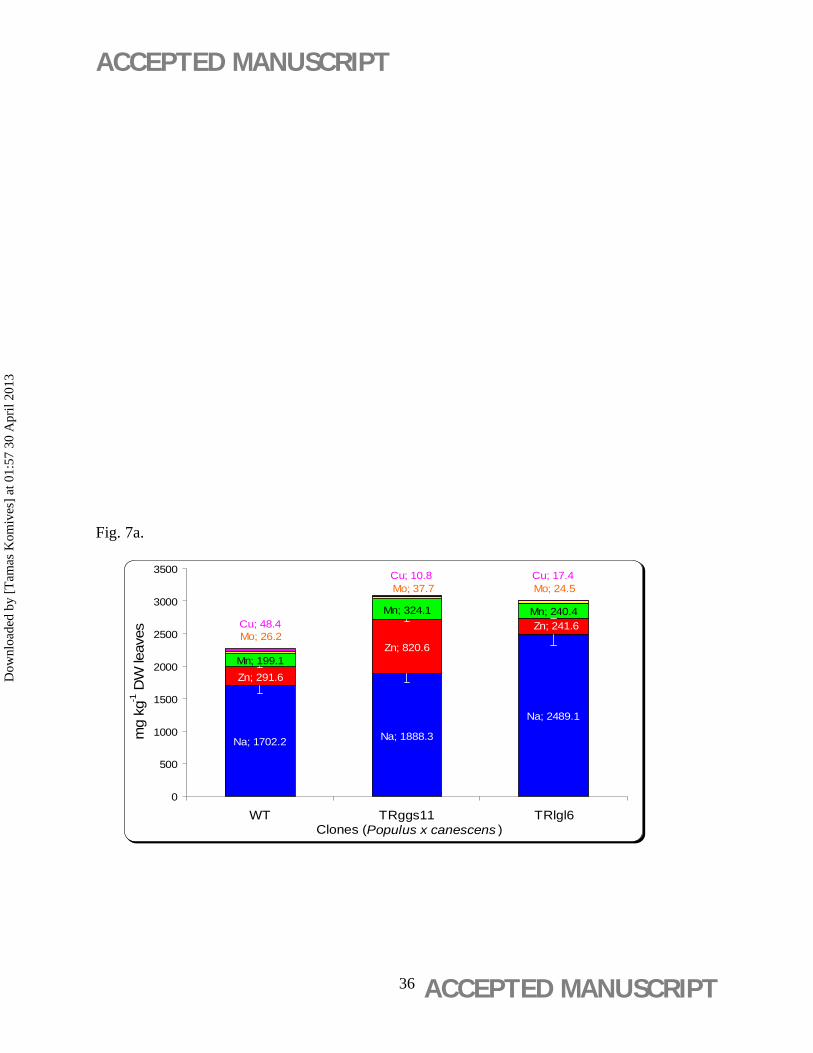
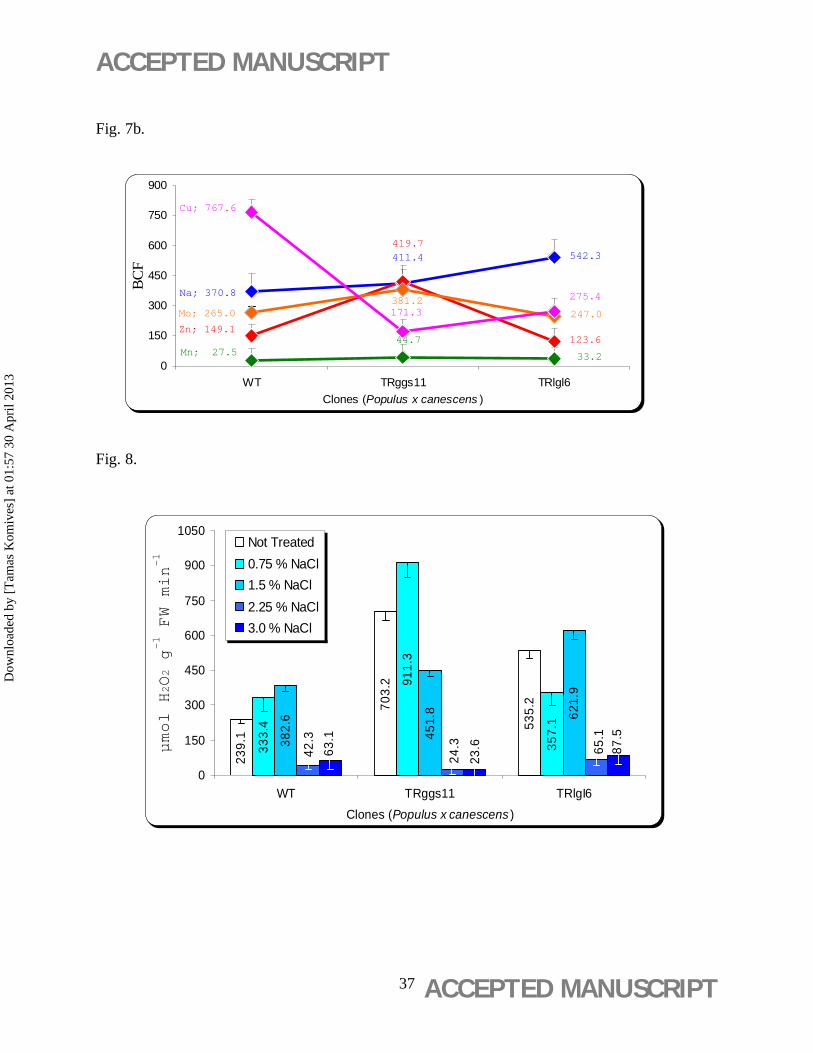
3.5. Total content of Zn, Na, Mn, Cu and Mo
The total contents (mg kg-1 DW) of Zn, Na, Mn, Mo and Cu measured in the leaf discs incubated
on basal WPM medium were the highest in the TRggs11 clone (3.3±0.1 g kg-1 DW leaf tissue)
(Figure 7a). This clone contained also the highest contents of Zn (0.82±0.03 g kg-1 DW leaf
tissue) (Figure 7a). TRlgl6 showed the highest Na content (2.49±0.9 g kg-1 DW leaf tissue). The
highest BCFCu value was observed in the WT (Figure 7b) with significantly higher Cu (48±1.9
mg kg-1 DW) content than in the two TR clones (Figure 7a).
3.6. Salt-induced CAT activity
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 12
The enzyme activity of CAT increased with the increasing concentration of NaCl (0.75% and
1.5%) (Figure 8) in all clones studied. However, significantly higher CAT activity was observed
in both transgenic lines compared with WT with an extreme level in the TRggs11 clone
(911.3±35 µmol H2O2 g-1 FW min-1) at 0.75% NaCl (Figure 8).
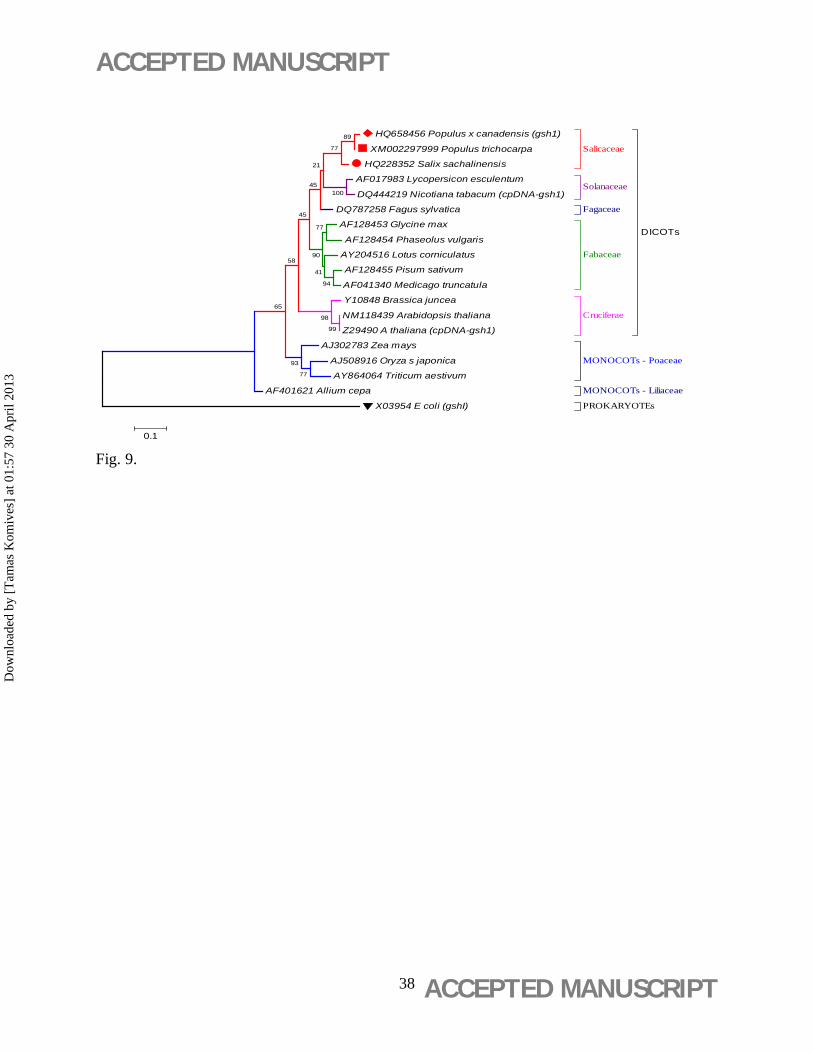
3.7. Multiple sequence alignment and phylogenetic analysis
The analysis revealed an evolutionarily conserved gsh1 cladogram (Figure 9), which strictly
followed the systematic order of plant families, and indicated that P. x canadensis and Salix
sachalinensis of Salicaceae family are genetically the closest to P. x canescens analyzed at this
gene (Figure 9).
4. Discussion
Genetic transformations provide unique plant materials for modeling plant responses to
environmental contaminations (Creissen et al., 1996; Czakó et al., 2006). Due to high growth
rate and water uptake capacity poplars became the main trees for not only the industrial (e.g.
paper production; Park et al., 2004) but also for environmental (e.g. phytoremediation) uses
(Peuke and Rennenberg, 2005; Bittsánszky et al., 2009). Transgenic poplar clones, used in our
study, were developed by genetic transformation of the bacterial (Escherichia coli) gene gshI
that encodes gamma-glutamylcysteine synthetase (Watanabe et al., 1986). It resulted in a dual
expression of poplar gsh1 and bacterial gshI genes with the endproduct of GSH, which
accumulated either in the cytosol by TRggs11 (Arisi et al., 1997) or in the chloroplasts by
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 13
TRlgl6 (Noctor et al., 1998) with increasing GSH production (Leplé et al., 1992). As plant
phytoextraction capacities show a direct correlation with their GSH content, these trees play a
potential role in phytoremediation and phytoextraction (Noctor et al., 1995; Gullner et al., 2001).
Owing to the presence of the sulfhydril (SH) group of Cys residue in the tripeptide GSH,
GSH is a reducing and electron donor agent. When GSH is oxidized, a disulfide bond is formed
between two GSH molecules and GSH is converted to the oxidized form of glutathione disulfide
(GSSG) with the release of an electron. GSSG can be reduced back to GSH by glutathione
reductase (GR) (EC 1.6.4.2) using NADPH as an electron donor. The ratio between reduced
GSH and oxidized GSSG is a sensitive indicator of cellular toxicity (Pompella et al., 2003). GHS
combined with another reductants dithiothreitol can further enhance reduction more than GSH
alone like in the case of arsenate used in vitro (Nagarajan and Ebbs, 2010).
Paraquat, as an effective electron acceptor, primarily affects the photosynthetic electron transport
chain (PETC) located in the chloroplasts (Will et al., 2001). For paraquat treatment in our study,
a reduced sucrose concentration (1%) was supplied to the agar media to stimulate the activity of
PETCs, which are switched off by higher sucrose concentrations (2-3%) (Medgyesi et al., 1986;
Lehoczki et al., 1992). Similar to our results, transgenic poplars did not show enhanced tolerance
to photo-oxidative stress caused by paraquat in a study of Will et al. (2001). The non bleaching
effect observed in the dark-incubated leaf discs (Figure 1) confirmed the mode of action of
paraquat, which primarily targets the PETC of chloroplasts without effecting the other organelle
electron transport chain METC (mitochondrial electron transport chain) (Gyulai et al., 2005;
Bittsánszky et al., 2006).
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 14
As a result of the dual expression of the poplar gene gsh1 and the prokaryotic transgene gshI, the
GSH content in both transgenic lines significantly increased compared with the GSH content of
WT (Figure 2). Due to the different transgene cassette of TRlgl6 (Noctor et al., 1998), the
chloroplast-directed GSH production was higher in TRlgl6 (296%) clone then in TRggs11
(190%) compared to WT (100%) (Figure 2). Nevertheless, under paraquat treatment (5 x 10-7
M), as a result of the paraquat-response plant defense mechanism, the GSH content of TRggs11
clone showed extreme high level (Figure 2). Similar to this GSH overproduction, the expression
level of the gshI gene (Bittsánszky et al., 2006) and the enzyme activity of glutathione S-
transferase (Gullner et al., 2001) also showed significant increment under paraquat stress in the
transgenic poplars studied here.
NO is a gaseous, water- and lipid-soluble reactive aerial nitrogen species known to act as a
biological messenger in cell signaling. In mammals, NO is released by the activity of nitric oxide
synthase (NOS, EC 1.14.13.39) (1433 amino acids), which catalyses the formation of citrulline
from L-arginine, and acts on the cardiovascular system as a vasodilator in mammals (Bian and
Murad, 2003). In plants, the role of NO was first studied more than a decade ago (Delledonne et
al., 1998; Durner et al., 1998) indicating that there are at least two main enzymatic routes for NO
production: the NOS-dependent L-arginine pathway, similar to animals, and the nitrite-
dependent pathway. The NOS enzymes, mapped first onto Arabidopsis chromosome 3
(Salanoubat et al., 2000), are functionally well characterized (Ötvös et al., 2005; Grün et al.,
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 15
2006). The nitrite-dependent pathway mainly involves nitrate reductase (NR) (EC 1.7.1.3), which
catalyses the reduction of nitrate to NO (Yamasaki et al., 2001).
NaNP, which releases NO when illuminated by a high light intensity (35 mol
photons/m2/s at 20°C for 15 h), release 5–15 µM photolytic NO (Floryszak-Wieczorek et al.,
2006). In the present study the lower level of NO release might probably due to the nutritive
media used and the relatively low light intensity applied for leaf discs grow (Figure 3).
NaNP-treatment tripled the dry weight of leaf discs of the TRggs11 at a concentration of
10-7 M (Figure 4). In a study of Wang et al. (2005), NaNP, applied exogenously up to 500 µmol
L-1, also elevated the metabolism of poplar leaves by increasing the photosynthesis rate,
photochemical efficiency of PSII, and the water content.
NO-treatment increased the GSH production of TRlgl6 and WT (Figure 5a). Although,
NO decreased the GSH content of TRggs11 clone, and the content of the GSH precursor amino
acid Cys, which showed the highest level in this clone without NO-treatment (Figure 5b). These
contradictory results might be caused by the two different transgene cassettes in TRggs11 and
TRlgl6 (Noctor et al., 1998).
Nitric oxide, released in the presence of an electron acceptor such as paraquat, binds to
thiol groups of GSH, Cys and sulphur-containing proteins, resulting in S-nitrosylated derivatives,
which can interact with cellular proteins, especially the stress-responsive signaling proteins
(Grün et al., 2006). This NO-responsive GSH production was observed in the TRggs11 clone,
which produced the highest level of GSH at 10-7 M paraquat combined with 10-7 M NaNP
(Figure 5c).
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 16
Zn homeostasis, in poplar, is maintained primarily in roots and leaves, and not in the transporting
organs of shoots and stems (Adams et al., 2011). Growth of poplars at early developmental
stages was found highly sensitive to environmental Zn, but was relatively resilient in the adult
trees (Adams et al., 2011). Unlike poplars, metallocrops like oat, barley and Indian mustard took
up high level of Zn in vivo (Ebbs and Kochian, 1998).
In our study, Zn uptake in all clones showed negative correlations with the decreasing
concentrations of ZnSO4 from 10-2 M (Figure 6). The molar concentrations of Zn and S in the
leaf discs showed not the expected 1:1 ratio, coupled with high levels of Zn exclusions, which
result indicates a separate and strictly regulated plant uptake of cationic (Zn2+) and anionic (SO42-
) moieties (Figure 6). However, TRggs11 showed a tripled Zn uptake capacity compared with
TRlgl6 and WT (Figure 7a) with the highest BCFZn value (Figure 7b).
Significantly high S uptake observed by the TRlgl6 clone at 10-3 M ZnSO4 concentration
(Figure 6), supplied sulfur for the high level of GSH synthesis with a further increment under
NO treatment (Figure 5a). Leafy cut clones of TRggs11, also showed a 1.6-fold sulfate uptake
capacity compared with WT (Herschbach et al., 2000). For a further study of Zn- and S uptake of
poplar a split-root technique (Waters et al., 1980; Marsh et al., 1985) would provide additional
information.
Plant mineral uptake, in general, relies on S-rich peptides such as GSH, metallothioneins, metal
transporter proteins and phytochelatins (Rennenberg and Brunold, 1994; Rennenberg and
Herschbach, 1995; Roosens et al., 2005). Total content of the elements measured in the poplar
leaf discs was the highest in the TRggs11 clone (Figure 7a). TRlgl6 showed the highest Na
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 17
content, which was a 133% increment compared to an in situ data of Teuber et al. (2008). The
highest BCFCu value was observed in the WT (Figure 7b). All these patterns of minerals uptake
were effected by the chelating agent EDTA supplied to the aseptic media and the plant hormones
applied, which increased the whole plant metabolisms in the leaf-disc. Without Fe-EDTA
(Haberlandt, 1902; White, 1943) and hormones (Murashige and Skoog, 1962; Gyulai et al.,
1992) plants can not grow in vitro due to chlorosis. In pot experiments (Ebbs and Kochian,
1998), using a Zn-contaminated soil, the addition of EDTA to the soil significantly increased Zn
accumulation by B. juncea but not by oat or barley. Nevertheless, barley accumulated >2 mg of
Zn plant-1, it was 2−4 times more Zn than what was observed in Indian mustard in the presence
of EDTA (Ebbs and Kochian, 1998).
As the result of the high level of Na content of each of the poplar clones (Figure 7a), CAT
activity, as a stress indicator (McClung, 1997), was measured in the leaf discs exposed to NaCl
(Figure 8). CAT scavenges H2O2, the reaction product of superoxide dismutase, which is also
generated during the processes of mitochondrial electron transport and β-oxidation of fatty acids
during the photorespiratory oxidation (McClung, 1997). In animals a single CAT isoform is
encoded by a single gene. In contrast, CAT in plants is present as multiple isoforms encoded by
three genes of a multigene super family (Yang and Poovaiah, 2002).
Due to the plant oxidative stress response mechanisms (McClung, 1997; Yang and
Poovaiah, 2002), the activity of CAT in our study increased with the increasing concentration of
NaCl (Figure 8). Significantly higher CAT activity was observed in both transgenic lines
compared with WT with an extreme level in the TRggs11 clone at 0.75% NaCl (Figure 8). This
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 18
result indicates that the presence of gshI transgene also improves effectively the plant oxidative
stress response capacity by the overproduction of GSH (Figure 2 and 5a). In poplar, a salt-
induced accumulation of GSH was also observed in poplar leaves as a result of enhanced
photorespiration (Herschbach et al., 2010).
Multiple sequence alignments and hierarchical cluster analyses (Moylan et al., 2004; Gyulai et
al., 2012a; Alzohary et al., 2012, 2013) of nucleotide sequences for gsh1 genes predicted
intensive GSH production for P. x canadensis and Salix sachalinensis similar to poplar, with
potential use for broadening the tools of phytoremediation (Figure 9),
5. Conclusions
To conclude, due to the aseptic in vitro conditions, the results presented gave insight into the
absolute genetic potential of WT and TR poplars exposed to common salt, zinc sulfate, the
herbicide paraquat and nitric oxide. The study of the EDTA-stimulated mineral uptake potentials
in vitro also excluded the environmental cross reactions, and indicated that WT and the two 35S-
gshI transgenic poplars might be useful for different phytoextraction purposes in situ: WT poplar
for Cu uptake; TRggs11 for uptake of Zn, Mn and Mo; and TRlgl6 for uptake of sulfate and Na.
Comparative in silico DNA sequence analyses of plant gsh1 genes indicated further species of
the Salicaceae family with a putatively high GSH production capacity and their possible uses for
phytoextraction under conditions where P. x canescens cannot be grown. Our results may
contribute to an integrated approach of soil remediation and the development and
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 19
implementation of new bioremediation technologies that make use of plants to remove toxic
substances from contaminated sites.
Acknowledgements
Poplar clones were kindly provided by Dr. C.H. Foyer (Newcastle-upon-Tyne, UK) and Dr. H.
Rennenberg (Freiburg University, Germany). The project was supported in part by grants of the
Hungarian Scientific Research Fund OTKA-72926. OTKA-K77641, OTKA-K72926 and
OTKA-PD-75169. All authors wish to thank the Editor-in-Chief and the two anonymous
reviewers for their valuable suggestions.
References
Adams JP, Adeli A, Hsu CH, Harkess RL, Page GP, de Pamphilis CW, Schultz EB, Yuceer C.
2011. Poplar maintains zinc homeostasis with heavy metal genes HMA4 and PCS1. J Exp
Bot 62: 3737–3752.
Aebi H. 1984. Catalase in vitro. Methods Enzymol 105: 121–126.
Altschul SF, Madden TL, Schaffer AA, Zhang JH, Zhang Z, Miller W, Lipmand DJ. 1997.
Gapped BLAST and PSI-BLAST: a new generation of protein database search programs.
Nucleic Acids Res 25: 3389–3402.
Alzohary AM, Yousef MA, Edris S, Kerti B, Gyulai G, Bahieldin A. (2012). Detection of LTR
Retrotransposons reactivation induced by in vitro environmental stresses in barley (Hordeum
vulgare) via RT-qPCR. Life Science Journal 9: 5019–5026.
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 20
Alzohary AM, Gyulai G, Jansen RK, Bahieldin A. 2013. Transposable elements domesticated and
neofunctionalized by eukaryotic genomes. Plasmid 69: 1–15.
Arisi ACM, Noctor G, Foyer CF, Jouanin L. 1997. Modification of thiol contents in poplars (Populus
tremula x P. alba) overexpressing enzymes involved in glutathione synthesis. Planta 203: 362–
372.
Bian K, Murad F. 2003. Nitric oxide (NO) – biogeneration, regulation, and relevance to human
diseases. Front Biosci 8: 264–278.
Binh DQ, Heszky LE, Gyulai G, Csillag A. 1992. Plant regeneration of NaCl-pretreated cells
from long-term suspension culture of rice (Oryza sativa L.) in saline conditions. Plant Cell,
Tissue Organ Cult 29: 75–82.
Bittsánszky A, Gyulai G, Gullner G, Kiss J, Szabó Z, Kátay Gy, Heszky L, Kőmíves T. 2009. In
vitro breeding of grey poplar (Populus × canescens) for phytoremediation purposes. J Chem
Technol Biotechnol 84: 890–894.
Bittsánszky A, Gyulai G, Humphreys M, Gullner M, Csintalan Zs, Kiss J, Szabó Z, Lágler R, Tóth Z,
Rennenberg, H, Heszky L, Kőmíves T. 2006. RT-PCR analysis and stress response capacity of
transgenic gshI-poplar clones (Populus x canescens) in response to paraquat exposure. Z
Naturforsch, C: J Biosci 61: 699–730.
Bittsánszky A, Kőmíves T, Gullner G, Gyulai G, Kiss J, Heszky L, Radimszky L, Rennenberg
H. 2005. Ability of transgenic poplars with elevated glutathione content to tolerate Zinc (2+)
stress. Environ Int 31: 251–254.
Czakó M, Feng X, He Y, Liang D, Márton L. 2006. Transgenic Spartina alterniflora for
phytoremediation. Environ Geochem Health 28: 103–110.
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 21
Combet C, Blanchet C, Geourjon C, Deléage G. 2000. NPS@: network protein sequence
analysis. Trends Biochem Sci 25: 147–150.
Creissen G, Broadbent P, Stevens R, Wellburn AR, Mullineaux P. 1996. Manipulation of glutathione
metabolism in transgenic plants. Biochem Soc. Trans 24: 465–469.
Csillag F, Pásztor L, Biehl L. 1993. Spectral band selection for the characterization of salinity status
of soils. Remote Sens Environ 43: 231–242.
Delledonne M, Xia Y, Dixon RA, Lamb C. 1998. Nitric oxide functions as a signal in plant disease
resistance. Nature 394: 585–588.
Durner J, Wendehenne D, Klessig DF. 1998. Defense gene induction in tobacco by nitric oxide,
cyclic GMP, and cyclic ADP-ribose. Proc Natl Acad Sci USA 95: 10328–10333.
Ebbs SD, Kochian LV. 1998. Phytoextraction of zinc by oat (Avena sativa), barley (Hordeum
vulgare), and Indian mustard (Brassica juncea). Environ Sci Technol 32: 802–806.
Floryszak-Wieczorek J, Milczarek G, Arasimowicz M, Ciszewski A. 2006. Do nitric oxide donors
mimic endogenous NO-related response in plants? Planta 224: 1363–1372.
George EF, Puttock DJM, George HJ. 1987. Plant culture media. Vol 1. Formulations and Uses.
Edington Exegetics Limited.
Gleba D, Borisjuk NV, Borisjuk LG, Kneer R, Poulev A, Skarzhinskaya M, Dushenkov S, Logendra
S, Gleba YY, Raskin I. 1999. Use of plant roots for phytoremediation and molecular farming.
Proc Natl Acad Sci USA 96: 5973–5977.
Grün S, Lindermayr C, Sell S, Durner J. 2006. Nitric oxide and gene regulation in plants. J Exp
Botany 57: 507–516.
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 22
Gullner G, Kömives T, Rennenberg H. 2001. Enhanced tolerance of transgenic poplar plants
overexpressing -glutamylcysteine synthetase towards chloroacetanilide herbicides. J Exp Bot
52: 971–979.
Gyulai G, Humphreys M, Bittsánszky A, Skøt K, Kiss J, Skøt L, Gullner G, Heywood S, Szabó Z,
Lovatt A, Radimszky L, Roderick H, Abberton M, Rennenberg R, Kőmíves T, Heszky L. 2005.
AFLP analysis and improved phytoextraction capacity of transgenic gshI-poplar clones (Populus
canescens L.) in vitro. Z Naturforsch, C: J Biosci 60: 523–537.
Gyulai G, Janovszky J, Kiss E, Lelik L, Csillag A, Heszky LE. 1992. Callus initiation and plant
regeneration from inflorescence primordia of the intergeneric hybrid Agropyron repens (L.)
Beauv. x Bromus inermis Leyss. cv. nanus on a modified nutritive medium. Plant Cell Rep 11:
266–269.
Gyulai G, Bittsánszky A, Gullner G, Heltai Gy, Pilinszky K, Molnár E, Kömíves T. 2012a. Gene
reactivation induced by DNA demethylation in Wild Type and 35S-gshI-rbcS transgenic poplars
(Populus x canescens). Novel plant sources for phytoremediation. J Chem Sci Tech 1: 9–13.
Gyulai G, Bittsánszky A, Pilinszky K, Gullner G, Heltai Gy, Anton A, Kőmíves T. 2012b. Boron
and Zinc uptake of Cucurbits – Field test and in silico approach. Acta Phytopathol Entomol
Hung 47: 2 75–284.
Haberlandt G. 1902. Kulturversuche mit isolierten Pflanzenzellen. Sb Akad Wiss Wien, Math-Nat
Cl, 111 Abt 1: 69–91.
Hall TA. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis
program for Windows 95/98/NT. Nucleic Acids Sym Ser 41: 95–98.
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 23
Heltai Gy, Percsich K, Fekete I, Barabás B, Józsa T. 2000. Speciation of waste water sediments.
Microchem J 67: 43–51.
Herschbach C, van der Zalm E, Schneider A, Jouanin L, De Kok LJ, Rennenberg H. 2000.
Regulation of sulfur nutrition in wild-type and transgenic poplar over-expressing γ-
glutamylcysteine synthetase in the cytosol as affected by atmospheric H2S. Plant Physiol 124:
461-474.
Herschbach C, Teuber M, Eiblmeier M, Ehlting B, Ache P, Polle A, Schnitzler JP, Rennenberg H.
2010. Changes in sulphur metabolism of grey poplar (Populus × canescens) leaves during salt
stress: a metabolic link to photorespiration. Tree Physiol 30: 1161–1173.
Hillis DM, Huelsenbeck JP, Swofford DL. 1994. Hobgoblin of phylogenetics? Nature 369: 363–
364.
Kalendar R, Lee D, Schulman AH. 2009. FastPCR Software for PCR Primer and Probe Design and
Repeat Search. Genes, Genomes and Genomics 3: 1–14.
Kampfl Gy, Kristóf K; Algaidi AA, Bayoumi HHEAF, Heltai Gy. 2007. Study of NOx and CO2
production of cultivated soil in closed microcosm experimental system. Microchem J 85: 31–38.
Kim KY, Ko HJ, Kim HT, Kim YS, Roh YM, Lee CM, Kim CN. 2007. Monitoring of aerial
pollutants emitted from swine houses in Korea. Environ Monit Assess 133: 255–266.
Koprivova A, Kopriva S, Jager D, Will B, Jouanin L, Rennenberg H. 2002. Evaluation of transgenic
poplars over-expressing enzymes of glutathione synthesis for phytoremediation of cadmium.
Plant Biol 4: 664–670.
Lehoczki E, Laskay G, Gaál I, Szigeti Z. 1992. Mode of action of paraquat in leaves of paraquat-
resistant Conyza canadensis (L.) Cronq. Plant Cell Environ 15: 531–539.
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 24
Leplé JC, Brasileiro MAC, Michel MF, Delmotte F, Jouanin L. 1992. Transgenic poplars:
expression of chimeric genes using four different constructs. Plant Cell Rep 11: 137–141.
Lloyd G, McCown BH. 1980. Commercially feasible micropropagation of mountain laurel, Kalmia
latifolia, by use of shoot-tip culture. Proc Int Plant Prop Soc 30: 421–427.
Mackay D. 1982. Correlation of bioconcentration factors. Environ Sci Technol 6: 274–278.
Marsh DB, Waters L, Ascher PD. 1985. The use of split-root technique to study Zn movement
and activity in a nodulating cowpea root system. HortScience 20: 425–427.
Metternicht GI, Zinck JA. 2003. Remote sensing of soil salinity: potentials and constraints.
Remot Sens of Environ 85: 1–20.
McClung CR. Regulation of catalases in Arabidopsis. 1997. Free Radical Biol Med 23: 489–96.
Medgyesy P, Páy A, Márton L. 1986. Transmission of paternal chloroplasts in Nicotiana. Mol
Gen Genet 204: 195–198.
Moylan EC, Bennett JR, Carine MA, Olmstead RG, Scotland RW. 2004. Phylogenetic
relationships among Strobilanthes s. L. (Acanthaceae): evidence from ITS nrDNA, trnl-f
cpDNA, and morphology. Am J Bot 91: 724–735.
Murashige T, Skoog F. 1962. A revised medium for rapid growth and bio assays with tobacco
tissue cultures. Physiol Plant 15: 473–497.
Nagarajan VK, Ebbs SD. 2010. Arsenate reductase activity in roots from the arsenic
hyperaccumulator Pteris vittata utilizes both glutathione and dithiothreitol as reductants. Plant
Biosys 14: 857–859.
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 25
Noctor G, Arisi ACM, Jouanin L, Kunert KJ, Rennenberg H, Foyer CH. 1998. Glutathione:
biosynthesis, metabolism and relationship to stress tolerance explored in transformed plants.
J Exp Botany 49: 623–647.
Ötvös K, Pasternak TP, Miskolczi P, Domoki M, Dorjgotov D, Szőcs A, Bottka S, Dudits D, Fehér
A. 2005. Nitric oxide is required for, and promotes auxin-mediated activation of, cell division
and embryogenic cell formation but does not influence cell cycle progression in alfalfa cell
cultures. Plant J 43: 849–860.
Park YW, Baba K, Furuta Y, Iida I, Sameshima K, Arai M, Hayashi T. 2004. Enhancement of
growth and cellulose accumulation by overexpression of xyloglucanase in poplar. FEBS Lett
564: 183–187.
Peuke AD, Rennenberg H. 2005. Phytoremediation. EMBO Rep 6: 497–501.
Peuke AD, Kopriva S, Stroukova L, Ivanova L, Baumbach R, Tintemann H, Krauss GJ, Wennrich R,
Rennenberg H. 2012. Metal accumulation in wildtype and transgenic poplars overproducing
glutathione on different contaminated field sites: results from a release experiment in Germany
and Russia. Environmental and Experimental Botany (submitted) EEB-D-11-00501.
Pompella A, Visvikis A, Paolicchi A, De Tata V, Casini AF. 2003. The changing faces of
glutathione, a cellular protagonist. Bioche Pharmaco 66: 1499–503.
Rennenberg H, Brunold C. 1994. Significance of glutathione metabolism in plants under stress.
Prog Bot 55: 142–156.
Rennenberg H, Herschbach C. 1995. Sulfur nutrition of trees: a comparison of spruce (Picea
abies L) and beech (Fagus sylvatica L). J Plant Nutr Soil Sci 158: 513–517.
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 26
Roosens NH, Bernard C, Leplae R, Verbruggen N. 2005. Adaptive evolution of metallothionein
3 in the Cd/Zn hyperaccumulator Thlaspi caerulens. Z Naturforsch, C: J Biosci 60: 224–227.
Salanoubat M, Lemcke K, Rieger M, Ansorge W, Unseld M, Fartmann B, Valle G, Blocker H,
Perez-Alonso M, Obermaier B, et al. 2000. Sequence and analysis of chromosome 3 of the
plant Arabidopsis thaliana. Nature 408: 820–822.
Strohm M, Jouanin L, Kunert KJ, Pruvost C, Polle A, Foyer CH, Rennenberg H. 1995. Regulation of
glutathione synthesis in leaves of transgenic poplar (Populus tremula × P. alba) overexpressing
glutathione synthetase. Plant J 7: 141–145.
Tamura K, Dudley J, Nei M, Kumar S. 2007. MEGA4: Molecular Evolutionary Genetics
Analysis (MEGA) software version 4.0. Mol Biol Evol 24: 1596–1599.
Teuber M, Zimmer I, Kreuzwieser J, Ache P, Polle A, Rennenberg H, Schnitzler JP. 2008. VOC
emissions of grey poplar leaves as affected by salt stress and different N sources. Plant Biol 10:
86–96.
Thompson JD, Higgins DG, Gibson TJ. 1994. CLUSTAL W: improving the sensitivity of
progressive multiple sequence alignment through sequence weighting, positions-specific gap
penalties and weight matrix choice. Nucleic Acids Res 22: 4673–4680.
Watanabe K, Yamano Y, Murata K, Kimura A. 1986. The nucleotide sequence of the gene for
gamma-glutamylcysteine synthetase of Escherichia coli. Nucleic Acids Res 14: 4393–4400.
Wang M, Li Q, Fu S, Dong B. 2005. Effects of exogenous nitric oxide on photosynthetic
characteristics of poplar leaves under water stress. Yingyong Shengtai Xuebao 6: 218–22.
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 27
Waters L Jr, Breen PJ, Mack HJ. 1980. Translocation of 14C-photosynthate, carbohydrate content,
and nitrogen fixation in Phaseolus vulgaris L. during reproductive development. J Amer Soc
Hort Sci 105: 424–427.
White PR. 1943. Nutrient deficiency studies and an improved inorganic nutrient medium for
cultivation of excised tomato roots. Growth 7: 53–65.
Will B, Jouanin L, Rennenberg H. 2001. Protection from paraquat-mediated photo-oxidative stress
by glutathione in poplar (Populus tremula x P. alba) plants. Plant Biol 3: 272–278.
Yamaguchi T, Blumwald E. 2005. Developing salt-tolerant crop plants: challenges and opportunities.
Trends Plant Sci,10: 615–620.
Yamasaki H, Shimoji H, Ohshiro Y, Sakihama, Y. 2001. Inhibitory effects of nitric oxide on
oxidative phosphorylation in plant mitochondria. Nitric Oxide 5: 261–270.
Yang T, Poovaiah BW. 2002. Hydrogen peroxide homeostasis: Activation of plant catalase by
calcium/calmodulin. Proc Natl Acad Sci USA 99: 4097–4102.
Zarcinas BA, Cartwright B, Spouncer LR. 1987. Nitric acid digestion and multi-element analysis of
plant material by inductively coupled plasma spectrometry. Commun Soil Sci Plant Anal 18:
131–146.
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 28
Figure Captions
Figure 1. Effects of light and dark incubation and sucrose concentration on bleaching of
poplar leaf discs by paraquat (PQ) (4 x 10-7 and 4 x 10-6 M). Leaf discs of wild-type (WT) and
the 35S-gshI transgenic poplar (Populus × canescens) lines TRggs11 and TRlgl6 were treated for
21 days on aseptic WPM media.
Figure 2. Glutathione (GSH) production in response to paraquat (PQ) exposure in poplar
leaf discs. GSH content was measured in leaf discs of wild-type (WT) and 35S-gshI transgenic
poplar (Populus × canescens) lines TRggs11 and TRlgl6 after treatment with 10-8 to 5 × 10-7 M
paraquat (PQ) for 21 days on aseptic WPM media. Mean values ± standard deviation are
indicated (n = 3).
Figure 3. Time course of nitric oxide [NO] release from NaNP (10-7 M) supplied in aseptic
WPM nutritive media and incubated under either a 16 h/8 h (light/dark) photoperiod or in
the dark. Mean values ± standard deviation are indicated (n = 3).
Figure 4. Concentration-dependent effect of NaNP on the leaf disc dry weight (DW). Leaf
discs of wild-type (WT) and 35S-gshI transgenic poplar (Populus × canescens) lines TRggs11
and TRlgl6 were treated with 10-7 to 10-4 M NaNP for 21 days added to aseptic WPM media.
Mean values ± standard deviation (n = 3) are indicated.
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 29
Figure 5. Glutathione (GSH) and cysteine (Cys) contents in leaf discs in response to [NO]
released from NaNP and paraquat exposure. (a) Glutathione (GSH) content and (b) cysteine
(Cys) content in leaf discs of wild-type (WT) and 35S-gshI transgenic poplar (Populus ×
canescens) lines TRggs11 and TRlgl6 exposed to 10-7 M NaNP for 21 days. (c) Effect of
combined treatment of NaNP (10-7 M) and paraquat (PQ) (10-8 to 5 × 10-7 M) on GSH
production after 21 days treatment on aseptic WPM medium. Mean values ± standard deviation
are indicated (n = 3).
Figure 6. Zinc (Zn) and sulfur (S) contents, and bioconcentration factor (BCF) ratios for
Zn and S. Leaf discs of wild-type (WT) and 35S-gshI transgenic poplar (Populus × canescens)
lines TRggs11 and TRlgl6 were exposed to a concentration series of ZnSO4 (10-2 to 10-4 M) for
21 days on aseptic WPM medium. Basal WPM medium contained 1.955 mg L-1 Zn2+ and 14.36
meq L-1 SO42-. Mean values ± standard deviation indicated (n = 3).
Figure 7. Mineral uptake capacity and bioconcentration factor (BCF) ratios of poplar leaf
discs. (b) Mineral uptake capacity of leaf discs of wild-type (WT) and 35S-gshI transgenic
poplar (Populus × canescens) lines TRggs11 and TRlgl6. Mean values ± standard deviation (n =
5) are indicated. (b) BCF ratios of Na, Zn, Mn, Cu, and Mo, and Basal WPM nutritive media
contained Na (4.59 mg L-1), Zn (1.955 mg L-1), Mn (7.248 mg L-1), Mo (0.099 mg L-1) and Cu
(0.063 mg L-1).
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13

ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 30
Figure 8. Catalase activity of leaf discs of wild-type (WT) and 35S-gshI transgenic poplar
(Populus × canescens) lines TRggs11 and TRlgl6 exposed to different concentrations of
NaCl for 21 days on aseptic WPM media. Mean values ± standard deviation (n = 3) are
indicated.
Figure 9. Bootstrap consensus ML (Maximum Likelihood) (Hillis et al., 1994) dendrogram
(Tamura et al., 2004) derived from nucleotide sequences of plant gsh1 genes with outgroup
of prokaryotic (E. coli) trangene gshI (Watanabe et al., 1986). GeneBank accession numbers
are specified for each accession. Bootstrap support values from 1000 replicates are provided
below each node. The scale bar (0.1) represents relative genetic distances with the numbers of
nucleotide substitutions per site of sequences. Plant families are indicated.
Fig. 1.
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 31
Fig. 2.
Light
Dark
Sucrose 1.0 % Sucrose 2.0 %
Not Treated Not Treated 4.0 x 10-7 M PQ 4.0 x 10-6 M PQ 4.0 x 10-7 M 4.0 x 10-6 M PQ
TRggs11
TRlgl6
WT
TRggs11
TRlgl6
WT
264.5174.1
298.1504.8 687.9504.8 328.6
579.3
380.1354.5270.8
331.2
625.3946.5
1932.1
0
400
800
1200
1600
2000
Not Treated 10-8 M PQ 5 x 10-8 M PQ 10-7 M PQ 5 x 10-7 M PQ
nmol
GSH
x g
-1 F
W le
af t
issu
e WTTRlgI6TRggs11
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 32
Fig. 3.
250
032
204
11497
0
149
195
0
225
35
0
41
300
217
00
236
191
220233
5982
102
188
63
33
0
50
100
150
200
250
300
350
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27Days
[NO
] rel
ease
(ng
L-1)
LightDark
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 33
Fig. 4.
0.51.3
3.83.03.0
3.9
8.1
10.2
0.4
2.33.9
3.03.30.61.8
0
2
4
6
8
10
12
Not Treated 10-7 M 10-6 M 10-5 M 10-4 M NaNP [NO]
Leaf
dis
cs (m
g D
W)
WTTRggs11TRlgl6
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 34
Fig. 5a.
Fig. 5b.
174.1
331.2
504.8
272.3 245.1
728.6
0
100
200
300
400
500
600
700
800
WT TRggs11 TRlgI6
Clones (Populus x canescens )
nmol
GSH
g-1
FW
leaf
tis
sue Not Treated
NaNP [NO] 10-7 M
17.6
27.7
18.1
27.2
10.8
23.6
0
5
10
15
20
25
30
35
WT TRggs11 TRlgI6Clones (Populus x canescens )
nmol
Cys
g-1
FW
leaf
tis
sue
Not TreatedNaNP [NO] 10-7 M
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 35
Fig. 5c.
Fig. 6.
397
1985
491
15140
410
1327
410
281374
11711165
906
255
889
340 36
0.612 2.81.30.8220.68
3 413
3
106
88
117
0
200
400
600
800
1000
1200
1400
10-2 10-3 10-4 10-2 10-3 10-4 10-2 10-3 10-4
WT TR (ggs11) TR (lgl6)
ZnSO4 (mol)
- nm
ol Z
n an
d S
g-1
DW
leaf
dis
c -
0
20
40
60
80
100
120
140
- BC
F -
Zn (nmol)S (nmol)BCF-ZnBCF-S
791.0
263.0324.0277.0
429.0432.0
689.0
1093.0
589.0
608.0514.0
425.0
0
200
400
600
800
1000
1200
10-8 M PQ and 10-7 M NaNP
5x10-8 M PQ and 10-7 M NaNP
10-7 M PQ and 10-7 M NaNP
5x10-7 M PQ and10-7 M NaNP
nmol
GSH
x g
-1 F
W l
eaf
tissu
e
WTTRggs11TRlgl6
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 36
Fig. 7a.
Na; 2489.1
Na; 1888.3Na; 1702.2
Zn; 820.6
Zn; 241.6
Zn; 291.6
Mn; 240.4Mn; 324.1
Mn; 199.1
Mo; 26.2
Mo; 37.7 Mo; 24.5
Cu; 48.4
Cu; 10.8 Cu; 17.4
0
500
1000
1500
2000
2500
3000
3500
WT TRggs11 TRlgl6Clones (Populus x canescens )
mg
kg-1 D
W le
aves
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 37
Fig. 7b.
Fig. 8.
542.3411.4
Na; 370.8
123.6
419.7
Zn; 149.1
33.2Mn; 27.5 44.7
247.0381.2
Mo; 265.0
275.4171.3
Cu; 767.6
0
150
300
450
600
750
900
WT TRggs11 TRlgl6Clones (Populus x canescens )
BCF
239.
1
703.
2
535.
2
333.
4
911.
3
357.
1
382.
6
451.
8 621.
9
65.1
42.3
24.363
.1
23.6 87
.5
0
150
300
450
600
750
900
1050
WT TRggs11 TRlgI6Clones (Populus x canescens )
µmol
H2O
2 g-1 F
W mi
n-1
Not Treated0.75 % NaCl1.5 % NaCl2.25 % NaCl3.0 % NaCl
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13
ACCEPTED MANUSCRIPT
ACCEPTED MANUSCRIPT 38
HQ658456 Populus x canadensis (gsh1)
XM002297999 Populus trichocarpa
HQ228352 Salix sachalinensis
Salicaceae
AF017983 Lycopersicon esculentum
DQ444219 Nicotiana tabacum (cpDNA-gsh1)Solanaceae
Fagaceae DQ787258 Fagus sylvatica
AF128453 Glycine max
AF128454 Phaseolus vulgaris
AY204516 Lotus corniculatus
AF128455 Pisum sativum
AF041340 Medicago truncatula
Fabaceae
Y10848 Brassica juncea
NM118439 Arabidopsis thaliana
Z29490 A thaliana (cpDNA-gsh1)
Cruciferae
DICOTs
AJ302783 Zea mays
AJ508916 Oryza s japonica
AY864064 Triticum aestivum
MONOCOTs - Poaceae
MONOCOTs - Liliaceae AF401621 Allium cepa
PROKARYOTEs X03954 E coli (gshI)
99
98
89
100
77
93
77
65
94
77
90
41
58
45
21
45
0.1
Fig. 9.
Dow
nloa
ded
by [
Tam
as K
omiv
es]
at 0
1:57
30
Apr
il 20
13





![Effect of heat-shock and bile salts on protein synthesis of Bifidobacterium longum revealed by [ 35S]methionine labelling and two-dimensional gel electrophoresis](https://img.dokumen.tips/doc/110x75/631df96d56cbbb4750055315/effect-of-heat-shock-and-bile-salts-on-protein-synthesis-of-bifidobacterium-longum.jpg)

![Functional correlation between allopregnanolone and [35S]-TBPS binding in the brain of rats exposed to isoniazid, pentylenetetrazol or stress](https://img.dokumen.tips/doc/110x75/635e0f7d095e4caf22060a5f/functional-correlation-between-allopregnanolone-and-35s-tbps-binding-in-the-brain.jpg)









![Potential problems in using [35S]-dATP-tailed oligonucleotides for detecting mRNAs in certain cells of the immune system](https://img.dokumen.tips/doc/110x75/635a64f94028fc5adf013378/potential-problems-in-using-35s-datp-tailed-oligonucleotides-for-detecting-mrnas.jpg)











