Embed Size (px)
Citation preview

1
UNIVERSITY OF CAPE COAST
SCHOOL OF BIOLOGICAL SCIENCES
PHYLOGENETIC ANALYSES OF THE BEE TRIBE ANTHIDIINI AND REVISION OF THE AFROTROPICAL GENUS
Anthidiellum (COCKERELL)
BY
ROFELA COMBEY
Thesis submitted to the Department of Entomology and Wildlife of the School of Biological Sciences, University of Cape Coast in partial
fulfillment of the requirements for the award of Doctor of Philosophy in Zoology
JUNE 2008

2
DECLARATION
Candidate’s declaration
I hereby declare that this thesis is the result of my own original work
and that no part has been presented for another degree in this
university or elsewhere.
Candidate’s Signature:_____________________
Name: Rofela Combey …………………….Date:_________________
Supervisors’ Declaration
We hereby declare that the preparation and presentation of the thesis
were supervised in accordance with the guidelines on supervision of
thesis laid down by the University of Cape Coast.
Principal Supervisor’s Signature:____________________
Name: Dr. Peter Kofi Kwapong Date:_____________
Co-Supervisor’s Signature:__________________________
Name: Professor (Mrs) Mary Botchey Date:____________
Co-Supervisor’s Signature:___________________
Name: Dr. Connal Desmond Eardley Date:___________

3
CHAPTER ONE
INTRODUCTION
The Concept of Phylogeny
There are so many different kinds of organisms on Earth that are
believed to exist as a result of evolution. If this evolutionary history or the
phylogeny of an organism is traced back, it connects through shared ancestors
to lineages of other organisms (Lipscomb, 1998). The scientific method that
analysizes and groups organisms that share derived characters is called
cladistic or phylogenetic systematics. This important aspect of systematics
was first published by Henning (1966) who argued that only shared derived
characters could possibly give us information about phylogeny.
The principle behind phylogenetics is that, organisms should be
classified according to their evolutionary relationships, and these relationships
are analysized based on characters occurring in either the primitive or advance
state. Primitive (plesiomorphic) characters are those characters in which all
members of the taxa possess. However these characters do not reveal the
relationship within a taxa. Advanced (apomorphic) characters on the other
hand are derived traits which appear in some members of the taxa and reveal
phylogenetic relationships among the taxa (groups). Taxa that share many
derived characters are grouped more closely together than those that do not
share similarly derived characters. The relationships are shown in a branching
hierarchical tree called a cladogram (Lipscomb, 1998; Michener, 2000).

4
This phylogenetic tree or cladogram provides the basis of
evolutionary interpretation; it also provides the classification by which other
biologists communicate information about an organism; the cladogram and its
corresponding classification predicts properties of newly discovered or poorly
known organisms (Lipscomb, 1998; Michener, 2000).
Why Bees?
Among the hymenopteran insects, bees occupy a unique position in the
world of Arthropods. They are entwined into most aspects of human culture
and mythology, not to mention our agriculture, economy and general ecology.
Several centuries ago, bees were used to instil in man ideal notions of
industriousness, loyalty, obedience and sacrifice not through denigration but
through almost holy reverence to a greater good. Even the famous parable
from the Bible teaching us to “go to the ant” originally included reference to
the labour of bees (“Go to the bee, and learn how diligent she is, and what a
noble work she produces, whose labours kings and private men use for their
use, she is desired and honoured by all, and though weak in strength she
values wisdom and prevails:” Proverbs, Chapter 6, Septuagint version-i.e.,
oldest Greek version and perhaps the oldest extant version of the Holy Bible).
It is believed that such emotions and concepts connected to bees flow through
peoples’ common natural heritage; even the great “Mother Goddess”
uncovered at the root of most Western religions were symbolized by bees
(Engel, 2000, 2001).

5
Today, bees are known by far to be major contributors to essential
ecological services including pollination. Globally, pollination and pollinator
issues have occupied keystone position in maintenance of biodiversity, in both
natural and agro-ecosystem. Pollination services provided by most bee
pollinators appear to be different from most other essential ecosystem services
in that, it is biodiversity dependent; for example many plant species require
specific pollinators and similarly many pollinator species require specific food
plant (Constanza, 1987; O’Toole, 1993; Kevan, et. al., 2002 and Fact sheet,
2004). Bees are almost widely used in pollination management in agriculture
(Prescott-Allen and Prescott-Allen, 1990; Ingram et. al., 1996; Kenmore and
Krell, 1998). Aside this essential service, hive products provide enormous
economic value to mankind.
In view of these essential roles played by bees, it is important to study
the systematics of existing bee fauna and document their role in ecosystem
maintenance.
Taxonomy of Bees
Bees are insects of the superfamily Apoidea of which there are seven
major families. They are among the major groups of aculeate hymenopterans
(ants and wasps), in which most females have a modified ovipositor as a
stinging apparatus for defence or hunting or both. They differ from the other
hymenopteran insects by possessing small, distinct pronotal lobe that is well
seperated from and is below the tegula, and an extension of the pronotum
ventro-laterally as a pair of processes, one on each side that encircle or nearly

6
encircle the mesosoma behind the forecoxa (Michener and Griswold, 1994;
Michener, 2000). The major families of bees include Stenotritidae, Colletidae,
Andrenidae, Halictidae, Melittidae, Megachilidae and Apidae (Michener,
2000). Several subfamilies, tribes, genera and species have been erected for
the bees of the world. Globally, over 30,000 species have so far been named,
and there are more yet to be identified.
The Tribe Anthidiini Fabricus
The Anthidiini is one of four tribes of the subfamily Megachilinae and
family Megachilidae. The others are: Megachiliini, Dioxyini and Osmiini.
This is an enormous subfamily with thousands of species that are commonly
called leaf-cutter, carder and mason bees. The anthidine bees exhibit
considerable variation in form from heriadiform, hoplitiform,
chalicodomiform, megachiliform to anthophoriform. Members of this tribe
mostly have yellow integumental markings (O’Toole and Raw, 1991;
Michener and Griswold, 1994; Michener, 2000).
According to Michener (2000), the nests of the Anthidiini are mostly
on exposed rocks, stems and leaves, or in pre-existing burrows and in their
own burrows. The entire nests (either exposed or in cavities) are constructed
from resin, sometimes mixed with or cementing together with soil particles,
pieces of leaves, chaff and plant hairs or fibers (Plate 1).

7
Plate 1a, b, c, d, e, f, g and h. Anthidine bee nests and cell (s). (a) Nest of
Serapista denticulata (arrows indicating nest material made of pieces of
leaves, chaff and plant hairs). (b and c) Dorsal and anterior views of the nest
of S. denticulata (arrow indicating nest entrance). (d and e) Nests of
Afranthidium immaculatum showing nest material made of soil cemented
together with resin and plant fibers. (f, g and h) Arrows showing cells of A.
immaculatum covered with plant fibers.
a
b
c d
e
f
g h
d

8
The tribe consist of thirty seven genera (see appendix 1), each with between
one and twelve subgenera.
Anthidine bees are widely distributed, occurring on all continents,
except Antarctica, and fairly common, except with only one species known
from Australia. The tribe largely comprises pollen collecting bees; the few
parasitic groups (Afrostelis, Euaspis, Hoplostelis and Larinostelis) have
narrow ranges of host taxa within the families Apidae (tribe Euglossini) and
Megachilidae (genus Megachile, Latreille) (Michener and Griswold, 1994,
Michener, 2000).
The non-parasitic genera are solitary, pollinating bees. Armbuster and
Steiner (1992) recorded two species of Pachyanthidium (Vachal) as the
principal pollinators of four species of the plants genus Dalechampia in South
Africa. Hence, it is essential to assess the phylogenetic relationship among
members of this tribe, in order to ascertain its classification status.
Justification of Research
Central to any biodiversity-dependent research is a sound knowledge
of what species are involved, particularly in pollination and pollinator studies.
This is the most basic pre-requisite for understanding broader ecological
systems as well as characterizing attributes of identified species.
However, Africa which is one of the biodiversity-rich continents in the
tropics, where a great deal of work on conservation and sustainable use of
biodiversity is urgently required is limited in resources, including taxonomic
expertise, to find out what species live within its boundary. This situation is

9
not different in Ghana, where data on bee fauna seem to be non-existent. Few
of such data that exist are limited to only individual institutions within the
country. Bee pollinators like many other ecologically important animals, have
not received the needed taxonomic attention due to lack of capacity to
effectively and accurately identify existing fauna. Furthermore, virtually
nothing is known about the taxonomy and ecology of the bees that pollinate
Ghana’s food crops and wild plant species.
In the Western Palaearctic Region and South Africa, members of the
bee tribe Anthidiini have been recorded as principal pollinators of certain
plant families including the Labiatae, the Compositae, the Leguminosae and
the Zygophyllaceae (Armbuster and Steiner, 1992; Müller, 1996). In Ghana,
members of this bee tribe are known to be the principal pollinator of plants
belonging to the mint family (Labiatae) and probably many other food crops
and wild plants.
Unfortunately, many authors such as Pasteels (1984), Michener and
Griswold (1994) and Michener (2000) have questioned the generic status of
most members of the tribe. In addition, data on the phylogeny of all the
anthidine genera of the world is fragmentary and very limited to few
geographic regions. It is therefore important to study the phylogenetic
relationship among members of this tribe, in order to ascertain and probably
postulate a better classification system for the tribe.

10
Objectives
Based on these taxonomic gaps in the world and Ghana in particular, the
research was principally conducted under 5 spectrums, in which the research
would seek to first and foremost, ascertain the monophyly of the Anthidiini
and evaluate the relationships among the genera as well as, revise the generic
classification of the tribe. In addition, cladistic analysis for the Anthidiini of
the world will be conducted in order to postulate their probable phylogeny or
evolutionary history. Furthermore, this research would revise classification of
the Afrotropical species of the genus Anthidiellum Cockerell and provide a
taxonomic key for the identification of the Afrotropical species of the genus
Anthidiellum.

11
CHAPTER TWO
LITERATURE REVIEW
History of Classification of the Anthidiini within the Apiformes
The history of the classification of this group of closely related bees
appears in zoological records in the following chronology. An early attempt to
classify the group into higher taxa was made by Fabricus (1804). He named
the first anthidine genus, as Anthidium, and placed it in the solitary division of
the family Apiariae. Later, Lepeletier (1835, 1841) placed Anthidium
(Fabricus) with the solitary-nesting bees of the family Gastrilegides, along
with Chelostoma (Latreille), Lithurgus (Berthold) and Megachile. Schenck
(1861, 1869) subsequently placed the genus in the subfamily Megachilinae
(family Megachilidae) together with Lithurgus and Megachile. Thompson
(1872) placed Anthidium, Coelioxys (Latreille), Megachile and Stelis (Panzer)
in the tribe Megachilini (subfamily Megachilinae).
Schmiedeknecht (1882) and Friese (1895), however, used a system of
classification similar to Lepeletier (1841), placing the genus Anthidium in the
section with solitary nest-making bees of the family Megachilidae, and Stelis
with the parasitic bees in the family Stelidae. Ashmead (1899) greatly
modified Schmiedeknecht’s system, separating Anthidium from the subfamily
Megachilinae (Lithurgus, Megachile and Ctenoplectra Kirby, which is today
regarded as being in the Apidae) and placed it in the subfamily Anthidiinae of

12
the family Megachilidae. He also placed Stelis and Coelioxys in separate
subfamilies, namely Stelidinae and Coelioxinae, both in the family
Megachilidae.
Robertson (1903 and 1904) in his classification of bees placed
Anthidium, Stelis and Trachusa Panzer in the family Stelidae (Anthidium and
Stelis in the subfamily Anthidiinae, and Trachusa in the subfamily
Trachusinae), both in the division Apygidialia. He based his classification on
the assumption that the pygidial plate has been lost repeatedly and
independently. In the same year, Bingham, (1903) synonymized Anthophora
gastrica (Illig), Stelis rufiventris Lepeletier, Anthidium abdominale (Smith)
and Euaspis abdominalis (Gerstaecker). E. abdominalis is presently the valid
name.
Michener (1944) provided a comprehensive classification of these
bees. He placed Anthidium, Dioxys Lepeletier and Serville, and Stelis in the
tribe Anthidiini (Megachilidae: Megachilinae). Michener and Griswold (1994)
and Michener (2000), however excluded Dioxys from the tribe Anthidiini. The
present genera of tribe Anthidiini are listed in appendix 1, and have been
grouped according to the number of female mandibular dentition.
Based on what appears to be inconsistency in the classification of the
tribe, it is vital to review the present classification system of the Anthidiini
using phylogenetic analysis.

13
Overview of all the Anthidine Genera
Various authors have worked on the tribe in different geographical
regions. An account on general shape of taxa, foraging status (parasitic/non-
parasitic), history of classification, geographic distribution, as well as generic
status may be outlined as follows for each of the thirty seven genera.
Genus Acedanthidium is a slender, megachiliform, non-parasitic genus
which was first described by Michener and Griswold (1994). Later, Michener
(2000) provided brief diagnosis on this monotypic genus, which is endemic to
India.
Afranthidium is non-parasitic, small to medium sized, megachiliform
genus which was first described by Michener (1948). Later, Michener (2000)
recorded 11 subgenera in this genus; all are Afrotropical, and two of these also
occur in the Palaearctic Region.
Afrostelis is one of the anthidine bees which is endemic to Africa. This
cleptoparasitic, hoplitiform genus is monotypic. It was first described by
Cockerell (1931b). Later, Pasteels (1969, 1984), Baker (1996a), Michener and
Griswold (1994), Michener (2000) and Pauly (2001), provided diagnoses of
the genus..
The genus Anthidiellum was first described by Cockerell (1904a).
Later, Pasteels (1969) recorded six subgenera; two of these are endemic to the
Afrotropical Region, one is Afrotropical and Oriental, two are endemic to the
Orient and one is Mesoamerica and Holarctic. This genus consists of small,
robust, megachiliform to bombiform, non-parasitic bees.

14
Anthidioctes--This genus was first described by Holmberg (1887,
1903). Later, Moure (1947a) redescribed an additional subgenus. It is known
that this small hoplitiform to slender megachiliform genus of pollen-collecting
bees is endemic to the Neotropical Region with a few species also occurring
in southern Africa.
Anthidioma--This non-parasitic, chalicodomiform genus is represented
by two species; namely, A. chalicodomoides Pasteels and A. murinum
Pasteels. These were first described by Pasteels (1984). Later, Michener and
Griswold (1994) and Michener (2000), provided a brief diagnosis of this
genus. The genus is endemic to southern Africa.
Fabricus (1804) first described the genus Anthidium from the type
species Apis manicata (Linnaeus). Michener (2000) listed seven subgenera.
Three of these subgenera are Palaearctic, two are endemic to Africa, one
occurs in the Nearctic Region, and the other subgenus occurs on all
continents, except Australia and it also does not occur in the Indo-Malayan
tropics. This genus comprises of small to medium size, megachiliform, pollen-
collecting bees.
Apianthidium--This megachiliform, non-parasitic genus was first
described by Pasteels (1969). It is monotypic, and occurs in Borneo.
Aspidosmia--This monotypic genus was first described by Brauns
(1926) as a subgenus of genus Osmia due to its striking morphological
similarity with species of Osmia s. str. Later, Peters (1972a) gave Aspidosmia
generic status. This genus comprises mostly of robust megachiliform, non-
parasitic bees that are endemic to southern Africa.

15
Atropium--This robust megachiliform, non-parasitic genus was first
described by Smith (1853). It is known by a single female specimen of
Atropium atropos (Smith), which is endemic to Gambia.
Aztecanthidium--The genus was described first by Michener and
Ordway (1964). The only three representative species were revised by
Snelling (1987). Michener (2000) later provided a brief diagnosis on the
genus. This large-sized, chalicodomiform, pollen-collecting genus is endemic
to Mexico.
The genus Bathanthidium was first described by Mavromoustakis
(1953). Later, other authors, such as Pasteels (1968a, 1969), Romankova
(1988), and Michener and Griswold (1994) worked on the genus. Michener
(2000) listed three subgenera. Two of these subgenera are endemic to the
Oriental Region and the other occurs in the eastern Palaearctic Region. This
genus consists of small hoplotiform to slender megachiliform, non-parasitic
bees.
Pasteels (1969) described the genus Benanthis for a species that was
originally described by Benoist (1962) in the genus Anthidium. It is still the
only described species in this genus of medium to large megachiliform, non-
parasitic bees. It is endemic to Madagascar.
Cyphanthidium--This non-parasitic, medium sized, megachiliform
genus is represented by two species. It was first described by Pasteels (1969),
and it is known to comprise two representative species. Cyphanthidium is
endemic to southern Africa.

16
Dianthidium--This genus was first described by Cockerell (1900).
Later, other authors such as, Moure (1965), Griswold and Michener (1988),
and Michener (1942, 2000) worked on the group. Michener (2000) recorded
four subgenera. One is endemic to the Nearctic Region; two are only known
from Mexico, and two occur in South Africa. It consists of mostly small to
medium sized, chalicodomiform, pollen-collecting bees.
Duckeanthidium--This non-parasitic, robust, chalicodomiform genus
was first described by Moure and Hurd (1960). Later, five species were
described by Urban (1995). Michener (2000) synonymized Grafanthidium and
Duckeanthidium. All of the species are endemic to Brazil.
Eoanthidium--Popov (1950) first described this non-parasitic, small
megachiliform to slender bombiform genus. Michener (2000) listed four
subgenera, two of which are endemic to Africa and the Palaearctic Regions.
The other two occur in India.
Epanthidium--Moure (1947a) first described this non-parasitic genus.
Later, Stange (1983), Moure and Urban (1990) Urban (1991) and Michener
(2000) worked on this genus. Michener (2000) recorded three subgenera, two
are endemic to South America and the other occurs in both South America
and Mexico. It comprises small chalicodomiform bees.
Euaspis--This cleptoparasitic, medium to large, chalicodomiform
genus was first described by Gerstaecker (1857). Since then, other authors,
such as Fairmaire (1858), Vachal (1903), Friese (1909a, 1912b, 1914, and
1921) and Cockerell (1933e), worked on this genus. Michener (2000)
recorded 12 species two were listed as Afrotropical. The other ten are
Oriental.

17
Gnathanthidium--This non-parasitic genus was first described by
Pasteels (1969). It is monotypic and endemic to eastern and southern Africa.
Hypanthidiodes--This genus was first described by Moure (1947a).
Michener (2000) listed ten subgenera, all of which are Neotropic. This non-
parasitic genus comprises small to medium-sized chalicodomiform or
hoplitiform bees.
The genus Hypanthidium was first described by Cockerell (1904b).
Later, Urban (1994a) revised this genus. Michener (2000) listed two
subgenera, all of which are endemic to the Neotropical Region. It consists of
large chalicodomiform bees.
Hoplostelis--This cleptoparasitic genus was first described by
Dominique (1898). Other authors, such as Griswold and Michener (1988),
Moure and Urban (1994) and Michener (2000), worked on this genus.
Michener (2000) listed three subgenera, all of which are Neotropical. This
genus consists of mostly medium to large sized, hoplitiform to
chalicodomiform bees.
Icteranthidium--This genus was first described by Latreille (1809).
Later, other authors, such as Michener (1948) and Pasteels (1969, 1984)
worked on this genus. According to Michener (2000), there are about 25
species, 15 species occur in the West Palaearctic Region, two are endemic to
the Afrotropical Region and the other eight, are Oriental.
Indanthidium was first described by Michener and Griswold (1994).
This monotypic genus consists of small to medium-sized, megachiliform,
pollen-collecting bees that are endemic to southern India.

18
Larinostelis--This genus was first described by Michener and
Griswold (1994). It is represented by only one described species Larinostelis
scapulata Michener and Griswold that is endemic to Kenya
The genus Neanthidium was first described by Pasteels (1969). It is
represented by one species Neanthidium octodentatum (Pasteels), which
occurs in North Africa (Algeria, Morocco).
Notanthidium--This slender, hoplitiform genus of pollen-collecting
bees was first described by Isensee (1927). Later, Moure (1947a) and Urban
(1997b) worked on the genus. Michener (2000) listed three subgenera, all of
which are endemic to South America.
Pachyanthidium was first described by Friese (1905). Since then, other
authors, such as Cockerell (1930b), Pasteels (1969) and Michener and
Griswold (1994), worked on this group. Michener (2000) recorded four
subgenera. Two are endemic to Africa, one occurs in both Africa and Arabia,
while the other occurs in both Africa and the Orient. This genus comprises
non-parasitic, robust megachiliform bees.
Paranthidium was first described by Cockerell and Cockerell (1901).
Michener (1948) described an extra two subgenera in this genus (namely
Rapanthidium and Mecanthidium), but later Michener (2000) moved
Mecanthidium to the genus Dianthidium. One of the subgenera is endemic to
Mesoamerica and the other occurs in both Mesoamerica and the Nearctic
Region.

19
Pleisianthidium--This genus was first described by Cameron (1905).
Since then, other authors, such as Mavromoustakis (1951), Pasteels (1969,
1984) and Michener and Griswold (1994), have revised the group. Michener
(2000) listed four subgenera, all of which are endemic to South Africa.
Pseudoanthidium was first described by Friese (1898). Michener
(2000) recorded six subgenera, two of which are endemic to Africa, three are
known from the Palaearctic and Orient regions. The other subgenus occurs in
both Africa and Arabia.
The genus Rhodanthidium was first described by Iseense (1927).
Michener (2000) recognized three subgenera, all of which occur in the
Palaearctic Region, however, one also occurs in the Oriental Region.
Serapista--This robust, megachiliform to chalicodomiform, pollen-
collecting genus was first described by Smith (1854) as Serapis. However,
because Serapis was preoccupied under phyllum mollusca, Cockerell (1904)
renamed this genus as Serapista Cockerell. Other authors, such as
Mavromoustakis (1940) and Pasteels (1969, 1984), worked on this genus.
Pasteels (1984) recorded four species all of which are endemic to Africa.
Stelis--This cleptoparasitic genus was first described by Panzer (1806).
Since then several other authors, such as Provancher (1888), Ashmead (1898),
Robertson (1902), Noskiewicz (1961) and Michener and Griswold (1994),
revised this genus. Michener (2000) listed six subgenera. Two subgenera are
endemic to the Orient, two occur in Mesoamerica, with one of these also
occurs in the Palaearctic Region, which has an additional endemic genus. The

20
last subgenus is Holarctic. Michener (2000), however, mentions that its
distribution extends into Kenya, yet Eardley and Urban (in preparation) do not
record a Kenyan species but did record one species from South Africa.
Trachusa--This parasitic genus was first described by Panzer in 1806.
Since then several other authors, such as Provancher (1888), Ashmead (1898),
Robertson (1902), Noskiewicz (1961) and Michener and Griswold (1994),
revised this genus. Michener (2000) listed six subgenera. Two subgenera are
endemic to the Orient, two occur in Mesoamerica, with one of these also
occurs in the Palaearctic Region, which has an additional endemic genus. The
last subgenus is Holarctic. Michener (2000), however, mentions that its
distribution extends into Kenya, yet Eardley and Urban (in preparation) do not
record a Kenyan species but did record one species from South Africa.
Trachusoides--This non-parasitic, large megachiliform genus was first
described by Michener and Griswold (1994). This monotypic genus is
endemic to southern India.
Phylogenetic Position of Bees
History and Evolution of the Apoidea
Among the bee genera, phylogenetic procedures have been employed
to provide evidence of close relationship to the sphecoid wasps Comstock
(1942). Fossil record on bees is known to be limited. It has been said that data
has been biased towards resin-collecting bees which were occasionally
trapped in the resin and were later fossilized in amber (a hard translucent
fossil resin varying in colour from yellow to light brown). Until the latter part

21
of 2006, the oldest fossil bee known was Cretotrigona pricsa Michener and
Grimaldi, from the New Jersey amber (Fig. 1; Pl. 2 (Michener and Grimaldi,
1988; Engel, 2000; Michener, 2000)).
However, latter in the year 2006, a new oldest fossilized bee
[Melittosphex burmensis Poinar and Danforth, Melittosphecidae] was
described from the early Cretaceous Burmese amber (over 100 million years
ago (Fig. 2 & Pl. 3)). M. burmensis from the Burmese amber is 35-45 million
years older than C. prisca. This new fossil provides insights into the
morphology of the earliest bees and provides a new minimum date for the
antiquity of bees and bee-mediated pollination Poinar and Danforth (2006).

22
Plate 2. Cretotrigona pricsa Michener and Grimaldi from the New Jersey
amber (after Engel, 2000)
Plate 3. Melittosphex burmensis Poinar and Danforth from the Burmese amber
(after Poinar and Danforth, 2006)
A
A

23
Figure 1. Cretotrigona pricsa (Illustration after Michener and Grimaldi 1988)
Figure 2. Melittosphex burmensis (Illustration after Poinar and Danforth,
2006).
Fig 2
Fig 2
Fig 2
Fig 2
Fig 2
Fig 2
Fig 2
Fig 2

24
Danforth (2004) in his review on bee phylogeny stated that
“Additional insights into the timing of bee pollination can be gained from
mapping extant pollinator relationships onto the phylogeny of the
angiosperms (Soltis, et al., 1999, Qiu, et al., 1999; the Missouri Botanical
Garden’s Angiosperm Phylogeny web site for the most up-to-date information
on this subject). Alone, this approach is fraught with problems. First,
pollination syndromes are highly variable within higher categories and it is
difficult to assign a state unambiguously to angiosperm families or orders.
Second, there is no guarantee that plant/pollinator relationships have remained
the same from the origins of the angiosperms to the present. Nevertheless,
specialized relationships between bees and angiosperms are not likely to have
existed prior to the common ancestor of the eudicots because extant
magnoliids (monocots, Winterales, Laurales, Magnoliales, Chloranthales,
Piperales, etc.) are, for the most part, not bee pollinated (Thien, et al., 2000).
The eudicots have recently been estimated to be between 147 and 131 million
years old based on combined fossil and DNA evidence (Wikstrцm, et al.,
2001). Finally, molecular estimates of angiosperm origins push the date of the
angiosperm/gymnosperm split back to approximately the Triassic/Jurassic
boundary (210 Ma; Wolfe, et al., 1989, Laroche, et al., 1995, Goremykin, et
al., 1996, and Sanderson, 1997). Needless to say, this is considerably earlier
than the oldest fossil angiosperms (early Cretaceous, 145 Ma) and casts even
more doubt on the belief that bees necessarily arose in the mid- to late
Cretaceous”.

25
The few anthidine fossil bees known are Anthidium exhumatum
Cockerell, Anthidium scudderi Cockerell and “Dianthidium” terttarium
(Cockerell). These fossil bees are believed to be of late Eocene- early
Oligocene period (Engel and Perkovsky, 2006)
Overview of Phylogenetic Studies in Apoidea
Roig-Alsina and Michener (1993) conducted the first phylogenetic
analysis on all the known long-tongued bee families. Two years later, the
phylogenetic analysis on short-tongued bees was conducted by Alexander and
Michener (1995). Various authors have worked on some of the bee tribes and
families. Some of these works can be found in Rozen (1970) and Engel (2002)
on the systematic and phylogenetic relationships of the Fideline bees of the
family Megachilidae. It is also known that, Brooks (1988), Alexander (1990;
1994), Roig-Alsina (1991) and Rozen (1996) have worked on the
phylogenetic studies of the Anthophorine bees whereas Rozen and McGinley
(1974) conducted phylogenetic studies on the bee family Melittidae. The
cladistic analysis on the subfamily Meliponinae was studied by Camargo and
Menezes-Pedro (1992). More research works on the phylogenetic studies of
bees can be found in Pignata and Diniz-Filho (1995) who conducted studies
on the phylogenetic autocorrelation and evolutionary constraints in worker
body size of some neotropical stingless bees. The phylogeny of the

26
Ammobatini was studied by Eardley and Brothers (1997) whiles the molecular
phylogeny and historical biogeography of the large carpenter bees, genus
Xylocopa (Hymenoptera: Apidae) have been done by Leys, et al., (2002).
Phylogenetic Position of the Anthidiini
The phylogenetic position of the Anthidiini within the family
Megachilidae (Subfamily Megachilinae) was studied by Roig-Alsina and
Michener (1993) in their cladistic analysis of long-tongued bees based on
adult and larval characters. The tribe Lithurgini emerged as the sister group of
tribes Anthidiini, Megachilini and Osmiini. The topology was supported by
six non-homoplasious synapomorphies, including the Labrum being longer
than broad, the thickening of secondary tentorial bridge merging with the
eutentorial arm before reaching the head wall, a reduced fan-shaped posterior
sheets of tentorium and the gonostylus more or less indistinguishably fused to
the gonocoxite. Furthermore, the volsella of the male is absent or fused to the
gonocoxite. Presently the tribe Dioxyini has been included within the
subfamily Megachilinae (Michener and Griswold, 1994). The monophyly of
the Anthidiini was supported by five non-homoplasious synapomorphies. The
five synapomorphies includes the anterior surface of the labrum of female
without basal polished area, the lateral expansion of internal thickening above
the epistomal ridge reduced and less than half width of socket diameter, a
lateral carina separating exposed part of the propleuron from part hidden by
pronotum is absent and the trunk of the anterior tibial spur with low expansion
at right angles to the velum, which curves apically into the spine of omaulus.

27
In addition, the stigma of the forewing is small, about as long as broad to
nearly being absent. This analysis was based on character from two anthidine
genera Roig-Alsina and Michener (1993).
In this study, a phylogenetic concept including a hypothetical
evolutionary scenario is presented for the Anthidiini of the world based on
cladistic analysis of adult morphology.

28
CHAPTER THREE
MATERIALS AND METHODS
Sources of the Bees Examined
A total of 990 bee specimens belonging to 35 genera were obtained
from various sources from August, 2005 to January, 2008. The genera and the
total number of specimens examined (in parenthesis) are as follows: Lithurgus
(2), Megachile (2), Acedanthidium (2), Afranthidium (111), Afrostelis (29),
Anthidiellum (162), Anthidioma (2), Anthidium (63), Anthidioctes (2),
Apianthidium (1), Aspidosmia (2), Aztecanthidium (2), Bathanthidium (1),
Benanthis (1), Cyphanthidium (3), Dianthidium (16), Duckeanthidium (2),
Eoanthidium (37), Epanthidium (4), Euaspis (40), Gnathanthidium (3),
Hoplostelis (2), Hypanthidiodes (2), Hypanthidium (6), Icteranthidium (2),
Indanthidium (2), Neanthidium (2), Notanthidium (2), Pachyanthidium (49),
Paranthidium (2), Plesianthidium (245), Pseudoanthidium (140),
Rhodanthidium (7), Serapista (32), Trachusa (13). The detailed biogeographic
information, collector’s name and location of depository of each specimen are
provided under the material examined (Appendix 2). The material studied
were loaned specimens obtained from the following individuals, their
institution, cities and acronyms are given in parenthesis. These include
Barbara Dombarosky of the Transvaal Museum of South Africa (TMSA),
Pretoria, South Africa, Christina Lebean, John Ascher and Jerry Rozen of the
American Museum of Natural History (AMNH), New York, USA, Eliane

29
DeConick of the Musee Royal de l'Afrique Centrale (MRAC) Tervuren,
Belgium, Frank Kock Museum Naturkunde für Humboldt (ZMHB)
Universität, Berlin, Germany); George Else formerly of the Natural History
Museum (NHM), London, United Kingdom, Jennifer Thomas of the Snow
Entomological Museum Collection (SEMC), University of Kansas, United
States of America and Margie Cockrane of the South African Museum
(SAMC), Cape Town, South Africa. Other depositories included Mary
Gikungu of the Kenya National Museum (KNM), Nairobi, Kenya, Peter
Kwapong University of the Cape Coast Entomology Museum (UCCEM),
Cape Coast, Ghana and Ros Urban of the South African National Collection
of Insects (SANC), Pretoria, South Africa.
Preparation of Specimens
Immediately the loaned specimens were received from the various
museums, they were quarantined and curated. Quarantine procedures involved
storage of specimens in freezers at temperatures of -4ºC to -15 ºC for a
minimum period of two weeks to eliminated possible museum pests such as
mites. In certain instances, direct hand picking method was used to remove
these pests. Curating of specimens was carried out to ensure that the detailed
information on each specimen conformed to what was provided by the
literature, as well as ensuring that each specimen was accurately placed into
the appropriate taxon.

30
Examination of Specimens
The morphological examination of the specimens in this present study
was on visual observation of external morphological features. Specimens were
examined, and illustrations and measurements made using an Olympus
dissection microscope fitted with a drawing tube and a graticule. Photographic
images of the specimens including certain features that were used for their
diagnoses were captured using camera lucida. The Combined ZM computer
software was used to edit captured images. Male genitalia and hidden sterna
of some relaxed specimens were removed using a pair of forceps and
macerated in 10% KOH for 12–24 hours at room temperature, depending on
the thickness of the cuticle of each structure. After clearing the genitalia and
sterna were stored and examined in 75% glycerine.
Morphological terminologies, Abbreviations and Measurements
The terminology used in this study follows that of Michener (1944,
2000). Different morphological terms are explained in the text. Vestiture
generally refers to the relatively fine hairs and where setae are specifically
mentioned, these are thick hairs. Pubescent hair refers to branched hairs, while
setose hair refers to unbranched hairs. Small punctures, as referred to in the
descriptions, measure 0.8 mm or less in diameter, whereas large punctures are
1.0 or more mm in diameter. Terminology such as antennal scape groove was
used to describe the large fovea or depression above the antennal sockets of
the female of genus Notanthidium (Fig. 3b).

31
In the revision of the genus Anthidiellum, the terms outer and inner
carinae were used to describe one or two ridge (s) on the mandible of the
female, the outer carina originates from the mesal region of the mandible and
terminates on the outer tooth, the inner carina originates from the basal region
of the mandible and terminates in the mesal region. Illustrations of the legs
exclude the coxa and the anterior region of trochanter. The abbreviations T
and S are used for the metasomal terga and sterna, respectively (e.g. T1 and
S1 refers to the first metasomal tergum and sternum, respectively). In the
illustrations of the male genitalia, the posterior end is directed towards the top
of the page. Furthermore, all labels on both photographs and illustrations
follow that of figure 3 and 4.
Phylogenetic Studies of the tribe Anthidiini
Selection of Taxa within the Tribe
During the cladistic studies of the anthidine genera, the type species of
the genus and the type species of the nominative subgenus (for large genera)
were selected and examined. However, for some genera the type species were
not available, hence other species belonging to those genera were selected for
the analysis.Two separate cladistic studies were undertaken. The first study
encorporated 35 species representing 33 anthidine genera (ingroup, listed in
the matrix) and two outgroups namely Lithurgus pullatus (Vachal) and
Megachile semiflava (Cockerell).
In the second analysis, 17 species of the genus Anthidiellum and two
outgroup taxa were studied. These represent the Afrotropical species of the

32
genus (listed in the cladogram) and the genera chosen as outgroups were
(Trachusa aquilifilum (Strand) and Eoanthidium turnericum
(Mavromoustakis)).
Roig-Alsina and Michener (1993) postulated that these outgroups used
for the phylogenetic studies, represent the recent common ancestors and the
most recent common ancestors of the tribe Anthidiini and genus Anthidiellum.
Hence, these two outgroups revealed the probable transformation series of the
character states within the subfamily Megachilinae.
Selection and Coding of Characters and Character States
Adults of each species were examined based on 51 external characters,
and each character for which distinct states occurred in different species was
included in the matrices. A number of the characters used in the present
phylogenetic analysis of the tribe were taken or modified from Roig-Alsina
and Michener (1993). Annotated lists of the characters used in the
phylogenetic reconstructions can be found in page 31. The states of each
character within the ingroup were coded as 0 representing primitive; 1, 2 and
3 referring to successive derived states. All character states in the outgroup
were considered to be pleisomorphic. Additive binary coding was employed.
Unknown character states were coded with "?" whereas inapplicable
characters were coded with "-". All characters were weighted equally. Four
characters of the male genitalia were deactived during the analysis.

33
Characters used for the Cladistic Analyses of the Anthidiini
The following characters and character states were used in the cladistic
analysis of Anthidiini.
Head
1. Paraocular carina. (0) Present. (1) Absent.
2. Dorsal region of preoccipital area, in female. (0) Gently rounded or
angulate. (1) Carinate.
3. Lateral region of preoccipital area, in female. (0) Gently rounded. (1)
Carinate.
4. Preoccipital area. (0) Without lamella. (1) Lamellate.
5. Juxtantennal carina. (0) Absent. (1) Present.
6. Subantennal suture. (0) More or less straight. (1) Curved.
7. Distance between antennal sockets. (0) More than two antennal socket
diameters. (1) Subequal to or less than two antennal socket diameters.
8. Antennal scape, posterior surface. (0) Densely pubescent. (1) Sparsely
pubescent to impubescent
9. Antennal scape groove of female. (0) Absent. (1) Present.
10. Mediolongitudinal region of face. (0) Slightly raised. (1) Strongly
raised.
11. Ventral margin of clypeus. (0) Without projections, smooth. (1)
Unevenly margined, with projections.
12. Ventro-lateral region of female clypeus. (0) Flat. (1) Tuberculate.
13. Dorsal region of clypeus of female. (0) Flat to gently convex. (1)

34
Highly elevated.
14. Mandibular teeth of female. (0) Three teeth. (1) Four or more teeth.
15. Malar area. (0) With one lower tubercle. (1) Possess several tubercles.
16. Facial vestiture. (0) Densely pubescence. (1) Sparse to impubescence.
Mesosoma
17. Dorso-lateral angle of pronotum. (0) Well developed. (1) Absent or
weakly developed.
18. Distal region of pronotal lobe. (0) Round or gently curved. (1) Sharply
pointed.
19. Omaulus. (0) Rounded. (1) Carinate.
20. Scutum length. (0) As long as broad. (1) Elongated, twice as long as
broad.
21. Scuto-scutellar suture. (0) Similar to scuto-axilla suture, closed. (1)
More open than scuto-axilla suture, foveate.
22. Posterior margin of axilla, dorsal view. (0) Does not extend laterally
beyond lateral margin of scutum. (1) Extend beyond lateral margin of
scutum.
23. Posterior region of axilla. (0) Without spine. (1) Possess spine.
24. Scutellum, dorsal view. (0) Not overhanging and concealing
propodeum. (1) Obscures propodeum, at least medially.
25. Scutellum , lateral view. (0). Rounded posteriorly, without lamella. (1)
Lamellate posteriorly.
26. Scutellum, posterior margin, dorsal view: (0) Entire. (1) Slightly

35
notched postero-medially. (2) Deeply notched postero-medially.
27. Apex of outer hind tibia spur. (0) More or less straight. (1) Strongly
curved.
28. Shape of female hind basitarsus. (0) Slender, subequal to or slightly
more than half width of hind tibia. (1) Broader than hind tibia.
29. Length of hind basitarsus. (0) Short, subequal to or less than half
length of hind tibia. (1) Long, subequal length of hind tibia.
30. Vestiture on dorsal surface of hind basitarsus. (0) Sparse to
impubescent. (1) Dense.
31. Basitibial plate in male. (0) well defined. (1) Very small or absent.
32. Tarsal claws of female. (0) Simple. (1) Cleft, inner ramus sometimes a
tooth.
33. Apex of marginal cell position. (0) Seperated from wing margin. (1)
On wing magin.
34. Apex of marginal cell, shape. (0) Rounded. (1) Pointed.
35. Stigma. (0) Broad, twice as wide as prestigma. (1) Slender, as wide as
prestigma or absent.
36. First recurrent vein of fore wing. (0) Extends distally behind first
transverse cubital vein. (1) Meets first transverse cubital vein distally
or enters first submarginal cell.
37. Second recurrent vein of fore wing. (0) Meets second transverse
cubital vein distally or extend distally behind second transverse cubital
vein. (1) Enters second submarginal cell midway.

36
38. Second recurrent vein, in profile. (0) Enters second submarginal cell or
meet second transverse cubital vein distally. (1) Extend distally behind
second transverse cubital vein.
39. Second cubital cell of fore wing. (0) Subequal half length of first
cubital cell or less. (1) More than half or subequal length of first
cubital cell.
40. Jugal lobe. (0) Long, more than half length of vannal lobe. (1) Short,
half as long as or less than half length of vannal lobe.
41. Apex of jugal lobe: (0) Broadly rounded; (1) Narrowly rounded to
slightly pointed.
42. Vein Cu-v of hind wing. (0) Straight. (1) Slants.
43. Vein Cu-v of hind wing (length). (0) Short, much less than half as long
as second absissa of vein M+Cu. (1) Long, subequal to half length of
second absissa of vein M+Cu. (2) Longer than second absissa of
M+Cu.
Metasoma
44. Scopa. (0) Present. (1) Absent.
45. Fovea on anterior region of T1. (0) Present. (1) Absent.
46. Hairy subapical bands on postgraduli on T1-T5. (0) Present. (1)
Absent.
47. Pygidial plate on female T6. (0) Present. (1) Absent.

37
48. Sternal setae of male. (0) Absent. (1) Present.
49. Gonostylus. (0) Narrow less than half width of gonocoxite. (1)
Expanded more than half width of gonocoxite.
50. Penis valves. (0) Narrowly seperated. (1) Widely seperated.
51. Apical region of penis valves. (0) Without notch on margin. (1)
Notched
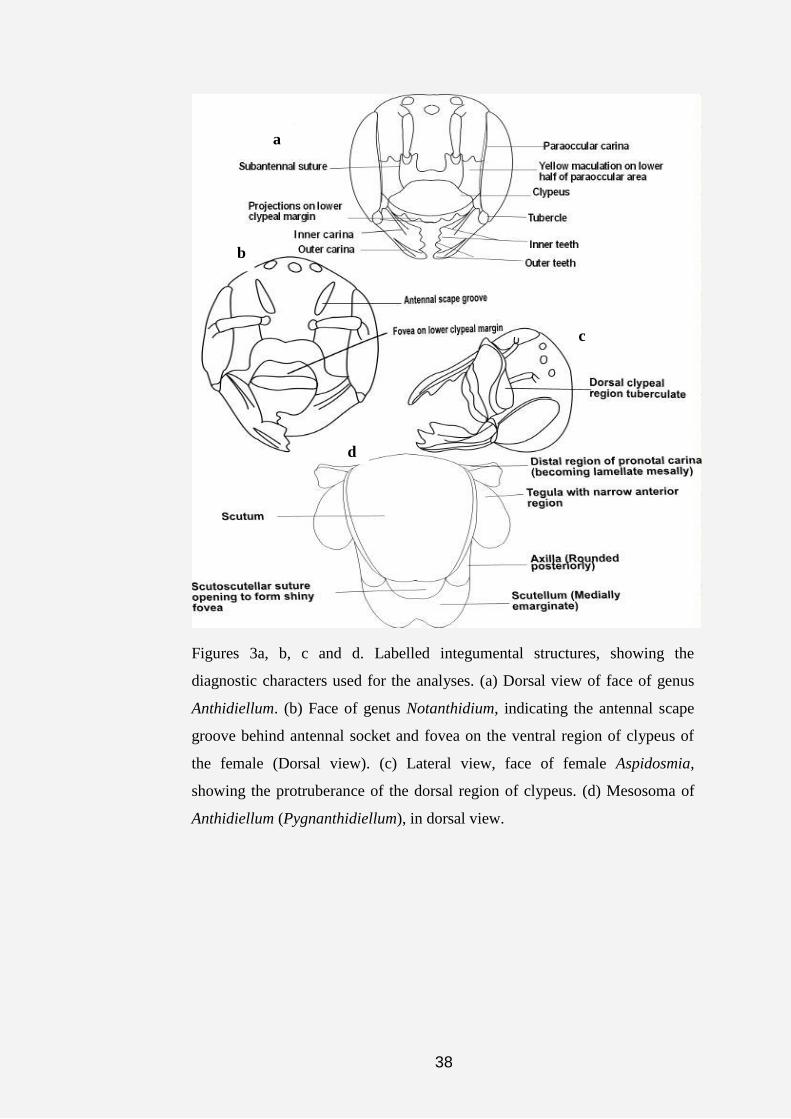
38
Figures 3a, b, c and d. Labelled integumental structures, showing the
diagnostic characters used for the analyses. (a) Dorsal view of face of genus
Anthidiellum. (b) Face of genus Notanthidium, indicating the antennal scape
groove behind antennal socket and fovea on the ventral region of clypeus of
the female (Dorsal view). (c) Lateral view, face of female Aspidosmia,
showing the protruberance of the dorsal region of clypeus. (d) Mesosoma of
Anthidiellum (Pygnanthidiellum), in dorsal view.
b
a
c
d
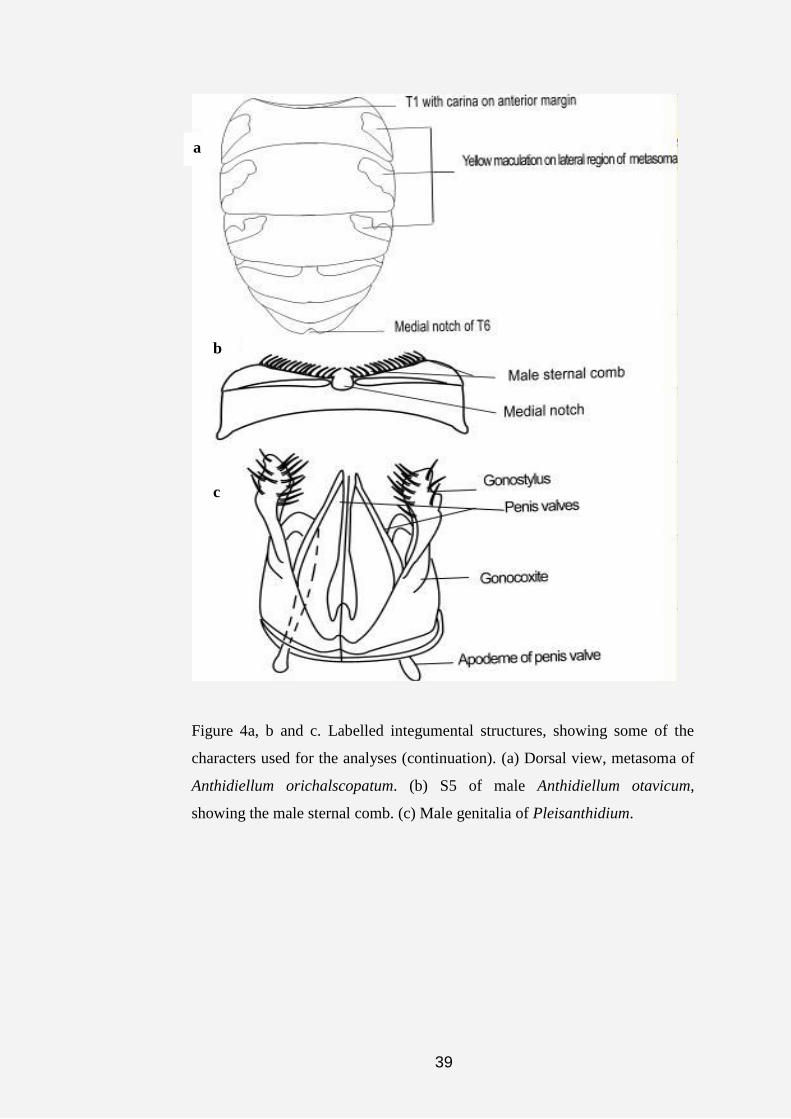
39
Figure 4a, b and c. Labelled integumental structures, showing some of the
characters used for the analyses (continuation). (a) Dorsal view, metasoma of
Anthidiellum orichalscopatum. (b) S5 of male Anthidiellum otavicum,
showing the male sternal comb. (c) Male genitalia of Pleisanthidium.
a
c
b

40
Characters used for the Cladistic Analyses of the Genus Anthidiellum
During the cladistic studies on the genus Anthidiellum, the following
characters and character states were used:
Head
1. Distance between antennal sockets: (0) More than two antennal socket
diameters. (1) Subequal or less than two antennal socket diameters.
2. Length of subantennal suture: (0) Subequal or less than half clypeus
length. (1) More than half clypeus length.
3. Profile of subantennal suture: (0) More or less straight. (1) Strongly
arcuate.
4. Juxtantennal carina. (0) Absent. (1) Present.
5. Projections on female clypeal margin (Number): (0).Six or more. (1)
Four or less.
6. Clypeus mid-region profile: (0) Greatly raised. (1) Slightly raised.
7. Paraoccular carina: (0) Strongly developed. (1) Weakly developed.
8. Preoccipital carina (orientation): (0) Present dorsally, or without
carina. (1) Dorsolaterally present. (2) Present laterally.
9. Mandible punctation (size). (0) Small puntures on entire dorsal
surface. (1) Small at apex, large towards base.
10. Mandibular punctation (density): (0) Sparsely dense. (1) Moderately
dense.
11. Malar area of female. (0) Two tubercles. (1) Only one is present.

41
12. Mandibular carina of female (Size). (0) Large. (1) Reduced.
13. Mandibular carina of female . (0) Present with two carinae. (1) Present
with one carina.
Mesosoma
14. Pronotum with dorsal, anterior margin: (0) Strongly convex. (1)
Gently convex.
15. Profile of pronotal lobe, with respect to distal region: (0) Pronotal
carina small mesally, becoming progressively lamellate distally. (1)
Lamellate.
16. Pronotal lamella or carina: (0) Extend little mesad from lateral margin
of scutum as laterad (1) Extend nearly as far mesad from lateral
margin of scutum as laterad.
17. Tegula (structure): (0) Narrow anteriorly, lateral margin slightly
concave. (1) Not narrowed anteriorly, with lateral margin convex.
18. Axilla (posterior margin): (0) Extend laterally beyond lateral margin of
scutum. (1) Does not extend laterally beyond lateral margin of scutum.
19. Scuto-axilla suture: (0) Similar to scuto-axilla suture, closed. (1)
Dissimilar to scuto-axilla suture, open to form shiny fovea.
20. Lateral margin of scutum (in dorsal view). (0) Does not extend
longitudinally concave. (1) Extend longitudinally concave, groove
separate extended surface with carina next to tegula.
21. Scutellum in dorsal view, with respect to shape. (0) Medially

42
emarginate. (1) Truncate.
22. Hamuli of hind wing (Number). (0) Eighteen or more. (1) Eleven and
less than eighteen. (2) Ten or less.
23. Vein cu-v of hind wing. (0) Slants. (1) Straight.
24. Second recurrent vein of forewing. (0) Meet or nearly meets second
transverse cubital vein. (1) Does not meet second transverse cubital
vein (extend 0.5mm distally).
25. Vestiture on hind basitarsus. (0) Very dense. (1) Moderate to sparsely
dense.
26. Hind basitarsus (size): (0) Broad, more than half as broad as hind tibia,
or as broad as hind tibia. (1) Slender, subequal or half as broad as hind
tibia.
Metasoma
27. Metasoma punctation (Density): (0) Sparsely dense. (1). Moderately
dense.
28. Vestiture on T1-T6 of female. (0) At least moderately dense on T6. (1)
Sparsely dense.
29. S5 of males with comb. (0) Present (1) Absent
30. S5 of males with comb or setae. (0) Medial. (1) Lateral.
31. S5 of males with lateral comb (number of setae). (0) Very long, more
than twenty setae long. (1) Short, less than twenty and more than ten
setae long. (2) Very short, less than ten setae long.
32. Male gonostyli (structure). (0) Narrowed apically. (1) Expanded

43
apically.
33. Gonostylus (Profile of margin). (0) Notched inside, or without
marginal notch. (1). Notched outside.
34. Penis valves. (0) Narrowly seperated. (1) Widely seperated.
35. Apical region of penis valves. (0) Straight. (1) Curved.
Phylogenetic Procedures
Character matrices for cladistic analyses were constructed using the
WinClada computer software, version 1.00.08 Nixon, 1999-2002). The
phylogeny was created in NONA computer software (Goloboff, 1993) using
an unconstrained heuristic search strategy having Multiple TBR+TBR (mult*
max*). The search parameters were 100,000 maximum trees to keep
(Default=100), 500 replication (Default=1), 500 starting tree per replication
(Default=1) and zero (0) random time. Common cladogram measures such as
the consistency index (CI) and the retention index (RI) were used to evaluate
the fitness of the data to the cladogram. Analysis of characters as well as
character optimization using unambiguous optimization modus was performed
with the WinClada software.
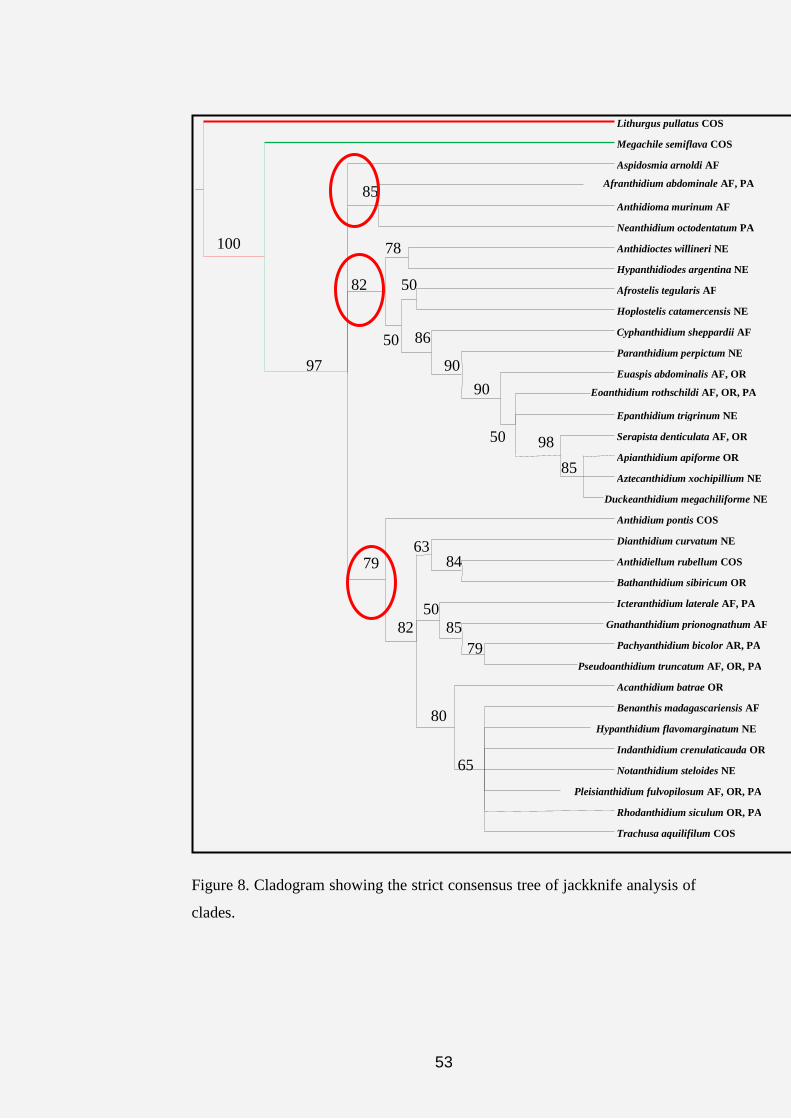
Bootstrap analysis (Felsenstein, 1985) and jackknife sampling
(Lanyon, 1985; Siddall, 1996) were used to assess evidential support for
clades of cladograms. Values were calculated based on 100 replicates with 50
random sequence additions per replicate. Presentation of trees as well as
character mapping was carried out in the WinClada.

44
The search parameters stated were derived from series of preliminary
analyses by NONA on various anthidine genera based on that were several
characters selected and coded. Few characters that were observed to have
lower constituency index and retention index were replaced with new ones the
provided high CI and RI, as well as resolved collapsed nodes on the
cladogram.
Procedure for Studies on Systematic of the Taxa
During the systematic studies of the tribe Anthidiini and the genus
Anthidiellum, lengths were measured to confirm the size variations of these
bee species as cited in literature. These were the head lengths which were
measured from dorsal region of the preoccipital area to the ventral region of
the clypeus, the scutum and scutellum lengths were measured from the antero-
mesal region of the scutum to the postero-mesal region of the scutellum. The
forewing lengths were measurements made from the basal region to the tip of
the apical region of the forewing while, body lengths were measured from the
ventral region of the clypeus to the postero-mesal region of tergites six (in
females) and seven (in males).
In addition, various attributes of vestiture on the integument were
described. These attributes included variations in vestiture colour, density, and
distribution, the position of female scopa on sternite and the presence or
absence of the male sternal comb. Furthermore, descriptive and diagnostic
characters that could be used in the identification of the genera were recorded
based on the following features. These included colour distribution on the

45
integument, number of mandibular dentition in the female, number of
projections on lower clypeal margin of the female, the presence or absence of
carinae on face, curvature of the subantennal suture and hind tibial spur as
well as aperture of scuto-scutellar suture. Other features included the
orientation and shape of the posterior region of the scutellum, the presence or
absence of arolia, the number of lateral spines on metasoma, the shape of the
apical region of the male gonostylus and width of seperation between penis
valves of the male genitalia.
In addition, under each genus, detailed information on previous works
stated in zoological records and comment have been made on the general
shape of taxa, foraging status (parasitic/non-parasitic), geographic
distribution, as well as generic status of each of the thirty three genera
examined.

46
CHAPTER FOUR
RESULTS
Cladograms and Tree topologies of the Anthidiini
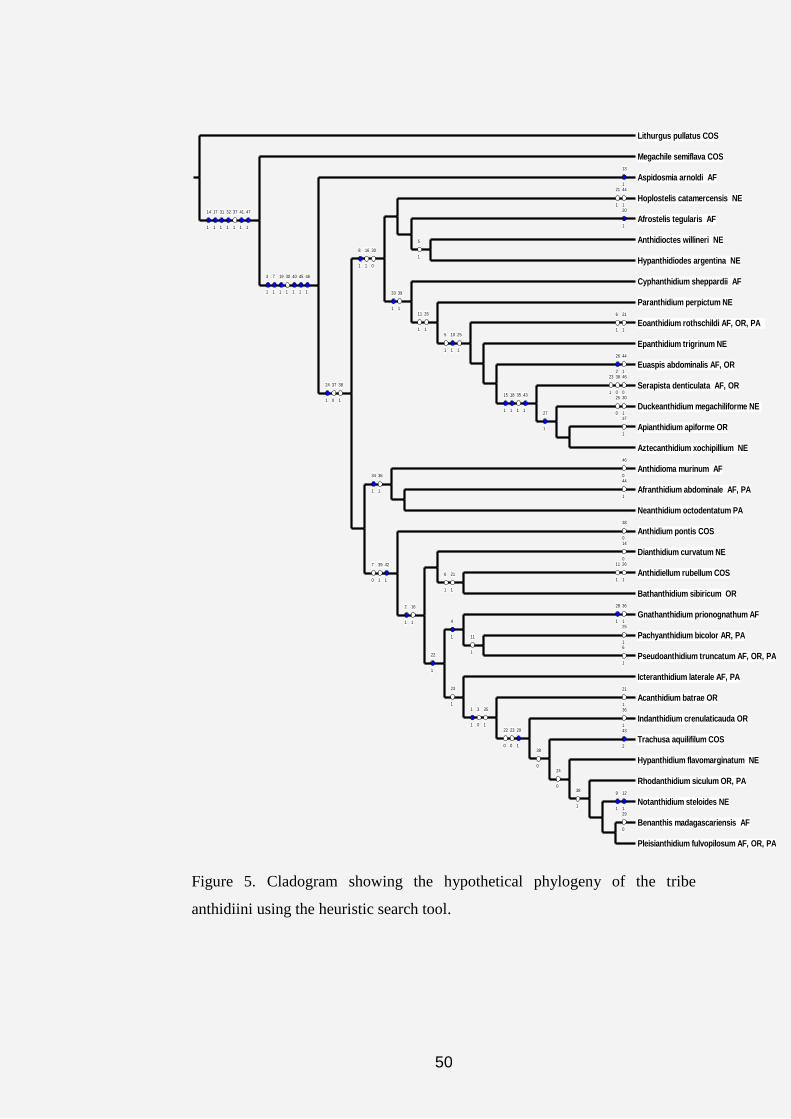
The results obtained from the cladistic analysis of the data matrix
(Table 1) using the heuristic search tool yielded 191 equally most
parsimonious trees with a length (L) of 86 steps, the consistency index (CI) of
56 and the retention index (RI) of 83. Figure 5 shows a typical tree topology of
most of the 191 most parsimonious trees obtained during analysis of
characters and character states by NONA computer software. However, the
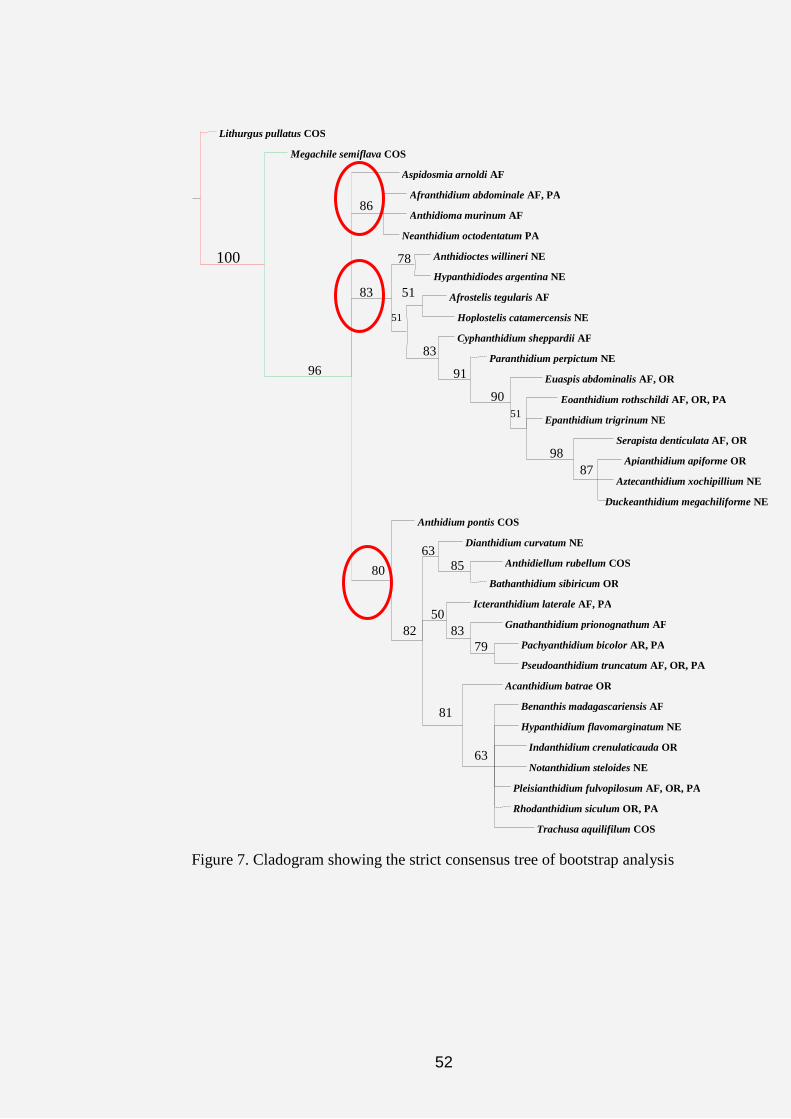
strict consensus tree in figure 11 yielded L: 93 steps, CI: 52% and RI: 80%.
On the strict consensus tree 17 unsupported nodes were collapsed into several
polytomies. In addition, bootstrap and jackknife search strategies resulted in
maximum branch length, consistency and retension indices of 90/ 89 steps, 54/
55 % and 81/ 81 %, respectively (Figures 7 and 8).
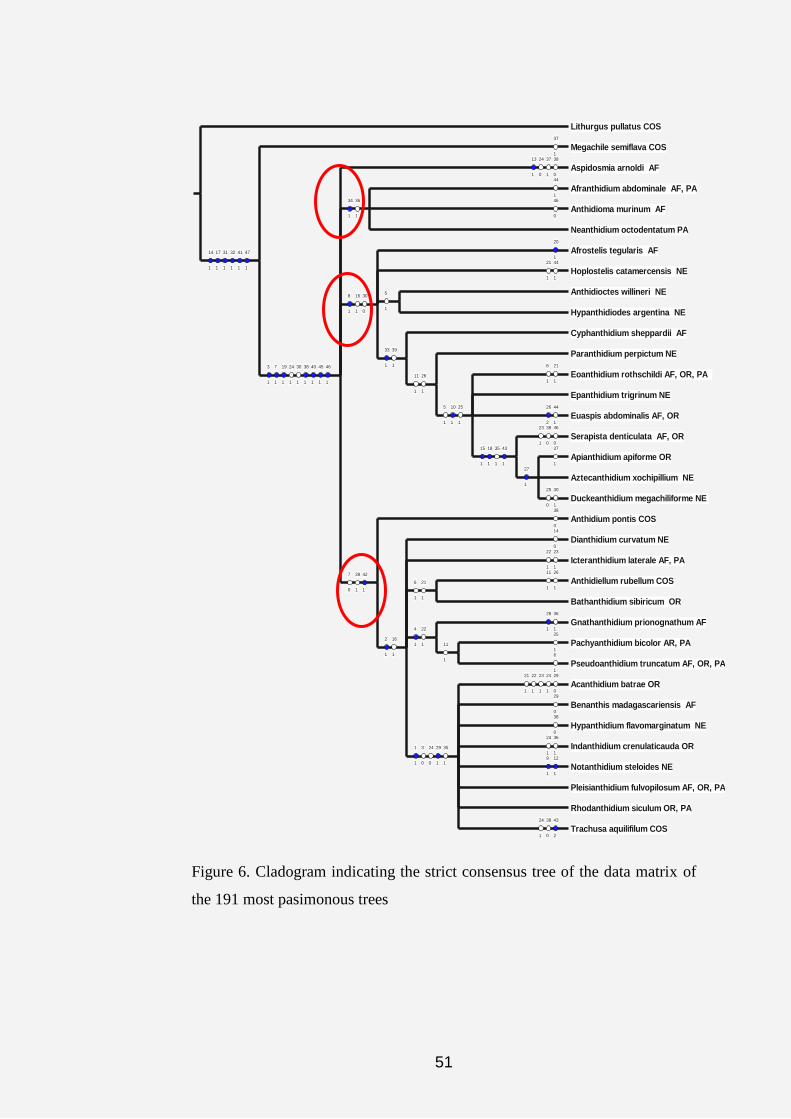
Three major clades parallel to the outgroups (Lithurgus and
Megachile) emerged from the cladograms (Figs. 6-8) as follows:
(1) Aspidosmia, Afranthidium, Anthidioma and Neanthidium.
(2) Afrostelis, Hoplostelis, Anthidioctes, Hypanthidiodes,
Cyphanthidium, Paranthidium, Eoanthidium, Epanthidium, Euaspis,
Serapista, Apianthidium, Aztecanthidium and Duckeanthidium.

47
(3) Anthidium, Dianthidium, Icteranthidium, Anthidiellum,
Bathanthidium, Gnathanthidium, Pachyanthidium, Pseudoanthidium,
Acedanthidium, Benanthis, Hypanthidium, Indanthidium, Notanthidium,
Pleisanthidium, Rhodanthidium and Trachusa.
On the cladograms in figures 5 and 6, the symbols“○”and “●”
represent the homoplasious and non-homoplasious states in which the
characters occurred after the analysis. The tree yielded 27 of the non-
homoplasious characters. The three red circles on figures 6, 7 and 8 are
representing the three major islands revealed by the cladograms.
The acronyms attached to each taxa in all cladograms give an
indication of the biogeographic data of that genus. Thus, AF represents the
Afrotropical Region (Sub-Saharan, East and West Africa and Madagascar);
PA refers to the Palaearctic Region (North Africa and Europe); OR represents
the Oriental Region (Asian countries); NE represents the Neotropical Regions
(South America) and COS refers to the Cosmopolitan regions (refers to those
genera that occur in all the geographical areas).
During the statistical analysis of the data matrix by NONA (computer
software), all the 51 characters possessing equal weights and were of additive
coding, with 308 maximum steps and 53 minimum steps of character change.
However, 47 out of the 51 external morphological characters were actually
employed (activated), while the remaining four characters on male genitalia
were deactivated. In addition, 4 characters of male genitalia of species
Anthidioma murinum resulted as inapplicable characters "-" due to the fact
that, this genus is known by only female species. 18 characters states on males

48
from five genera were revealed as missing characters. The five genera
included Icteranthidium, Bathanthidium, Gnathanthidium, Aztecanthidium and
Duckeanthidium. The 18 characters states were examined on the genitalia of
the male specimens of these genera. However, only type specimens of those
genera were available for examination hence, examinations were not
conducted on hidden structures such as the male genitalia that require
dissection procedures. In genera Bathanthidium and Gnathanthidium only
female specimens were available. During the preliminary analysis of the tribe
by NONA, the absence of these characters was observed to have caused the
collapse of several nodes into polytomies. Hence, all those characters were
deactivated during the analysis of the tribe in the course of the main research
analysis by NONA.
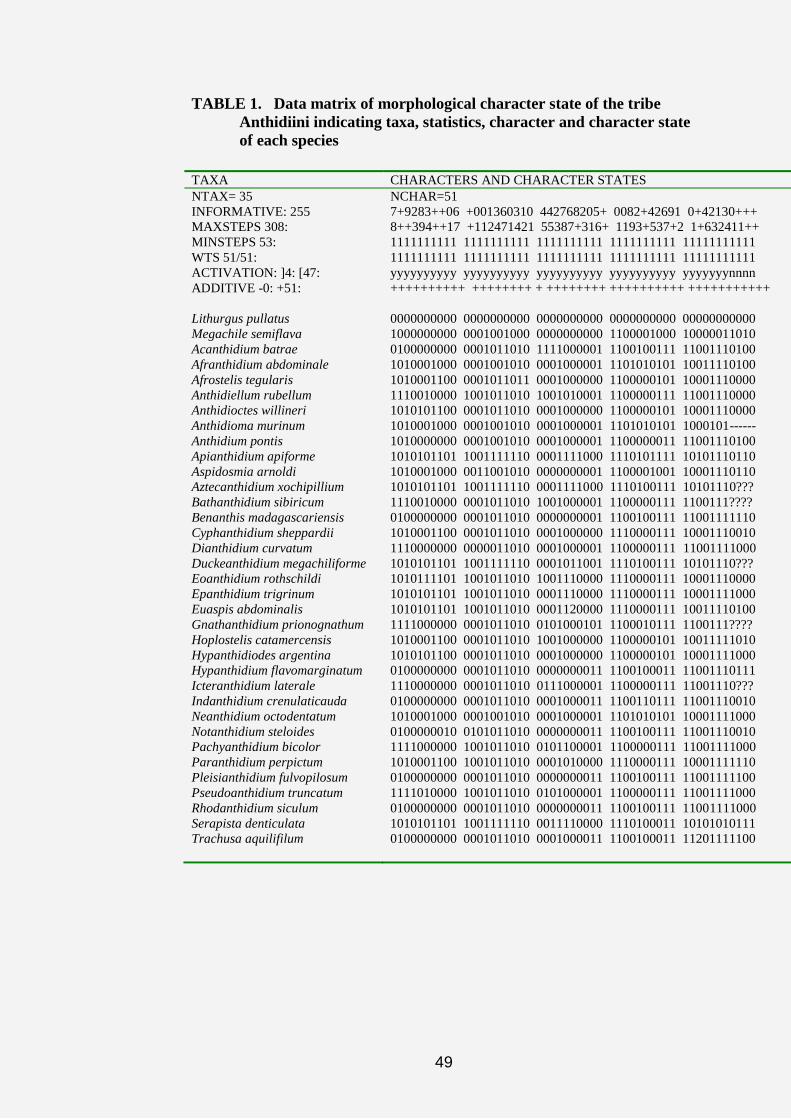
49
TABLE 1. Data matrix of morphological character state of the tribe
Anthidiini indicating taxa, statistics, character and character state
of each species
TAXA CHARACTERS AND CHARACTER STATES
NTAX= 35
INFORMATIVE: 255
MAXSTEPS 308:
MINSTEPS 53:
WTS 51/51:
ACTIVATION: ]4: [47:
ADDITIVE -0: +51:
NCHAR=51
7+9283++06 +001360310 442768205+ 0082+42691 0+42130+++
8++394++17 +112471421 55387+316+ 1193+537+2 1+632411++
1111111111 1111111111 1111111111 1111111111 11111111111
1111111111 1111111111 1111111111 1111111111 11111111111
yyyyyyyyyy yyyyyyyyyy yyyyyyyyyy yyyyyyyyyy yyyyyyynnnn
++++++++++ ++++++++ + ++++++++ ++++++++++ +++++++++++
Lithurgus pullatus
Megachile semiflava
Acanthidium batrae
Afranthidium abdominale
Afrostelis tegularis
Anthidiellum rubellum
Anthidioctes willineri
Anthidioma murinum
Anthidium pontis
Apianthidium apiforme
Aspidosmia arnoldi
Aztecanthidium xochipillium
Bathanthidium sibiricum
Benanthis madagascariensis
Cyphanthidium sheppardii
Dianthidium curvatum
Duckeanthidium megachiliforme
Eoanthidium rothschildi
Epanthidium trigrinum
Euaspis abdominalis
Gnathanthidium prionognathum
Hoplostelis catamercensis
Hypanthidiodes argentina
Hypanthidium flavomarginatum
Icteranthidium laterale
Indanthidium crenulaticauda
Neanthidium octodentatum
Notanthidium steloides
Pachyanthidium bicolor
Paranthidium perpictum
Pleisianthidium fulvopilosum
Pseudoanthidium truncatum
Rhodanthidium siculum
Serapista denticulata
Trachusa aquilifilum
0000000000 0000000000 0000000000 0000000000 00000000000
1000000000 0001001000 0000000000 1100001000 10000011010
0100000000 0001011010 1111000001 1100100111 11001110100
1010001000 0001001010 0001000001 1101010101 10011110100
1010001100 0001011011 0001000000 1100000101 10001110000
1110010000 1001011010 1001010001 1100000111 11001110000
1010101100 0001011010 0001000000 1100000101 10001110000
1010001000 0001001010 0001000001 1101010101 1000101------
1010000000 0001001010 0001000001 1100000011 11001110100
1010101101 1001111110 0001111000 1110101111 10101110110
1010001000 0011001010 0000000001 1100001001 10001110110
1010101101 1001111110 0001111000 1110100111 10101110???
1110010000 0001011010 1001000001 1100000111 1100111????
0100000000 0001011010 0000000001 1100100111 11001111110
1010001100 0001011010 0001000000 1110000111 10001110010
1110000000 0000011010 0001000001 1100000111 11001111000
1010101101 1001111110 0001011001 1110100111 10101110???
1010111101 1001011010 1001110000 1110000111 10001110000
1010101101 1001011010 0001110000 1110000111 10001111000
1010101101 1001011010 0001120000 1110000111 10011110100
1111000000 0001011010 0101000101 1100010111 1100111????
1010001100 0001011010 1001000000 1100000101 10011111010
1010101100 0001011010 0001000000 1100000101 10001111000
0100000000 0001011010 0000000011 1100100011 11001110111
1110000000 0001011010 0111000001 1100000111 11001110???
0100000000 0001011010 0001000011 1100110111 11001110010
1010001000 0001001010 0001000001 1101010101 10001111000
0100000010 0101011010 0000000011 1100100111 11001110010
1111000000 1001011010 0101100001 1100000111 11001111000
1010001100 1001011010 0001010000 1110000111 10001111110
0100000000 0001011010 0000000011 1100100111 11001111100
1111010000 1001011010 0101000001 1100000111 11001111000
0100000000 0001011010 0000000011 1100100111 11001111000
1010101101 1001111110 0011110000 1110100011 10101010111
0100000000 0001011010 0001000011 1100100011 11201111100
50
Lithurgus pullatus COS
Megachile semiflava COS
Acanthidium batrae OR
Afranthidium abdominale AF, PA
Afrostelis tegularis AF
Anthidiellum rubellum COS
Anthidioctes willineri NE
Anthidioma murinum AF
Anthidium pontis COS
Apianthidium apiforme OR
Aspidosmia arnoldi AF
Aztecanthidium xochipillium NE
Bathanthidium sibiricum OR
Benanthis madagascariensis AF
Cyphanthidium sheppardii AF
Dianthidium curvatum NE
Duckeanthidium megachiliforme NE
Eoanthidium rothschildi AF, OR, PA
Epanthidium trigrinum NE
Euaspis abdominalis AF, OR
Gnathanthidium prionognathum AF
Hoplostelis catamercensis NE
Hypanthidiodes argentina NE
Hypanthidium flavomarginatum NE
Icteranthidium laterale AF, PA
Indanthidium crenulaticauda OR
Neanthidium octodentatum PA
Notanthidium steloides NE
Pachyanthidium bicolor AR, PA
Paranthidium perpictum NE
Pleisianthidium fulvopilosum AF, OR, PA
Pseudoanthidium truncatum AF, OR, PA
Rhodanthidium siculum OR, PA
Serapista denticulata AF, OR
Trachusa aquilifilum COS
21
1
44
1
20
1
26
1
11
1
46
0
38
0
37
1
13
1
29
0
14
0
30
1
25
0
21
1
6
1
44
1
26
2
36
1
28
1
44
1
21
1
36
1
12
1
9
1
25
16
1
46
0
38
0
23
1
43
2
38
1
24
0
38
0
29
1
23
0
22
0
35
1
3
0
1
1
23
1
11
1
4
1
22
1
21
1
6
1
16
1
2
1
42
1
39
1
7
0
36
1
34
1
5
1
27
1
43
1
35
1
18
1
15
1
25
1
10
1
5
1
26
1
11
1
39
1
33
1
30
0
16
1
8
1
38
1
37
0
24
1
46
1
45
1
40
1
30
1
19
1
7
1
3
1
47
1
41
1
37
1
32
1
31
1
17
1
14
1
Figure 5. Cladogram showing the hypothetical phylogeny of the tribe
anthidiini using the heuristic search tool.
51
Lithurgus pullatus COS
Megachile semiflava COS
Acanthidium batrae OR
Afranthidium abdominale AF, PA
Afrostelis tegularis AF
Anthidiellum rubellum COS
Anthidioctes willineri NE
Anthidioma murinum AF
Anthidium pontis COS
Apianthidium apiforme OR
Aspidosmia arnoldi AF
Aztecanthidium xochipillium NE
Bathanthidium sibiricum OR
Benanthis madagascariensis AF
Cyphanthidium sheppardii AF
Dianthidium curvatum NE
Duckeanthidium megachiliforme NE
Eoanthidium rothschildi AF, OR, PA
Epanthidium trigrinum NE
Euaspis abdominalis AF, OR
Gnathanthidium prionognathum AF
Hoplostelis catamercensis NE
Hypanthidiodes argentina NE
Hypanthidium flavomarginatum NE
Icteranthidium laterale AF, PA
Indanthidium crenulaticauda OR
Neanthidium octodentatum PA
Notanthidium steloides NE
Pachyanthidium bicolor AR, PA
Paranthidium perpictum NE
Pleisianthidium fulvopilosum AF, OR, PA
Pseudoanthidium truncatum AF, OR, PA
Rhodanthidium siculum OR, PA
Serapista denticulata AF, OR
Trachusa aquilifilum COS
37
1
29
0
24
1
23
1
22
1
21
1
44
1
20
1
26
1
11
1
46
0
38
0
37
1
38
0
37
1
24
0
13
1
29
0
14
0
30
1
25
0
21
1
6
1
44
1
26
2
36
1
28
1
44
1
21
1
38
0
23
1
22
1
36
1
24
112
1
9
1
25
16
1
46
0
38
0
23
1
43
2
38
0
24
1
27
1
5
1
43
1
35
1
18
1
15
1
25
1
10
1
5
1
35
1
29
1
24
0
3
0
1
1
26
1
11
1
11
1
39
1
33
1
22
1
4
1
21
1
6
1
16
1
2
1
42
1
39
1
7
0
30
0
16
1
8
1
36
1
34
1
46
1
45
1
40
1
38
1
30
1
24
1
19
1
7
1
3
1
47
1
41
1
32
1
31
1
17
1
14
1
Figure 6. Cladogram indicating the strict consensus tree of the data matrix of
the 191 most pasimonous trees
52
Figure 7. Cladogram showing the strict consensus tree of bootstrap analysis
Lithurgus pullatus COS
Megachile semiflava COS
Acanthidium batrae OR
Afranthidium abdominale AF, PA
Afrostelis tegularis AF
Anthidiellum rubellum COS
Anthidioctes willineri NE
Anthidioma murinum AF
Anthidium pontis COS
Apianthidium apiforme OR
Aspidosmia arnoldi AF
Aztecanthidium xochipillium NE
Bathanthidium sibiricum OR
Benanthis madagascariensis AF
Cyphanthidium sheppardii AF
Dianthidium curvatum NE
Duckeanthidium megachiliforme NE
Eoanthidium rothschildi AF, OR, PA
Epanthidium trigrinum NE
Euaspis abdominalis AF, OR
Gnathanthidium prionognathum AF
Hoplostelis catamercensis NE
Hypanthidiodes argentina NE
Hypanthidium flavomarginatum NE
Icteranthidium laterale AF, PA
Indanthidium crenulaticauda OR
Neanthidium octodentatum PA
Notanthidium steloides NE
Pachyanthidium bicolor AR, PA
Paranthidium perpictum NE
Pleisianthidium fulvopilosum AF, OR, PA
Pseudoanthidium truncatum AF, OR, PA
Rhodanthidium siculum OR, PA
Serapista denticulata AF, OR
Trachusa aquilifilum COS
51
78
85
79
87
86
63
83
50
98
51
90
63
91
81
83
51
83
82
80
96
100
53
Figure 8. Cladogram showing the strict consensus tree of jackknife analysis of
clades.
Lithurgus pullatus COS
Megachile semiflava COS
Acanthidium batrae OR
Afranthidium abdominale AF, PA
Afrostelis tegularis AF
Anthidiellum rubellum COS
Anthidioctes willineri NE
Anthidioma murinum AF
Anthidium pontis COS
Apianthidium apiforme OR
Aspidosmia arnoldi AF
Aztecanthidium xochipillium NE
Bathanthidium sibiricum OR
Benanthis madagascariensis AF
Cyphanthidium sheppardii AF
Dianthidium curvatum NE
Duckeanthidium megachiliforme NE
Eoanthidium rothschildi AF, OR, PA
Epanthidium trigrinum NE
Euaspis abdominalis AF, OR
Gnathanthidium prionognathum AF
Hoplostelis catamercensis NE
Hypanthidiodes argentina NE
Hypanthidium flavomarginatum NE
Icteranthidium laterale AF, PA
Indanthidium crenulaticauda OR
Neanthidium octodentatum PA
Notanthidium steloides NE
Pachyanthidium bicolor AR, PA
Paranthidium perpictum NE
Pleisianthidium fulvopilosum AF, OR, PA
Pseudoanthidium truncatum AF, OR, PA
Rhodanthidium siculum OR, PA
Serapista denticulata AF, OR
Trachusa aquilifilum COS
50
78
84
79
85
85
63
85
50
98 50
90
65
90
80
86 50
82
82
79
97
100

54
Systematics of the tribe Anthidiini
General diagnosis of the tribe
Based on the examination of 990 bee specimens, a catalogue on the
diagnostic features of the tribe Anthidiini is presented in telegraphic writing as
follows:
Length: head 1.2-3.0 mm; scutum and scutellum 1.3-5.2 mm; forewing 5.5-
11.2 mm; body 3.9-17.5 mm. Vestiture: mostly white, except greyish in genus
Anthidioma, reddish-brown to black in most parasitic genera and on tarsi of
most taxa; sparse to moderately pubescent; setosed hairs on tibia, basitarsi and
tarsi; female scopa usually on S2-S6, absent in parasitic genera; male sternal
comb on variable number of sclerites in Hypanthidioctes, Neanthidium,
Paranthidium, Rhodanthidium, Trachusa, except Anthidiellum, Benanthis,
Cyphanthidium, Epanthidium, Hoplostelis, Pachyanthdium, Pleisianthidium,
Pseudoanthidium, but absent in Afranthidium, Afrostelis, Anthidium,
Apianthidium, Aspidosmia, Dianthidium, Eoanthidium, Euaspis,
Hypanthidium, Notanthidium, Serapista. Integument: black with cream,
yellow, or reddish maculations, except in some few species with yellow to
reddish-brown, maculations absent in genera Afrostelis, Aspidosmia, Euaspis
and Pleisianthidium; integument punctations moderate to very dense, usually
small in most genera, shiny between punctures; most genera have various
degrees of carinae on paraocular area, preoccipital area, mandible, pronotal
lobe, postero-lateral regions of axilla and scutum; in genera Gnathanthidium
and Pachyanthidium carinae is lamellate; female mandible multidentate,

55
usually four or more teeth, except female of genus Dianthidium with three
teeth, male usually tridentate, except in genus Hypanthidium with mandible
bidentate, clypeus usually broad, dorsal region gently convex, except in genus
Aspidosmia with strongly elevated protruberance; subantennal suture mostly
straight, arcuate in Anthidiellum, Eoanthidium and Pseudoanthidium; omaulus
carinate except in Bathanthidium and Paranthidium; scutum broader than
long, uniquely elongate in genus Afrostelis; axilla rarely extend beyond lateral
regions of scutum, except in genera Acedanthidium, Serapista and
Icteranthidium; scutellum slightly overhungs propodeum in most taxa;
metasoma possess spines on lateral region in a few genera, S5 or S6 and
posterior margin of T6 of some females; T7 of male usually bifid or trifid;
hind tibia subequal or broader than hind basitarsus, except in genus
Gnathanthidium with hind basitarsus broader than hind tibia; hind tibial spur
straight, except in genera Apianthidium, Aztecanthidium and
Duckeranthidium; inner ramus of claw simple, except cleft in genus
Aspidosmia; arolia present in most genera, absent in Anthidium, Trachusa,
Hypanthidiodes and Apianthidium; male gonostylus generally more or less
fused to gonocoxite, forming gonobase.
Genus Acedanthidium Michener and Griswold
Acanthidium Michener and Griswold, 1994: 305. Type species: Acanthidium
batrae Michener and Griswold, 1994, by original designation;
Michener, 2000: 480.

56
Comment
This slender, megachiliform, non-parasitic genus was first described by
Michener and Griswold (1994) using the type species Acanthidium batrae.
Later, Michener (2000) provided brief diagnosis on this monotypic genus,
which is endemic to India.
Diagnosis
Length: head 1.8-1.9 mm; scutum and scutellum 1.8-2.1 mm; forewing
5.5-5.8 mm; body 6.7-7.0 mm. Vestiture: white; moderately dense on head and
mesosoma, sparse on tergites; female scopa on S2-S5; male sternal comb
absent. Integument: black, except yellow maculations on most of lower half of
face, most of paraocular area, entire preoccipital margin, dorsal margin of
tegula, antero-lateral region of scutum, posterior regions of axilla and
scutellum, most of dorsal surfaces of legs, subapical bands on lateral regions
of T1-T6 (slightly narrow on T1-T4, wide on T5-T6 (Pl. 4a)); female mandible
with four teeth; lower clypeus margin with three projections; subantennal
suture straight; juxtantennal carina absent; preoccipital area with dorsal carina;
postero-lateral region of axilla slightly extend beyond lateral magin of scutum
(Pl. 5); scuto-scutellar suture slightly open, forming narrow fovea, scuto-axilla
suture closed; scutellum, in dorsal view, rounded posteriorly, slightly obscures
propodeum; arolia present; hind tibial spur curved; lateral spines on T5, T6;
male gonostylus expanded apically; penis valves narrowly seperated, arcuate
apically (Figure 9a).
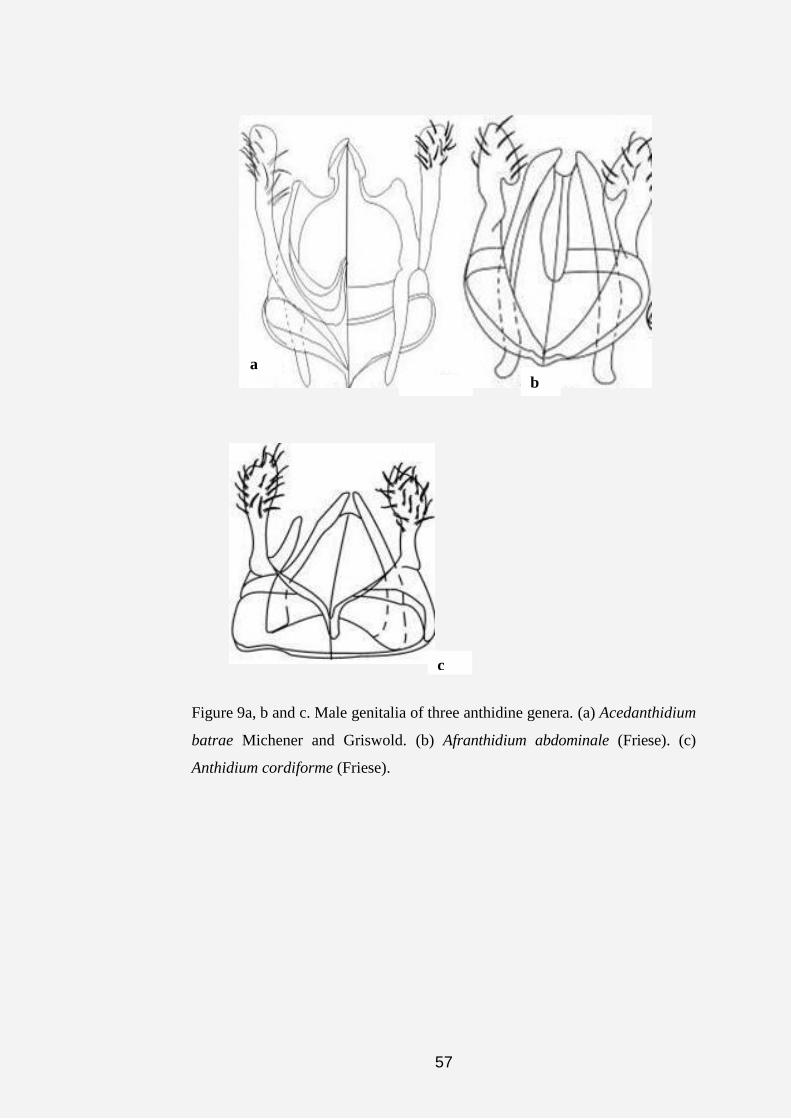
57
Figure 9a, b and c. Male genitalia of three anthidine genera. (a) Acedanthidium
batrae Michener and Griswold. (b) Afranthidium abdominale (Friese). (c)
Anthidium cordiforme (Friese).
a
b Fig. 15
c
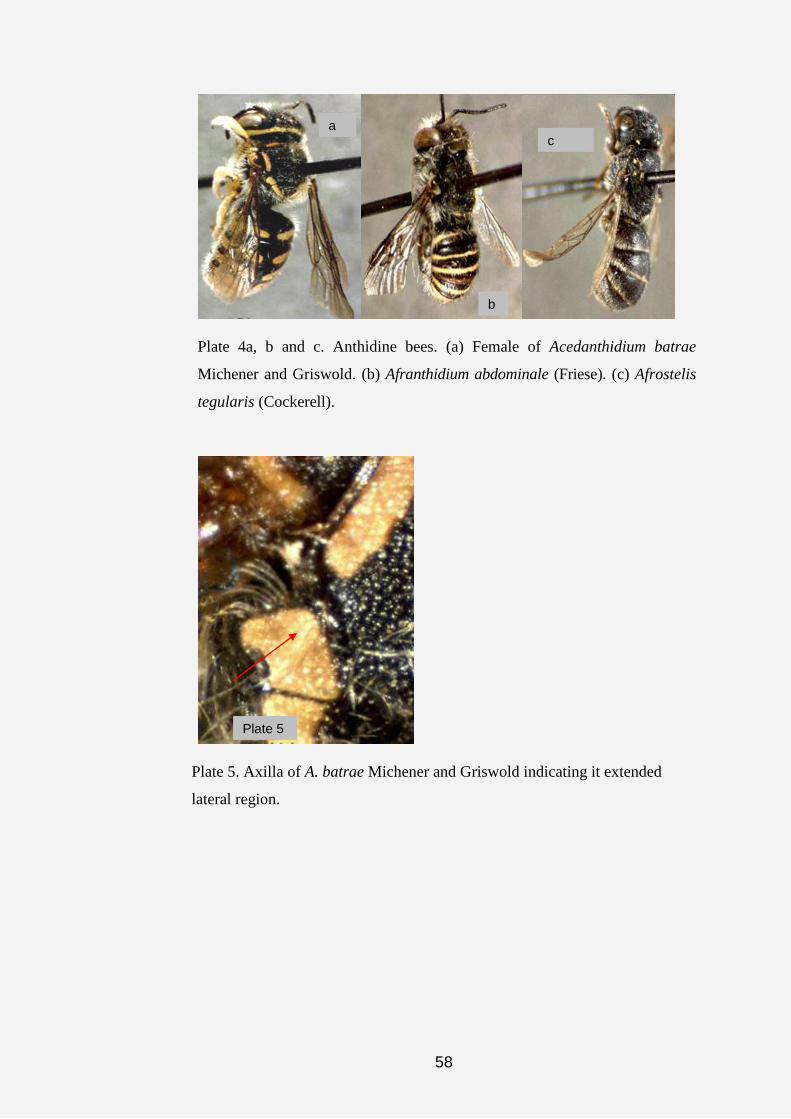
58
Plate 4a, b and c. Anthidine bees. (a) Female of Acedanthidium batrae
Michener and Griswold. (b) Afranthidium abdominale (Friese). (c) Afrostelis
tegularis (Cockerell).
Plate 5. Axilla of A. batrae Michener and Griswold indicating it extended
lateral region.
Plate 5
a
b
c

59
Genus Afranthidium Michener
Subgenus Afranthidium Michener s. str.
Anthidium (Afranthidium) Michener, 1948: 24. Type species: Hypanthidium
haplophilum Cockerell, 1936 by original designation.
Afranthidium (Afranthidium) Michener: Pasteels, 1984: 76-78; Michener and
Griswold, 1994: 309; Michener, 2000: 482.
Subgenus Branthidium Pasteels
Branthidium Pasteels, 1969: 88. Type species: Anthidium braunsi Friese,
1904, by original designation.
Honanthidium Pasteels, 1969: 88. Type species: Anthidium honestum
Cockerell, 1936, by original designation; Pasteels, 1984: 59, syn.
Afranthidium (Branthidium) Pasteels: Michener and Griswold 1994: 309;
Michener 2000: 482.
Subgeus Capanthidium Pasteels
Capanthidium Pasteels, 1969: 85. Type species: Anthidium capicola Brauns,
1905, by original designation; Pasteels, 1984: 64.
Afranthidium (Capanthidium) Pasteels: Michener and Griswold, 1994: 310;
Michener, 2000: 482.
Subgenus Domanthidium Pasteels
Domanthidium Pasteels, 1969: 95. Type species: Anthidium abdominale
Friese, 1904, by original designation.
Afranthidium (Domanthidium) Pasteels: Michener, 2000: 483.
Subgeus Immanthidium Pasteels
Immanthidium Pasteels, 1969: 89. Type species: Anthidium immaculatum
Smith, 1854, by original designation; Pasteels, 1984: 51.

60
Afranthidium (Immanthidium) Pasteels: Michener, 2000: 483.
Subgenus Mesanthidiellum Pasteels
Mesanthidiellum Pasteels, 1969: 83. Type species: Mesanthidiellum amoenum
Pasteels, 1969, by original designation; Pasteels, 1984: 71.
Afranthidium (Mesanthidiellum) Pasteels: Michener, 2000: 483.
Subgenus Mesanthidium Pasteels
Mesanthidium Popov, 1950: 316. Type species: Anthidium pentagonum
Gussakovskiji, 1930, by original designation.
Afranthidium (Mesanthidium) Popov: Michener, 2000: 484.
Subgenus Nigranthidium Pasteels
Melanthidium Pasteels, 1969: 90 (not Cockerell 1947). Type species:
Anthidium concolor Friese, 1913, by original designation.
Nigranthidium Pasteels, 1984: 57. Replacement for Melanthidium Pasteels,
1969.
Warnckeia Pagliano and Scaramozzino, 1990: 6. Replacement for
Melanthidium Pasteels, 1969; Pasteels, 1984: 57.
Afranthidium (Nigranthidium) Pasteels: Michener and Griswold, 1994: 310;
Michener, 2000: 484.
Subgenus Oranthidium Pasteels
Oranthidium Pasteels, 1969: 95. Type species: Anthidium oraniense Brauns,
1905, by original designation; Pasteels, 1984: 83.
Afranthidium (Oranthidium) Pasteels: Michener, 2000: 484.
Subgenus Xenanthidium Pasteels
Xenanthidium Pasteels, 1984: 33. Type species: Xenanthidium biserratum

61
Pasteels, 1984, by original designation.
Afranthidium (Xenanthidium) Pasteels: Michener and Griswold, 1994: 310;
Michener, 2000: 484.
Subgenus Zosteranthidium Michener and Griswold
Afranthidium (Zosteranthidium) Michener and Griswold, 1994: 310. Type
species: Nigranthidium tergofasciatum Pasteels, 1984, by original
designation; Michener, 2000: 485.
Comment
This non-parasitic, small to medium sized, megachiliform genus was
first described by Michener (1948). Later, Michener (2000) recorded 11
subgenera in this genus; all are Afrotropical, and two of these also occur in the
Palaearctic Region. The species chosen for the cladistic analysis were
Afranthidium abdominale. This is because it is the type species of the most
diverse subgenera.
DIAGNOSIS
Length: head 1.5-2.2 mm; scutum and scutellum 1.4-2.6 mm; forewing
4.0-5.8 mm; body 5.5-8.1 mm. Vestiture: white to reddish-brown; sparse to
impubescent, except moderately dense on head and mesosoma in subgenus
Immanthidium; setose hair on tibia; scopa on female S2-S5; male sternal comb
absent. Integument: black (metasoma brown in A. folliculosum) with pale
yellow maculations on entire posterior margins of female T1-T5 and male T1-
T6 (Pl. 4b); lower clypeal margin with six projections; female mandible with 5

62
to 18 sharp teeth; subantennal suture straight; juxtantennal carina absent;
preoccipital area carinate laterally, except subgenus Branthidium with
preoccipital area rounded; scuto-scutellar and scuto-axilla sulci closed;
scutellum, in dorsal view, truncate posteriorly, sometimes obscures
propodeum; arolia absent; T1-T3 without teeth, female T4-T5 with reduced
lateral teeth, male T4-T5 variable; female T6 with posterior margin
denticulate; gonostylus usually expanded apically, inner margin slightly
notched; penis valves slender apically, mostly narrowly separated (Figure 9b).
Genus Afrostelis Cockerell
Afrostelis Cockerell, 1931b: 340. Type species: Afrostelis tegularis Cockerell,
by original designation; Michener and Griswold, 1994: 312; Baker,
1996a: 1-3; Michener, 2000: 485.
Stelis (Afrostelis) Cockerell, 1969: 57; Pasteels, 1984: 134.
Comment
This cleptoparasitic, hoplitiform genus is monotypic. It was first
described by Cockerell (1931b). Later, Pasteels (1969, 1984), Baker (1996a),
Michener and Griswold (1994), Michener (2000) and Pauly (2001), provided
diagnoses of the genus. The genus is endemic to Africa.
Diagnosis
Length: head 1.2-2.1 mm; scutum and scutellum 1.3-1.8 mm; forewing
5.5-6.2 mm; body 3.9-5.6 mm. Vestiture: white; mostly sparse to impubescent;

63
female lacks scopa; male sternal comb absent. Integument: entirely black (Pl.
4c); female mandible with four dissimilar teeth; lower clypeal margin with
three projections; subantennal suture straight; juxtantennal carina absent;
preoccipital area carinate dorsolaterally; scutum uniquely elongated (1:1.5X
length of mesosoma); scuto-scutellar suture closed, similar to scuto-axilla
suture; scutellum, in dorsal view, rounded posteriorly; arolia present;
metasoma lacks lateral teeth; gonostylus of male narrows apically, slightly
notched on outer margin; penis valves narrowly separated.
Genus Anthidiellum Cockerell
Subgenus Ananthidiellum Pasteels
Anthidiellum (Ananthidiellum) Pasteels, 1969: 49-50. Type species: Anthidium
anale Friese, 1914, by original designation; Michener and Griswold,
1994: 312; Michener, 2000: 486.
Subgenus Anthidiellum Cockerell s. str.
Anthidium (Anthidiellum) Cockerell, 1904b: 3. Type species: Trachusa
strigata Panzer, 1805, by original designation.
Anthidium (Cerianthidium) Friese, 1932a: 304. Type species: Trachusa
strigata Panzer, 1805. Isogenotypic with subgenus Anthidiellum.
Anthidiellum (Anthidiellum) Cockerell: Pasteels, 1969: 45; Pasteels, 1984: 92;
Michener and Griswold, 1994: 312; Michener, 2000: 486.
Subgenus Chloranthidiellum Mavromoustakis
Anthidiellum (Chloranthidiellum) Mavromoustakis, 1963a: 491. Type species:
Anthidium flavescens Friese, 1925, by original designation.

64
Anthidiellum (Chloranthidium) [!] Mavromoustakis: Pasteels, 1969: 48.
Anthidiellum (Chloranthidiellum) Mavromoustakis: Pasteels, 1984: 93. Type
species: Anthidiellum minutulum flavescens Friese, 1925; Pasteels,
1984: 92; Michener and Griswold, 1994: 312; Michener, 2000: 487.
Subgenus Clypanthidium Pasteels
Bathanthidium (Clypanthidium) Pasteels, 1968a: 1060; 1969: 53. Type
species: Anthidium ruficeps Friese, 1914, by original designation.
Anthidiellum (Clypanthidium) Pasteels: Michener and Griswold, 1994: 312;
Michener, 2000: 487.
Subgenus Pycnanthidium Krombein
Pycnanthidium Krombein, 1951: 292. Type species: Pycnanthidium solomonis
Krombein, 1951, by original designation.
Pygnanthidium [!] Krombein: Mavromoustakis, 1963: 491.
Pygnanthidium [!] (Pygnanthidiellum) Mavromoustakis, 1963a: 492. Type
species: Anthidium zebra Friese, 1904, by original designation.
Anthidiellum (Pycnanthidium) Krombein: Pasteels, 1969: 45; Michener and
Griswold, 1994: 312; Michener, 2000: 487.
Anthidiellum (Pygnanthidiellum) Mavromoustakis: Pasteels, 1984: 94.
Subgenus Ranthidiellum Pasteels
Anthidiellum (Ranthidiellum) Pasteels, 1969: 48. Type species:
Protoanthidium rufomaculatum Cameron, 1902, by original
designation; Michener and Griswold, 1994: 312; Michener, 2000: 487.
Anthidiellum (Rhanthidiellum) [!] Pasteels: Pasteels, 1972: 102.

65
Comment
The genus Anthidiellum was first described by Cockerell (1904a).
Later, Pasteels (1969) recorded six subgenera; two of these are endemic to the
Afrotropical Region, one is Afrotropical and Oriental, two are endemic to the
Orient and one is Mesoamerica and Holarctic. This genus consists of small,
robust, megachiliform to bombiform, non-parasitic bees.
Diagnosis
Length: head 1.2-1.6 mm; scutum and scutellum 1.3-2.2 mm; forewing
3.2-5.0 mm; body 4.0-7.0 mm. Vestiture: white to reddish-black; sparse to
impubescent; female scopa densely setose on S2-S5; male S5 with comb,
except in nominative subgenus. Integument: mostly black (Chloranthidiellum
with brown metasoma), limited yellow maculation usually on lower half of
face, lateral region of metasoma; mandible of female with four teeth; lower
clypeal margin with four or six projections; subantennal suture strongly
arcuate; juxtantennal carina absent; preoccipital carina present dorsally, except
in nominative subgenus with preoccipital area carinate dorso-laterally and in
subgenus Chloranthidiellum where carina occurs laterally only; scuto-scutellar
suture open, forms shiny fovea; axilla-scutellar suture closed; arolia present;
metasoma lacks lateral spines; gonostylus usually expanded apically, mostly
notched on outer margin; penis valves narrowly separated.

66
Genus Anthidioctes Moure
Subgenus Anthidioctes Holmberg s. str.
Anthidioctes Holmberg, 1887: 36, nomen nudum.
Anthidioctes Holmberg, 1903: 435. Type species: Anthidioctes megachiloides
Holmberg, 1903, designation of Cockerell, 1927: 2; Michener, 1948:
20
Nananthidium Moure, 1947a: 26. Type species: Nananthidium bettyae Moure,
1947, by original designation; Moure, 1947a: 26, syn.
Anthidioctes (Nananthidium) Moure: Griswold and Michener, 1988: 30;
Michener, 2000, syn.
Anthidioctes (Anthidioctes) Holmberg: Michener, 2000: 491.
Subgenus Bothranthidium Moure
Bothranthidium Moure, 1947a: 23. Type species: Bothranthidium lauroi
Moure, 1947, by original designation
Anthidioctes (Bothranthidium) Moure: Michener, 1948: 21; Michener, 2000:
491.
Comment
This genus was first described by Holmberg (1887, 1903). Later,
Moure (1947a) redescribed an additional subgenus. It is known that this small
hoplitiform to slender megachiliform genus of pollen-collecting bees is
endemic to the Neotropical Region with a few species also occuring in
southern Africa.

67
Diagnosis
Length: head 1.2-1.4 mm; scutum and scutellum 1.5-1.8 mm; forewing
3.4-3.9 mm; body 4.5-5.6 mm. Vestiture: white to yellow; sparse to
impubescent; scopa on female S2-S5; male sternal comb absent. Integument:
mostly black or yellowish brown on head, mesosoma and metasoma mostly
black, yellowish-brown pronotum, tegula and legs, yellow on anterolateral
margin of scutum, most of axilla and scutellum, lateral regions of T1-T2, sub-
apical regions of T3-T5 (Plate 6a); female mandible with four dissimilar teeth;
lower clypeal margin with four projections; subantennal suture straight;
juxtantennal carina well developed; preoccipital area carinate laterally; scuto-
scutellar suture open, forming shiny fovae, dissimilar to scutoaxilla suture;
scutellum, in dorsal view, truncate posteriorly; arolia present; metasoma lacks
lateral spines; gonostylus of male slender apically; penis valve widely
seperated.
Genus Anthidioma Pasteels
Anthidioma Pasteels, 1984: 34. Type species: Anthidioma chalicodomoides
Pasteels, 1984, by original designation.
Comment
This non-parasitic, chalicodomiform genus is represented by two
species; namely, A. Chalicodomoides Pasteels and A. Murinum Pasteels. These
were first described by Pasteels (1984). Later, Michener and Griswold (1994)
and Michener (2000), provided a brief diagnosis of this genus. The genus is
endemic to southern Africa.

68
Diagnosis
Male: Unknown. Length: head 2.1-2.4 mm; scutum and scutellum 2.6-
3.0 mm; forewing 5.5-6.0 mm; body 7.4-8.3 mm. Vestiture: grayish white,
moderately dense; postgradular margins of T2-T5 with dense hair bands;
female scopa on S2-S6. Integument: black, except yellow maculations on most
of lower half of face, entire dorsal margin of tegula, dorsolateral margins of
axilla and scutellum, fore, middle and hind tibiae and basitarsi, subapical
bands on T1-T5 (entire width on T1-T4, medial third on T5 (Plate 6b)); female
mandible with seven teeth; lower clypeus margin with four reduced
projections; subantennal suture straight; uxtantennal carina absent;
preoccipital area gently rounded; scuto-scutellar suture open, forming shiny
fovae, with scuto-axilla suture closed; scutellum, in dorsal view, rounded
posteriorly, slightly obscures propodeum; arolia absent; metasoma lacks
lateral spines; posterior margin of female T6 denticulate.
Genus Anthidium Fabricus
Subgenus Anthidium Fabricus s. str.
Anthidium Fabricus, 1804: 364. Type species: Apis manicata Linnaeus, 1758,
by designation of Latreille, 1810: 439.
Anthidium (Anthidium) Fabricus: Pasteels, 1984: 11-17; Michener and
Griswold, 1994: 314; Michener, 2000: 489-490.
Anthidium (Stenanthidium) Moure, 1947a: 16. Type species: Anthidium
espinosai Ruiz, 1938, by original designation; Michener, 1948: 16;

69
Griswold and Michener, 1988: 29, syn.
Anthidium (Tetranthidium) Moure, 1947a: 15. Type species: Anthidium latum
Schrottky, 1902, by original designation; Michener, 1948: 16;
Griswold and Michener, 1988: 29, syn.
Anthidium (Melanthidium) Cockerell, 1947: 106, not Melanthidium Pasteels,
1969. Type species: Melanthidium carri Cockerell, 1947, by original
designation; Michener, 1948: 16; Griswold and Michener, 1988: 299,
syn.
Anthidium (Melanoanthidium) Tkalců, 1967: 91. Type species: Anthidium
montanum Morawitz, 1864, by original designation; Tkalcu: 1967: 91,
syn.
Anthidium (Echinanthidium) Pasteels, 1969: 101. Type species: Anthidium
echinatum Klug, 1864, by original designation; Pasteels, 1969:101,
syn.
Anthidium (Pontanthidium) Pasteels, 1969: 105. Type species: Anthidium
pontis Cockerell, 1933, by original designation; Pasteels, 1969: 105,
syn.
Anthidium (Ardenthidium) Pasteels, 1969: 103. Type species: Anthidium
ardens Smith, 1879, by original designation; Pasteels, 1969: 103, syn.
Anthidium (Morphanthidium) Pasteels, 1969: 423. (Synonym of
Ardenthidium, because two of the included species were clearly
intended by Pasteels to be the same as Ardenthidium, Michener, 2000).
Subgenus Callanthidium Cockerell
Callanthdium Cockerell Cockerell, 1925: 365. Type species: Anthidium

70
illustre Cresson, 1879, by original designation.
Anthidium (Callanthidium) Cockerell: Michener, 2000: 490.
Subgenus Gulanthidium Pasteels
Anthidium (Gulanthidium) Pasteels, 1969: 101. Type species: Anthidium
anguliventre Morawitz, 1888, by original designation; Michener and
Griswold, 1994: 314; Michener, 2000: 490.
Subgenus Nivanthidium Pasteels
Anthidium (Nivanthidium) Pasteels, 1969: 106. Type species: Anthidium
niveocinctum Gerstaecker, 1857, by original designation; Michener
and Griswold, 1994: 313; Michener, 2000: 490.
Subgenus Proanthidium (Friese)
Anthidium (Proanthidium) Friese, 1898: 101. Type species: Anthidium
oblongatum Latreille, 1809, by designation of Cockerell, 1909: 269;
Michener and Griswold, 1994: 314; Michener, 2000: 490-491.
Subgenus Severanthidium Pasteels
Anthidium (Severanthidium) Pasteels, 1969: 106. Type species: Anthidum
severini Vachal, 1903, by original designation; Pasteels, 1984: 18-26;
Michener and Griswold, 1994: 313; Michener, 2000: 491.
Subgenus Turkanthidium Pasteels
Anthidium (Turkanthidium) Pasteels, 1969: 103. Type species: Anthidium
unicum Morawitz, 1875, by original designation; Michener and
Griswold, 1994: 314; Michener, 2000: 491.

71
Comment
Fabricus (1804) first described the genus Anthidium from the type
species Apis manicata (Linnaeus). Several authors have worked on the group
since then. The most recent author being Michener (2000) who listed seven
subgenera. Three of these subgenera are Palaearctic, two are endemic to
Africa, one occurs in the Nearctic Region, and the other subgenus occurs on
all continents, except Australia and it also does not occur in the Indo-Malayan
tropics. This genus comprises of small to medium size, megachilform, pollen-
collecting bees.
Diagnosis
Length: head 2.4-3.1 mm; scutum and scutellum 2.1-3.2 mm; forewing
4.5-7.7 mm; body 5.8-13.8 mm. Vestiture: white; sparse to moderately
pubescent, except densely setose on basitarsus; scopa on female S2-S6; male
sternal comb absent. Integument: black with extensive pallid maculation on
most of lower half of face, dorsal surface of preoccipital area, T1-T5 with
broken cross-bands, continuous in subgenus Anthidium, (maculation yellow,
except reddish in subgenera Anthidium and Severanthidium and white in
subgenus Nivanthidium); female mandible with 5-10 teeth; lower clypeal
margin with one lateral projection; subantennal suture nearly straight;
juxtantennal carina absent; preoccipital area gently rounded; scuto-scutellar
and scuto-axilla suture closed; scutellum, in dorsal view, truncate posteriorly,
sometimes obscures propodeum; arolia absent; metasomal spines variable,
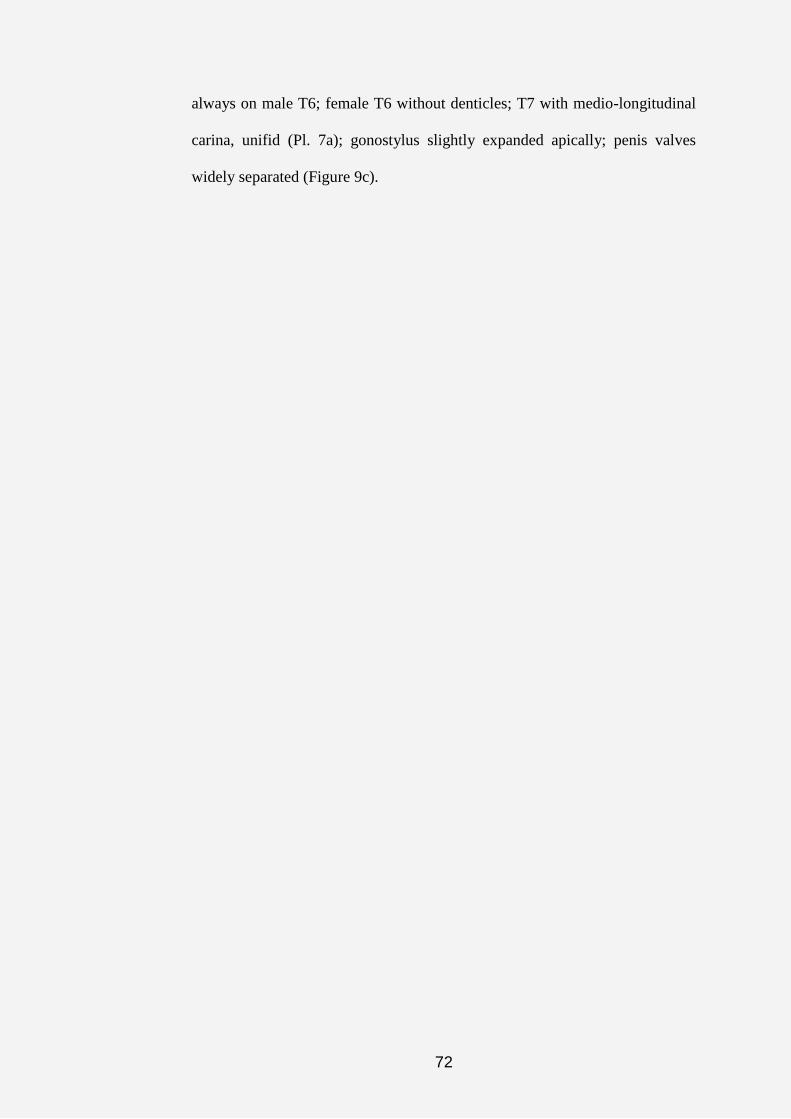
72
always on male T6; female T6 without denticles; T7 with medio-longitudinal
carina, unifid (Pl. 7a); gonostylus slightly expanded apically; penis valves
widely separated (Figure 9c).
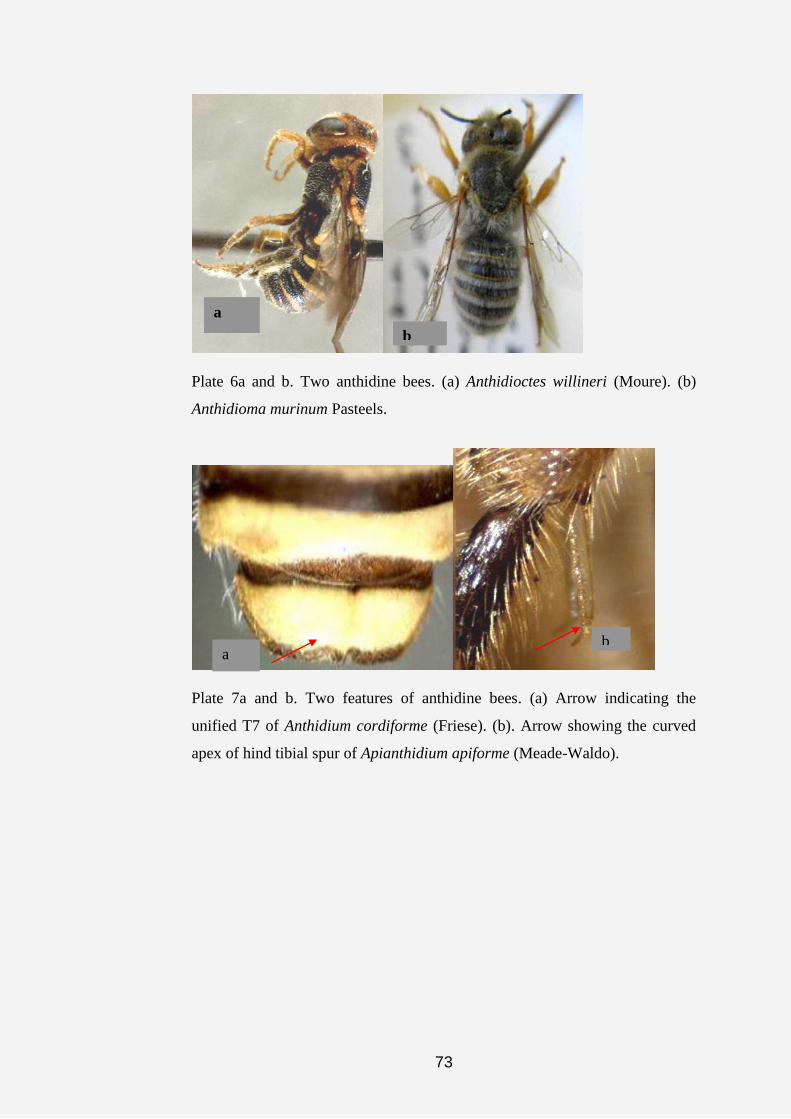
73
Plate 6a and b. Two anthidine bees. (a) Anthidioctes willineri (Moure). (b)
Anthidioma murinum Pasteels.
Plate 7a and b. Two features of anthidine bees. (a) Arrow indicating the
unified T7 of Anthidium cordiforme (Friese). (b). Arrow showing the curved
apex of hind tibial spur of Apianthidium apiforme (Meade-Waldo).
a
b
b a

74
Genus Apianthidium Pasteels
Apianthidium Pasteels, 1969: 41. Type species: Anthidium apiforme Meade-
Waldo, 1914, by original designation; Michener, 2000: 492.
Comment
This megachiliform, non-parasitic genus was first described by
Pasteels (1969) and later by Michener (2000). It is monotypic, and occurs in
Borneo.
Diagnosis
Length: head 2.6 mm; scutum and scutellum 3.2 mm; forewing 6.5
mm; body 10.3 mm. Vestiture: cream to white; sparse to impubescent; male
sternal comb absent; thick setae on tibia absent. Integument: brownish yellow,
except reddish brown maculations apex of mandible; most of supra- antennal
area, medio-longitudinal region of scutum, posterior regions of T1-T7; lower
clypeal margin with 5 projections (1 medial, 2 lateral); subantennal suture
more or less straight; juxtantennal carina absent; preoccipital area gently
rounded; scuto-scutellar suture closed; scutellum, in dorsal view, emarginate
medio-posteriorly; arolia absent; hind tibial spur curved apically (Plate 7b);
metasoma lacks lateral spines; gonostylus expand apically, notched on outer
margin; penis valves narrow apically, widely separated (Figure 10a).
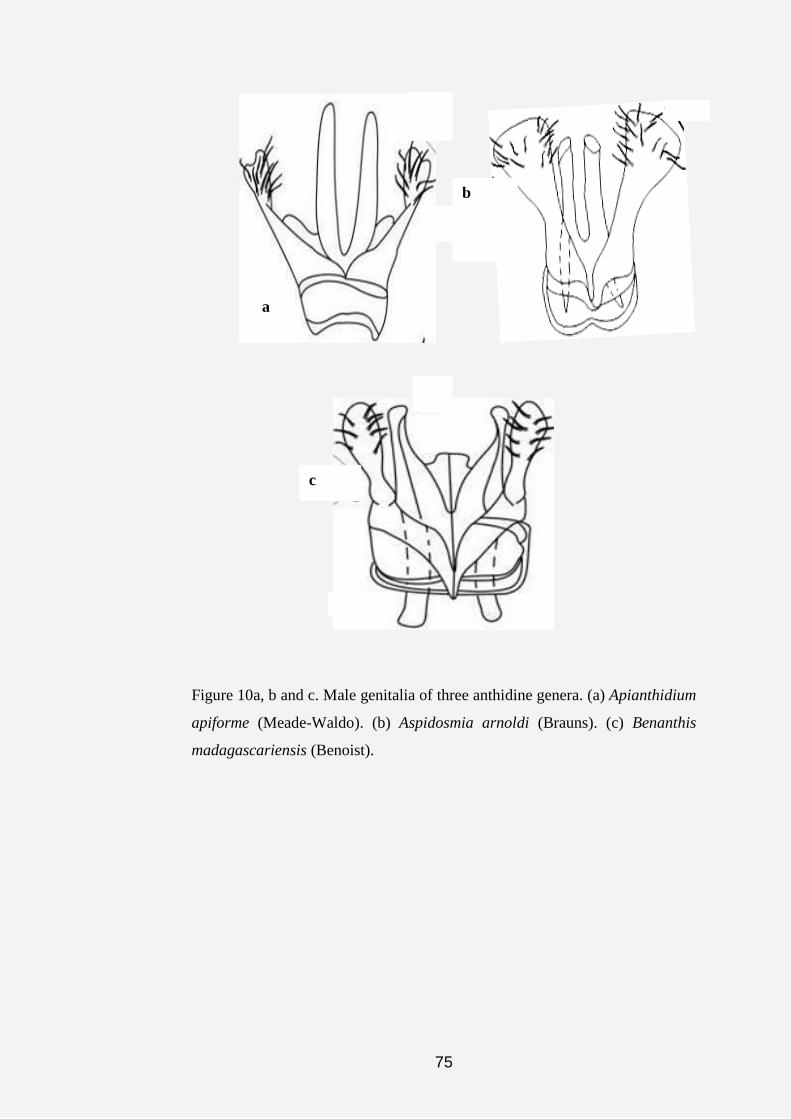
75
Figure 10a, b and c. Male genitalia of three anthidine genera. (a) Apianthidium
apiforme (Meade-Waldo). (b) Aspidosmia arnoldi (Brauns). (c) Benanthis
madagascariensis (Benoist).
Pl. 1
a
2
Fig
. 17
b
Fig
. 17
c
Fig. 17
Fi
g.
19
Fig.
17
Fig
. 17

76
Genus Aspidosmia (Brauns)
Osmia (Aspidosmia) Brauns, 1926a: 208. Type species: Osmia arnoldi Brauns
1926a, monobasic.
Aspidosmia Brauns: Peters, 1972a: 167-169, 170, 171, 173, 185; Peters, 1984:
359, 362; Struck, 1992: 71, 76, 80; Struck, 1994a: 15; Michener, 2000:
492.
Comment
This monotypic genus was first described by Brauns (1926) as a subgenus of
genus Osmia due to its striking morphological similarity with species of
Osmia s. str. Later, Peters (1972a) gave Aspidosmia generic status. This genus
comprises mostly of robust megachiliform, non-parasitic bees that are endemic
to southern Africa.
Diagnosis
Length: head 2.2-2.5 mm; scutum and scutellum 2.5-2.8 mm; forewing
5.5-6.1 mm; body 8.1-10.0 mm. Vestiture: white; mostly moderately dense to
very dense; female scopa on S2-S5; male sternal comb absent. Integument:
mostly dark brown to black, yellow on male clypeus; female mandible with
three similar teeth; lower clypeal margin without projection, female clypeus,
in lateral view, uniquely tuberculate (Figure 3c); preoccipital area gently
rounded; subantennal suture straight; juxtantennal carina absent; scuto-
scutellar suture closed, similar to scuto-axilla suture; scutellum, in dorsal
view, rounded posteriorly; arolia present; inner ramus of tarsal claw uniquely
Fig.
17

77
reduced and adjacent to outer claw; metasomal without lateral teeth;
gonostylus greatly expanded apically, without marginal notch; penis valves
narrowly separated (Figure 10b).
Genus Atropium Smith
Atropium Pasteels 1984: 131-32. Type species: Megachile atropos Smith,
1853, by original designation; Michener 2000: 493.
Comment
This robust megachiliform, non-parasitic genus was first described by
Smith (1853). It is known by a single female specimen of Atropium atropos
(Smith), which is endemic to Gambia. This genus was not available for
examination during this study, due to the fact that, it is a rare specimen and
therefore, the museum was unwilling to loan it out.
Genus Aztecanthidium Michener and Ordway
Aztecanthidium Michener and Ordway, 1964: 70. Type species:
Aztecanthidium xochipillium Michener and Ordway, 1964, original
designation; Michener, 2000: 493.
Comment
The genus was described first by Michener and Ordway (1964). The only
three representative species were revised by Snelling (1987). Michener (2000)
later provided a brief diagnosis on the genus. This large-sized,
chalicodomiform, pollen-collecting genus is endemic to Mexico.

78
Diagnosis
Length: head 2.6-2.9 mm; scutum and scutellum 3.4-3.9 mm; forewing 7.7-8.1
mm; body 10.3-11.6 mm. Vestiture: cream to white; sparse to impubescent;
female scopa on S2-S6; male sternal comb absent; thick setae absent on tibiae.
Integument: mostly black with reddish or yellow maculations on most of
lower half of face of female, only on most of dorsal region of clypeus in male
(mandible black), antero-lateral region of scutum, posterior region of
scutellum, lateral regions of T1-T2, medio-posterior margin T3-T4; lower
clypeal margin with numerous projections; subantennal suture more or less
straight; marginated carina absent; preoccipital area with dorso-lateral
carina; scuto-scutellar and scuto-axilla sulci closed; posterior region of axilla
slightly produced backward (Plate 8a); scutellum, in dorsal view, emarginated
medio-posteriorly; arolia present; hind tibial spur curved apically (Plate 8b);
T5-T6 with two lateral spines per segment, T7 bilobed.
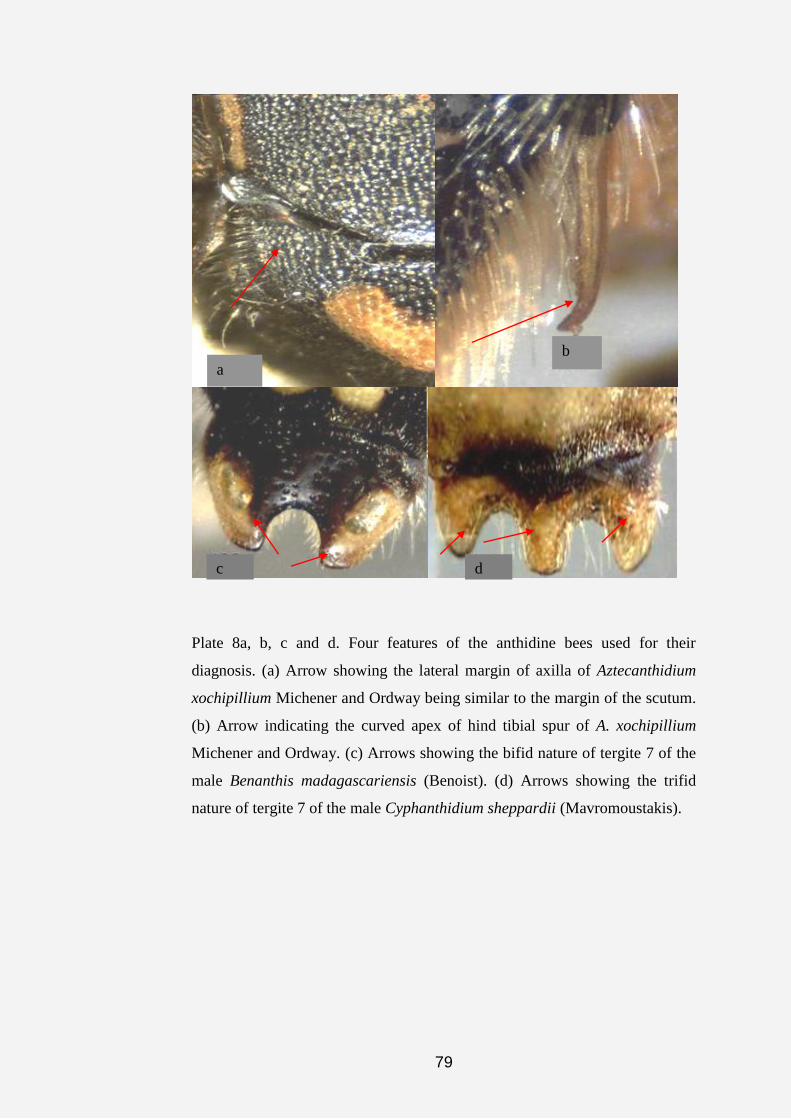
79
Plate 8a, b, c and d. Four features of the anthidine bees used for their
diagnosis. (a) Arrow showing the lateral margin of axilla of Aztecanthidium
xochipillium Michener and Ordway being similar to the margin of the scutum.
(b) Arrow indicating the curved apex of hind tibial spur of A. xochipillium
Michener and Ordway. (c) Arrows showing the bifid nature of tergite 7 of the
male Benanthis madagascariensis (Benoist). (d) Arrows showing the trifid
nature of tergite 7 of the male Cyphanthidium sheppardii (Mavromoustakis).
a
b
c d

80
Genus Bathanthidium Mavromoustakis
Subgenus Bathanthidium Mavromoustakis s. str.
Dianthdium (Bathanthidium) Mavromoustakis, 1953: 837. Type species:
Dianthidium bifoveolatum Alfken, 1937, by original designation;
Bathanthidium (Bathanthidium) Mavromoustakis: Michener and Griswold,
1994: 314; Michener, 2000: 494.
Subgenus Manthidium Pasteels
Manthidium Pasteels, 1969: 43. Type species: Anthidium binghami Friese,
1901, by original designation; Michener, 2000: 494.
Bathanthidium (Manthidium) Pasteels: Michener and Griswold, 1994: 314;
Michener, 2000: 494.
Subgenus Stenanthidiellum Pasteels
Bathanthidium (Stenanthidium) Pasteels, 1968a: 1059. Type species:
Anthidium sibiricum Eversmann, 1852, by original designation.
? Lasanthidium Romankova, 1988: 26. Type species: Stelis malaisei Popov,
1941, by original designation, syn.
Bathanthidium (Stenanthidiellum) Pasteels; Michener and Griswold; 1994:
314; Michener, 2000: 494
Comment
The genus Bathanthidium was first described by Mavromoustakis
(1953). Later, other authors, such as Pasteels (1968a, 1969), Romankova
(1988), and Michener and Griswold (1994) worked on the genus. Michener
(2000) listed

81
three subgenera. Two of these subgenera are endemic to the Oriental Region
and the other occurs in the eastern Palaearctic Region. This genus consists of
small hoplotiform to slender megachiliform, non-parasitic bees.
Diagnosis
Length: head 1.8 mm; scutum and scutellum 1.6 mm; forewing 4.1
mm; body 5.8 mm. Vestiture: white, sparse to impubescent; female scope on
S2-S6. Integument: mostly black, with limited yellow maculations paraoccular
area, lateral regions of metasoma; female mandible with four dissimilarly
teeth; lower clypeal margin without projection; subantennal suture straight;
juxtantennal carina absent; preoccipital area carinate dorsally; scuto-scutellar
and scuto-axilla suture slightly open, forming fovea; scutellum, in dorsal view,
emarginate medio-posteriorly; arolia present; metasoma lacks lateral spines.
Genus Benanthis (Pasteels)
Benanthis Pasteels, 1969: 61, 63. Type species: Anthidium madagascariensis
Benoist, 1963, by original designation; Pasteels, 1984: 133; Michener
and Griswold, 1994: 301, 314; Michener, 1997b: 10; Michener, 2000:
495; Pauly et. al., 2001: 226-228.
Comment
Pasteels (1969) described the genus Benanthis for a species that was
originally described by Benoist (1962) in the genus Anthidium. It is still the
only described species in this genus of medium to large megachiliform, non-

82
parasitic bees. It is endemis to Madagascar
Diagnosis
Female unknown. Length: head 2.0 mm; scutum and scutellum 2.1 mm;
forewing 6.1 mm; body 7.1 mm. Vestiture: white; sparse to impubescent; male
sternal comb present on S6. Integument: mostly black, tegula and legs brown,
yellow maculation on most of lower half of face, paraoccular area, dorsolateral
region of preoccital area, part of anteior region of scutum, lateral margins of
axilla and scutellum, lateral regions of T1-T6 (narrows medially on T1-T4);
lower clypeal margin without projection; subantennal suture more or less
straight; juxtantennal carina absent; preoccipital area rounded; scuto-scutellar
suture open, forming shiny fovae, scutoaxilla suture closed; scutellum, in
dorsal view, rounded posteriorly; arolia present; metasomal lateral spine
absent, T7 with unique medio-longitudinal carina, bifid (Plate 8c); gonostylus
expanded apically; penis valves widely separated, apex slightly expanded
(Figure 10c).
Genus Cyphanthidium Pasteels
Cyphanthidium Pasteels, 1969: 57. Type species: Cyphanthidium intermedium
Pasteels, 1969, by original designation; Michener and Griswold 1994:
314; Michener 2000: 495.
Trianthidiellum Pasteels, 1969: 58. Type species: Hypanthidium sheppardi
Mavromoustakis, 1937b, by original designation; Pasteels: 1969: 58,
syn.

83
Anthidiellum (Trianthidiellum) Mavromoustakis: Pasteels 1984: 113-114, syn.
Comment
This non-parasitic, medium sized, megachiliform genus is represented by two
species. It was first described by Pasteels (1969), and it is known to comprise
two representative species. Cyphanthidium is endemic to southern Africa.
Diagnosis
Length: head 2.3 mm; scutum and scutellum 2.9 mm; forewing 5.7
mm; body 8.8 mm. Vestiture: yellow to brown; sparse to impubescent; male
sternal comb absent. Integument: black, except yellow maculations on most of
lower half of face, paraoccular area, spot below medial ocellus, distal region
of pronotum, anterior margin of tegula, anterolateral margin of scutum, lateral
margin of axilla, posterolateral margin of scutellum, legs, lateral region of T1,
band on anterior surfaces of T2-T6, most of T6; female mandible with four
dissimilar teeth; lower clypeal margin with four projections; subantennal
suture more or less straight; juxtantennal carina absent; preoccipital area
gently rounded; pronotal carina uniquely lamellate; scuto-scutellar and scuto-
axilla sulci closed; scutellum, in dorsal view, truncate posteriorly; arolia
present; metasoma with lateral spines, T7 trifid (Plate 8d); gonostylus slender
apically; penis narrow apically (Figure 11a).
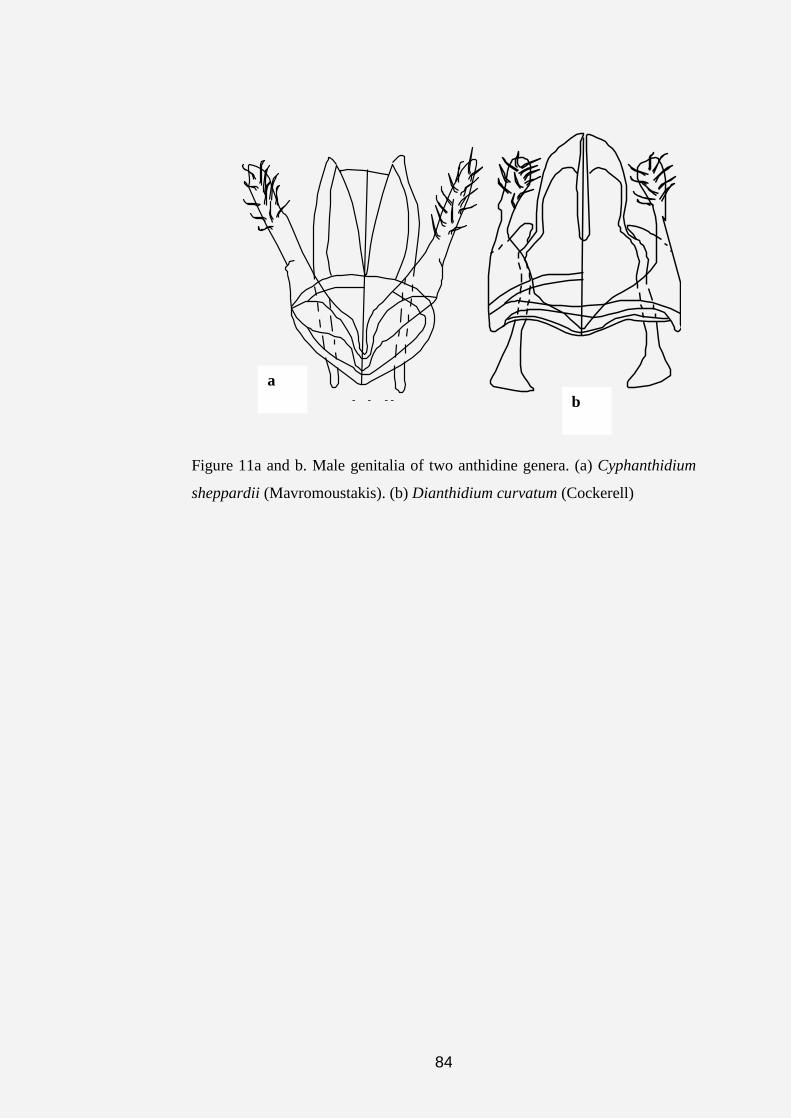
84
Cyphanthidium Dianthidium
Figure 11a and b. Male genitalia of two anthidine genera. (a) Cyphanthidium
sheppardii (Mavromoustakis). (b) Dianthidium curvatum (Cockerell)
a
b

85
Genus Dianthidium (Cockerell)
Subgenus Adanthidium Moure
Adanthidium Moure, 1965: 29. Type species: Anthidium texanum Cresson,
1878, by original designation
Dianthidium (Adanthidium) Moure: Michener, 2000: 496.
Subgenus Deranchanthidum Griswold and Michener
Dianthidium (Deranchanthidium) Griswold and Michener, 1988: 34. Type
species: Dianthidium chamela Griswold and Michener, 1988, by
original designation; Michener, 2000: 496.
Subgenus Dianthidium Cockerell s. str.
Anthidium (Dianthidium) Cockerell, 1900: 412. Type species: Anthidium
curvatum Smith, 1854, by original designation.
Dianthidium (Dianthidium) Cockerell: Michener, 2000: 496.
Subgenus Mecanthidium Michener
Paranthidium (Mecanthidium) Michener, 1942: 278. Type species:
Paranthidium sonorum Michener, 1942, by original designation;
Michnener, 1944: 257.
Dianthidium (Mecanthidium) Michener: Michener, 2000: 496.
Comment
This genus was first described by Cockerell (1900). Later, other
authors such as, Moure (1965), Griswold and Michener (1988), and Michener
(1942, 2000) worked on the group. Michener (2000) recorded four subgenera.

86
One is endemic to the Nearctic Region; two are only known from Mexico, and
two occur in South Africa. It consists of mostly small to medium sized,
chalicodomiform, pollen-collecting bees.
Diagnosis
Length: head 1.6-2.7 mm; scutum and scutellum 1.8-3.7 mm; forewing
4.2-7.7 mm; body 5.2-10.6 mm. Vestiture: white; sparse to impubescent;
female scopa on S2-S5; male sternal comb absent, except in Adanthidium and
Dianthidium with comb on S5; thick setae. Integument: black, except reddish
or yellow maculations on most of lower half of face, medial region of
supraclypeal area, entire paraoccular area, dorsolateral margin of preoccipital
ridge, distal margin of scutum, dorsolateral margin of axilla and scutellum,
metasoma (variable as in Plate 9a); female mandible with four dissimilar teeth;
lower clypeal margin with four reduced projections; subantennal suture more
or less straight; juxtantennal carina reduced; preoccipital ridge reduced
dorsally in most subgenera; scuto-scutellar suture closed; scutellum, in dorsal
view, truncate posteriorly; arolia present except in Deranchanthidium; lack
lateral metasomal spines; female T6 lacks carina; gonostylus narrow apically;
penis valves greatly expanded apically, narrowly separated (Figure11b, Plate
9b).
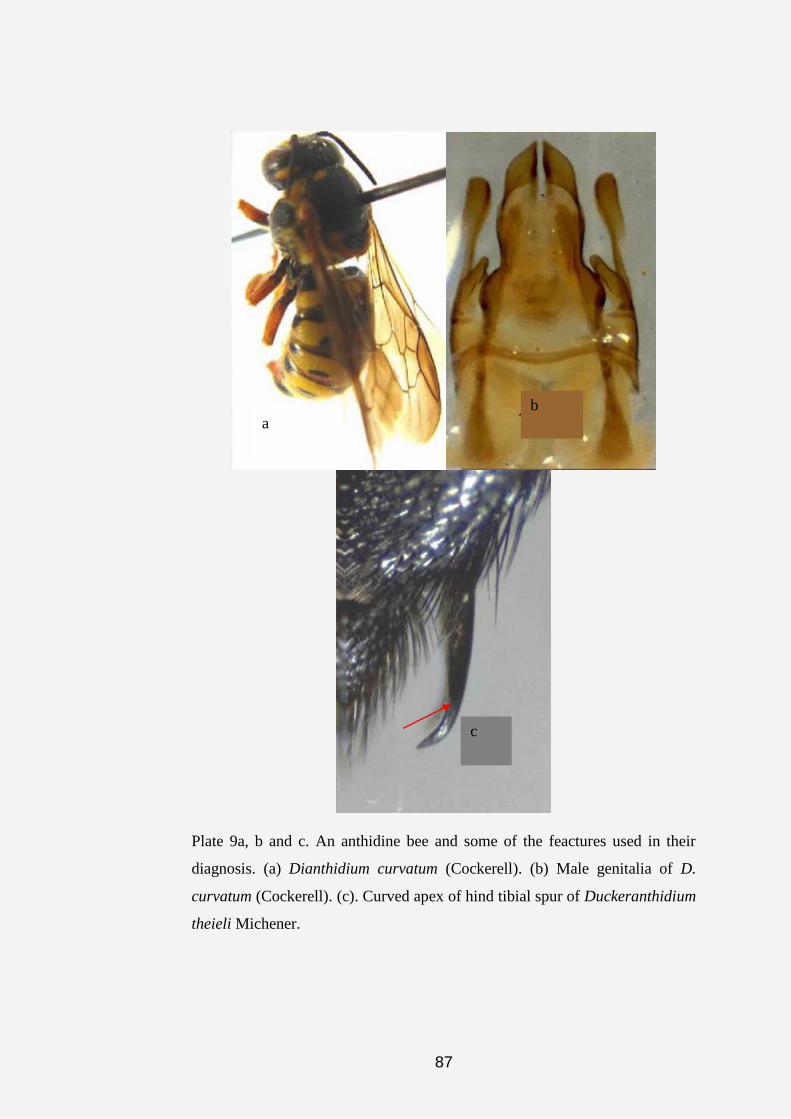
87
Plate 9a, b and c. An anthidine bee and some of the feactures used in their
diagnosis. (a) Dianthidium curvatum (Cockerell). (b) Male genitalia of D.
curvatum (Cockerell). (c). Curved apex of hind tibial spur of Duckeranthidium
theieli Michener.
b a
c

88
Genus Duckeanthidium Moure and Hurd
Duckeanthidium Moure and Hurd, 1960: 2. Type species: Anthidium
megachiliforme Ducke, 1907, by original designation; Michener, 2000:
496-497.
Grafanthidium Urban, 1995: 435. Type species: Grafanthidium amazonense
Urban, 1995, by original designation; Urban; 1995: 435, syn.
Comment
This non-parasitic, robust, chalicodomiform genus was first described
by Moure and Hurd (1960). Later, five species were described by Urban
(1995). Michener (2000) synonymized Grafanthidium and Duckeranthidium.
All of the species are endemic to Brazil.
Diagnosis
Length: head 3.2-3.5 mm; scutum and scutellum 3.9-4.5 mm; forewing
7.7-9.2 mm; body 10.5-12.3 mm. Vestiture: black; moderately dense, except
uniquely long and dense on fore, middle and hind barsitarsi and tarsi of male;
male sternal comb absent; female scopa on S2-S6. Integument: black with
reduced integumental maculation on lower region of male clypeus, lateral
regions of T1-T4; female mandible with four teeth; lower clypeal margin with
6 projections (1 medial, 2 lateral); subantennal suture more or less straight;
juxtantennal carina well developed; preoccipital area lacks carina; scuto-
scutellar suture open, forming narrow fovea; scutellum, in dorsal view,

89
emarginate medio-posteriorly; arolia present in male, absent in female; hind
tibial spur strongly curved apically (Plate 9c); T6 of male possess spine of
lateral third region, T7 with strong lateral lobe and a median spine.
Genus Eoanthidium Popov
Subgenus Clistanthidium Michener and Griswold
Eoanthidium (Clistanthidium) Michener and Griswold, 1994: 315. Type
species: Dianthidium turnericum Mavromoustakis, 1934, by original
designation; Michener and Griswold, 1994: 315; Michener, 2000: 497.
Subgenus Eoanthidium Popov s. str.
Dianthdium (Eoanthidium) Popov, 1950: 316. Type species: Anthidium
insulare, Morawitz, 1873, by original designation.
Eoanthidium (Eoanthidium) Pasteels, 1969: 51. Type species: Anthidium
elongatum Friese, 1897, by original designation; Michener and
Griswold, 1994: 315; Michener, 2000: 497.
Subgenus Hemidiellum Pasteels
Eoanthidium (Hemidiellum) Pasteels, 1972: 112. Type species: Eoanthidium
semicarinatum Pasteels, 1972, monobasic; Michener and Griswold,
1994: 315; Michener, 2000: 498.
Subgenus Salemanthidium Pasteels
Eoanthidium (Salemanthidium) Pasteels, 1969: 51. Type species:
Hypanthidium salemanse Cockerell, 1919, by original designation;
Michener and Griswold, 1994: 315; Michener, 2000: 498.

90
Comment
Popov (1950) was the first to have described this non-parasitic, small
megachiliform to slender bombiform genus. Several authors have worked on
this genus since 1950. Michener (2000) listed four subgenera, two of which
are endemic to Africa and the Palaearctic Regions. The other two occur in
India.
Diagnosis
Length: head 1.9-2.6 mm; scutum and scutellum 2.3-2.7 mm; forewing
4.8-6.3 mm; body .5.8-10.5 mm. Vestiture: yellow to brown; sparse to
impubescent; female scopa on S2-S6; male lacks sternal comb. Integument:
mostly black, Clistanthidium metasoma brownish, extensive yellow
maculation on face, dorsal margins of scutum and scutellum, entire legs,
meso-lateral regions of T1-T3 (Plates 10a & b); female mandible with four
similar teeth; lower clypeal margin with one ventro-lateral projection;
subantennal suture strongly arcuate; juxtantennal carina well developed (Plate
11a); preoccipital area gently rounded (Plate 11b); scuto-scutellar suture open,
forming shiny fovae (closed in Salemanthidium); scutellum, in dorsal view,
rounded posteriorly; arolia absent, except in Salemanthidium; T6 of male with
lateral spine; gonostylus slender apically; penis valves narrow apically (Figure
12a).
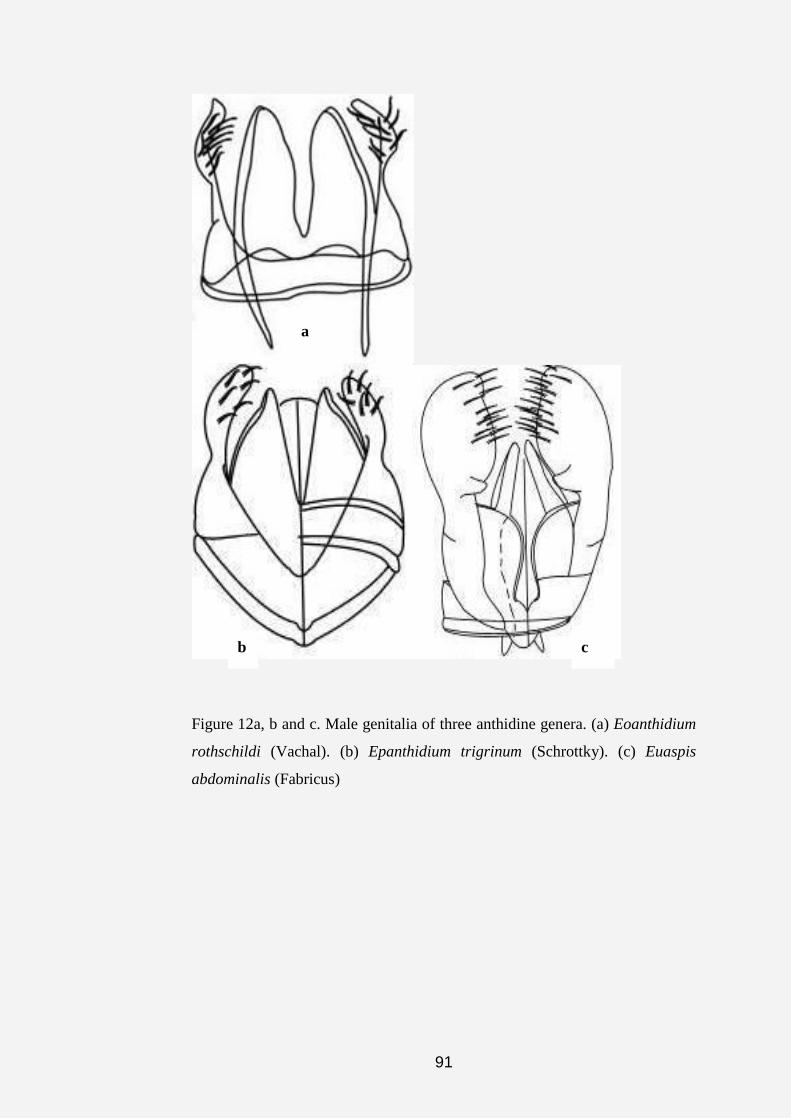
91
Figure 12a, b and c. Male genitalia of three anthidine genera. (a) Eoanthidium
rothschildi (Vachal). (b) Epanthidium trigrinum (Schrottky). (c) Euaspis
abdominalis (Fabricus)
a
c b

92
Plate 10a, b and c. Three anthidine bees, in dorsolateral view. (a) Eoanthidium
rothschildi (Vachal). (b) Eoanthidium turnericum (Mavromoustakis). (c)
Euaspis abdominalis (Fabricus).
Plate 11a, b, c and d. Four diagnostic features of anthidine bees. (a) Face of
Eoanthidium rothschildi (Vachal) with arrows indicating the juxtantennal
carina and actuate nature of the subantennal suture. (b) Postero-lateral region
of E. rothschildi (Vachal) with arrow showing the gently rounded preoccipital
area. (c) Arrow showing the straight apex of the hind tibial spur of Euaspis
abdominalis (Fabricus). (d) Arrows showing the trifid nature of tergite 7 of the
male of Euaspis abdominalis (Fabricus).
a b
a
b
c d
c

93
Genus Epanthidium Moure
Subgenus Ananthidium Urban
Ananthidium Urban, 1991: 73. Type species: Anthidium inerme Friese, 1908,
by original designation.
Epanthidium (Ananthidium) Urban: Michener, 2000: 498.
Subgenus Carloticola Moure and Urban
Carloticola Moure and Urban, 1990: 90. Type species: Dianthidium
paraguayense Schrottky, 1908, by original designation.
Epanthidium (Carloticola) Moure and Urban: Michener, 2000: 499.
Subgenus Epanthidium Moure s. str.
Epanthidium Moure, 1947a: 33. Type species: Hypanthdium tigrinum
Schrottky, 1905, by original designation; Stange, 1983: 281: 297.
Epanthidium (Epanthidium) Moure: Michener, 2000: 499.
Comment
Moure (1947a) first described this non-parasitic genus. Later, Stange
(1983), Moure and Urban (1990) Urban (1991) and Michener (2000) worked
on this genus. Michener (2000) recorded three subgenera, two are endemic to
South America and the other occurs in both South America and Mexico. It
comprises small chalicodomifom bees.
Diagnosis
Length: head 2.0-2.7 mm; scutum and scutellum 2.0-2.7 mm; forewing
4.9-6.8 mm; body 6.5-9.0 mm. Vestiture: white; moderate to sparse, except
dense on tibiae; female scopa on S2-S5; male sternal comb on S3.

94
Integument: black; limited yellow maculation on face, dorsal margin of
preoccipital rigde, most of tegula, spots on dorsal surfaces of scutum, axilla,
scutellum and metasoma; mandible of female with five dissimilar teeth; lower
clypeal margin without projection; subantennal suture straight; juxtantennal
carina well developed; preoccipital area gently rounded; scuto-scutellar scuto-
axilla suture closed; scutellum, in dorsal view, rounded posteriorly; arolia
present; metasomal lateral spines absent, except in nominative subgenus;
female with unique medial carina on T6; female S6 with unique submarginal
carina, sometimes multidentated; S7 of male trifid (subgenus Epanthidium) or
bifid (subgenera Ananthidium and Carloticola); gonostylus of male slightly
narrow apically; penis valves narrows at apex, widely separated (Figure 12b).
Genus Euaspis Gerstaecker
Euaspis Gerstaecker, 1857: 460. Type species: Thynnus abdominalis
Fabricius, 1793, by original designation; Vachal, 1903: 376; Bingham,
1903: 60; Friese, 1909a: 88, 100, 101, 325, 343, 440, 442; Strand,
1912a: 129; Strand, 1912b: 311; Strand, 1914: 67; Friese, 1914: 294;
Friese, 1921: 1104; Cockerell, 1933d: 267-268; Cockerell, 1933e: 1;
Pasteels, 1969: 64; Gess, 1981: 26, 61; Medler, 1980: 482; li Enkulu,
1988: 212; Michener, 1997: 23; Pauly, 1998: 28.
Dilobopeltis Fairmaire, 1858: 266. Type species: Diolobopeltis fuscipennis
Fairmaire, 1858 (= Anthidium abdominale Fabricius 1793), by original
designation, Thomson, 1858: 266, syn.
Parevaspis Ritsema, 1874: 1xxi. Type species: Parevaspis basalis Ritsema,

95
1874, designated by Sandhouse, 1943: 585; Baker, 1995: 282, syn.
Comment
This cleptoparasitic, medium to large, chalicodomiform genus was first
described by Gerstaecker (1857). Since then, other authors, such as Fairmaire
(1858), Vachal (1903), Friese (1909a, 1912b, 1914, and 1921) and Cockerell
(1933e), worked on this genus. Michener (2000) recorded 12 species two were
listed as Afrotropical. The other ten are Oriental.
Diagnosis
Length: head 2.7-3.2 mm; scutum and scutellum 3.7-5.2 mm; forewing
7.2-10.3 mm; body 7.5-15.7 mm. Vestiture: reddish brown to black; sparse to
impubescent; female scopa absent; male sternal comb absent. Integument:
head and mesosoma black, metasoma reddish-brown, yellow maculation
absent (Plate 10c); female mandible with four dissimilar teeth; lower clypeal
margin with six projections; subantennal suture striaght; juxtantennal carina
strongly developed, uniquely extending a little behind antennal socket;
mediolongitudinal margin of face strongly carinate; preoccipital carina
strongly developed laterally; scuto-scutellar and scuto-axilla suture closed;
scutellum, in dorsal view, emarginate posteromedially, with two flat, plate-like
lobes projecting backwards; hind tibial spur straight (Plate 11c); arolia present;
metasomal spines absent; T7 trilobed (Plate 11d); male gonostylus notched on
inner margin, apical region greatly expanded; penis valves reduced, narrowly
separated (Figure 12c).

96
Genus Gnathanthidium Pasteels
Gnathanthidium Pasteels, 1969: 92-93. Type species: Pachyanthidium
prionognathum Mavromoustakis, 1935, original designation; Pasteels
1984: 36; Michener 2000: 499-500.
Comment
This non-parasitic genus was first described by Pasteels (1969). It is
monotypic and endemic to eastern and southern Africa.
Diagnosis
Length: head 2.4-2.7 mm; scutum and scutellum 2.7-3.1 mm; forewing
6.7-7.2 mm; body 7.7-9.4 mm. Vestiture: white; sparse to impubescent; female
scopa on S2-S5. Integument: black, with reddish-brown legs; yellow or
reddish maculations on dorsolateral portion of preoccipital ridge (Plate 12c);
lateral regions of T1-T5 (Plate 12a); female mandible with 13 dissimilar teeth
(Plate 12b); lower clypeal margin with six projections; subantennal suture
straight; juxtantennal carina reduced; preoccipital area carinate or and
lamellate dorsolaterally (Plate 12c); scuto-scutellar suture closed, similar to
scutoaxilla suture; scutellum, in dorsal view, strongly truncate; arolia present;
hind barsitarsus unusually broad (Plate 12d); metasomal lateral spines absent.
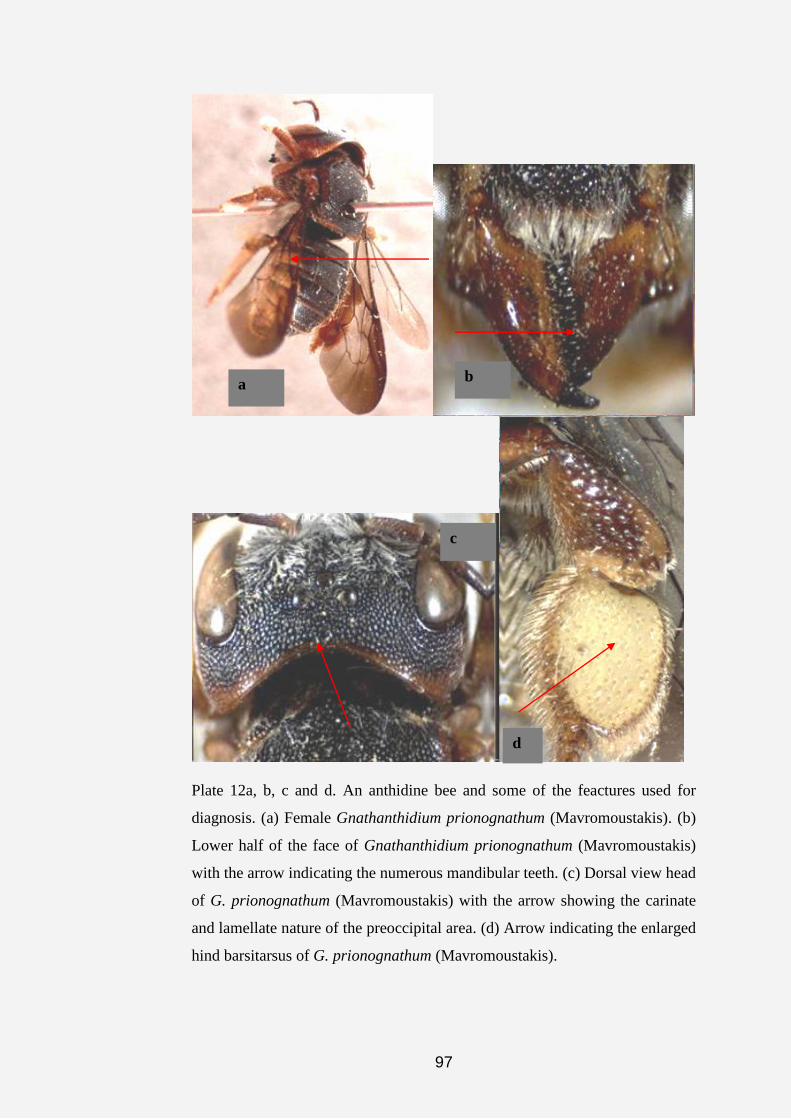
97
Plate 12a, b, c and d. An anthidine bee and some of the feactures used for
diagnosis. (a) Female Gnathanthidium prionognathum (Mavromoustakis). (b)
Lower half of the face of Gnathanthidium prionognathum (Mavromoustakis)
with the arrow indicating the numerous mandibular teeth. (c) Dorsal view head
of G. prionognathum (Mavromoustakis) with the arrow showing the carinate
and lamellate nature of the preoccipital area. (d) Arrow indicating the enlarged
hind barsitarsus of G. prionognathum (Mavromoustakis).
b
d
c
a

98
Genus Hoplostelis Dominique
Subgenus Austrostelis Michener and Griswold
Hoplostelis (Austrostelis) Michener and Griswold, 1994: 676. Type species:
Stelis aliena Cockerell, 1919, by original designation; Michener, 2000:
500.
Subgenus Hoplostelis Dominique s. str.
Hoplostelis Dominique, 1898: 60. Type species: Stelis abnormis Friese, 1925,
designated by Griswold and Michener, 1988: 36.
Odontostelis Cockerell, 1931c: 542. Type species: Stelis abnormis Friese,
1925. Isogenotypic with Hoplostelis.
Hoplostelis (Hoplostelis) Dominique: Griswold and Michener, 1988: 36;
Michener, 2000: 500-501.
Subgenus Rhynostelis Moure and Urban
Rhynostelis Moure and Urban, 1994: 297. Type species: Anthidium
multiplicatum Smith, 1879, by original designation.
Hoplostelis (Rhynostelis) Urban: Michener, 2000: 501.
Comment
This cleptoparasitic genus was first described by Dominique (1898).
Other authors, such as Griswold and Michener (1988), Moure and Urban
(1994) and Michener (2000), worked on this genus. Michener (2000) listed
three subgenera, all of which are Neotropical. This genus consists of mostly
medium to large sized, hoplitiform to chalicodomiform bees.
b
c

99
Diagnosis
Length: head 1.5-1.8 mm; scutum and scutellum 1.8-2.3 mm; forewing
2.9-5.2 mm; body 4.8-7.1 mm. Vestiture: white, sparse to impubescent; female
lacks scopa; male sternal comb on S3. Integument: black, except for limited
yellow to reddish maculation on preoccipital area, legs, lateral regions of
female T1-T5, male T1-T6; female mandible usually with three dissimilar
teeth; lower clypeal margin with four small projections; subantennal suture
straight; juxtantennal carina well developed; preoccipital area slightly carinate
dorsally; scuto-scutellar suture open, forming shiny fovea; scuto-axilla suture
closed; scutellum, in dorsal view, truncate posteriorly; arolia present; S5-S6
with lateral teeth; male T7 uniquely bilobed; gonostylus slender apically; penis
valves widely seperated (Figure 13a).
GENUS Hypathidiodes MOURE
Subgenus Anthidulum Michener
Dianthidium (Anthidulum) Michener, 1948: 19. Type species: Dianthidium
currani Schwarz, 1933, by original designation.
Hypanthidiodes (Anthidulum) Michener: Griswold and Michener, 1988: 31;
Michener, 2000: 502.
Subgenus Ctenanthidium Urban
Ctenanthidium Urban, 1993a: 85. Type species: Ctenanathidium gracile
Urban, 1991, by original designation.
Hypanthidiodes (Ctenanthidium) Urban: Michener, 2000: 502.

100
Subgenus Dichanthidium Moure
Dichanthidium Moure, 1947a: 30; [also described as new later that year by
Moure, 1947b: 235]. Type species: Dichanthidium exile Moure, by
original designation.
Hypanthidiodes (Dichanthidium) Moure: Michener, 2000: 502.
Subgenus Dicranthidium Moure
Dicranthidium Moure and Urban, 1975: 837. Type species: Anthidium
arenarium Ducke, 1907, by original designation.
Hypanthidiodes (Dicranthidium) Moure and Urban: Michener, 2000: 502.
Subgenus Hypanthidiodes Moure s. str.
Hypanthidiodes Moure, 1947a: 35. Type species: Anthidium flavofasciatum
Schrottky, 1902, by original designation.
Hypanthidioides [!] Moure: Michener, 1948: 22; Moure and Urban, 1975: 837;
Urban, 1993b: 28.
Hypanthidiodes (Hypanthidiodes) Moure: Michener, 2000: 502-503.
Subgenus Larocanthidium Urban
Larocanthidium Urban, 1997a: 299. Type species: Larocanthidium
emarginatum Urban, 1997, by original designation.
Hypanthidiodes (Larocanthidium) Urban: Michener, 2000: 503.
Subgenus Michanthidium Urban
Gnathanthidium Urban, 1994a: 337 (not Pasteels, 1969). Type species:
Gnathanthidium sakagamii Urban, 1994a, by original designation.
Michanthidium Urban, 1994a: 281. Replacement for Gnathanthidium Urban,
1994a.
Hypanthidiodes (Michanthidium) Urban: Michener, 2000: 503.

101
Subgenus Mielkeanthidium Urban
Mielkeanthidium Urban, 1996: 121. Type species: Mielkeanthidium nigripes
Urban, 1996, by original designation.
Hypanthidiodes (Mielkeanthidium) Urban: Michener, 2000: 503.
Subgenus Moureanthidium Urban
Moureanthidium Urban, 1995: 37. Type species: Dianthidium subarenarium
Schwarz, 1933, by original designation.
Hypanthidiodes (Moureanthidium) Urban: Michener, 2000: 503.
Subgenus Saranthidium Moure and Hurd
Saranthidium Moure and Hurd, 1960: 6. Type species: Anthidium flavopictum
Smith, by original designation; Griswold and Michener, 1988: 31-32.
Hypanthidiodes (Saranthidium) Moure and Hurd, 1960: 6; Michener, 2000:
503-504.
Comment
This genus was first described by Moure (1947a). Michener (2000) listed
ten subgenera, all of which are Neotropic. This non-parasitic genus comprises
small to medium-sized chalicodomiform or hoplitiform bees.
Diagnosis
Length: head 1.4-2.0 mm; scutum and scutellum 1.8-2.5 mm; forewing
2.9-5.5 mm; body 4.8-7.6 mm. Vestiture: white; sparse dense to impubescent;
female scopa on S2-S5; male sternal comb on S3, S4, or S5. Integument:
black, yellow maculations on most of lower half of face, sometimes a spot on
dorsal

102
region of preoccipital area; anterior margin of scutum, posterior margin of
scutellum, extensively on legs and metasoma; female mandible with four
dissimilar teeth; lower clypeal margin with four reduced projections;
subantennal suture straight; juxtantennal carina well developed; preoccipital
area gently rounded; scuto-scutellar suture sightly, forming reduced fovea;
scuto-axilla suture closed; scutellum, in dorsal view, rounded or lamellate
posteriorly; arolia absent, except in subgenera Ctenanthidium and
Dichanthidium; metasomal spines absent; gonostylus of male very slender,
lacks notch on margin; penis valves slender, widely separated.
Genus Hypanthidium Cockerell
Subgenus Hypanthidium Cockerell
Hypanthidium Cockerell, 1904b: 292. Type species: Anthidium
flavomarginatum Smith, 1879, by original designation.
Hypanthdium (Hypanthidium) Cockerell: Michener, 2000: 504.
Subgenus Tylanthidium Urban
Tylanthidium Urban, 1994a: 277. Type species: Tylanthidium tuberigaster
Urban, 1994, by original designation.
Hypanthidium (Tylanthidium) Urban: Michener, 2000: 504.
Comment
The genus Hypanthidium was first described by Cockerell (1904b). Later,
Urban (1994a) revised this genus. Michener (2000) listed two subgenera, all
of which are endemic to the Neotropical Region. It consists of large
chalicodomiform bees.

103
Diagnosis
Length: head 2.9-3.2 mm; scutum and scutellum 3.2-3.5 mm; forewing
4.2-7.7 mm; body 9.7-12.9 mm. Vestiture: white to brownish; moderate to
sparse; female scopa present on S2-S5; male sternal comb absent. Integument:
mostly black, legs yellow to brownish; yellow maculations on most of lower
half of face; dorsal portion of preoccipital ridge; lateral margin of scutum,
axilla, posterolateral half of scutellum; most of anteromedial half of female
T1-T5, and male T6 (mediolongitudinal margin black (Plate 13a)); female
mandible with four blunt teeth, male mandible bidentate; lower clypeal margin
without projection; subantennal suture straight; juxtantennal carina absent;
preoccipital area gently rounded; scuto-scutellar suture closed, similar to
scuto-axilla suture; scutellum, in dorsal view, subtruncate posteriorly; arolia
present; T6 with lateral spines, female; male T7 trifid (Plate 14b); male
gonostylus slight expanded apically; penis valves with apex deeply notched,
widely separated (Figure 13b).
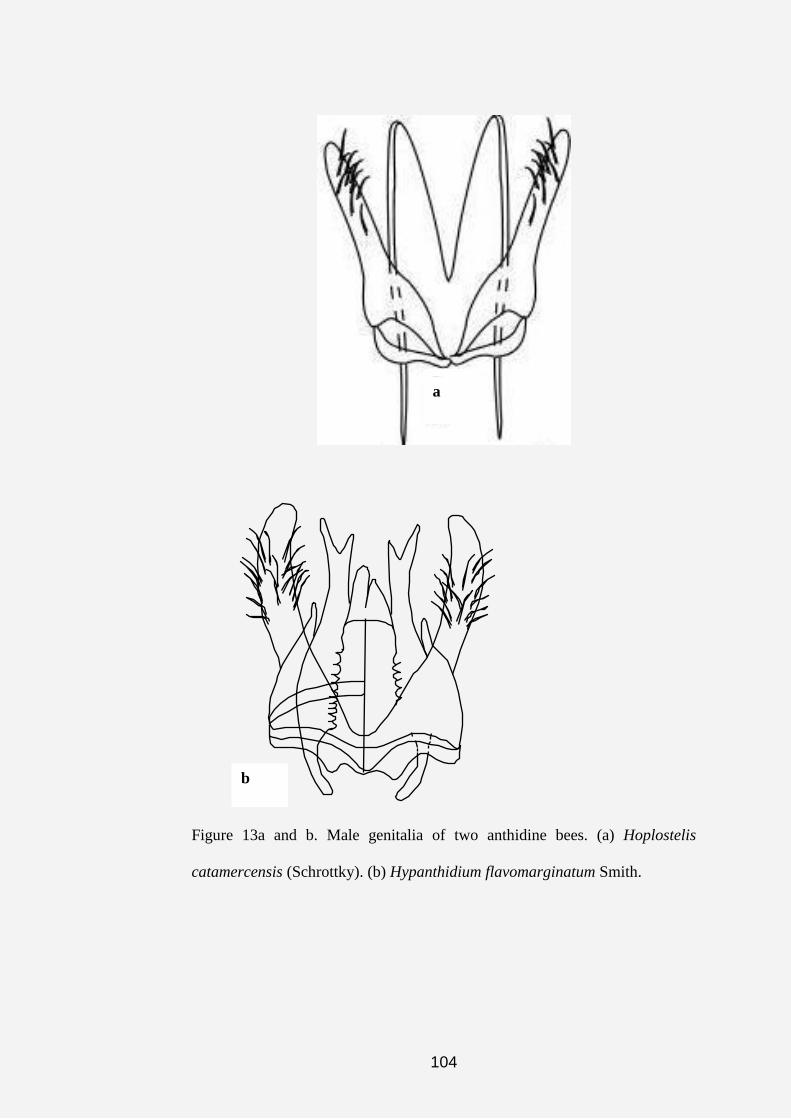
104
Hypanthidium
Figure 13a and b. Male genitalia of two anthidine bees. (a) Hoplostelis
catamercensis (Schrottky). (b) Hypanthidium flavomarginatum Smith.
Fig.
25
b
a

105
Plate 13a, b and c. Three species of the tribe Anthidiini. (a) Hypanthidium
flavomarginatum Smith. (b) Female Indanthidium crenulaticauda Michener
and Griswold. (c) Female Notanthidium steloides (Spinola).
Plate 14a, b and c. Three diagnostic features used for diagnosis of the genera.
(a) Arrow showing the lateral margin of the axilla of Icteranthidium laterale
Latreille extending beyond scutum margin. (b) Arrows showing the trifid
nature of tergite 7 of the male of Hypanthidium flavomarginatum Smith. (c)
Arrows indicating the quadrifid nature of tergite 7 of the male of Neanthidium
octodentatum (Pérez).
a
b
c
c
b a

106
Genus Icteranthidium Latreille
Icteranthidium Michener, 1948: 25. Type species: Anthidium limbiferum
Morawitz, 1875, by original designation; Pasteels 1969: 67-69; 1984:
136, 156; Michener, 2000: 504.
Comment
This genus was first described by Latrielle (1809). Later, other authors,
such as Michener (1948) and Pasteels (1969, 1984) worked on this genus.
According to Michener (2000), there are about 25 species, 15 species occur in
the West Palaearctic Region, two are endemic to the Afrotropical Region and
the other eight, are Oriental.
Diagnosis
Length: head 2.9-3.1 mm; scutum and scutellum 2.9-3.5 mm; forewing
7.4-9.9 mm; body 11.0-12.0 mm. Vestiture: white; pubescence moderate to
sparsely dense; female scopa on S2-S6; male sternal comb absent; thick setae
on tibia absent. Integument: black, except yellow maculations on lower half of
face (female mandible black), lateral region of preoccipital area, most of
dorsal surfaces of tegula and legs, lateral regions of T1-T6 (narrow on T1-T2);
mandible of female with four teeth; lower clypeal margin with 5 projections (1
medial, 2 lateral); subantennal suture more or less straight; juxtantennal carina
absent; preoccipital area lacks carina; scuto-scutellar suture closed; posterior
margin of axilla with strong spine (Plate 14a); scutellum, in dorsal view,
subtruncate posteriorly; arolia absent; T6 and T7 with reduced spines; T7
trifid.

107
Genus Indanthidium Michener and Griswold
Indanthidium Michener and Griswold, 1994: 315. Type species: Indanthidium
crenulaticauda Michener and Griswold, 1994, by original designation;
Michener, 2000: 504-505.
Comment
Genus Indanthidium was first described by Michener and Griswold
(1994). This monotypic genus consists of small to medium-sized,
megachiliform, pollen-collecting bees that are endemic to southern India.
Diagnosis
Length: head 2.0-2.2 mm; scutum and scutellum 1.8-2.0 mm; forewing
3.9-5.0 mm; body 5.1-7.1 mm. Vestiture: white; sparse to impubescent; female
scopa on S2-S6; male sternal comb absent; thick setae on tibia absent.
Integument: black, except yellow maculations on lower half of face (mandible
black), spot on dorsal region of preoccipital area, anterior margin of scutum,
dorso-lateral regions of axilla and scutellum, lateral and medial third regions
od T1-T5, medial region of T6 (Plate 13b); female mandible with seven teeth;
lower clypeal margin without projection; subantennal suture straight;
juxtantennal carina absent; preoccipital area with lateral carina; scuto-scutellar
suture closed; scutellum, in dorsal view, emarginate medio-posteriorly; arolia
present; hind tibial spur straight apically; T7 of male trifid and uniquely broad,
subequal T6; gonostylus slender apically; penis valves narrow apically, widely
separated (Figure 14a).
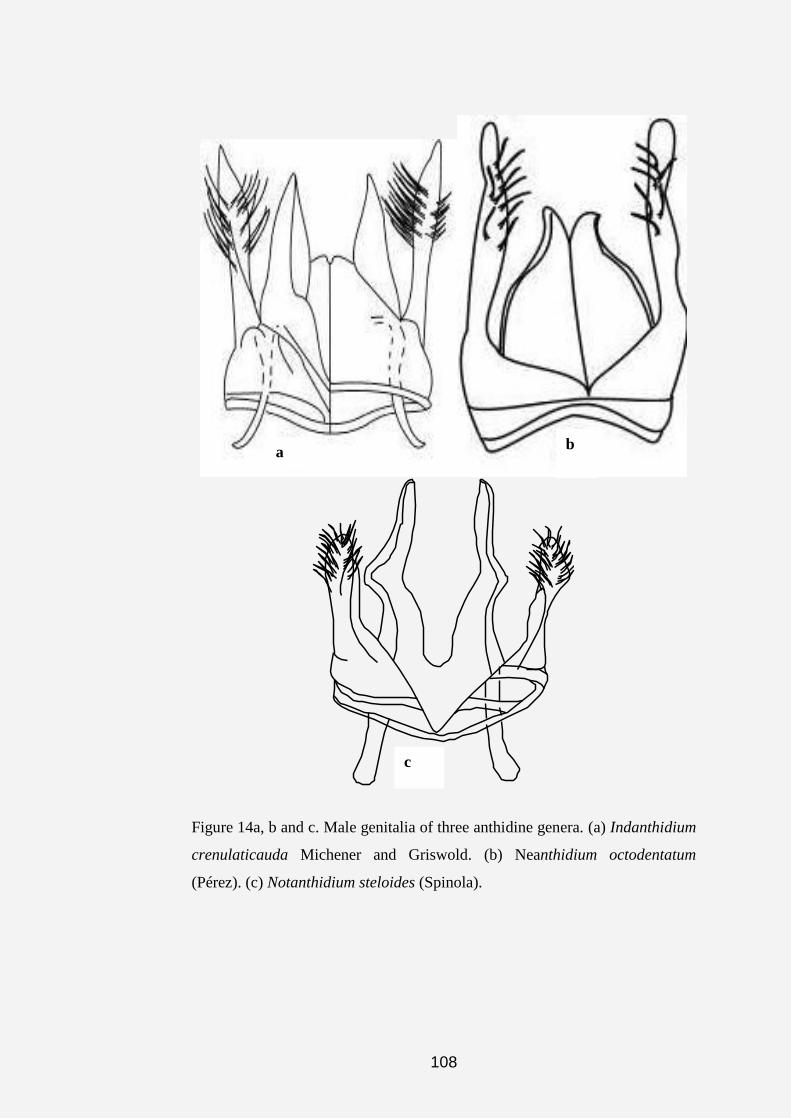
108
Notanthidium
Figure 14a, b and c. Male genitalia of three anthidine genera. (a) Indanthidium
crenulaticauda Michener and Griswold. (b) Neanthidium octodentatum
(Pérez). (c) Notanthidium steloides (Spinola).
a b
c

109
Genus Larinostelis Michener and Griswold
Larinostelis scapulata Michener and Griswold, 1994: 317. Type species:
Larinostelis scapulata Michener and Griswold, 1994, by original
designation; Michener 1997: 32; Michener 2000: 505.
Comment
This genus was first described by Michener and Griswold (1994). It is
represented by only one described species Larinostelis scapulata Michener
and Griswold that is endemic to Kenya. This genus was not available for
examination during this study, due to the fact that, it is a rare specimen and the
museum was unwilling to loan then out.
Genus Neanthidium Pasteels
Neanthidium Pasteels, 1969: 93. Type species: Anthidium octodentatum Pérez,
1895, by original designation; Michener, 2000: 505.
Comment
The genus Neanthidium was first described by Pasteels (1969). It is
represented by one species Neanthidium octodentatum (Pasteels), which
occurs in North Africa (Algeria, Morocco).
Diagnosis
Length: head 2.6-2.9 mm; scutum and scutellum 2.3-2.9 mm; forewing 4.8-7.1
mm; body 7.1-10.6 mm. Vestiture: white to yellow; moderate to sparse;
female scopa on S2-S5; male sternal comb on S4 and S5. Integument: black,

110
yellow or reddish-yellow maculation on most of lower half of face, dorsal
margin of preoccipital area, most of tegula, spots on dorsal surfaces of axilla
and scutellum, lateral regions of T1-T5 of female, T1-T2 of male; mandible of
female with five dissimilar teeth; lower clypeal margin with 3-5 projections, a
median and one-two lateral; subantennal suture straight; juxtantennal carina
absent; preoccipital area gently rounded; scuto-scutellar and scuto-axilla
suture closed; scutellum, in dorsal view, rounded posteriorly; arolia absent;
lateral region of T6 of female with small metasomal spines, T6 and T7 of male
with strong spines on posterior regions, multidentated (Plate 14c); S7 of male
trifid; gonostylus of male slender apically; penis valves narrows at apex,
narrowly separated (Figure 14b).
Genus Notanthidium (Isensee)
Subgenus Allanthidium Moure
Allanthidium Moure, 1947a: 21. Type species: Anthidium rodolfi Ruiz, 1938,
by original desination.
Trichanthidium Moure, 1947b: 209 (not Cockerell, 1930). Type species:
Anthidium subpetiolatum Schrottky, 1910, by original designation,
Moure, 1947a: 21, syn.
Notanthidium (Allanthidium) Moure: Michener, 2000: 505-506.
Subgenus Chrisanthidium Urban
Chrisanthidium Urban, 1997b: 181. Type species: Anthidium bidentatum
Friese, 1908, by original designation.
Notanthidium (Chrisanthidium) Moure: Michener, 2000: 506.

111
Subgenus Notanthidium Isensee s. str.
Dianthidium (Notanthidium) Isensee, 1927: 373. Type species: Anthidium
steloides Spinola, 1851, by original designation.
Notanthidium (Notanthidium) Moure: Michener, 2000: 506.
Comment
This slender, hoplitiform genus of pollen-collecting bees was first
described by Isensee (1927). Later, Moure (1947a) and Urban (1997b) worked
on the genus. Michener (2000) listed three subgenera, all of which are
endemic to South America.
Diagnosis
Length: head 1.6-2.6 mm; scutum and scutellum 2.7-3.3 mm; forewing
5.8-6.1 mm; body 7.9-10.2 mm. Vestiture: reddish brown to black; sparse
dense to impubescent; female scopa on S2-S6; male sternal comb absent.
Integument: mostly black, antenna, lateral region of preoccipital ridge, tegula,
legs reddish-brown, yellow maculations on lateral regions of T1-T3, medial
region of T4 (Plate 13c); female mandible with five teeth; lower clypeal
margin with large groove, lower margin without projection; subantennal suture
straight; juxtantennal carina absent; female with unique, distinct, antennal
scape groove, (Fig. 4); preoccipital area carinate dorsally; scutellum, in dorsal
view, rounded posteriorly; arolia present; metasomal lateral spines absent;
gonostylus slender, apically narrow; penis valve widely separated (Figure
14c).

112
Genus Pachyanthidium Vachal
Subgenus Ausanthidium Pasteels
Ausanthidium Pasteels 1969: 60, 132-133. Type species: Anthidiellum ausense
Mavromoustakis, 1934; Pasteels 1984: 130, by original designation.
Pachyanthidium (Ausanthidium) Pasteels: Michener and Griswold, 1994: 319;
Michener, 2000: 507.
Subgenus Pachyanthidium Friese s. str.
Anthidium (Pachyanthidium) Friese, 1905b: 66-67, 69; Friese, 1910: 158.
Type species: Anthidium bicolor Lepeletier, 1841, designated by
Cockerell, 1920: 298.
Pachyanthidium (Pachyanthidium): Mavromoustakis, 1963: 490; Pasteels
1968b: 482-483; Pasteels, 1984: 138-139, 141-145, 147-148, Michener
and Griswold, 1994: 319; Michener, 2000: 507.
Subgenus Trichanthidiodes Michener and Griswold
Pachyanthidium (Trichanthidiodes) Michener and Griswold, 1994: 319. Type
species: Pachyanthidium semilutum Pasteels, 1981, by original
designation; Michener 2000: 507.
Subgenus Trichanthidium Cockerell
Pachyanthidium (Trichanthidium) Cockerell, 1930b: 52. Type species:
Pachyanthidium uxtanten Cockerell, 1930, monobasic; Pasteels,
1984: 151; Michener and Griswold, 1994: 317; Michener, 2000: 507.
Comment
Genus Pachyanthidium was first described by Friese (1905). Since

113
then, other authors, such as Cockerell (1930b), Pasteels (1969) and Michener
and Griswold (1994), worked on this group. Michener (2000) recorded four
subgenera. Two are endemic to Africa, one occurs in both Africa and Arabia,
while the other occurs in both Africa and the Orient. This genus comprises
non-parasitic, robust megachiliform bees.
Diagnosis
Length: head 1.6-2.7 mm; scutum and scutellum 1.5-2.9 mm; forewing
4.3-5.8 mm; body 5.4-9.0 mm. Vestiture: white, sparse to impubescent; female
scope on S2-S6; male sternal comb on S5. Integument: mostly black, with
limited yellow maculations on most of lower half of face (Plate 16a), posterior
margin of preoccipital area, laterally on scutum, axilla and scutellum,
metasoma (Plate 15); female mandible 5 to 10 dissimilar teeth; lower clypeal
margin without projection; subantennal suture straight; suxtantennal carina
reduced; preoccipital area carinate or lamelate dorsolaterally; scuto-scutellar
and scuto-axilla suture closed; scutellum, in dorsal view, truncate (Plate 16d);
arolia absent, except in male of subgenus Trichanthidium and both sexes of
subgenus Ausanthidium; T1-T6 with lateral spines, T7 of male trifid;
gonostylus apically expanded; penis valves narrowly separated (Figure 15a).

114
Plate 15. Female of Pachyanthidium bicolor (Lepeletier).
Plate 16a, b, c and d. Four diagnostic features of two anthidine genera.
(a) Face of P. bicolor (Lepeletier), with arrows indicating the straight
nature of the subantennal suture and regions of yellow integumental
maculations. (b and c) Lower region of the face of Paranthidium
perpictum (Cockerell), arrows indicating the mandibular dentition,
projections on lower clypeal margin and subantennal suture. (d) Arrow
indicating lamella nature of the scutellum of P. bicolor (Lepeletier).
a
c
b
d
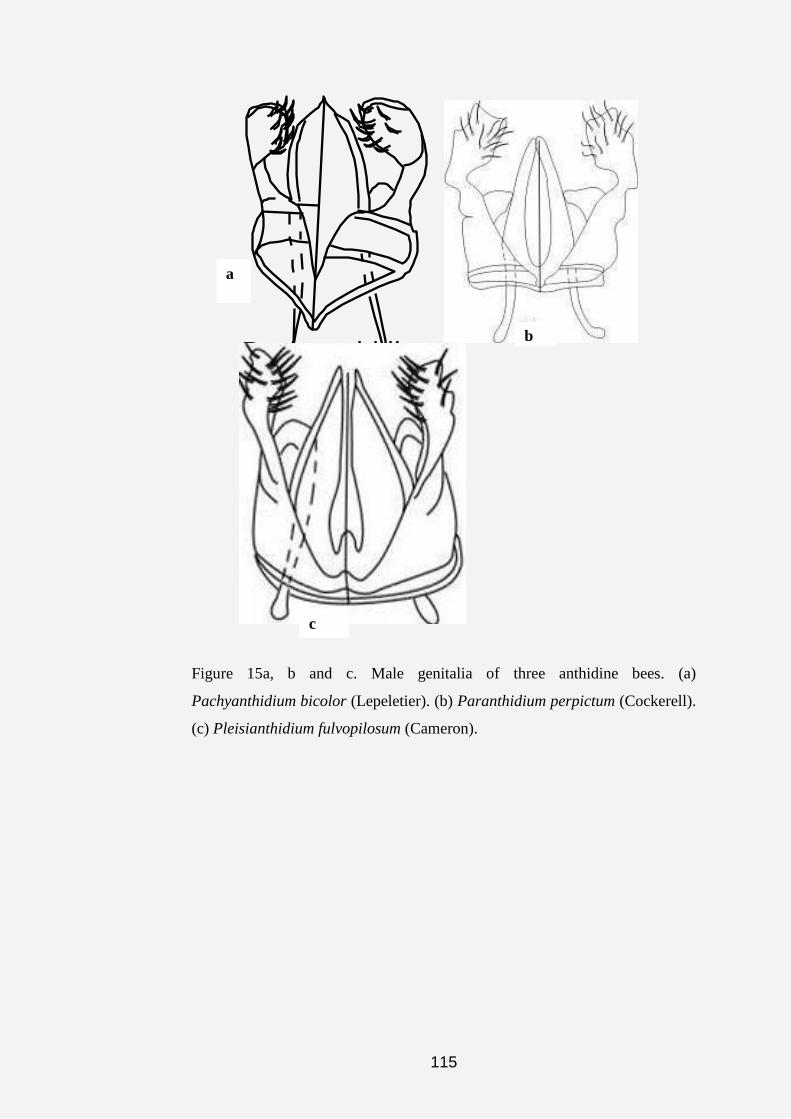
115
Pachyanthidium
Figure 15a, b and c. Male genitalia of three anthidine bees. (a)
Pachyanthidium bicolor (Lepeletier). (b) Paranthidium perpictum (Cockerell).
(c) Pleisianthidium fulvopilosum (Cameron).
a
c
b

116
Genus Paranthidium Cockerell and Cockerell
Subgenus Paranthidium Cockerell and Cockerell s. str.
Paranthidium Cockerell and Cockerell, 1901: 50. Type species: Anthidium
perpictum Cockerell, 1898, by original designation.
Paranthidium (Paranthidium) Cockerell and Cockerell: Michener, 2000: 507.
Subgenus Rapanthidium Michener
Paranthidium (Rapanthidium) Michener, 1948: 11. Type species: Anthidium
vespoides Friese, 1925, by original designation; Michener, 2000: 507.
Comment
Genus Paranthidium was first described by Cockerell and Cockerell
(1901). Michener (1948) described an extra two subgenera in this genus
(namely Rapanthidium and Mecanthidium), but later Michener (2000) moved
Mecanthidium to the genus Dianthidium. One of the subgenera is endemic to
Mesoamerica and the other occurs in both Mesoamerica and the Nearctic
Region.
Diagnosis
Length: head 2.0-2.4 mm; scutum and scutellum 2.0-2.6 mm; forewing
6.1-7.1 mm; body 7.1-9.0 mm. Vestiture: yellowish to white; sparse to
impubescent; female scopa on S2-S6; male sternal comb on S4-S5.
Integument: black, except for yellow maculation on face, dorsal margin of
preoccipital area, entire margin of scutum, axilla and scutellum, spots on
dorsal surfaces of femora and tibiae, lateral regions of T1-T3 (shorter and

117
wider on T3), most of T4-T5 (mediolongitudinal region black), entire T6-T7;
female mandible with four dissmilar teeth; lower clypeal margin with six
projections (Plate 16b and c); subantennal suture straight; juxtantennal carina
absent; preoccipital area rounded; scuto-scutellar suture closed, similar to
scuto-axilla suture; scutellum, in dorsal view, medially emarginate, obscures
propoduem; arolia present; metasomal lateral spines absent, except male T7
trifid, median spine larger than lateral spine; gonostylus of male expanded
apically, slightly notched on outer margin; penis valves reduced apically,
widely separated (Figure 15b).
Genus Pleisianthidium Cameron
Subgenus Carinanthidium Pasteels
Carinthanthidium Pasteels, 1969: 42-43. Type species: Megachile
cariniventris Friese, 1904, by original designation.
Plesianthidium (Carinthanthidium) Pasteels; Michener, 2000: 508.
Subgenus Plesianthidium Cameron s. str
Pleisianthidium Cameron, 1905: 256-257. Type species: Pleisianthidium
fulvopilosum Cameron, 1905, by original designation.
Pleisianthidium (Pleisianthidium) Cameron: Michener and Griswold, 1994:
301, 319; Michener, 2000: 508.
Subgenus Spinanthidiellum Pasteels
Spinanthidium (Spinanthidiellum) Pasteels, 1969: 55, 59-60. Type species:
Anthidium volkmanni Friese, 1909, by original designation; Pasteels,
1984: 128-129.

118
Pleisanthidium (Spinanthidiellum) Pasteels: Michener, 2000: 508.
Subgenus Spinanthidium Mavromoustakis
Dianthidium (Spinanthidium) Mavromoustakis, 1951: 977-980. Type species:
Anthidium trachusiforme Friese, 1913, by original designation.
Pleisianthidium (Spinanthidium) Mavromoustakis: Michener, 2000: 508-509.
Comment
This genus was first described by Cameron (1905). Since then, other
authors, such as Mavromoustakis (1951), Pasteels (1969, 1984) and Michener
and Griswold (1994), have revised the group. Michener (2000) listed four
subgenera, all of which are endemic to South Africa.
Diagnosis
Length: head 2.5-3.6 mm; scutum and scutellum 2.4-4.3 mm; forewing
5.4-8.2 mm; body 7.3-12.9 mm. Vestiture: white (in Spinanthidium), yellow or
reddish-brown (in nominative subgenus and Spianthidiellum), black (in
Carinanthidium); moderate to dense; female scopa on S2-S5; male S5 with
sternal comb, except in Spinanthidium. Integument: black, except brown
integumental bands in Spinanthidium (Plate 17a); female mandible with four
dissmilar teeth; lower clypeal margin without projection; subantennal suture
straight; juxtantennal carina absent; preoccipital ridge carinate dorsally; scuto-
scutellar suture closed; similar to scuto-axilla suture; scutellum usually
rounded posteriorly; arolia present; metasomal lateral spines absent; T7 of
male trifid (Plate 18c); gonostylus expanded apically; penis valves narrowly
separated (Figure 15c; Plate 18a).

119
Plate 17a and b. Two genera of the tribe Athidiini. (a) Pleisianthidium
fulvopilosum (Cameron). (b) Rhodanthidium siculum (Spinola).
Plate 18a, b and c. Some feactures used for the diagnosis of the genera. (a)
Male genitalia of P. fulvopilosum (Cameron). (b) Male genitalia of R. siculum
(Spinola). (c) Arrows indicating the trifid nature of tergite 7 of the male of R.
siculum (Spinola).
a
b
a b
c

120
Genus Pseudoanthidium Smith
Subgenus Exanthidium Pasteels
Exanthidium Pasteels, 1969: 82. Type species: Anthidium eximum Giraud,
1863, by original designation.
Pseudoanthidium (Exanthidium) Pasteels: Michener and Griswold, 1994: 320;
Michener, 2000: 509.
Subgenus Micranthidium Cockerell
Pachyanthidium (Micranthidium) Cockerell, 1930b: 45-46, 48. Type species:
Anthidium truncatum Smith, 1854, by original designation.
Pseudoanthidium (Micranthidium) Cockerell: Michener, 2000: 510.
Subgenus Pseudoanthidium Friese s. str.
Anthidium (Pseudoanthidium) Friese, 1898: 101. Type species: Anthidium
alpinum Morawitz, 1873, designated by Sandhouse, 1943: 593.
Pseudoanthidium (Pseudoanthidium) Pasteels: Michener and Griswold, 1994;
Michener, 2000: 510.
Paranthidiellum Michener, 1948: 25. Type species; Anthidium cribratum
Morawitz, 1875, by original designation; Michener and Griswold, 1994:
320, syn.
Pseudoanthidium (Paraanthidiellum) [!] Michener: Pasteels, 1969: 79.
Psuedoanthidium (Carinellum) Pasteels, 1969: 80. Type species: Anthidium
ochrognathum Alfken, 1932, by original designation; Pasteels, 1969: 80,
syn.
Trachusa (Orientotrachusa) Gupta, 1993: 50. Type species: Anthidium
orientale Bingham, 1897, by original designation; Michener, 2000: 510,
syn.

121
Subgenus Royanthidium Pasteels
Royanthidium Pasteels, 1969: 86. Type species: Anthidium melanurum Klug,
1873, by original designation.
Reanthidium Pasteels, 1969: 87. Type species: Anthidium reticulatum
Mocsáry, 1884, by original designation; Michener and Griswold, 1994:
320, syn.
Pseudoanthidium (Royanthidium) Pasteels: Michener and Griswold, 1994:
320; Michener, 2000: 510.
Subgenus: Semicarinella Pasteels
Pseudoanthidium (Semicarinella) Pasteels, 1984: 32. Type species:
Pseudoanthidium lattarse Pasteels, by original designation; Michener and
Griswold, 1994: 320; Michener, 2000: 510.
Subgenus Tuberanthidium Pasteels
Tuberanthidium Pasteels, 1969: 87-88. Type species: Anthidium
tuberculiferum Brauns, 1905, by original designation; Pasteels, 1984:
60-63; Gess, 1996: 290.
Pseudoanthidium (Tuberanthidium) Pasteels: Michener and Griswold, 1994:
320; Michener, 2000: 510-511.
Comment
Genus Pseudoanthidium was first described by Friese (1898). Michener
(2000) recorded six subgenera, two of which are endemic to Africa, three are
known from the Palaearctic and Orient regions. The other subgenus occurs in
both Africa and Arabia.

122
Diagnosis
Length: head 2.1-3.4 mm; scutum and scutellum 1.5-3.2 mm; forewing
4.2-7.1 mm; body 5.2-9.4 mm. Vestiture: white to reddish-black; sparse to
impubescent; female scopa on S2-S6; male sternal comb on S3. Integument:
black, with extensive yellow or reddish maculations on most of lower half of
face, lateral regions of T1-T5 (yellow areas increase progressively from front
to back), T6; Tuberanthidium mostly black; mandible of female with 5 or
more teeth; lower clypeal margin without projection; subantennal suture
arcuate; juxtantennal carina absent; preoccipital area carinate laterally; scuto-
scutellar suture slightly open, forming shiny fovae; scuto-axilla suture closed;
scutellum, in dorsal view, truncate posteriorly; arolia absent; metasomal lateral
spines absent; gonostylus of male slightly expanded apically; penis valve
narrow apically (Figure 16a).
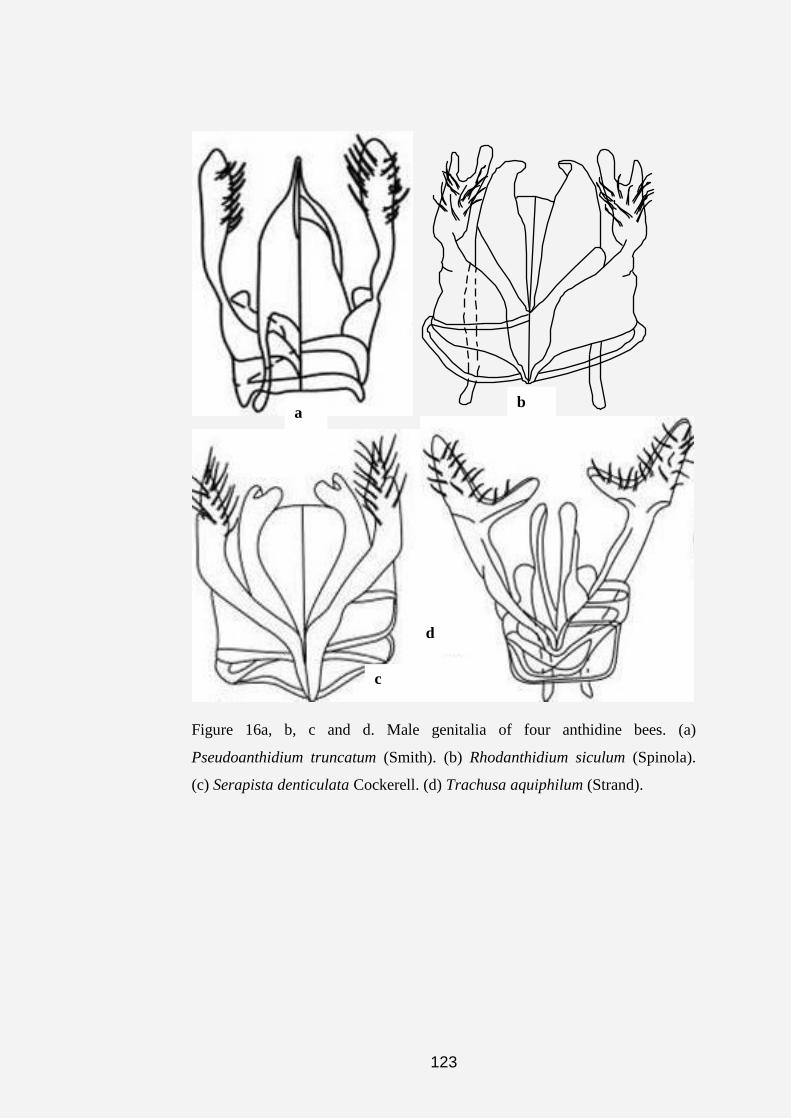
123
Rhodanthidium
Figure 16a, b, c and d. Male genitalia of four anthidine bees. (a)
Pseudoanthidium truncatum (Smith). (b) Rhodanthidium siculum (Spinola).
(c) Serapista denticulata Cockerell. (d) Trachusa aquiphilum (Strand).
b a
c
d

124
Genus Rhodanthidium Spinola
Subgenus Asianthidium Popov
Meganthidium (Asianthidium) Popov, 1950: 315. Type species: Anthidium
glasunovii Morawitz, 1894, by original designation.
Rhodanthidium (Asianthidium) Popov: Michener and Griswold, 1994: 322;
Michener, 2000: 511.
Rhodanthidium (Trianthidium) Mavromoustakis, 1958: 435. Type species:
Anthidium caturigense Giraud, 1863, by original designation;
Warncke, 1980: 119-209, syn.
Meganthidium (Oxyanthidium) Mavromoustakis, 1963a: 653. Type species:
Anthidium aculeatum Klug, 1832, by original designation; Warncke,
1980: 119-209, syn.
Axillanthidium Pasteels, 1969: 39. Type species: Axillanthidium axillare
Pasteels, 1969, by original designation; Warncke, 1980: 119-209, syn.
Subgenus Meganthidium Popov
Meganthidium Popov, 1950: 315. Type species: Anthidium christophi
Morawitz, 1876, by original designation; Pasteels, 1969: 31.
Rhodanthidium (Meganthidium) Popov: Michener and Griswold, 1994: 322;
Michener, 2000: 511-512.
Subgenus Rhodanthidium Isensee s. str.
Anthidium (Rhodanthidium) Isensee, 1927; 374. Type species: Anthidium
siculum Spinola, 1838, by original designation.
Bellanthidium Pasteels, 1969: 38. Type species: Anthidium infuscatum
Erichson, 1835, by original designation; Michener and Griswold, 1994:
322, syn.

125
Rhodanthidium (Rhodanthidium) Isensee: Michener and Griswold, 1994: 322;
Michener, 2000: 512.
Comment
The genus Rhodanthidium was first described by Iseense (1927).
Michener (2000) recognized three subgenera, all of which occur in the
Palaearctic Region, however, one also occurs in the Oriental Region.
Diagnosis
Length: head 2.9-3.4 mm; scutum and scutellum 3.9-5.0 mm; forewing
9.2-11.2 mm; body 11.2-17.1 mm. Vestiture: white to reddish-brown, sparse to
moderate, very dense on legs; female scopa on S2-S5; male S4-S5 with medial
sternal comb. Integument: mostly black, (Meganthidium mostly yellow) with
extensive yellow to reddish brown maculations on face and metasoma (Pl.
12b); mandible of female with four dissimilar, large teeth; lower clypeal
margin without projection; subantennal suture more or less straight;
juxtantennal carina absent; preoccipital area gently rounded; scuto-scutellar
suture closed, similar to scuto-axilla suture; scutellum, in dorsal view, rounded
posteriorly; arolia present; metesoma with reduced lateral spines; T7 of male
trifid (Pl. 12e); male gonostylus slender with outer margin deeply notched;
penis vales narrowly seperated, with their apices curved inward (Figure 16b;
Pl. 12d).

126
Genus Serapista Cockerell
Serapis Smith, 1854: 218-219 (not Link, 1839). Type species: Serapis
denticulatus Smith, 1854, monobasic.
Serapista Cockerell 1904b: 357. Replacement for Serapis Smith, 1854;
Cockerell 1932: 360; Cockerell 1933c: 357; Cockerell 1935: 237;
Mavromoustakis, 1940a: 65; Pasteels 1969: 107; Pasteels 1984: 38;
Michener, 2000: 512.
Comment
Serapista--This robust, megachiliform to chalicodomiform, pollen-
collecting genus was first described by Smith (1854) as Serapis. However,
because Serapis was preoccupied under phyllum mollusca, Cockerell (1904)
renamed this genus as Serapista Cockerell. Other authors, such as
Mavromoustakis (1940) and Pasteels (1969, 1984), worked on this genus.
Pasteels (1984) recorded four species all of which are endemic to Africa.
Diagnosis
Length: head 2.6-2.9 mm; scutum and scutellum 3.1-3.9 mm; forewing
6.7-8.1 mm; body 9.8-11.5 mm. Vestiture: mostly white with unique lateral
bands on T1-T6 (increasing in size pregressively from T1 to T5, reduced on
T6, Plate 19a); brownish on legs; sparse to impubescent; female scopa on S2-
S5; male sternal comb absent. Integument: entirely black, female mandible
with eleven dissimilar teeth; lower clypeal margin with six projections;
subantennal suture straight; juxtantennal carina well developed;
mediolongitudinal region of face

127
strongly crinate; preoccipital area gently rounded; scuto-scutellar suture
closed, similar to scuto-axilla suture; posterior region of axilla with strong
spine (Plate 20a); scutellum, in dorsal view, truncate posteriorly; arolia absent;
female T1-T6, male T1-T6 with lateral spines (Plate 19a); T7 of male trifid
(Plate 20b); gonostylus narrows anteriorly; penis valve arcuate in profile,
notched anteriorly, widely seperated (Figure 16c).

128
Plate 19a and b. Two anthidine bees. (a) Arrow indicating the lateral spines on
the metasoma of Serapista denticulata Cockerell. (b) Female of Trachusa
aquiphilum (Strand).
Plate 20a and b. Two features used for diagnosis of Serapista denticulata
Cockerell. (a) Arrows indicating lateral extension of axilla. (b) Arrows
showing the trifid nature of tergite 7 of the male.
a
b
d c

129
Genus Stelis Panzer
Subgenus Dolichostelis Parker and Bohart
Dolichostelis Parker and Bohart, 1979: 138. Type species: Stelis laticincta,
Cresson, 1878, by original designation.
Stelis (Dolichostelis) Parker and Bohart: Michener, 2000: 513.
Subgenus Malanthidium Pasteels
Malanthidium Pasteel, 1969: 26. Type species: Anthidium malaccense Friese,
1914, by original designation.
Stelis (Malanthidium) Pasteels: Michener and Griswold, 1994: 322-323;
Michener, 2000: 514.
Subgenus Protostelis Friese
Protostelis Friese, 1895: 25. Type species: Stelis: freygessneri Friese, 1885, by
designation of Popov, 1938.
Stelis (Protostelis) Friese: Michener, 1944: 258-259, syn; Michener, 1948: 4;
Michener and Griswold, 1994: 323; Michener, 2000: 514-515.
Heterostelis Timberlake, 1941: 125. Type species: Stelis anthidiodes
Timberlake, 1941, by original designation; Griswold and Michener,
1988: 37, syn.
Doxanthidium Pasteels, 1969: 28. Type species: Anthidium paradoxum
Mocsáry, 1884, by original designation; Pasteels, 1969: 28, syn
Subgenus Pseudostelis Popov
Pseudostelis Popov, 1956: 167. Type species: Stelis strandi Popov, 1935, by
original designation.
Stelis (Pseudostelis) Popov: Michener and Griswold, 1994: 322; Michener,
2000: 515.

130
Subgenus Stelidomorpha Morawitz
Stelidomorpha Morawitz, 1875: 131. Type species: Anthidium nasutum
Latreille, 1809, monobasic.
Stelis (Stelidomorpha) Morawitz: Michener and Griswold, 1994: 322;
Michener, 2000: 515.
Subgenus Stelis Panzer s. str.
Trachusa Jurine, 1801: 164 (not Panzer 1801). Type species: Apis aterrima
Panzer, 1798, designated by Morice and Durrant, 1915: 426.
Suppressed by commission Opinion 135, 1939 (Direction 4).
Stelis Panzer, 1806: 264. Type species: Apis aterrima Panzer, 1798 (not
Christ, 1791), monobasic. Isogenotypic with Trachusa Jurine.
Stelis (Stelis) Panzer: Michener and Griswold, 1994: 322; Michener, 2000:
515-516.
Gyrodroma Klug, 1807: 198. Type species: Apis aterrima Panzer, 1798
[Sandhouse incorrectly considered Gyrodroma to be monobasic,
however, two species were listed by Klug, 1807: 225]. Isogenotypic
with Stelis.
Gymnus Spinola, 1808: 9. Type species: Apis aterrima Panzer, 1798.
Isogenotypic with Stelis.
Ceraplastes Gistel, 1848: x, unjustified replacement name for Stelis Panzer,
1806. Type species: Apis aterrima Panzer, 1798. Isogenotypic with
Stelis.
Chelynia Provancher, 1888: 296, 322. Type species: Chelynia labiata
Porvancher, 1888, monobasic; Michener, 1944: 258, syn; Michener,

131
1997: 15.
Melanostelis Ashmead, 1898: 283. Type species: Melanostelis betheli
Ashmead, 1898, by original designation; Michener, 1948: 4, syn.
Stelidium Robertson, 1902: 323. Type species: Stelidium trypetinum
Robertson, 1902, monobasic; Robertson, 1902: 323, syn.
Microstelis Robertson, 1903: 170, 175. Type species: Stelis lateralis Cresson,
1864, by original designation; Michener, 1944, syn; Michener 1948:
4; Michener 2000: 515.
Stelis (Pavostelis) Sladen, 1916: 313. Type species: Stelis montana Cresson,
1864, monobasic; Michener, 1944: 258, syn; Michener, 1948: 4.
Stelis (Stelidina) Timberlake, 1941: 131. Type species: Stelis hemirhoda
Linsley, 1939, by original designation; Michener, 1944: 258, syn;
Michener, 1948:4.
Stelis (Stelidiella) [!] Timberlake: Michener, 1944: 258-259.
Stelis (Stelidium) Robertson, 1902: 323. Type species: Stelidium trypetinum
Robertson, 1902, by original designation; Michener, 1944: 258-259,
syn; Michener, 1948: 4.
Stelis (Leucostelis) Noskiewicz, 1961: 126, 132. Type species: Gyrodroma
ornatula Klug, 1807, by original designation; Noskiewicz: 1961:
132, syn.
Comment
This cleptoparasitic genus was first described by Panzer (1806). Since
then several other authors, such as Provancher (1888), Ashmead (1898),

132
Robertson (1902), Noskiewicz (1961) and Michener and Griswold (1994),
revised this genus. Michener (2000) listed six subgenera. Two subgenera are
endemic to the Orient, two occur in Mesoamerica, with one of these also
occurs in the Palaearctic Region, which has an additional endemic genus. The
last subgenus is Holarctic. Michener (2000), however, mentions that its
distribution extends into Kenya, yet Eardley and Urban (in preparation) do not
record a Kenyan species but did record one species from South Africa. This
genus was not available for examination during this study, due to the fact that,
it is a rare specimen and the museum was unwilling to loan them out.
Genus Trachusa Panzer
Subgenus Archianthidium Mavromoustakis
Archianthidium Mavromoustakis, 1939: 91. Type species: Anthidium laticeps
Morawitz, 1873, by original designation.
Trachusa (Archianthidium) Mavromoustakis: Michener and Griswold, 1994:
323; Michener, 2000: 517.
Subgenus Congotrachusa Pasteels
Trachusa (Congotrachusa) Pasteels, 1969: 24. Type species: Anthidium
schoutedeni Vachal, 1901, by original designation; Pasteels, 1984:
117; Michener and Griswold, 1994: 323; Michener, 2000: 517.
Subgenus Hetereanthidium Cockerell
Anthidium (Heteranthidium) Cockerell, 1904b: 292. Type species: Anthidium
dorsale Lepeletier, 1841, by original designation.
Trachusa (Heteranthdium) Cockerell: Griswold and Michener, 1988: 27-28;

133
Michener, 2000: 517-518.
Subgenus Legnanthidium Griswold and Michener
Trachusa (Legnanthidium) Griswold and Michener, 1988: 27. Type species:
Anthidium ridingsii Cresson, 1878, by original designation; Michener,
2000: 518.
Subgenus Massanthidium Pasteels
Trachusa (Massanthiidium) Pasteels, 1969: 24. Type species: Trachusa
flavorufula Pasteels, 1969, by original designation; Pasteels, 1984:
118; Michener and Griswold, 1994: 323; Michener, 2000: .518.
Subgenus Metatrachusa Pasteels
Trachusa (Metatrachusa) Pasteels, 1969: 22. Type species: Anthidium
pendleburyi Cockerell, 1927, by original designation; Michener and
Griswold, 1994: 323; Michener, 2000: 518.
Subgenus Orthanthidium Mavromoustakis
Paraanthidium (Orthanthidium) Mavromoustakis, 1953: 837. Type species:
Anthidium formosanum Friese, 1917, by original designation.
Trachusa (Orthanthidium) Mavromoustakis: Pasteels, 1969: 23; Michener and
Griswold, 1994: 323; Michener, 2000: 518.
Subgenus Paraanthidium Friese
Anthidium (Paraanthidium) Friese, 1898: 101. Type species: Apis interrupta
Fabricius, 1781, designated by Cockerell, 1909: 269.
Trachusa (Paraanthidium) Friese: Pasteels, 1984: 114-115; Michener and

134
Griswold, 1994: 323; Michener, 2000: 518-519.
Protanthidium Cockerell and Cockerell, 1901: 49. Type species: Megachile
steloides Bingham, 1896, by original designation; Mavromoustakis,
1939: 90, syn.; Michener, 1948: 23, 26.
Trachusa (Philotrachusa) Pasteels, 1969: 22. Type species: Anthidium
aquilifilum Strand, 1912, by original designation; Pasteels: 1969: 22,
syn.
Subgenus Trachusa Panzers. Str.
Trachusa Panzer, 1804: 14-15. Type species: Trachusa serratulae Panzer,
1804, designated by Sandhouse, 1943: 605.
Trachusa (Trachusa) Panzer: Pasteels, 1969: 21; Michener and Griswold,
1994: 325; Michener, 2000: 519.
Diphysis Lepeletier, 1841: 307. Type species: Diphysis pyrenaica Lepeletier,
1841 (= Trachusa serratulae). Isogenotypic with Trachusa.
Megachileoides Radoszkowski, 1874: 132. Type species: Trachusa serratulae
Panzer, 1804, designated by Michener, 1995: 375; Isogenotypic with
Trachusa.
Megachiloides [!] Radoszkowski: Saussure, 1890: 35; Michener, 2000: 519.
Subgenus Trachusomimus Popov
Trachusomimus Popov, 1964: 406. Type species: Trachusa perdita Cockerell,
1904, by original designation.
Trachusa (Trachusomimus) Popov: Griswold and Michener, 1988: 25;
Michener, 2000: 519.

135
Subgenus Ulanthidium Michener
Ulanthidium Michener, 1948: 13. Type species: Ulanthidium mitchelli
Michener, 1948, by original designation.
Ulanthidium (Olmecanthidium) Peters, 1972b: 377. Type species:
Ulanthidium interdisciplinaris Peters, 1972, by original designation;
Peter, 1972: 13, syn.
Trachusa (Ulanthidium) Michener: Griswold and Michener, 1988: 26;
Michener, 2000: 519.
Comment
This non-parasitic genus was first described by Panzer in 1806. Since
then several other authors, such as Provancher (1888), Ashmead (1898),
Robertson (1902), Noskiewicz (1961) and Michener and Griswold (1994),
revised this genus. Michener (2000) listed six subgenera. Two subgenera are
endemic to the Orient, two occur in Mesoamerica, with one of these also
occurs in the Palaearctic Region, which has an additional endemic genus. The
last subgenus is Holarctic. Michener (2000), however, mentions that its
distribution extends into Kenya, yet Eardley and Urban (in preparation) do not
record a Kenyan species but did record one species from South Africa.
Diagnosis
Length: head 2.5-3.1 mm; scutum and scutellum 3.0-4.0 mm; forewing
6.1-8.4 mm; body 9.0-11.6 mm. Vestiture: pale white to brownish; sparse to
moderately dense pubescent; female scopa on S2-S5; male sternal comb on
S4-S5. Integument: mostly black, with limited to extensive pallid maculation

136
on face, scutellum and metasoma (Plate 19b); female mandible with four
dissimilar teeth, sometimes five or six, as in subgenera Massanthidium and
Congotrachusa; lower clypeal margin with six projections; subantennal suture
straight; juxtantennal carina absent; preoccipital area gently rounded; scuto-
scutellar suture closed, similar to scuto-axilla suture; scutellum, in dorsal
view, truncate or emarginate postero-medially; arolia absent, sometimes
reduced in subgenus Congotrachusa; metasomal without lateral spine;
gonostlyus expanded apically, deeply notched on inner margin; penis valves
reduced in size and narrowly separated (Figure 16d).
Genus Trachusoides Michener and Griswold
Trachusoides Michener and Griswold, 1994: 324. Type species: Trachusoidse
simplex Michener and Griswold, 1994, by original designation;
Michener and Griswold, 1994: 324; Michener, 2000: 519-520.
Comment
This non-parasitic, large megachiliform genus was first described by
Michener and Griswold (1994). This monotypic genus is endemic to southern
India. The representative species of this genus was not available for
examination during this study, due to the fact that, it is a rare specimen and the
museum was unwilling to loan it out.

137
CLADOGRAMS AND TREE TOPOLOGIES OF GENUS Anthidiellum
In the cladistic analysis of the genus Anthidiellum with heuristic
search, 385 most parsimonious trees were found, with L: 39 steps, CI: 79%
and RI: 81%. Figure 17 depicts a typical cladogram topology of most of the
385 most parsimonious trees. The strict consensus tree, on the other hand, had
L: 51, CI: 60%, RI: 53% collapsed 14 unsupported nodes (Fig. 18). The strict
consensus values using bootstrap and jackknife search tool were found to be:
L: 48/ 48, CI: 64/ 64%, RI: 60/ 60% (Figure 19-20).
The analyses of the ground-plan of those trees revealed three major
clades outside the outgroups (Trachusa and Eoanthidium) and these consist of:
(1) Anthidiellum flavescens and A. rubellum. 2) A. eritrinum, A. absonulum, A.
apicatum, A. auriscopatum, A. frontoreticulatum, A. kimberleyana, A.
orichalciscopatum, A. polyochrum, A. krugerense, A. sternale, A.
tegwaniense, A. zebra, A. bulawayense, A. otavicum, A. bipectinatum, A.
spilotum. The probable phylogenetic implications have been dealt with in the
discussion.
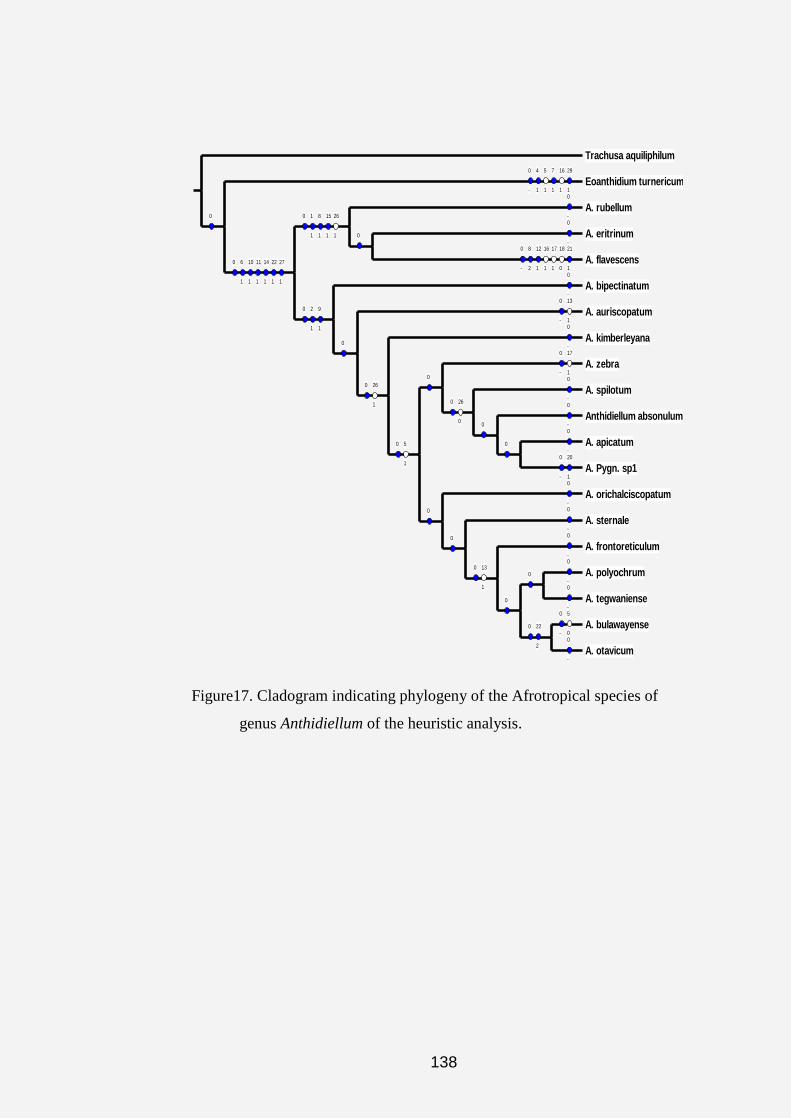
138
Trachusa aquiliphilum
Eoanthidium turnericum
Anthidiellum absonulum
A. apicatum
A. auriscopatum
A. bipectinatum
A. bulawayense
A. eritrinum
A. rubellum
A. flavescens
A. frontoreticulum
A. kimberleyana
A. orichalciscopatum
A. otavicum
A. polyochrum
A. Pygn. sp1
A. spilotum
A. sternale
A. tegwaniense
A. zebra
29
1
16
1
7
1
5
1
4
1
0
-
0
-0
-
13
1
0
-
0
-
5
0
0
-
0
-
0
-
21
1
18
0
17
1
16
1
12
1
8
2
0
-
0
-
0
-
0
-
0
-
0
-
20
1
0
-
0
-
0
-
0
-
17
1
0
-
0
0
26
0
0
0
0
22
2
0
0
13
1
0
0
0
5
1
0
26
1
0
0
9
1
2
1
0
0
26
1
15
1
8
1
1
1
0
27
1
22
1
14
1
11
1
10
1
6
1
0
0
Figure17. Cladogram indicating phylogeny of the Afrotropical species of
genus Anthidiellum of the heuristic analysis.

139
Figure 18. Cladogram indicating the strict concensus tree of the Afrotropical
species of genus Anthidiellum
Trachusa aquiliphilum
Eoanthidium turnericum
A. absonulum
A. apicatum
A. auriscopatum
A. bipectinatum
A. bulawayense
A. eritrinum
A. rubellum
Anthidiellum flavescens
A. frontoreticulum
A. kimberleyana
A. orichalciscopatum
A. otavicum
A. polyochrum
A. krugerense
A. spilotum
A. sternale
A. tegwaniense
A. zebra
83
70
70
92
90
99
100
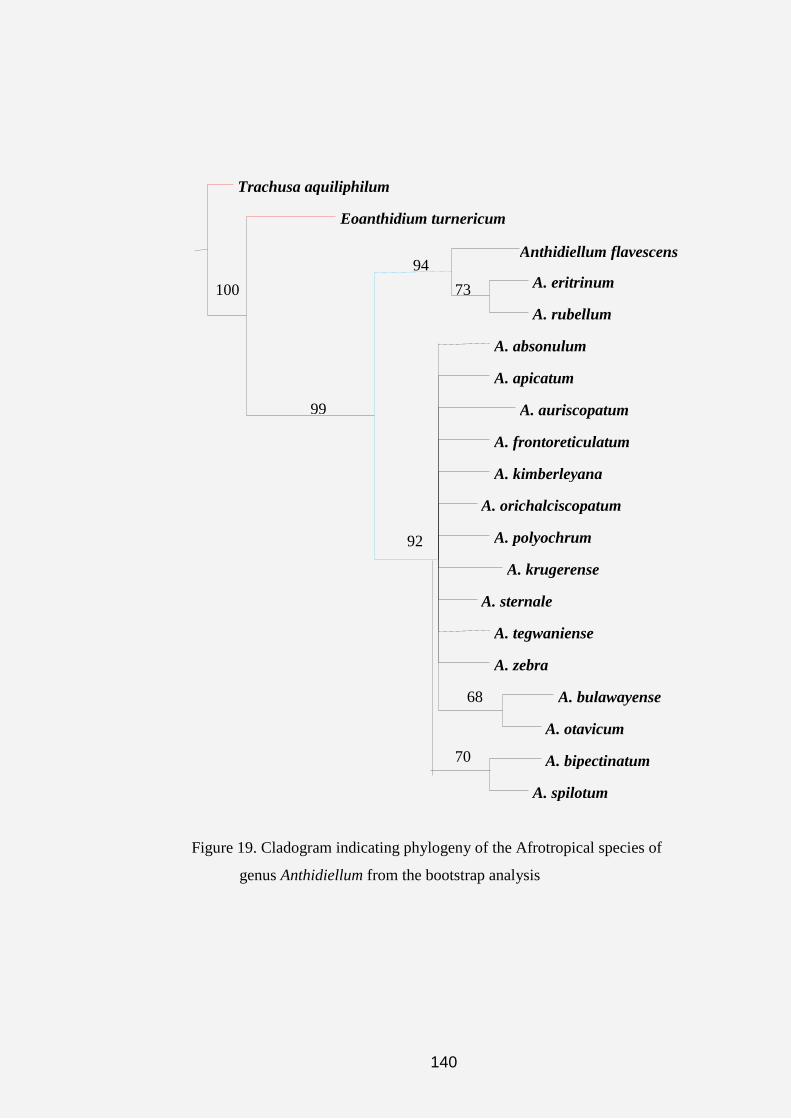
140
Figure 19. Cladogram indicating phylogeny of the Afrotropical species of
genus Anthidiellum from the bootstrap analysis
Trachusa aquiliphilum
Eoanthidium turnericum
A. absonulum
A. apicatum
A. auriscopatum
A. bipectinatum
A. bulawayense
A. eritrinum
A. rubellum
Anthidiellum flavescens
A. frontoreticulatum
A. kimberleyana
A. orichalciscopatum
A. otavicum
A. polyochrum
A. krugerense
A. spilotum
A. sternale
A. tegwaniense
A. zebra
73
68
70
94
92
99
100

141
Figure 20. Cladogram indicating phylogeny of the Afrotropical species of
genus Anthidiellum from the jackknife analysis
Trachusa aquiliphilum
Eoanthidium turnericum
A. absonulum
A. apicatum
A. auriscopatum
A. bipectinatum
A. bulawayense
A. eritrinum
A. rubellum
Anthidiellum flavescens
A. frontoreticulatum
A. kimberleyana
A. orichalciscopatum
A. otavicum
A. polyochrum
A. krugerense
A. spilotum
A. sternale
A. tegwaniense
A. zebra
73
70
70
95
89
100
100

142
SYSTEMATIC REVISION OF THE AFROTROPICAL SPECIES OF
THE GENUS Anthidiellum (COCKERELL)
Genus Anthidiellum Cockerell
Anthidium (Anthidiellum) Cockerell, 1904c: 3. Type species: Trachusa
strigata Panzer, 1805, by original designation.
Anthidiellum Cockerell: Pasteels, 1969: 45; Pasteels, 1984: 92; Michener and
Griswold, 1994: 312; Michener, 2000: 486.
Comment
This genus was described as a subgenus of Anthidium and later given
generic status by Pasteels (1969). However, the taxon appears be defined by
only two homoplasious apomorphic characters within the tribe Anthidiini. Its
arcuate subantennal suture and fovea of the scuto-axilla suture are also found
in Bathanthidium and Eoanthidium, though fovea is narrow in these genera, as
well as in Acanthidium. The fovea that is margined by a carina behind the
propodeal spiracle differs from that in Duckeanthidium and Stelis. However,
results in the analysis reveals two major clades of trees within the genus. This
apparently is not compatible with the subgeneric classification of genus by
Michener (2000). For the purpose of this study, the subgeneric classification
of Michener (2000) has been retained.
The Afrotropical genus comprises three subgenera. Two of these
subgenera are endemic species, while the third subgenus occurs in both Orient
and Afrotropical regions. 17 species are currently known to occur in
Afrotropical Region.

143
Keys to the Species
A taxonomic key is provided below for all the Afrotropical species that were
available for examination during this study.
1. Mandible with three teeth, apical region narrow; antennal scape
long, subequal 7X first flagella segment (male) -----------------------------2.
— Mandible with four teeth, apical region broad; antennal scape short, 3X
first flagella segment (female) -------------------------------------------------15.
2. Tegula broad anteriorly, lateral margin convex; pronotal lobe lamella------3.
— Tegula narrows anteriorly, lateral margin concave; pronotal lobe carinate
mesally, becoming progressively lamellate distally---------------------------5.
3. Sternal comb present medially on S5, head, mesosoma black, metasoma
yellowish brown --------------------------------------.Anthidiellum flavescens.
— Sternal comb absent, integument mostly reddish brown to black with
yellow integumental maculations -----------------------------------------------4.
4. Mesosoma entirely black, with large, very dense punctures; 12 hamuli on
hind wing--------------------------------------------------------------A. eritrinum.
— Mesosoma black, except yellow to reddish brown integumental maculation
on axilla and scutellum, punctations small and moderately dense; 15
hamuli on hind wing -------------------------------------------------A. rubellum.
5. Gonostylus narrows apically------------------------------------------------------6.
— Gonostylus broadens apically ----------------------------------------------------7.
6. Penis valve broad apically ----------------------------------------A. tegwaniense.
—. Penis valves narrow apically -------------------------------------------.A. zebra.
7. Ventral margin of clypeus with four projections -------------------------------8.

144
— Ventral margin of clypeus with five to six projections ----------------------11.
8. Hind wing with 10 hamuli on anterior margin ---------------------A. otavicum.
— Hind wing with 11 or more hamuli on anterior margin -----------------------9
9. Sternal comb very long (about 40 setae long) ----------------A.
frontoreticulatum.
— Sternal comb short (12 or less) -------------------------------------------------10.
10. Penis valve narrow apically, gonostylus slightly notched on outer margin -
---------------------------------------------------------------------------A. spilolum.
— Penis valve broad apically, gonostylus deeply notched on outer margin------
---------------------------------------------------------------------------.A. sternale.
11. Hind wing with 10 hamuli on anterior margin --------------A. bulaweyense.
— Hind wing with 11 or more hamuli on anterior margin ---------------------12.
12. Hind basitarsus slender, subequal half width of hind tibia ---A. absonulum.
—Hind basitarsus as broad as hind tibia-------------------------------------------13.
13. Vein cu-v of hind wing oblique (slants) -----------------------A. polyochrum.
— Vein cu-v of hind wing perpendicular (straight) -------------A. bipectinatum.
14. Tegula broad anteriorly, lateral margin convex; pronotal lobe lamellate;
distance between antennal sockets narrow, subequal two antennal socket
diameters -------------------------------------------------------------------------15.
— Tegula narrows anteriorly, lateral margin concave; pronotal lobe carinate
mesally, becoming progressively lamellate distally; distance between
antennal sockets wide, more than two antennal socket diameters --------17.
15. Preoccipita1 area carinate laterally; mesosoma yellowish brown; scutellum

145
(in dorsal view) truncate posteriorly -----------------------------A. flavescens.
— Preoccipital area carinate dorso-laterally; mesosoma reddish brown to
black with yellow or reddish integumental maculations; scutellum (in
dorsal view), emarginate medio-posteriorly --------------------------------16.
16. Mesosoma entirely black, with large, very dense punctures; 12 hamuli on
hind wing ------------------------------------------------------------.A. eritrinum.
— Mesosoma black, except yellow to reddish brown integumental maculation
on axilla and scutellum, punctations small and moderately dense; 15
hamuli on hind wing -------------------------------------------------A. rubellum.
17. Ventral margin of clypeus with four projections----------------------------18.
— Ventral margin of clypeus with five or more projections ------------------27.
18. Mandible with outer carina only -----------------------------------------------19.
— Mandible with both outer and inner carinae ----------------------------------22.
19. Hind wing with 10 hamuli on anterior region --------------------A. otavicum.
— Hind wing with 11 or more hamuli --------------------------------------------20.
20. Vein cu-v of hind wing oblique (curves back); 11 hamuli on hind wing ----
-----------------------------------------------------------------A. frontoreticulatum.
— Vein cu-v of hind wing perpendicular (straight); 12 hamuli on hind wing---
--------------------------------------------------------------------------------------21.
21. Hind basitarsus slender, subequal half as broad as hind tibia -----------------
----------------------------------------------------------------------A. tegwaniense.
— Hind basitarsus broad, as wide as hind tibia or broader than hind tibia.------
--------------------------------------------------------------------A. auriscopatum.

146
22. Hind wing with eleven hamuli on anterior margin -------------------------23.
— Hind wing with twelve hamuli on anterior region -----A. orichalciscopatum.
23. Vein cu-v of hind wing straight.------------------------------------------------24.
— Vein cu-v of hind wing oblique (curves back) -------------------------------25.
24. Tibiae with thick stout setae.---------------------------------------------A. zebra.
— Tibiae lack thick stout setae ----------------------------------------.A. apicatum.
25. Scutum (in dorsal view) extend laterally and longitudinally, forming
groove with carina next to tegula----------------------------------A. krugerense
— Scutum (in dorsal view) rounded, lack lateral extension --------------------26.
26. Tibiae with thick stout setae; mandible with very sharp outer teeth ----------
----------------------------------------------------------------------------A. sternale.
— Tibiae lacks thick stout setae; mandible with blunt outer teeth ----------------
---------------------------------------------------------------------------A. spilotum.
27. Mandible with outer carina only.------------------------------A. bulawayense.
— Mandible with both outer and inner carinae ----------------------------------28.
28. Vein cu-v of hind wing straight ------------------------------------------------29.
— Vein cu-v of hind wing oblique (curves back) -------------------------------30.
29. Hind wing with 11 hamuli.-----------------------------------------------A. zebra
— Hind wing with 12 hamuli--------------------------------------.A. kimberleyana.
30. Mandible with entirely large punctures ------------------------A. polyochrum.
— Mandible with large punctures basally, small punctures apically -------------
----------------------------------------------------------------------A. bipectinatum.

147
Subgenus Anthidiellum Cockerell s. str.
Anthidium (Anthidiellum) Cockerell, 1904c: 3. Type species: Trachusa
strigata Panzer, 1805, by original designation.
Anthidium (Cerianthidium) Friese, 1932: 304. Type species: Trachusa strigata
Panzer, 1805. Isogenotypic with subgenus Anthidiellum.
Anthidiellum (Anthidiellum) Cockerell: Pasteels, 1969: 45; Pasteels, 1984: 92;
Michener and Griswold, 1994: 312; Michener, 2000: 486.
Comment
This subgenus comprises two Afrotropical species, Anthidiellum
eritrinum (Friese) and Anthidiellum rubellum (Friese). A. eritrinum is endemic
Eritrea, whereas A. rubellum is only known from South Africa.
General Diagnosis of the Genus
Length: head 2.0-2.2 mm; scutum and scutellum 1.7-2.3 mm; forewing
3.8-5.4 mm; body 5.4-5.8 mm. Vestiture: cream to reddish brown; sparse to
impubescent; male sternal comb absent; upper edge of inner surface of hind
tibia not evidently marked by limited area of keirotrichiate. Integument: black,
except yellow or reddish maculation on most of face, entire margin of
preoccipital area, margins of scutum, axilla and scutellum, lateral regions of
T1-T4 (in A. eritrinum); preoccipital carina present dorso-laterally; paraocular
carina well developed; vertex convex, 2X ocellus diameters above eye;
distance between antennal sockets subequal two antennal sockets diameter;
subantennal suture strongly curved, subequal clypeus length; scape long, 5X

148
length of first flagellar segment; antennal flagellum of male long (0.70-0.84
head length); apex of female mandible twice as broad as base; pronotal lamella
well developed, extends mesad well past edge of scutum; tegula not narrowed
anteriorly, lateral margin convex; omaular distinct, weakly lamellate,
extending ventrally, not reaching midline; axilla extending laterally beyond
lateral margin of scutum; rows of pits across upper margin of propodeum
weakly developed, not carinate; corbiculate sculpture on propodeum confined
to basolateral region; entire surface of fore wing with numerous hair and
papillae; jugal lobe of hind wing about half as long as vannal lobe; T2-T5, and
T6 in dorsolateral view, gently curved towards apex; T5-T6 in female, T6-T7
of males curved weakly downward and appears swollen.
Anthidiellum (Anthidiellum) eritrinum (Friese)
Anthidium eritrinum Friese, 1915: 275, 295-296, female holotype (ZMHB)
Eritrea; Friese, 1915: 275, 295-296.
Anthidiellum eritrinum (Friese): Mavromoustakis, 1936a: 44, 45.
Anthidiellum (Anthidiellum) eritrinum (Friese): Pasteels, 1984: 92-93, 95, 156.
Diagnosis
Male. Unknown. Female. Length: head 2.1 mm; scutum and scutellum
2.3 mm; forewing 5.4 mm; body 5.8 mm. Vestiture: white to pale yellow,
except orange on vertex, mesosomal dorsum and under tarsi; mostly sparse,
tibiae with fairly dense pubescent, without thick, stout setae. Integument: head
black with postero-lateral regions of vertex and lower face (Plate 22a) mostly

149
yellow; mesosoma mostly black, pronotal lobe and tegula partly yellow, legs
almost entirely yellow (blackish proximally and orangish distally); metasoma
black to reddish-black with following areas yellow: dorso-lateral region of T1,
wide cross-bands on T2-T5, most of T6 (Plate 21a; Figure 21a); lower clypeal
margin with five projections (Figure 21a); mandible with large punctures
basally (shiny between punctures), fine sculpture near apex, two ventro-distal
sutures; two dissimilar, large outer teeth, two very small inner teeth; lateral
margin of scutum, dorsal view, straight; scuto-scutellar suture open, forming
narrow shiny fovea; scutellum, dorsal view, generally gently convex
posteriorly, concave postero-medially, obscures propodeum; 12 long hamuli
on hind wing; vein cu-v slants; hind basitarsus narrow, more than half as wide
as hind tibia; T2, dorsal view, gently convex laterally.
Distribution
Anthidiellum eritrinum (Friese) is endemic to Eritrea.
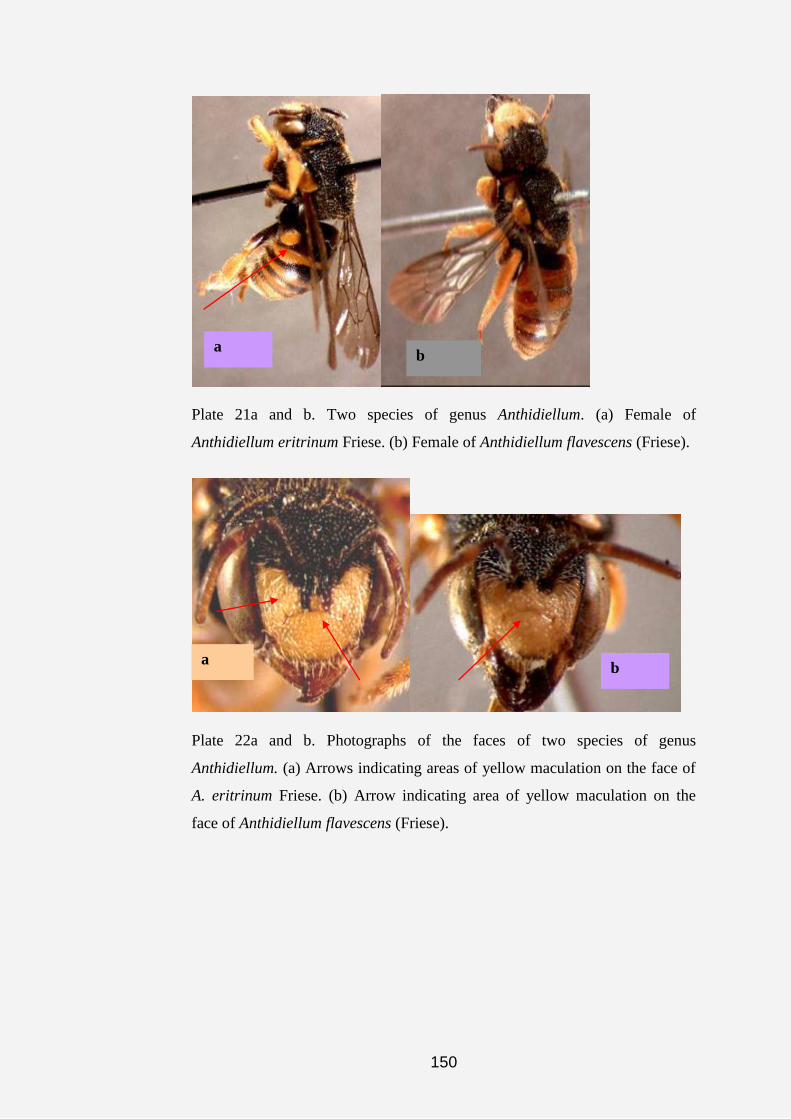
150
Plate 21a and b. Two species of genus Anthidiellum. (a) Female of
Anthidiellum eritrinum Friese. (b) Female of Anthidiellum flavescens (Friese).
Plate 22a and b. Photographs of the faces of two species of genus
Anthidiellum. (a) Arrows indicating areas of yellow maculation on the face of
A. eritrinum Friese. (b) Arrow indicating area of yellow maculation on the
face of Anthidiellum flavescens (Friese).
a b
a b
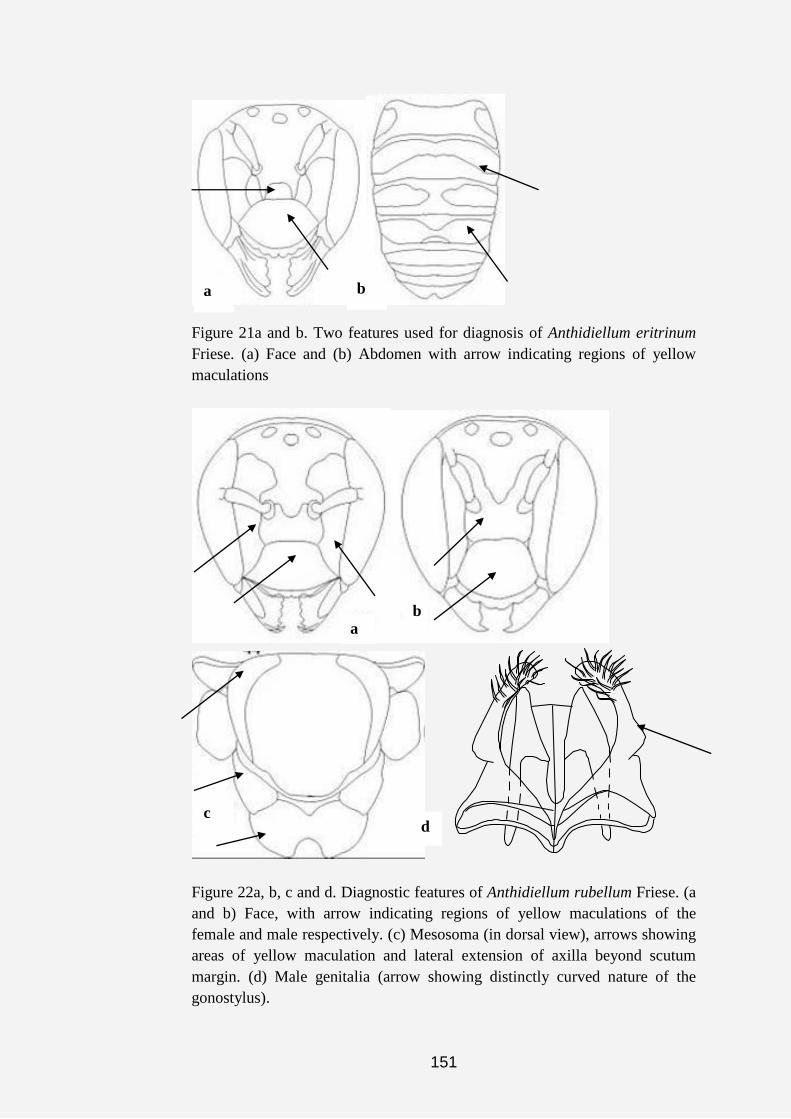
151
Figure 21a and b. Two features used for diagnosis of Anthidiellum eritrinum
Friese. (a) Face and (b) Abdomen with arrow indicating regions of yellow
maculations
A. rubellum
Figure 22a, b, c and d. Diagnostic features of Anthidiellum rubellum Friese. (a
and b) Face, with arrow indicating regions of yellow maculations of the
female and male respectively. (c) Mesosoma (in dorsal view), arrows showing
areas of yellow maculation and lateral extension of axilla beyond scutum
margin. (d) Male genitalia (arrow showing distinctly curved nature of the
gonostylus).
a b
c d
a b

152
Anthidiellum (Anthidiellum) rubellum (Friese)
Anthidium rubellum Friese, 1905e: 74, female holotype (TMSA) South Africa;
Friese, 1909a: 400, 401, 412; Anonymous, 1958: 33.
Anthidiellum (Anthidiellum) rubellum (Friese): Pasteels, 1969: 45.
Diagnosis
Female. .Length: head 2.2 mm; scutum and scutellum 1.9 mm;
forewing 4.0 mm; body 5.9 mm. Vestiture: white, except reddish to black on
fore tarsi; sparse to impubescent; thick stout setae absent on tibiae.
Integument: head, mesosoma black, metasoma brownish, except yellow or
reddish-brown maculation on clypeus (ventral margin black, Figure 22a),
ventral region of supraclypeus, entire paraoccular area, preoccipital region and
gena, dorso-lateral surface of scutum, entire axilla, most of scutellum (antero-
median region black), dorsal region of mesepisternum and mesepimeron,
entire legs; mandible with large punctures basally, small punctures apically,
shiny between punctures, with outer and inner carinae, two, similar, large
outer teeth and two, similarly very reduced teeth; clypeus margin with three
projections on each side (one medial and one lateral); scuto-scutellar suture
open, forming shiny fovea; lateral region of sutum rounded; scutellum, in
dorsal view, truncate posteriorly, obscures propodeum (Figure 22c); 15 short
hamuli on hind wing; vein cu-v slants; hind basitarsus broad, more than half as
wide as hind tibia.
Male. Length: head 2.0 mm; scutum and scutellum 1.7 mm; forewing
3.8 mm; body 5.4 mm. Vestiture and integument: similar to female, except for

153
sex limit characters and moderately dense lateral fringes on S2-S4, S5 lacks
sternal comb; apex of mandible half as wide as in female (Figure 22b), 3 very
small teeth; gonostylus wide apically, without a notch on outer margin; penis
valves narrowly separated (Figure 22d).
Distribution
Anthidiellum rubellum Friese is endemic to South Africa.
Subgenus Chloranthidiellum Mavromoustakis
Anthidiellum (Chloranthidiellum) Mavromoustakis, 1963b: 491. Type species:
Anthidium minutulum flavescens Friese, 1925, by original designation;
Pasteels, 1984: 92; Michener and Griswold, 1994: 312; Michener,
2000: 487.
Anthidiellum (Chloranthidium) [!] Mavromoustakis: Pasteels, 1969: 48.
Comment
This subgenus is monotypic, comprising Anthidiellum flavescens
(Friese), endemic to East and southern Africa.
This study agree with the position of Anthidiellum hessi (Cockerell), as
synonym of A. flavescens based on similar morphological characters such as
integument colour and vestiture, carina on head and genitalia of male.

154
Anthidiellum flavescens (Friese)
Anthidium minutulum var. flavescens Friese, 1925a: 509, male holotype
(NMBZ) Zimbabwe.
Anthidiellum flavescens (Friese): Mavromoustakis, 1936b: 605-606; Michener,
2000: 487.
Anthidiellum (Chloranthidiellum) flavescens (Friese): Mavromoustakis,
1963b: 491; Pasteels, 1984: 93-95.
Anthidiellum (Chloranthidiellum) flavescens nigrescens Pasteels, 1984: 94,
female holotype (SEMC) Kenya.
Anthidiellum hessei Cockerell, 1932c: 523-524, male syntypes (SAMC or
NHML) Namibia; Cockerell, 1936c: 456; Mavromoustakis, 1936b:
605, syn.
Diagnosis
Female. Length: head 1.4-1.8 mm; scutum and scutellum 1.9-2.1 mm;
forewing 4.5-4.6 mm; body 5.6-5.8 mm. Vestiture: white, except reddish to
black on tarsi; sparse to impubescent; tibiae without thick, stout setae.
Integument: head and mesosoma black, except yellow maculations on most of
lower half of face (Plate 22b), a stripe on dorsal corner of preoccipital area,
spot on doral surface of axilla, entire dorsal and lateral surfaces of legs (Figure
23e); anterior metasoma orange to reddish-brown with yellow maculations on
anterior regions of T1-T6 (Figure 23b; Plate 21b); preoccipital carina present
laterally, absent dorsally; paraocular carina well developed; vertex convex, 2X
ocellus diameters above eye; distance between antennal sockets subequal two

155
antennal socket diameter; subantennal suture strongly curved, subequal to
clypeus length; scape long (first flagellar segments less than 3X scape length);
antennal flagellum short (about half head length); lower clypeal margin with
five very small papillae (one median, two lateral); apex of mandible 1.5X as
wide as base; mandible finely sculptured with small punctures, shiny between
punctures, with two carinae (outer and inner carinae), two large similar outer
teeth and two small dissimlar inner teeth; pronotal lobe strongly lamellate,
lamella tapers, extending nearly as far mesad from lateral margin of scutum as
laterad; tegula, not narrowed anteriorly, with lateral margin convex; omaular
distinct, weakly lamellate, extending ventrally, not reaching midline; axilla
extending laterally beyond lateral margin of scutum; scuto-scutellar suture
open, forming shiny fovae; scutellum, in dorsal view, truncate posteriorly,
hides median region of propodeum (Figure 23a); rows of pits across the upper
margin of propoduem strongly developed, evident more horizontally, lateral
margin with carina; 12 hamuli on hind wing; vein cu-v slants; hind barsitarsi
nearly as long as broad.
Male: Length: head 1.5-1.6 mm; scutum and scutellum 1.6-1.7 mm;
forewing 4.2-4.3 mm; body 4.8-5.0 mm. Vestiture and integument: Similar to
female (Figure 23c), except apical region of S5 with medial comb of about
thirty similar setae; clypeus entirely yellow; mandibles mostly bright yellow
with apex dark brown; gonstylus expanded apically, deeply notched on outer
margin; penis valves narrowly separated (Figure 23d).
156
Distribution
Anthidiellum flavescens (Friese) is widespread in Kenya and southern
Africa.
157
A. flavescens
A. flavescens
Figure 23a, b, c, d and e. Diagnostic features of Anthidiellum flavescens
(Friese). (a) Mesosoma (in dorsal view), arrows showing areas of yellow
maculation and lateral extension of axilla beyond scutum margin. (b and c)
Metasoma in dorsal view, with arrows indicating regions of yellow
maculations of the female and male, respectively. (d) Male genitalia. (e) Hind
leg (arrows showing regions of integumental maculations).
a
b
d
c
e

158
Subgenus Pycnanthidium Krombein
Pycnanthidium Krombein, 1951: 292. Type species: Pycnanthidium solomonis
Krombein, 1951, by original designation.
Pygnanthidium [!] Krombein, 1951: Mavromoustakis, 1963b: 491.
Pygnanthidium [!] (Pygnanthidiellum) Mavromoustakis, 1963b: 492. Type
species: Anthidium zebra, 1904, by original designation.
Anthidiellum (Pycnanthidium) Krombein: Pasteels, 1969: 45; Michener and
Griswold, 1994: 312; Michener, 2000: 487.
Anthidiellum (Pygnanthidiellum [!]) Mavromoustakis: Pasteels: 1984: 94.
Comment
This is the most diverse group among the Afrotropical subgenera.
Pasteels (1984) listed 13 Afrotropical species. This subgenus also occurs in
Asia and Australia. Currently, two Madagascan species have been included,
bringing the number of species to 15 Afrotopical species. These consist of
Anthidiellum absonulum (Cockerell)= A. aspidopodum Mavromoustakis, A.
apicatum (Smith), A. bipectinatum Pasteels, A. bulawayense Mavromoustakis,
A. madli Pauly, A. mediale Pasteels, A. micheneri Pauly, A. orichalciscopatum
(Strand) = A. auriscopatum, A. otavicum (Cockerell), A. polyochrum
Mavromoustakis, A. spilotum (Cockerell) = A. Kimberleyana (Friese), A.
sternale, A. tegwaniense (Cockerell), A. transversale Pasteels, A. zebra
(Friese) = Anthidiellum medionigrum Cockerell.

159
Diagnosis
Length: head 1.3-2.2 mm; scutum and scutellum 1.5-2.2 mm; forewing
2.5-5.0 mm; body: 3.5-6.0 mm. Vestiture: white, sparse to impubescent,
moderately long and dense around antennal sockets; male sternal comb on
apical region of S5; thick stout setae sometimes on tibiae; upper edge of inner
surface of hind tibia evidently marked by limited area of keirotrichiate.
Integument: black with yellow maculation on face, dorsal region of
preoccipital area, sometimes on margins of scutum, axilla and scutellum,
meso-lateral regions of metasoma; preoccipital area carinate dorsally;
paraoccular carina very reduced; vertex convex, 2X ocellus diameters above
eye; distance between antennal sockets more than two antennal socket
diameters; subantennal suture weakly curved, nearly 0.6X clypeus length;
scape long (7X first flagellar segment); antennal flagellum short (subequal to
head length); apical region of female mandible twice as broad as base;
pronotal carina small mesally, becoming progressively lamellate distally;
tegula narrow anteriorly, lateral margin slightly concave, except in A.
krugerense where lateral region of scutum becomes strongly carinate (Fig.
102); omaular weakly carinate, lamellate laterally, extends ventrally to midline
(left and right omaular join); axilla not extending laterally beyond lateral
margin of scutum; rows of pits across upper margin of propoduem very
reduced, evident only laterally, not carinate; T2-T5 of females, and to T6 of
males, in dorsolateral view, gently curved towards apex; propodeum
scrobiculate dorsolaterally; female T1 carinate antero-dorsally; T2-T5, in
dorsolateral view, curved gently downwards

160
Anthidiellum (Pycnanthidium) absonulum (Cockerell)
Anthidium absonulum Cockerell, 1932b: 526-527, male holotype (NHML)
Democratic Republic of the Congo; Mavromoustakis, 1937a: 232;
Mavromoustakis, 1937b: 267; Mavromoustakis, 1963b: 499.
Anthidium absonulum Cockerell [!]: Mavromoustakis, 1934: 43;
Mavromoustakis, 1963b: 499.
Anthidiellum absonulum (Cockerell): Cockerell, 1936c: 455.
Anthidiellum (Pycnanthidium) absonulum (Cockerell): Pasteels, 1984: 96, 99-
100.
Anthidiellum aspidopodium Mavromoustakis, 1936a: 45-47, female holotype
(NMBZ or NHML) Zimbabwe.
Pygnanthidium (Pygnanthidium) absonulum (Cockerell): Mavromoustakis,
1963b: 492.
Pygnanthidium (Pygnanthidium [!]) aspidopodium (Mavromoustakis):
Mavromoustakis, 1963b: 492; Pasteels, 1984: 99, syn.
Diagnosis
Female. Length: head 1.5-1.9 mm; scutum and scutellum 1.5-2.1 mm;
forewing 3.2-4.6 mm; body 4.5-5.9 mm. Vestiture: white; sparse, dense on
tibiae and tarsi; thick stout setae on tibiae. Integument: black, except yellow
maculation mesad to antennal socket, below antennal sockets, lower
paraocular area, most of clypeus (ventral clypeal margin black, Figure 24a), a
spot on dorso-lateral margin of preoccipital area, proximal end of dorsal
surface of fore tibia, dorsal surface of hind tibia (varying from basal spot to

161
almost entire dorsal surface, Figure 24g), meso-lateral regions of T1-T4 (wider
laterally on T3, medially interrupted cross-band on T4), continuous cross-band
on T5, almost entire T6 (Figure 24d); distal tarsal segments orange; mandible
punctate basally, finely sculptured apically, shiny between punctures, with
inner and outer carinae, two large outer teeth, two small inner teeth; lower
clypeal margin with four very small papillae (two on either side of mid-line);
lateral margin of scutum, in dorsal view, gently rounded; scuto-scutellar suture
open, forming shiny fovea; scutellum, in dorsal view, emarginate medio-
posteriorly, slightly obscures propodeum (Figure24c); 11 hamuli on hind
wing; vein cu-v straight; hind basitarsus broad, much broader than middle
basitarsus, long, half as long as hind tibia.
Male. Length: head 1.5-1.8 mm; scutum and scutellum 1.6-2.0 mm;
forewing 4.5-4.8 mm; body 5.2 mm. Vestiture and integument: similar to
female, except differ in the following characters: S5 with distal comb,
narrowly interrupted medially, about 28 similar setae; yellow maculation on:
entire clypeus, most of mandible, apex black (Figure 24b), ventral surfaces of
fore and middle femura, dorsal surfaces of fore and middle tibiae and basitarsi,
proximal and distal ends of dorsal surface of hind tibia, hind basitarsus (Figure
24f), medially interrupted cross-band on T3, continuous or medially
interrupted cross-band on T4, most of T5-T7, distal margins reddish; apex of
mandible half as wide as female, three subequal teeth (Figure 24b); S3 with
short, median notch; gonostylus expanded apically, slightly notched on outer
margin; penis valves seperated narrowly (Figure 24e).
162
Distribution
Anthidiellum absonulum is widespread throughout central, eastern and
southern regions of Africa.
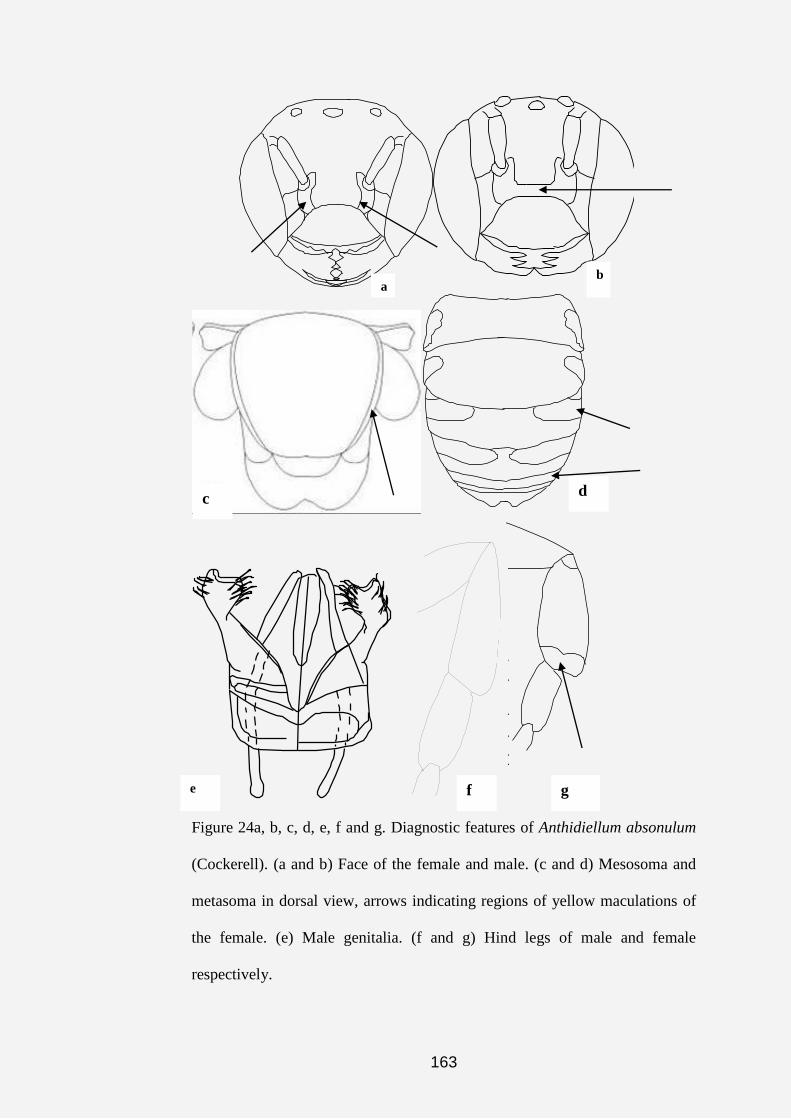
163
Fig A1 Fig. A2
A. absonulum (female)
A. bipectinatum (female)A. absonulum
A. absonulum
A. absonulum
(male)
Figure 24a, b, c, d, e, f and g. Diagnostic features of Anthidiellum absonulum
(Cockerell). (a and b) Face of the female and male. (c and d) Mesosoma and
metasoma in dorsal view, arrows indicating regions of yellow maculations of
the female. (e) Male genitalia. (f and g) Hind legs of male and female
respectively.
A. a
bso
nulu
m
A. a
bso
nu
lum
(ma
le)
g
d
a
f
c
b
e

164
Anthidiellum (Pycnanthidium) apicatum (Smith)
Anthidium apicatum Smith, 1879: 84, female holotype (NHML) South Africa.
Anthidium nigripes Friese, 1904b: 105, female holotype (ZMHB)
Mozambique; Friese 1905b: 67; Friese 1909a: 400, 409; Friese 1909b:
158, 166; Strand 1912a: 143; Strand 1912b: 307; Pasteels 1984: 98,
syn.
Anthidium (Pygnanthidiellum [!]) apicatum Smith: Friese 1905b: 66; Friese
1909a: 400, 404; Friese 1909b: 158.
Anthidiellum (Pygnanthidiellum [!]) apicatum (Smith): Pasteels 1984: 98, 165.
Dianthidium melanocephalum Cockerell, 1920: 298-299, 300, three female
syntypes (NHML) South Africa; Pasteels 1984: 98, syn.
Anthidiellum nigripes var. rhodesinum Mavromoustakis, 1937a: 231, female
holotype (TMSA) Zimbabwe.
Anthidiellum nigripes var. rhodesianum [!] Mavromoustakis: Anonymous
1958: 33.
Pygnanthidium (Pygnanthidiellum) apicatum (Smith): Mavromoustakis
1963a: 492.
Diagnosis
Male. Unknown. Female. Length: head 1.7-2.4 mm; scutum and
scutellum 1.6-2.0 mm; forewing 3.7-4.4 mm; body 4.8-5.7 mm. Vestiture:
white; sparse; thick stout setae absent on tibiae. Integument: black, tegula, legs
Figure 25b) and metasoma dark brown, except yellow maculations on meso-
lateral regions of T1-T3 (yellow area increases progressively from a lateral

165
spot on T1 to occupying lateral third of T3), antero-lateral region of T4
yellow, medial regions of T5-T6 (Figure 25a); mandible with large basally,
small punctures apically, shiny between punctures, outer and inner carinae,
two large outer teeth and two similar inner teeth subequal to outer teeth;
clypeus margin with four projections, two on each side; lateral region of
scutum, in dorsal view, rounded; scuto-scutellar suture open, forming shiny
fovea; scutellum, in dorsal view, emarginate medio-posteriorly, hides median
region of propodeum; 11 long hamuli on hind wing; vein cu-v straight; hind
basitarsus broad, nearly as wide as base of hind tibia.
Anthidiellum (Pycnanthidium) auriscopatum (Strand)
Anthidium auriscopatum Strand, 1912b: 308, two female holotypes (HUMB)
Equatorial Guinea; Strand, 1920: 102.
Anthidiellum auriscopatum (Strand): Mavromoustakis, 1936a: 44, 45;
Pasteels, 1984: 99, syn.
Diagnosis
Male. Unknown. Female. Length: head 1.9-2.1 mm; scutum and
scutellum: 1.9-2.1 mm; forewing 3.7-3.9 mm; body 5.4-6.1 mm. Vestiture:
mostly white, except black to reddish on fore tibia; sparse; tibiae without thick
stout setae. Integument: black, brownish on legs, except yellow to reddish-
brown maculations on most of lower half of face (mandible mostly black,
Plate 23a), lateral region of axilla, dorso-lateral region of scutellum, basi-
dorsal and distal regions of tibiae (Plate 23b), entire surface of hind basitarsus
(figure 26b), meso-lateral regions of T1-T2 (yellow area increases

166
progressively from a lateral spot on T1 to occupying lateral third of T2); entire
anterior regions of T3-T5, T6 entirely yellow; mandible with large punctures
basally, small punctures apically, shiny between punctures, with outer carina
only, two, similar, large outer teeth and two, similarly small teeth; clypeus
margin with six small projections (one on median, two on lateral (Figure 26a);
lateral region of scutum rounded; scuto-scutellar suture open, forming shiny
fovea; scutellum weakly emarginated medio-posteriorly; 12 small hamuli on
hind wing, vein cu-v straight; hind basitarsus broad, as wide as base of hind
tibia.
Distribution
Anthidiellum auriscopatum is endemic to Congo and Equatorial
Guinea.
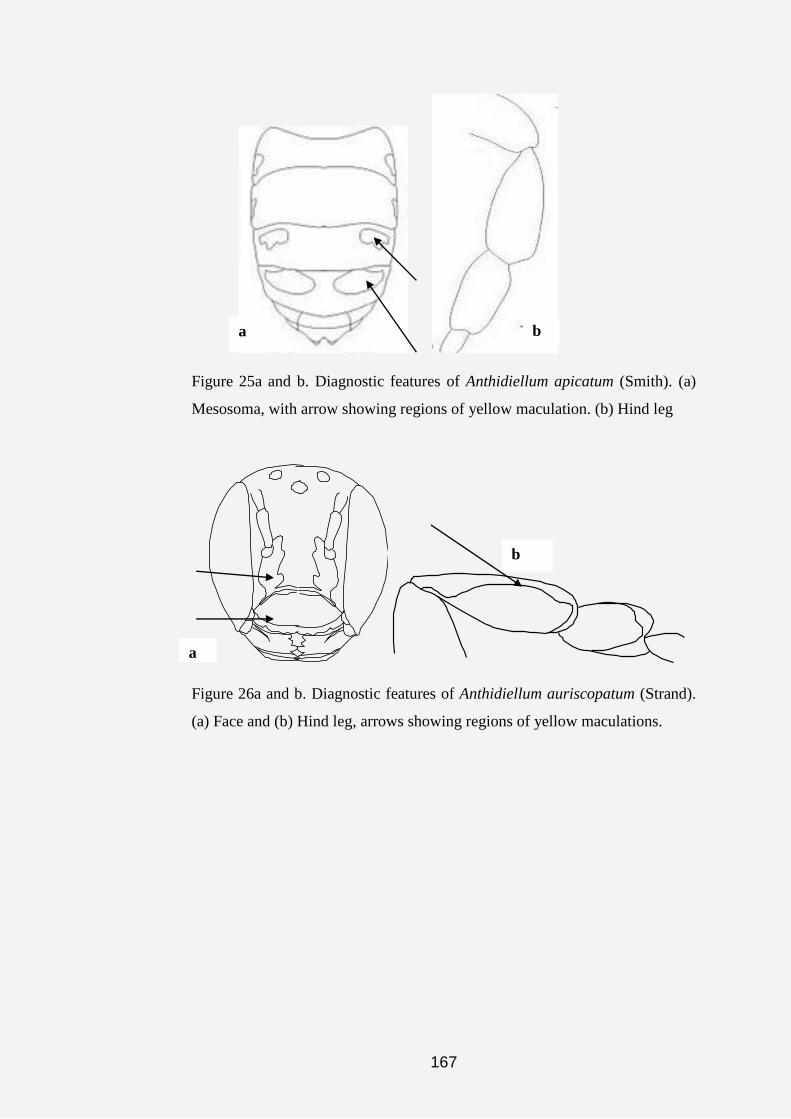
167
Figure 25a and b. Diagnostic features of Anthidiellum apicatum (Smith). (a)
Mesosoma, with arrow showing regions of yellow maculation. (b) Hind leg
A. auriscopatum (Female)
A. auriscopatum
Figure 26a and b. Diagnostic features of Anthidiellum auriscopatum (Strand).
(a) Face and (b) Hind leg, arrows showing regions of yellow maculations.
a b
Fig.
27
a
b

168
Plate 23a and b. Diagnostic features of Anthidiellum auriscopatum (Strand).
(a) Face and (b) Hind leg of with arrows indicating regions of yellow
maculation.
Plate 24a and b. Anthidiellum bipectinatum Pasteels. (a) Adult Female in
dorsolateral view. (b) Arrow showing regions of yellow integumental
maculation on the face.
c
d
a
b

169
Anthidiellum (Pycnanthidium) bipectinatum Pasteels
Anthidiellum (Pygnanthidiellum) bipectinatum Pasteels, 1984: 97, 105-106,
male holotype (AMNH); South Africa.
Diagnosis
Female. Length: head 1.5-1.9 mm; scutum and scutellum 1.8-2.2 mm;
forewing 3.9-4.7 mm; body 5.3-6.3 mm. Vestiture: white, sparse to
impubescent; tibiae without thick stout setae. Integument: black to dark brown
(lighter on tegula, legs and metasoma), with yellow maculations on most of
lower half of face (mandible and ventral margin of clypeus black, Figure 27a,
Plate 24a and b), proximal end of fore tibia, spot near base of middle tibia,
most of dorsal surface of hind tibia, entire dorsal surface of hind basitarsus
(Figure 27d), meso-lateral regions of T1-T4 (yellow area increases
progressively from a lateral spot on T1 to occupying lateral third of T4); entire
T5-T6 yellow (Figure 27c); mandible with large punctures at base and small
punctures towards apex, shiny between punctures, with both outer and inner
carinae, two, similar, large outer teeth and two, similarly small teeth; lower
clypeal margin with three projections, a median and two lateral; lateral margin
of scutum, in dorsal view, rounded; scuto-scutellar suture forms shiny fovea;
scutellum, in dorsal view, emarginate medio posteriorly, slightly obscures
propodeum; 11 small hamuli on hind wing, vein cu-v slants; hind basitarsus
broad, nearly as broad as base of hind tibia.
Male. Length: head 1.6-1.9 mm; scutum and scutellum 1.9-2.0 mm;
forewing 3.9-4.7 mm; body 5.8-6.3 mm. Vestiture and integument: similar to

170
female, except for gender limited characters and S3-S5 with short, distal
fringes; S5 with lateral comb widely seperated medially, with about eighteen
setae; clypeus entirely yellow (without black ventral margin), most of
mandible yellow, apex black (Figure 27b), ventral surface of fore femur,
dorsal surface of fore tibia, part of dorsal surfaces of middle and hind tibiae,
entire dorsal surfaces of basitarsi, dorsolaterally on T3, dorsally on T4, most
of T5-T7, distal margins reddish; apex of mandible half as wide as female,
teeth very small.
Distribution
Anthidiellum bipectinatum is endemic to southern Africa.
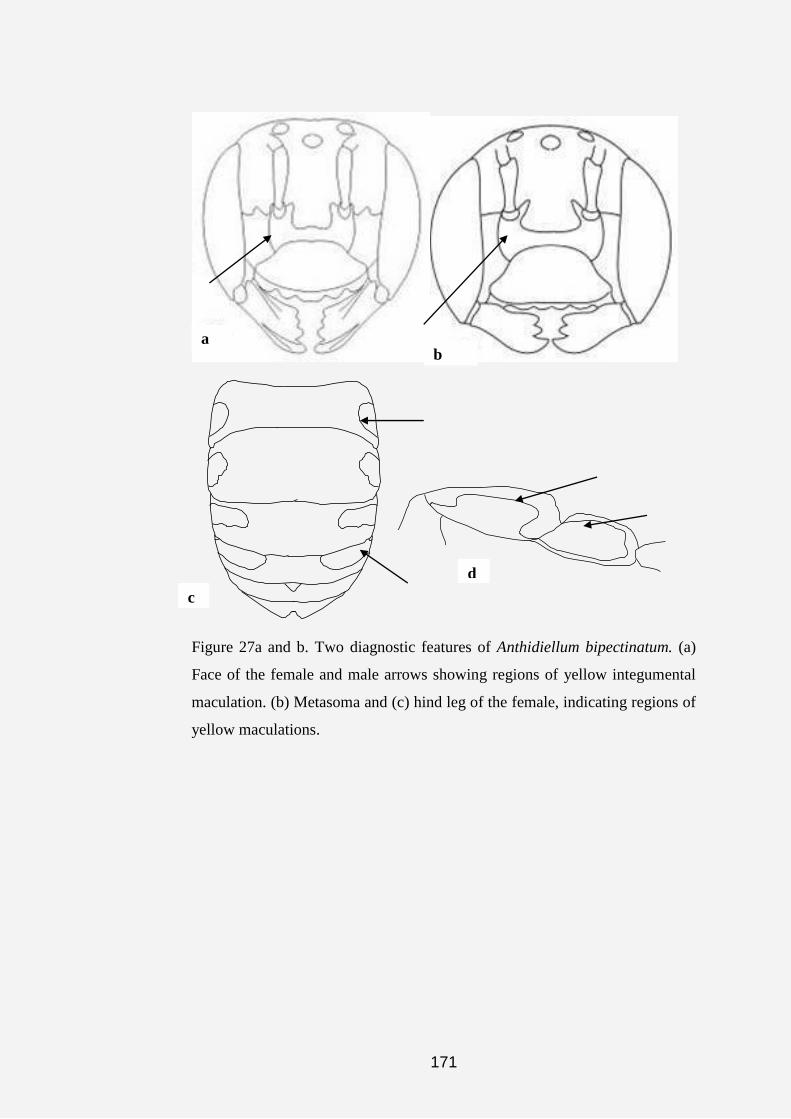
171
A. absonulum (female)
A. bipectinatum (female)
Figure 27a and b. Two diagnostic features of Anthidiellum bipectinatum. (a)
Face of the female and male arrows showing regions of yellow integumental
maculation. (b) Metasoma and (c) hind leg of the female, indicating regions of
yellow maculations.
A. bipectinatum(female)
a b
c
d

172
Anthidiellum (Pycnanthidium) bulawayense Mavromoustakis
Anthidiellum bulawayense Mavromoustakis, 1937a: 231-232, male holotype
(TMSA) Zimbabwe.
Anthidiellum (Pygnanthidiellum) bulawayense Mavromoustakis: Pasteels,
1984: 96, 99.
Diagnosis
Female. Unknown. Male. Length: head 1.3-1.5 mm; scutum and
scutellum 1.6-1.8 mm; forewing 3.9-4.2 mm; body 4.6-4.9 mm. Vestiture:
white; sparse to impubescent, S2-S3 with short fringe on distal margin; S4
with long fringe, S5 with a lateral comb, (widely seperated medially) with
about fourteen subequal setae, thick stout setae absent on tibiae. Integument:
black, extremities reddish-black, yellow maculation on most of lower half of
face, black on dorso-medial region of supraclypeus (Figure 28), small yellow
spot on vertex, part of pronotal lobe and medio-lateral region of scutum,
narrow postero-lateral margin of scutellum, entire or distal part of ventral
surface of fore femur, dorsal surface of fore tibia, proximal and distal ends of
middle and hind tibiae, entire dorsal surface of middle and hind basitarsi,
meso-lateral region of T1 (small lateral spot), T2 similar to T1 or black, T3
similar to T4 or black, T4 (small yellow area in middle of lateral half),
posterior margin of T5 yellow, entire surfaces of T6-T7 (Plate 25); mandible
with large punctures basally and small punctures apically, shiny between
punctures, outer carina only, 1 large outer tooth, 2 similar, small inner teeth
(Figure 27); lower clypeal margin with 3-5 projections, one median, one-two

173
lateral; lateral margin of scutum, in dorsal view, rounded; scuto-scutellar
suture open, forming shiny fovea; scutellum, in dorsal view, emarginate
medio-posteriorly, hides median region of propodeum; 10 small hamuli on
hind wing; vein cu-v straight; hind basitarsus slender, half as long as hind
tibia.
Distribution
Anthidiellum bulaweyense is known from tropical Africa and dry
savannah in Zimbabwe.
Anthidiellum (Pycnanthidium) frontoreticulatum sp. nov.
Anthidiellum (Pycnanthidium) frontoreticulatum, female holotype (MRAC)
Kenya.
Description
Female. Length: head 1.5-1.6 mm; scutum and scutellum: 1.5-1.8 mm;
forewing 3.5-3 8 mm; body 4.8-5.0 mm. Vestiture: mostly white, brownish on
tarsi; sparse to impubescent, moderately dense around antennal socket,
mesopleuron, tibiae and tarsi; tibiae without thick stout setae. Integument:
mostly black, except yellow maculations on lower region of face (mandible
entirely black, Plate 26a), lateral regions of T1-T2, meso-lateral regions of T3-
T4 (small on T3), medial region of T6 (Figure 29a); mandible with large
punctures basally, small punctures apically, shiny between punctures, with
outer carina only, two, similar, large outer teeth and two, similarly small teeth;

174
clypeus margin with four small projections (two on each side); lateral region
of scutum rounded; scuto-scutellar suture open, forming shiny fovea;
scutellum, in dorsal view, truncate posteriorly; 11 small hamuli on hind wing,
vein cu-v slants; hind basitarsus broad, nearly as wide as base of hind tibia
(Figure 29c).
Male. Length: head 1.4-1.5 mm; scutum and scutellum 1.5-1.6 mm;
forewing 3.9-4.1 mm; body 4.2-4.5 mm. Vestiture: similar to female, except
for gender limited characters and S5 with lateral comb narrowly seperated
medially, with about 40 similar setae (c.f. Fig. 8). Integument: similar to
female, except clypeus entirely yellow (without black ventral margin), most of
mandible yellow, apex black, basal region of dorsal surfaces of fore, middle
and tibia, lateral region of T1; apex of mandible half as wide as female, teeth
very small; gonostylus apically expanded, notched on outer margin; penis
valves narrow apically, widely separated (Figure 29b)
Distribution
Anthidiellum frontoreticulatum sp. nov is endemic to Kenya.

175
A. bulaweyense(male)
Figure 28. Face of Anthidiellum bulawayense Mavromoustakis with arrows
showing regions of yellow maculation
A. frontorecticulumA. frontoreticulum
A. frontorecticulum
Figure 29a, b and c. Diagnostic features of Anthidiellum frontoreticulatum sp.
nov. (a) Metasoma, arrows showing regions of yellow maculations. (b) Male
genitalia. (c) Hind leg, arrow showing region of yellow maculation.
a
c
b

176
Plate 25. Male of Anthidiellum bulawayense Mavromoustakis.
Plate 26a, b and c. Diagnostic features two species of the genus Anthidiellum.
(a) Face of Anthidiellum frontoreticulatum sp. nov. with arrow showing
regions of yellow maculations. (b and c) Face and hind leg (b) of Anthidiellum
kimberleyana (Friese).
a
b
c
a

177
Anthidiellum (Pycnanthidium) kimberleyana (Friese)
Anthidium kimberleyana Friese, 1922: 30-31, female holotype (HUMB) South
Africa; Cockerell, 1936a: 3.
Anthidiellum kimberleyana (Friese): Mavromoustakis, 1936a: 44-45.
Anthidiellum (Pygnanthidiellum) kimberleyana (Friese): Pasteels, 1969: 46-
47; Gess, 1981: 25, 61; Pasteels, 1984: 106.
Diagnosis
Male. Unknown. Female. Length: head 1.9 mm; scutum and scutellum
2.0 mm; forewing 4.5 mm; body 5.0 mm. Vestiture: white, except reddish to
black on tarsi; sparse, tibiae without thick, stout setae. Integument: black, legs
reddish-brown to black (Figure 30c; Plate 26c), except yellow maculations on
paraocular area (Figure 30a; Plate 26b), a stripe on dorsal margin of
preoccipital area, antero-laterally on T1-T2, meso-laterally on T3-T4, T5 with
small yellow spots dorso-laterally (Figure 30b); mandible with large punctures
basally, shiny between punctures, with two carinae (outer and inner carinae),
two large dissimilar outer teeth and two small dissimilar inner teeth; lower
clypeal margin with five very small papillae (one median, two lateral); lateral
margin of scutum, in dorsal view, rounded; scuto-scutellar suture open,
forming shiny fovea; scutellum, in dorsal view, emarginated medio-
posteriorly, hides median region of propodeum; 12 hamuli on hind wing; vein
cu-v straight; hind basitarsi nearly as long as broad.
DISTRIBUTION
Anthidiellum kimberleyana (Friese) is endemic to South Africa.
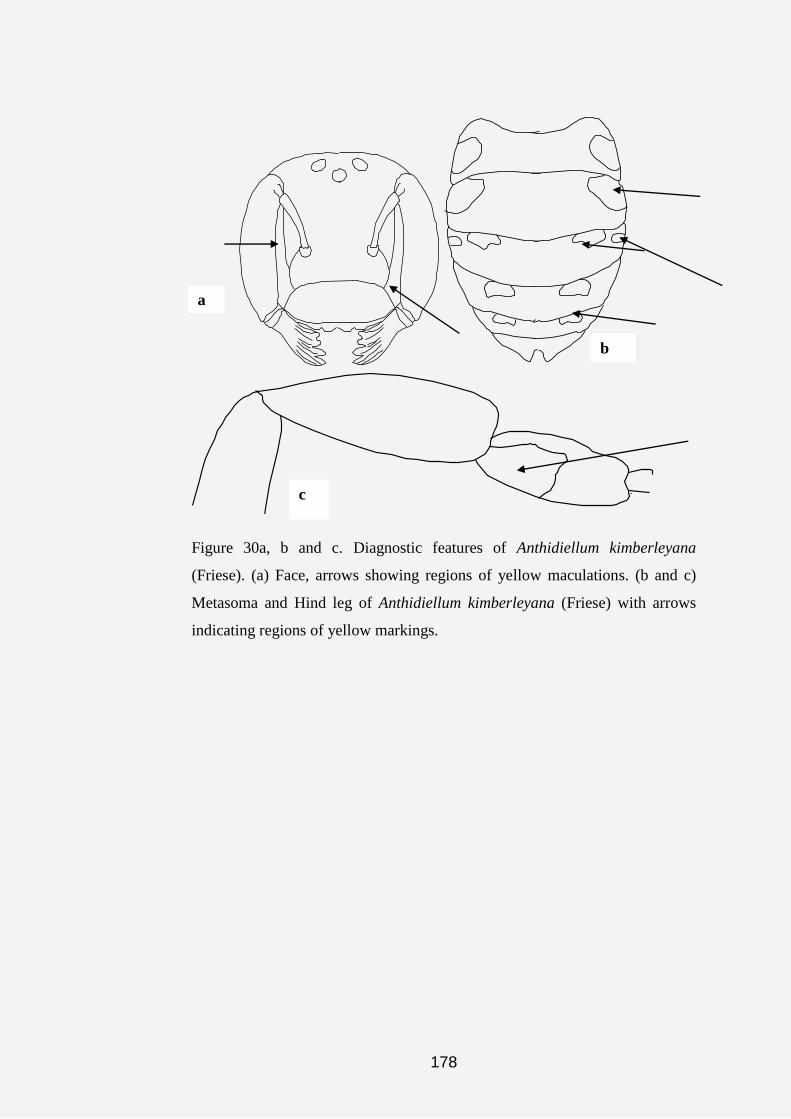
178
A. kimberlyana(female) A. kimberlyana
A. kimberlyana (female)
Figure 30a, b and c. Diagnostic features of Anthidiellum kimberleyana
(Friese). (a) Face, arrows showing regions of yellow maculations. (b and c)
Metasoma and Hind leg of Anthidiellum kimberleyana (Friese) with arrows
indicating regions of yellow markings.
Fig
. 71 a
c
b

179
Anthidiellum (Pycnanthidium) madli Pauly
Anthidiellum (Pycnanthidium) madli Pauly [In] Pauly et. al., 2001: 229, male
holotype (NHMV) Madagascar.
Comment
This genus endemic to Madagascar was not available for examination
during this study, due to the fact that, it is a rare specimen unwilling to loan
them out.
Anthidiellum (Pycnanthidium) mediale Pasteels
Anthidiellum (Pygnanthidiellum) mediale Pasteels, 1984: 96, 101-103, male
holotype (EC) South Africa.
Comment
This genus was not available for examination during this study, due to
the fact that, it is a rare specimen and the museum was unwilling to loan then
out.
Anthidiellum (Pycnanthidium) micheneri Pauly
Anthidiellum micheneri Pauly [In] Pauly et al., 2001: 226, 228-229, female
holotype (SEMC) Madagascar.

180
Comment
This genus was not available for examination during this study, due to
the fact that, it is a rare specimen and the museum was unwilling to loan then
out.
Anthidiellum (Pycnanthidium) orichalciscopatum (Strand)
Anthidium orichalciscopatum Strand, 1912b: 307-308, female holotype
(ZMHB) Equatorial Guinea.
Anthidiellum orichalciscopatum (Strand): Mavromoustakis, 1936a: 44.
Anthidiellum (Pygnanthidiellum) orichalciscopatum (Strand): Pasteels, 1984:
99.
Diagnosis
Male: Unknown. Female. Length: head 1.9-2.2 mm; scutum and
scutellum 1.8-2.2 mm; forewing 4.5-5.0 mm; body 5.6-5.8 mm. Vestiture:
white, except brownish on tarsi; sparse to impubescent, moderate around
antennal socket and on mesopleuron; tibiae without thick, stout setae.
Integument: black, except extensive yellow maculation on most of lower half
of face, a spot on dorso-lateral margin of preoccipital area (Figure 31a), lateral
regions of scutum, axilla, posterior margin of scutellum, dorsal surface of fore
tibia, base of dorsal surface of middle tibia, hind mostly black (Plate 28),
meso-lateral regions of T1-T2, dorso-lateral and lateral regions of T3, short
continuous cross-bands on T4-T5 (Figure 31b; Plate 27a); mandible with large
punctures basally, small punctures apically, shiny between punctures, two

181
carinae (outer and inner carinae), two, similar, large outer teeth and two,
similarly small teeth; lower clypeal margin with four small papillae (two on
each side); lateral margin of scutum, in dorsal view, rounded; scuto-scutellar
suture open, forming shiny fovea; scutellum, in dorsal view, emarginate
medio-posteriorly, hides median region of propodeum; 12 hamuli on hind
wing; vein cu-v slants; hind basitarsus broad, nearly as wide as hind tibia.
Distribution
This species is widely distributed from central Africa to the southern
Cape of the coast of Africa.
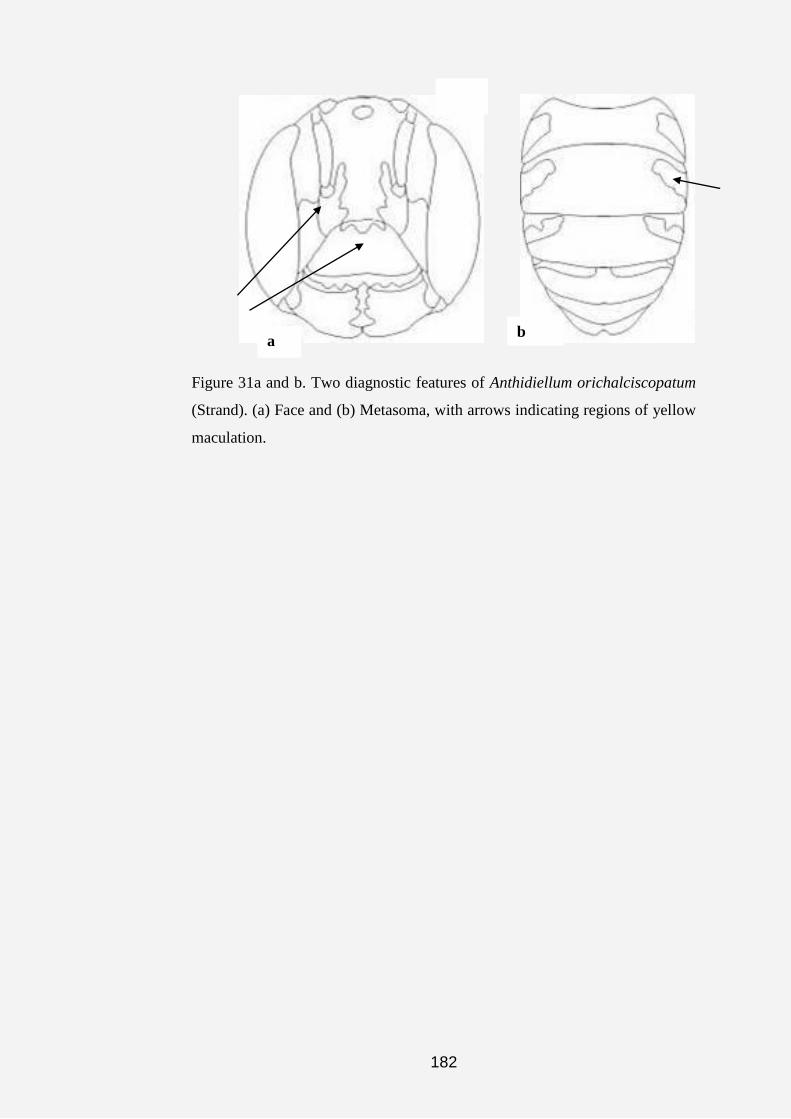
182
Figure 31a and b. Two diagnostic features of Anthidiellum orichalciscopatum
(Strand). (a) Face and (b) Metasoma, with arrows indicating regions of yellow
maculation.
a b
Fig.
72

183
Plate 27a, b and c. Three species of genus Anthidiellum. (a) Anthidiellum
orichalciscopatum (Strand). (b) Anthidiellum otavicum (Cockerell). (c)
Anthidiellum polyochrum Mavromoustakis.
Plate 28. Hind leg of Anthidiellum orichalciscopatum (Strand), showing
reduced integumental markings.
a
b
28
c

184
Anthidiellum (Pycnanthidium) otavicum (Cockerell).
Dianthidium otavicum Cockerell, 1936c: 452-453, male holotype (NHML)
Namibia.
Anthidiellum (Pygnanthidiellum) otavicum (Cockerell): Pasteels, 1984: 96,
101, 103.
Diagnosis
Female. Length: head 1.5-1.6 mm; scutum and scutellum: 1.8-2.0 mm;
forewing 3.5-3.8 mm; body 4.5-4.7 mm. Vestiture: white, except black to
reddish on fore tibia; sparse; tibiae without thick stout setae. Integument:
black, except for yellow maculations on lateral and medio-dorsal regions of
clypeus, ventral region of paraocular area (Figure 32a), meso-lateral third of
T3-T5 (yellow spots on T3 smaller than on T4-T5, Plate 27b); mandible with
large punctures basally, small punctures apically, shiny between punctures,
with outer carina only, two, similar, large outer teeth and two, similarly small
teeth; lower clypeal margin with four small projections (two on each side
(Figure 32a)); lateral region of scutum rounded; scuto-scutellar suture open,
forming shiny fovea; scutellum weakly emarginate medio-posteriorly; 10
small hamuli on hind wing, vein cu-v straight; hind basitarsus broad, as wide
as base of hind tibia.
Male. Length: head 1.5-1.7 mm; scutum and scutellum: 1.6-1.7 mm;
forewing 3.7-4.0 mm; body 5.0-5.4 mm. Vestiture and integument: similar to
female, except gender limited characters and S3-S5 with short, distal fringe;
S5 with a lateral comb, with about 20 similar setae, seperated narrowly by
Fig.
72

185
medial notch (Figure 4b); clypeus entirely yellow, ventro-lateral regions of
supraclypeus, mandible yellow with apex black; ventral surfaces of fore and
middle femora; dorsal surfaces of fore, middle and hind tibiae, dorsal surfaces
of basitarsi, lateral regions of T1-T3; cross band on T4 interrupted medially by
narrow blackish area; continuous cross band; T5-T7 mostly yellow; apex of
mandible half as wide as female, teeth very small; S5 with a small medial
notch; gonostylus expanded apically, margin with reduced notch; penis valve
narrowly seperated (Figure 32b).
Distribution
Anthidiellum otavicum (Cockerell) is endemic to south- western
Africa.

186
A. otavicum A. otavicum
Figure 32. Diagnostic features of Anthidiellum otavicum (Cockerell). (a) Face
and (b) Male genitalia, arrows indicate regions of yellow markings.
A. polyogrum
Figure 33. Hind leg of Anthidiellum polyochrum Mavromoustakis (arrows
showing regions of yellow markings)
a b
33

187
Anthidiellum (Pycnanthidium) polyochrum Mavromoustakis
Anthidiellum polyochrum Mavromoustakis, 1937b: 266-267, female holotype
(SAMC)
Anthidiellum (Pygnanthidiellum) polyochrum Mavromoustakis: Pasteels,
1984: 97, 102-103, 105.
Diagnosis
Female. Length: head 1.5-1.6 mm; scutum and scutellum 1.5-1.7 mm;
forewing 3.4-3.5 mm; body 3.8-4.2mm. Vestiture: white; sparse to
impubescent, except moderately dense around antennal socket and on tarsi;
tibiae without thick stout setae. Integument: mostly black, except yellow
maculation on most of lower face, basal region of mandible, dorsal region of
preoccipital area, dorsal surfaces of fore tibia and basitarsus, basal part of
dorsal surfaces of middle, hind tibia and most of hind basitarsus (Figure 33),
meso-lateral regions of T1-T2, T3 with small lateral spot and larger dorso-
lateral spot, T4-T5 with dorso-lateral spot, T6 mostly yellow (Plate 27c);
mandible with large punctures basally, shiny between punctures, outer and
inner carinae, 2 large outer tooth, 2 similar, small inner teeth; lower clypeal
margin with 3-5 projections, one median, one-two lateral, lateral margin of
scutum, in dorsal view, rounded; scuto-scutellar suture open, forming shiny
fovea; scutellum, in dorsal view, emarginate medio-posteriorly, hides median
region of propodeum; 11 hamuli on hind wing; vein cu-v slants; hind
basitarsus broad, subequal to width of hind tibia.

188
Male. Length: head 1.3 mm; scutum 1.6 mm; forewing 2.5 mm; body
3.5 mm. Vestiture and integument: similar to female, except for sex limited
characters and S5 with lateral comb with 30 similar setae; apex of mandible
half as wide as in female, teeth very small.
Distribution
Anthidiellum polyochrum Mavromoustakis is endemic to Namibia.
Anthidiellum (Pycnanthidium) spilotum (Cockerell)
Dianthidium spilotum Cockerell, 1920: 299, female holotype (NHML) South
Africa; Cockerell, 1936a: 1, 6.
Anthidiellum (Pyganthidiellum) spilotum (Cockerell): Pasteels, 1984: 97, 106-
108.
Anthidium capuzinum Friese, 1925: 510-511; Pasteels, 1984: 106, syn.
Diagnosis
Female. Length: head 1.7-1.9 mm; scutum and scutellum 1.8-2.1 mm;
forewing 4.3-4.9 mm; body 5.6-5.9 mm. Vestiture: white, except reddish-
black on dorsal surface of tibiae; sparse to impubescent; thick stout setae
absent on tibiae. Integument: black, tegula and legs dark brown to black,
yellow maculation on ventro-lateral region of clypeus, basal region of
mandible (Figure 34a), most of paraocular area, dorso-lateral region of
preoccipital area, lateral region of scutellum, most of dorsal surfaces of fore,
middle and hind femora, antero-dorsal region of fore and middle tibiae,

189
proximal and distal ends of hind tibia, entire dorsal surfaces of fore and hind
basitarsus (Figure 34g; Plate 30a), meso-lateral regions of T1-T5 (narrow on
T1-T2, wide on T3-T5), medial region of T6 (Figure 34a; Plate 29); mandible
with large punctures basally, small punctures apically, shiny between
punctures, two carinae (outer and inner carinae), four distinct teeth (outer teeth
largest, third and fourth progressively smaller Figure 34a); lower clypeal
margin with six projections, one median, two on each side; lateral region of
scutum, in dorsal view, rounded; scuto-scutellar suture open, forming shiny
fovea; scutellum, in dorsal view, emarginate medio-posteriorly, hides median
region of propodeum; 11 long hamuli on hind wing; vein cu-v slants; hind
basitarsus broad, more than half as wide as hind tibia.
Male.Length: head 1.5-1.6 mm; mesosoma 1.5-1.7 mm; forewing 4.4-
4.9 mm; body 4.8-5.6 mm. Vestiture and integument: similar to female, except
gender limited characters and S2-S4 with short fringes; S5 with a lateral comb
with about 10 similar setae (latertal combs widely seperated medially;
scutellum and axilla entirely black, legs brownish, yellow maculation on most
of lower half of face (without black ventral margin), most of mandible (black
apically (Figure 34b)), anterior and distal regions of femora, basal regions of
fore and middle tibiae, proximal and distal regions of hind tibia (Figure 34f),
entire surface tarsi, wider on dorso-lateral region of T4, entire anterior region
of T6 (Figure 34d); apex of mandible half as wide, teeth very small;
gonostylus expanded apically, slightly notched on outer margin; penis valves
widely separated (Figure 34e).
190
Distribution
Anthidiellum spilotum Cockerell is endemic to south-western and
southern Africa.
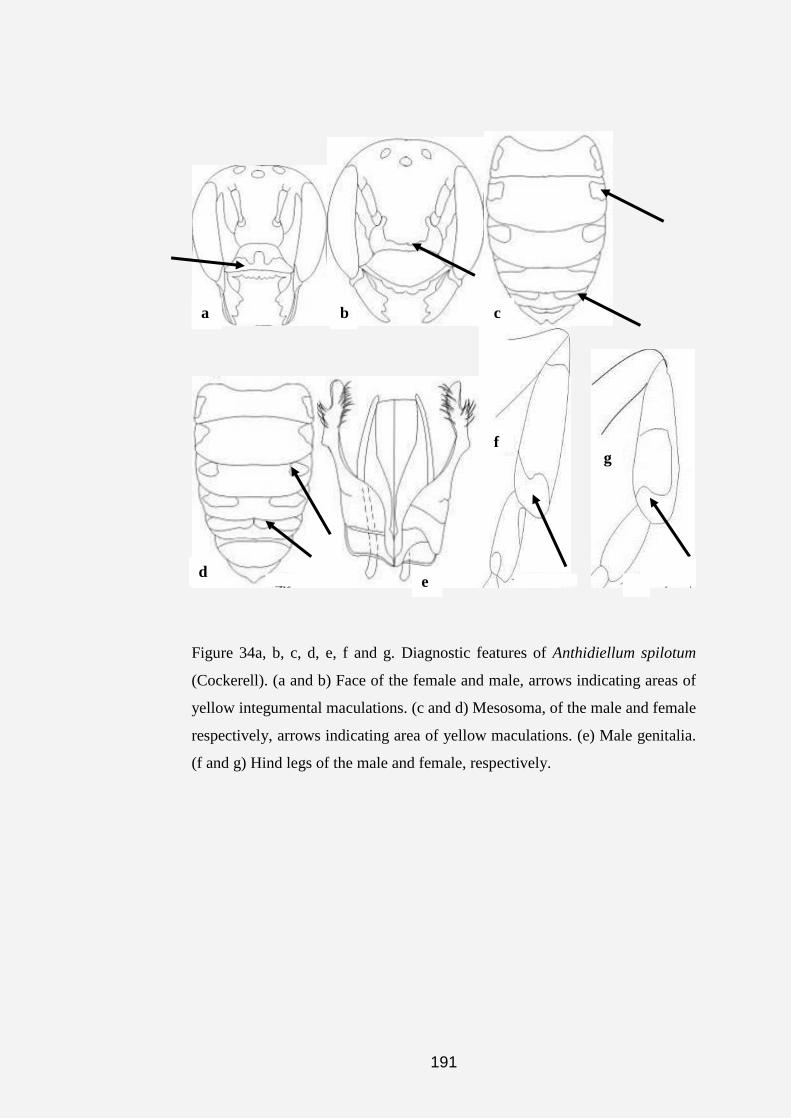
191
Figure 34a, b, c, d, e, f and g. Diagnostic features of Anthidiellum spilotum
(Cockerell). (a and b) Face of the female and male, arrows indicating areas of
yellow integumental maculations. (c and d) Mesosoma, of the male and female
respectively, arrows indicating area of yellow maculations. (e) Male genitalia.
(f and g) Hind legs of the male and female, respectively.
Fig
.
78
a b c
g
d e
f
F
i
g
.
7
8
Fig. 81
Fig.
81 F
i
g
.
8
1

192
Plate 29. Female Anthidiellum spilotum (Cockerell).
Plate 30a and b. Two diagnostic features of genus Anthidiellum. (a) Hind leg
of A. spilotum (Cockerell), arrows indicating regions of yellow maculations.
(b) Male genitalia of Anthidiellum sternale Pasteels.
a
b
29

193
Anthidiellum (Pycnanthidium) sternale Pasteels
Anthidiellum (Pygnanthidiellum[!]) sternale Pasteels, 1984: 96, 104-105, 155,
male holotype (TMSA) South Africa.
Diagnosis
Female. Length: head 1.7 mm; scutum and scutellum 1.8 mm;
forewing 4.4 mm; body 5.9 mm. Vestiture: white; sparse to impubescent; thick
stout setae on tibiae. Integument: black, antennal flagella and tegula dark
brownish, except yellow maculation on most of clypeus (dorsal and ventral
margins black), lateral region of supraclypeal area, lower half of paraocular
area (Figure 35a), dorsal region of preoccipital area, most of surfaces of
femora, entire tibiae and tarsi (Figure 35g), meso-lateral regions of T1-T2,
dorso-lateral regions of T3-T4 (mediolongitudinal black areas broad on T3,
narrow on T4), entire T5-T6 (Figure 35c); mandible with large punctures
basally, small punctures apically, shiny between punctures, with two carinae
(outer and inner carinae), two, similar, large outer teeth and two, similarly
small teeth; lower clypeal margin with four projections (two on each side);
lateral region of sutum, in dorsal view, rounded; scuto-scutellar suture open,
forming shiny fovea; scutellum, in dorsal view, emarginate medio-posteriorly,
hides median region of propodeum; 11 long hamuli on hind wing; vein cu-v
slants; hind basitarsus broad, more than half as wide as hind tibia.
Male. Length: head 1.5-1.7 mm; scutum and scutellum 1.7-1.8 mm;
forewing 3.5-4.3 mm; body 5.0-5.6 mm. Vestiture and integument: similar to
female, except sex limited character and S5 with lateral comb of about 12

194
similar setae (combs widely seperated medially); apex of mandible half as
wide as in female, 3 very small teeth (Figure 35b), medial region of hind tibia
(Figure 35f); lateral region of T5 (Figure 35d); gonostylus slender apically,
deeply notched on outer margin; penis valves slightly wide apically, narrowly
separated (Figure 35e; Plate 30b).
Distribution
This species is endemic to South Africa.
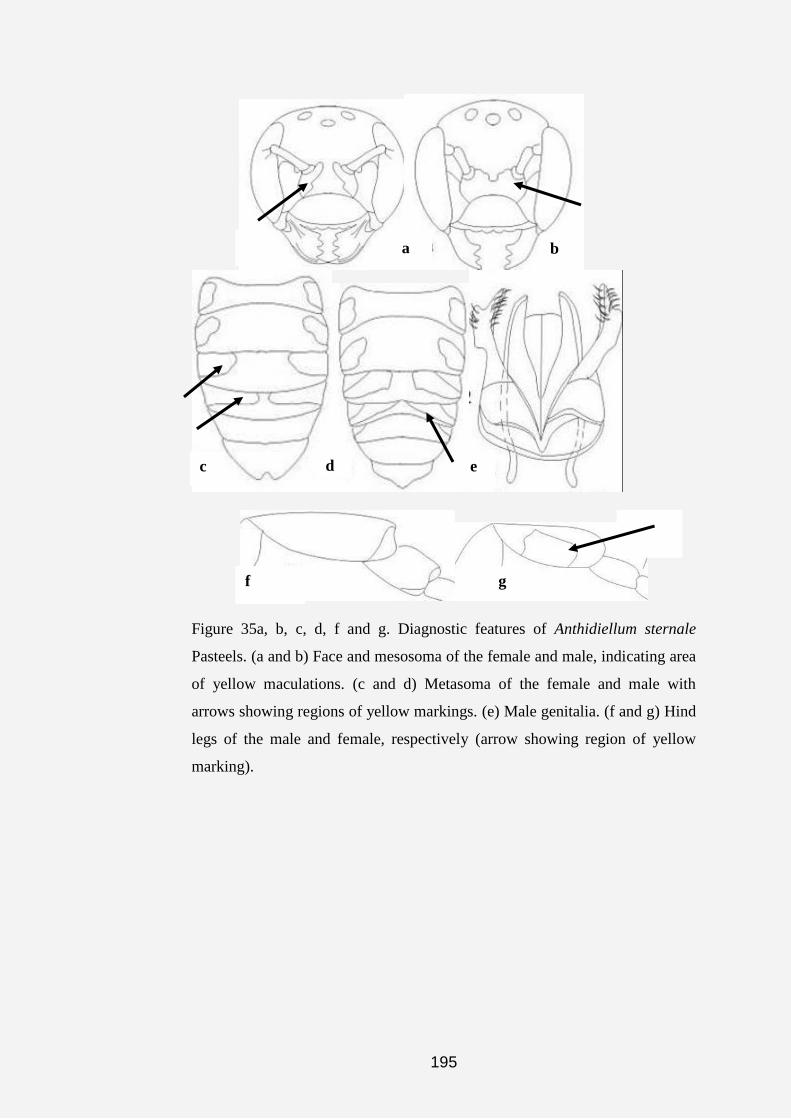
195
Figure 35a, b, c, d, f and g. Diagnostic features of Anthidiellum sternale
Pasteels. (a and b) Face and mesosoma of the female and male, indicating area
of yellow maculations. (c and d) Metasoma of the female and male with
arrows showing regions of yellow markings. (e) Male genitalia. (f and g) Hind
legs of the male and female, respectively (arrow showing region of yellow
marking).
e
a a b
c d
f
F
i
g
.
7
6
Fig. 76
g

196
Anthidiellum (Pycnanthidium) tegwaniense (Cockerell)
Dianthidium tegwaniense Cockerell, 1914a: 278, female holotype (NHML)
South Africa.
Anthidiellum tegwaniense (Cockerell): Cockerell, 1932c: 524; Cockerell,
1936c: 455; Mavromoustakis, 1936a: 46.
Anthidiellum (Pygnanthidiellum) tegwaniense (Cockerell): Pasteels, 1984:
107-108.
Anthidiellum swalei Mavromoustakis, 1936b: 603-605, female holotype
(NHML) Zimbabwe; Pasteels, 1984: 107-108, syn.
Pygnanthidiellum (Pygnanthidiellum) swalei (Mavromoustakis):
Mavromoustakis, 1963b: 492-493.
Diagnosis
Female. Length: head 1.6-1.7 mm; scutum and scutellum 1.8-2.0 mm;
forewing 4.3-4.6 mm; body 5.4-5.8 mm. Vestiture: white; sparse to
impubescent; tibiae without thick stout setae. Integument: black, antenna,
tegula and legs dark brown, yellow maculations on most of lower half of face
(ventral margin of clypeus dark brown), basal region of mandible (apex
black), dorso-lateral region of preoccipital area, lateral regions of scutum and
axilla, dorso-lateral region of scutellum, entire legs (Figure 36d), meso-lateral
regions of T1-T4 (smaller on T1-T2, wider on dorso-lateral regions of T3-T4),
entire surfaces of T5-T6 (Figure 36b); mandible with large punctures basally,
shiny between punctures, outer carina only, two, similar, large, outer teeth and
two, similarly, small teeth; lower clypeal margin with four projections (two on
each side; lateral region of

197
scutum, in dorsal view, rounded; scuto-scutellar suture open, forming shiny
fovae; scutellum, in dorsal view, emarginate medio-posteriorly, hides median
region of propodeum; 12 long hamuli on hind wing; vein cu-v straight; hind
basitarsus broad, nearly as wide as base of hind tibia.
Male. Length: head 1.5-1.8 mm; mesosoma 1.6-1.8 mm; forewing 3.5-
3.7 mm; body 4.9-5.4 mm. Vestiture and integument: similar to female, except
for gender limited characters and S5 with 30 similar setae laterally; yellow
maculation on entire clypeus, mandible (apex black, figure 36a), dorsal
surface of fore tibia, medial region of hind tibia (Figure 36c), smaller on T3,
most of T4-T7; apex of mandible half as wide, 3 very small teeth; gonostylus
narrow apically, deeply notched on outer margin; penis valves expanded
apically, widely separated (Figure 36e).
Distribution
Anthidiellum tegwaniense (Cockerell) is widespread throughout Malawi,
South Africa and Zimbabwe.
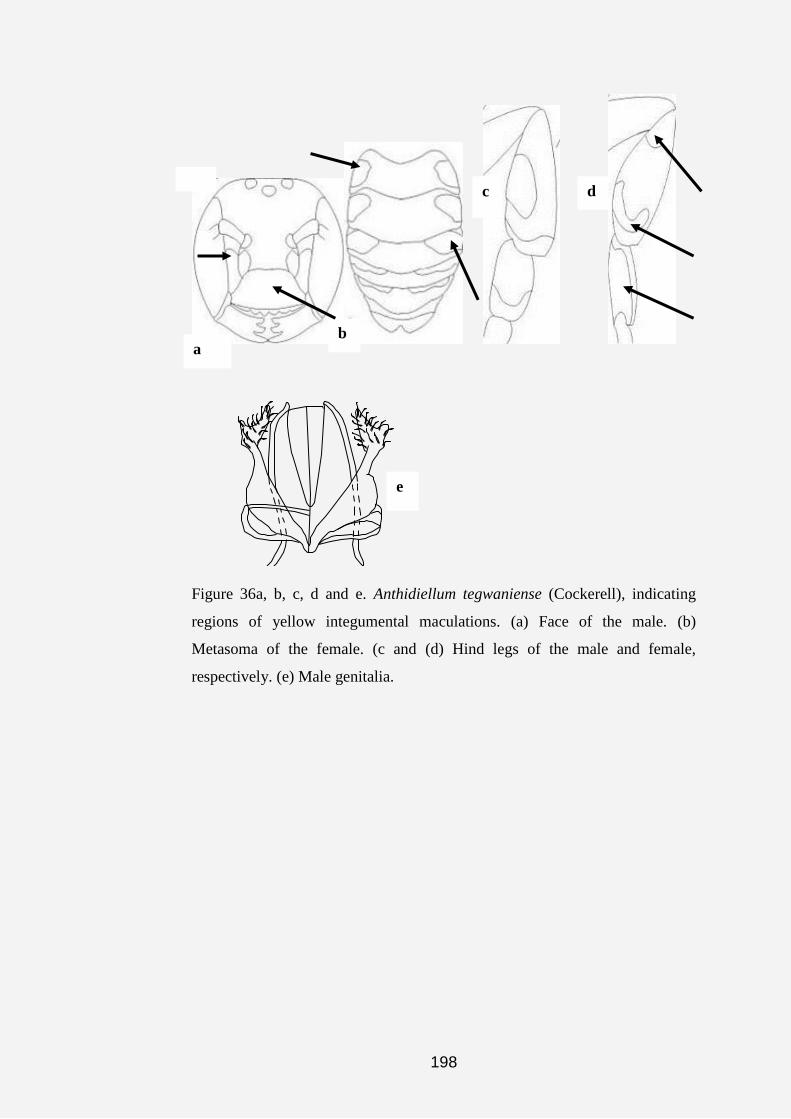
198
A. tewaniense
Figure 36a, b, c, d and e. Anthidiellum tegwaniense (Cockerell), indicating
regions of yellow integumental maculations. (a) Face of the male. (b)
Metasoma of the female. (c and (d) Hind legs of the male and female,
respectively. (e) Male genitalia.
Fig.
76
a b
c d
e

199
Anthidiellum transversale Pasteels
Anthidiellum (Pygnanthidiellum) transversale Pasteels, 1984: 104, 106, male
holotype (RMNH) Zimbabwe.
Comment
This genus was not available for examination during this study, due to
the fact that, it is a rare specimen and the museum was unwilling to loan then
out.
Anthidiellum (Pycnanthidium) zebra (Friese)
Anthidium zebra Friese, 1904b: 103, male and female syntypes (TMSA) South
Africa; Friese, 1905b: 67-68, 70, 73; Friese, 1909a: 400, 401, 402, 409,
412; Friese, 1909b: 158, 160-161; Strand, 1912a; 142; Cockerell,
1932b: 527; Cockerell, 1936a: 6; Anonymous, 1958: 33.
Dianthidium zebra (Friese): Cockerell, 1909b: 261; Cockerell, 1920b: 299;
Cockerell, 1936a: 6; Cockerell, 1936c: 453.
Anthidiellum zebra (Friese): Mavromoustakis, 1936b: 605; Mavromoustakis,
1937a: 232; Mavromoustakis, 1937b: 267;
Pygnanthidiellum (Pygnanthidiellum) zebra (Friese): Mavromoustakis, 1963a:
492-493.
Anthidiellum (Pygnanthidiellum) zebra (Friese): Pasteels, 1984: 95-99, 101,
104, 106.
Anthidiellum medionigrum Cockerell, 1933a: 464, male holotype (NHML)
South Africa; Cockerell, 1936c: 455; Mavromoustakis, 1936a: 46;
Pasteels, 1984: 97, syn.

200
Diagnosis
Female. Length: head 1.5-2.0 mm; scutum and scutellum 1.8-1.9 mm;
forewing 4.3-4.5 mm; body 3.9-4.8 mm. Vestiture: white; sparse to
impubescent; thick stout setae on tibiae. Integument: black, tegula, legs and
metasoma dark brown, yellow maculations on most of lower half of face, most
of mandible (ventral margin of clypeus dark brown, Figure 37a), dorsal region
of preoccipital area, lateral regions of scutum and axilla, dorso-lateral region
of scutellum, small on ventral and distal regions of fore and middle femur,
proximal regions dorsal surfaces of fore and middle tibia, most of dorsal
surface of hind tibia, entire hind basitarsus (Figure 37g), meso-lateral regions
of T1-T4 (yellow area increases progressively from a lateral spot on T1 to
occupying lateral third of T4), entire surfaces of T5-T6 (Figure 37c); mandible
with large punctures basally, shiny between punctures, with two carinae (outer
and inner carinae), two, dissimilar, large outer teeth and two, similarly small
teeth; lower clypeal margin with four projections, two on each side; lateral
region of scutum, in dorsal view, rounded; scuto-scutellar suture open,
forming shiny fovea; scutellum, in dorsal view, emarginate medio-posteriorly,
hides median region of propodeum; 11 small hamuli on hind wing, vein cu-v
straight; hind basitarsus broad, subequal base of hind tibia.
Male. Length: head 1.5-1.9 mm; scutum and scutellum 1.7-1.8 mm;
forewing 4.1-4.3 mm; body 4.6-5.5 mm. Vestiture and integument: similar to
female, except for sex limited characters and S2 with lateral fringe, S3-S4 with
narrow fringes, S5 with lateral comb of about 30 setae; yellow maculation on
entire dorsal surface of fore tibia, medial region of hind tibia (Figure 37f);

201
smaller on T4, most of T5-T7 mostly yellow, distal margins of T5-T6 reddish
to dark brown (Figure 37d); apex of mandible half as wide, 3 very small teeth
(Figure 37b); gonostylus slender aically, deeply notched on outer margin;
penis valves widely separated (Figure 37e).
Distribution
This species is endemic to southernmost part of Eastern and South
Africa

202
Figure 37a, b, c, d, f and g. Diagnostic features of Anthidiellum zebra (Friese).
(a and b) Face of the female and male, arrows indicating regions of yellow
integumental maculations. (c and d) Metasoma of the female and the male
respectively (arrow indicate regions of yellow markings). (e) Male genitalia. (f
and g) Hind legs of the male and female, respectively.
a b c d
F
i
g
.
7
6
e
Fig. 76
Fig. 76
f
g
Fig. 76

203
Anthidiellum (Pycnanthidium) krugerense sp. nov.
Anthidiellum (Pycnanthidium) krugerense, female holotype (SANC), South
Africa.
Description
Female: Length: head 1.9 mm; scutum and scutellum 1.8 mm; forewing 4.0
mm; body: 4.8 mm. Vestiture: white; sparse to impubescent; thick stout setae
on tibiae. Integument: black, tegula, lateral regions of axilla and scutellum
dark brown, except for yellow maculations on most of lower half of face
(ventral clypeal margin black, Figure 38a), dorso-lateral region of preoccipital
area, entire legs, meso-lateral regions of T1-T4 (yellow area increases
progressively from a lateral spot on T1 to occupying lateral third of T4), entire
surfaces of T5-T6; mandible with small, distinct punctures, shiny between
punctures, outer carina only, two large similar outer teeth and two small
dissimlar inner teeth; lower clypeal margin with four projections, two on each
side; lateral margin of scutum, in dorsal view, extend longitudinally concave,
groove separate extended surface with carina next to tegula (Figure 38b);
scuto-scutellar suture open, forming shiny fovea; scutellum, in dorsal view,
emarginate medio-posteriorly, hides median region of propodeum; 12 long
hamuli on hind wing; vein cu-v straight; hind basitarsus broad, nearly as wide
as base of hind tibia.
Distribution
This species is endemic to South Africa.

204
Figure 38a and b. Descriptive features of Anthidiellum krugerense sp. nov. (a)
Face, arrows showing region of yellow markings. (b) Scutum, arrow
indicating the carina on the lateral region of scutum extending beyond axilla
margin.
b a

205
CHAPTER FIVE
DISCUSSION
Phylogenetic analysis of the tribe Anthidiini
The tribe Lithurgiini is reported as the sister group of the Osmiini,
Anthidiini, Megachiliini and Dioxyini which constitute the Megachilinae, with
the tribe Megachilini known to the closest sister group of the Anthidiini (Riog-
Alsina and Michener, 1993). These authors adequately demonstrated the
Megachilinae to be monophyletic by the presence of several unique
synapormorphies. However, they did not demonstrate the monophyly of the
Anthidiini using most of the genera of the tribe. The inclusion of Lithurge
pullatus (Vachal) and Megachile semiflava (Cockerell) in this study as two
outgroups clearly confirmed the monophyly of the Anthidiini. This is
supported by nine synapormorphies, seven of which are non-homoplasious
namely, (a) Lateral region of preocciptal area of female carinate (level of
sclerotization variable. (b) Distance between antennal sockets at least subequal
to two antennal sockets diameters. (c) Omaulus carinate (sometimes entirely)
(d) Scutellum, in dorsal view, obscures propodeum, at least medially. (e)
Vestiture on dorsal surface of hind basitarsus dense. (f) Second recurrent vein,
usually extends distally behind second transverse cubital vein. (g) Jugal lobe
short, less than half length of vannal lobe. (i) Anterior region of T1 without
fovea. (j) Metasoma without subapical hair bands. Characters (d) and (e) show

206
reversals in higher tree topology and therefore probably do not strictly
characterize the Anthidiini as a whole. The Anthidiini is clearly supported by
several unambiguous synapormorphies and by very high bootstrap and
jackknife values of 96% and 97% respectively, hence confirms the tribe as a
monophyletic taxon.
The second major island comprises relatively small genera, namely
Aspidosmia, Afranthidium, Anthidioma, Neanthidium, with very high
boothstrap and jackknife valves of 86% and 85% in the strict consensus trees.
Aspidosmia forms the first major clade within this island. Its monophyly is
demonstrated by an autoapormorphy, that is, the dorsal region of the female
clypeus being highly elevated. This uniquely derived autoapomorphy is
supported by another character state namely, second recurrent vein of the
forewing entering the second submarginal cell. The clade therefore supports
the relationship of genus Aspidosmia to the rest of the anthidine bees which
confirms previous classification works including Peter (1972), Riog-Alsina
and Michener (1993), Michener and Griswold (1994) and Michener (2000).
The genera Afranthidium, Anthidioma and Neanthidium formed the next sub-
clade within this island. The monophyly of this clade was demonstrated by
two synapomorphies: apex of marginal cell mostly rounded and the first
recurrent vein of the fore wing meeting the first transverse cubital vein distally
or entering the first submarginal cell. The high evidential support of this clade,
gives an indication of a very close relationship between these three genera.
However, in the absence of the male specimen of Anthidioma, examination of
the male genitalia could not be done to confirm the ascertion by Michener and

207
Griswold (1994) and Michener (2000) that this genus should be placed as a
subgenus of Afranthidium. Hence Afranthidium and Anthidioma have been
retained as separate genera. In addition, due to the considerable variations in
the number of spines on T6 and T7 of male, structure of the male genitalia and
in the mandibular dentition of females of Afranthidium and Neanthidium, the
two genera were also retained as separate anthidine taxa as had been
documented in Michener (2000).
Two large groups constitute the remaining major islands within the
Anthidiini. One of these comprises three parasitic genera and nine non-
parasitic. The parasitic genera include Afrostelis, Euaspis and Hoplostelis
while the non-parasitic genera comprise Anthidioctes, Hypanthidiodes,
Cyphanthidium, Paranthidium, Eoanthidium, Epanthidium, Serapista,
Duckeanthidium, Apianthidium and Aztecanthidium. This island is clearly
supported by high bootstrap and jackknife values of 83% and 82%
respectively and gives an indication that these genera form a close sister
group. In addition, two synapomorphies including a non-homplasious
character also support this island namely, the posterior surface of the antennal
scape being sparsely pubescent and the facial vestiture also being sparse to
impubescent. The latter reversed in higher tree topology, probably due to the
fact that, that character is commonly shared by other genera. In the analysis of
this island of trees, the clade [Anthidioctes, Hypanthidiodes, Afrostelis and
Hoplostelis] is not defined by any unique synapomorphy. However, Afrostelis
and Hoplostelis are shown clearly as monophyletic, defined primarily by the
elongated scutum and scuto-axilla suture slightly opening to form a fovea as

208
well as absence of a scopa of the female. These parasitic genera supported by
low bootstrap and jackknife values (51/ 50%), clearly show little support of
this group. On the other hand, Anthidioctes and Hypanthidiodes are defined by
high bootstrap and jackknife values (78/ 78%), hence have high evidential
support of this group. Synonymization of the Neotropic Anthidioctes with
Neotropic Hypanthidiodes is suggested in this present study. Cyphanthidium
forms the third sub-clade of this island. Inspite of its high bootstrap and
jackknife values (83/ 86%), the genus however is not defined by any
autapomorphy. Thus, in this study, the monophyly of this genus was not
established. The monophyly of genus Paranthidium, which constitute the fifth
sub- clade was not demonstrated in this study because of the absence of an
autapomorphy. However, the combination of two homoplasious character
states defined this clade, i.e. a lower clypeal margin with projections and a
slight notch in the scutellum postero-medially. This clade as sister group is
supported by very high bootstrap and jackknife values (91/ 90%). The clade
Euaspis, Eoanthidium and Epanthidium forms the sixth sub-clade within this
island. The parasitic genus Euaspis, is strongly supported by bootstrap and
jackknife values (90/ 90%) and is characterized primarily by the scutellum, in
dorsal view, being deeply notched postero-medially. Inspite of the presence of
two homoplasious synapomorpies in defining genus Eoanthidium as
monophyletic, it forms polytomy with Neotropic genus Epanthidium which is
not characterized by an autoapomorphy. These two genera are weakly
supported evidentially by low bootstrap and jackknife values 51/ 50%.
Inadequate information could probably account for the poor resolution of the

209
clade. The monophyly of genus Serapista is strongly supported by a single
homoplasious apomorphy inwhich the posterior region of the axilla has a
spine. The high bootstrap and jackknife values (98/ 98 %) support the clade
that includes Serapista and three other genera namely, Duckeanthidium,
Apianthidium and Aztecanthidium. The clade formed by these three genera is
primarily defined by a single synapomorphy inwhich the apex of the outer
hind tibia spur is strongly curved. Furthermore, a high bootstrap and jackknife
values 87/ 85% evidentially support this clade. The monophyly of
Duckeanthidium was defined by one homoplasious character state, i.e.
vestiture on the dorsal surfaces of tibiae and basitarsi is densely pubescent. In
addition, Apianthidium was defined by a single apomorphy inwhich the
second recurrent vein of the fore wing enters the second submarginal cell.
Aztecanthidium on the other hand was not characterized by any derived
character state, hence its monophyly was not established in this study.
The remaining major island of the Anthidiini comprised Anthidium,
Dianthidium, Anthidiellum, Bathanthidium, Icteranthidium, Gnathanthidium,
Pachyanthidium, Pseudoanthidium, Acedanthidium, Benanthis, Hypanthidium,
Indanthidium, Notanthidium, Pleisianthidium, Rhodanthidium and Trachusa.
This island is supported by high bootstrap and jackknife values (80/ 79 %) and
is also defined by a combination of two characters namely, the second cubital
cell of the fore wing is more than half or subequal length to the length of the
first cubital cell and also, vein Cu-v of the hind wing is slanted. The genus
Anthidium forms a clade within this island and is seperated from the rest by
two synapomorphies: (a) dorsal region of preoccipital area, in female carinate

210
and b) facial vestiture sparse to impubescence. High bootstrap and jackknife
values of 82/ 82% clearly support this clade. Anthidium was establisted as
monophyletic, defined by a single homoplasious character state in which the
second recurrent vein meets the second transverse cubital vein distally. The
next sub-clade within that island consists of the genera Dianthidium,
Anthidiellum and Bathanthidium. This clade is not defined by any
synapomorphy, although it is highly supported by bootstrap and jackknife
values of 63/ 63 %. The monophyly of Dianthidium is defined primarily the
female having three mandibular teeth. Anthidiellum and Bathanthidium
formed a subclade strongly supported by very high bootstrap and jackknife
(85/ 84%) and defined by two homoplasious synapomorphies, i.e. a curved
subantennal suture and foveate scuto-scutellar suture. In the absence of male
characters of Bathanthidium in this study, synonymization the two taxa are in
this study was not considered as was suggested in Michener (2000). The
monophyly of Anthidiellum was demonstrated by two homoplasious
apomorphies, i.e. projections on the ventral margin of the clypeus and a
slighty notch on the scutellum postero-medially. A single non-homoplasious
synapomorphy separated this clade from the rest, i.e. posterior margin of the
axilla extending beyond the lateral margin of the scutum. The genera
Gnathanthidium, Pachyanthidium and Pseudoanthidium formed the next sub-
clade, and were characteristically defined by a single non-homoplasious
synapomorphy in which the preoccipital area is lamellate. Low bootstrap and
jackknife values (50/ 50%) weakly supported this clade to be considered as
sister group. However, the subclade Gnathanthidium, Pachyanthidium and

211
Pseudoanthidium was clearly supported by high bootstrap and jackknife
values (83/ 85%, and 79/ 79%). All three genera are defined by at least one
apomorphy, which demonstrates their monophyletic status. Icteranthidium
was however not defined by any uniquely derived character hence its
monophyly was not confirmed in this present study. This clade was separated
from the remaining clade by one non-homoplasious character state and two
homoplasious character states, as well as, high bootstrap and jackknife values
(81/ 80%) supporting this clade as sister group. The remaining clade which is
weakly supported by bootstrap and jackknife values (63/ 65%) had several
synapomorphies defining the monophyly of the genera. This clade comprises
Acedanthidium, Benanthis, Hypanthidium, Indanthidium, Notanthidium,
Pleisianthidium, Rhodanthidium and Trachusa. The collapse of several nodes
within this clade could probably be due to inadequate characters to clearly
resolve the clade. The genera Rhodanthidium, Hypanthidium and
Pleisianthidium were not supported by any autoapomorphy to ascertain their
monophyly. However, these genera differ significantly in the structure of the
male genitalia, hence, could be maintained as separate taxa.
In the analysis of the genus Anthidiellum, the first clade consists of
Anthidiellum flavescens, A. eritrinum and A. rubellum. The clade is
evidentially supported by very high bootstrap and jackknife values (94/ 95%)
and three synapomorphies namely, the distance between antennal sockets
subequal or less than two antennal socket diameters and a preoccipital carina
occurring dorso-laterally or laterally, as well as a distally lamellate pronotal
lobe. However, the two species of the nominative subgenus formed a sub-

212
clade and was highly supported by bootstrap and jackknife values of 73/ 73%.
Hence, this study clearly confirms the two clades as separate subgenera
(Anthidiellum and Chloranthidiellum) within the genus as in Michener (2000).
The second major clade is made of A. absonulum, A. apicatum, A.
auriscopatum, A. frontoreticulatum, A. kimberleyana, A. polyochrum, A.
krugerense, A. sternale, A. tegwaniense, A. zebra, A. bulawayense, A.
otavicum, A. bipectinatum and A. spilotum. The clade is supported by very
high bootstrap and jackknife values (92/ 89%), and two synapomorphies
inwhich the length of the subantennal suture is more than half the length of the
clypeus and mandibles having small punctures apically and large punctures
basally. Several nodes collapsed to form polytomies within this clade. The
reason for this could be due to the inavailability of several specimens of most
taxa to adequately resolve the cladogram.
In spite of a number of missing genera and character states during the
analysis of the tribe, at least the following classification, based on available
information, is postulated for the tribe. Most of the generic classification of
Michener (2000) is retained, except that, three main subtribes are erected
within the tribe. The genus Hypanthidiodes is synonymized with Anthidioctes
(the latter being the older available name). In addition, synonymization of the
genus Bathanthidium with genus Anthidiellum was considered, however, they
were retained as separate taxa. The tribe Anthidiini may therefore be placed
under family Megachilidae and subfamily Megachilinae with three subtribes,
namely Afranthidina, Anthidina and Euaspina. The subtribe Afranthidina
comprises non-parasitic, small to medium size, megachiliform to

213
anthophoriform and hairy bees which are all endemic to the African continent.
The four genera that occur in this subtribe include Afranthidium, Anthidioma,
Aspidosmia and Neanthidium. The Anthidina comprises mainly non-parasitic
bees with wide morphological variation. Its size ranges from medium to large
and varies in shape from hoplitiform, megachiliform, chalicodomiform to
anthophoriform. It is the only subtribe with three cosmopolitan genera
(Anthidium, Anthidiellum and Trachusa) while the other genera
(Acedanthidium, Bathanthidium, Benanthis, Dianthidium, Gnathanthidium,
Hypanthidium, Icteranthidium, Indanthidium, Notanthidium, Pachyanthidium,
Pleisianthidium and Pseudoanthidium) are known to have narrow range of
distribution. The subtribe Euaspina constitute both parasitic and non-parasitic
genera, just as in Anthidina, it varies from small to large size hoplitiform to
anthophoriform bees, as well as small heriadiform bees. The subtribe is made
up of genera with limited geographic distribution among the continents. These
genera include, Anthidioctes, Afrostelis, Apianthidium, Aztecanthidium,
Cyphanthidium, Duckeanthidium, Eoanthidium, Epanthidium, Euaspis,
Hoplostelis and Serapista. The classification of the Anthidiini into three
subtribes enormously facilitates its study and provides useful information on
these genera. The name of each subtribe was derived from the oldest available
name within the genera.
Biogeography and Evolution of Anthidiini
Based on the results of the present analyses, three main lineages of
Anthidiini are distinguished. The first lineage comprises a relatively small
clade of mostly Afrotropical genera (Afranthidium, Anthidioma and

214
Aspidosmia), and a Palaearctic genus Neanthidium (Algeria and Morocco).
The second lineage is made up of a distinctly large clade with three
cosmopolitan genera (Anthidium, Anthidiellum and Trachusa), three genera
endemic to the new world (Neotropic and Nearctic- Dianthidium,
Hypanthidium and Notanthidium), and ten genera occurring in the old world
(Afrotropical and Palaearctic: Icteranthidium, Pachyanthidium,
Gnathanthidium, Benanthis; Palaearctic and Orient: Rhodanthidium,
Bathanthidium, Acedanthidium, Indanthidium; Pleisanthidium is widespread
throughout the old world). The third clade also comprises a relatively large
clade of the remaining genera, with endemic biogeographical distribution;
eight genera of the new world (Neotropic and Nearctic: Anthidioctes,
Hoplostelis, Paranthidium, Aztecanthidium, Duckeanthidium), two
Afrotropical genera (Afrostelis, Cyphanthidium), one Oriental genus,
Apianthidium, and three genera with relatively extensive distribution within
the old world (Euaspis, Eoanthidium, Serapista).
These biogeographical data at the subtribe and generic levels were
used to obtain information about probable evolutionary processes and events
that might have taken place within the Anthidiini. The Afrotropical and
Oriental regions exhibit high bee richness within the Anthidiini at the generic
level, with 16 and 13 genera, respectively. One subtribe is limited to the Africa
continent. High bee richness at the generic level and basal groups of primitive
forms are enough evidence to postulate that, the origin and center of radiation
for the Anthidiini probably occurred in southern Pantropic Region (Southern
Africa to South East Asia). These geographic locations, before the split of the

215
world’s continent, formed South-eastern block of the southern hemisphere,
super-continent Gondwanaland (during the early Cretaceous over 100 million
years before present, Raven and Axelrod, 1974).
The discovery of the oldest fossil bee, Melittosphex burmensis
(Melittosphecidae) (Pionar and Danforth, 2006), from the early Cretaceous
Burmese amber (~100 million years ago) which is 35-45 million years older
than the previously known fossil bee, Cretotrigona prisca by Michener and
Grimaldi, 1988 a & b), tend to suggest that the evolution of bees might have
occurred earlier than previously indicated, possibly, as early as during the
evolution and diversification of angiosperms in early Cretaceous. M.
burmensis shares several characters with most members of present day
anthidine genera. Visual observation clearly shows that the forewing of this
bee possess strong first and second transverse cubital veins that forms two
complete submarginal cells. Further to this, this fossil resin-collecting bee has
a heriadiform shape which is typical of small size anthidine bees. Hence, it is
probable to also assume that the present day anthidine genera existed about the
same era as M. burmensis.
It is also known that, it was only in the tertiary (66.4 to 1.6 million
years ago) during the late Paleocene and early Eocene era that bilaterally
symmetrical organization of floral organs coevolved with animal behaviour
including bee sociality (Danforth, 2004; Dilcher, 2006). Based on the
discovery of the new fossil bee (after publication of Engel and Perkovsky,
2006), and data from host-plant specialization in western anthidine bees by
Müller (1996), as well as information of the evolution and biogeography of

216
bees, it is probable to assume that the basal lineages of Anthidiini happened
much earlier, possibly arose during the early Cretaceous, a period when the
continental connection of Africa and South-eastern Asia still existed. Further,
it is however possible that, more diversification within the tribe took place
much later (Eocene-Oligocene period), especially of genus Anthidium,
accounting for some of the fossils recorded in Engel and Perkovsky (2006).
A second and independent migration to the Neotropical region by the
probable ancestors of current Neotropic genera is conceivable. Such an event
probable took place when the gap between the drifted continents of South
America and Africa was narrow enough to permit short insect flight along its
corridors and similar climatic conditions still existed that provides similar
forage resources for these bees.
Evolution of parasitism within the tribe Anthidiini probably followed
this scenario. The parasitic genera probably evolved ones within the lineages
of the Anthidiini. This lineage comprises mainly two Afrotropical and Oriental
genera (Afrostelis and Euaspis), and a Neotropic genus Hoplostelis.
Revision of the Afrotropical Genus Anthidiellum
In the course of this present study, examination of all the Afrotropical
species of genus Anthidiellum (except A. madli Pauly, A. mediale Pasteels, A.
micheneri Pauly and A. transversale Pasteels) revealed two undescibed species
Anthidiellum (Pycnanthidium) frontoreticulatum spec nov and Anthidiellum
(Pcynanthidium) krugerense spec nov. A. frontoreticulatum spec nov, was

217
collected by Michener in 1967 from Kenya. However, he did not describe and
published this species as spec. nov, hence the present study has provided
description on this new species. A. krugerense spec nov, was found among the
bee collections of the South Africa National Collection. The only specimen
found was collected from Kruger National Park, South Africa, hence this new
species has been named after the place of collection. A. krugerense differ from
other Anthidiellum species by having lateral margin of scutum (in dorsal view)
extended longitudinally into small carinae, with a groove separating the
extended surface with a larger carina next to the tegula.
During the revision of this genus, Anthidiellum auriscopatum was
found to differ significantly from Anthidiellum orichalciscopatum (males of
both species unknown) in the following characters. These include: A)
Extensive bright yellow maculation on face, legs and metasoma of A.
auriscopatum than in A. orichalciscopatum Antero-distal region of pronotum
gently rounded and non-lamellate, than strongly angulate and lamellate
pronotal lobe as in A. orichalciscopatum. C) Lower clypeal margin of A.
auriscopatum with six small papillae (projections), four projections on lower
clypeal margin of A. orichalscopatum. D) Hind basitarsus much short and
broader than hind tibia in A. auriscopatum, however in A. orichalciscopatum,
hind barsitarsus is long and nearly as broad as hind tibia. Based on the
variation of these characters between these species, I considered A.
auriscopatum and A. orichalciscopatum as two different species within the
subgenus Pycnanthidium Krombein.

218
In addition, Anthidiellum kimberleyana and A. spilotum which were
considered synonym by Pasteels (1984), have been re-stated into separate taxa
based on differences existing in the following characters: A) Reduced
integumental maculations on T3-T6 of A. kimberleyana than in A. spilotum. B)
Legs entirely brownish-black, lacking integumental maculations in A.
kimberleyana, which occurs in A. spilotum. C) Inner carina of mandible
distinct well developed in A. kimberleyana. D) Outer teeth of mandible appear
blunt in A. kimberleyana than in A. spilotum. E) Apical region of hind tibia
much broader in A. kimberleyana than in A. spilotum. During the revision of
the Afrotropical genus, the cladogram revealed two major clades of trees
within the genus. This apparently is not compatible with the subgeneric
classification of the genus (Michener, 2000). For the purpose of this study, the
subgeneric classification of Michener (2000) has been retained. The
Afrotropical species of genus Anthidiellum is postulated to be 17, excluding
the four species that were not available during the course of this study. A
taxonomic key to the Afrotropical species of the genus Anthidiellum has been
provided for the 17 species examined during this study.

219
CHAPTER SIX
CONCLUSIONS AND RECOMMENDATIONS
Conclusions and Recommendations
Of great interest to this current research are the possible contributions
of postulated re-classification and phylogeny of the tribe Anthidiini, as well as
providing a review of the Afrotropical species of genus Anthidiellum.
The monophyly of the Anthidiini and most of its genera were
confirmed. Further to this, three subtribes: Afranthidina, Anthidina and
Euaspina have been erected for the world’s anthidine genera. These comprise
the following: Subtribe Afranthidina—Afranthidium, Anthidioma, Aspidosmia,
Neanthidium; Subtribe Anthidina—Acedanthidium, Anthidiellum, Anthidium,
Bathanthidium, Benanthis, Dianthidium, Gnathanthidium, Hypanthidium,
Icteranthidium, Indanthidium, Notanthidium, Pachyanthidium,
Pleisianthidium, Pseudoanthidium, Trachusa; Subtribe Euaspina—
Anthidioctes, Afrostelis, Apianthidium, Aztecanthidium, Cyphanthidium,
Duckeanthidium, Eoanthidium, Epanthidium, Euaspis, Hoplostelis, Serapista.
Based on the result of the cladistic analysis and the biogeographic data
of the Anthidiini, an evolutionary scenario was postulated in which the
Anthidiini probably evolved in the early Cretaceous (over 100 million years
before present), with high diversification within the tribe occurring in the
Eocene era.

220
The Afrotropical and Oriental regions exhibit high bee richness within
the Anthidiini at the generic level, with 16 and 13 genera, respectively. One
subtribe, Afranthidina is limited to the African continent. High bee richness at
the generic level and basal groups of primitive forms are enough evidence to
postulate that, the origin and center of radiation for the Anthidiini probably
occurred in southern Pantropic Region (Southern Africa to South East Asia).
A second and independent migration to the Neotropical region by the
probable ancestors of current Neotropic genera of the Anthidiini is
conceivable.
The parasitic genera probably evolved ones within the lineages of the
Anthidiini. This lineage comprises mainly two Afrotropical and Oriental
genera (Afrostelis and Euaspis), and a Neotropic genus Hoplostelis.
Further, two new species Anthidiellum (Pcynanthidium)
frontoreticulatum sp nov and Anthidiellum (Pcynanthidium) krugerense sp
nov. have been described. Two synonyms A. auriscopatum = A.
orichalciscopatum and Anthidiellum kimberleyana = A. spilotum were re-
instated to their species status as separate taxa. A total of 17 species of
Anthidiellum were found to occur in Afrotropical Region and a taxonomic key
to these species has been provided as well.
Some gaps were identified during the course of this study that
affected analysis of the results, and these centred basically on the limited

221
number of specimens available for the analysis. It is therefore recommended
that
1. A global survey should be conducted for the world’s anthidine bees, in
order to ascertain all existing fauna, as well as increasing the numbers
of museum collections.
2. Phylogenetic studies of the Anthidiini, involving all known genera,
subgenera and possibly species, need to be conducted to accurately
confirm postulated theories in this present study.
3. Phenetic studies on these groups of closely related bees could be
conducted to establish their relationship at the molecular level.

222
REFERENCES
Alexander, B. 1990. A Cladistic analysis of the nomadine bees (Apoidea).
Systematic Entomology 15, 121-152.
Alexander, B. 1994. Species-groups and cladistic analysis of the
cleptoparasitic bees of genus Nomada (Hymenoptera: Apoidea). The
University of Kansas. Science Bulletin 55 (6), 175-238.
Alexander, B. and C. D. Michener. 1995. Phylogenetic studies of the families
of short-tongued bees (Hymenoptera: Apoidea). The University of
Kansas. Science Bulletin 55 (11), 377-424.
Alfken, J. D. 1932. Die chilenischen Arten der Gatteung Caenohalictus. Cam.
Archiv für Naturgeschichte (n.f.) 1: 654-659.
Alfken, J. D. 1937. Zur Unterscheidung der Bienengatttegen Crocisa. Jur. und
Melecta Latr. Konovia 16: 172-173.
Anonymous, 1958. A list of zoological and botanical types preserved in
collections in southern and east Africa. Vol. 1 Zoology, Published by
the South African Museum's Association, Pretoria. Part I, 147pp.
Armbuster, W. S. and K. E. Steiner. 1992. Pollination Ecology of four
Dalechampia species (Euphorbiaceae) in northern Natal, South Africa.
American Journal of Botany 79 (3), 306-313.
Ashmead, W. H. 1899. Classification of the bees of the superfamily Apoidea.
Transactions of the American Entomologial Society 26, 49-100.
Baker, D. B. 1995. A review of the Asian species of the genus Euaspis
Gerstäcker (Hymenoptera: Apoidea Megachilidae). Zoologische

223
Mededelingen (Leiden) 69, 282-302.
Baker, D. B. 1996a. An annotated list of the nominal species asigned to the
genus Afrostelis Cockerell. Deutsche Entomologische Zeitschrift 43,
155-157.
Baker, D. B. 1996b. Hymenoptera collections of Boyer de Fonscolombe:
Apoidea in the University Museum, Oxford. Journal of Natural
History 30, 537-550.
Benoist, R. 1962. Nouvelles espèces d'Apides malgaches (Hym.). Bulletin de
la Sociétè entomologique de Frances 67, 214-223.
Bingham, C. T. 1897. The fauna of British India including Caylon and Burma,
Hymenoptera, Vol 1. Wasp and Bees XXIX +577. 4pls. Taylor and
Francis [Reprinted in 1975, New Dehli: Today and Tomorrow]
Bingham, C. T. 1903. On the Hymenoptera collected by Mr. W.L. Distant in
the Transvaal, South Africa, with Descriptions of supposed new
Species. Annals and Magazine of Natural History (ser. 12) 7, 46-69.
Brauns, H. 1905. Eine neue Serapis. (Hym.). Zeitschrift für systematische
Hymenopterologie und Dipterologie 5, 75-76.
Brauns, H. 1912. Zusätze und Berichtigungen zu Dr. H. Friese: Die Bienen
Afrikas. (Hym.). Deutsche entomologishe Zeitschrift 1912, 598-603.
Brauns, H. 1926. V. Nachtrag zu `Friese, Bienen Afrikas'. Zoologische
Jahrbücher 52, 187-230.
Brooks, R. W. 1988. Systematics and Phylogeny of the Anthophorine Bees
(Hymenoptera; Anthophoridae: Anthophorini). The University of

224
Kansas. Science Bulletin 53 (9), 436-575.
Camargo, J. M. F. and S. R. Menezes-Pedro. 1992. Systematics, phylogeny
and biogeography of the Meliponinae (Hymenoptera, Apidae): a mini-
review. Apidologie 23, 509-522.
Cameron, P. 1902. Description of a new genera and species of Hymenoptera
collected by Major C. G. Nurse at Deesa, Simla and Ferozepore, Part
II. Journal of Bombay Natural History Society 14: 419-449, 1 pl.
Cameron, P. 1905. On some new genera and species of Hymenoptera from
Cape Colony and Transvaal. Transactions of the South African
Philosophical Society 15, 195-257.
Cockerell, T. D. A. 1898. On some panurgine and other bees. Transactions of
the American Entomological Society 25: 185-198.
Cockerell, T. D. A. 1900. Observations on Bees collected at Las Vegas, New
Mexico and in the adjacent mountains. Annals and Magazine of
Natural History (ser.7) 5, 412-414.
Cockerell, T. D. A. 1904a The bees of southern California – 1. Bulletin
Southern California Academy of Science 3, 3-6.
Cockerell, T. D. A. 1904b. The bee genus Apista etc. Canadian Entomology.
36 (12), 357.
Cockerell, T. D. A. 1904c. New records of bees. Entomologist 37: 231-236.
Cockerell, T.D.A. 1908. LIII-Descriptions and Records of Bees.-XIX. Annals
and Magazine of Natural History ser 8 (1), 337-344.
Cockerell, T. D. A. 1909. Some Anthidiine Bees. Entomological News. 20,
261-262.

225
Cockerell, T. D. A. 1910. New and Little-Known Bees. Transactions of the
American Entomological Society 36 (3), 199-249.
Cockerell, T. D. A. 1914. Descriptions and Records of Bees.-LVII. Annals and
Magazine of Natural History (ser. 8) 13, 277-286.
Cockerell, T. D. A. 1916. XIV.-Report on a collection of South African Bees
chiefly from Natal. Annals of the Durban Museum 1 (3), 188-216.
Cockerell, T. D. A. 1919. Bees in the collection of the united States National
Museum -3. Proceedings of the United States National Museum 55,
167-221.
Cockerell, T. D. A. 1920. XXV. On South African Bees, chiefly collected in
Natal. Annals of the Durban Museum 2, 286-318.
Cockerell, T. D. A. 1922. XLIV.-Descriptions and Records of Bees.-XCIII.
The Annals and Magazine of Natural History (ser. 9) 9, 360-367.
Cockerell, T. D. A. 1925. Anthidiine bees in the collection of the California
Academy of Sciences. Proceedings of the California Academy of
Sciences (4) 14, 345-367.
Cockerell, T. D. A. 1927. Megachilid bees from Bolivia collected by the
Mulford biological expedition, 1921-22. Proceedings of the United
States National Museum 71 (12): 1-22.
Cockerell, T. D. A. 1930a. L.-African Bees of the Genus Megachile. Annals
and Magazine of Natural History (ser. 10) 6, 512-519.
Cockerell, T. D. A. 1930b. African Bees of the family Megachilidae,
(Anthidiinae, Serapista and Lithurgus) mainly from the Belgian Congo
and Liberia (Hymenoptera, Apoidea). Revue de zoologie et de
botanique africaines 19 (1), 43-55.

226
Cockerell, T. D. A. 1931a. African Bees of the family Megachilidae,
(Gronoceras and Megachile) from Liberia and Belgian Congo. Revue
de zoologie et de botanique africaines 20 (2), 132-179.
Cockerell, T. D. A. 1931b. Heriadine and related Bees from Liberia and the
Belgian Congo (Hymenoptera, Apoidea). Revue de zoologie et de
botanique africaines 20 (4), 331-341.
Cockerell, T. D. A. 1931c. Descriptions and Records of Bees.-CXXX. Annals
and Magazine of Natural History 8 (10), 537-553.
Cockerell, T.D.A. 1932a. XXXIX.-Descriptions and Records of
Bees.-CXXXI. Annals and Magazine of Natural History (ser. 10) 9,
355-365.
Cockerell, T. D. A. 1932b. LIV.-Descriptions and Records of Bees.-CXXXIII.
Annals and Magazine of Natural History (ser. 10) 9, 518-527.
Cockerell, T. D. A. 1932c. Bees of the Genus Nomia from Africa. Stylops 1,
159-165.
Cockerell, T. D. A. 1933a. L.-Descriptions and Records of Bees.-CXLIII.
Annals and Magazine of Natural History (ser. 10) 11, 456-468.
Cockerell, T. D. A. 1933b. XII.-Descriptions and Records of Bees.-CXLV.
Annals and Magazine of Natural History (ser. 10) 12, 126-136.
Cockerell, T. D. A. 1933c. XXXV.-Descriptions and Records of
Bees.-CXLVI. Annals and Magazine of Natural History (ser. 10) 12,
357-368.
Cockerell, T. D. A. 1933d. Bees from the Belgian Congo. The Genus Euaspis,
Gerstäecker. Revue de zoologie et de botanique africaines 23 (2), 267-
268.

227
Cockerell, T. D. A. 1933e. Records of the African Bees. II. American Museum
Novitiates No. 615, 1-8.
Cockerell, T. D. A. 1934. VI-Descriptions and Records of Bees.-CXLVII.
Annals and Magazine of Natural History ser. 10 (13), 109-119.
Cockerell, T. D. A. 1935a. XXXII.-Descriptions and Records of Bees.-CL.
Annals and Magazine of Natural History (ser. 10) 16, 343-350.
Cockerell, T. D. A. 1935b. LVII.-Descriptions and Records of Bees.-CLI.
Annals and Magazine of Natural History (ser. 10) 16, 555-562.
Cockerell, T. D. A. 1936a. Some African Anthidiine Bees. American Museum
Novitates No. 855, 1-12.
Cockerell, T. D. A. 1936b. XXXIII.-Descriptions and Records of Bees.-CLIII.
Annals and Magazine of Natural History (ser. 10) 17, 346-360.
Cockerell, T. D. A. 1936c. -Descriptions and Records of Bees.-CLV. Annals
and Magazine of Natural History (ser. 10) 8, 446-456.
Cockerell, T. D. A. 1937a. XLVIII.-Descriptions and Records of Bees.-CLX.
Annals and Magazine of Natural History (ser. 10) 19, 495-505.
Cockerell, T. D. A. 1937b. African Bees of the Genera Ceratina, Halictus and
Megachile. Wiliam Clowes and Sons, Limited, London and Beccles.
254.
Cockerell, T. D. A. 1944. Some African Megachilid Bees. Proceedings of the
Entomological Society of Washington 46 (9), 246-249.
Cockerell, T. D. A. 1947. A new genus of bees from Honduras. Proceedings
of the Entomological Society of Washington 49, 106.

228
Cockerell, T. D. A. and W. P. Cockerell. 1901. Contributions from the New
Mexico Biological Station-IX. On certain genera of bees. Annals and
Magazine of Natural History (7) 7, 46-50.
Comstock, J. H. 1924. An Introduction to Entomology. Xix+1044. Ithaca, New
York: Comstock Publishing Company.
Constanza, R. 1987. The valve of the world’s ecosystem services and natural
capital. Nature, 387.
Cression, E. T. 1864. On the North America species of several genera of
Apidae. Proceedings of the Entomological Society of Philadelphia 4,
1-200.
Cression, E. T. 1878. Description of new North American Hymenoptera in the
collection of the American Entomological Society. Transactions of the
American Entomological Society 7, 61-136.
Danforth, B. N. 2004. Bee fossil and the antiquity of the bees.
http://www.entomology.cornell.edu/Beephylogeny.
Dilcher, D. L. 2006. Paleobotany and evolution.
http://www.britannica.com/oscar/article-73133.
Dominique, J. 1898. Coup d’oeil sur les mellifères sud-américains du muséum
de Nantes. Bulletin de la Société des Sciences Naturelles de l’Ouest de
la France [Nantes] 8 (1), 57-63.
Ducke, A. 1907. Bietrag zur kenntnis der Solitärbienen Brasiliens. Zeitschrift
für systematische Hymenopterologie und Dipterologie 7, 361-368.
Eardley, C. D. and R. W. Brothers. 1997. Phylogeny of the Ammobatini and
Revision of the Afrotropical genera (Hymenoptera: Apidae:
Nomadinae). Journal of Hymenoptera Research 6 (2), 353-418.

229
Engel, M. S. 2000. A new interpretation of the oldest fossil bee (Hymenoptera:
Apidae). American Museum Novitates, 3296, 1-11.
Engel, M. S. 2001. A monograph of the Baltic amber bees and the evolution of
the Apoidea (Hymenoptera). Bulletin of the American Museum of
Natural History, 259, 1-192.
Engel, M. S. 2002. Phylogeny of the bee tribe Fideliini (Hymenoptera:
Megachilidae), with a description of a genus from southern Africa.
African Entomology 10 (2), 305-313.
Engel, M. S. and E. E. Perkovsky, 2006. An Eocene Bee in Rovno Amber,
Ukraine (Hymenoptera: Megachilidae). American Museum Novitates
3506, 1-12.
Fabricius, J. C. 1793. Entomologia Systematica Emendata et Aucta. 2, viii,
519. Copenhagen: Christ. Gottl. Proft.
Fabricius, J. C. 1804. Systema Piezatorum…xiv + [15] – [440] + [1]- 30.
Brunsvigae: Reichard. [The index (pp. [1] – 30) appeared in 1805,
according to Hedicke, 1941, Mitteilungen der Deutschen
Entomologischen Gesellschaft 10: 82-83.
Fact Sheet, 2004. Pollinator Diversity.
http:/www.albany/edu/nature/natweb/dispoll.html
Fairmaire, L. 1858. Ordre Hyménoptères, in J. Thomson, Voyage au Gabon,
Histoire Naturelle des Insectes et des Arachnides. Archives
Entomologiques 2, 263-267.
Felsenstein, J. 1985. Confidence limit on phlogenies: an approach using the
boostrap. Evolution, 39, 783-791.

230
Friese, H. 1895. Die Bienen Europa’s, Vol. 3, Theil I. 128. Belin: Friedländer.
Friese, H. 1898. Die Bienen Europas (Apidae Europaeae) Teil IV. Solitäre
Apiden: Akademische Druk- u. Verlagsanstalt, Graz. 304.
Friese, H. 1902. Beiträge zur kenntniss der insektenfauna von Kamerun.
Entomologisk Tidskrift 23 (3), 225-231.
Friese, H. 1904a. Neue afrikanische Megachile-Arten 111. (Hym.). Zeitschrift
für systematische Hymenopterologie und Dipterologie 4, 330-336.
Friese, H. 1904b. Neue Anthidium-Arten aus der neotropischen Region.
(Hym.) Zeitschrift für systematische Hymenopterologie und
Dipterologie 4, 101-106.
Friese, H. 1905a. Neue afrikanische Bienenarten. (Hym.). Zeitschrift für
systematische Hymenopterologie und Dipterologie 5, (1), 18-19.
Friese, H. 1905b. Die Wollbienen Afrikas. Genus Anthidium (Hym.).
Zeitschrift für systematische Hymenopterologie und Dipterologie 5,
65-75.
Friese, H. 1908. Neue Bienenarten aus Ostafrika. Deutsche entomologische
Zeitschrift Friese, 1908, 567-572.
Friese, H. 1909a. Die Bienen Afrikas nach dem Stande unserer heutigen
Kenntnisse. In: Schultze, L. Zoologische und Anthropologische
Ergebenisse einer Forschungsreise im westlichen und zentralen
Südafrika ausgeführt in den jahren 1903-1905, Band 2. Denkschriften
der Medizinisch-naturwissenschaftlichen Gesellschaft zu Jena 14,
83-476, pls. IX-X.

231
Friese, H. 1909b. 8. Hymenoptera. 5. Apidae. In: Sjostedts, B. Y.
Kilimandjaro-Meru Expedition. Stockholm 8 (5), 119-168.
Friese, H. 1910. Apidae, 119-168 in Y. Sjöstedt, Wissenschaftliche Ergebnisse
der schwedischen zoologischen Expedition nach dem Kilimanjaro, dem
Meru und den umgebenden Massaisteppen Deutsch- Ostafrikas 1905-
1906….Band 2, Abt. 8 (Hymenoptera). Stockholm: Palmquist.
Friese, H. 1911. Nachtrag zu `Bienen Afrikas'. Zoolgische Jahrbücher 30,
651-670.
Friese, H. 1912. Neue und wenig bekannte Bienen Süd-Afrikas. Archiv für
naturgeschichte, Abt A 78 (5): 181-189.
Friese, H. 1913a. II Nachtrag zu 'Bienen Afrika'. Zoologische Jahrbücher 35,
581-598.
Friese, H. 1913b. Neue Bienenarten aus Afrika. Deutsche entomologische
Zeitschrift 1913, 573-578.
Friese, H. 1914. XIV. Fam. Apidae. In: Hymenopteren aus Kamerun.
Gesammelt von Rothkirch, Oberleutnant der Schutztruppe. von Dr. A.
v. Schulthefs, Zurich. Deutsche Entomologische Zeitschrift 1914, 292-
297.
Friese, H. 1915. Zur Bienenfauna von Abessinien. (Hym.). Deutsche
entomologische Zeitschrift 1915, 265-298.
Friese, H. 1916. Hymenoptera. 5. Apidae. In: Michaelsen, U., Beitrage zur
Kenntnis der Land-und Süsswasserfauna Deutsch-
Südwestafrikanischen Studierreise 1911, 415-452.
Friese, H. 1920. On South African Bees, chiefly collected in Natal. Annals of
the Durban Museum 12, 286-318.

232
Friese, H. 1921. Ergebnisse der "Zweiten Deutschen Zentral- Afrika-
Expedition 1910-1911, Apidae. Hamburgische Wissenschaft Zeitung 1,
1091-1112.
Friese, H. 1922. III. Nachtrag zu `Bienen Afrikas'. Zoologischer Jahrbücher
46, 1-42.
Friese, H. 1923. Die Europäischen Bienen. xvii + 456 pp., 33 pls Berlin and
Leipzig: Gruyter.
Friese, H. 1925. III. Nachtrag zu `Bienen Afrika'. Zoologischer Jahrbücher
49, 501-512.
Friese, H. 1926. Forward in H. Brauns, V. Nachtrag zu `Bienen Afrikas.
Zoologische Jahrbücher 52, 187-189.
Friese, H. 1941(1939). Apidae aus Süd-Ost-Africa gesammelt von Missionar
Henri A. Junod Dr phil. h.c. (1863-1934) (Hym. Apid.) Eos Madrid 15,
95-107.
Gerstaecker, C. E. A. 1857. Diagnosen der von Peters in Mossambique
gesammelten Käfer und Hymenoptera (Apiariae, Vespariae). Bericht
über die Verhandlungen der Klg. Sächsischen Gesellschaft (Akademie)
der Wissenschaften zu Leipzig, Math.-Phys. Kl. 1857, 460-464.
Gerstaecker, C. E. A. 1862. In Peters, W.C.H., Naturwissenschaftliche Reise
nach Mossambique. Hymenoptera. Hautflügler, 1862, 439-526. 4 pl.
Gess, F. W. 1981. Some aspects of an ethological study of the aculeate wasps
and the bees of a karroid area in the vicinity of Grahamstown, South
Africa. Annals of the Cape Provincial Museums of Natural History 14
(1), 1-80.

233
Gess, S. K. 1996. The Pollen Wasps. Ecology and Natural History of the
Masarinae. Harvard University Press. 340.
Gess, F. W. and S. K. Gess. 1994. Potential pollinators of the Cape Group of
Crotalarieae (sensu Polhill) (Fabales: Papilionaceae), with implications
for seed production in cultivated rooibos tea. African Entomology 2
(2), 97-106.
Giraud, J. 1863. Hyménoptères recueillis aux environs de Suse, en Piémont, et
dans le départment des Hautes-Alpes, en France; et description de
quinze espéces nouvelles, Verhandlungen der Zoologisch-Botanischen
Gesellschaft in Wien 13, 11-46.
Gistel [Gistl], J. N. F. X. 1848. Naturgeschichte des Thierreichs, für Höhere
Schulen. Xvi + 216 pp., 32 pls. Stuttgart: Hoffmann.
Goloboff, P. A. 1993. NoName (NONA), version 2.0. Program and
documentation. Tucumán, Argentina: Fundación e Instituto Miguel
Lillo.
Gribodo, G. 1895. Memorie della Reale Accademia delle Scienze dell'Istituto
di Bologna, Sezione dell Scienze Naturali 5 (5), 327, 328.
Grigarick, A. A. and L. A. Stange. 1968. The pollen-collecting bees of the
Anthidiini of California. Bulletin of the California Insect Survey no. 9,
1-113.
Grimaldi, D. and M. S. Engel. 2005. Evolution of Insects. Cambridge
University Press. XV + 755.
Griswold, T. L. and C. D. Michener. 1988. Taxonomic Observations on
Anthidiini of the Western Hemisphere (Hymenoptera: Megachilidae).
Journal of the Kansas Entomological Society. 61 (1). 22-45.

234
Gupta, R. K. 1993. Taxonomic Studies on the Megachilidae of North-Western
India. [4] + 288 pp. New Dehli: Indian Council of Agricultural
Research. Also published 1998, Jodhpur: Scientific Publishers (India).
Henning, W. 1966. Phylogenetic Systematic. University of Illinois Press:
Urbana, Illinois.
Holmberg, E. L. 1887. Viaje á Misiones. Boletin de la Academia National de
Ciecias en Córdoba 10,5-391.
Holmberg, E. L. 1903. Delectus Hymenopterologicus Argentinus…Anales del
Museo Nacional de Buenos Aires 2 (3), 377-517.
Ingram, M., Nabhan, G. P. and S. L. Buchman. (With assistance from board
Advisors of the forgotten pollinators). 1996. Ten essential reasons to
protect the birds and the bees. Arizona-Sonara Desert Mesuem,
Tuscom AZ.
Isensee, R. 1927. A study of the male genitalia of certain anthidine bees.
Annals of the Carnegie Museum 17, 371-382, pl. xxxii.
Jurine, L. 1801. In: Panzer, G.W.F. Nachricht von einem entomolischen
Werke, des Hrn Prof. Jurine in Geneve (Beschluss). Intelligenzblatt der
Litteratur-Zeitung [Erlangen] 1, 161-165.
Kenmore, P. and R. Krell. 1998. Global perspectives on pollination in
agriculture and agroecosystem management. International workshop
on the conservation and sustainable use of pollinators in agriculture,
with emphasis on bees. October 7-8, Sao Paulo, Brazil.

235
Kevan, P. G. and V. L. Imperatriz-Fonseca.(eds.) 2002. Pollinating bees: The
conservation link between agriculture and nature. Ministry of
Environment, Brazil.
Klug, J. C. F. 1807. Species apiarium familiae novas, descripsit, generumque
characteres adjecit. Magazin des Gesellschaft naturforscher Freunde
zu Berlin 1, 263-265.
Krombein, K. V. 1951. Additional notes on the bees of the Solomon Islands
(Hymenoptera: Apoidea). Proceedings of the Hawaiian Entomological
Society 14, 277-295.
Lanyon, S. M. 1985. Detecting internal inconsistencies in distance data.
Systematic Zoology, 34, 397-403.
Latreille, P. A. 1809. Genera Crustaceorum et Insectorum Secundum Ordinem
Naturalem in Familias Disposita, Iconibus Exemplisque Plurimis
Explicata. König, Paris et Argentorati, Tome 4, 399.
Latreille, P. A. 1810. Considérations générales sur l'ordre naturel des
animaux composant les classes des Crustacé, des Arachnides et des
Insectes; avec un tableau méthodique de leurs genres, disposés en
familles. VII + 444. Paris.
Lepeletier De Saint-Fargeau, A. L. M. 1835. Histoire naturelle des insects.
Hyménoptères 1835, Vol. 1-547; 1841, Vol. 2, 1-680. Paris.
Lepeletier De Saint-Fargeau, A. L. M. 1841. Histoire naturelle des insects.
Hyménoptères Suites à Buffon. Roret 2, 1-680pp. Paris.
Leys, R., S. J. B. Cooper and M. P. Schwarz. 2002. Molecular phylogeny and
historical biogeography of the large carpenter bees, genus Xylocopa

236
(Hymenoptera: Apidae). Biological Journal of the Linnean Society, 77,
249–266.
Linnaeus, C. 1758. Systema Naturae, Vol. 1, ed. 10. 824. Holmiae: Salvii.
Lipscomb, D. 1998. Basics of cladistic analysis. George Washington
University, Washington D.C. 20052 USA.
Mavromoustakis, G. A. 1934. New Aethiopian Bees of the Subfamily
Anthidiinae (Apoidea). Annals and Magazine of Natural History (ser.
10) 14, 35-48.
Mavromoustakis, G. A. 1936a. New and little known African Bees of the
Subfamily Anthidiinae (Apoidea) Part 1. Annals and Magazine of
Natural History (ser. 10) 17, 31-47.
Mavromoustakis, G. A. 1936b. New and little known African Bees of the
Subfamily Anthidiinae (Apoidea) Part II. Annals and Magazine of
Natural History (ser. 10) 17, 600-606.
Mavromoustakis, G. A. 1937a. Some New African Bees of the Subfamily
Anthidiinae (Apoidea). Annals of the Transvaal Museum 17 (4), 229-
232.
Mavromoustakis, G. A. 1937b. Report on Some Anthidiinae Bees (Apoidea)
in the South African Museum. Annals of the South African Museum 32
(3), 265-267.
Mavromoustakis, G. A. 1938. New African Anthidiinae (Apoidea) Part 1.
Annals of the Transvaal Museum 18 (2), 147-150.
Mavromoustakis, G. A. 1939. New and little known African Bees of the
Subfamily Anthidiinae (Apoidea) Part 3. Annals and Magazine of
Natural History (ser. 11) 3, 346-352.

237
Mavromoustakis, G. A. 1940a. Some bees of the genera Serapista
(Hymenoptera, Apoidea). Annals of the Transvaal Museum 20 (2), 65-
69.
Mavromoustakis, G. A. 1940b. Descriptions of New African Anthidiinae
(Apoidea). Annals of the South African Museum 32 (6), 663-667.
Mavromoustakis, G. A. 1945. New and little known African Bees of the
Subfamily Anthidiinae (Apoidea) Part 4. Annals and Magazine of
Natural History (ser. 11) 12, 180-186.
Mavromoustakis, G. A. 1951. Further contributions to our knowledge of the
Ethiopean Anthidiinae (Hymenoptera, Apoidea) and their
classification. Annals and Magazine of Natural History (ser. 12) 14,
962-981.
Mavromoustakis, G. A. 1953. New and little- known bees of the subfamily
Anthidiinae (Apoidea) – Part VI. Annals and Magazine of Natural
History 6 (12), 834-840.
Mavromoustakis, G. A. 1958. Bees (Hymenoptera, Apoidea) of Attica
(Greece), Part 1. Eos 32, 215-229.
Mavromoustakis, G. A. 1963a. On the bees (Hymenoptera, Apoidea) of
Lebanon, Part III. Annals and Magazines of Natural History 5 (13),
647-655.
Mavromoustakis, G. A. 1963b. Further contributions to our knowledge of the
Ethiopean Anthidiinae (Hymenoptera, Apoidea) and their
classification.- Part 2. Annals and Magazine of Natural History (ser.
13) 6, 481-499.

238
Meade-Waldo, G. 1914. Notes on the Hymenoptera in the collection of the
British Museum, with descriptions of new species. Annals and
Magazine of Natural History 13 (8), 450-464.
Medler, J. T. 1980. Insects of Nigeria - Check List and Bibliography. Memoirs
of the American Entomological Institute 30, 1-919.
Meunier, M. F. 1890. (Communication in meeting of 26 November 1890).
Bulletin de la Societé Entomologique de France 115-117, 201-202.
Michener, C. D. 1942. Taxonomic observations on bees with descriptions of
new genera and species (Hymenoptera: Apoidea). Journal of the New
York Entomological Society, New York 50, 273-282.
Michener, C. D. 1944. Comparative external morphology, phylogeny, and a
classification of the bees. Bulletin of the American Museum of Natural
History 82 (6), 151-326.
Michener, C. D. 1948. The Generic Classification of the Anthidiine Bees
(Hymenoptera, Megachilidae). American Museum Novitates 1381, 1-
29.
Michener, C. D. 1968a. Nests of some African Megachilid Bees, with
Description of a New Hoplitis (Hymenoptera, Apoidea). Journal of the
Entomological Society of southern Africa 31 (2), 337-359.
Michener, C. D. 1968b. Heriades spiniscutis, a bee that facultatively omits
partitions between rearing cells (Hymenoptera, Apoidea). Journal of
the Kansas Entomological Society 41 (4), 484-493.
Michener, C. D. 1995. Some genus-group names of Bees. Journal of Kansas
Entomological Society 67 (1994): 373-377.
Michener, C. D. 1997. Genus-group names of bees and supplemental family

239
group names. Scientic Papers, Natural History Museum, The
University of Kansas 1, 1-81.
Michener, C. D. 2000. The Bees of the World. The John Hopkins University
Press. Baltimore and London. 913.
Michener, C. D. and E. Ordway. 1964. Some anthidiine bees from Mexico.
Journal of the New York Entomological Society 72: 70-78.
Michener, C. D. and D. A. Grimaldi. 1988a. A Trigona from late Cretaceous
amber of New Jersey. American Museum Novitates no. 2917: 1-10.
Michener, C. D. and D. A. Grimaldi. 1988b. The oldest fossil bee: Apoid
history, evolutionary stasis, and anthiquity of social behaviour.
Proceedings of the National Academy of Sciences USA 85, 6424-6426.
Michener, C. D. and T. L. Griswold. 1994. The Classification of Old World
Anthidiini (Hymenoptera, Megachilidae). The University of Kansas
Science Bulletin 55 (9), 299-327.
Mocáry, A. 1894. E fauna apidarum hungariae. Természetrajzt Füztek 17, 34-
37.
Morawitz, F. 1875. Pchely (Mellifera Latr.), Tetrad 1, Apidae Genuinae, pp.
1-160 in A.P. Fedtschenko, Puteshestvie v Turkestan (Reisen in
Turkestan), Zoogeografischeskaya Izsledovaniya, part V, sec. 7. [Also
as Izvestiya Imperatorskago Obshchestva Lyubiteley Yestestvoznaniya
Antropologii I Etnografii 19 (2): 1-160].
Morawitz, F. 1876. Zur Bienenfauna der Caucasusländer. Horae Societatis

240
Entomologicae Rossicae 12, 3-69.
Moure, J. S. 1947a. Novos agrupamentos genéricos e algumas espécies novas
de abelhas sulamericanas. Museu paranaense Publicações Avulsas, 1:
1-37.
Moure, J. S. 1947b. Notas sobre algunas abejas de la provincia de Salta.
Revisita de la Sociedad Entomológica Argentina 13, 218-253.
Moure, J. S. 1965. New placements for some species of Paranthidium.
Proceedings of the Entomological Society of Washington 67, 29-31.
Moure J. S. and P. D. Hurd, Jr. 1960. On the systematic position of three
anthidine bees described byAldof Ducke. Boletim de Museu Paraense
Emilio Goeldi. (Nova Série), Zoologia. 28, 1-13.
Moure, J. S. and D. Urban. 1975. Dicranthidium novo subgenero de
Hypanthidioides [sic] Moure, 1947. Revista Brasileria de Biologia 35,
837-842.
Moure, J. S. and D. Urban. 1990. Carloticola gen. n. e Carloticola trichura sp.
N. da Argentina. Acta Biologica Paranaense 19, 89-99.
Moure, J. S. and D. Urban. 1994. Rhynostelis, gen. n. e notas sobre Hoplostelis
Dominique, Revista Brasileira de Zoologia 11, 297-302.
Müller, A. 1996. Host plant specialization in western palearctic anthidiine
bees. Ecological Monographs 66, 235-257.
Nixon, K. C. 1999-2002. WinClada ver. 1.0000 Published by the author,
Ithaca, NY, USA.
Noskiewicz, J. 1961. Beiträge zur Kenntnis der palärktischen Arten der
Gattung Stelis. Polskie Pismo Entomologiczne 31, 113-133.

241
O’Toole, C. 1993. Diversity of native bees and agroecosystems, In: LaSalle, J.
and Gauld, I. (eds.) Hymenoptera and Biodiversity. Commonwealth
Agricultural Bureau International, London. 69-106.
O’Toole, C and A. Raw. 1991. Bees of the World.192. London: Blanford.
Pagliano, G. and P. Scaramozzino. 1990. Elenco dei generi di Hymenoptera
del mondo. Memoria della Società Entomologica Italiana 68 (1989):
1-210.
Panzer, G. W. E. 1804. Faunae Insectorum Germancae, heft 86. Nürnberg:
Felssecker. [According to Coommission Opinion 151 (1944), the
correct date for this heft is [1801-1802], but Sherborn (ref. supra),
gives the date as 1804.]
Panzer, G. W. E. 1806. Kristische Revision der Insektenfauna Deutschlands,
Vol., 2. [14] + 271, 2 pls. Nürnberg: Felssecker.
Parker, F. D. and G. E. Bohart. 1979. Dolichostelis, a new genus of parasitic
bees. Journal of the Kansas Entomological Society 52, 138-153.
Pasteels, J. J. 1968a. Statut, affinities et origins des Anthidiines parasites.
Naturalist Canadien 95, 1055-1063.
Pasteels, J. J. 1968b. Revision des Megachilidae (Hymenoptera Apoidea) de
L'Afrique Noire. II. Le Genre Coelioxys. Musee Royal de L'Afrique
Centrale- Tervuren, Belgique Annales, Serie In-8- Sciences
Zoologiques 167, 1-139.
Pasteels, J. J. 1968c. A new species of Pachyanthidium (Hymenoptera:
Megachilidae). Journal of the Kansas Entomological Society 41 (4),
481-483.

242
Pasteels, J. J. 1969. La systématique générique et subgénérique des
Anthidiinae (Hymenoptera, Apoidea, Megachilidae) de l'Ancien
Monde. Mem Soc r. ent. Belg. 31, 3-148.
Pasteels, J. J. 1972. Révision des Anthidiinae (Hymenoptera Apoidea) de la
region Indo-Malaise. Bulletin et Annuales de la société Royale Belge
d’Entomologie 108,72-128.
Pasteels, J. J. 1977. Une revue comparative de l'éthologie des Anthidiinae
nidificateurs de l'ancien monde (Hymenoptera, Megachildae). Annales
de la Société Entomologique de France. (n.s.) 13 (4), 651-667.
Pasteels, J. J. 1980. Revision du Genre Euaspis Gerstaecker (Hymenoptera,
Apoidea, Megachilidae) Bulletin et Annales de la Société Royale Belg
d’Entomologie. 116, 73-89.
Pasteels, J. J. 1984. Révision des Anthidiinae (Hymenoptera, Apoidea,
Megachilidae) de l'Afrique subsaharienne. Mémoires de la classa des
Sciences XIX, 1-165.
Pauly, A. 1998. Hymenoptera Apoidea du Gabon. Musée Royal de l’Afrique
Centrale Tervuren, Belgique. Annales Sciences Zoologique 282, 1-121.
Pauly, A. 2001. Notes on phoretic deutonymphs of mites (Acari) associated with
Old World Megachilidae and Anthophoridae (Insecta Hymenoptera),
mainly Madagascar. 1. Families Chaetodactylidae, Acaridae,
Histiostomatidae and Winterschmidtiidae (Astigmata). Belgian Journal
of Entomology 3, 125-142.
Pauly, A., R. W. Brooks, L. A. Nilsson, Y. A. Pesenko, C. D. Eardley, M.
Terzo, T. Griswold, M. Schwarz, S. Patiny, J Munzinger and Y.

243
Barbier. 2001. Hymenoptera Apoidea de Madagascar et des Iles
Voisines.Musee Royal de L'Afrique Centrale- Tervuren, Belgique
Annales Sciences Zoologiques 286, 1-390, 16 planche.
Peters, D. S. 1972a. Über die stellung von Aspidosmia Brauns 1926 nebst
allgemeinen erörterungen der phylogenetischen systematik der
Megachilidae (Insecta, Hymenoptera, Apoidea). Apidologie 3 (2), 167-
186.
Peters, D. S. 1972b. Drei neue Arten der Megachilinae aus Mexico.
Senckenbergiana Biologica 53 373-382.
Peters, D. S. 1984. Revision der von H. Brauns beschriebenen oder
behandelten afrikanischen Heriades- und Osmia-Arten (Hymenoptera,
Apoidea). Entomofauna 5 (32), 359-390.
Poinar, G. O. (Jr) and B. N. Danforth 2006. Bee fossil, DNA genera a buzz.
http://www.news.bbc.co.uk/go/fr/-/2/hi/science/nature/6084974.stm.
Popov, V. B. 1935. Beiträge zur Bienenfauna von Tadjikstan. Trudy
Akademiya Nauk SSSR, Tadzhikskoi Bazo no. 5: 351-408. [In Russian,
Germany summary]
Popov, V. B. 1938. Notiz über die Gattung Chelynia Pro. und einige
Untergattungsgruppierungen der Gattung Stelis Panz. Konowia 17, 36-
41.
Popov, V. B. 1941. Notes on Dianthidium sibericum (Eversm.) and a new a
new species of Stelis Panz. Entomologisk Tidskrift 62, 222-224.
Popov, V. B. 1950. Generic groupings of the mid-Asian bees of the subfamily
Anthidiinae. Doklady Akademii Nauk SSSR 70, 315-318. [In Russian].

244
Popov, V. B. 1956. New and little-known bees from central Asia.
Entomologicheskoe Obozrenie 35, 159-171. [In Russian].
Popov, V. B. 1964. On the bee genera Trachusa Panzer and Trachusomimus,
gen. n. Entomologicheskoe Obozrenie 43, 405-417. [In Russian ;
English translation in Entomological Review 43, 207-214.]
Prescott-Allen, R. and C. Prescott-Allen. 1990. How many crops plants feed
the world? Conservation Biology, 4, 3650374.
Prins, A. J. 1978. Hymenoptera. In Biogeography and Ecology of southern
Africa. Ed. M.J.A. Werger and A.C. van Bruggen. W. Junk Publishers.
The Hague.
Provancher, L. 1888. Additions and corrections au volume II de la faune
entomologique du Canada Traitant des Hyménoptères. 148. C.
Darveau, Quebec.
Radoszkowski, O. 1874. Supplément indispensable à l’article publié par M.
Gerstaecker en 1869, sur quelques genres d’hyménoptères. Bulletin de
la Société Imperiale des Naturalistes de Moscou 46 (3), 33-151.
Raven, P. H., and D. I. Axelrod. 1974. Angiosperm biogeography and past
continental movements. Annals of the Missouri Botanical Garden. 61,
539-673.
Ritsema, C. 1874. Aanteekeningen betreffende eene kleine collectie
Hymenoptera van Neder-Guinea, en beschrijving van de nieuwe
soorten. Tijdschrift voor Entomologie, s"Gravenhage, Amsterdam. 17,
175-211.

245
Robertson, I. A. D. 1902. Some new or little known bees- 11. Canandian
Entomologist. 34 48-49.
Robertson, I. A. D. 1903. Synopsis of Megachilidae and Bombinae.
Transactions of the American Entomological Society 29, 163-178.
Robertson, I. A. D. 1904. Synopsis of Anthophila. Canandian Entomologist
35, 284-288.
Robertson, I. A. D. 1969. A list of Hymenoptera (Symphyta and Aculeata)
collected in Tanzania. Journal of the Entomological Society of
Southern Africa 32 (2), 479-480.
Roig-Alsina, A. 1991. Cladistic Analysis of the Nomadinae s. str. With
Description of a new genus (Hymenoptera: Anthophoridae). Journal of
the Kansas Entomological Society 64 (1), 23-37.
Roig-Alsina, A. and C. D. Michener, 1993. No. 4. Studies of the Phylogeny
and Classification of Long-Tongued Bees (Hymenoptera: Apoidea).
The University of Kansas Science Bulletin 55 (4 & 5), 123-162.
Romankova, T. G. 1988. A new far-eastern bee of the tribe Anthidiini. Vestnik
Zoologii 1988 (4): 25-30. [In Russian.]
Rozen, J. G. 1970. Biology, Immature stages and Phylogenetic relationship of
Fideliinae bees, with description of a new species of Neofidelia
(Hymenoptera: Apoidea). American Museum Novitates 2427, 1-25.
Rozen, J. G. 1996. Phylogenetic analysis of the cleptoparasitic bees belonging
to the Nomadinae based on matured larvae (Apoidea: Apidae).
American Museum Novitates 3309, 1-39.

246
Rozen, J. G. and R. McGinley. 1974. Phylogeny and systematics of the
Melittidae based on the matured larvae (Insecta. Hymenoptera:
Apoidea). American Museum Novitates 2545, 1-3.
Sandhouse, G.H. 1943. The type species of the genera and subgenera of bees.
Proceeding of the United States National Museum , 512-619.
Saussure, H. De. 1890. Histoire Naturelle des Hymènoptéres, Vol. 20, xxi +
590, 27 pls., in A. Granndidier, Histoire Physique, Naturelle et
Politique de Madagascar. Paris: Imprimerie Nationale. [Actually
published in 1891]
Schenck, A. 1861. Die Naussauischen Bienen. Jahrbűcher des Vereins fűr
Naturkunde im Herzogthum Nassau 14 1-414.
Schenck, A. 1869. Beschreibung der naussauischen Bienen, Zweiter Nachtrag.
Jahrbűcher des Naussauischen Vereins fűr Naturkunde 21-22, 1[269]-
114[328].
Schmedeknecht, H. L. O. 1882-1884. Apidae Europaeae [Die Bienen
Europa’s], Vol. 1, [xiv] + 866, pls. 1-15. Belin: Gumperdae and
Berolini. [Pp. 1-314 published in 1882; 315-550, 1883; and 551-866,
1886.]
Schrottky, C. 1902. Ensaio sobre as abelhas soltarias do Brazil, Revista do
Museo Paulista 5, 330-613, pls. XII-XIV.
Schwarz, H. F. 1926. North American Dianthidium Anthidiellum, and
Paranthidium. American Musuem Novitates no. 253, 1-17.

247
Siddal, M. E. 1996. Another monophyly index: revisiting the jackknife.
Cladistics, 11, 33-56.
Sladen, F. W. L. 1916. Canadian species o the bee genus Stelis Panz.
Canadian Entomologist 48, 312-314.
Snelling, R. R. 1987. A revision of the genus Aztecanthidium. Pan-Pacific
Entomologist 63, 165-171.
Smith, F. 1853. Catalogue of Hymenopterous Insects in the Collection of the
British Museum. London, Part I, Andrenidae and Apidae. 197pp., pls.
I-VI.
Smith, F. 1854. Catalogue of Hymenopterous Insects in the collection of the
British Museum. London, Part II, Apidae. pp. 198-465, pls. VII-XII.
Smith, F. 1879. Descriptions of new species of Hymenoptera in the collection
of the British Museum. xxi + 240.London, British Museum.
Spinola. M. 1808. Insectorum Liguriae Species Novae aut Raviores..., 2-4,
262. Genuae: Printed for the author.
Spinola, M. 1838. Compte- rendu des Hyménoptères recuellis par M. Fischer
pendant son voyage en Egypt. Annales de la Sociétè Entomologique de
France 7 (1838): 437-546.
Spinola, M. 1851. Hymenópteros, pp. 153-569. In C. Gay, Historia Fisica y
Politica de Chile.., Zoologia, 6. Paris: Casa del autor.
Stadelmann, H. 1898. Die Hymenopteren Ost-Afrikas. In: Möbius, K., Die
Thierwelt Ost-Afrikas und der Nachbargebiete 4 (3), 1-74.

248
Stange. L. A. 1983. A synopsis of the genus Epanthidium Moure with the
description of a new species from north–eastern Mexico. Pan-Pacific
Entomologist 59: 281-297.
Strand, E. 1911a. Faunistische und systematische Notizen über afrikanische
Bienen. Wiener entomologische Zeitung 30 (6, 7), 135-159.
Strand, E. 1911b. Neue afrikanische Arten der Bienen-gattungen Anthophora,
Eriades, Anthidium, Coelioxys und Trigona. Entomologische
Rundschau 28, 119-120, 122-124.
Strand, E. 1912a. Ein neueres Werk über afrikanische Bienen Kritisch
besprochen. Archiv für Naturgeschichte 78 (1), 126-144.
Strand, E. 1912b. Zoologische Ergebisse der Expedition des Herrn G.
Tessmann nach Süd-Kamerun und Spanisch-Guinea. Bienen.
Mitteilungen aus dem Zoologische Museum in Berlin 6, 265-312.
Strand, E. 1914. Ueber einige afrikanische Bienen des Deutschen
Entomologischen Museums. Archiv für Naturgeschichte 80A. (9), 61-
67.
Strand, E. 1917. Über einige Apidae des Deutschen Entomologischen
Museums. Archiv für Naturgeschichte. 83A, 57-71.
Strand, E. 1920 (1921). Notes sur quelques Apides de Congo belge. Revue de
zoologie et de botanique africaines 8, 87-106.
Struck, M. 1992. Pollination Ecology in the arid winter rainfall Region of
southern Africa: a case study. Mitteilungen aus dem Institut fuer
Allgemeine Botanik Hamburg 24, 61-90.

249
Struck, M. 1994a. A check-list of flower visiting insects and their host plants
of the Goegab Nature Reserve, Northwestern Cape, South Africa.
Bontebok 9, 11-21.
Struck, M. 1994b. Flowers and their insect visitors in the arid winter rainfall
region of southern Africa: observations on permanent plots.
Composition of the anthophilous insect fauna. Journal of Arid
Environments 28, 45-50.
Taylor, J. S. 1965. Notes on some South African bees (Hymenoptera:
Colletidae, Megachilidae). The Pan-Pacific Entomologist 41, 173-179.
Timberlake, P. H. 1941. Ten new species of Stelis from California. Journal of
the New York Entomological Society 49, 123-137.
Tkalcu, B. 1967. Bemerkungen zur Taxonomie einiger paläarktischen Arten
der Familie Megachilidae. Acta Entomologica Bohemoslovaca 64, 91-
104.
Thomsom, C. G. 1872. Skandinaviens Hymenoptera, 2,1-286. Lund: Berling.
Urban, D. 1991. Ananthidium, um gênero novo di Dianthidiini neotropical.
Revista Brazileira de Zoologia 7, 73-78
Urban, D. 1993. Ctenanthidium, gen. n. de Dianthidium com quarto espécies
novas da América do Sul. Revista Brasileira de Zoologia 8 (1992): 11-
28.
Urban, D. 1994a. Tylanthidium, gen, n. de Anthidiinae da América do Sul
(Hymenoptera, Megachilidae) e nota taxonômica. Revista Brasileira de
Zoologia 11, 277-281.
Urban, D. 1994b. Gnanthanthidium, gen. n. de Anthidiinae da América do Sul.
Revista Brasileira de Zoologia 9 (1992): 337-343.

250
Urban, D. 1995. Moureanthidium, gen. n. de Dianthidiini do Brasil. Revista
Brasileira de Zoologia 12, 37-45.
Urban, D. 1996. Mielkeanthidium, gen. n. de Dianthidiini da América do Sul.
Revista Brasileira de Zoologia 13, 121-125.
Urban, D. 1997a. Larocanthidium, gen. n. de Anthidiinae do Brasil. Revista
Brasileira de Zoologia 14, 299-317.
Urban, D. 1997b. Chrisanthidium, um novo gênero sulamericano de
Dianthidiini. Revista Brasileira de Zoologia 14, 181-185.
Vachal, J. 1903. Hyménoptères du Congo Francais Rapportés par l'ingénieur J.
Bouyssou. Mellifera. Annales de la Société entomologique de France
72, 358-400. Vachal, J. 1910a. Collections recueilles par M. le Baron
Maurice de Rothschild dans l’Afrique orientale Insectes.
hyménoptéres: Mellifres. Bulletin du Muséum (national) d’histoire
naturelle 15, 529-534.
Vachal, J. 1910b. Diagnoses d'insectes nouveaux recuellis dans le Congo
belge par le Dr. Sheffield-Neave: Hymenoptera, Apidae. Annales de la
Sociéte entomologique de Belgique 54, 306-328.
Warncke, K. 1980. Die Bienengattung Anthidium Fabricius 1804 in der
Westpaläarktis und im turkestanischen Becken. Entomofauna 1, 119-
209.
Williams, P. H. 1994. Phylogenetic relationships among bumble bees
(Bombus Latr.): a reappraisal of morphological evidence - Systematic
Entomology- Blackwell Synergy. 1. The Natural History Museum,
London.

251
APPENDICIES
APPENDIX 1
Genera of Anthidiini in Each Series (** indicate parasitic groups)
Series A Series B
Acedanthidium Euaspis ** Afranthidium
Afrostelis ** Hoplostelis ** Anthidioma
Anthidiellum Hypanthidiodes Anthidium
Anthodioctes Hypanthidium Gnathanthidium
Apianthidium Icteranthidium Indanthidium
Aspidosmia Larinostelis** Neanthidium
Atropium Notanthidium Pseudoanthidium
Aztecanthidium Pachyanthidium Serapista
Bathanthidium Paranthidium
Benanthis Pleisanthidium
Cyphanthidium Rhodanthidium
Dianthidium Stelis
Duckeanthidium Trachusa
Eoanthidium Trachusoides
Epanthidium
(Michener, 2000)

252
APPENDIX 2
This give detailed information of the locality labels on each specimen examined
in this research. They have been captioned as material examined.
MATERIAL EXAMINED
Genus Acedanthidium Michener and Griswold
Type material: Acanthidium batrae Michener and Griswold, male and
female paratype: India, Uttar Pradesh, S.W.T. Batra (SEMC).
Genus Afranthidium Michener
Total specimens: 66 females and 45 males.
Afranthidium abdominale (Friese)
4 females & 2 males. SOUTH AFRICA: Pretoria, ii.1949, J.H. Stiglingh (1
female- SANC); Christiana, 9.i.1965, H.N. Empey (1 female- SANC);
Middlesburg, Kaoo, 11.ii.1971, H.N. Empey (1 female, 1 male- SANC); Tussen
Die Rivere Residence near Bethulie, 30º30'S 26º12'E, 30.iii.-3.iv.1987, C.D.
Eardley (1 female,1 male- SANC).
Afranthidium braunsi (Friese)
36 females & 16 males. BOTSWANA: Serowe, 4.xi.1984, P. Forchhammer (1
male- SANC); Serowe, 4.ix.1986, P. Forchhammer (1 female, 1 male- SANC);
Serowe, 1988, P. Forchhammer (2 males- SANC). NAMIBIA: Near Orange
River, 14 Km S Rosh Pinash, 28º17'S 16º55'E, 27.x.1974, R.H. Watmough (1
female- SANC). SOUTH AFRICA: Beaufort West, Cape Town, 24.xi.1933,
J.Ogilvie (1 female- SANC); Mondeor, Johannesburg, 4.xii.1961, H.N. Empey
(1 female- SANC); Wepener, 18.i.1965, H.N. Empey (1 female- SANC);
Middleburg, 16.xi.1972, 24.i.1973, E. Holm (5 females- SANC); Oudtshoorn

253
Experiment Farm, 4.iii.1981, R.H. Watmough (3 female- SANC); Petrsville,
30º05'S 24º14'E, 2.xi.1981, H. Horn (1 female- SANC); Pretoria District,
Soutpan, 25º24'S 28º06'E, 13.xii.1982, C.D. Eardley (1 female, 3 males-
SANC); Rooiheuwel, 5 Km E Oudtshoorn, 33º36'S 22º17'E,5.x.1981, xi.1982,
2.xii.1983, 2.xii.1984, R.H. Watmough (8 females, 5 males- SANC);
Magaliesberg, Castle Gorge, 25º48'S 27º34'E, 17.ii.1993, R. Urban (1 male-
SANC); Adullam Farm, 28º32'S 28º28'E, 15-18.i.1986, 17-18.i.1994, C.D.
Eardley (4 females- SANC); 20 km E Waterpoot, 22º52'S 29º47'E, 6.i.2004,
C.D. Eardley (1 male- SANC); Guateng, Soutpan, 25º24'S 28º06'E, C.D.
Eardley (1 male- SANC).
Afranthidium folliculosum (du Buysson)
7 females and 14 males. KENYA: Central Island, Lake Turkana, 5.vii.1980, 23-
25.vii.1980 (12 males- KNM). NAMIBIA: Okahandija District, Erichsfelde 44,
21º35'S 16º56'E, 20-22.iii.2003, A.H. Kirk-Spriggs (1 female- SMWN).
SOUTH AFRICA: Alberton, Transvaal, 28.i.1966, H.N. Empey (1 male-
SANC); D’ Nyala Nature Reserve, Ellisras District, 23º45'S 27º49'E, 10-
14.xi.1986, V.M. Uys (1 female- SANC); D’ Nyala Nature Reserve, Ellisras
District, 23º45'S 27º49'E, 8-12.xii.1989, C.D. Eardley (3 females, 1 male-
SANC); Hotazel, 27º13'S 22º57'E, C.D. Eardley (1 female- SANC).
Afranthidium immaculatum (Smith)
7 females and 6 males. KENYA: Makalia Bridge Area, Lake Narkura,
28.xi.1977, E. Mlen (1 female- KNM); Gilgil, Aloe-Lalashwa grassland,
9.v.1982 (1 female, 1 male- KNM). SOUTH AFRICA: Graskop, E. Transvaal,
6.v.1964, H.N. Empey (1 male- SANC); Orange Free State, Marquad,
26.xii.1975, H.N. Empey (1 male- SANC); Long Tom Pass, 25º07'S 30º35'E,

254
i.1977, E.F.Whiteside (1 female- SANC); Tussen Die Rivere Residence near
Bethulie, 30º30'S 26º12'E, 30.iii.-3.iv.1987, 20-26.xii.1994, C.D. Eardley (2
females- SANC); Lesotho, 12km NE of Mamohau, 23.i.1992, H. Geertsema (1
female-SANC); Tussen Die Rivere Residence near Bethulie, 30º30'S 26º12'E,
21.i.1994, R. Adam (1 male- SANC). ZIMBABWE: Chimanimani Mountain
Hut, 4.iv.1972, R.H. Watmough (1 female- SANC).Afranthidium minutulum
(Friese)
12 females and 7 males. NAMIBIA: Ugab Road, 2 km W Brandberg, 20º57'S
14º06'E, 22-24.x.1998, E. Marais and A. Kirk- Spriggs (1 female, 2 males-
SMWN). SOUTH AFRICA: Augrabies National Reserve, 11.xii.1974, H.N.
Empey (1 female, 1male- SANC); Winheok, 22º34'S 17º06'E, xii.1983,
28.xiii.1983, R. Oberprieler (7 females, 4 males- SANC); 20 km E of
Waterpoort, 22º52'S 29º47'E, 6.i.2004, C.D. Eardley 2 females- SANC);
Richsterveld National Park, near Oena Mines, 28º03'S 17º03'E, R. Combey (1
female-SANC).
Genus Afrostelis Cockerell
Total specimens examined: 21 females and 10 males.
Afrostelis aethiopica (Friese)
19 females and 7 males. DEMOCRATIC REPUBLIC OF CONGO:
Bulawayo, S. Rhodesia, xii.1912, G. Arnold (1 female- TMSA). SOUTH
AFRICA: Lady Grey, 13.xii.1924 (1 male- NHML); Katberg, 15-30.i.1933,
R.E. Turner (1 female- NHML); Discovery Florida Transvaal, 16.ix.1958, H.N.
Empey (1 male- SANC); Mogoto National Reserve, Zebediela, 24º15'S
29º13'E, 22-25.x.1979, C.D. Eardley (1 male- SANC); Groenfontein, 35 km E

255
Thabazimbi, 24º34'S 27º45'E, 27.xi.1980, C. Kok (1 male- SANC);
Rustenburg Nature Reserve, 25º40'S 27º12'E, 17-20.iii.1980, C.D. Eardley (1
female, 1 male- SANC); Duiweiskloof, 23º42'S 30º06'E, 12-14.i.1987, C.D.
Eardley (1 male- SANC); Molteno Pass, near Beaufort West, 32º12'S 22º33'E,
14.xii.1988, C.D. Eardley (1 female- SANC); Karoo National Park, Beaufort
West, 32º20'S 22º30'E, 13.ii.1991, C.D. Eardley (1 male- SANC); Adullam
Farm, 28º32'S 28º28'E, 15-18.i.1986, 17-18.i.1994, C.D. Eardley (4 females, 1
male-SANC); Adullam Farm, 28º32'S 28º28'E, 18.i.1994, V.M.Uys (1 female-
SANC).
Afrostelis tegularis (Cockerell)
Type material: Afrostelis tegularis (Cockerell), female holotype:
Kamaiola, Congo, MRAC.
Additional material: CONGO: Bukama, 6.v.1911, Dr Bequaert (1 male-
MRAC); Luluabourg, 12.v.1919, P.Callewaert (1 male- MRAC).
Genus Anthidiellum Cockerell
Total material examined: 91 females & 68 males
Type material: Anthidiellum eritrinum Friese, female holotype, Gheleb
Eritrea, J. Pasteels 1963’, (HUMB).
A rubellum
1 female and 1 male. SERBIAN: (1female- HUMB). TUNISIA:
Tunis, .1976 (1 male- HUMB ‘labelled as type’).
A. flavescens

256
23 females and 8 males. ANGOLA: Bruco, 26.ii.-2.iii.1972 (1 male-
NHML). DEMOCRATIC REPUBLIC OF CONGO: Lonely Mine, S.
Rhodesia, 26.ii.1913, 21-24.v.1913, 13.xii.1913, 18.ii.1914, 26.xii.1914,
15.ii.1915, A.H. Swali (14 females- NHML). NAMIBIA: Kaoko Otavi, (1
female, 1 male- MRAC). ZIMBABWE: Bulawayo, 21.xii.1919, 28.xii.1924 (1
female, 2 males- SAMC, TMSA); Turk Mine, 16.xii.1957, H. Empey (5
females, 1 male- SANC); Premier Mine, 14.xii.1941 (1 female, 1 male-
SAMC).
A. absonulum
16 females & 17 males. BOTSWANA: Serowe, 17.ii.1984, 1988, P.
Forchhammer (2 females- SANC). DEMOCRATIC REPUBLIC OF CONGO:
Elizabethville, 11-17.ix.1931, J. Ogilvie (2 females- AMNH, MRAC),
3.xi.1933, M. Bequaert (1 male- MRAC); Haut-Uele: Paulis Mholi, iv.1947,
P.L.G. Benoit (1 female- MRAC). NAMIBIA: Salambala Forest, 17º09'57'S
20º52' 55'E, 26-27.xii.1998, A. Kirk-Spriggs & E. Marais, (1 female-
SMWN); idem, 23-29.xii.2002, A.H. & M.K. Kirk-Spriggs (7 females, 13
males- SMWN). SOUTH AFRICA: Mogol Nature Reserve, Ellisras District,
23º58'S 27º45'E, 25-26.i.1982, C.D. Eardley (1 female- SANC); idem,
21.xii.1987, B. Grobbelaar (1 female- SANC); D’ Nyala Nature Reserve,
Ellisras District, 23º45'S 27º49'E, 8.xii.1989, C.D. Eardley (1 male- SANC);
Schoongelegen, 24º12'S 27º45'E, 14.i.1991, V.M.Uys (2 males- SANC).

257
A. apicatum
7 females. SOUTH AFRICA: Happy Rest Nature Reserve, 22º55'57'S
29º44'06'E, 8.i.2004, C. D. Eardley (4 females- SANC). ZIMBABWE: Cashel,
30.xii.1947 (3 females- MRAC, SAMC).
Type material: Anthidiellum auriscopatum Strand, female holotype:
Spanish Guinea, Hinterland-Makomo, S.G. Tessman (HUMB).
Additional material: 2 females. DEMOCRATIC REPUBLIC OF THE
CONGO: Elizabethville, Piége Harris, 8.ix.1909, 11-17; idem, ix.1934, P.
Quarré (2 females- MRAC).
A. bipectinatum
5 females & 1 male: BOTSWANA: Serowe, 17.ii.1988, P.
Forchhammer (1 female- SANC). SOUTH AFRICA: Roodeplaat, xi.1978,
Pretoria District, C.D. Eardley (1 female- SANC); Soutpan, 25º24'S 28º06'E,
16.xi.1983, C.D. Eardley (2 females- SANC); Rustenburg Nature Reserve,
25º40'S 27º12'E, 8.xii.1983, C. D. Eardley (1 male- SANC).
A. bulaweyense
2 males. DEMOCRATIC REPUBLIC OF CONGO: Uamgebiet,
Bosum, 21.iii.1931, S.G. Tessmann (1 male- HUMB). ZIMBABWE:
Bulawayo, xii.1938 (1 male- SAMC).
Type material: Anthidiellum frontoreticulatum (sp. nov), holotype
female: Kenya, 26.v.1967, C.D. Michener, MRAC; Anthidiellum
frontoreticulatum (sp. nov), allotype male: Kenya, 26.v.1967, C.D. Michener,
MRAC; idem paratypes 2 females, 1 male, SEMC.

258
Type material: Anthidiellum kimberleyana (Friese), holotype female:
Kimberley, South Africa, Bro. Power (HUMB).A. orichalciscopatum
1 female. SOUTH AFRICA: Mossel Bay, i.1940 (1 female- SAMC).
A. otavicum
6 females and 3 males. KENYA: South Melindi, 26.v.1967, C. D.
Michener (4 female, 2 males- MRAC, AMNH). NAMIBIA: Otavi,
27.xii.1933, 16.i.1934, J. Ogilvie (2 females, 1 male- NHML).
Type material: Anthidiellum (Pycnanthidium) polyochrum
Mavromoustakis, holotype female: Kaross, South Africa, SAMC.
Additional material: 1 female, 1 male: NAMIBIA: 40 km North
Usakos, 18.ii.1977, J. G. & B. L. Rozen (1 male- AMNH); Ameib Farm,
19mls, NW Karibib, 31.i.-2.ii.1972 (1 female- NHML).
A. spilotum
5 females and 7 males: ANGOLA: Bruco, 26.ii.-2.iii.1972 (1 female-
NHML). BOTSWANA: Serowe, 17.ii.1988, P. Forchhammer (1 female-
SMWN). SOUTH AFRICA: Mossel Bay, i.1940 (1 female- SAMC); Crystal
Waters, Breedsnek, 4.xi.1962, H.N. Empey (2 females, 5 males- SANC);
Queenstown, 16.xii.1979 H.N. Empey (1 female, 1 male- SANC);
Saartjiesnek, 25º41'S 27º54'E, 12.xi.1983, C. Bellamy (1 male- SANC).
Type material: Anthidiellum (Pycnanthidiellum) sternale Pasteels,
holotype male and 1 paratype male: Breedsnek Transvaal, South Africa,

259
TMSA; Anthidiellum (Pycnanthidiellum) sternale Pasteels, 1 paratype male:
Mogal Nature Reserve, South Africa SANC).
Additional material: 1 female & 1 male. SOUTH AFRICA: Nylsvley
Nature Reserve, 24º39'S 28º42'E, 10-11.xii.1979, C.D. Eardley (1 male-
SANC); Mogal Nature Reserve, Ellisras District, 23º58'S 27º45'E, 25-
26.i.1982, C.D. Eardley (1 female- SANC).
A. tegwaniense
2 female and 2 males. ZIMBABWE: Bulawayo, 11.iv.1923, R.
Stevenson (1 male- TMSA); Khami, 28.x.1928 (1 female- SAMC); Khami,
27.x.1938 (1 female, 1 male- SAMC).
A. zebra
17 females and 21 males. BOTSWANA: Serowe, 17.ii.1984, 1988,
Forchhammer (1 male- SANC). SOUTH AFRICA: Willomore, 5.i.1905, H.
Brauns (2 female, 2 male- HUMB); Ben Alberts Nature Reserve, 24º37'S
27º23'E, 24-28.xi.1980, C.D. Eardley (1 male- SANC); Mogal Nature
Reserve, Ellisras District, 23º58'S 27º45'E, 25-26.i.1982, 27-29.ii.1984, C.D.
Eardley (1 female, 3 males- SANC); D’ Nyala Nature Reserve, Ellisras
District, 23º45'S 27º49'E, 19.xii.1987, 8.xii.1989, M. W. Mansell (6 females, 5
males- SANC), idem, 10-14.xi.1986, B. Grobbelaar (1 male); Breedsnek,
Crystal Waters, 17.xi.1962, (1 female, 3 males- SANC), H.N. Empey (1
female, 2 males- SANC), Pretoria, Soutpan, 25º24'S 28º06'E, 6.xi.1983, C. D.
Eardley (3 males- SANC); Happy Rest Nature Reserve, 22º55'57'S 29º44'06'E,
8.i.2004, C. D. Eardley (1 male- SANC).

260
Type material: Anthidiellum (Pcynanthidium) krugerense, holotype female-
SANC, South Africa, Kruger National Park, Skukuza, 24º59'S 31º35'E, 14-
17.i.1985, G.L. Prinsloo.
Genus Anthidioctes Moure
Anthidioctes willineri (Moure)
1 female and 1 male. BOLIVIA: Santa Cruz, Santiago, xi.1959 (SAMC).
Genus Anthidioma Pasteels
2 females. NAMIBIA: Diamond Area N º1, Pomona, 20.ix.1980, W.B.
Whitethead (SAMC).
Genus Anthidium Fabricus
Total specimens examined: 33 females and 30 males.
Anthidium Cockerelli (Schwarz)
1 female and 1 male. UNITED STATE OF AMERICA: California, Nev Clark
Co, 3.3 rd mi SE Juanita Spr Ranch S of Riverside, 11-21.v.1983, F.D. Parker
and J.H. Parker (SANC).
Anthidium cordiforme (Friese)
9 females and 11 males. BOTSWANA: Farmers Brigade, 5 km SE of Serowe
Hiiside, i.1986, P. Forchhammer (1 males- SANC); Serowe, vi.1990,
Forchhammer (1 male- SANC). SOUTH AFRICA: Pretoria, 6-14.x.1971, E.
Holm (1 female- SANC); East Transvaal, Wolkberg, 30.viii.1975, R.H.
Watmough (1 female- SANC); Buysdor, 23º01'S 29º22'E, 16.i.1980, C.Kok (1
female- SANC); Wylliespoort, Ingwe Motel, 22º58S 29º57'E, 20-22.i.1982,

261
C.D. Eardley (1 female- SANC); Eureka City, Louw Creek SE , 3.iv.1983,
S.L. Chown (1 female- SANC); Rustenburg Nature Reserve, 25º40'S 27º12'E,
14.iii.1984, M. Mansell (1 female- SANC); 14-29 km W Alldays, 22º44'S
28º57'E, 14.iii.1990, C.D. Eardley (2 males- SANC)Farm Carpe Diem, near
Trichardtsdal, 24º10'S 30º26'E, 30-31.ix.1994, C.D. Eardley & M. Mansell (1
female- SANC); Faerie Glen, Pretoria, 25º44'S 28º13'E, 25.ii.1998, M. Smit (1
male- SANC); Gauteng, Soutpan, 25º24'S 28º06'E, 11.iii.1998, S. Muller (1
male- SANC); Northern Province, Thabahaswa, Groenkom Farm, near
Potgietersrus, 24º03'S 29º02'E, 21-23.ii.2001, M. Stiller (1 female- SANC);
Roodeplaat Research Station, 25º35'S 28º21'E, iii.2004, C.D. Eardley (1 male-
SANC); Gauteng, Soutpan, 25º24'S 28º06'E, 2.iii.2005, C.D. Eardley (1
female, 2 males- SANC); Happy Rest Nature Reserve, 22º55'S 29º44'E,
8.i.2004, C.D. Eardley (1 male- SANC).
Anthidium echinatum (Klug)
1 female and 1 male. MAURITANIA: Akjoujt, Biological Integrated Lucust
Control, xi.1993-ii.1994, J. Tabel (SANC).
Anthidium niveocinctum (Gerstaecker)
4 females and 3 males. SOUTH AFRICA: Waterpoort, SE 29º22'DC, iv.1978,
M. Botha (1 male-SANC); Groothoek Mine, 24º30 Ca, 6.iv.1980, A.P. du Toit
(1 female, 1 male- SANC); Langjan Nature Reserve, 23º52'S 29º14'E, 23-
24.i.1982, C.D. Eardley (1 female- SANC); Ranium, 25º05'S 28º17'E,
3.iii.1989, P. Snyman (1 female- SANC); Schuitdrift Road, 9 km SW
Tshipise, 22º40'S 30º04'E, 4.iii.1990, C.D. Eardley (1 female- SANC).
TANZANIA: Mkomazi Game Reserve, Ibaya, 03º58'S 37º48'E, 8.v.1995, A.
Russell (1 male- SANC).

262
Anthidium paroselae (Cockerell)
1 female and 1 male. UNITED STATE OF AMERICA: California, Nevada,
Clark Co, St Charles Gap, 16.v.1984, F.D. Parker (SANC).
Anthidium pontis (Cockerell)
3 females and 1 male. SOUTH AFRICA; Wilsonfontein, 22º40'S 15º40'E,
8.vii.1975, R.H. WAtmough (1 female-SANC); Langjan Nature Reserve,
23º52'S 29º14'E, 2.ii.1984, C.D. Eardley (1 female- SANC); Rus-en-Vrede,
near Oudtshoorn, 23º24'S 22º23'E, 18.xii.1986, M.W. Mansell (1 female-
SANC); Namaqualand, Hester Malan, 5.xi.1987, M. Struck (1 male-SANC).
Anthidium rodecki (Schwarz)
1 female and 1 male. UNITED STATES OF AMERICA: California, Nevada,
Humbodt Co, 10 mi N Winnemucca, 19.vi.1985, F.D. Parker (SANC).
Anthidium severini (Vachal)
13 females and 13 males. MALAWI: Senga Bay, 20 km NE of Salima, 7-
8.iii.1987, J. & A. Londt Woodland. SOUTH AFRICA: Mondeor,
Johannesburg, 21.x.1961, H.N. Empey (1 female- SANC); Retiefs Kloof,
Rustenberg, 13.iv.1963, H.N. Empey (1 female- SANC); Discovery Florida,
23.xii.1966, H.N. Empey (1 male- SANC); Kruger National Park, Tshokwana,
26.v.1969, L.C. Starke (1 female- SANC); Strubens Valley, florida, 15.xi.1970,
H.N. Empey (1 male- SANC); Pretoria, 9-17.xi.1971, E. Holm (1 male-SANC);
Nylsvley Reserve, x.1978, C.D. Eardley (1 female- SANC); Hennops River, 20
km W Pretoria, 25º74'S 27º55'E, 5.iii.1980, C.D. Eardley (1 male- SANC);
Groenfontein, 35 km E Thabazimbi, 24º34'S 27º45'E, 27.xi.1980, C. Kok (1
female, 1 male- SANC); Diamant, near Bulge Riviere, 24º07'S 27º33'E,
18.i.1983, M.W. Mansell (1 female, 2 males- SANC); Near Vivo, 22º56'S

263
29º33'E, 3.iii.1984, M.W. Mansell (1 male- SANC); Blyderiverspoortdam
Nature Reserve, 24º32'S 30º47'E, 25-26.x.1984, C.D. Eardley (1 female-
SANC); O.T.K Reserve, near Loskopdam, 25º27'S 29º24'E, 9.xii.1985, C.D.
Eardley (1 male- SANC); 5 km E of Thaba Nchu, 29º26'AB, 25.iv.1986, B.
Struckenberg & J. Londt (1 male- SANC); Malta Forest near The Downs,
24º10'S 30º14'E, 3-6.iii.1986, C.D. Eardley (1 male- SANC); Mac Mac Falls,
10 km N Sabie, 25º02'S 30º48'E, 27.ii.1986, B. Grobbelaar (1 female- SANC);
Swadini, Blydepoort Nature Reserve, 24º32'S 30º54'E, 26-29.i.1987, B.
Grobbelaar (1 male- SANC); Langjan Nature Reserve, 23º52'S 29º14'E,
10.iii.1990, C.D. Eardley (1 male- SANC); 14-29 km W Alldays, 22º44'S
28º57'E, 14.iii.1990, C.D. Eardley (4 female- SANC).
Genus Apianthidium Pasteels
1 male. MALAYSIA: Borneo, Sarawak, Mt Matang, 13.xii.1913, G.E.
Bryant (NHML).
Genus Aspidosmia (Brauns)
Type material: Aspidosmia arnoldi (Brauns), lectotype female and
male paralectotype. Willowmore, South Africa, TMSA.
Genus Aztecanthidium Michener and Ordway
Type material: Aztecanthidium tenochtitlanicum Snelling, male and
female paratype: Mexico, Jalisco, C.D. Michener (SEMC).

264
Genus Bathanthidium Mavromoustakis
1 female. SIBERIA: Kudia River, Amagu, vii.1923 (NHML).
Genus Benanthis (Pasteels)
Benanthis madagascariensis (Benoist), 1 male, A. Sekrig, MRAC.
Genus Cyphanthidium Pasteels
Cyphanthidium sheppardii (Mavromoustakis).
1 male. DEMOCRATIC REPUBLIC OF CONGO: Bassin Lukuga,
1934, De Saeger (MRAC).
Genus Dianthidium (Cockerell)
Total specimens examined: 8 females and 8 males.
Dianthidium curvatum (Cockerell)
1 female and 1 male. UNITED STATES OF AMERICA: Utah, Delta,
3.viii.1951, G. E. Bohart (1 female- SANC); Utah, Cacheco, 5 km W Paradise,
4.ix.1987, D. K. Broemeling (1 male, SANC).
Dianthidium cressonii (Dalla Torre)
1 female and 1 male. UNITED STATES OF AMERICA: Utah, Joab, Eureka,
10.viii.1965, G.L. Bohart (1 female- SANC); Utah, Farmington Cyn,
10.vi.1977, F.D. Parker (1 male- SANC).
Dianthidium dubium (Schwarz)
1 female and 1 male. UNITED STATES OF AMERICA: California, Shasta
CO, 2 km W Shingletown 2750´, 16-25.vii.1986, R. Miller (SANC).

265
Dianthidium pudicum (Cresson)
1 female and 1 male. UNITED STATES OF AMERICA: Utah, Farmington
Cyn, 30.vi.1970, P.F. Torchio (1 male- SANC); Utah, Uintah Co, SW
Bonanza, 5000'-5600 T10S R24E, 28.viii.1981, M. Schwartz & R. Miller (1
female- SANC).
Dianthidium subparvum (Swenk)
1 female and 1 male. UNITED STATES OF AMERICA: California, Lassen
Co, 45 m NW, Susanville, 23.vii.1990, P.F. Torchio, G. Trostle (SANC).
Dianthidium ulkei perterritum (Cockerell)
2 females and 2 males. UNITED STATES OF AMERICA: Arizona, Cochise
Co Cave Cr.., 5 km SW, Portal, 4.ix.1986, F. Parker (1 female, 1 male-
SANC); California, Shasta CO, 20 mi N Burney, 25.vii.1990, P.F.Torchio &
G. Trostle (1 female, 1 male- SANC).
Dianthidium parvum (Cresson)
1 female and 1 male. UNITED STATES OF AMERICA: Utah, Uintah Co,
7.ix.1965, R.W. Rust (SANC).
GENUS Duckeanthidium MOURE AND HURD
Type material: Duckeranthidium theieli Michener, male and female
paratype: Costa Rica, R. Thiele (SEMC).
GENUS Eoanthidium POPOV
Total number of specimens examined: 20 females and 13 males.

266
Eoanthidium armaticeps (Friese)
3 female and 2 males. DEMOCRATIC REPUBLIC OF CONGO:
Elizabethville, 31.v.1935, P. Quarré (1 female- MRAC); Elizabethville,
29.iii.1938, H.J. Brédo, (1 female- MRAC). ZIMBABWE: Springvale, S.
Rhodesia, 26.xii.1911 (1 male- SAMC); Lonely Mine, S. Rhodesia, 9.ii.1919
(1 female- SAMC); Sawills, S. Rhodesia, 12.x.1923 (1 male- SAMC).
Eoanthidium bituberculatum (Pasteels)
1 female and 5 males. NAMIBIA: Brandberg, Snusib Ravine, 21º04'S 14º36'E,
2.xii.2000, E. Marais & W. Mey (1 female- SMWN). SOUTH AFRICA:
Spritzkoppe, 17 km Usakos, 21º59'S 15º35'E, 5.vii.1975, 18.vii.1976, R.
Watmough (4 males- SANC); Richtersveld Paradys Kloof, 28º19'S 17º00'E,
11.v.1990, M. Stiller (1 male- SANC).
Eoanthidium rothschildi (Vachal)
14 females and 4 males. NAMIBIA: Etosha, Ondundo Hills, 19º15'S 15º43'E,
10.x-30.xi.1986, E. Griffin. SOUTH AFRICA: Mogal Nature Reserve, Ellisras
Dist, 23º58'S 27º45'E 27-29.ii.1984, C.D. Eardley (1 female- SANC);
Rustenburg Nature Reserve, 22º40'S 27º12'E, 23-26.ii.1981, C.D. Eardley (1
female- SANC); Presidentsrus, near Witbank, 25º41'S 29º22'E, 20.iii.1986,
V.M. Swain (1 female- SANC); Kruger National Park, Pafuri, 22º26'S 31º12'E,
9.iv.1986, L.E.O. Braack (1 female- SANC); 14-29 km W Alldays, 22º44'S
28º57'E, 14.iii.1990, C.D. Eardley (9 females, 3 males- SANC); Between
Nylstroom and Warmbaths, 24º43'S 28º18'E, 18.i.1991, M. Harney (1 female-
SANC).

267
Eoanthidium (Clistanthidium) turnericum (Mavromoustakis)
2 females and 2 males. SOUTH AFRICA: South West Africa, Zesfontein (1
female, 1 male- SAMC); South West Africa, Namib Park, 23º28'S 15º19'E,
16.iii.1983, C.D. Eardley (1 male- SANC); Richtersveld National Park,
28º18'S 16º58'E, 12-14.ix.2001, C.D. Eardley (1 female- SANC).
GENUS Epanthidium MOURE
Epanthidium trigrinum (Schrottky)
1 female and 1 male. ARGENTINA: Salta, El Allsal, i.1990, iv.1991, Fritz
(AMNH).
GENUS Euaspis GERSTAECKER
Total number of specimens examined: 35 females and 5 males.
Euaspis abdominalis (Fabricus)
34 females and 4 males. GHANA: University of Cape Coast, Cape Coast,
05º07'N 01º15'W, 13.ii.2003, N. Akrasi (2 females- SANC). SOUTH
AFRICA: Tongaat, viii.1909, H.C. Burnup (1 female, 1 male- SANC);
Knauzl, 21.xi.1916, Marley (1 female- SANC); Maritzburg, 12.i.1917, C.
Akorman (2 female-SANC); Pretoria, Transvaal, 4.xii.1919 (1 female-SANC);
Gever, Transvaal, 12.xii.1931 (1 female- SANC); Pietersburg, Pretoria,
xii.1961, H. Botha (1 female- SANC); Strubens Val. Transvaal, 5.xii.1961,
H.N. Empey (1 female- SANC); Stanger, Natal, 14.iii.1963, H.N. Empey (1
male- SANC); Nelspriut, xii.1962, iv.1964 (2 females-SANC); Mpumalanga,
Graskop, Mrt. 1969 (2 females- SANC); Pretoria, Retief, i.1970, S. Kohmeyes
(1 female- SANC); Pretoria, P.A. Battey, xii. 1972 (1 female- SANC); Mogoto

268
National Reserve, Zebediela, 24º15'S 29º13'E, 22-25.x.1979, C.D. Eardley (1
female- SANC); Mogoto National Reserve, Zebediela, 24º15'S 29º13'E, 12-
14.iii.1980, G.L. Prinsloo (1 female- SANC); Charters Creek, St Lucia,
28º12'S 32º25'E, 14-16.i.1981, M.W. Mansell (1 female- SANC); Fanies
Island Camp, 28º10'S 32º25'E, 14-16.i.1981, G.L. Prinsloo (1 male- SANC);
O.T.K. Reserve, near, Loskopdam, 25º27'S 29º24'E, 9-11.xii.1985, C.D.
Eardley (1 female- SANC); Cathedral Peak Area above Mike 1s Pass, 28º59'S
29º14'E, 28-29.iii.1985, C.D. Eardley (1 female- SANC); Ludwig, Pretoria,
6.xii.1986 (1 female- SANC); Kosi Bay, 26º58'S 32º48'E, 10-11.ii.1990, C.D.
Eardley (3 females, 1 male- SANC); Kosi Bay, 26º58'S 32º48'E, 10-
11.ii.1990, N. Verheijen (2 females- SANC); Happy Rest, 22º55'S 29º44'E,
8.i.2004, C.D. Eardley (1 female-SANC). ZIMBABWE: Bindura, Rhodesia,
1938 (1 female- SANC); Salisbury, Rhodesia, A. Watsham (1 female-
SAMC).
Euaspis erythros (Meunier)
1 female and 1 male. SOUTH AFRICA: Natal, 1.xii.1930 (1 male- SANC);
Barberton, Transvaal, xii.1978, C.D. Eardley (1 female- SANC).
GENUS Gnathanthidium PASTEELS
3 females. KENYA: Shimoni, iv.1955 (1 female- MRAC).
ZIMBABWE: Bulawayo, S. Rhodesia, 28.xii.1924 (1 female- MRAC);
Nyamandhlovu, S. Rhodesia, 9.i.1960 (1 female- MRAC).

269
GENUS Hoplostelis DOMINIQUE
1 female and 1 male. ARGENTINA: Salta, Cafayate, i. 1992, Fritz
(AMNH).
GENUS Hypathidiodes MOURE
Hypanthidioctes argentina (Urban).
1 female and 1 male. ARGENTINA: Salta, Rosario Lerma, xi. 1993, Fritz, (1
male- AMNH); Salta, El Allsal, iii.1992, Fritz (AMNH).
Hypanthidioctes emarginatum (Urban)
1 female and 1 male. UNITED STATE OF AMERICA: Utah, Uintah Co, SW
Bonanza, 5000'-5600 T10S R24E, 28.viii.1981, M. Schwartz & R. Miller
(SANC).
GENUS Hypanthidium COCKERELL
Hypanthidium flavomarginatum Smith
2 females and 2 males. ARGENTINA: Mendora, 20.x.1908 (TMSA).
Genus Icteranthidium Latreille
Icteranthidium laterale Latreille
1 female & 1 male. ITALY: Ulten, Tirol, Sclett (1 male-SEMC). RUSSIA:
Chotin-Slov, 4.viii.1950, C. Pàdr (1 female-SEMC).
Genus Indanthidium Michener And Griswold
1 female & 1 male. INDIA: Poona, Lonavia, 20.i.1959, F.L. Wain (SEMC)

270
Genus Neanthidium Pasteels
1 female and 1 male. LIBYA: Jebel Soda, Wadi Ghodaifa, 3iii.1952,
4.iii.1952, K.M. Guichard (NHML).
Genus Notanthidium (Isensee)
Notanthidium steloides (Spinola)
1 female and 1 male. CHILE: Canelo, Santiago, 31.i.1949, L.E. Pena (1 male-
SAMC); Ceigual, 1400-1640 m E, 13.i.1955, L.E. Pena (1 female- SAMC).
Genus Pachyanthidium Vachal
Total number of specimens examined: 14 females & 35 males.
Pachyanthidium ausense (Vachal)
3 females & 6 males. BOTSWANA: Farmers Brigade, 5 km SE of Serowe
Hiiside, 1988, xi.1989, P. Forchhammer (2 females- SANC). NAMIBIA: Near
Orange River, 14 Km S Rosh Pinash, 28º17'S 16º55'E, 27.x.1974, R.H.
Watmough (5 male- SANC). SOUTH AFRICA: Richterveld National Park,
Hellskloof Pass, 9.x.1974, H.D. Brown (1 female- SANC); Namaqualand
2917 DB, 26.ix.1987, N.R. Hester Malan, M. Struck (1 female- SANC).
Pachyanthidium benguelense (Vachal)
6 females & 12 males. GHANA: Kakum Conservation Area, 05º20'N
001º22'W, 17.ii.2005, R. Combey (1 female- UCCEM). DEMOCRATIC
REPUBLIC OF CONGO: Elizabethville, 20.ix.23, 8.iv.1933, M. Bequaert (2
males- MRAC); Umtali District, Witchwood Valley, 27.i.1959, A.C. van
Bruggen (1 female- SANC); Salisbury, A. Watsham (1 female- SANC).
MALAWI: Fort Lister, Mt Mulanje, 100m SE, 15.35 DC, 2-4-xii.1983 (1

271
female- SANC). SOUTH AFRICA: Silverton, 6.i.1962, H.N. Empey (1 male-
SANC); Swartkops, 19.x.1963, H.N. Empey (1 female- SANC); Natal,
Milands, Escourt, 5.ii.1975, H.N. Empey (1 male- SANC); Schoemanville,
Dam Wall, 12.x.1975, H.N. Empey (2 males- SANC); Drakenberg, Cathkin
Peak District, 26.x.1976, H.N. Empey (1 male- SANC); Nylsvley Reserve,
x.1978, C.D. Eardley (1 female- SANC); Mogol Nature Reserve, Ellisras
District, 23º58'S 27º45'E, 25-26.i.1982, C.D. Eardley (1 female, 1 male-
SANC); idem, 3.xii.1985, J.S. Donaldson (1 male- SANC); Eastern Transvaal
Cooperative Reserve, near Loskopdam, 25º27'S 29º24'E, 9-11.xii.1985 C.D.
Eardley (1 male- SANC); Guateng, Soutpan, 25º24'S 28º06'E, 2.iii.2005, C.D.
Eardley (1 male- SANC).
Pachyanthidium bicolor (Lepeletier)
1 female & 3 males. GHANA: Kakum Conservation Area, 05º20'N 001º22'W,
18.i.2005, R. Combey (1 male- UCCEM). UGANDA: Entebbe, 30.vii.1913,
C.C. Gowdey (1 male- SANC); 20 km W Kampala, v.1952, E. Pinbey (1
male- KNM); Bwamba, W. Uganda, ii-iii.1957, R. Carcasson (1 female-
KNM).
Pachyanthidium cordatum (Smith)
4 females & 15 males. DEMOCRATIC REPUBLIC OF CONGO: Salisbury,
A. Watsham (1 female- SANC). SOUTH AFRICA: Maritzburg, 23.ii.1917, C.
Akerman (1 female- SANC); Pretorius Kop, 27.x.1950, H.K.Munro (1 male-
SANC); Scottburgh, 15.iii.1963, H.N. Empey (1 female, 1 male- SANC);
Stijdom Tunnel, 19.ii.1969, L.C. Starke, (1 male- SANC); Natal, Tugela
Ferry, 13.xi.1971, H.N. Empey (1 male- SANC); Umtentwini, 20.iv.1973,
H.N.Empey (2 males- SANC); Natal, Hluhluwe Game Reserve, 27º54'S

272
32º22'E, 8.ii.1974, R.H. Watmough (1 male- SANC); Assegaaihoutkloof, near
Groblersdal, 25º15'S 29º25'E, 4.xi.1979, R.H. Watmough; Mogol Nature
Reserve, Ellisras District, 23º58'S 27º45'E, 19-23.xi.1979, S.J. van Tonder, C.
Kok, G.L. Prinsloo & M.W. Mansell (1 male- SANC); Leboea, Mahlasi, 18
km NE Burgersfort, 24º31'S 30º25'E, 24.xi.1981, G.L. Prinsloo (1 male-
SANC); Kuleni Farm, Hluhluwe, 27º54'S 32º22'E, 14.ii.1990, C.D. Eardley (1
male-SANC); Mfongesi, Zululand, W.E.Jones (2 males-SANC); Tang Terr,
Ukerewe, Father Conrads (1 male- SANC). UGANDA: 14 Falls, Thika,
xi.1959, R. Carcasson (2 males- KNM).
Genus Paranthidium Cockerell and Cockerell
1 female and 1 male. UNITED STATES OF AMERICA: Arizonia,
Onion Saddle, Chiricahua Mts., Cochise, 4.ix.1967 (1 male- SANC); Arizona,
Cochise Co., South Western Research Station, 5 mi W Portal, 24.viii.1973 (1
female- AMNH).
GENUS Pleisianthidium CAMERON
Total number of specimens examined: 87 females & 158 males.
Plesianthidium bruneipes (Friese)
5 females & 2 males. SOUTH AFRICA: Namaqualand, Hester Malan,
20.viii.1985, 13.ix.1987, M. Struck (2 females, 1 male- SANC); Kamiesberg
Pass, 30º12'S 17º59'E, 11. ix.1987, C.D. Eardley (1 female, 1 male- SANC);
Farm Dassiefontein, near Kamieskroon, 30º09'S 17º59'E, 1.x.1990, C.D.
Eardley (2 females- SANC).

273
Plesianthidium calescens (Cockerell)
14 females & 21 males. SOUTH AFRICA: Clanwilliam District, Biedouw
Valley, 32º08'S 19º14'E, 5-7.ix.1987, C.D. Eardley (6 females, 5 males-
SANC); Namaqualand, Hester Malan, 20.viii.1985, 14.x.1987, M. Struck (2
females, 3 males- SANC); Pakhuis Pass, 32º08'S 19º02'E, 7.ix.1987, C.D.
Eardley (1 female, 5 males- SANC); Farm Dassiefontein, near Kamieskroon,
30º09'S 17º59'E, 1.x.1990, C.D. Eardley (5 females, 3 males- SANC);
Western Cape Province, near Skurfkop, 31º55'S 18º37'E, 20.ix.2005, C.D.
Eardley (1 male- SANC).
Plesianthidium cariniventre (Friese)
5 females & 16 males. SOUTH AFRICA: Pakhuis Pass, 32º08'S 19º02'E,
21.ix.1985, C.D. Eardley (1 female, 5 males- SANC); Namaqualand, Hester
Malan, 20.viii.1985, 2.ix.1986, 13.ix.1987, M. Struck (1 female,4 males-
SANC); Kamiesberg Pass, 30º12'S 17º59'E, 11. ix.1987, C.D. Eardley (1
female, 5 males- SANC); Nieuwoudtville Botanic Gardens, 31º22'S 19º07'E,
4.ix.1987, C.D. Eardley (1 male- SANC); Wuppertal, 32º17'S 19º13'E,
8.ix.1987, C.D.Eardley (1 male- SANC); Farm Dassiefontein, near
Kamieskroon, 30º09'S 17º59'E, 1.x.1990, C.D. Eardley (2 females- SANC);
Clanwilliam District, Biedouw Valley, 32º08'S 19º14'E, 27.ix.1990,
19.ix.2001, C.D. Eardley (1 female, 2 males- SANC).
Pleisianthidium fulvopilosum (Cameron)
3 females & 4 males. SOUTH AFRICA: Namaqualand, Hester Malan, 20-
28.viii.1985, 5.ix.1986, 12.viii.1986, 13.ix.1987, 16.ix.1987, 9.x.1987, M.
Struck (3 females, 4 males- SANC).

274
Pleisianthidium neli (Brauns)
5 females & 4 males. SOUTH AFRICA: 10 km W of Algeria, Clanwilliam
road, 32º21'S 19º03'E, 4.ix.1987, C.D. Eardley (1 female- SANC);
Kamiesberg Pass, 30º12'S 17º59'E, 11. ix.1987, C.D. Eardley (2 females, 3
males- SANC); Farm Dassiefontein, near Kamieskroon, 30º09'S 17º59'E,
1.x.1990, C.D. Eardley (1 female- SANC); Leliefontein, 30º23'S 18º16'E,
4.x.2003, C. Mayer (1 female, 1 male- SANC).
Pleisianthidium (Spinanthidium) trachusiforme (Friese)
28 females & 64 males. SOUTH AFRICA: Namaqualand, Hester Malan,
20.viii.1985, 2.ix.1986, 13.ix.1987, M. Struck (7 males- SANC); Clanwilliam
District, Biedouw Valley, 32º08'S 19º14'E, 5-7.ix.1987, C.D. Eardley (4
females- SANC); Kamiesberg Pass, 30º12'S 17º59'E, 11. ix.1987, C.D.
Eardley (1 male- SANC); O’okiep, 29º37'S 17º55'E, 13.ix.1987, C.D. Eardley
(2 males- SANC); Farm Dassiefontein, near Kamieskroon, 30º09'S 17º59'E,
1.x.1990, 16-17.ix.2001, C.D. Eardley (22 females, 46 males- SANC); Farm
Arkoep, 6 km N Kamieskroon, 30º19'S 17º56'E, 1-2.x.1990, C.D. Eardley (1
female, 3 males- SANC); 32 km N Nieuwoudtville, 31º16'S 19º46'E,
2.ix.2004, C.D. Eardley (1 female- SANC).
Plesianthidium (Spinanthidiellum) volkmanni (Friese)
27 females & 47 males. SOUTH AFRICA: Stellenbosch, 21.xii.1915, A.
Nicholson (4 females, 2 males- SANC); 5 km from Kuboos, 9.ix.1974, R.H.
Watmough (1 male- SANC); Gydo Pass, 33º14'S 19º20'E, 19.xi.1982,
26.ix.1990, C.D. Eardley (1 female, 4 males- SANC); Near Silverton, 33º34'S
18º23'E, 14.xi.1984, C.D. Eardley (1 male- SANC); Clanwilliam District,
Biedouw Valley, 32º08'S 19º14'E, 30.ix.1986, M.W. Mansell & J.H. Hoffman

275
(1 female, 1 male- SANC); Namaqualand, Hester Malan, 20.viii.1985,
2.ix.1986, 31.viii.1987, 13.ix.1987, M. Struck (7 females, 9 males- SANC);
Nieuwoudtville, 31º20'S 18º53'E, 6.ix.1986, H.C. Hatting (1 female- SANC);
Nieuwoudtville Botanic Gardens, 31º22'S 19º07'E, 9.ix.1987, C.D. Eardley (1
male- SANC); Wuppertal, 32º17'S 19º13'E, 8.ix.1987, C.D.Eardley (1 male-
SANC); Kamiesberg Pass, 30º12'S 17º59'E, 11. ix.1987, C.D. Eardley (1
male- SANC); 15 km E Citrusdal, 32º37'S 19º08'E, 4.x.1990, C.D. Eardley (1
female, 1 male- SANC); Farm Dassiefontein, near Kamieskroon, 30º09'S
17º59'E, 1.x.1990, 16-17.ix.2001, C.D. Eardley (3 females, 4 males- SANC);
Clanwilliam District, Biedouw Valley, 32º08'S 19º14'E, 22.ix.2001, C.D.
Eardley (1 male- SANC); Vanrhynsdorp, near Water Fall, , 31º47'S 18º45'E,
15.ix.2005, C.D. Eardley (5 males- SANC); Western Cape Province, near
Skurfkop, 31º55'S 18º37'E, 20.ix.2005, C.D. Eardley (3 males- SANC);
Western Cape, near Redeinghuys, 32º32'S 18º27'E, C.D. Eardley (1 female, 7
male- SANC); Die Berg, 32º32'S 18º27'E, 18.ix.2005, C.D. Eardley (3 males-
SANC).
Genus Pseudoanthidium Smith
Total number of specimens examined: 78 females & 62 males.
Pseudoanthidium lanificum (Smith)
4 females & 2 males. DEMOCRATIC REPUBLIC OF CONGO: Bena Bendi,
v.1915, R. Mayné (1 female- MRAC); Abumombazi, 18-26.ii.1932, H.J.
Brédo (1 male- MRAC); Eala, 1932, A. Corbisier (1 female- MRAC);
Bambesa, 1933, H.J. Brédo (1 male- MRAC); Eala, xi.1935, viii.1938, J.
Ghesquiére (2 females- MRAC).

276
Pseudoanthidium tertium (Pasteels)
6 females & 4 males. KENYA: Nairobi, E. Shirley (1 female- KNM); Doho
Kosele, Kisii, xi.1950, Mrs Adamson (1 female- KNM); Makneni, v.1951,
B.T. Persons (1 female- KNM). SOUTH AFRICA: Katberg Mt, 32º31'S
26º38'E, 2.xii.1983, G.L. Prinsloo & N.C. Grobbelaar (1 male- SANC);
Lesotho, Katse, 24-28.xii.1991, H. Gertsema (1 female, 2 males- SANC);
Adullam Farm, 28º32'S 28º28'E, 17-18.i.1994, C.D. Eardley (1 female-
SANC); Kaffervoetpad, 15km S of Lydenburg, 25º14'S 30º29'E, 9-
10.xii.1995, R. Stals (1 female- SANC); Hlogoma, 29º48'S 29º32'E, 4-
7.xii.2001, S. van Noort (1 male- SANC).
Pseudoanthidium truncatum (Smith)
53 females & 25 males. DEMOCRATIC REPUBLIC OF CONGO: Eala,
28.ii.1939, J. Ghesquiére (1 female- MRAC); Ubangi, Nouvelle Anvers,
9.xii.1952, P. Basilewsky (1 female- MRAC); Tshuapa, Bokuma, vi, 1952,
R.P. Lootens (1 female- MRAC); Tshuapa, Bamania, ix.1954, R.P. Hulstaert
(1 female- MRAC). GABON: Gamba, Ogoué, 02º42'S 10º01'E, Syssou,
Ngoma, Moussavo (1 female, 1 male- SANC); GHANA: University of Cape
Coast, 05º07'N 01º25'W, N. Akrasi (3 females, 2 males- SANC); Cape Coast,
Antonio Gardens, 05º07'N 01º15'W, 3.xii.2003, C.D. Eardley (1 female, 1
male- SANC); Kakum Conservation Area, 05º20'N 001º23'W, 05º20'N
001º22'W, 18.i.2005, R. Combey (3 males- UCCEM). KENYA: Stoneham, E.
Surr Estate, 4.v.1943, 23.vi.1943, 15.vii.1948 (3 females, 1 male- KNM);
Amani, E. Usambara, iii.1950, E. Pinhey (1 male- KNM); Kakamega Forest
Reserve, v.1949, R.W. Barney (1 female- KNM); Kakamega Forest Reserve,
v.1950, Mrs. Adamson (1 female, 1 male- KNM); Yatta Kitui, iv.1960, R.H.

277
Carcasson (1 female- KNM); Karura, Nairobi, i.1972, M.P. Clifton (1 female-
KNM); Kakamega Forest Reserve, i.1980, M.P. Clifton (3 females- KNM);
Kakamega Forest Reserve, 00º24'N 34º50'E, v.2000, M. Kraemer & G. Velten
(1 male-KNM); Eastern Kerio Valley, 00º44'N 35º34'E, 3.vii.2003, D.
Martins, W. Kinuthia, C.D. Eardley & L. Njoroge (1 male-KNM); Nairobi
City Park, 31.vi.2006, M. Leah (1 male- KNM). SOUTH AFRICA: letaba
Estate, 3.viii.1947 (7 females- SANC); Water Val-Onder, 6.x.1962, H.N.
Empey (1 male- SANC); Malta Forest, 8.ix.1965, 13.ix.1965, 18.x.1974, H.N.
Empey (1 female, 1 male- SANC); Balloon Forest, 24º12'S 30º20'E, 8-
11.iv.1977, D.H. Jacobs (1 female- SANC); Scheeferskloof, near Loskop
Dam, 25º24'S 29º22'E, 3.vi.1978, r.h. Watmough (3 females- SANC); Kloof,
viii.1971, J. Yates (1 female- SANC); Nylsvley Reserve, x.1978, C.D Eardley
(2 females, 6 males- SANC); Barberton, xii, 1978, C.D. Eardley (1 female-
SANC); Pelgrimsrus, 24,30 DC, iv.1979, D.R. Swart (1 female- SANC);
Umzimkulu, 24.xii.1980 (1 female- SANC); De Kuilen, Lydenburg District,
25º10'S 30º32'E, 12.ii.1981, C. Moolman & W. Harrop (1 female- SANC);
Blyderiverspoortdam Nature Reserve, 24º32'S 30º47'E, 25-26.x.1984, C.D.
Eardley (1 female-SANC); Hoedspruit, 24º22'S 30º57'E, 23.xi.1985, S. King
(1 female- SANC); Duiwelskloof, 23º42'S 30º06'E, 15.xii.1985, J.S.
Donaldson (1 male- SANC); Entabeni Forest Reserve, 33º00'S 30º16'E, 7-
11.i.1987, C.D. Eardley (1 male- SANC); Ludwig, Pertorius, 17.i.1987 ( 1
female- SANC); Welgevonden, 24º52S 30º34'E, 11-20.xi.1990, V.M. Vys (1
female- SANC); Nelspruit, 30’25 BD, 11.iv.1994, J.D. Greeff (1 female-
SANC); Tembe Elephant Reserve, 27º03'S 32º25'E, 2.xi.1995. M. W. Mansell
(1 female- SANC); Roodeplaat, 25º41'S 28º18'E, 14.ii.1995, C.D. Eardley (1

278
female- SANC); Kiepersol, 26º10'S 31º07'E, 20.ii.1998, J. Smith (1 female-
SANC); Thabaphaswa, near Potgietersrus, 24º03'S 29º02'E, 21-23.ii.2001, M.
Stiller (1 male- SANC). UGANDA: Bwamba Forest, viii.1928, G.D.H.
Carpenter (1 female- KNM); Bwamba Forest, iii.1948, J.G. Williams (1
female- KNM).
Pseudoanthidium tuberculiferum (Friese)
15 females & 31 males. DEMOCRATIC REPUBLIC OF CONGO: 18 mi E of
Toues River, xii.1962 (1 female- MRAC). SOUTH AFRICA: Lesotho,
Teyateyaneng, 30.xii.1964 C. Jacot-Guillarmod (1 male-SANC); Cedarberg,
20.xii.1972, R.H. Watmough (1 female-SANC); Cathkin Peak, 26.x.1976,
H.N. Empey (1 male- SANC); Adullam Farm, 28º32'S 28º28'E, 20-26.ii.1980,
S.J. van Tonder (1 female- SANC); Natal, Underberg, 29º47'S 29º30'E,
17.xi.1981, S.J. van Tonder (1 male-SANC); Rooiheuwel, 5 km E
Oudtshoorn, 33º36'S 22º17'E, xi.1982, R. H. Watmough (1 male- SANC);
Georgida, 33º26'S 23º20'E, 25.xi.1983, G.L. Prinsloo & N.G. Grobbelaar (2
males- SANC); 20 km N of Silverton, 33º24'S 18º16'E, 14.xi.1984, C.D.
Eardley (4 males- SANC); Near Algeria Forest Reserve, 32º22'S 19º03'E,
14.xi.1984, C.D. Eardley (1 femlae, 1 male- SANC); Tweeling, 27º33'S
28º31'E, 15.i.1986, C.D. Eardley (1 male- SANC); Adullam Farm, 28º32'S
28º28'E, 15-18.i.1986, C.D. Eardley (3 females- SANC); Richtersveld
National Park, 5.x.1986, M. Struck (1 female- SANC); Verlatenkloof, near
Sutherland, 32º33'S 20º37'E, 10.xii.1988, C.D. Eardley (1 female, 2 males-
SANC); Mountain View, Karoo National Park, 32º15'S 22º32'E, 14.xii.1988,
C.D. Eardley (4 male- SANC); Gydo Pass, 33º14'S 19º20'E, 8.xii.1988, C.D.
Eardley (1 female, 1 male- SANC); Montagu Botanical Gardens, 33º47'S

279
20º09'E, 18.xii.1988, C.D. Eardley (1 female, 1 male- SANC); Doring River,
near Nieuwoudtville, 31º18'S 19º09'E, 17.xi.1984, 28.ix.1990, C.D. Eardley (1
female, 1 male- SANC); Arkoep Farm, 6 km N kamiekroon, 30º19'S 17º56'E,
1-2.x.1990, C.D. Eardley (1 female, 1 male- SANC); Pakhuispas, near
Clanwilliam, 32º08'S 18º58'E, 27.ix.1990, C.D. Eardley (1 female- SANC).
Genus Rhodanthidium Spinola
Rhodanthidium septemdentatum (Latrielle)
1 female and 1 male. FRANCE: Var: Plan, vii.1979, 19.vii.1986, F. Schwartz
(AMNH).
Rhodanthidium siculum (Spinola)
2 females and 1 male. MOROCCO: Tanger (1 male-TMSA); Mogador,
ii.1905, escalera. UNITED STATES OF AMERICA: Florida, Bartagena, J.
Cac (1 female- TMSA).
Rhodanthidium fedtschericoi
1 female and 1 male. KAZAKHSTAN: Karataugeb, bei Djulek, Belamurun,
Koshantschilkow (1 female- AMNH). SERBARIA: Achtsha-Kujma,
7.vii.1934, V. Popov (1 male- AMNH).
GENUS Serapista COCKERELL
12 females & 20 males. KENYA: Stoneham, Nerang, 13.v.1952 (1
female-KNM); Bred, Ngong Escarb, xi.1957, P.R.O. Balley (1 male- KNM).
SOUTH AFRICA: Umtata, 15.xi.1920 (1 female, 7 males- SANC); Eshowe,
vi.1925 (1 female, 3 males- SANC); Pretoria, i.1944, Mr. Mogg (1 female-
SANC); Pertoria, i.1964, M.E.du T (1 male- SANC); 8 km N Nieuwodtville,

280
28.ix.1972, H.D. Brown, E. Koster & A. Prinsloo (1 female- SANC); Entabeni
Forest Reserve, 33º00'S 30º16'E, 7-11.i.1987, C.D. Eardley (1 female-
SANC);Groblershoop, 28º53'S 21º59'E, C. Moolman (1 female- SANC);
Langjan, SE 22’ 29 CC, 27.ii.1988, K. Kappmeier (1 male- SANC); Summit
of Mt Gilboa, KwaZulu-Natal, ii.2000, S.D. Johnson (1 female- SANC); 10
km N Concordia, 29º31'S 17º57'E, 23-24.viii.2006, R. Combey (2 females, 7
males- SANC). ZIMBABWE: Rhodesia, Chisawasha, 11.x.1979, A. Watsham
(1 male- SANC); Salisbury, A. Watsham (1 female- SANC).
GENUS Trachusa PANZER
Total number of specimens: 4 females and 7 males.
Trachusa aquilifilum (Strand)
3 females and 1 male.NAMIBIA: South West Africa, Otjimbumbe, iii.1923,
R. Kunene (2 females-SAMC, 1 male- MRAC); South West Africa, Ngosib
Koppie, Etosha National Park, 19º10'S 16º40'E, 9.iv.1989, E. Marais and J.
Irish (1 female- SAMC).
Trachusa schutedeni (Vachal)
2 males. CONGO: Costermansville, 1939, Dr. Hautmann (MRAC)
Trachusa serratula (Panzer)
1 female and 4 males. CHILE: Rostoch, Meche, 4.vi.1968, 28.v.1988,
20.vii.1988, Dr Brauns (TMSA).





























