Embed Size (px)
Citation preview

2004;64:180-188. Cancer Res Thierry Massfelder, Herve Lang, Eric Schordan, et al. for the von Hippel-Lindau Tumor Suppressor GeneFactor for Human Clear Cell Renal Carcinoma and a Target Parathyroid Hormone-Related Protein Is an Essential Growth
Updated version
http://cancerres.aacrjournals.org/content/64/1/180
Access the most recent version of this article at:
Cited Articles
http://cancerres.aacrjournals.org/content/64/1/180.full.html#ref-list-1
This article cites by 37 articles, 17 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/64/1/180.full.html#related-urls
This article has been cited by 11 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on January 9, 2014. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on January 9, 2014. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from

[CANCER RESEARCH 64, 180–188, January 1, 2004]
Parathyroid Hormone-Related Protein Is an Essential Growth Factor for HumanClear Cell Renal Carcinoma and a Target for the von Hippel-LindauTumor Suppressor Gene
Thierry Massfelder,1 Herve Lang,1,2 Eric Schordan,1 Veronique Lindner,1,3 Sylvie Rothhut,1 Sandra Welsch,1
Patricia Simon-Assmann,4 Mariette Barthelmebs,1 Didier Jacqmin,2 and Jean-Jacques Helwig1
1Section of Renovascular Pharmacology and Physiology, Institut National de la Sante et de la Recherche Medicale-University Louis Pasteur, University Louis Pasteur School ofMedicine, Strasbourg; Departments of 2Urology and 3Pathology, University Hospital, Strasbourg; and 4Institut National de la Sante et de la Recherche Medicale U381,Strasbourg-Hautepierre, France
ABSTRACT
Clear cell renal carcinoma (CCRC) is responsible for 2% of cancer-related deaths worldwide and is resistant to virtually all therapies,indicating the importance of a search for new therapeutic targets.Parathyroid hormone-related protein (PTHrP) is a polyprotein derivedfrom normal and malignant cells that regulates cell growth. In thecurrent study, we show that blocking PTHrP with antibodies or an-tagonizing the common parathyroid hormone (PTH)/PTHrP receptor,the PTH1 receptor, dramatically blunts the expansion of human CCRCin vitro by promoting cell death. Importantly, in nude mice, anti-PTHrP antibodies induced complete regression of 70% of the im-planted tumors by inducing cell death. In addition, we demonstratethat the von Hippel-Lindau tumor suppressor protein, which functionsas a gatekeeper for CCRC, negatively regulates PTHrP expression atthe post-transcriptional level. These studies indicate that PTHrP is anessential growth factor for CCRC and is a novel target for the vonHippel-Lindau tumor suppressor protein. Taken together, these resultsstrongly suggest that targeting the PTHrP/PTH1 receptor system mayprovide a new avenue for the treatment of this aggressive cancer inhumans.
INTRODUCTION
Clear cell renal carcinoma (CCRC) represents 75% of all renaltumors. It is responsible for 2% of cancer-related deaths worldwide(200,000 cases and 100,000 deaths/year; Refs. 1, 2). The mortalityrate is 30% at 5 years for localized CCRC, but this rate increases to95% when metastases are present. At the time of diagnosis, about20–30% of patients present with metastases, and 50–60% of patientsdevelop metachronous metastatic disease (1, 2). Nephrectomy, themost widely used therapy for localized CCRC, is associated with poorlong-term survival. Metastatic CCRC is highly resistant to chemo-therapy, radiotherapy, hormonal therapy, and, to a lesser degree,immunotherapy (3). Treatment with IFNs, interleukins (ILs), or both,not only are toxic, but produced brief responses, limited to 20% ofpatients. Thus, effective and nontoxic therapy for CCRC is stilllacking.
Parathyroid hormone-related protein (PTHrP) was initially identi-fied as the factor responsible for the paraneoplasic syndrome, humoralhypercalcemia of malignancy (4). PTHrP has been proven to be apolyprotein normally expressed throughout the body where it displaysa variety of effects, including the regulation of cellular growth,differentiation, and death (5). The perinatal lethality of PTHrP or
PTH1 receptor (PTH1R) knockout mice (4) emphasizes the biologicalimportance of this peptide system. In addition to the skeleton, thekidney appears as a specific target for PTHrP. In this organ, PTHrP isexpressed in vessels, glomeruli, and tubules, and regulates vasculartone, glomerular filtration rate, as well as cell growth and differenti-ation (6).
PTHrP is abundantly expressed by most malignant human tumors,whether they are associated with hypercalcemia or not. Thus, inaddition to its systemic hypercalcemic properties, PTHrP may haveother effects in cancers. Its growth factor-like properties, togetherwith the complex modulation of its expression by a number of growthas well as angiogenic factors such as interleukins, tumor-derivedgrowth factor-� (TGF-�), platelet-derived growth factor (PDGF), andvascular endothelial-derived growth factor (VEGF), point towardpotential roles for PTHrP in the regulation of tumor growth andinvasion (4–6). Recent studies in breast, prostate, and lung cancer(7–9) suggest such roles, and highlight the therapeutic potential ofPTHrP-targeting strategies in human cancer.
CCRC is a highly vascularized tumor, which originates from therenal proximal tubular epithelium (1, 2), a target tissue for PTHrPproliferation effects (10, 11). Moreover, PTHrP is expressed in95% of CCRC in humans, whether they are associated with hy-percalcemia or not (12, 13). In 1990, Burton et al. (14) showed thatPTHrP regulates the proliferation of a human PTHrP-secretingCCRC cell line (SKRC-1) in vitro. Collectively, these studiesstrongly suggest that PTHrP might be involved in CCRC growthand invasion.
Biallelic inactivating mutations of the von Hippel-Lindau (VHL)tumor suppressor gene on chromosome 3p25–26 occur in patientswith the VHL syndrome, an autosomal dominantly inherited syn-drome associated with CCRC, and in most (�75%) patients withsporadic CCRC (1, 15). The products of the VHL gene (pVHL) areVHL (1–213) and VHL (54–213), the latter being generated byinternal translation initiation from an in-frame methionine at codon54. These proteins have been demonstrated to possess identical gate-keeper properties in renal proximal tubular cells. In support of this,reintroduction of the VHL gene in VHL�/� CCRC cells results ingrowth suppression of CCRC tumors in nude mice (16, 17). At themolecular level, pVHLs are components of a ubiquitin-ligase complexinvolved in the down-regulation of several angiogenic and growthfactors, such as VEGF and TGF-�, that contribute to CCRC devel-opment and growth (18).
Herein, we show that PTHrP, interacting with the PTH1R, acts asan essential growth and survival factor for CCRC, both in vitro and invivo. Most importantly, we also show that inhibiting the PTHrP/PTH1R pathway produces substantial regression of CCRC in vivothrough activation of cell death. Finally, we provide evidence thatPTHrP is a novel target for pVHL, and that pVHLs regulate PTHrPexpression at the post-transcriptional level.
Received 7/3/03; revised 10/22/03; accepted 11/3/03.Grant support: Institut National de la Sante et de la Recherche Medicale, University
Louis Pasteur, the French Ligue Contre le Cancer (Comites du Bas-Rhin et du Haut-Rhinet Comite National), and ARC (Association pour la Recherche sur le Cancer).
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
Requests for reprints: Thierry Massfelder, Pharmacologie et Physiologie Renovas-culaires (Equipe Mixte INSERM-ULP 0015), 11 rue Humann, Batiment 4, 1er etage,F67085 Strasbourg Cedex, France. Phone: 333-90-24-34-56; Fax: 333-90-24-34-59;E-mail: [email protected].
180
Research. on January 9, 2014. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from

MATERIALS AND METHODS
Cells, Cell Culture, and Stable Transfection
Normal human (h) proximal tubular cell line HK-2 (a gift from Dr. CarolineSilve, Faculte de Medecine Xavier Bichat, Paris, France) and CCRC cell lines786–0 (American Type Culture Collection, Manassas, Virginia), UOK-126,and UOK-128 (19) were maintained in RPMI 1640 or DMEM medium(Invitrogen, Cergy-Pontoise, France) supplemented with 10% fetal bovineserum. All of the CCRC cell lines are derived from human sporadic CCRCsand encode inactive pVHL. Unless otherwise specified, cells were used at70–80% confluence. 786–0 cells [786–0 wild-type (wt)] were transfectedwith PCR3.1-Uni vector alone (786–0 V) or containing the hVHL cDNAhemagglutinin-tagged at the COOH terminus (786–0 VHL), using Lipo-fectamine (Invitrogen). Stable clones were selected using G418 (500 �g/ml).
RNA Extraction and Reverse Transcription-PCR (RT-PCR) Analysis
Total RNA was extracted from cultured cells using the TRIzol reagent(Invitrogen), according to the manufacturer’s protocol. The relative abundanceof hVHL, hPTHrP, and hPTH1R transcripts was analyzed by RT-PCR usinghuman glyceraldehyde-3-phosphate dehydrogenase (hGAPDH) expression asa loading control. Primers for hVHL, hPTHrP, and hPTH1R were describedpreviously (20–22). Primers for hGAPDH are: sense, 5�-GGAAGGTGAAG-GTCGGAGTC-3�; and antisense, 5�-GCAGTGATGGCATGGACTG-3�.
Analysis of PTHrP Expression
PTHrP concentrations (expressed in pM) in the conditioned medium ofcultured cells were determined by RIA (Bachem, Voisins-le-Bretonneux,France) as described (21).
PTHrP and PTH1R Immunofluorescence
Cells were processed as detailed (23). Affinity-purified polyclonal rabbitantibodies directed against either hPTHrP (1–34; N-term Ab; Bachem) orhPTH1R (Eurogentec, Angers, France) were used at 5 �g/ml. A tetramethyl-rhodamine isothiocyanate-conjugated antirabbit secondary antibody was usedfor detection. As a competition control, the primary antibody was preincubatedovernight at 4°C with 10�6 M PTHrP (1–36) peptide (Bachem) for PTHrPstaining or with 10�6 M of TLDEAERLTEEELH peptide (peptide IV; Euro-gentec) for PTH1R staining. As an additional control, nonimmune rabbit IgG(Sigma-Aldrich, St. Quentin Fallavier, France) was used instead of primaryantibody.
Western Blot Analysis
Protein expression was analyzed as described (22) using 10–30 �g ofprotein and a monoclonal mouse antihemagglutinin antibody (Roche Diagnos-tics, Meylan, France) to detect pVHL or the anti-hPTH1R antibody (Eurogen-tec). A polyclonal mouse anti-� actin antibody (Sigma-Aldrich) was used forvisualization of protein gel loading.
Cell Proliferation Measurements
CCRC cell proliferation was assessed by counting adherent cells and meas-urement of bromodeoxyuridine incorporation (Roche Diagnostics), as de-scribed (21). Cells were grown in serum-free medium for 48 h before stimu-lation and then for a second 48 h with test substances at the concentrationsindicated in the figures. Test substances included rabbit anti-PTHrP antibodiesdirected against either the NH2 terminus (N-term Ab; Bachem), the mid-region(VWR International, Strasbourg, France), or the COOH terminus (a gift fromDr. Pedro Esbrit, Fundacion Jimenez Diaz, Madrid, Spain); the PTH1R an-tagonist [Asn (10), Leu (11), and D-Trp (12)] PTHrP (7–34)amide (Bachem);and hPTHrP (1–36; Bachem), hPTHrP (38–94)amide (a gift from Prof.Andrew Fyfe Stewart, University of Pittsburgh School of Medicine, Pittsburgh,Pennsylvania), or hPTHrP (107–139; Bachem).
Cell Death/Apoptosis Analysis
Cells were treated with control medium alone or with either the N-term Abat 1.5 �g/ml or the PTH1R antagonist at 1 �M for 48 h. Acridine orange and
ethidium bromide were dissolved in PBS buffer and added at 2 �g/ml to theculture medium. Cells were viewed and photographed using fluorescencemicroscopy, either for FITC (acridine orange) or tetramethylrhodamine iso-thiocyanate (ethidium bromide). Internucleosomal DNA fragmentation wasdetected by DNA laddering assay. Cells were resuspended in 300 �l of lysisbuffer [25 mM EDTA, 1% SDS, and 1 mg/ml proteinase K (pH 8.0)] andincubated overnight at 50°C. DNA was extracted using phenol-chloroform,precipitated with ethanol, and analyzed by agarose gel electrophoresis.
PTHrP Promoter Activity Measurement
786–0 wt cells, and 786–0 V and 786–0 VHL clones were transientlytransfected with a construct containing the 5�-flanking promoter regions (pro-moters P1, P2, and P3) of the hPTHrP gene ligated to a promoterless bacterialchloramphenicol acetyltransferase (CAT) gene (24). Transfection was carriedout 24 h after cell seeding with 1 �g/cm2 of the PTHrP promoter-CAT plasmidor the promotorless CAT plasmid, using Lipofectamine (Invitrogen). Cellswere analyzed 48 h after transfection for CAT enzyme expression using a CATenzyme ELISA kit (Roche Diagnostics). Results were normalized and ex-pressed as pg CAT enzyme/mg of protein.
Actinomycin D Experiments
786–0 wt cells, and 786–0 V and 786–0 VHL clones were exposed to 5�g/ml actinomycin D (Sigma) for 0–30 h. Total RNA was isolated and PTHrPexpression analyzed by RT-PCR using hGAPDH for normalization of PCRreactions as described above. Lane intensity was quantified using SigmaScansoftware (Jandel Scientific, Erkrath, Germany) and ratio of hPTHrP onhGAPDH calculated for each time point. The relative intensity of the PTHrPmRNA signal (hPTHrP:hGAPDH ratio) in each lane was then expressed as apercentage of the signal obtained at the time of actinomycin D addition (0 h)set to 100%.
Tumor Model
Implantation. All of the animal studies were in compliance with Frenchanimal use regulations. Seven-week-old male Swiss nu/nu nude mice (Iffa-Credo, St. Germain sur l’Arbresle, France) were given s.c. injections of 106
786–0 cells into the skin of the back, either on both sides (n � 10) or on theright side only (n � 20). Tumor size was measured using calipers. Two weeksafter injection, mice bearing two tumors or one tumor were separated each intwo groups. Mice bearing two tumors were injected i.p. daily with 40 �g of theN-term Ab or with nonspecific IgG (Sigma). Mice bearing 1 tumor wereinjected i.p. daily with 40 �g of the PTH1R antagonist or with the diluent. Atthe end of treatment, mice were anesthetized and blood was harvested formeasurement of plasma electrolytes, creatinine, and PTHrP concentrations(Laboratoires Universitaires d’Analyses Medicales, Strasbourg, France). Tu-mors were then removed, fixed in formalin, and paraffin-embedded. Four�m-thick sections were used. Some sections were stained with H&E and otherswith the PTHrP N-term Ab or the anti-hPTH1R antibody using standardmethods (21).
Proliferative/Apoptotic Index. The proliferative index was determined bystaining tumor sections with a mouse monoclonal anti-hKi67 antibody (Dako,Trappes, France) using standard methods (21). An apoptosis detection kit,based on the terminal deoxynucleotidyl transferase-mediated nick end labelingmethod (Roche Diagnostics), was used to measure the apoptotic index onsections. Total and stained cells in 15 fields (0.25 mm2 each) were counted todetermine both indices, which were then expressed as a percentage of stainedcells to total cells.
Factor VIII and Microvascular Density. Tumor sections were stained forendothelial cells with a rabbit polyclonal antihuman factor VIII antibody(Dako) using a standard immunohistochemistry method (21). Microvesseldensity was determined by counting, for each tumor, both vessel intersectingpoints and the total number of vessels in four to five fields (0.25 mm2 each)showing the highest vascular density.
Statistics
All of the values are expressed as mean � SE. Values were compared usingmultifactorial ANOVA followed by Student-Newman-Keul’s test for multiplecomparisons. P � 0.05 was considered significant.
181
PTHrP IN RENAL CELL CARCINOMA
Research. on January 9, 2014. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from
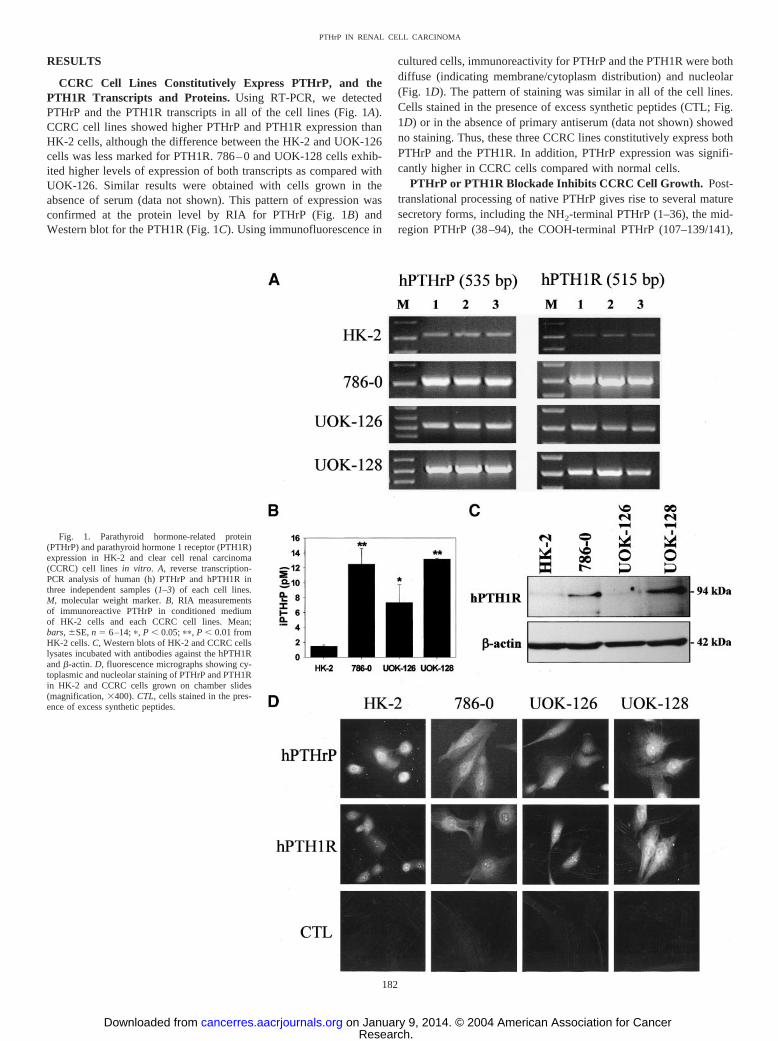
RESULTS
CCRC Cell Lines Constitutively Express PTHrP, and thePTH1R Transcripts and Proteins. Using RT-PCR, we detectedPTHrP and the PTH1R transcripts in all of the cell lines (Fig. 1A).CCRC cell lines showed higher PTHrP and PTH1R expression thanHK-2 cells, although the difference between the HK-2 and UOK-126cells was less marked for PTH1R. 786–0 and UOK-128 cells exhib-ited higher levels of expression of both transcripts as compared withUOK-126. Similar results were obtained with cells grown in theabsence of serum (data not shown). This pattern of expression wasconfirmed at the protein level by RIA for PTHrP (Fig. 1B) andWestern blot for the PTH1R (Fig. 1C). Using immunofluorescence in
cultured cells, immunoreactivity for PTHrP and the PTH1R were bothdiffuse (indicating membrane/cytoplasm distribution) and nucleolar(Fig. 1D). The pattern of staining was similar in all of the cell lines.Cells stained in the presence of excess synthetic peptides (CTL; Fig.1D) or in the absence of primary antiserum (data not shown) showedno staining. Thus, these three CCRC lines constitutively express bothPTHrP and the PTH1R. In addition, PTHrP expression was signifi-cantly higher in CCRC cells compared with normal cells.
PTHrP or PTH1R Blockade Inhibits CCRC Cell Growth. Post-translational processing of native PTHrP gives rise to several maturesecretory forms, including the NH2-terminal PTHrP (1–36), the mid-region PTHrP (38–94), the COOH-terminal PTHrP (107–139/141),
Fig. 1. Parathyroid hormone-related protein(PTHrP) and parathyroid hormone 1 receptor (PTH1R)expression in HK-2 and clear cell renal carcinoma(CCRC) cell lines in vitro. A, reverse transcription-PCR analysis of human (h) PTHrP and hPTH1R inthree independent samples (1–3) of each cell lines.M, molecular weight marker. B, RIA measurementsof immunoreactive PTHrP in conditioned mediumof HK-2 cells and each CCRC cell lines. Mean;bars, �SE, n � 6–14; �, P � 0.05; ��, P � 0.01 fromHK-2 cells. C, Western blots of HK-2 and CCRC cellslysates incubated with antibodies against the hPTH1Rand �-actin. D, fluorescence micrographs showing cy-toplasmic and nucleolar staining of PTHrP and PTH1Rin HK-2 and CCRC cells grown on chamber slides(magnification, �400). CTL, cells stained in the pres-ence of excess synthetic peptides.
182
PTHrP IN RENAL CELL CARCINOMA
Research. on January 9, 2014. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from
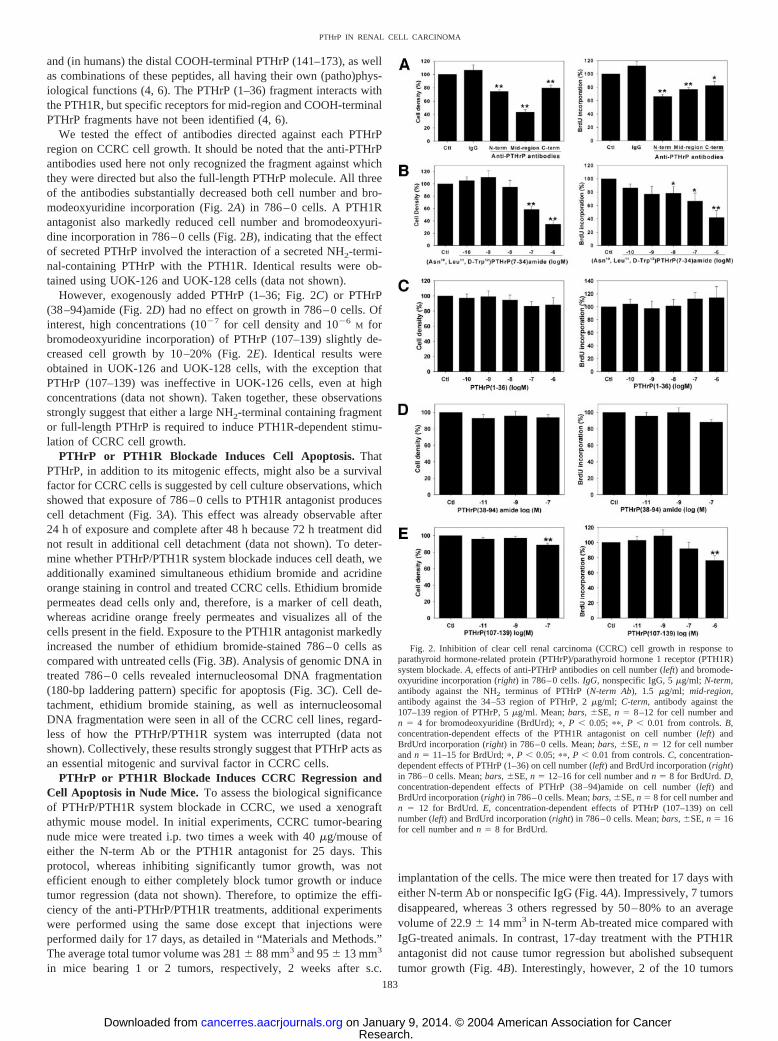
and (in humans) the distal COOH-terminal PTHrP (141–173), as wellas combinations of these peptides, all having their own (patho)phys-iological functions (4, 6). The PTHrP (1–36) fragment interacts withthe PTH1R, but specific receptors for mid-region and COOH-terminalPTHrP fragments have not been identified (4, 6).
We tested the effect of antibodies directed against each PTHrPregion on CCRC cell growth. It should be noted that the anti-PTHrPantibodies used here not only recognized the fragment against whichthey were directed but also the full-length PTHrP molecule. All threeof the antibodies substantially decreased both cell number and bro-modeoxyuridine incorporation (Fig. 2A) in 786–0 cells. A PTH1Rantagonist also markedly reduced cell number and bromodeoxyuri-dine incorporation in 786–0 cells (Fig. 2B), indicating that the effectof secreted PTHrP involved the interaction of a secreted NH2-termi-nal-containing PTHrP with the PTH1R. Identical results were ob-tained using UOK-126 and UOK-128 cells (data not shown).
However, exogenously added PTHrP (1–36; Fig. 2C) or PTHrP(38–94)amide (Fig. 2D) had no effect on growth in 786–0 cells. Ofinterest, high concentrations (10�7 for cell density and 10�6
M forbromodeoxyuridine incorporation) of PTHrP (107–139) slightly de-creased cell growth by 10–20% (Fig. 2E). Identical results wereobtained in UOK-126 and UOK-128 cells, with the exception thatPTHrP (107–139) was ineffective in UOK-126 cells, even at highconcentrations (data not shown). Taken together, these observationsstrongly suggest that either a large NH2-terminal containing fragmentor full-length PTHrP is required to induce PTH1R-dependent stimu-lation of CCRC cell growth.
PTHrP or PTH1R Blockade Induces Cell Apoptosis. ThatPTHrP, in addition to its mitogenic effects, might also be a survivalfactor for CCRC cells is suggested by cell culture observations, whichshowed that exposure of 786–0 cells to PTH1R antagonist producescell detachment (Fig. 3A). This effect was already observable after24 h of exposure and complete after 48 h because 72 h treatment didnot result in additional cell detachment (data not shown). To deter-mine whether PTHrP/PTH1R system blockade induces cell death, weadditionally examined simultaneous ethidium bromide and acridineorange staining in control and treated CCRC cells. Ethidium bromidepermeates dead cells only and, therefore, is a marker of cell death,whereas acridine orange freely permeates and visualizes all of thecells present in the field. Exposure to the PTH1R antagonist markedlyincreased the number of ethidium bromide-stained 786–0 cells ascompared with untreated cells (Fig. 3B). Analysis of genomic DNA intreated 786–0 cells revealed internucleosomal DNA fragmentation(180-bp laddering pattern) specific for apoptosis (Fig. 3C). Cell de-tachment, ethidium bromide staining, as well as internucleosomalDNA fragmentation were seen in all of the CCRC cell lines, regard-less of how the PTHrP/PTH1R system was interrupted (data notshown). Collectively, these results strongly suggest that PTHrP acts asan essential mitogenic and survival factor in CCRC cells.
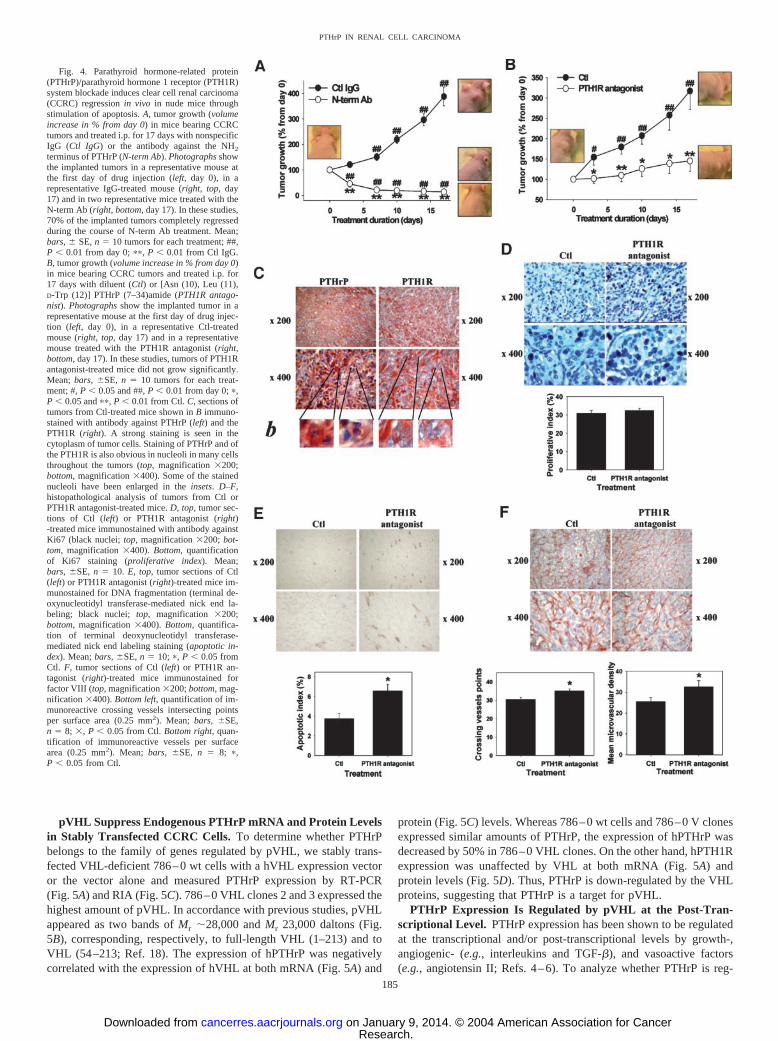
PTHrP or PTH1R Blockade Induces CCRC Regression andCell Apoptosis in Nude Mice. To assess the biological significanceof PTHrP/PTH1R system blockade in CCRC, we used a xenograftathymic mouse model. In initial experiments, CCRC tumor-bearingnude mice were treated i.p. two times a week with 40 �g/mouse ofeither the N-term Ab or the PTH1R antagonist for 25 days. Thisprotocol, whereas inhibiting significantly tumor growth, was notefficient enough to either completely block tumor growth or inducetumor regression (data not shown). Therefore, to optimize the effi-ciency of the anti-PTHrP/PTH1R treatments, additional experimentswere performed using the same dose except that injections wereperformed daily for 17 days, as detailed in “Materials and Methods.”The average total tumor volume was 281 � 88 mm3 and 95 � 13 mm3
in mice bearing 1 or 2 tumors, respectively, 2 weeks after s.c.
implantation of the cells. The mice were then treated for 17 days witheither N-term Ab or nonspecific IgG (Fig. 4A). Impressively, 7 tumorsdisappeared, whereas 3 others regressed by 50–80% to an averagevolume of 22.9 � 14 mm3 in N-term Ab-treated mice compared withIgG-treated animals. In contrast, 17-day treatment with the PTH1Rantagonist did not cause tumor regression but abolished subsequenttumor growth (Fig. 4B). Interestingly, however, 2 of the 10 tumors
Fig. 2. Inhibition of clear cell renal carcinoma (CCRC) cell growth in response toparathyroid hormone-related protein (PTHrP)/parathyroid hormone 1 receptor (PTH1R)system blockade. A, effects of anti-PTHrP antibodies on cell number (left) and bromode-oxyuridine incorporation (right) in 786–0 cells. IgG, nonspecific IgG, 5 �g/ml; N-term,antibody against the NH2 terminus of PTHrP (N-term Ab), 1.5 �g/ml; mid-region,antibody against the 34–53 region of PTHrP, 2 �g/ml; C-term, antibody against the107–139 region of PTHrP, 5 �g/ml. Mean; bars, �SE, n � 8–12 for cell number andn � 4 for bromodeoxyuridine (BrdUrd); �, P � 0.05; ��, P � 0.01 from controls. B,concentration-dependent effects of the PTH1R antagonist on cell number (left) andBrdUrd incorporation (right) in 786–0 cells. Mean; bars, �SE, n � 12 for cell numberand n � 11–15 for BrdUrd; �, P � 0.05; ��, P � 0.01 from controls. C, concentration-dependent effects of PTHrP (1–36) on cell number (left) and BrdUrd incorporation (right)in 786–0 cells. Mean; bars, �SE, n � 12–16 for cell number and n � 8 for BrdUrd. D,concentration-dependent effects of PTHrP (38–94)amide on cell number (left) andBrdUrd incorporation (right) in 786–0 cells. Mean; bars, �SE, n � 8 for cell number andn � 12 for BrdUrd. E, concentration-dependent effects of PTHrP (107–139) on cellnumber (left) and BrdUrd incorporation (right) in 786–0 cells. Mean; bars, �SE, n � 16for cell number and n � 8 for BrdUrd.
183
PTHrP IN RENAL CELL CARCINOMA
Research. on January 9, 2014. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from
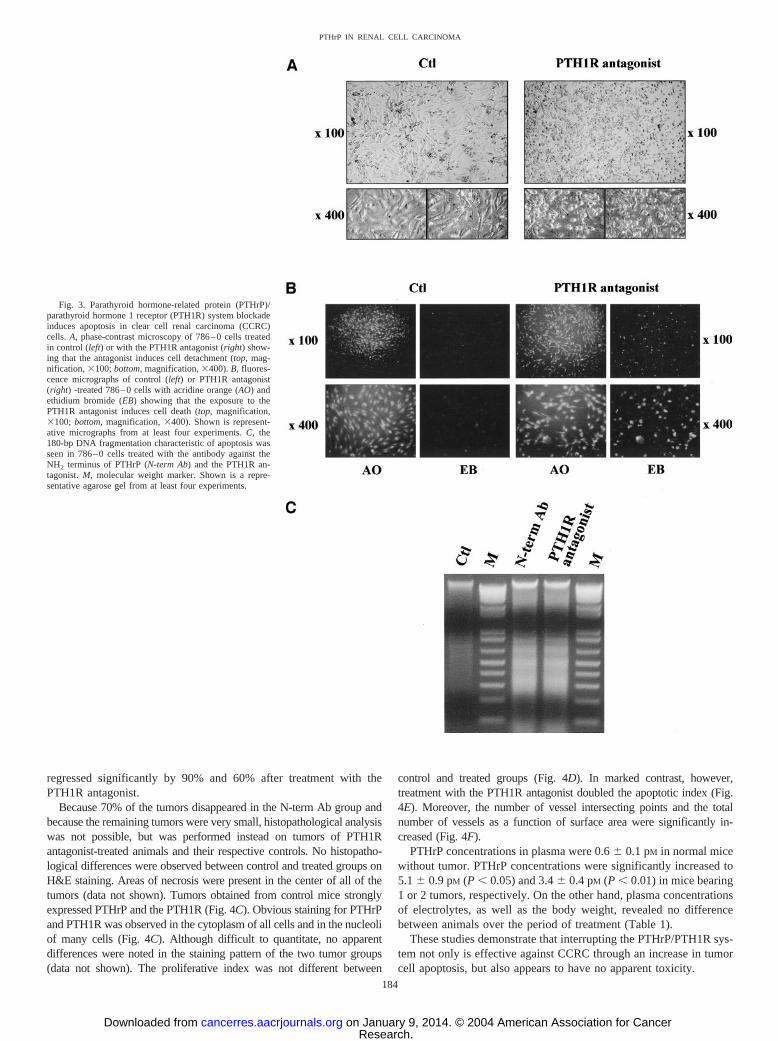
regressed significantly by 90% and 60% after treatment with thePTH1R antagonist.
Because 70% of the tumors disappeared in the N-term Ab group andbecause the remaining tumors were very small, histopathological analysiswas not possible, but was performed instead on tumors of PTH1Rantagonist-treated animals and their respective controls. No histopatho-logical differences were observed between control and treated groups onH&E staining. Areas of necrosis were present in the center of all of thetumors (data not shown). Tumors obtained from control mice stronglyexpressed PTHrP and the PTH1R (Fig. 4C). Obvious staining for PTHrPand PTH1R was observed in the cytoplasm of all cells and in the nucleoliof many cells (Fig. 4C). Although difficult to quantitate, no apparentdifferences were noted in the staining pattern of the two tumor groups(data not shown). The proliferative index was not different between
control and treated groups (Fig. 4D). In marked contrast, however,treatment with the PTH1R antagonist doubled the apoptotic index (Fig.4E). Moreover, the number of vessel intersecting points and the totalnumber of vessels as a function of surface area were significantly in-creased (Fig. 4F).
PTHrP concentrations in plasma were 0.6 � 0.1 pM in normal micewithout tumor. PTHrP concentrations were significantly increased to5.1 � 0.9 pM (P � 0.05) and 3.4 � 0.4 pM (P � 0.01) in mice bearing1 or 2 tumors, respectively. On the other hand, plasma concentrationsof electrolytes, as well as the body weight, revealed no differencebetween animals over the period of treatment (Table 1).
These studies demonstrate that interrupting the PTHrP/PTH1R sys-tem not only is effective against CCRC through an increase in tumorcell apoptosis, but also appears to have no apparent toxicity.
Fig. 3. Parathyroid hormone-related protein (PTHrP)/parathyroid hormone 1 receptor (PTH1R) system blockadeinduces apoptosis in clear cell renal carcinoma (CCRC)cells. A, phase-contrast microscopy of 786–0 cells treatedin control (left) or with the PTH1R antagonist (right) show-ing that the antagonist induces cell detachment (top, mag-nification, �100; bottom, magnification, �400). B, fluores-cence micrographs of control (left) or PTH1R antagonist(right) -treated 786–0 cells with acridine orange (AO) andethidium bromide (EB) showing that the exposure to thePTH1R antagonist induces cell death (top, magnification,�100; bottom, magnification, �400). Shown is represent-ative micrographs from at least four experiments. C, the180-bp DNA fragmentation characteristic of apoptosis wasseen in 786–0 cells treated with the antibody against theNH2 terminus of PTHrP (N-term Ab) and the PTH1R an-tagonist. M, molecular weight marker. Shown is a repre-sentative agarose gel from at least four experiments.
184
PTHrP IN RENAL CELL CARCINOMA
Research. on January 9, 2014. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from
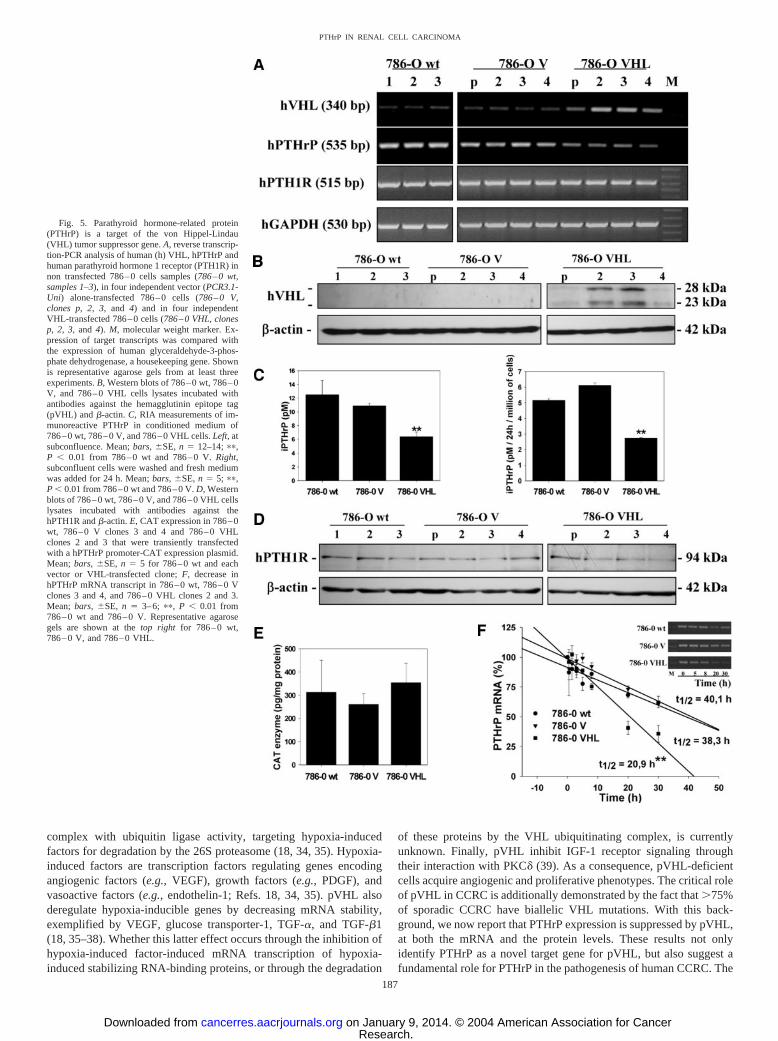
pVHL Suppress Endogenous PTHrP mRNA and Protein Levelsin Stably Transfected CCRC Cells. To determine whether PTHrPbelongs to the family of genes regulated by pVHL, we stably trans-fected VHL-deficient 786–0 wt cells with a hVHL expression vectoror the vector alone and measured PTHrP expression by RT-PCR(Fig. 5A) and RIA (Fig. 5C). 786–0 VHL clones 2 and 3 expressed thehighest amount of pVHL. In accordance with previous studies, pVHLappeared as two bands of Mr �28,000 and Mr 23,000 daltons (Fig.5B), corresponding, respectively, to full-length VHL (1–213) and toVHL (54–213; Ref. 18). The expression of hPTHrP was negativelycorrelated with the expression of hVHL at both mRNA (Fig. 5A) and
protein (Fig. 5C) levels. Whereas 786–0 wt cells and 786–0 V clonesexpressed similar amounts of PTHrP, the expression of hPTHrP wasdecreased by 50% in 786–0 VHL clones. On the other hand, hPTH1Rexpression was unaffected by VHL at both mRNA (Fig. 5A) andprotein levels (Fig. 5D). Thus, PTHrP is down-regulated by the VHLproteins, suggesting that PTHrP is a target for pVHL.
PTHrP Expression Is Regulated by pVHL at the Post-Tran-scriptional Level. PTHrP expression has been shown to be regulatedat the transcriptional and/or post-transcriptional levels by growth-,angiogenic- (e.g., interleukins and TGF-�), and vasoactive factors(e.g., angiotensin II; Refs. 4–6). To analyze whether PTHrP is reg-
Fig. 4. Parathyroid hormone-related protein(PTHrP)/parathyroid hormone 1 receptor (PTH1R)system blockade induces clear cell renal carcinoma(CCRC) regression in vivo in nude mice throughstimulation of apoptosis. A, tumor growth (volumeincrease in % from day 0) in mice bearing CCRCtumors and treated i.p. for 17 days with nonspecificIgG (Ctl IgG) or the antibody against the NH2
terminus of PTHrP (N-term Ab). Photographs showthe implanted tumors in a representative mouse atthe first day of drug injection (left, day 0), in arepresentative IgG-treated mouse (right, top, day17) and in two representative mice treated with theN-term Ab (right, bottom, day 17). In these studies,70% of the implanted tumors completely regressedduring the course of N-term Ab treatment. Mean;bars, � SE, n � 10 tumors for each treatment; ##,P � 0.01 from day 0; ��, P � 0.01 from Ctl IgG.B, tumor growth (volume increase in % from day 0)in mice bearing CCRC tumors and treated i.p. for17 days with diluent (Ctl) or [Asn (10), Leu (11),D-Trp (12)] PTHrP (7–34)amide (PTH1R antago-nist). Photographs show the implanted tumor in arepresentative mouse at the first day of drug injec-tion (left, day 0), in a representative Ctl-treatedmouse (right, top, day 17) and in a representativemouse treated with the PTH1R antagonist (right,bottom, day 17). In these studies, tumors of PTH1Rantagonist-treated mice did not grow significantly.Mean; bars, �SE, n � 10 tumors for each treat-ment; #, P � 0.05 and ##, P � 0.01 from day 0; �,P � 0.05 and ��, P � 0.01 from Ctl. C, sections oftumors from Ctl-treated mice shown in B immuno-stained with antibody against PTHrP (left) and thePTH1R (right). A strong staining is seen in thecytoplasm of tumor cells. Staining of PTHrP and ofthe PTH1R is also obvious in nucleoli in many cellsthroughout the tumors (top, magnification �200;bottom, magnification �400). Some of the stainednucleoli have been enlarged in the insets. D–F,histopathological analysis of tumors from Ctl orPTH1R antagonist-treated mice. D, top, tumor sec-tions of Ctl (left) or PTH1R antagonist (right)-treated mice immunostained with antibody againstKi67 (black nuclei; top, magnification �200; bot-tom, magnification �400). Bottom, quantificationof Ki67 staining (proliferative index). Mean;bars, �SE, n � 10. E, top, tumor sections of Ctl(left) or PTH1R antagonist (right)-treated mice im-munostained for DNA fragmentation (terminal de-oxynucleotidyl transferase-mediated nick end la-beling; black nuclei; top, magnification �200;bottom, magnification �400). Bottom, quantifica-tion of terminal deoxynucleotidyl transferase-mediated nick end labeling staining (apoptotic in-dex). Mean; bars, �SE, n � 10; �, P � 0.05 fromCtl. F, tumor sections of Ctl (left) or PTH1R an-tagonist (right)-treated mice immunostained forfactor VIII (top, magnification �200; bottom, mag-nification �400). Bottom left, quantification of im-munoreactive crossing vessels intersecting pointsper surface area (0.25 mm2). Mean; bars, �SE,n � 8; �, P � 0.05 from Ctl. Bottom right, quan-tification of immunoreactive vessels per surfacearea (0.25 mm2). Mean; bars, �SE, n � 8; �,P � 0.05 from Ctl.
185
PTHrP IN RENAL CELL CARCINOMA
Research. on January 9, 2014. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from

ulated by pVHL at the transcriptional level, we transiently transfected786–0 wt cells, as well as 786–0 V clones 3 and 4, and 786–0 VHLclones 2 and 3 with a CAT expression vector containing the 5�-flanking hPTHrP promoter regions, and measured CAT enzyme incell extracts. The reintroduction of the VHL gene in 786–0 cells didnot alter hPTHrP promoter transcriptional activity (Fig. 5E). Theseresults suggest that the regulation could be post-transcriptional, mostlikely through an effect on mRNA stability. Analysis of the decay ofhPTHrP mRNA in 786–0 wt cells, 786–0 V clones 3 and 4, and786–0 VHL clones 2 and 3 exposed to the transcriptional inhibitoractinomycin D, indeed indicated that the hPTHrP mRNA half-life was38.3 � 3 h and 40.1 � 3.4 h in 786–0 wt and 786–0 V, respectively,but was significantly decreased by 50% to 20.9 � 2.6 h in 786–0VHL (Fig. 5F; P � 0.01). Therefore, pVHL decreased PTHrP ex-pression in CCRC by destabilizing PTHrP mRNA.
DISCUSSION
Previous findings focusing on breast, prostate, and lung cancers(see “Introduction”) strongly indicated that PTHrP not only behavesas a tumor hypercalcemic factor, but also as a factor regulating tumorgrowth. Here we report that the expression of both PTHrP and PTH1Rare common features of CCRC cells and that PTHrP expression issignificantly increased in tumor cells versus normal cells. Most im-portantly, the findings reported herein strongly support the view thatthe PTHrP/PTH1R system is essential for CCRC cell survival. Indeed,the addition of either PTHrP antibodies or blockade of the PTH1Rrapidly induced apoptosis in cultured CCRC cells. Moreover, theCCRC-derived PTHrP species, which acts as a survival factor, is mostlikely an NH2-terminal-containing PTHrP fragment, consistent withthe major circulating PTHrP species in humoral hypercalcemia ofmalignancy (4). Additionally, the COOH-terminal PTHrP fragmentslightly decreased tumor cell growth in two CCRC cell lines. Theinhibitory effect of both the COOH-terminal anti-PTHrP antibody andthe exogenously added COOH-terminal PTHrP fragment we observedon CCRC growth are discrepant. However, it should be stressed firstthat the effect of the COOH-terminal fragment was only observed intwo cell lines and at high concentrations, and second, that the COOH-terminal anti-PTHrP antibody used here not only recognizes theCOOH-terminal part of the PTHrP molecule but also the full-lengthPTHrP. Taken together with the results obtained with the PTH1Rantagonist, these results suggest that the CCRC-derived PTHrP spe-cies, which acts as a survival factor, may contain both NH2- andCOOH-terminal determinants, and that this species may be the majorCCRC-derived PTHrP fragment. Interestingly, investigating COOH-terminal PTHrP (109–141) expression in human CCRC by immuno-histochemistry using a specific antibody, Iwamura et al. (13) haveshown that the expression of the COOH-terminal PTHrP (109–141) inCCRC may be associated with lower CCRC recurrence after radicalnephrectomy. They hypothesized that the COOH-terminal PTHrPfragment might have an inhibitory effect on CCRC cell growth. Ourresults are in accordance with these assumptions, suggesting the
existence of a receptor recognizing a COOH-terminal PTHrP coupledto CCRC growth inhibition. They suggest, however, that the COOH-terminal fragment, if secreted, is a minor secretory form of PTHrP inhuman CCRC. The results obtained by Burton et al. (14) in 1990 onone CCRC cell line (SKRC-1 cells), suggesting that PTHrP canfunction as an autocrine growth factor in human renal cell carcinoma,also support this interpretation. It will be interesting in future studiesto specifically identify the PTHrP species secreted by CCRC cells.
In CCRC cells, PTHrP and the PTH1R were localized to both thecytosol and nucleolar compartments. Nuclear/nucleolar localization ofPTHrP not only has been described in various cell types includingnormal and tumoral cells, but also has been associated with cellsurvival and/or proliferation (6, 23, 25). On the other hand, themolecular targets for PTHrP in the nucleus have not yet been identi-fied. Although the PTH1R has also been seen in the nuclear/nucleolarcompartment in a number of tissues (6, 26, 27), its role in the nucleus,as well as its possible interaction with nuclear PTHrP, is incompletelyunderstood. It is tempting to speculate that, upon secretion by CCRCcells, PTHrP binds to PTH1R, followed by internalization of thereceptor/ligand complex and nuclear entry, with subsequent inhibitionof cell death. However, such a sequence of events has never beendescribed in any cell type. In rat UMR-106.01 osteoblastic cells,PTHrP is targeted to the nucleus through binding to the PTH1R (28).In other cell types, the nuclear presence of PTHrP is a consequence ofeither alternative translational initiation or non-PTH1R-mediated en-docytosis (6, 27). Alternatively, because the PTH1R is coupled toprotein kinase A (PKA) and protein kinase C (PKC) pathways inproximal tubular cells, the antiapoptotic effect of PTHrP in CCRCcells might be related to PKA/PKC stimulation (10). However, thequestion as to whether stimulation of the PKA/PKC pathways byPTHrP might induce proximal tubular cell survival has not yet beenclarified. Specific experiments are required to define the cellularpathways responsible for the apoptotic effect of PTHrP/PTH1Rinhibition.
Another significant finding in the current study was the increasedmicrovessel density in CCRC tumors grown in nude mice in responseto PTHrP/PTH1R system inhibition. This latter observation stronglysuggests that PTHrP is an antiangiogenic factor in CCRC. In supportof this, PTHrP has been shown to behave as an angiogenic or anti-angiogenic factor, depending not only on the cell type but also onwhether it acts in an auto/paracrine or intracrine manner (29–31). Asa rule, vascularization leads to a phase of rapid growth of a giventumor, a prerequisite for the development of invasive processes. Most,but not all, studies show that CCRC is a highly vascularized tumor(32, 33). Indeed, it has been proposed that in CCRC tumors with poorprognosis, decreased microvessel density is associated with the de-velopment of large vessels, which facilitate metastatic spread (33).Thus, the angiogenic effect, i.e., increased microvessel density, inresponse to the anti-PTHrP/PTH1R maneuvers might be beneficialtherapeutically, together with the induction of apoptosis.
pVHL interact with cullin 2, and elongin B and C, forming a
Table 1 Anti-PTHrP/PTH1Ra treatments in nude mice do not affect plasma electrolytes
Untreated mice and mice treated for 17 days according to the different protocols were anesthetized and blood was collected. Concentrations of sodium, phosphate, calcium, andcreatinine in plasma were measured. Animal weights were measured at the beginning (day 0) and at the end (day 17) of each treatment.
Group Sodium mM Phosphate mM Calcium mM Creatinine �MWeight g at
Day 0Weight g at
Day 17 n
Untreated 154.67 � 0.6667 2.84 � 0.2990 2.11 � 0.0568 10.80 � 1.1576 N/A N/A 6Ctl diluent 159.50 � 2.4422 2.36 � 0.3628 2.27 � 0.0619 13.56 � 1.0289 33.18 � 2.9974 33.04 � 2.7394 10PTH1R antagonist 154.44 � 1.5194 2.81 � 0.2017 2.26 � 0.621 13.30 � 0.9551 32.47 � 2.6962 32.89 � 2.7325 10Ctl IgG 155.50 � 0.8660 2.88 � 0.1296 2.21 � 0.0272 8.50 � 0.9574 28.40 � 0.6782 28.60 � 0.6782 5N-term Ab 156.50 � 0.8660 3.27 � 0.0944 2.18 � 0.0263 8.25 � 0.9465 28.40 � 1.2083 29.00 � 1.3038 5
a PTHrP, parathyroid hormone-related protein; PTH1R, parathyroid hormone receptor; N/A, not applicable.
186
PTHrP IN RENAL CELL CARCINOMA
Research. on January 9, 2014. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from
complex with ubiquitin ligase activity, targeting hypoxia-inducedfactors for degradation by the 26S proteasome (18, 34, 35). Hypoxia-induced factors are transcription factors regulating genes encodingangiogenic factors (e.g., VEGF), growth factors (e.g., PDGF), andvasoactive factors (e.g., endothelin-1; Refs. 18, 34, 35). pVHL alsoderegulate hypoxia-inducible genes by decreasing mRNA stability,exemplified by VEGF, glucose transporter-1, TGF-�, and TGF-�1(18, 35–38). Whether this latter effect occurs through the inhibition ofhypoxia-induced factor-induced mRNA transcription of hypoxia-induced stabilizing RNA-binding proteins, or through the degradation
of these proteins by the VHL ubiquitinating complex, is currentlyunknown. Finally, pVHL inhibit IGF-1 receptor signaling throughtheir interaction with PKC� (39). As a consequence, pVHL-deficientcells acquire angiogenic and proliferative phenotypes. The critical roleof pVHL in CCRC is additionally demonstrated by the fact that �75%of sporadic CCRC have biallelic VHL mutations. With this back-ground, we now report that PTHrP expression is suppressed by pVHL,at both the mRNA and the protein levels. These results not onlyidentify PTHrP as a novel target gene for pVHL, but also suggest afundamental role for PTHrP in the pathogenesis of human CCRC. The
Fig. 5. Parathyroid hormone-related protein(PTHrP) is a target of the von Hippel-Lindau(VHL) tumor suppressor gene. A, reverse transcrip-tion-PCR analysis of human (h) VHL, hPTHrP andhuman parathyroid hormone 1 receptor (PTH1R) innon transfected 786–0 cells samples (786–0 wt,samples 1–3), in four independent vector (PCR3.1-Uni) alone-transfected 786–0 cells (786–0 V,clones p, 2, 3, and 4) and in four independentVHL-transfected 786–0 cells (786–0 VHL, clonesp, 2, 3, and 4). M, molecular weight marker. Ex-pression of target transcripts was compared withthe expression of human glyceraldehyde-3-phos-phate dehydrogenase, a housekeeping gene. Shownis representative agarose gels from at least threeexperiments. B, Western blots of 786–0 wt, 786–0V, and 786–0 VHL cells lysates incubated withantibodies against the hemagglutinin epitope tag(pVHL) and �-actin. C, RIA measurements of im-munoreactive PTHrP in conditioned medium of786–0 wt, 786–0 V, and 786–0 VHL cells. Left, atsubconfluence. Mean; bars, �SE, n � 12–14; ��,P � 0.01 from 786–0 wt and 786–0 V. Right,subconfluent cells were washed and fresh mediumwas added for 24 h. Mean; bars, �SE, n � 5; ��,P � 0.01 from 786–0 wt and 786–0 V. D, Westernblots of 786–0 wt, 786–0 V, and 786–0 VHL cellslysates incubated with antibodies against thehPTH1R and �-actin. E, CAT expression in 786–0wt, 786–0 V clones 3 and 4 and 786–0 VHLclones 2 and 3 that were transiently transfectedwith a hPTHrP promoter-CAT expression plasmid.Mean; bars, �SE, n � 5 for 786–0 wt and eachvector or VHL-transfected clone; F, decrease inhPTHrP mRNA transcript in 786–0 wt, 786–0 Vclones 3 and 4, and 786–0 VHL clones 2 and 3.Mean; bars, �SE, n � 3–6; ��, P � 0.01 from786–0 wt and 786–0 V. Representative agarosegels are shown at the top right for 786–0 wt,786–0 V, and 786–0 VHL.
187
PTHrP IN RENAL CELL CARCINOMA
Research. on January 9, 2014. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from

half-life of VEGF, glucose transporter-1, TGF-�, and TGF-�1 tran-scripts has been proven to be markedly increased in VHL-deficientCCRC cells compared with other cell types (36–38), and it has beenproposed that a common mechanism of stabilization of these mRNAsexists in VHL-deficient cells, which takes place at the level of theregulation of specific mRNA-binding stabilizing protein (38).Whether PTHrP belongs to the family of genes directly regulated bysuch mechanisms remains an open question.
CCRC is highly resistant to all of the currently available therapies.The need for new agents for clinical evaluation remains a high priorityin this refractory disease. The current studies argue that agents tar-geting the PTHrP/PTH1R system in human CCRC may have thera-peutic potential.
ACKNOWLEDGMENTS
We thank Dr. Patrick Anglard (Institut of Genetic and Molecular andCellular Biology, Illkirch-Graffenstaden, France) for giving us the UOK-126and UOK-128 cell lines. We also thank Prof. Peter John Ratcliffe (The HenryWellcome Building of Genomic Medicine, Oxford, United Kingdom) forsending us the PCR3.1-Uni vector containing the human hemagglutinin-taggedVHL gene, and Dr. William Philbrick (Department of Internal Medicine, YaleUniversity School of Medicine, New Haven, CT) for the hPTHrP promoters-CAT expression vector. We thank INSERM U381 (Dr. Michele Kedinger,Strasbourg, France) for housing nude mice. We thank Francoise Thirion andFabienne Reymann for technical assistance in immunohistochemistry studies.
REFERENCES
1. Motzer, R. J., Bander, N. H., and Nanus, D. M. Renal-cell carcinoma. N. Engl.J. Med., 335: 865–875, 1996.
2. Pantuck, A. J., Zisman, A., and Belldegrun, A. S. The changing natural history ofrenal cell carcinoma. J. Urol., 166: 1611–1623, 2001.
3. Motzer, R. J., and Russo, P. Systemic therapy for renal cell carcinoma. J. Urol., 163:408–417, 2000.
4. Philbrick, W. M., Wysolmerski, J. J., Galbraith, S., Holt, E., Orloff, J. J., Yang, K. H.,Vasavada, R. C., Weir, E. C., Broadus, A. E., and Stewart, A. F. Defining the rolesof parathyroid hormone-related protein in normal physiology. Physiol. Rev., 76:127–173, 1996.
5. Martin, T. J., Moseley, J. M., and Williams, E. D. Parathyroid hormone-relatedprotein: hormone and cytokine. J. Endocrinol., 154: 23–37, 1997.
6. Massfelder, T., and Helwig, J. J. The parathyroid hormone-related protein system:more data but more unsolved questions. Curr. Opin. Nephrol. Hypertens., 12: 35–42,2003.
7. Luparello, C., Romanotto, R., Tipa, A., Sirchia, R., Olmo, N., Lopez de Silanes, I.,Turnay, J., Lizarbe, A., and Stewart, A. F. Midregion parthyroid hormone-relatedprotein inhibits growth and invasion in vitro and tumorigenesis in vivo of humanbreast cancer cells. J. Bone Miner. Res., 16: 2173–2181, 2001.
8. Dougherty, K. M., Blomme, E. A., Koh, A. J., Henderson, J. E., Pienta, K. J., Rosol,T. J., and Mc Cauley, L. K. Parathyroid hormone-related protein as a growth regulatorof prostate carcinoma. Cancer Res., 59: 6015–6022, 1999.
9. Hastings, R. H., Burton, D. W., Quintana, R. A., Biederman, E., Gujral, A., andDeftos, L. J. Parathyroid hormone-related protein regulates the growth of orthotopichuman lung tumors in athymic mice. Cancer (Phila.), 92: 1402–1410, 2001.
10. Garcia-Ocana, A., De Miguel, F., Penaranda, C., Albar, J. P., Sarasa, J. L., and Esbrit,P. Parathyroid hormone-related protein is an autocrine modulator of rabbit proimaltubule cell growth. J. Bone Miner. Res., 10: 1875–1884, 1995.
11. Garcia-Ocana, A., Galbraith, S. C., Van Why, S. K., Yang, K., Golovyan, L., Dann,P., Zager, R. A., Stewart, A. F., Siegel, N. J., and Orloff, J. J. Expression and role ofparathyroid hormone-related protein in human renal proximal tubule cells duringrecovery from ATP depletion. J. Am. Soc. Nephrol., 10: 238–244, 1999.
12. Gotoh, A., Kitazawa, S., Mizuno, Y., Takenaka, A., Arakawa, S., Matsumoto, O.,Kitazawa, R., Fujimori, T., Maeda, S., and Kamidono, S. Common expression ofparathyroid hormone-related protein and no correlation of calcium level in renal cellcarcinomas. Cancer (Phila.), 71: 2803–2806, 1993.
13. Iwamura, M., Wu, W., Muramoto, M., Ohori, M., Egawa, S., Uchida, T., and Baba,S. Parathyroid hormone-related protein is an independent prognostic factor for renalcell carcinoma. Cancer (Phila.), 86: 1028–1034, 1999.
14. Burton, P. B., Moniz, C., and Knight, D. E. Parathyroid hormone related peptide canfunction as an autocrine growth factor in human renal cell carcinoma. Biochem.Biophys. Res. Commun., 167: 1134–1138, 1990.
15. Friedrich, C. A. Von Hippel-Lindau syndrome. A pleomorphic condition. Cancer(Phila.), 86: 2478–2482, 1999.
16. Gnarra, J. R., Zhou, S., Merrill, M. J., Wagner, J. R., Krumm, A., Papavassiliou, E.,Oldfield, E. H., Klausner, R. D., and Linehan, W. M. Post-transcription regulation ofvascular endothelial growth factor mRNA by the VHL tumor suppressor geneproduct. Proc. Natl. Acad. Sci. USA, 93: 10589–10594, 1996.
17. Iliopoulos, O., Kibel, A., Gray, S., and Kaelin, W. G. Tumor suppression by thehuman Von Hippel-Lindau gene product. Nat. Med., 1: 822–826, 1995.
18. Kondo, K., and Kaelin, W. G. The Von Hippel-Lindau tumor suppressor gene. Exp.Cell Res., 264: 117–125, 2001.
19. Anglard, P., Trahan, E., Liu, S., Latif, F., Merino, M. J., Lerman, M. I., Zbar, B., andLinehan, W. M. Molecular and cellular characterization of human renal cell carci-noma cell lines. Cancer Res., 52: 348–356, 1992.
20. Fukuzumi, M., Hamakawa, H., Onishi, A., Sumida, T., and Tanioka, H. Geneexpression of GLUT isoforms and VHL in oral squamous cell carcinoma. CancerLett., 161: 133–140, 2000.
21. Massfelder, T., Taesch, N., Endlich, N., Eichinger, A., Escande, B., Endlich, K.,Barthelmebs, M., and Helwig, J. J. Paradoxical actions of exogenous and endogenousparathyroid hormone-related protein on renal vascular smooth muscle cell prolifera-tion: reversion in the SHR model of genetic hypertension. FASEB J., 15: 707–718,2001.
22. Massfelder, T., Taesch, N., Fritsch, S., Eichinger, A., Barthelmebs, M., Stewart, A. F.,and Helwig, J. J. Type 1 parathyroid hormone receptor expression level modulatesrenal tone and plasma renin activity in spontaneously hypertensive rat. J. Am. Soc.Nephrol., 13: 639–648, 2002.
23. Massfelder, T., Dann, P., Wu, T. L., Vasavada, R., Helwig, J. J., and Stewart A. F.Opposing mitogenic and anti-mitogenic actions of parathyroid hormone-related pro-tein in vascular smooth muscle cells: a critical role for nuclear targeting. Proc. Natl.Acad. Sci. USA, 94: 13630–13635, 1997.
24. Vasavada, R. C., Wysolmerski, J. J., Broadus, A. E., and Philbrick, W. M. Identifi-cation and characterization of a GC-rich promoter of the human parathyroid hormone-related peptide gene. Mol. Endocrinol., 7: 273–282, 1993.
25. Henderson, J. E., Amizuka, N., Warshawsky, H., Biasotto, D., Lanske, B. M.,Goltzman, D., and Karaplis, A. C. Nucleolar localization of parathyroid hormone-related peptide enhances survival of chondrocytes under conditions that promoteapoptotic cell death. Mol. Cell Biol., 15: 4064–4075, 1995.
26. Watson, P. H., Fraher, L. J., Hendy, G. N., Chung, U. I., Kisiel, M., Natale, B. V., andHodsman, A. B. Nuclear localization of the type I PTH/PTHrP receptor in rat tissues.J. Bone Miner. Res., 15: 1033–1044, 2000.
27. Fiaschi-Taesch, N., and Stewart, A. F. Parathyroid hormone-related protein as anintracrine factor-trafficking mechanisms and functional consequences. Endocrinol-ogy, 144: 407–411, 2003.
28. Lam, M. H., C. M. House, T. Tiganis, Mitchelhill, K. I., Sarcevic, B., Cures, A.,Ramsay, R., Kemp, B. E., Martin, T. J., and Gillespie, M. T. Phosphorylation at thecyclin-dependent kinases site (Thr85) of parathyroid hormone-related protein nega-tively regulates its nuclear localization. J. Biol. Chem., 274: 18559–18566, 1999.
29. Akino, K., Ohtsuru, A., Kanda, K., Yasuda, A., Yamamoto, T., Akino, Y., Naito, S.,Kurokawa, M., Iwahori, N., and Yamashita, S. Parathyroid hormone-related peptideis a potent angiogenic factor. Endocrinology, 141: 4313–4316, 2000.
30. Esbrit, P., Alvarez-Arroyo, M. V., De Miguel, F., Martin, O., Martinez, M. E., andCaramelo, C. C-terminal parathyroid hormone-related protein increases vascularendothelial growth factor in human osteoblastic cells. J. Am. Soc. Nephrol., 11:1085–1092, 2000.
31. Bakre, M. M., Zhu, Y., Yin, H., Burton, D. W., Terkeltaub, R., Deftos, L. J., andVarner, J. A. Parathyroid hormone-related peptide is a naturally occuring, proteinkinase A-dependent angiogenesis inhibitor. Nat. Med., 8: 995–1003, 2002.
32. MacLennan, G. T., and Bostwick, D. G. Microvessel density in renal cell carcinoma:lack of prognostic significance. Urology, 46: 27–30, 1995.
33. Delahunt, B., Bethwaite, P. B., and Thornton, A. Prognostic significance of micro-scopic vascularity for clear cell renal cell carcinoma. Br. J. Urol., 80: 401–404, 1997.
34. Lonergan, K. M., Iliopoulos, O., Ohh, M., Kamura, T., Conaway, R. C., Conaway,J. W., and Kaelin, W. G., Jr. Regulation of hypoxia-inducible mRNAs by the VonHippel Lindau tumor suppressor protein requires binding to complexes containingelongins B/C and Cul2. Mol. Cell Biol., 18: 732–741, 1998.
35. Ivan, M., and Kaelin, W. G. The Von Hippel-Lindau Tumor suppressor protein. Curr.Opin. Genet. Dev., 11: 27–34, 2001.
36. Levy, A. P., Levy, N. S., and Goldberg, M. A. Hypoxia-inducible protein binding tovascular endothelial growth factor mRNA and its modulation by the Von Hippel-Lindau protein. J. Biol. Chem., 271: 25492–25497, 1996.
37. Knebelmann, B., Ananth, S., Cohen, H. T., and Sukhatme, V. P. Transforming growthfactor � is a target for the Von Hippel-Lindau tumor suppressor. Cancer Res., 58:226–230, 1998.
38. Ananth, S., Knebelmann, B., Gruning, W., Dhanabal, M., Walz, G., Stillman, I. E.,and Sukhatme, V. P. Transforming growth factor �1 is a target for the Von Hippel-Lindau tumor suppressor and a critical growth factor for clear cell renal carcinoma.Cancer Res., 59: 2210–2216, 1999.
39. Datta, K., Nambudripad, R., Pal, S., Zhou, M., Cohen, H. T., and Mukhopadhyay, D.Inhibition of insulin-like growth factor I-mediated cell signalling by the Von Hippel-Lindau gene product in renal cancer. J. Biol. Chem., 275: 20700–20706, 2000.
188
PTHrP IN RENAL CELL CARCINOMA
Research. on January 9, 2014. © 2004 American Association for Cancercancerres.aacrjournals.org Downloaded from





























