Embed Size (px)
Citation preview

Palgtontologische Zeitschrift 2008, Vol. 82/4, p. 373-384, 31-12-2008
Oldest record of freshwater sponges (Porifera: Spongillina)- spiculite finds in the Permo-Carboniferous of Europe THOMAS SCHINDLER, SpabrOcken; MICHAEL WUTTKE, Mainz & MARKUS POSCHMANN, Mainz
with 7 figures and 1 table
SCHINDLER, TH.; WUTTKE, M. & POSCHMANN, M. 2008. Oldest record of freshwater sponges (Porifera: Spongillina) - spiculite finds in the Permo-Carboniferous of Europe. - Pal~iontologische Zeitschrift 82 (4): 373-384, 7 figs.. 1 tab., Stuttgart, 31. 12. 2008.
Abstract: Up to now the oldest freshwater siliceous sponges are known from Jurassic deposits of Europe and North America. In this paper stratigraphically older Permo-Carboniferous freshwater siliceous sponges are reviewed/re- ported. Largely overlooked occurrences originate from the French Massif Central (Stefanian B), new discoveries come from the Saar-Nahe Basin (Stefanian C and Autunian) in south-west Germany. All Perrno-Carboniferous finds originate from freshwater lake deposits situated hundreds of kilometers away from the sea. Preserved are monaxone spicules (oxeas) of various length/diameter ratios partly concentrated to spiculites. Gemmoscleres have not been found. A taxonomic classification on family level is not possible. Reasons for the immigration of marine euryhaline sponges into the freshwater might be the conquerence of the newly developed freshwater biotopes on Pangaea after the Variscan Orogeny. Possible migration routes are rivers connecting inland basins with the sea.
Keywords: Porifera ° freshwater sponges ° palaeobiogeography • Upper Carboniferous • Lower Permian ° Europe
Kurzfassung: Bis heute gelten die Stil3wasser-Kieselschw~imme aus jurassischen Ablagerungen yon Europa und Nordamerika als die Altesten. In dieser Arbeit werden stratigraphisch ~ltere Siirwasser-Kieselschw~imme vorgestellt. Weitgehend unbeachtete Funde stammen aus dem franzrsischen Zentralmassiv (Stefan B), neue Funde kommen aus dem siJdwestdeutschen Saar-Nahe-Becken (Stefan C und Autunium). Alle permo-karbonischen Funde stammen aus Seeablagerungen, hunderte von Kilometer vom Meer entfernt. Es liegen monaxone Nadeln (Oxeen) mit variablem L~inge/Durchmesser-Verhgltnis vor, z. T. zu Spiculithen angereichert. Eine taxonomische Zuordnung auf Familienni- veau ist nicht mrglich. Ursachen far die Einwanderung mariner euryhaliner Schw~imme in das StiBwasser kOnnte die Eroberung der nach der Variszischen Gebirgsbildung entstandenen Stigwasser-Biotope sein. Mrgliche Migrations- wege sind Fltisse, die Binnenbecken mit dem Meer verbinden.
Schl/isselwOrter: Porifera • Sti6wasserschwamme • Pal~iobiogeographie ° Ober-Karbon • Unter-Perm • Europa
In t roduct ion
According to widespread opinion (REITNER & WOR- HEIDE 2002) the stratigraphically oldest freshwater sili- ceous sponges are known from Mesozoic deposits (Ju- rassic, no details of location and taxon: DE LAUBENFELS 1955, GRUBER 1994, Eospongilla morrisonensis DUNA- GAN, 1999; Upper Jurassic, North America: WALD- SCHMIDT & LEROY 1944, DUNAGAN 1999, Spongilla purbeckensis HINDE, 1883; Upper Jurassic, England: HINDE 1883, GREENSMITH 1978, Palaeospongilla chu-
butensis OTT d~; VOLKHEIMER, 1972; Lower Cretaceous, South America: YOUNG 1878, OTT & VOLKHEIMER 1972, VOLKMER-RIBEIRO & REITNER 1991, Spongilla patagonica VOLKMER-RIBEIRO & REITNER, 1991; Lo- wer Cretaceous, South America). But CAYEUX (1929: pl. 17 figs. 64-65) described sponge spicules from Up- per Carboniferous freshwater deposits of the French Massif Central preserved in a material similar to chert. This discovery was largely overlooked. Only GEYER (1962) and ZIMMERLE ( 1991) mentioned these findings again without providing further details.
Addresses of the authors: Thomas Schindler, Btiro fiir Pal~iontologie, Stratigraphie und Geotopschutz, Am Wald 11, 55595 Spabrti- cken; e-mail <[email protected]>. - Michael Wuttke & Markus Poschmann. Generaldirektion Kulturelles Erbe Rhein- land-Pfalz, Direktion Landesarch~iologie, Referat Erdgeschichte, Groge Langgasse 29, 55116 Mainz; e-mail <erdgeschichte@ gdke.rlp.de>.
0031-0220/08/0082-373 $ 5.85 © 2008 E. Schweizerbart'sche Verlagsbuchhandlung, D-70176 Stuttgart
374 THOMAS SCHINDLER et al.
A
3 0 °
o
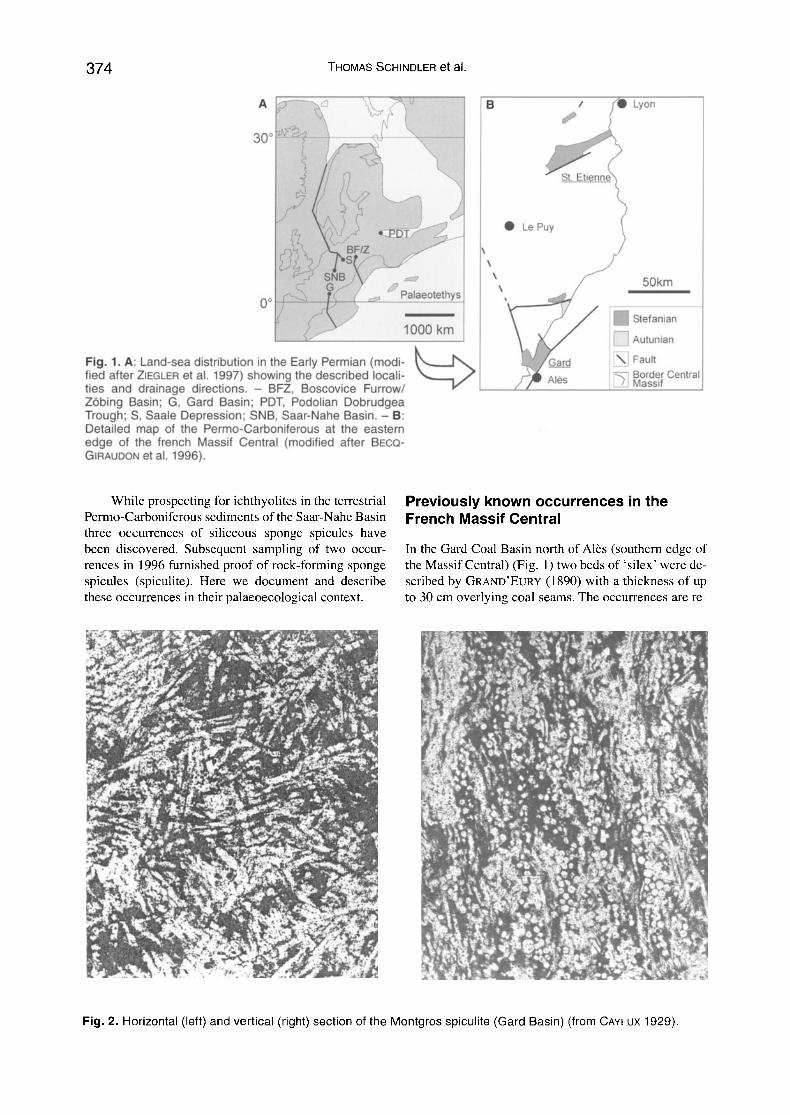
k . Fig. 1. A: Land-sea distribution in the Early Permian (modi- ~ , " , , . . , d~ fied after ZIEGLER et al. 1997) showing the described locali- t y ties and drainage directions. - BFZ, Boscovice Furrow/ Z6bing Basin; G, Gard Basin; PDT, Podolian Dobrudgea Trough; S, Saale Depression; SNB, Saar-Nahe Basin. - B: Detailed map of the Permo-Carboniferous at the eastern edge of the french Massif Central (modified after BECQ- GIRAUDON et al. 1996).
B
\
~:,' ~.~'/0 Lyon
• LeP
50km
• 3 Stefanian
.F~ Autunlan Fau,
~ Border Central Massif
While prospecting for ichthyolites in the terrestrial Permo-Carboniferous sediments of the Saar-Nahe Basin three occurrences of siliceous sponge spicules have been discovered. Subsequent sampling of two occur- rences in 1996 furnished proof of rock-forming sponge spicules (spiculite). Here we document and describe these occurrences in their palaeoecological context.
Previously known occurrences in the French Massif Central
In the Gard Coal Basin north of Albs (southern edge of the Massif Central) (Fig. 1) two beds of 'silex' were de- scribed by GRAND'EURY (1890) with a thickness of up to 30 cm overlying coal seams. The occurrences are re-
Fig. 2. Horizontal (left) and vertical (right) section of the Montgros spiculite (Gard Basin) (from CAYEUX 1929).

Oldest record of freshwater sponges (Spongillina) - spiculite in the Permo-Carboniferous of Europe 3 7 5
corded at Montgros, Gouffre Veyret, La Crouzille and the area of the hard coal concession Doulovy (no further details of location) (CAYEUX 1929). The layers belong to the 'Zone ~ Cordaites' (GRAND'EURY 1877) which corresponds to the Stefanian B (BECQ-GIRAUDON et al. 1996: 238). The associated flora of the coal and adjacent rocks comprises calamites, ferns, seedferns, lepidoden- drons, sigillarians and cordaites (GRAND'EURY 1877). Associated fauna is unknown.
CAYEUX (1929: 299) describes samples of the 'si- lex' from the Doulovy concession as (translated) 'black, massive, compact, translucent at the rims, very finely scaled at the fracture and without granularity, under cor- neous and waxy appearance'. In the outcrop the layer shows bedding planes (CAYEUX 1929).
According to CAYEUX'S (1929) description the chert is a spiculite with thoroughly matted spicules aligned parallel to bedding. The sediment bearing the spicules consists of quartz grains, phyllite clasts, rare pyrite and an additional ore mineral. There are spots of organic material that is cautiously described by CAYEUX ( 1929:301) as an ' unknown form of sapropelitical coal'. Fig. 2 shows a reproduction of the two thin sections (CAYEUX 1929: pl. 17 figs. 64-65).
New discoveries in the Saar-Nahe Basin (SW Germany)
In the Saar-Nahe Basin sponge spicules occur at three localities of different age (Figs. 3-4). These sites are de- scribed in ascending stratigraphic order.
Lemberg near Oberhausen/Nahe, Former Hard Coal Pit 'Heinrichssegen'
Various ruined galleries and fault pits at the western end of the Lemberg near Oberhausen/Nahe (49°46'55,4 '' N, 7°45'28,7 '' E) belong to a disused coal mine. According to JOSTEN (1956) the coal is 'bright coal', i.e. anthracite coalification caused by the rhyodacitic Lemberg intru- sion (Fig. 3).
The succession consists of coal and associated sili- ciclastic sediments of the middle Breitenbach Forma- tion (Ottweiler Subgroup; Upper Carboniferous; ATZ- BACH 1984) (Fig. 4B). Plant remains from the stockpile (layers 9 and 11 a) (Tab. 1) indicate a Late Carboniferous (Stefanian) age (H. KERP pers. comm. 1996). The spiculite bearing horizon (layer 4; Fig. 4B) contains plant debris and faunal elements (Tab. 1).
Thin sections of layer 4 vertical to bedding (Figs. 5A-E) show a black matrix consisting of siliciclastics with much organic detritus. The organic detritus is un- structured and could not be determined microscopically in more detail. In the ground mass there are tightly packed siliceous spicules aligned parallel to bedding. Single calcitic ostracod shells and some fine-grained plant debris are present. Individual bigger quartz aggre- gates represent bone fragments almost completely re- placed by authigenic quartz.
Outcrop at the path to the former lime pit south- east of Friedelhausen
This locality on the edge of the Palatine Anticline (Fig. 3) represents deposits of palaeo-lake Friedelhausen
Fig. 3. Geological map of the Saar-Nahe Basin with detailed maps of Lemberg (left) and of Donnersberg (right) (modified after ATZBACH 1984; HANEKE & LORENZ 2000).
Devontan
Carboniferous Saarbrucken and Ottweder Subgroup
Permocarbonfferous G~an Subgroup
Permian Nahe Subgroup
I Permian Intrusiva
~ Ottweller Subgroup' Sreltenbach Formation
r - ~ - - Glan Subgroup Remlglusberg to
. ' . Wahnwegen Formation ~ Glan Subgroup
Qulrnbach and Lauterecken Formation
~ Glan-Subgroup" Mefsen helm to Thalhchtenberg Formation
Nahe Subgroup l Donnersberg Formation
• , Kuseht
;+ +i L_t~ +
- ll,- i ~ ~ r b r 0 c k e n 1 0 k m i
l l i i i [ T ....... To,t,, L
l .au~;4 ,_~ . . . . d
I ''//'~"'. ~ * ÷ + ~ ÷ + ~] , # / / ~ + + + + + . /
I ;| '." Lemberg / ~,, + + + , • . .+ + ! 'k' .422.1m 1
+ + + + + + + +
+ + 4- + + + + + +
~ + + + + + + ÷ + + ÷ + + + + . ÷ +
Rhyohthe/Dacite - _ - Quaternary Fault i Coal-mining " ~ • Ji Street,Railroad

376 THOMAS SCHINDLER et al.
A
Medard ~lDgns ~
Odenbach ~ berg
Friedelhausen
Legend
[ ~ Coal ,,~
Limestone
[ ] Claystone
m Black Shale
[ ~ Siltstone
~ ] fineqrained Sandstone
~t~ medium to coarse ~ , grained Sandstone
Conglomerate
volcanic Ash
[ ] Lamination
Bioturbation
Mud Crack
Pebble [ ~
Limestone Concretion
Plant Debris
Plant Stem
Sponge Spicule
Leave
Conchostracan
Syncarid Shrimp
Triodus lauterensis
Acanthodian
Elonichthyid Scale
Paramblypterus Scale
Scale indet.
Coprolite
Bone indet.
Godelhausen/Dirmingen Coal LL Lemberg Altenkirchen/Niede rlinxweiler Coal
Habach/Breitenbach Coal Fig. 4. A: Columnar section of Breitenbach to Lauter- ecken formations of the Saar-Nahe Basin with marking of the three finding levels (modified after BoY 1989; CLAUS- ING 1990; SCHINDLER & POSCHMANN 2001). -- B: Detailed section of the findspot Lemberg; spiculite = layer 4.
within the upper Remigiusberg Formation (BoY & SCHINDLER 2000; Fig. 3) (49°32'30,2 '' N, 7o30 ' 16,8" E). Sample FH-I-14 consists of a bioturbated wackestone with cm-size intraclasts that is partially silicified. The flora includes banded calcareous algae and little to very little organic material neither of which can be determined further. The fauna consists of one monaxone crystal-clear siliceous sponge spicule, many arthropod fragments, one indeterminable xenacanthid tooth, smooth as well as sculptured but indeterminable actinopterygian scales, and indeterminable bones and coprolites.
Donnersberg, coal stockpile at Mordkammerhof
This locality on the north-western edge of the Donners- berg (Figs. 3-4) (49°37'23,6 '" N, 7°53'36,4 ' ' E) repre- sents sinks of former galleries and a shaft in hard coal. Due to the Donnersberg Intrusion (Fig. 3) older rocks be- longing to the Quirnbach and Lauterecken formations were uplifted. HANEKE et al. (1983) regarded the coal- bearing part of the succession as belonging to the Lauter- ecken Formation. Due to the associated calcareous rocks the choice for the stratigraphic level of the spiculite bear- ing horizon is limited to the Odenbach Bed or the Medard

O l d e s t record of f reshwater s p o n g e s (Spongi l l ina) - spicul i te in the P e r m o - C a r b o n i f e r o u s of E u r o p e 377
B
T I
J
0 0 0 0 0 0 0 ' ~ 0 0 0 0 0 0 0 0
I o o o o o o , O 0 0 0 0 0 d ~ooooooI oooooo~
r . . . . . ) OOOoOOoO 1 z_- - - - - ' - -E . . . . . . . . . . . . . . . . . . .
1 5 i - - - - - - ~
10
o o "g l I' --T - = . . . . . . . " ' T ' " "1
C Si f S m S c S f G m G c G
C \
i
i, \k
.- Jungenwald Conglomerate Bed
5O
>
5 0
[cml z
E
~' C Si IS mS 5 1-
6~ --' II PY
, , r - - - - - - .
5, i F z z z z z 2 1
P y I ' ]
i ~ ~ C S i f S mS
..... p y
Bed. The profile is no longer accessible. On the stockpile there is coal and calcareous claystone containing smooth actinopterygian scales. There are rare records of oncolitic marlstone (see l~ I s 1921) as well as dark grey marlstone with macroscopic siliceous spicules, ostracod shells, smooth actinopterygian scales and scale-bearing flat oval coprolites. Furthermore there is grey siltstone with nu- merous calamite stems, Asterophyllites and pecopterid remains, as well as grey silty fine-grained sandstone and light grey coarse-grained sandstone.
Thin sections of marlstones bearing sponge spi- cules (Figs. 6 A - D ) show that the matrix is brown and
consists of siliciclastics in silt fraction with many black, opaque, spherical to angular fragments. Some appear to be pyrite crystals, but most are detrital or- ganic substance. The organic detritus is unstructured and could not be determined. In the ground mass there are many sponge spicules tightly packed and not aligned. In addition many calcareous ostracods, few tiny plant remains (up to 500 p,m) and some vertebrate bones and actinopterygian scales are present. All o f the latter contain authigenic quartz that occurs as a selvage but can also replace the ichthyolites completely.

378 THOMAS SCHINDLER et al.
Tab. 1. Findspot Lemberg, fossil content of layers 4, 9 and 11a.
Fossils Layer 4 Layers 9 and l l a
Plants
cf. Codonotheca sp. indet. Pecopteris sp. indet. Pseudomariopteris busqueti Annularia sphenophylloides
Remia pinnatifida Barthelopteris germari
Odontopteris minor/seilleri
Aphlebia indet.
Alethopteris seilleri
Animals
Spongitlina indet.
Syncarid crustaceans: aff. Palaeocaris
Conchostracans indet.
Ostracods indet.
Xenacanthid sharks: Triodus lauterensis, Xenacanthi- dae indet.
Actinopterygians: Elonichthyidae indet., smooth scales of Amino- pterygii indet.
Acanthodii indet.
Coprolites
Taphonomy and diagenesis
laminated sediments from the Saar-Nahe Basin (SCHAFER & STAMM 1989).
In contrast to the strongly silicified cherty spicu- lites of Gard, the Saarland-Palatinate occurrences are only slightly silicified. At Lemberg the slight indura- tion of the siltstones could even be the result of contact metamorphism due to the above mentioned intrusion. The authigenic quartz in the ichthyolites at Lemberg and Donnersberg, however, shows initial silicification by silicic acid released from the spicule opal.
Gard We could only judge the French material from the pub- lished evidence. According to CAYEUX (1929: 300) the sample shows a typical spiculite with spicules oriented in all directions, or grouped in stacks showing a perfect parallelism. The spicules show traces of a central canal (see Lemberg for further remarks).
Lemberg The skeleton material consists of fine-grained quartz that seems almost monocrystalline. The originally opaline skeleton substance was diagenetically trans- formed within the already consolidated sediment via opal-CT to more stable quartz, while the central canal of almost all spicules is preserved (Figs. 5A-B, 5D). This indicates a successive substitution (RICHTER & BASZIO 2000). Due to intergrowth with clay minerals of the sed- iment the outline of the sponge is preserved only vaguely (Fig. 5F). The conversion into quartz probably took place late in diagenesis, but still before the final com- paction of the sediment, as the quartz grains are broken repeatedly due to vertical pressure.
The skeleton of extant freshwater sponges of the sub- order Spongillina consists of isolated spicules and a variable content of spongin, causing these sponges to disintegrate soon after death (RACEK & HARRISON 1974). Thus the spiculites of Gard, Lemberg and Don- nersberg are most likely parautochthonous to alloch- thonous concentrations in the shallow marginal facies of lakes. Decayed sponges were drifted from their pu- tative habitat near the lake shore (see below) and occur as spicule concentrations with partly preserved spicule bundles (Gard, Lemberg). They differ, however, in the degree of alignment. Spicules from the Lemberg local- ity show significant alignment, whereas this is not ob- served in the Gard and Donnersberg occurrences. These articulated spicule bundles (Figs. 5C, E) show that fragments of sponge tissue remained intact until they were buried.
The time interval needed for the formation of a spiculite of a few centimetres in thickness, like the one encountered at Lemberg, is estimated to have ranged from years to decades. This estimate relies on sedimen- tological data available for the formation of lacustrine
Donnersberg Mineralogically the spicules consist of quartz, rarely of calcite. All of them are aggregates of many relatively coarse crystals. Rarely a central canal is evident. The originally opaline skeleton was probably completely dissolved in the already consolidated sediment and the mould later filled with quartz or calcite (see also RICH- TER ~; BASZIO 2000).
Morphological description and supra- generic classification of sponge remnants
Phylum Porifera GRANT, 1836 Class Demospongiae SOLLAS, 1885 Order Haplosclerida TOPSENT, 1928
Suborder Spongillina MANCONI & PRONZATO, 2002
gen. et sp. indet . Figs. 5-6
Oldest record of f reshwater sponges (Spongil l ina) - spicul i te in the Permo-Carboni ferous of Europe 379
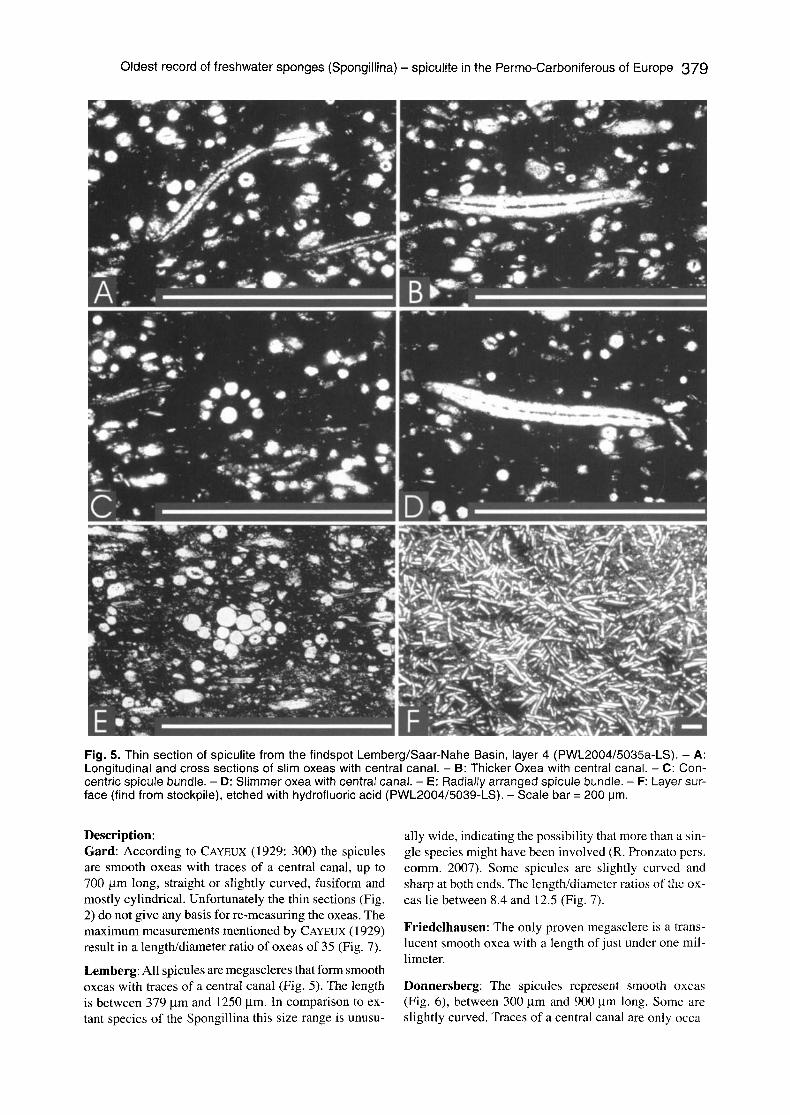
Fig. 5. Thin section of spiculite from the findspot Lemberg/Saar-Nahe Basin, layer 4 (PWL2004/5035a-LS). - A: Longitudinal and cross sections of slim oxeas with central canal. - B: Thicker Oxea with central canal. - C: Con- centric spicule bundle. - D: Sl immer oxea with central canal. - E: Radially arranged spicule bundle. - F: Layer sur- face (find from stockpile), etched with hydrofluoric acid (PWL2004/5039-LS). - Scale bar = 200 t3m.
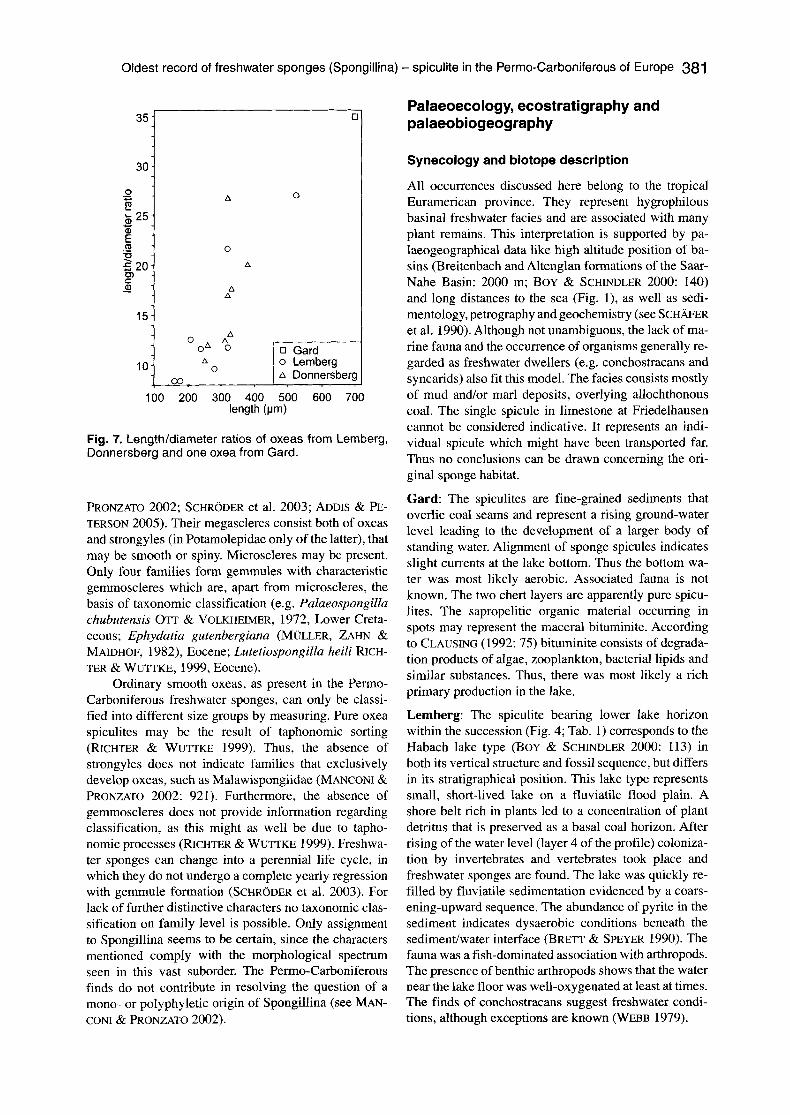
Description: Gard: According to CAYEUX (1929: 300) the spicules are smooth oxeas with traces of a central canal, up to 700 gtm long, straight or slightly curved, fusiform and mostly cylindrical. Unfortunately the thin sections (Fig. 2) do not give any basis for re-measuring the oxeas. The maximum measurements mentioned by CAYEUX (1929) result in a length/diameter ratio of oxeas of 35 (Fig. 7).
Lemberg: All spicules are megascleres that form smooth oxeas with traces of a central canal (Fig. 5). The length is between 379 ~tm and 1250 ~tm. In comparison to ex- tant species of the Spongillina this size range is unusu-
ally wide, indicating the possibility that more than a sin- gle species might have been involved (R. Pronzato pers. comm. 2007). Some spicules are slightly curved and sharp at both ends. The length/diameter ratios of the ox- eas lie between 8.4 and 12.5 (Fig. 7).
Friedelhausen: The only proven megasclere is a trans- lucent smooth oxea with a length of just under one mil- limeter.
Donnersberg: The spicules represent smooth oxeas (Fig. 6), between 300 p.m and 900 ktm long. Some are slightly curved. Traces of a central canal are only occa-
380 THOMAS SCHINDLER et al.
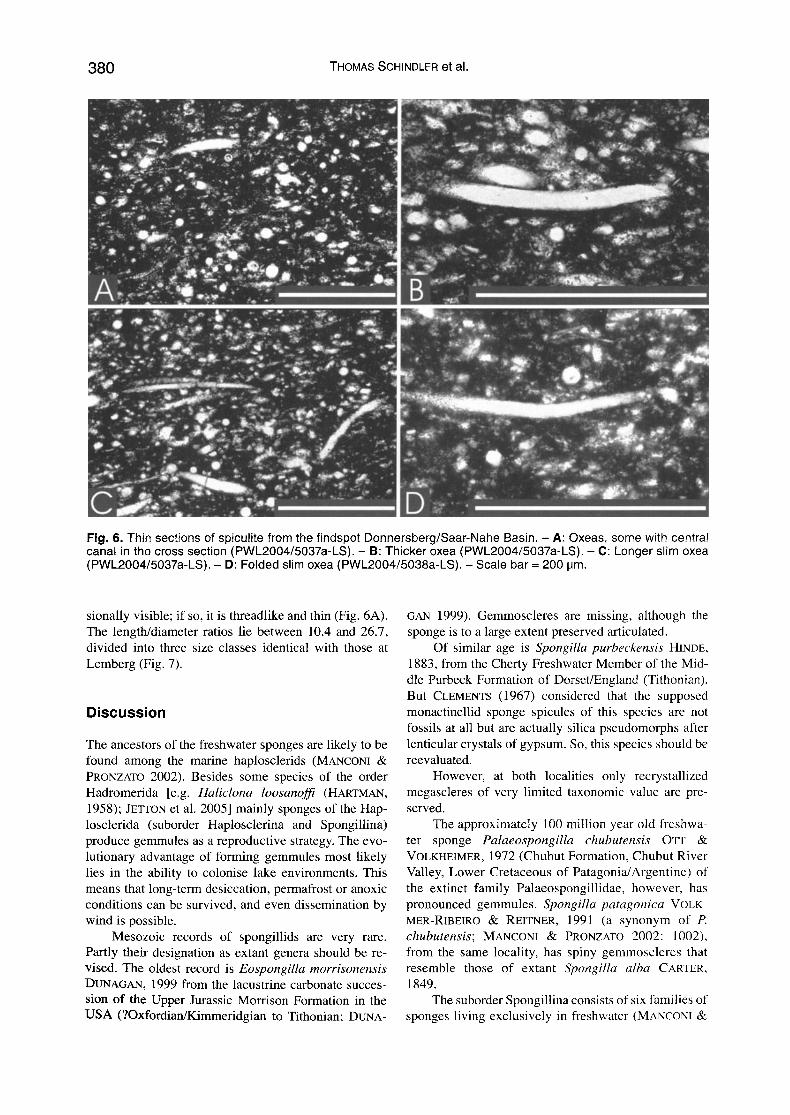
Fig. 6. Thin sect ions of spicul i te f rom the f indspot Donnersberg /Saar -Nahe Basin. - A: Oxeas, some with central canal in the cross sect ion (PWL2004/5037a-LS). - B: Thicker oxea (PWL2004/5037a-LS). - C: Longer sl im oxea (PWL2004/5037a-LS). - D: Folded slim oxea (PWL2004/5038a-LS). - Scale bar = 200 IJm.
sionally visible; if so, it is threadlike and thin (Fig. 6A). The length/diameter ratios lie between 10.4 and 26.7, divided into three size classes identical with those at Lemberg (Fig. 7).
Discussion
The ancestors of the freshwater sponges are likely to be found among the marine haplosclerids (MANCONI 8,~ PRONZATO 2002). Besides some species of the order Hadromerida [e,g. Haliclona loosanoffi (HARTMAN, 1958); JETTON et al. 2005] mainly sponges of the Hap- losclerida (suborder Haploscterina and Spongillina) produce gemmules as a reproductive strategy. The evo- lutionary advantage of forming gemmules most likely lies in the ability to colonise lake environments. This means that long-term desiccation, permafrost or anoxic conditions can be survived, and even dissemination by wind is possible.
Mesozoic records of spongillids are very rare. Partly their designation as extant genera should be re- vised. The oldest record is Eospongilla morrisonensis DUNAGAN, 1999 from the lacustrine carbonate succes- sion of the Upper Jurassic Morrison Formation in the USA (?Oxfordian/Kimmeridgian to Tithonian; DUNA-
GAN 1999). Gemmoscleres are missing, although the sponge is to a large extent preserved articulated.
Of similar age is Spongilla purbeckensis HINDE, 1883, from the Cherty Freshwater Member of the Mid- dle Purbeck Formation of Dorset/England (Tithonian). But CLEMENTS (1967) considered that the supposed monactinellid sponge spicules of this species are not fossils at all but are actually silica pseudomorphs after lenticular crystals of gypsum. So, this species should be reevaluated.
However, at both localities only recrystallized megascleres of very limited taxonomic value are pre- served.
The approximately 100 million year old freshwa- ter sponge Palaeospongilla chubutensis OTT VOLKHE~MER, 1972 (Chubut Formation, Chubut River Valley, Lower Cretaceous of Patagonia/Argentine) of the extinct family Palaeospongillidae, however, has pronounced gemmules. Spongilla patagonica VOLK- MER-RIBEIRO & REITNER, 1991 (a synonym of P. chubutensis; MANCONI & PRONZATO 2002: 1002), from the same locality, has spiny gemmoscleres that resemble those of extant Spongilla alba CARTER, 1849.
The suborder Spongillina consists of six families of sponges living exclusively in freshwater (MANCONI &
Oldest record of freshwater sponges (Spongillina) -spicul i te in the Permo-Carboniferous of Europe 381
35
30
0
15
0 0 ~ 0
lO o
100 200 300
O
25 E
20 e.-
[ ]
Zx O
rn Gard o Lemberg
400 500 600 700 length (IJm)
Fig. 7. Length/diameter ratios of oxeas from Lemberg, Donnersberg and one oxea from Gard.
PRONZATO 2002; SCHRODER et al. 2003; ADDIS & PE- TERSON 2005). Their megascleres consist both of oxeas and strongyles (in Potamolepidae only of the latter), that may be smooth or spiny. Microscleres may be present. Only four families form gemmules with characteristic gemmoscleres which are, apart from microscleres, the basis of taxonomic classification (e.g. Palaeospongilla chubutensis OTT & VOLKHEIMER, 1972, Lower Creta- ceous; Ephydatia gutenbergiana (MOLLER, ZAHN & MAIDHOF, 1982), Eocene; Lutetiospongilla heili RICH- TER & WUTTKE, 1999, Eocene).
Ordinary smooth oxeas, as present in the Permo- Carboniferous freshwater sponges, can only be classi- fied into different size groups by measuring. Pure oxea spiculites may be the result of taphonomic sorting (RICHTER & WUTTKE 1999). Thus, the absence of strongyles does not indicate families that exclusively develop oxeas, such as Malawispongiidae (MANCONI & PRONZATO 2002: 921). Furthermore, the absence of gemmoscleres does not provide information regarding classification, as this might as well be due to tapho- nomic processes (RICHTER & WUTTKE 1999). Freshwa- ter sponges can change into a perennial life cycle, in which they do not undergo a complete yearly regression with gemmule formation (SCHRODER et al. 2003). For lack of further distinctive characters no taxonomic clas- sification on family level is possible. Only assignment to Spongillina seems to be certain, since the characters mentioned comply with the morphological spectrum seen in this vast suborder. The Permo-Carboniferous finds do not contribute in resolving the question of a mono- or polyphyletic origin of Spongillina (see MAN- CONI & PRONZATO 2002).
Palaeoecology, ecostratigraphy and palaeobiogeography
Synecology and biotope description
All occurrences discussed here belong to the tropical Euramerican province. They represent hygrophilous basinal freshwater facies and are associated with many plant remains. This interpretation is supported by pa- laeogeographical data like high altitude position of ba- sins (Breitenbach and Altenglan formations of the Saar- Nahe Basin: 2000 m; BoY & SCHINDLER 2000: 140) and long distances to the sea (Fig. 1), as well as sedi- mentology, petrography and geochemistry (see SCHAFER et al. 1990). Although not unambiguous, the lack of ma- rine fauna and the occurrence of organisms generally re- garded as freshwater dwellers (e.g. conchostracans and syncarids) also fit this model. The facies consists mostly of mud and/or marl deposits, overlying allochthonous coal. The single spicule in limestone at Friedelhausen cannot be considered indicative. It represents an indi- vidual spicule which might have been transported far. Thus no conclusions can be drawn concerning the ori- ginal sponge habitat.
Card : The spiculites are fine-grained sediments that overlie coal seams and represent a rising ground-water level leading to the development of a larger body of standing water. Alignment of sponge spicules indicates slight currents at the lake bottom. Thus the bottom wa- ter was most likely aerobic. Associated fauna is not known. The two chert layers are apparently pure spicu- lites. The sapropelitic organic material occurring in spots may represent the maceral bituminite. According to CLAUSING (1992: 75) bituminite consists of degrada- tion products of algae, zooplankton, bacterial lipids and similar substances. Thus, there was most likely a rich primary production in the lake.
Lemberg: The spiculite bearing lower lake horizon within the succession (Fig. 4; Tab. 1) corresponds to the Habach lake type (BoY & SCHINDLER 2000: 113) in both its vertical structure and fossil sequence, but differs in its stratigraphical position. This lake type represents small, short-lived lake on a fluviatile flood plain. A shore belt rich in plants led to a concentration of plant detritus that is preserved as a basal coal horizon. After rising of the water level (layer 4 of the profile) coloniza- tion by invertebrates and vertebrates took place and freshwater sponges are found. The lake was quickly re- filled by fluviatile sedimentation evidenced by a coars- ening-upward sequence. The abundance of pyrite in the sediment indicates dysaerobic conditions beneath the sediment/water interface (BRETT & SPEYER 1990). The fauna was a fish-dominated association with arthropods. The presence of benthic arthropods shows that the water near the lake floor was well-oxygenated at least at times. The finds of conchostracans suggest freshwater condi- tions, although exceptions are known (WEBB 1979).

382 THOMAS SCHINDLER et al.
Donnersberg: At this locality, a large shallow lake de- veloped on a fluviatile flood plain by a rise of the water level (transgression) (J. BOY pers. comm. 1994). Abun- dant hygrophilous shore vegetation led to concentration of allochthonous plant detritus, evidenced by coal and plant-bearing siltstone. Due to further rising of the wa- ter level and an absence of siliciclastic supply calcare- ous sediments like marlstone and oncolites formed. Here freshwater sponges and benthic animals like ostra- cods are found, indicating aerobic lake bottom condi- tions. Nektonic animals include actinopterygians (most likely Paramblypterus sp.) and piscivorous fish such as freshwater sharks that are only represented by their co- prolites. The fauna is an arthropod-bearing amblypterid- dominated fish association. It is, however, less varied than at the Lemberg locality. The benthos comprises an abundance of a single species of ostracod.
The ecological parameters of all of these freshwater lakes in a marginal tropical climate (BoY & SCHINDLER 2000) correspond to the requirements of extant lacustrine freshwater sponges such as Nudospongilla ANNANDALE, 1918 (MANCONI & PRONZATO 2002: fig. 47).
Autoecology of Permo-Carboniferous freshwater sponges Sponges are filter feeders. It can be assumed that this is also true for the Permo-Carboniferous freshwater sponges. Extant freshwater sponges colonise hard grounds like rocks, wood and bivalve shells (MANCONI & PRONZATO 2002). Therefore the fossil spicules pre- sumably originated from the shore zone and were trans- ported to deeper water.
The Permo-Carboniferous pelitic or calcareous lake deposits occur transgressively above allochthonous or parautochthonous coal. Rocky hard grounds were several kilometres away. Thus, the sponges probably grew on submerged or drifting plant parts at the lake shore or river delta respectively.
Ecostratigraphy In the context of the ecostratigraphic division of the Eu- ropean Permo-Carboniferous (BOY & SCHINDLER 2000) freshwater siliceous sponges appear in the upper Stephanian predominantly fluviatile phase (Gard, Lem- berg) and persist throughout the Stefano-Autunian (Friedelhausen) and the lower Autunian predominantly lacustrine phases (Donnersberg). In sediments of the up- per Autunian and Saxonian they are missing. The hitherto oldest records (Stefanian B) occur in the Gard Basin in the west. The record from Lemberg (Saar-Nahe Basin) is slightly younger (Stefanian C), but belongs to the same phase. In the Saar-Nahe Basin, however, freshwater sili- ceous sponges persist over several floral ('Stefanian' to 'Autunian' association) and faunal changes (Breitenbach phase to Quirnbach phase; BoY & SCHINDLER 2000: fig. 7). During this time span of approximately three mil-
lion years (calculated after BOY & SCHINDLER 2000: 129) their ecological requirements (occurrences in shallow coal beating lakes) remained the same.
Palaeobiogeography The French and German localities are some hundred kilo- metres apart (Fig. 1). The Gard Basin most likely drained southwards to the Palaeotethys (BOY & SCHINDLER 2000) (distance approx. 600 km; LORENZ & N1CHOLLS 1976; ZONENSHAYN et al. 1988; ZIEGLER et al. 1997). The Saar- Nahe Basin drained via the Hessian Depression, the North German Basin and the area of today's North Sea into the Panthalassa Ocean beyond Norway (BoY & SCHINDLER 2000: 121) (distance approx. 2400 km; ZONENSHAYN et al. 1988; ZIEGLER et al. 1997). Hydraulic connections via the Franconian basins or the Elbe Zone to the Paiaeo- tethys can be ruled out, as these basins also drained north- wards (e.g. HELMKAMPF et al. 1982). Only the Podolian Dobrudgea Basin (SCHNEIDER 1994) located further east and possibly the Boskovice Basin and the Zrbing Basin (ScHINDLER & HAMPE 1996) drained to the Palaeotethys. The drainage directions mentioned are contrary to SCHULTZE & SOLER-GIJON (2004) who, due to suppo}ed identical' marine' immigrants to Europe and North Artier- ica, claimed a connection to the Palaeotethys for all Euro- pean basins.
Marine organisms could only adapt to freshwater environments via an intermittent brackish stage ('estu- ary effect'; PARK & GIERLOWSK1-KORDESCH 2007). The almost simultaneous initial immigration into widely separated limnic basins (Gard, Saar-Nahe Basin) might have taken place from marine shelf areas via rivers. Subsequent distribution can be explained by spreading (e.g. aquatic tetrapods) or, if gemmules were formed, by wind dissemination. The distances to the seas are calcu- lated for the uppermost Stefanian and Autunian. Assum- ing a long-lasting migration period the sponges could have already immigrated to the rivers in the Westfalian or lower Stefanian and persisted within the limnic ba- sins. In the lower Stefanian at least the Saar-Nahe Basin was nearer to marine environments (distance to the ma- rine North German Depression approx. 700 km; LORENZ & NICHOLLS 1976). The migration route of the sponges has never been proven by 'intermediate stops' in time and space although appropriate deposits exist. This, however, could be due to a lack of targeted searching. The method of preparing ichthyolites, which led to the discoveries from the Saar-Nahe Basin, has also been carried out in the Central German Saale Basin (SCHNEI- DER et al. 1984; GEBHARDT 1988) and in the Lower Austrian ZObing Basin (SCHINDLER & HAMPE 1996). There, sponges are absent.
A lithologically striking silicified spiculite has been documented so far only in the French localities. This might be another reason why no other Palaeozoic fresh- water sponges are known.

Oldest record of freshwater sponges (Spongillina) - spiculite in the Permo-Carboniferous of Europe 383
C o n c l u s i o n s
The Permo-Carboniferous immigration of sponges into the limnic environment was most likely a continent- wide event, as indicated by their nearly contemporane- ous occurrence in French and German basins.
The Variscan Orogeny that finally resulted in the formation of the super-continent Pangaea probably lead to isolation of many marine environments that subse- quently became fresh water. In the case of the Saar- Nahe Basin this happened at the Lower/Upper Carbon- iferous boundary (FALKE & KNEUPER 1972). Conse- quently, the fauna, possibly comprising the putative an- cestors of freshwater sponges, either had to emigrate, adapt or become extinct. Another possibility is that after the Variscan Orogeny euryhaline sponges of the order Haplosclerida conquered the newly developed freshwa- ter biotopes via river systems.
Although not without problems, we prefer this lat- ter scenario. Presumably already in the Palaeozoic the gemmule-forming Haplosclerida were typical inhabit- ants of shallow-marine and intertidal habitats (nowa- days e.g. also mangroves), that were exposed to chang- ing salinity. Thus, euryhaline representatives of the Hap- losclerida may have adapted to a freshwater environ- ment. At the end of the Palaeozoic these sponges again disappeared. It is unclear whether this reflects the cli- matic changes (increasing aridization and vanishing la- custrine biotopes) in central Pangaea in the latest Early Permian, or whether the sponges succumbed to other factors that also led to the greatest of all mass extinc- tions at the Permo-Triassic boundary. According to the known fossil record it was only in the Jurassic that sponges once again invaded freshwater. However, only further sampling of older Mesozoic sites will show whether this gap in the record is an artefact or if the co- lonization of the limnic realm has really taken place more than once.
A c k n o w l e d g e m e n t s
We are t hank fu l for m a n y f o l d he lp by A. Brt ickner (Frankfur t a. M.), C. Ecker t (Mainz) , K. G r i m m (Mainz) , Dorte J a n u s s e n (Frankfur t a. M.) , K a u f m a n n (Mainz) , M. M a u s (Mainz) , W. E.G. Mti l ler (Mainz) , D. S chmi d t (B ingen) and I. Wes t (Sou thampton) . For technical suppor t we w o u l d like to thank the Inst i tut ftir S te in fo r schung (Mainz) . W e thank J. Rei tner (G6t t ingen) for crit ical rev is ion o f an ear ly vers ion o f the pa- per and for va luab le hints . H. Kerp (Mfinster) gene rous ly d e -
t e r m i n e d the L e m b e r g flora. P. S labon ( L a n d e s a m t fiir Geolo- gie und Be rgbau , Mainz) a l lowed acces s to an old gal lery at Lemberg . D .E .G. Br iggs (New Haven ) k ind ly i mproved the Eng l i sh and g a v e va luable c o m m e n t s . T h e initial f inds were real ized du r ing the G e r m a n R esea r ch F o u n d a t i o n projects Bo 553/6 and Bo 553/8, for wh i ch we w o u l d like to thank J,A. Boy (Mainz) and the G e r m a n R esea r ch Founda t ion . We thank R. Pronza to (Genoa) and an a n o n y m o u s r ev iewer for their cri- tical r ev iew o f the manusc r ip t and va luab le c o m m e n t s .
R e f e r e n c e s
ADDIS, J.S. & PETERSON, K.J. 2005. Phylogenetic relationships of freshwater sponges (Porifera, Spongillina) inferred from ana- lyses of 18S rDNA, CO1 mtDNA, and ITS2 rDNA sequences. Zoologica Scripta 34 (6): 549-557.
ANNANDALE, N. 1918. Zoological results of a tour in the Far East. - Memoires of the Asiatic Society of Bengal 6:199-216.
ATZBACH, O. 1984. Geologische Karte von Rheinland-Pfalz 1:25.000. - Blatt 6212 Meisenheim, Mainz (Geologisches Landesamt Rheinland-Pfalz).
BECQ-GIRAUDON, J.-F.: MONTENAT, C. & VAN DEN DRIESSCHE, J. 1996. Hercynian high-altitude phenomena in the French Massif Central: tectonic implications. - Palaeogeography, Palaeoclima- tology, Palaeoecology 122: 227-241.
BoY, J.A. 1989. Zur Lithostratigraphie des tiefsten Rotliegend (?Ober- Karbon - ?Unter-Perm) im Saar-Nahe-Becken (SW-Deutsch- land). - Mainzer Geowissenschaftliche Mitteilungen 18: 9 4 2 .
BOY, J.A. & SCHINDLER, W. 2000. (3kostratigraphische Bioevents im Grenzbereich Stefanium/Autunium (h6chstes Karbon) des Saar- Nahe-Beckens (SW-Deutschland) und benachbarter Gebiete. - Neues Jahrbuch flit Geologie und Palaontologie, Abhandlungen 216: 89-152.
BRETT, C.E. 8z SPEYER, S.E. 1990. Taphofacies. - In: BRIGGS, D.E.G. & CROWTHER, P.R,, eds., Palaeobiology II: 258-263, Oxford (Blackwell).
CARTER, H.J. 1849. A descriptive Account of the Freshwater Sponges (genus Spongilla) in the Island of Bombay, with Observations on their Structure and Development. - Annals and Magazine of Na- tural History (2) 4 (20): 81-100.
CAYEUX, L. 1929. Les roches srdimentaires de France: Roches siliceu- ses. - Mrmoires de la Carte Grologique de France [1929]: 1-774~
CLAUS1NG, A. 1990. Mikrofazies lakustriuer Karbonathorizonte d e s Saar-Nahe-Beckens (Unterperm, Rotliegend, SW-Deutschland). - Facies 23: 121-140.
CLAUSING, m. 1992. Fluoreszenzmikroskopisehe Untersuchungen an Seesedimenten des Saar-Nahe-Beckens (Ober-Karbon - Unter- Penn; SW-Deutschland). - Leica. Mitteilungen fiir Wissenschaft und Technik 10 (3): 72-79.
CLEMENTS, R.G. 1967. Some notes on the Purbeck Beds. - Proceedings of the Dorset Natural History and Archaeological Society 88: 43- 44.
DE LAUBENFELS, M.W. 1955. Porifera. - In: MOORE, R.C., ed., Treatise on Invertebrate Paleontology, Part E, Archaeocyatha and Pori- fera: E21-EI12, Lawrence, Kans. (University of Kansas) & Boulder, Colo. (The Geological Society of America).
DUNAGAN, S.P. 1999. A North American freshwater sponge (Eospon- gilla morrisonensis new genus and species) from the Morrison Formation (Upper Jurassic), Colorado. -Journal of Paleontology 73: 389-393.
FALKE, H. 8z KNEUPER, G. 1972. Das Karbon in limnischer Entwick- lung. - Compte Rendu Septi~me Congr~s International de Strati- graphie et de GEologie du Carbonifrre 1:49 67.
GEBHARDT, U. 1988. Mikrofazies und Paltiontologie biogener Karbo- hate der Unteren Mansfelder Schichten (Oberkarbon, Stefan). Hallesches Jahrbuch fiir Geowissenschaften 13:5-21.
GEYER, O.F. 1962. Uber Schwammgesteine (Spongiolith, Tuberolith, Spiculit und Gaizit). - In: Hermann-Aldinger-Festschrift: 51-59, Stuttgart (Schweizerbart).
GRAND'EURY, M.EC. 1877. Flore carbonifrre du department de la Loire. - Mrmoires de l 'Academie des Sciences de l'Institut Na- tional de France 24 (1): 1-624.
GRAND'EURY, M.EC. 1890. Grologie et palrontologie du bassin houiller du Gard. - 354 p., St. l~tienne (Throlier et C'e).
GRANT, R.E. 1836. Animal Kingdom. - In: TODD, R.B., ed., The Cyc- lopaedia of Anatomy and Physiology 1:107-118, London (Shcr- wood, Gilbert & Piper).
GREENSMITH. J.T. 1978. Textbook of petrology. 2: Petrology of the se- dimentary rocks. 6 ~h ed. -241 p., London (Allen & Unwin).

384 THOMAS SCHINDLER et al.
GRUBER, G. 1994. Kieselschwamm-Nadeln aus dem Mitteleoz~in des Eckfelder Maares, Eifel (Porifera: Demospongiae: Spongillidae). - Mainzer Naturwissenschaftliches Archiv 32: 1-6.
HANEKE, J. & LORENZ, V. 2000. Geologische Karte yon Rheinland- Pfalz 1:25.000. - Blatt 6313 Dannenfels, Mainz (Geologisches Landesamt Rheinland-Pfalz).
HANEKE, J.; LORENZ, V. & STAPF, K.R.G. 1983. Geologie und Grund- wasser des Landschaftsschutzgebiets Donnersberg. - In: STAPF, K.R.G., ed., Das Landschaftsschutzgebiet Donnersberg in der Nordpfalz. - Pollichia-Buch 11: 41-66.
HARTMAN, W.D. 1958. Natural history of the marine sponges of southern New England. - Bulletin of the Peabody Museum of Natural History 12:1-155.
HELMKAMPF, K.E; KUHLMANN, J. & KAISER, D. 1982. Das Rotlie- gende im Randbereich der Weidener Bucht. - Geologica Bava- rica 83: 167-186.
HINDE, G.J. 1883. Catalogue of the fossil sponges in the Geological De- partment of the British Museum (Natural History). - 248 p., Lon- don (Taylor & Francis).
JETTON, T.L.; FELL, P.E. & HARRISSON, KW. 2005. Cytological and cy- tochemical investigations of development from dormant gem- mules of the marine sponge, Haliclona loosanoffi. - Journal of Morphology 1: 99-116.
JOSTEN, K.-H. 1956. Die Kohlen im Pfiilzer Bergland. - Notizblatt des Hessischen Landesamtes ftir Bodenforschung zu Wiesbaden 84: 300-327.
LORENZ, W. & NICHOLLS, I.A. 1976. The Permocarboniferous Basin and Range Province of Europe. An application of plate tectonics. - In: FALKE, H., ed., The Continental Permian in Central, West, and South Europe: 313-342, Dordrecht (Reidel).
MANCONI, R. & PRONZATO, R. 2002. Suborder Spongillina subord. nov.: Freshwater sponges. - In: HOOPER, J.N.A. & VAN SOEST, R.W.M., eds., Systema Porifera: A guide to the classification of sponges: 921-1019, New York (Kluwer).
MfJLLER, W.E.G.; ZAHN, R.K. & MAIDHOF, A. 1982. Spongilla guten- bergiana n. sp. ein St~Bwasserschwamm ans dem Mittel-Eoz~in von Messel. - Senckenbergiana 63: 465-472.
OTr, E. & VOLKHEIMER, W. 1972. Paleospongilla chubutensis n.g. et n.sp. - ein StiBwasserschwamm aus der Kreide Patagoniens. - Neues Jahrbuch ftir Geologie und Palaontologie, Abhandlungen 140: 49-63.
PARK, L.E. & GIERLOWSKI-KORDESCH, E.H. 2007. Paleozoic lake faunas: Establishing aquatic life on land. -Palaeogeography, Pa- laeoclimatology, Palaeoecology 248 (1/2): 160-179.
RACEK, A.A. & HARRISON, F.W. 1974. The systematic and phylogene- tic position of Palaeospongilla chubutensis (Porifera: Spongilli- dae). - Proceedings of the Linnean Society of New South Wales 99:157-165.
REIS, O.M. 1921. Erl~iuterungen zu dem Blatte Donnersberg (21) der Geognostischen Karte von Bayern (1:100.000). - 320 p., Miin- chen (Piloty & Loehle).
REITNER, J. & WORHEIDE, G. 2002. Non-lithistid fossil Demospongiae - Origins of their palaeobiodiversity and highlights in history of preservation. - In: HOOPER, J.N.A. & VAN SOEST, R.W.M., eds., Systema Porifera: A guide to the classification of sponges: 52- 68, New York (Kluwer).
RICHTER, G. & BASZIO, S. 2000. Zur Taphonomie yon Spongilliden (Porifera) - I. Korrosionserseheinungen an fossilen Skleren der Grube Messel. - S enckenbergiana lethaea 80:445-461.
RICHTER, G. & WUTrKE, M. 1999. Lutetiospongilla heili n. gen. n. sp. und die Spongillidenfauna yon Messel. - Courier Forschungs-In- stitut Senckenberg 216:183-195.
SCH~,FER, A. & STAMM, R. 1989. Lakustrine Sedimente im Permokar- bon des Saar-Nahe-Beckens. - Zeitschrift der Deutschen Geolo- gischen Gesellschaft 140: 259-276.
SCH.g, FER, m.; RAST, U. & STAMM, R. 1990. Lacustrine paper shales in the Permocarboniferous Saar-Nahe Basin (West Germany) - de-
positional enviromnent and chemical characterization. - In: HE- LING, D.; ROTHE, P.; FORSTNER, U. & STOFFERS, P., eds., Sedi- ments and environmental geochemistry: 220-238, Berlin (Sprin- ger).
SCHINDLER, T. & HAMPE, O. 1996. Eine erste Fischfauna (Chondrich- thyes, Acanthodii, Osteichthyes) aus dem Permokarbon Nieder- 6sterreichs (Z6bing, NE Krems) mit pal~io6kologischen und bio- stratigraphischen Anmerkungen. - Beitr/ige zur Pal/iontologie 21: 93-103.
SCHINDLER, T. & POSCHMANN, M. 2001. Das Profil einer Pipeline- Trasse bei Odenbach und seine lithostratigraphischen Leithori- zonte (Lauterecken-Formation, Unterperm; Saar-Nahe-Gebiet, SW-Deutschland). - Mainzer Geowissenschaftliche Mitteilun- gen 30: 91-104.
SCHNEIDER, J. 1994. Environment, biotas and taphonomy of the Lower Permian lacustrine Niederhaslich limestone, D6hlen basin, Ger- many. - Transactions of the Royal Society of Edinburgh: Earth Sciences 84: 453-464.
SCHNEIDER, J.; SIEGESMUND, S. & GEBHARDT, U. 1984. Pal~iontologie und Genese limnischer Schill- and Algenkarbonate in der Rand- fazies der kohlefiihrenden Wettiner Schichten (Oberkarbon, Ste- fan C) des NE-Saaletroges. - Hallesches Jahrbuch (fir Geowis- senschaften 9:35-51.
SCHRODER, H.C.; EFREMOVA, S.M.; ITSKOVICH, V.B.; BELIKOV, S.; MASUDA, Y.; KRASKO, A.; MOLLER, I.M. & MOLLER, W.E.G. 2003. Molecular phylogeny of the freshwater sponges in Lake Baikal. - Journal of Zoological Systematic and Evolution Re- search 41: 80-86.
SCHULTZE, H.-E & SOLER-GIJON, R, 2004. A xenacanth clasper fcom the ?uppermost Carboniferous - Lower Permian of Buxlere-les- Mines (Massif Central, France) and the palaeoecology of the Eu- ropean Permo-Carboniferous basins. - Neues Jahrbuch far Geo- logie und Pal/iontologie, Abhandlungen 232: 325-363.
SOLLAS, W.J. 1885. A classification of the sponges. - Annals and Ma- gazine of Natural History (5) 16 (95): 395.
TOPSENT, E. 1928. Spongiaires de l'Atlantique et de la M6diterran6e provenant des croisi~res du Prince Albert Ier de Monaco. - R~- sultats des campagnes scientifiques accomplies par le Prince Al- bert I. Monaco 74: 1-376.
VOLKMER-RIBEIRO, C. & REITNER, J. 1991. Renewed study of the type material of Palaeospongilla chubutensis Ott and Vokheimer (1972). - In: REITNER, J. & KEUPP, H., eds., Fossil and Recent sponges: 121-133, Berlin (Springer).
WALDSCHMIDT, W.A. & LEROY, LW. 1944. Reconsideration of the Morrison Formation in the type area, Jefferson County, Colo- rado. - Geological Society of America Bulletin 55:1097-1114.
WEBB, J.A. 1979. A reappraisal of the palaeoecology of conchostracans (Crustacea: Branchiopoda). - Neues Jahrbuch fiir Geologie und Palaontologie, Abhandlungen 158: 259-275.
YOUNG, J.T. 1878. On the occurence of a freshwater sponge in the Pur- beck Limestone. - Geological Magazine 5:220-221.
ZIEGLER, A.M.; HULVER, M.L. & ROWLEY, D.B. 1997. Permian world topography and climate. - In: MARTINI, I.R, ed., Late glacial and postglacial environmental changes - Quaternary, Carboniferous- Permian and Proterozoic: 111-146, New York (Oxford Univer- sity Press).
ZIMMERLE, W. 1991. Stratigraphic distribution, lithological paragene- sis, depositional environment and diagenesis of fossil siliceous sponges in Europe. - In: REITNER, J. & KEUPP, H., eds., Fossil and Recent sponges: 554-577, Berlin (Springer).
ZONENSHAYN, L.R; KUZ'MIN, M.I. & KONONOV, M.V. 1988. Absolute reconstructions of the position of the continents during Paleozoic and Early Mesozoic time. - Geotectonics 21: 199-212.
Manuskripteingang / manuscript received 1.2. 2008: Manuskriptannahme / manuscript accepted 20. 8. 2008.





























