Embed Size (px)
Citation preview

Occurrence and phylogenetic significance of monoplastidic meiosis in liverworts
KAREN SUE RENZAGLIA~ Department of Biological Sciences, Box 70703, East Tennessee State University, Johnson City, TN 37601-0703, U.S.A.
ROY C. BROWN AND BETTY E. LEMMON Department of Biology, University of Southwestern Louisiana, Lnfayette, LA 70504-2451, U.S.A.
JEFFREY G. DUCKETT School of Biological Sciences, Queen Mary and Wes@eld College, Mile End Road, University of London,
London El 4NS, United Kingdom
AND
ROBERTO LIGRONE Dipartimento di Biologia Vegetale, Universita di Napoli, Via Foria, 223, Napoli 1-80139, ltaly
Received June 2 , 1993
RENZAGLIA, K.S., BROWN, R.C., LEMMON, B.E., DUCKETT, J.G., and LIGRONE, R. 1994. Occurrence and phylogenetic significance of monoplastidic meiosis in liverworts. Can. J. Bot. 72: 65-72.
Monoplastidic meiosis is reported for the first time in three seemingly unrelated liverworts, namely Blasia pusilla (Metzgeriales), Monoclea gottschei (Monocleales), and Haplornitrium blumei (Haplomitriales). A second species of Haplornitriurn, H. hookeri, is polyplastidic as previously reported. All three taxa represent isolated relicts of ancient liverwort lineages. Monoplastidy in these hepatics is evident in archesporial tissue and is maintained through successive sporogenous cell generations. In archesporial mitosis, the single plastid divides and the two resultant plastids are precisely positioned so that one is inherited by each daughter cell. In the nascent spore mother cell, the solitary plastid undergoes two successive divisions and the resulting four plastids become positioned in a tetrahedral arrangement. Concomitantly, the sporocyte assumes a quadrilobed shape, which is less exaggerated in Monoclea, and a single large plastid is situated in each lobe. Details of plastid ultrastructure and morphology vary slightly among the three taxa. Evidence is presented that Blasia and Monoclea share a common ancestry and represent pivotal taxa in the evolution of the two main lines of liverworts. Haplomitrium is suggested to occupy a more basal position in bryophyte phylogeny. Monoplastidy in meiosis of liverworts links the charo- phytes, the three bryophyte clades, and the lycopsid pteridophytes and supports a monophyletic interpretation of land plant phylogeny.
Key words: chloroplast, liverwort, meiosis, monoplastidy, phylogeny, sporogenesis.
RENZAGLIA, K.S., BROWN, R.C., LEMMON, B.E., DUCKETT, J.G., et LIGRONE, R. 1994. Occurrence and phylogenetic significance of monoplastidic meiosis in liverworts. Can. J . Bot. 72 : 65-72.
On rapporte pour la premibre fois la mkiose monoplaste chez trois hkpatiques apparemment sans parentk, nommkment le Blasiapusilla (Metzgkriales), le Monoclea gottschei (Monoclkales) et 1'Haplomitrium blumei (Haplomitriales). Une deuxibme espbce d'Haplomitriurn, 1'H. hookeri, est polyplaste comme dkjB rapportk. Les trois taxons reprksentent des vestiges isolks d'anciennes lignkes d'hkpatique. La monoplastidie chez ces hkpatiques est evidente dans les tissus archksporaux et se main- tient B travers les genkrations de cellules sporogbnes. Dans la mitose archksporale, le plaste unique se divise et les deux plastes qui en rksultent sont positionnkes avec prkcision, de sortes que chaque cellule fille en r e ~ o i v e un. Chez la jeune cellule mere des spores, le plaste unique subit deux divisions successives et il en rksulte quatre plastes qui se distribuent selon un arrange- ment tktrabdrique. Simultankment, le sporocyte prend une forme tktralobke qui est moins prononcke chez le Monoclea, et on retrouve un gros plaste isolk dans chaque lobe. I1 y a peu de variation dans l'ultrastructure et la morphologie entre les trois taxons. ~ e s auteurs donnent des preuves B l'effet que le Blasia et le Monoclea ont un ancktre commbn et reprksentent des taxons charnikres dans l'kvolution des deux lignkes principales d'hCpatique. On suggere que 1'Haplornitrium occupe une position plus prbs de la base dans la phylogknie des bryophytes. La monoplastidie dans la mkiose des hkpatiques relie les charophytes, les trois clades de bryophytes et les ptkridophytes lycopsidkes, ce qui supporte I'interprktation monophylktique de la phylogknie des plantes terrestres.
Mots cle's : chloroplastes, hkpatique, mkiose, monoplastidie, phylogknie, sporogtnbse. [Traduit par la rkdaction]
Introduction
Monoplastidic cell division in embryophytes not only pro- vides fundamental information on plastid-based cellular mor- phogenesis but also furnishes important clues to the early diversification of terrestrial organisms (Brown and Lemmon 1990a, 1992). Monoplastidic meristematic systems are char- acterized by precisely controlled plastid migration that pre- dicts the ensuing division plane and ensures that a plastid is inherited by each daughter cell. These morphogenetic events
are associated with highly specialized microtubule assemblages that exhibit variations among the monoplastidic taxa and thus provide valuable insights into evolutionary modifications of cellular differentiation processes.
Unmistakably an ancestral condition, monoplastidy is char- acteristic of cell division throughout the life cycle, both mito- sis and meiosis, in a variety of charophycean green algae, including Coleochaete (Pickett-Heaps 1975). The majority of land plants are polyplastidic or exhibit monoplastidy only in reproductive cell lineages (Brown and Lemmon 19886, 1990a, 1990~). For example, monoplasfidy is typical of
'Author to whom all correspondence should be addressed. spermatogenesis in all archegoniates that produce biflagellated Printed in Canada 1 Imprime au Canada
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
4/13
For
pers
onal
use
onl
y.
66 CAN. J . BOT. VOL. 72. 1994
male gametes, i.e., bryophytes, Lycopodium, and Selaginella (Renzaglia and Duckett 1988; Garbary et al. 1994).
The only archegoniates that exhibit monoplastidy in mitosis as well as meiosis are the hornworts, the moss Takakia (K.S. Renzaglia, unpublished data; Crandall-Stotler 1986), and the lycopsid pteridophytes (Brown and Lemmon 1990a; Uehara and Kurita 1991). In the majority of hornwort species, the mature cells remain monoplastidic throughout their cell cycle (Vaughn et al. 1992); this condition is characteristic of many algal taxa but is unparalleled in any other group of embryo- phytes.
Aside from the enigmatic Takakia, the remaining moss taxa exhibit monoplastidic meiosis and mitosis in sporogenous cells but are polyplastidic throughout the vegetative cell cycle. This is in contrast with the diverse organisms in the liverwort clade that reportedly undergo polyplastidic meiosis. Monoplastidic meiosis has been cited as a diagnostic character that decisively differentiates mosses and hornworts from liverworts (Crandall- Stotler 1984; Schuster 19846). Therefore, the presence of monoplastidic meiosis in hepatics would have profound phylo- genetic implications.
This study represents the first report of monoplastidic meiosis in liverworts. The phenomenon was discovered in three widely disparate hepatic taxa, namely Blasia (Jungermanniidae, Metz- geriales), Monoclea (Marchantiidae, Monocleales), and Hap- loinitrium (Jungermanniidae, Haplomitriales = Calobryales). The taxonomic status of each of these genera is controversial (Schuster 1984b; Renzaglia and Duckett 1987b), but it is generally agreed that each occupies either an ancestral and (or) a pivotal position in liverwort evolution. We set forth the idea that all three of these taxa possess a variety of ancestral characteristics and that they are remnants of plant lineages that were among the first to colonize land. We further suggest that the occurrence of monoplastidic meiosis in liverworts is a fundamental link between mosses, liverworts, hornworts, and lycopsids, and this single criterion provides strong support for the premise that land plants are monophyletic.
Materials and methods
Sporophytic plants of Blasia pusilla L. were collected in September 1990 and February 1991 from Triplett, Watauga Co., North Caro- lina, and in September 1988 and March 1989 from the Breadalbane Range, Perthshire, Scotland; Monoclea gottschei Lindb. from Sierra Madre Occidental, Oaxaca, Mexico in January 1990 and 1991; Haplomitrium blumei (Nees) Schust. (=Calobryutn blumei N. ab. E.) from Genting Highlands, Peninsular Malaya in March 1990 and Haplomitrium hookeri (Smith) Nees in August 1984 from the Breadal- bane Range, Perthshire, Scotland. Taxonomy of Haplomitriwn follows Schuster (1967, 1984b) who recognizes Calobryum as a subgenus of Haplomitrium.
Living material was observed and photographed using differential interference contrast optics or epifluorescence. When illuminated with UV light, native chlorophyll fluoresces red.
Materials processed for TEM were fixed either in 4% glutaralde- hyde in 0.05 M PIPES buffer (pH 7.2) or in 1.0 M phosphate buffer
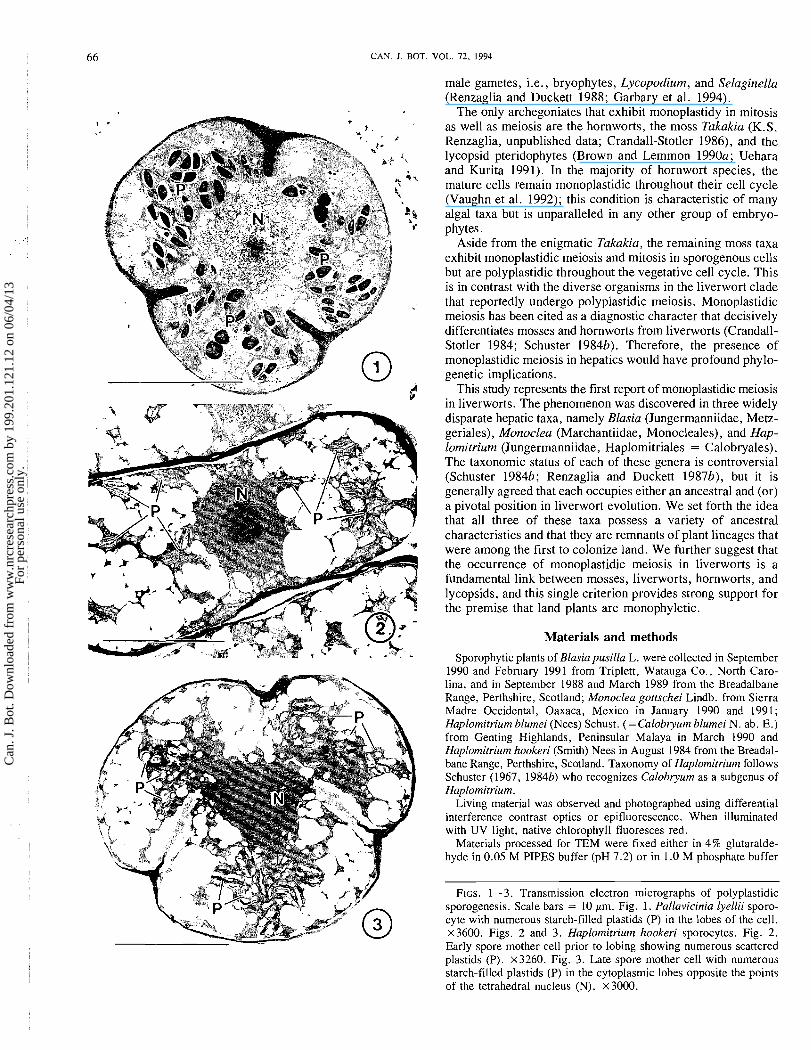
FIGS. 1-3. Transmission electron micrographs of polyplastidic sporogenesis. Scale bars = 10 pm. Fig. 1. Pallavicinia lyellii sporo- cyte with numerous starch-filled plastids (P) in the lobes of the cell. ~ 3 6 0 0 . Figs. 2 and 3. Haplomitriurn hookeri sporocytes. Fig. 2. Early spore mother cell prior to lobing showing numerous scattered plastids (P). x3260. Fig. 3. Late spore mother cell with numerous starch-filled plastids (P) in the cytoplasmic lobes opposite the points of the tetrahedral nucleus (N). ~ 3 0 0 0 .
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
4/13
For
pers
onal
use
onl
y.

RENZAGLIA ET AL. 67
(pH 6.9), washed in the appropriate buffer, and postfixed in 2% osmium tetroxide in either cacodylate or phosphate buffer. Following dehydration in a graded ethanol or acetone series, tissue was trans- ferred into propylene oxide and slowly infiltrated and embedded in Spurr's resin. Thin sections were stained with methanolic uranyl ace- tate followed by basic lead citrate and viewed with a Philips EM 201 or Hitachi 600 TEM.
In addition to the above materials, sporogenous tissues of 10 addi- tional genera of liverworts were examined for the occurrence of monoplastidy. Taxa surveyed are the following and include represen- tatives of all major hepatic groups from putatively primitive to advanced genera: Herberta, Anrhelia, Zoopsis, Diplophyllutn, Apo- trerrbia, Fossotnbronia, Pallavicinia, Petalophyllum, Mehgeria, and Conocephalum.
Observations Polyplastidic cell division in sporogenous tissue is the norm
for the majority of hepatics (Figs. 1-3). Young as well as old sporocytes contain an abundance of small plastids, each with starch grains and a well-developed membrane system. In prep- aration for meiosis, plastids aggregate in the cytoplasmic lobes, opposite the four points of the tetrahedral nucleus (Figs. 1, 3). Of the 14 hepatic species surveyed in this study, all but 3 were found to be polyplastidic in all stages of devel- opment. Sporocytes with multiple plastids are illustrated for the simple thalloid liverwort Pallavicinia lyelli (Fig. 1) and the leafy taxon Haplomitrium hookeri (Figs. 2, 3).
In contrast, sporogenous cells in Blasia pusilla, Haplo- mitrium blumei, and Monoclea gottschei are monoplastidic. Developing archesporial cells each contain a single flattened chloroplast that partially encircles the nucleus (Fig. 4). Before any signs of the onset of mitosis, the plastid divides by median constriction and the two resulting plastids migrate to the future spindle poles (Fig. 5). In Blasia and Monoclea, microbodies are frequently evident in the vicinity of the plastid cleavage furrow. Following telophase, a cell plate will isolate a single plastid and nucleus into each daughter cell.
In preparation for meiosis, the plastid anticipates the two nuclear divisions and subsequent quadripartitioning of the cytoplasm into a tetrad of spores by dividing twice in early meiotic prophase (Figs. 6-8, 11). After the first plastid division (Figs. 7, 8) the two plastids migrate to opposite hemispheres of the nucleus and develop a median constriction (Fig. 11). After the second plastid division, the four plastids migrate to tetrahedral positions around the central nucleus (Figs. 9, 10, 12- 14). Cytoplasmic lobing is more pronounced in Blasia (Figs. 12- 14) and Haplomitrium (Fig. 10) than in Monoclea (Fig. 15). Once the comprehensive polarity for meiosis has been established and prior to the initiation of the division process, the plastids of Monoclea divide (Fig. 15). In Blasia and Haplomitrium, plastid lobing and division occurs after meiosis is completed in the young spore (Figs. 16, 17).
TEM reveals that the plastids in sporocytes of all three hepatics are relatively complex with well-developed thylakoid systems that become progressively more complex during sporogenesis. It is interesting to follow changes in the plastids of Blasia that overwinters in meiotic prophase (Duckett and Renzaglia 1993). In archesporial cells of Blasia, the saucer- shaped plastid is devoid of staich grains and contains a rela- tively poorly developed membrane system of short grana and intergranal lamellae that frequently encircle the periphery of the stroma (Figs. 4, 5). By the time sporocytes are lobed, the spherical plastids are extremely large and contain an abun-
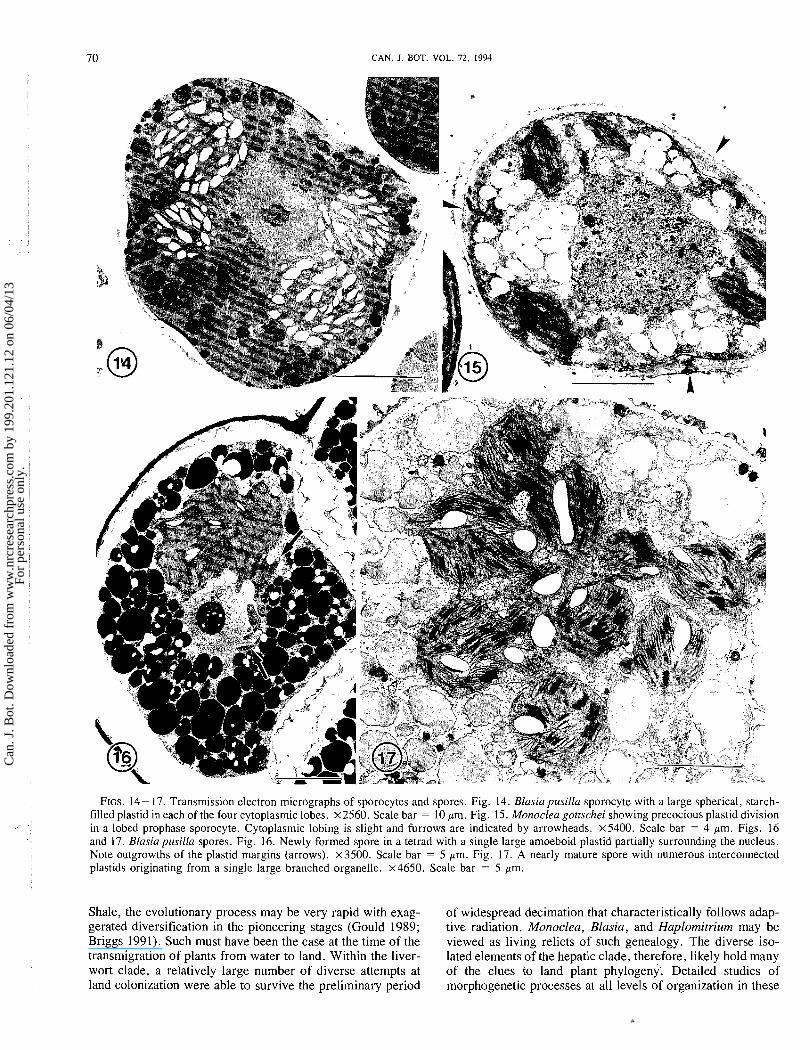
dance of starch and an elaborate membrane system of stacked thylakoids (Figs. 12- 14). The sporocyte overwinters in this stage. After meiosis is completed the following spring, young spores in tetrads (Fig. 16) contain a large pleomorphic plastid with reduced starch and abundant grana. The margins of these plastids and those of Monoclea are characterized by irregular outgrowths or lobes that lack grana and contain vesicular membranes and aggregates of tubules. Plastid proliferation in spores occurs rapidly as the chloroplast lobes and divides to produce numerous elliptical chloroplasts (Fig. 17).
Discussion The existence of monoplastidic meiosis in liverworts has
far-reaching phylogenetic implications. First, it represents a critical symplesiomorphy between all bryophyte clades, lycop- sid pteridophytes, and the coleochaetalean green algae, the modern lineage that likely contained the progenitor of land plants (Graham et al. 1991). The fundamental similarity of morphogenetic plastid migration in these major taxa provides strong support for a monophyletic interpretation of archegoni- ate evolution (Brown and Lemmon 1 9 9 0 ~ ) .
Second, the occurrence of both monoplastidic meiosis and polyplastidic meiosis within the liverwort assemblage as well as within a single genus Haplomitrium opens the way to a detailed scrutiny of evolutionary changes in sporogenesis. It is now possible to investigate changes in cellular organization and morphogenesis associated with the evolution of seemingly apolar polyplastidic sporocytes characteristic of Marchantii- dae (e.g., Conocephalum; Brown and Lemmon 1988a) and seed plants (Brown and Lemmon 1991) from polarized quadri- plastidic sporocytes of simple plants (for review, see Brown and Lemmon 1991). Of particular interest will be characteri- zation of the cytoskeletal system associated with plastid polar- ity in hepatics. In other embryophytes with monoplastidic meiosis, a plastid-based quadripolar microtubule system (QMS) is precursor to the meiotic spindle (Brown and Lemmon 1 9 9 0 ~ ) . Is a QMS produced in hepatics? It is pertinent to examine additional species of Haplomitrium to identify pos- sible intermediate conditions that will shed light on the trans- formation from monoplastidy to polyplastidy.
Within the liverwort clade, there exists an immense range of morphological, anatomical, ultrastructural, and molecular fea- tures (Crandall-Stotler 198 1 ; Schuster 1984a; Renzaglia 1982; Pike et al. 1992; Waters et al. 1990). Although no one has sug- gested that the hepatics are polyphyletic, it has proven difficult to organize the extant taxa into natural groupings (e.g., Schuster 1979, 19846; Waters et al. 1990; Mishler et al. 1992). There appear to be several small, isolated units whose affinities with other living genera are problematic. Monoclea, Blasia (including its sister genus Cavicularia), and Haplo- mitrium all fit into this category.
Among the hepatic genera, Monoclea and Haplomitrium are considered to be isolated relict taxa that occupy pivotal posi- tions in liverwort diversification and currently are held in monotypic orders. Because of its leafy, erect, radially sym- metrical habit, massive archegonia, and malleability in organog- raphy, Haplomitrium is viewed as a primitive hepatic (Schuster 1984a, 19846; Bartholomew-Began 1990, 1991). It shares the greatest number of attributes with certain metzgerialean taxa (Bartholomew-Began 1990, 1991) but is similar to members of the Jungermanniales in the possession of ti tetrahedral apical cell and three-ranked leaves. Like Monoclea and other Mar-
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
4/13
For
pers
onal
use
onl
y.
68 CAN. I. BOT. VOL. 72, 1994
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
4/13
For
pers
onal
use
onl
y.

RENZAGLlA ET AL. 69
chantiidae, Haplomitrium has a monostromatic sporophyte Thus, it appears that Monoclea occupies a pivotal position capsule wall that in at least one species (H. blumei; Campbell between the simple and complex thalloid liverworts. 1920) dehisces by a single longitudinal slit. The placental Blasia and its sister genus Cavicularia are extraordinary organization of Haplomitrium, characterized by the develop- metzgerialean liverworts that possess complicated ventral ment of transfer cells on both the sporophytic and gameto- auricles that are apically derived and house a Nostoc endo- phytic sides, is similar to that found in the Marchantiales and symbiont. Affinities between Blasia and the complex thalloid in putatively primitive members of the Metzgeriales but very taxa have been recognized only recently. Most notable are the different from that typical of the Jungermmaniales (Ligrone marchantialean-like spermatozoids (Renzaglia and Duckett et al. 1993). 1987a, 1987b) and the two rows of ventral scales (Renzaglia
It is interesting to note that certain features of Haplomitrium 1982). Blasia resembles Monoclea in the location of the such as leaf insertion and branching patterns have been described as mosslike. The commonalities shared by Haplo- mitrium and various bryophyte lineages coupled with the exis- tence of monoplastidy in meiosis are indicative of a basal position of Haplomitrium within the Hepaticae and suggest a more recent common ancestry between the liverwort and moss clades than has been postulated (Crandall-Stotler 1984, 1986; Mishler and Churchill 1985; Bremer et al. 1987). The fact that Takakia, a genus whose identity has recently been confirmed as a moss (Smith and Davison 1993), was traditionally placed in the Haplomitriales further provides skepticism as to the absolute separation of moss and hepatic lineages. Most likely, the simple mechanism of spore release in Takakia, Haplo- mitrium blumei, and Monoclea, through a solitary elongated suture, is plesiomorphic. This feature is also shared with primitive pteridophytes, including the lycopsids and rhynio- phytes (Bold et al. 1987) and represents an additional charac- ier link between these ancient lineages of archegoniates. Cladistic analyses based on spermatogenesis corroborate these evolutionary affinities with the depiction of mosses and liver- worts as monophyletic and the association of Selaginella, a lycophyte, with the bryophyte clade (Garbary et al. 1994). An unequivocal molecular link between the mosses, liverworts, and lycophytes is the common occurrence of a unique inver- sion in the chloroplast genome that is lacking in other pterido- phytes and seed plants (Raubeson and Jansen 1992).
Like Haplomitrium, Monoclea is an enigmatic taxon that shows affinities with more than one hepatic group and there- fore is not readily placed in any existing taxonomic unit. Monoclea shares key features with the complex thalloid line but possesses a well-developed sporophyte with an elongated seta that originates directly from a vegetative thallus devoid of air chambers; these features are shared with metzgerialean liverworts. Monoclea also possesses a number of peculiar characteristics, including the unique development of sporo- genous cells and a free-nuclear embryo (Campbell 1954; Johnson 1904; Cavers 1904; Brown and Lemmon 1992).
female sex organs, morphology of sporophyte and protective structures, and apical organization. Monoplastidy is an addi- tional feature uniting these two genera, both of which show affinities with the Marchantiales. Moreover, in both taxa the dividing plastids in sporogones are associated with micro- bodies and the sporocyte plastids exhibit similar unique marginal projections. Plastids associated with microbodies also occur in the hornworts and charophytes (Duckett and Renzaglia 1988; Graham and Kaneko 1991) and provide further evidence for a phylogenetic connection among these groups. These findings suggest that Blasia and Monoclea had a common origin from a proto-liverwort and raise questions as to the individuality of the two currently recognized liverwort lineages, the Marchan- tiidae or complex thalloids (including Monocleales, Sphaero- carpales, and Marchantiales) and the Jungermanniidae or simple thalloid and leafy liverworts (including Haplomitriales, Treubiales, Metzgeriales, and Jungermanniales). Certainly, the existence of the Carboniferous fossils Blasiites and ~reub i - ites attest to the antiquity of the unique Blasia habit (Schuster 1984b).
The plastids of the sporogenous tissues of Haplomitrium, Monoclea, and Blasia exhibit a more elaborate fine structure than those of bryopsid mosses (Brown and Lemmon 1988b; Gambardella and Alfano 1990), Lycopodium (Uehara and Kurita 1991), Selaginella (Brown and Lemmon 1985), or Isoetes (Brown and Lemmon 1989). Sporogones of putatively primitive mosses such as Andreaeales and Sphagnales (Brown and Lemmon 1990c), hornworts (Vaughn et al. 1992; Brown and Lemmon 1990b), and charophycean algae (Graham 1990) produce plastids that are comparable in complexity to those of monoplastidic liverworts. Plastid reduction in this case is most realistically viewed as a derived condition specific to certain lines of evolution within the archegoniates.
It is not surprising that this most informative condition of monoplastidy in sporogenesis was discovered in rather dispar- ate liverwort genera. As learned from the wide array of invertebrate fossils of problematic affinities in the Burgess
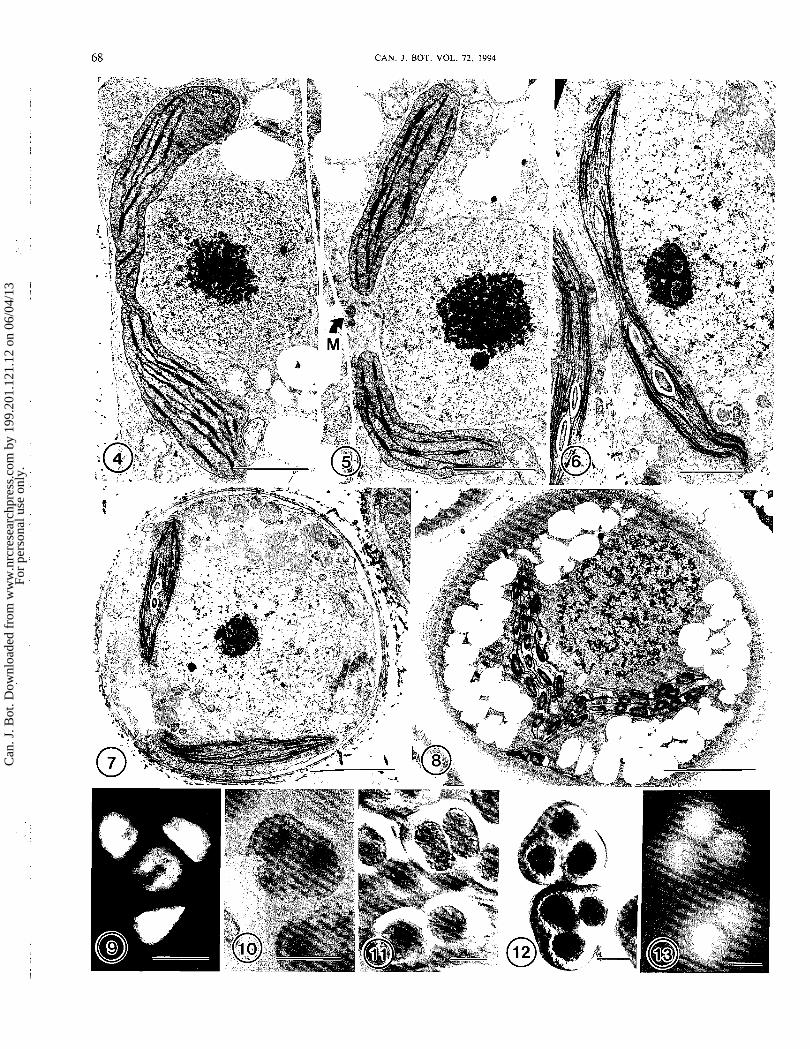
FIGS. 4-8. Transmission electron micrographs of plastids in sporogones. Figs. 4 and 5. Blasia pusilla archesporial cells. Scale bars = 2 pm. Fig. 4. A single bowl-shaped plastid partially encircles the nucleus. Note the central plastid constriction, narrow grana, and peripheral mem- branes. x9900. Fig. 5. Archesporial cell with recently divided plastids. A microbody (M) is visible between the plastids. x 10 500. Figs. 6 and 7. Young sporocytes of Monoclea gottschei. Fig. 6. Early sporocyte containing a single plastid with median constriction. X8300. Scale bar = 2 pm. Fig. 7. A slightly older sporocyte containing two newly divided plastids. x5600. Scale bar = 4 pm. Fig. 8. Young sporocyte of Haplomitrium blumei with two newly divided plastids. ~ 4 8 0 0 . Scale bar = 5 pm. FIG. 9. Epifluorescence image of Monoclea sporocyte in late meiotic prophase with four plastids in a tetrahedral position. x 1450. Scale bar = 10 pm. FIG. 10. Differential interference contrast image of a Haplomitrium blumei sporocyte showing three of the four plastids in the cytoplasmic lobes. x 850. Scale bar = 20 pm. FIGS. 11 - 13. Blasia pusilla sporocytes. x425. Scale bars = 20 pm. Fig. 11. A pair of sporocytes each with a single large plastid is shown in the lower portion of the micrograph. An older sporocyte in the upper portion of the micrograph has begun to lobe and contains two constricted plastids.
- - -
Differential interference contrast image. Fig. 12. Two sporocytes each containing four plastids (three are visible) in the cytoplasmic lobes. Differential interference contrast image. Fig. 13. Chlorophyll autofluorescence in sporocytes of Blasia pusilla at the same stage of meiotic prophase as those in Fig. 12.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
4/13
For
pers
onal
use
onl
y.
CAN. J. BOT. VOL. 72, 1994
FIGS. 14- 17. Transmission electron micrographs of sporocytes and spores. Fig. 14. Blasia p~tsilla sporocyte with a large spherical, starch- filled plastid in each of the four cytoplasmic lobes. ~ 2 5 6 0 . Scale bar = 10 pm. Fig. 15. Monoclea gottschei showing precocious plastid division in a lobed prophase sporocyte. Cytoplasmic lobing is slight and furrows are indicated by arrowheads. ~ 5 4 0 0 . Scale bar = 4 pm. Figs. 16 and 17. Blasiapusilla spores. Fig. 16. Newly formed spore in a tetrad with a single large amoeboid plastid partially surrounding the nucleus. Note outgrowths of the plastid margins (arrows). x3500. Scale bar = 5 pm. Fig. 17. A nearly mature spore with numerous interconnected plastids originating from a single large branched organelle. x4650. Scale bar = 5 pm.
Shale, the evolutionary process may be very rapid with exag- of widespread decimation that characteristically follows adap- gerated diversification in the pioneering stages (Gould 1989; tive radiation. Monoclea, Blasia, and Haplomitrium may be Briggs 1991). Such must have been the case at the time of the viewed as living relicts of such genealogy. The diverse iso- transmigration of plants from water to land. Within the liver- lated elements of the hepatic clade, therefore, likely hold many wort clade, a relatively large number of diverse attempts at of the clues to land plant phylogeny. Detailed studies of land colonization were able to survive the preliminary period morphogenetic processes at all levels of organization in these
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
4/13
For
pers
onal
use
onl
y.

RENZAGLIA ET AL. 71
plants will undoubtedly contribute significant pieces to the puzzle of the phyletic history of embryophytes.
Acknowledgements
This study was supported in part by the National Science Foundation grant No. DEB-9207646 to K.S.R. and J.G.D., and grants Nos. BSR-8610594 and RII-880219 to R.C.B. and B.E.L. R.L. gratefully acknowledges support from the Italian Ministry of University and Scientific Research and Tech- nology.
Bartholomew-Began, S.E. 1990. Classification of the Haplomitriales and Metzgeriales into the subclass Metzgeriidae, Subclass Nov. (Hepatophyta, Jungermanniopsida). Phytologica, 69: 464-466.
Bartholomew-Began, S.E. 1991. A morphogenetic re-evaluation of Haplotrzitriutrz Nees, Hepatophyta. Bryophytorum Bibl. No. 41.
Bold, H.C., Alexopoulos, C.J., and Delevoryas, T. 1987. Morphol- ogy of plants and fungi. Harper & Row, New York.
Bremer, K., Humphries, C.J., Mishler, B.D., and Churchill, S.P. 1987. On cladistic relationships in green plants. Taxon, 36: 339-349.
Briggs, D.E.G. 1991. Extraordinary fossils. Am. Sci. 79: 130-141. Brown, R.C., and Lemrnon, B.E. 1985. A cytoskeletal system
predicts division plane in meiosis of Selaginella. Protoplasma, 127: 101 - 109.
Brown, R.C., and Lemmon, B.E. 1988a. Cytokinesis occurs at boundaries of domains delimited by nuclear-based microtubules in sporocytes of Conocephalum conicum (Bryophyta). Cell Motil. Cytoskeleton, 11: 139- 146.
Brown, R.C., and Lemmon, B.E. 19886. Sporogenesis in bryo- phytes. 1988. Adv. Bryol. 3: 159-223.
Brown, R.C., and Lemmon, B.E. 1989. Morphogenetic plastid migration and microtubule organization during megagametogene- sis in Isoetes. Protoplasma, 152: 136- 147.
Brown, R.C., and Lemmon, B.E. 1990a. Monoplastidic cell division in lower land plants. Am. J. Bot. 77: 559-571.
Brown, R.C., and Lemmon, B.E. 1990b. The quadripolar micro- tubule system and meiotic spindle ontogeny in hornworts (Bryophyta: Anthocerotae). Am. J. Bot. 77: 1482- 1490.
Brown, R.C., and Lemmon, B.E. 1990c. Sporogenesis in bryo- phytes. In Microspores: evolution and ontogeny. Edired by S. Blackmore and R.B. Knox. Academic Press, London. pp. 55-94.
Brown, R.C., and Lemmon, B.E. 1991. The cytokinetic apparatus in meiosis: control of division plane in the absence of a preprophase band of microtubules. In The cytoskeletal basis of plant growth and form. Edited by C.W. Lloyd. Academic Press, London. pp. 260 -273.
Brown, R.C., and Lemmon, B.E. 1992. Polar organizers in mono- plastidic mitosis of hepatics (Bryophyta). Cell Motil. Cyto- skeleton, 22: 72-77.
Campbell, D.H. 1920. Studies on some Indian Hepaticae. Calobryum blurnei N. ab. E. Ann. Bot. (London), 31: 1 - 12.
Campbell, E.O. 1954. The structure and development of Monoclea forsteri Hook. Trans. R. Soc. N.Z. 82: 237-248.
Cavers, F. 1904. On the structure and development of Monoclea Forsteri Hooker. Rev. Bryol. 31: 69-80.
Crandall-Stotler, B.J. 1981. Morphology/anatomy of hepatics and anthocerotes. Adv. Bryol. 1: 315-398.
Crandall-Stotler, B.J. 1984. Musci, hepatics and anthocerotes-an essay on analogues. In New manual of bryology. Edired by R. Schuster. Hattori Botanical Laboratory, Nichinan, Japan. pp. 1093 - 1129.
Crandall-Stotler, B.J. 1986. Morphogenesis, developmental anatomy and bryophyte phylogenesis: contraindications of monophyly. J. Bryol. 14: 1-23.
Duckett, J.G., and Renzaglia, K.S. 1988. Ultrastructure and devel- opment of plastids in the bryophytes. Adv. Bryol. 3: 33-93.
Duckett, J.G., and Renzaglia, K.S. 1993. The reproductive biology of the liverwort Blasia pusilla L. J. Bryol. 17: 541 -552.
Gambardella, R., and Alfano, F. 1990. Monoplastidic mitosis in the moss Tirnmiella barbuloides (Bryophyta). Protoplasma, 156: 29-38.
Garbary, D.J. Renzaglia, K.S., and Duckett, J.G. 1994. The phylogeny of land plants: a cladistic analysis based on spermato- genesis. Plant Syst. Evol. In press.
Gould, S.J. 1989. Wonderful life-the Burgess Shale and the nature of history. W.W. Norton & Co., New York.
Graham, L.E. 1990. Meiospore formation in charophycean algae. In Microspores: evolution and ontogeny. Edired by S. Blackmore and B.R. Knox. Academic Press, London. pp. 43 -54.
Graham, L.E., and Kaneko, Y. 1991. Subcellular structures of rele- vance to the origin of land plants (embryophytes) from green algae. Crit. Rev. Plant Sci. 10: 323-342.
Graham, L.E., Delwiche, C.F., and Mishler, B.D. 1991. Phylo- genetic connections between the "green algae" and the "bryo- phytes." Adv. Bryol. 4: 213-244.
Johnson, D.S. 1904. The development and relationship of Monoclea. Bot. Gaz. 38: 185-205.
Liarone, R., Duckett, J.G., and Renzaglia, K.S. 1993. The gametophyte-sporophyte junction in landplants. Adv. Bot. Res. 19: 231-317.
Mishler, B.D., and Churchill, S.P. 1985. Transition to a land flora: phylogenetic relationships of the green algae and bryophytes. Cladistics, 1: 305-328.
Mishler, B.D., Thrall, P.H., Hopple, J.S., Jr., De Luna, E., and Vilgalys, R. 1992. A molecular approach to the phylogeny of bryo- phytes: cladistic analysis of chloroplast-encoded 16s and 23s ribo- somal RNA genes. Bryologist, 95: 172- 189.
Pickett-Heaps, J.D. 1975. Green algae: structure, reproduction and evolution in selected genera. Sinauer Associates, Inc., Sunderland, Mass.
Pike, L.M., Hu, A., Renzaglia, K.S., and Musich, P. 1992. Liver- wort genomes display extensive structural variations. Bot. J. Linn. SOC. 109: 1 - 14.
Raubeson, L.A., and Jansen, R.K. 1992. Chloroplast DNA evidence on the ancient evolutionary split in vascular plants. Science (Washington, D.C.), 255: 1697- 1699.
Renzaglia, K.S. 1982. A comparative developmental investigation of the gametophyte generation in the Metzgeriales (Hepatophyta). Bryophytorum Bibl. No. 24.
Renzaglia, K.S., and Duckett, J.G. 1987a. Comparative ultrastruc- tural studies of spermatogenesis in the Metzgeriales (Hepato- phyta). 11. The blepharoplast of Blasia pusilla. Am. J. Bot. 74: 53 -64.
Renzaglia, K.S., and Duckett, J.G. 19876. Spermatogenesis of Blasia pusilla from antheridial initial through mature spermato- zoid. Bryologist, 90: 468-501.
Renzaglia, K.S., and Duckett, J.G. 1988. Different developmental processes underlie similar spermatozoid architecture in mosses, hepatics and hornworts. J. Hattori Bot. Lab. 64: 2 19 -236.
Schuster, R.M. 1967. Studies on Hepaticae XV: Calobryales. Nova Hedwigia, 13: 1-63.
Schuster, R.M. 1979. The phylogeny of the Hepaticae. In Bryophyte systematics. Edited by G.C.S. Clarke and J.G. Duckett. Syst. Assoc. Spec. Vol. 14: 41 -82.
Schuster, R.M. 1984a. Comparative anatomy and morphology of the Hepaticae. In New manual of bryology. Edited by R. Schuster. Hattori Botanical Laboratory, Nichinan, Japan. pp. 760- 89 1.
Schuster, R.M. 19846. Evolution, phylogeny and classification of the Hepaticae. In New manual of bryology. Edired by R. Schuster. Hattori Botanical Laboratory, Nichinan, Japan. pp. 892- 1070.
Smith, D.K., and Davison, P.G. 1993. Antheridia and sporophytes in Takakia ceratophylla (Mitt.) Grolle: evidence for reclassifica- tion among mosses. J. Hattori Bot. Lab. 73: 263-271.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
4/13
For
pers
onal
use
onl
y.

72 CAN. J . BOT. VOL. 72, 1994
Uehara, K., and Kurita, S. 1991. Ultrastructural study on spore wall Waters, D.A., Buchheim, M.A., and Chapman, R.L. 1990. Bryo- morphogenesis in Lycopodium clavatum (Lycopodiaceae). Am. J. phyte phylogeny and cytoplasmic rRNA sequence data. Am. J. Bot. 78: 24-36. Bot. 77(Suppl.): 4-5.
Vaughn, K.C., Ligrone, R., Owen, H.A., Hasegawa, J., Campbell, E.O., Renzaglia, K.S., and Monge-Najera, J. 1992. The antho- cerote chloroplast. New Phytol. 120: 169- 190.
Can
. J. B
ot. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
4/13
For
pers
onal
use
onl
y.





























