Embed Size (px)
Citation preview

Nucleolar Binding Sequences of the Ribosomal Protein S6e Family Reside inEvolutionary Highly Conserved Peptide Clusters
Swarupa Kundu-Michalik, Marc-Angelo Bisotti, Edgar Lipsius, Andreas Bauche, Antonina Kruppa,Thomas Klokow, Gertrud Kammler,1 and Joachim KruppaCenter of Experimental Medicine, Institute of Molecular Cell Biology, Hamburg University, D-20246 Hamburg, Germany
Proteomic analyses of the nucleolus have revealed almost 700 functionally diverse proteins implicated in ribosomebiogenesis, nucleolar assembly, and regulation of vital cellular processes. However, this nucleolar inventory has notunveiled a specific consensus motif necessary for nucleolar binding. The ribosomal protein family characterized by theirbasic nature should exhibit distinct binding sequences that enable interactions with the rRNA precursor moleculesfacilitating subunit assembly. We succeeded in delineating 2 minimal nucleolar binding sequences of human ribosomalprotein S6 by fusing S6 cDNA fragments to the 5# end of the LacZ gene and subsequently detecting the intracellularlocalization of the b-galactosidase fusion proteins. Nobis1 (nucleolar binding sequence 1), comprising of 4 highly conservedamino acid clusters separated by glycine or proline, functions independently of the 3 authentic nuclear localization signals(NLSs). Nobis2 consists of 2 conserved peptide clusters and requires the authentic NLS2 in its native context. Similarly, wededuced from previous publications that the single Nobis of ribosomal protein S25 is also highly conserved. The functionalprotein domain organization of the ribosomal protein S6e family consists of 3 modules: NLS, Nobis, and the C-terminalserine cluster of the phosphorylation sites. This modular structure is evolutionary conserved in vertebrates, invertebrates, andfungi. Remarkably, nucleolar binding sequences of small and large ribosomal proteins reside in peptide clusters conservedover millions of years.
Introduction
The nucleolus is a highly dynamic nuclear structure(Andersen et al. 2005) formed around tandemly repeatedgenes of rDNA coding for preribosomal RNA (Hadijolov1985). The major function of the nucleolus is the assemblyof ribosomal subunits (Raska et al. 2006). During ribosomebiogenesis, the rRNA precursor is transcribed at the borderbetween the fibrillar center and dense fibrillar component(DFC), and in the DFC (Koberna et al. 2002). RibosomalRNA processing and base modifications as well as the as-sociation of rRNA with ribosomal proteins give rise to in-termediates of preribosomal particles (Grandi et al. 2002;Tschochner and Hurt 2003). These progress vectoriallythrough functional subdomains of the DFC into the granularcomponent (GC) for further maturation and completion ofribosomal subunit assembly (Scheer and Hock 1999; Gerbiet al. 2003). A myriad of small nucleolar ribonucleoproteinparticles (RNPs) assist the processing and modification ofthe primary rRNA transcript (Fromont-Racine et al. 2003;Granneman et al. 2004) so that ultimately mature 5.8S, 18S,and 25S rRNA molecules are produced in Saccharomycescerevisiae (Venema and Tollervey 1999).
The advent of proteomic analyses of the nucleolus(Andersen et al. 2002; Scherl et al. 2002) as well as of pre-ribosomal particles (Takahashi et al. 2003) have revealedcountless trans-acting factors participating in the complexpattern of preribosome maturation (Milkereit et al. 2003),which is accompanied by a dynamic change in the compo-sition and subnuclear distribution of ribosomal synthesisfactors (Leung et al. 2003; Hinsby et al. 2006). Fluorescene
recovery after photobleaching analyses have demonstratedthat some nucleolar components involved in ribosome bio-genesis rapidly shuttle between the nucleolus and the nu-cleoplasm (Chen and Huang 2001).
Ribosomes are complexes essential for cell prolifera-tion and cell growth present in all organisms (Mayer andGrummt 2006; Bernstein et al. 2007). In bacteria, the cat-alytically active rRNA forms the framework upon which 55ribosomal proteins are assembled, thereby stabilizing the3-dimensional structure of the ribosomal subunits (Schuwirthet al. 2005). Analogous to all other nuclear proteins, eukary-otic ribosomal proteins are synthesized in the cytoplasm,subsequently imported through the nuclear pore complexinto the nucleoplasm, and ultimately accumulate in the nu-cleolus (Fromont-Racine et al. 2003). Their associationwith the newly transcribed pre-rRNA generates a 90S preri-bosomal particle that is processed in a highly complexpattern into 40S and 60S ribosomal subunits, which areexported through the nuclear pore complex into the cyto-plasm (Grandi et al. 2002; Fromont-Racine et al. 2003;Tschochner and Hurt 2003). In mammalian cells, nuclearimport of ribosomalproteins (Schmidt et al. 1995) ismediatedby classical nuclear localization signals (NLSs) of eithermonopartite or bipartite nature (Dingwall and Laskey1991; Lipsius et al. 2005) and assisted by a variety of importfactors (Jakel and Gorlich 1998).
A specific mechanism for targeting proteins into thenucleolus is not needed because the nucleolus is not sur-rounded by a lipid bilayer (Carmo-Fonseca et al. 2000).Furthermore, analysis of the nucleolar proteome has dem-onstrated that a simple universal targeting motif shared byall nucleolar proteins does not exist (Andersen et al. 2002;Scherl et al. 2002) suggesting that they are instead seques-tered and retained by binding to other nucleolar compo-nents (Carmo-Fonseca et al. 2000). Within proteins ofretroviruses (Hatanaka 1990), short nucleolar targeting sig-nals of 12–20 amino acids can target a reporter protein fromthe cytoplasm into the nucleolus.
It is assumed that the ribosome arose on a single oc-casion as a particle composed completely of RNA capable
1 Present address: Department of Neurological Surgery, Center ofClinical Neurosciences, Hamburg University, D-20246 Hamburg,Germany.
Key words: ribosomal protein, nuclear import, nucleolar bindingsequence, nuclear localization signal, ribosome biogenesis.
E-mail: [email protected].
Mol. Biol. Evol. 25(3):580–590. 2008doi:10.1093/molbev/msn002Advance Access publication January 4, 2008
� The Author 2008. Published by Oxford University Press on behalf ofthe Society for Molecular Biology and Evolution. All rights reserved.For permissions, please e-mail: [email protected]
at Medical C
ollege of Wisconsin L
ibraries-Serials on July 3, 2015http://m
be.oxfordjournals.org/D
ownloaded from

of catalyzing peptide bond formation (Noller 1991; Woolet al. 1995). Numerous established rRNA sequences of dif-ferent phyla and species support this assumption by provid-ing compelling evidence for the homology of rRNAs of allspecies, which is more apparent in the comparison of sec-ondary structure than of nucleotide sequences. In the1970#s, Wool and Stoffler (1974) were already convincedthat ribosomal protein sequences from distant species arealso evolutionary related. Nowadays, genomic sequencecomparisons have confirmed the existence of homologousproteins in ribosomes of Archaea, Bacteria, and Eukarya(Lecompte et al. 2002; Mears et al. 2002). Presumably, evo-lutionary highly conserved clusters in these ribosomal pro-tein sequences fulfill a crucial role in ribosome assemblyand function.
Research on the biogenesis of ribosomal subunits con-centrating on rRNA transcription, processing, and matura-tion has neglected the influence of ribosomal proteins in theassembly process. However, in 2005, Milkereit’s group(Ferreira-Cerca et al. 2005) demonstrated that eukaryoticribosomal proteins play vital roles in the maturation andtransport of the pre-18S rRNA. Implied by this result isthe existence of specific amino acid sequences within ribo-somal proteins, which optimally interact with their corre-sponding nucleotide sequences so that distinct pre-rRNAprocessing steps can occur (Ferreira-Cerca et al. 2005).Unfortunately, the exact sequences and structural featuresof such binding domains in eukaryotic ribosomal proteinshave not yet been systematically investigated. Therefore,we characterized the topogenic sequences of human ribo-somal protein S6 in detail by delineating the regions re-quired for nuclear targeting and nucleolar binding inorder to understand the organization and interrelationshipof these functional domains (Fukami-Kobayashi et al.
2007). To this end, we demonstrate that S6 has 2 nucleolarbinding sequences —Nobis1 (nucleolar binding sequence 1)in the center functioning independently of the 3 authenticNLSs and Nobis2 in the C-terminal region depending onthe presence of NLS2—both of which reside in regionsof highly conserved peptide clusters. Furthermore, we showthat NLS3 of human S6 is a classical bipartite import signal.
Materials and MethodsConstructs for the Delineation of Nobis1 in the CentralRegion of S6
A set of deletion mutants was constructed to identifya Nobis in the central region of S6. To this end, 2 polymer-ase chain reaction (PCR) fragments were designed using theprimers S6 (A1) and S6 (Nhe170) as well as S6 (Nhe210)and S6 (H249) (table 1). The PCR fragments were digestedwith AflII/NheI and NheI/HindIII, respectively, and clonedtogether into the pASH vector (Lipsius et al. 2005), whichhad been restricted with AflII/HindIII. Since the PCR primerS6 (Nhe170) also hybridized to the S6 DNA sequence atthe nucleotides 414–420, an additional S6 fragment corre-sponding to amino acids 1–139 was synthesized so that asecond deletion mutant S6(1–249,D140–210) was obtained.In order to narrow down Nobis1, additional constructs weredesigned beginning at amino acid residues 53, 64, 72, 75,100, and 115, respectively, by employing the indicatedprimers (table 1) and the S6(1–249,D140–210) constructas a template.
Additional constructs with consecutively shorter DNAsegments were prepared containing the exogenous SV40NLS. The DNA fragments were generated by PCR usingthe plasmid pBluescript KS(�) S6 as a template and thecorresponding primers (table 1). The PCR products wereincubated with the restriction enzymes AflII and HindIIIand ligated into the pASH-SV40 vector describedpreviously (Lipsius et al. 2005).
Site-Directed Mutagenesis of NLS3 in the FragmentS6(215–240) and in the Tetrapeptide
Arginine 231 of NLS3 was mutated into valine,glycine, and glutamic acid. Hybrids were prepared fromsynthetic oligonucleotides as summarized in table 2 to ob-tain the tetrapeptides for fusion to b-galactosidase. Thesehybrids were ligated into the vector pKHlacZ (Anniloet al. 1998), which had been restricted with NotI andHindIII. The correct constructs of the clones obtainedwere verified by sequencing. Plasmid DNA was preparedfor transfections as previously described (Schmidt et al.1995).
The fragment S6(215–240) was subcloned into thevector M13mp19 and an oligonucleotide-dependent in vitromutagenesis was carried out as described by the manufac-turer (Amersham-Buchler, Braunschweig, Germany). A setof oligonucleotides was synthesized by using a mixture ofA, C, G, and T at position N in the sequence: 5#-ATTGC-GAAGGNACGCAGACTT-3#. Competent Escherichiacoli XL1-Blue cells were transformed with the mutated
Table 1PCR Primers for Characterizing Nucleolar Binding Sequences
Primer Sequence in 5# / 3#
S6 (A1) GAAAGGATCCCCTTAAGATGAAGCTGAACATCTCS6 (A53) TTGGATCCCTTAAGAGTGGTGGGAACGACAAACAAS6 (A64) TTGGATCCCTTAAGAAGCAGGGTGTCTTGACCCATS6 (A72) TTGGATCCCTTAAGCGTGTCCGCCTGCTACTGAGTS6 (A75) TTTTTATCTTAAGCTGCTACTGAGTAAGGGGCATTCS6 (A100) ATTTTATCTTAAGTGCATTGTGGATGAAATCTGAGCS6 (A115) TTTTTATCTTAAGAAAAAAGGAGAGAAGGATAT
TCCTGGS6 (A170) TTTGGATCCCTTAAGAGGACCAAAGCACCCAAGS6 (A173) TTTTTATCTTAAGGCACCCAAGATTCAGCGTCTT
GTTAS6 (A180) TTTGGATCCCTTAAGGTTACTCCACGTGTCCTGS6 (A192) GGATCCTTTCTTAAGATTGCTCTGAAGAAGCAGCS6 (H94) GGATCCAAAGCTTCTCTTTCTTTCTCCAGTTCTS6 (H98) GGATCCAAAGCTTCGAACTGATTTTCTCTTTCTS6 (H102) GGATCCAAAGCTTACAATGCAACCACGAACTGATTS6 (H120) GGATCCAAAGCTTATCCTTCTCTCCTTTTTTTACA
ATAAS6 (H203) GGATCCAAAGCTTTTATTTTTCTTGGTACGCTGCTTS6 (H210) GGATCCAAAGCTTGCATATTCTGCAGCCTCTTS6 (H229) GGATCCAAAGCTTGCAATTTGTTCCTGGCGCS6 (H249) GGATCCAAAGCTTTTCTGACTGGATTCAGACTTS6 (Nhe170) GGATCCGCTAGCAGGTTTCTTACCTTCTTTATTS6 (Nhe210) GGATCCGCTAGCAAACTTTTGGCCAAGAGAATG
NOTE.—The orientation in which the PCR primers have been used is indicated,
and the restriction sites are underlined.
Nucleolar Accumulation of Ribosomal Protein S6 581
at Medical C
ollege of Wisconsin L
ibraries-Serials on July 3, 2015http://m
be.oxfordjournals.org/D
ownloaded from

M13mp19 RF DNA that contained mutated S6 fragments.The clones were isolated and sequenced. Only 3 of the 4theoretically possible mutations were obtained, that is, va-line, glycine, and glutamic acid but not alanine. The indi-vidual S6 cDNA fragments were isolated from thecorresponding M13 constructs, sequenced, and subclonedinto pPb�S6 as previously described (Schmidt et al.1995).
The KR deletion mutants were constructed as follows:the fragment S6(215–240) containing the valine mutationwas excised from the corresponding pPb�S6 with EcoRIand HindIII and ligated into pUC19. To construct thedeletion mutants S6(215–240,D215/216) and S6(215–240,D223/224), the primers in table 2 were used in thesite-directed mutagenesis. The mutants were selected aftera ScaI restriction in the E. coli strain BMH71-18 mutS.After a second ScaI restriction, the isolated plasmids weretransformed into E. coli JM105 cells, and their correctsequence was verified.
A NotI linker had to be added to the excised S6fragment after filling up the cohesive ends with theKlenow fragment of DNA polymerase I. The modifiedconstructs were subcloned into the NotI/HindIII sites ofthe vector pKHlacZ where they were in frame with b-galactosidase. After ligation, the correct frame was verifiedby sequencing.
Construction of S6 Fragments for the Nucleolar BindingDomain (Nobis2) Encompassing NLS2
Starting from the S6 cDNA, several constructscontaining a single NLS, either NLS2 or NLS3, were de-signed and synthesized by PCR using the primers indicatedin table 1. All the fragments were purified by the QIAEX IIgel extraction kit (Qiagen, Hilden, Germany) beforerestriction digestion with AflII and HindIII and then sub-cloned into the corresponding sites of the expression vectorpASH. The final expression constructs were verified bysequencing.
Tagging of S6 Fragments Lacking the 3 Authentic NLSswith the Heterologous SV40 NLS
To produce the SV40 NLS–tagged S6 constructs, theoligonucleotides T-Ag1 and T-Ag2 (table 2) were hybrid-ized giving rise to a double-stranded DNA fragment with aNotI site at the 5# end and an EcoRI site at the 3# end. TheSV40 NLS tag was inserted into NotI and EcoRI sites of theplasmid KS-S6DNLS1-3, which contained the S6(3–240)fragment with the triple NLS deletion in its EcoRI andHindIII sites. The correct frame of the tag insertion wasverified by sequencing. After transformation of E. coliXL1-Blue cells, the plasmid was isolated and purified.The fragment containing the SV40 NLS tag and the S6sequence was excised by NotI and HindIII and ligatedinto the corresponding sites of pKHlacZ in order to fusethe S6 construct to the 5# end of the b-galactosidase gene.The hybrid gene was in the correct reading frame ofthe ATG codon of the pKHlacZ vector as was verifiedby sequencing.
The S6 deletion mutant lacking all 3 authentic NLSs ofS6 was the one published previously (Schmidt et al. 1995)corresponding to amino acids 3–240 of human ribosomalS6 protein. The DNA sequence was obtained as an EcoRI/HindIII fragment, which was subcloned into pUC18. To ob-tain the correct reading frame in the planned construct, anexcess single cytidine residue adjacent to the EcoRI site hadto be deleted. After the successful deletion, the fragmentwas inserted into the HindIII/EcoRI site of pBluescriptKS(�), and the correct sequence was verified. TheS6(3–240) sequence was excised from the pBluescript vec-tor with NotI/HindIII and cloned into the correspondingsites of pKHlacZ. Sequencing of the final construct showedthat the open reading frame of S6 was correctly initiatedwith the AUG codon of pKHlacZ giving rise to the ex-pected fusion protein. The cDNA fragment coding forS6(173–249,D1–3) was obtained by PCR from an S6 tem-plate lacking the 3 authentic NLSs using the primers listedin table 1. The PCR product was digested with AflII/HindIIIand ligated into the corresponding sites of pASH-SV40(Lipsius et al. 2005).
Cell Culture Techniques
Cell culture, transfection, and intracellular localizationof fusion proteins by indirect immunostaining as well ashistochemical localization of b-galactosidase activitywere carried out as described previously (Lipsius et al.2005).
ResultsDelineation of Nobis1 in the Central Region of S6
In prokaryotes, early ribosomal binding proteins typ-ically fold into globular structures that bind to multistemjunctions, thereby folding and fixing the conformation ofrRNA helices in their vicinity (Brodersen et al. 2002).Human ribosomal protein S6 is categorized as a primaryRNA-binding protein because it associates with early
Table 2Oligonucleotides Used for Site-Directed Mutagenesis andHybrid Formation
Constructs Sequence
NLS3 wt 5#-GGCCGCAAAAGAAGAAGAATA-3#NLS3 wt 3#-CGTTTTCTTCTTCTTATTCGA-5#R230G 5#-GGCCGCAAAGGAAGAAGAATA-3#R231G 3#-CGTTTCCTTCTTCTTATTCGA-5#R231V 5#-GGCCGCAAAGTAAGAAGAATA-3#R231V 3#-CGTTTCATTCTTCTTATTCGA-5#R231E 5#-GGCCGCAAAGAAAGAAGAATA-3#R231E 3#-CGTTTCTTTCTTCTTATTCGA-5#D215/216 5#-CCAGTGAATTCGGCCATGAAGGAGGCTAAG-3#D223/224 5#-AAGGAGGCTAAGGAGCAGGAACAAATTGCG-3#T-Ag1 5#-GGCCGCCCGAAAAAAAAACGCAAAGTGG-3#T-Ag2 3#-CGGGCTTTTTTTTTGCGTTTCACCTTAA-5#
NOTE.—Oligonucleotides were purchased from MWG Biotech AG (Ebersberg,
Germany) and used either for site-directed mutagenesis or hybrid formation. The
nucleotides in bold represent codons of site-directed mutagenesis.
582 Kundu-Michalik et al.
at Medical C
ollege of Wisconsin L
ibraries-Serials on July 3, 2015http://m
be.oxfordjournals.org/D
ownloaded from

precursors to the 18S rRNA in the nucleolus (Bernsteinet al. 2004; Ferreira-Cerca et al. 2005). Comparison of eu-karyotic S6 amino acid sequences revealed a high sequenceconservation between amino acids 54 and 154 implying thatthis region plays a vital role in the 40S subunit assemblyand/or ribosomal function.
The b-galactosidase fusion construct containing thefragment S6(3–183) was inefficiently targeted to the nu-cleus, which was attributed to the weak nature of NLS1(Schmidt et al. 1995). Appending the C-terminal segment,S6(210–249) containing the stronger NLS3 to the con-structs greatly increased the efficiency of nuclear importof the corresponding fusion proteins and made it possibleto examine the amino-terminal region of the S6 protein(1–170) for its nucleolar binding capability (fig. 1; supple-mentary fig. S1, Supplementary Material online for a colorversion). The first construct, S6(1–249,D171–209),directed the b-galactosidase reporter into the nucleolusimplying that the amino-terminal half of S6 contains a nu-cleolar binding domain (fig. 1).
A rough estimate of the location of this Nobis was ob-tained using the construct S6(1–249,D140–209) fused tob-galactosidase, which accumulated exclusively in the nu-cleolus (fig. 1). A series of amino-terminal deletions of S6
was studied to narrow down the region capable of nucleolarbinding: 115–139, 100–139, 75–139, 64–139, and 53–139.Only the b-galactosidase fusion proteins containingS6(53–249,D140–209) (data not shown) and S6(64–249,D140–209) (fig. 1) accumulated in the nucleolus, whereasall shorter fragments starting with S6(75–249,D140–209)were merely imported into the nucleoplasm (fig. 1). Theamino-terminal border of Nobis1 was clearly defined bythe construct S6(72–249,D140–209) that included thehighly conserved tripeptide RVR as an N-terminal exten-sion of the S6(75–249,D140–209) fragment (fig. 1 and sup-plementary fig. S3, Supplementary Material online).
To further narrow down Nobis1 by defining its C-terminal border, additional constructs containing theexogenous SV40 NLS were generated by consecutivelyshortening the C-terminus of the S6(72–139) fragment(fig. 1). The fragments S6(72–120) and S6(72–102) gaverise to a succinct nucleolar staining, whereas theS6(72–98) fragment showed, in addition to a distinct nucle-olar accumulation, a fainter staining of the nucleoplasm.Shortening by 4 amino acids at the C-terminus producedfragment S6(72–94), which was completely excluded fromthe nucleolus recognizable by the dark spots appearing inthe stained nucleoplasm (fig. 1).
FIG. 1.—Nobis1 in the central region of S6. A set of deletion mutants starting from S6(1–170) was constructed by employing the endogenousbipartite NLS3 to identify the minimal nucleolar binding sequence, Nobis1. Decreasingly shorter N-termini of the fragment S6(1–139) were tested fortheir nucleolar binding ability. A second set of constructs with an exogenous SV40 NLS attached to the N-terminus was generated by PCR, in which theC-terminus was consecutively shortened. All fusion proteins were expressed in COS cells. N, nuclear; No, nucleolar. Scale bar, 5 lm.
Nucleolar Accumulation of Ribosomal Protein S6 583
at Medical C
ollege of Wisconsin L
ibraries-Serials on July 3, 2015http://m
be.oxfordjournals.org/D
ownloaded from

In summary, the 27 amino acids of S6(72–98) repre-senting Nobis1 have the capability to lead to an accumula-tion of the fusion protein in the nucleolus. This S6 peptidesequence most probably interacts with the ribosomal 18SrRNA, thereby sequestering the fusion protein in the nucle-olus. These experiments demonstrate that a Nobis lackingany endogenous NLS resides in the central and evolution-ary highly conserved region of S6 (supplementary fig. S3,Supplementary Material online).
NLS3 of Human Ribosomal Protein S6 Is of BipartiteNature
Incorporation of the stronger NLS3 not only improvedthe efficiency of nuclear import but also surprisingly al-lowed for nucleolar accumulation, which prompted us tocarry out a detailed characterization of NLS3. The follow-ing substitutions in the tetrapeptide sequence 230KRRR233
of the arginine at position 231, R231V, R231G, and R231E,gave rise to nonfunctional NLSs unable to direct the fusedb-galactosidase into the nucleoplasm (fig. 2; supplementaryfig. S2, Supplementary Material online for a color version).This experimental result was predicted from the NLS con-sensus sequence K-R/K-X-R/K postulated by Chelsky et al.(1989). Remarkably, all 3 mutations were ineffective in theS6 context of the S6(215–240) fragment because the b-galactosidase fusion construct was still targeted into the nu-cleus (fig. 2). Hence, additional sequence elements of S6 aremost probably responsible for the unexpected highly effi-cient nuclear import unraveling the bipartite nature ofNLS3. Deletions of the dipeptides KR, which are 5 and13 amino acids upstream of the mutated tetrapeptide, re-vealed that 215KR216 is essential for nuclear targeting inthe presence of the mutated peptide KVRR (fig. 2). Thefragment S6(215–240) was incapable of accomplishing nu-cleolar binding and, therefore, suitable to use as a nuclearimport signal in the series of constructs presented in figure 1.
In short, NLS3 represents a bipartite signal in humanribosomal protein S6 in contrast to the previously delin-
eated monopartite NLS1 and NLS2 (Schmidt et al.1995). Interestingly, this bipartite signal has been con-served over almost 1 billion years because it is also presentin ribosomal protein S6A of S. cerevisiae (Lipsius et al.2005).
Nobis2: A Minimal Nucleolar Binding DomainEncompassing NLS2
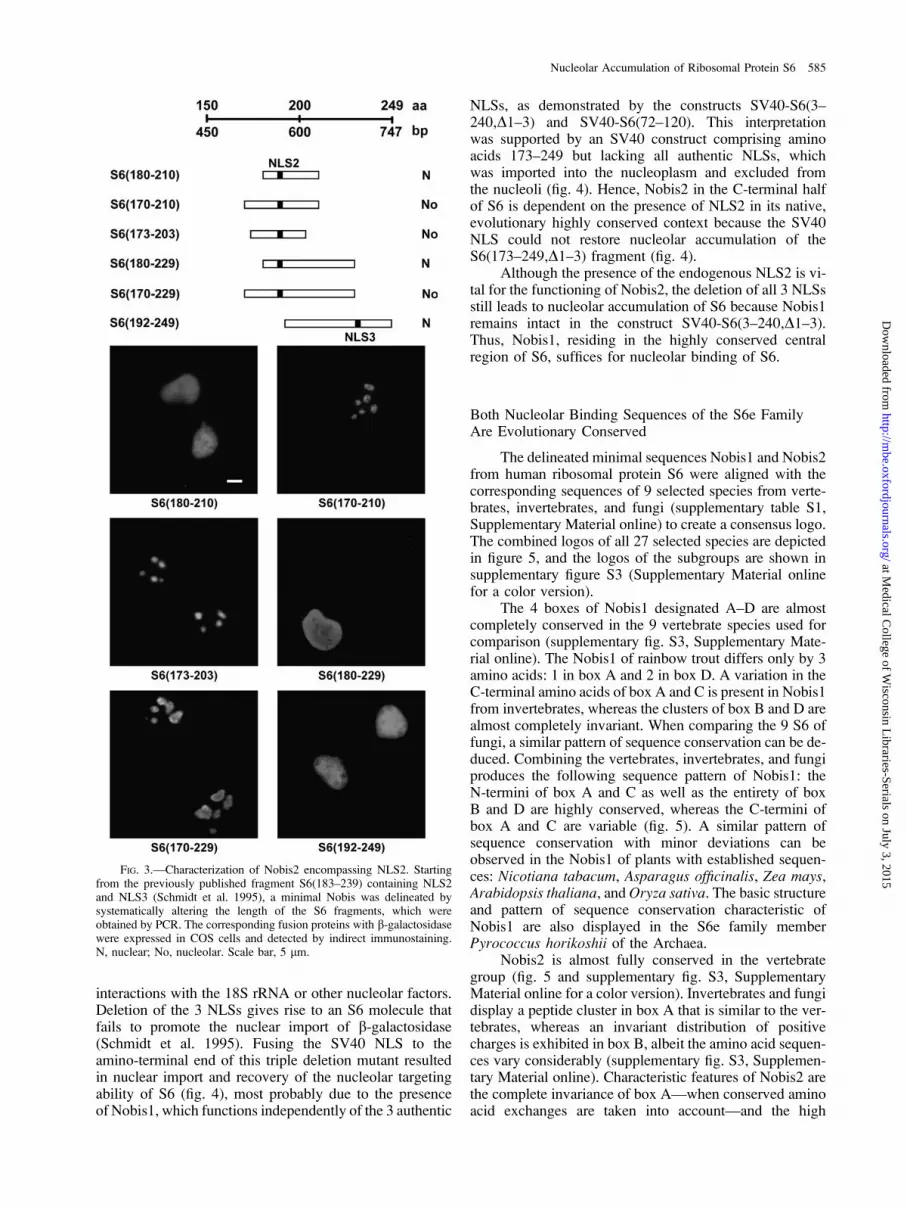
The sequence of human ribosomal protein S6 containsthe peptide 188KRRRIALKKQRTKKNK203, which ishighly reminiscent of the nucleolar localization signals ofthe retroviral proteins rex, rev, and tat (Hatanaka 1990).Interestingly, this peptide sequence is the core elementof the rather large fragments, S6(138–198) andS6(183–240), which targeted b-galactosidase into the nu-cleolus (Schmidt et al. 1995). Based on these observations,the existence of a shorter nucleolar binding domain in theregion of NLS2 was postulated. To test this hypothesis, theS6(180–210) fragment fused to the N-terminus of b-galac-tosidase was constructed (fig. 3). The corresponding fusionprotein was imported into the nucleus due to the presenceof NLS2, however, it was excluded from the nucleolus(fig. 3). Extension of this peptide sequence by 10 aminoacids at the N-terminus gave rise to the S6 fragmentS6(170–210) which, when fused to b-galactosidase, wasable to target the reporter protein exclusively into the nu-cleolus (fig. 3). In an attempt to define the C-terminalboundary of Nobis2, 12 amino acids were removed fromS6(170–210) giving rise to S6(170–198), which abolishednucleolar accumulation (data not shown).
A sequence comparison of the eukaryotic S6e familymembers called our attention to an evolutionary highly con-served S6 peptide cluster between 2 proline residues:174PKIQRLVTP182 (supplementary fig. S3, SupplementaryMaterial online). The shortest construct, S6(173–203), con-taining this peptide imported the fusion protein into the nu-cleolus (fig. 3). Presumably, the evolutionary conservedcluster between amino acids 174 and 182 functions as anessential domain vital for nucleolar binding of this S6 fu-sion protein (fig. 3). The fusion constructs, S6(180–229)and S6(170–229), support the observation that the highlyconserved peptide leads to an accumulation of the fusionproteins in the nucleolus (fig. 3). The fragment S6(192–249), which is an N-terminal extension of S6(210–249)used for the characterization of Nobis1, is also imported in-to the nucleus due to NLS3. A distinct nucleolar staining isabsent because NLS2 and the highly conserved peptidecluster were removed (fig. 3).
Taken together, the minimal Nobis2 reminiscent ofretroviral nucleolar targeting signals consists of a clusterof evolutionary highly conserved amino acids encompass-ing NLS2 as the core element (supplementary fig. S3, Sup-plementary Material online).
Nobis1 Suffices for Accumulation of S6 in the Nucleolus
The 13 basic amino acids of the 3 NLSs contribute 13positive charges, which may prove essential for ionic
FIG. 2.—NLS3 of S6 is a bipartite import signal. The tetrapeptide230KRRR233 and the fragment S6(215–240) were mutated by site-directedmutagenesis, fusion proteins with b-galactosidase were expressed in COScells, and the intracellular distribution of the constructs was visualized byX-gal staining. C, cytoplasmic; N, nuclear. Scale bar, 5 lm.
584 Kundu-Michalik et al.
at Medical C
ollege of Wisconsin L
ibraries-Serials on July 3, 2015http://m
be.oxfordjournals.org/D
ownloaded from
interactions with the 18S rRNA or other nucleolar factors.Deletion of the 3 NLSs gives rise to an S6 molecule thatfails to promote the nuclear import of b-galactosidase(Schmidt et al. 1995). Fusing the SV40 NLS to theamino-terminal end of this triple deletion mutant resultedin nuclear import and recovery of the nucleolar targetingability of S6 (fig. 4), most probably due to the presenceof Nobis1, which functions independently of the 3 authentic
NLSs, as demonstrated by the constructs SV40-S6(3–240,D1–3) and SV40-S6(72–120). This interpretationwas supported by an SV40 construct comprising aminoacids 173–249 but lacking all authentic NLSs, whichwas imported into the nucleoplasm and excluded fromthe nucleoli (fig. 4). Hence, Nobis2 in the C-terminal halfof S6 is dependent on the presence of NLS2 in its native,evolutionary highly conserved context because the SV40NLS could not restore nucleolar accumulation of theS6(173–249,D1–3) fragment (fig. 4).
Although the presence of the endogenous NLS2 is vi-tal for the functioning of Nobis2, the deletion of all 3 NLSsstill leads to nucleolar accumulation of S6 because Nobis1remains intact in the construct SV40-S6(3–240,D1–3).Thus, Nobis1, residing in the highly conserved centralregion of S6, suffices for nucleolar binding of S6.
Both Nucleolar Binding Sequences of the S6e FamilyAre Evolutionary Conserved
The delineated minimal sequences Nobis1 and Nobis2from human ribosomal protein S6 were aligned with thecorresponding sequences of 9 selected species from verte-brates, invertebrates, and fungi (supplementary table S1,Supplementary Material online) to create a consensus logo.The combined logos of all 27 selected species are depictedin figure 5, and the logos of the subgroups are shown insupplementary figure S3 (Supplementary Material onlinefor a color version).
The 4 boxes of Nobis1 designated A–D are almostcompletely conserved in the 9 vertebrate species used forcomparison (supplementary fig. S3, Supplementary Mate-rial online). The Nobis1 of rainbow trout differs only by 3amino acids: 1 in box A and 2 in box D. A variation in theC-terminal amino acids of box A and C is present in Nobis1from invertebrates, whereas the clusters of box B and D arealmost completely invariant. When comparing the 9 S6 offungi, a similar pattern of sequence conservation can be de-duced. Combining the vertebrates, invertebrates, and fungiproduces the following sequence pattern of Nobis1: theN-termini of box A and C as well as the entirety of boxB and D are highly conserved, whereas the C-termini ofbox A and C are variable (fig. 5). A similar pattern ofsequence conservation with minor deviations can beobserved in the Nobis1 of plants with established sequen-ces: Nicotiana tabacum, Asparagus officinalis, Zea mays,Arabidopsis thaliana, andOryza sativa. The basic structureand pattern of sequence conservation characteristic ofNobis1 are also displayed in the S6e family memberPyrococcus horikoshii of the Archaea.
Nobis2 is almost fully conserved in the vertebrategroup (fig. 5 and supplementary fig. S3, SupplementaryMaterial online for a color version). Invertebrates and fungidisplay a peptide cluster in box A that is similar to the ver-tebrates, whereas an invariant distribution of positivecharges is exhibited in box B, albeit the amino acid sequen-ces vary considerably (supplementary fig. S3, Supplemen-tary Material online). Characteristic features of Nobis2 arethe complete invariance of box A—when conserved aminoacid exchanges are taken into account—and the high
FIG. 3.—Characterization of Nobis2 encompassing NLS2. Startingfrom the previously published fragment S6(183–239) containing NLS2and NLS3 (Schmidt et al. 1995), a minimal Nobis was delineated bysystematically altering the length of the S6 fragments, which wereobtained by PCR. The corresponding fusion proteins with b-galactosidasewere expressed in COS cells and detected by indirect immunostaining.N, nuclear; No, nucleolar. Scale bar, 5 lm.
Nucleolar Accumulation of Ribosomal Protein S6 585
at Medical C
ollege of Wisconsin L
ibraries-Serials on July 3, 2015http://m
be.oxfordjournals.org/D
ownloaded from

density of positive charges distributed in 3 unequally sizedclusters that are separated by 2 doublets of uncharged,mostly aliphatic amino acids in box B (fig. 5). This patternis also present in the Nobis2 of plants (N. tabacum, A. offi-cinalis, Z. mays, A. thaliana, and O. sativa).
Interestingly, the S6e family members of Archaea,which have shorter protein sequences that are discontinuedsoon after Nobis1, do not contain an rRNA-binding se-quence comparable to Nobis2.
DiscussionDifferential Import Efficiencies of the 3 S6 NLSs
The efficiency of the S6 delivery to the nucleus is ex-ceptionally high because no free S6 protein, that is, not as-sembled into 40S subunits, can be detected in the cytoplasmduring interphase (Wool and Stoffler 1976). Apparently, the
import machinery flawlessly and rapidly translocates nearly3,000 S6 molecules per min per HeLa cell from the cyto-plasm into the nucleus (Kruppa A, Kruppa J, unpublisheddata).
Human ribosomal protein S6 has 3 NLSs each able todirect the reporter protein b-galactosidase into the nucleus,thereby leading to the accumulation of the correspondingfusion protein in the nucleolus (Schmidt et al. 1995).Detailed analysis revealed the bipartite nature of NLS3in contrast to the monopartite NLS1 and NLS2 (fig. 2).Human S6 and yeast S6A protein are endowed with 3distinct import signals differing in their transport efficien-cies (Schmidt et al. 1995; Lipsius et al. 2005), albeit a sin-gle NLS is sufficient to direct a reporter protein into thenucleus in vivo. Phosphorylation of the serine cluster atthe C-terminus may affect the adjacent NLS3 by decreasingthe affinity of the target sequence for the import factors.Hence, nuclear import would primarily have to rely onNLS2 because NLS1 is a weak import signal (Schmidt et al.1995). Eukaryotic cells possess different importins withdistinct NLS-binding specificities; therefore, the affinityof the importin–NLS interaction (Fontes et al. 2003) isthe critical parameter in determining transport efficiency.The limiting cellular concentration of these importins aswell as of the guanosine triphosphate–binding factor Raninfluence the extent of nucleocytoplasmic transport (Janset al. 2000).
Differences in import efficiencies within the 3 func-tionally distinct NLSs of ribosomal protein L5 have alsobeen observed (Claussen et al. 1999; Rosorius et al. 2000).Only L5-NLS1 binds to a number of importins in vitro.However, in vivo L5-NLS1 and L5-NLS3 mediate the nu-clear import of the 5S RNP. On the contrary, L5-NLS2 isincapable of translocating a heterologous 5S RNP into thenucleoplasm (Claussen et al. 1999).
Remarkably, the number, nature, and position of thetargeting signals in S6 and L5 are evolutionary conserved.Specifically, the number of NLSs—2 monopartite and 1bipartite—as well as their positions at the C-terminus of
FIG. 4.—Nucleolar binding of the S6 fusion proteins lacking the 3 authentic NLSs. The triple NLS deletion mutant, S6(3–240,D1–3), was taggedwith the SV40 NLS at the N-terminus, and the corresponding cDNA was cloned into pKHlacZ. S6(3–240) containing the 3 authentic NLSs is depictedas a control. In addition, a fragment containing Nobis1 located in the central region of S6 is shown, which is amino terminally tagged with the classicalSV40 NLS. The S6 cDNA fragment coding for S6(173–249,D1–3) was cloned into pASH-SV40, and the b-galactosidase fusion protein containing anSV40 NLS at the N-terminus was expressed in COS cells. N, nuclear; No, nucleolar. Scale bar, 5 lm.
FIG. 5.—Consensus sequences of Nobis1 and Nobis2 from theeukaryotic S6e family. The conserved Nobis1 (line 1) and Nobis2 (line 2)of 27 species listed in supplementary table S1 (Supplementary Materialonline) were aligned, and a consensus sequence logo was created usingWebLogo (Schneider and Stephens 1990; Crooks et al. 2004) available at:http://weblogo.berkley.edu/logo.cgi.
586 Kundu-Michalik et al.
at Medical C
ollege of Wisconsin L
ibraries-Serials on July 3, 2015http://m
be.oxfordjournals.org/D
ownloaded from

ribosomal protein S6 have been conserved in vertebrates,invertebrates, and fungi (Ruvinsky and Meyuhas 2006).
Characteristic Features of the 2 S6 Nucleolar BindingSequences Conserved throughout Evolution
After entering the nucleoplasm, the import complexesdissociate releasing the S6 protein. Given that there is nostructural or functional evidence for a membrane barrierseparating the nucleolus from the surrounding nucleo-plasm, the imported soluble S6 protein should migrate intothe nucleolar compartment where it associates with the pre-rRNA as an early binding protein required for efficient earlycleavages (Ferreira-Cerca et al. 2005). S6 has been local-ized to the small head region of the 40S subunit near themRNA/tRNA-binding site (Volarevic and Thomas 2001).Cross-linking studies have revealed that a region of theS6 molecule extends to the subunit interphase where itcross-links with L24 (Uchiumi et al. 1986). S6 interactswith components of the ternary initiation complex, thuscontributing to the formation of the P-site on the small sub-unit (Bommer et al. 1980).
For the prokaryotic 30S and 50S ribosomal subunits,the specific binding interactions between ribosomal pro-teins and the corresponding rRNA molecules have beenprecisely defined by X-ray crystallography (Brodersenet al. 2002; Klein et al. 2004). The atomic resolution struc-tures of these subunits have demonstrated that most ribo-somal proteins are located at the surface of therespective particle. The 3-dimensional structures of theseproteins display globular domains with either internal ex-tended a or b hairpin loops or long peptide extensions attheir N- or C-termini. These structural motifs deeply pen-etrate into the interior of the ribosomal subunit anchoringeach protein to the RNA core, thereby increasing thepercentage of protein surface interacting with the rRNA(Brodersen et al. 2002; Klein et al. 2004).
In analogy to the shape of the prokaryotic ribosomalproteins and their RNA interactions in stabilizing the 3-dimensional architecture of the subunits, eukaryotic ribo-somal proteins should also possess 1 or several regions thatserve as rRNA– and/or protein–protein binding domains(Ramakrishnan and Moore 2001) giving rise to nucleolarlocalization.
A single Nobis, either Nobis1 or Nobis2, is sufficientto lead to an exclusive accumulation of S6 in the nucleolus(figs. 1 and 3). The 27-amino-acid-long Nobis1—(G)RVRLLLSKGHSCYRPRRTGERKRKSVR(G)—con-tains 12 basic residues amounting to 41% in this oligopeptidecomparedwithonly27%basicresiduesintheentireS6protein.The N- and C-termini of Nobis1 are flanked by glycinesthat provide a certain conformational flexibility to Nobis1.Inherent to the amino acid sequence are several constraintsthat affect the 3-dimensional folding of Nobis1 preventingthe formation of an extended a-helical structure: 1) electro-static repulsion between successive basic residues (bold); 2)bulkiness of adjacent R-groups (underlined) in Ser, Cys,and Thr; and 3) occurrence of Pro and Gly residues (bolditalics).Thus, it is assumed that the specific featuresof thisNo-bisareaperfectmatchforthe18SrRNA;thisS6protein–rRNAinteraction occurs early during particle assembly in the GC
of the nucleolus (Kruger et al. 2007) giving rise to thesmall-subunit processome (Bernstein et al. 2004). Specificbinding of ribosomal protein S6 to distinct nucleolar proteinswas not detected during protein interaction mapping in S.cerevisiae (Gavin et al. 2006; Reguly et al. 2006).
Taken together, the sequence comparisons (fig. 5 andsupplementary fig. S3, Supplementary Material online)demonstrate that Nobis1 plays an important role in thephylogenetic domain of Eukarya and was already estab-lished in the Archaea, which lack a nuclear compartment,implying that the highly basic Nobis1 is interacting witha specific, presumably evolutionary conserved rRNA se-quence. It has not escaped our notice that nature employedexisting basic amino acid sequences as nucleolar bindingsequences after the appearance of the nucleus.
As mentioned in the results, Nobis2 is dependent onthe functional NLS2 (fig. 4). Nobis2—APKIQRLVTPRVLQHKRRRIALKKQRTKKNK—comprises of 31amino acids, 12 of which are basic in nature. The basic res-idues correspond to 39% in this oligopeptide comparedwith only 27% in the entire S6 protein. The structural con-straints mentioned for Nobis1 also hold true for Nobis2,thereby creating a positively charged, flexible binding se-quence capable of optimally interacting with the 18S rRNAbackbone.
Nobis1 and Nobis2 differ in 1 important aspect:Nobis1 resides in a region of the S6 protein sequence thatlacks any NLS, whereas the functioning of Nobis2 iscrucially dependent on NLS2 because deletion of NLS2in SV40-S6(173–249,D1–3) concomitantly results in a lossof nucleolar accumulation (fig. 4). Thus, integration of anNLS into the second Nobis of S6 ascertains the efficientrecognition of the newly synthesized ribosomal proteinby the nuclear import machinery, thereby leading to a rapidimport into the nucleoplasm. Clearing the cytoplasm of thehighly basic ribosomal proteins seems vital to avoid the del-eterious binding of ribosomal proteins to mRNAs thatwould otherwise interfere with the translation process(Wool and Stoffler 1976; Jakel and Gorlich 1998). Thenucleolar binding sequences are essential for driving thesubunit assembly in the nucleolus, thus positively influenc-ing the growth rate of cells. The presence of an NLS in aNobis increases the efficiency of the ribosomal proteindelivery into the nucleus, for example, for Nobis2 of S6eand for the Nobis of S25e. An NLS as an integral buildingblock is also found in the nucleolar binding sequences ofhuman S7 and L7a (table 3).
In essence, although nuclear targeting signals increaseimport efficiency, NLSs are superfluous because ribosomalproteins can diffuse through the nuclear pore into the nu-cleoplasm due to their low molecular weight, yet the pres-ence of a Nobis is indispensable for nucleolar binding,thereby facilitating subunit assembly.
Does the C-terminal Serine Cluster of S6 Contribute tothe 18S rRNA Binding?
The C-terminus of ribosomal protein S6 contains theserine cluster of phosphorylation sites, which play an im-portant role as determinants of cell size, cell proliferation,
Nucleolar Accumulation of Ribosomal Protein S6 587
at Medical C
ollege of Wisconsin L
ibraries-Serials on July 3, 2015http://m
be.oxfordjournals.org/D
ownloaded from

and glucose homeostasis (Ruvinsky et al. 2005; Ruvinskyand Meyuhas 2006). In vertebrates, the sequence of the ser-ine cluster with its 5 phosphorylation sites is completelyconserved, whereas the amino acid sequence varies in in-vertebrates (Ruvinsky and Meyuhas 2006). In contrast,fungi have to rely on a shorter C-terminus with just 2 serinephosphorylation sites (Lipsius et al. 2005).
By proton nuclear magnetic resonance spectroscopy,Katahira et al. (1996) obtained a solution structure forthe C-terminal end, S6(217–249), containing the serinecluster for phosphorylation. This peptide folds into ana-helix between E222 and R238, a distorted helical struc-ture for the following 3 residues, and a flexible tail lackingany secondary structure starting from S242 that must re-main freely accessible to the p70 S6 kinase, which usesthe tetrapeptide KRRR of NLS3 as its recognition sitefor the sequential phosphorylation of the 5 serine residuesin the C-terminal tail. Phosphatase 1, which sequentiallydephosphorylates the serine phosphates, must also have ac-cess to the S6 tail. Although this S6 C-terminal region isunstructured similar to the terminal extension of prokary-otic ribosomal proteins, this domain cannot anchor S6 tothe 18S rRNA because it is easily cleaved off by trypsin(Wettenhall and Cohen 1982). The last 13 amino acidsat the C-terminus of human S6, which are absent in theS6e members of fungi, cannot be involved in stabilizingthe S6 binding to the 18S rRNA because the C-tail hasto remain flexible to allow for enzymatic interactions.
Evolutionary Highly Conserved Nucleolar BindingSequences Occur in Numerous Ribosomal Proteins
Our detailed analyses clearly show that the nucleolarbinding sequences of S6 reside in evolutionary highly con-served peptide clusters (fig. 5 and supplementary fig. S3,Supplementary Material online). Does this observation alsohold true for the nucleolar binding sequences of other ribo-somal proteins that have been published? Our conclusion iscorroborated by the 2 publications (Kubota et al. 1999;Timmers et al. 1999) that delineated a single Nobis of yeastand human S25, respectively, which consist of a highlyconserved cluster that has been maintained over more than1 billion years of evolution (table 3 and supplementaryfig. S4, SupplementaryMaterial online). Previously, Annilo
et al. (1998) have identified a single Nobis in human S7containing a high percentage of basic amino acids that isbuilt from 2 clusters, which are evolutionary highly con-served, analogous to S6. In human S19, 2 short nucleolarbinding sequences have been characterized: 1 located at theN-terminus and the other at the C-terminus (Da Costa et al.2003). A single amino acid exchange in either Nobis fails tolocalize S19 in the nucleolus and leads to Diamond–Black-fan anemia (DBA). The C-terminal Nobis of S19 is insertedbetween 2 glycine residues similar to Nobis1 of S6. Themutations in S19 of DBA patients clearly demonstratethe importance of proper localization of ribosomal proteinsfor the development of a healthy organism.
Ribosomal proteins of the large subunit also containevolutionary highly conserved nucleolar binding sequencesas exemplified by human L22 (Shu-Nu et al. 2000) and ratL31 (Quaye et al. 1996), which are 6 and 5 residues long,respectively, whereas human L5 (Rosorius et al. 2000) dis-plays 2 nucleolar binding sequences, both of which have anintegrated NLS (table 3).
Supplementary Material
Supplementary table S1 and figures S1–S4 are avail-able at Molecular Biology and Evolution online (http://www.mbe.oxfordjournals.org/).
Acknowledgments
We thank Antonie Kruppa for her critical reading ofthe manuscript and her valuable suggestions.
Literature Cited
Andersen JS, Lam YW, Leung AK, Ong SE, Lyon CE,Lamond AI, Mann M. 2005. Nucleolar proteome dynamics.Nature. 433:77–83.
Andersen JS, Lyon CE, Fox AH, Leung AKL, Lam YW,Steen H, Mann M, Lamond AI. 2002. Directed proteomicanalysis of the human nucleolus. Curr Biol. 12:1–12.
Annilo T, Karis A, Hoth S, Rikk T, Kruppa J, Metspalu A. 1998.Nuclear import and nucleolar accumulation of the humanribosomal protein S7 depends on both a minimal nuclear
Table 3Nuclear Localization Signals and Nucleolar Binding Sequences in Ribosomal Proteins
Species RP
NLS Nobis
ReferenceNumber Length Type Number Length
Homo sapiens S6 3 4/4/17 2 monopartite/1 bipartite 2 27/31 This paperS7 1 21 Bipartite-like 1 21 Annilo et al. 1998S19 1 21 Bipartite 2 17/23 Da Costa et al. 2003S25 1 4 Monopartite 1 17 Kubota et al. 1999L5 3 17/11 Monopartite 2 17/11 Rosorius et al. 2000L7a 3 29/49/120 ND 1 49 Russo et al. 1997L22 1 4 Monopartite 1 6 Shu-Nu et al. 2000
Rattus norvegicus L31 1 6 Monopartite 1 6 Quaye et al. 1996Saccharomyces cerevisiae S6A 3 14/18/17 2 monopartite/1 bipartite 2 62/42 Lipsius et al. 2005
S25 2 26/9 Bipartite/Mata2-like 1 26 Timmers et al. 1999
NOTE.—RP, ribosomal protein; ND, not determined.
588 Kundu-Michalik et al.
at Medical C
ollege of Wisconsin L
ibraries-Serials on July 3, 2015http://m
be.oxfordjournals.org/D
ownloaded from

localization sequence and an adjacent basic region. BiochemBiophys Res Commun. 249:759–766.
Bernstein KA, Bleicherst F, Bean JM, Cross FR, Baserga SJ.2007. Ribosome biogenesis is sensed at the start cell cyclecheckpoint. Mol Biol Cell. 18:953–964.
Bernstein KA, Gallagher JE, Mitchell BM, Granneman S,Baserga SJ. 2004. The small-subunit processome is a ribosomeassembly intermediate. Eukaryot Cell. 3:1619–1626.
Bommer UA, Noll F, Lutsch G, Bielka H. 1980. Immunochem-ical detection of proteins in the small subunit of rat liverribosomes involved in binding of the ternary initiationcomplex. FEBS Lett. 111:171–174.
Brodersen DE, Clemons WM Jr, Carter AP, Wimberly BT,Ramakrishnan V. 2002. Crystal structure of the 30 Sribosomal subunit from Thermus thermophilus: structure ofthe proteins and their interactions with 16 S RNA. J Mol Biol.316:725–768.
Carmo-Fonseca M, Mendes-Soares L, Campos I. 2000. To be ornot to be in the nucleolus. Nat Cell Biol. 2:E107–E112.
Chelsky D, Ralph R, Jonak G. 1989. Sequence requirements forsynthetic peptide-mediated translocation to the nucleus. MolCell Biol. 9:2487–2492.
Chen D, Huang S. 2001. Nucleolar components involved inribosome biogenesis cycle between the nucleolus andnucleoplasm in interphase cells. J Cell Biol. 153:169–176.
ClaussenM,Rudt F, Pieler T. 1999. Functionalmodules in ribosomalprotein L5 for ribonucleoprotein complex formation andnucleocytoplasmic transport. J Biol Chem. 274:33951–33958.
Crooks GE, Hon G, Chandonia JM, Brenner SE. 2004. WebLogo:a sequence logo generator. Genome Res. 14:1188–1190.
Da Costa L, Tchernia G, Gascard P, Lo A, Meerpohl J,Niemeyer C, Chassis JA, Fixler J, Mohandas N. 2003.Nucleolar localization of RPS19 protein in normal cellsand mislocalization due to mutations in the nucleolarlocalization signals in 2 Diamond-Blackfan anemia patients:potential insights into pathophysiology. Blood. 101:5039–5045.
Dingwall C, Laskey RA. 1991. Nuclear targeting sequences—aconsensus? Trends Biochem Sci. 16:478–481.
Ferreira-Cerca S, Poll G, Gleizes PE, Tschochner H, Milkereit P.2005. Roles of eukaryotic ribosomal proteins in maturationand transport of pre-18S rRNA and ribosome function. MolCell. 20:263–275.
Fontes MRM, The T, Jans D, Brinkworth RI, Kobe B. 2003.Structural basis for the specificity of bipartite nuclearlocalization sequence binding by importin-alpha. J BiolChem. 278:27981–27987.
Fromont-Racine M, Senger B, Saveanu C, Fasiolo F. 2003.Ribosome assembly in eukaryotes. Gene. 313:17–42.
Fukami-Kobayashi K, Minezaki Y, Tateno Y, Nishikawa K.2007. A tree of life based on protein domain organizations.Mol Biol Evol. 24:1181–1189.
Gavin AC, Aloy P, Grandi P, et al. 2006. (32 co-authors).Proteome survey reveals modularity of the yeast cellmachinery. Nature. 440:631–636.
Gerbi SA, Borovjan AV, Lange TS. 2003. The nucleolus: a siteof ribonucleoprotein maturation. Curr Opin Cell Biol.15:318–325.
Grandi P, Rybin V, Bassler J, et al. (12 co-authors). 2002. 90Spre-ribosomes include the 35S pre-rRNA, the U3 snoRNP,and 40S subunit processing factors but predominantly lack60S synthesis factors. Mol Cell. 10:105–115.
Granneman S, Vogelzangs J, Luhrmann R, van Venrooij WJ,Pruijn GJ, Watkins NJ. 2004. Role of pre-rRNA base pairingand 80S complex formation in subnucleolar localization of U3snoRNP. Mol Cell Biol. 24:8600–8610.
Hadijolov AA. 1985. The nucleolus and ribosome biogenesis.Wien (Austria): Springer Verlag.
Hatanaka M. 1990. Discovery of the nucleolar targeting signal.Bioessays. 12:143–148.
Hinsby AM, Kiemer L, Karlberg EO, Lage K, Fausbøll A,Juncker AS, Andersen JS, Mann M, Brunak S. 2006. A wiringof the human nucleolus. Mol Cell. 22:285–295.
Jakel S, Gorlich D. 1998. Importin beta, transportin, RanBP5 andRanBP7 mediate nuclear import of ribosomal proteins inmammalian cells. EMBO J. 17:4491–4502.
Jans DA, Xiao CY, Lam MHC. 2000. Nuclear targeting signalrecognition: a key control point in nuclear transport?Bioessays. 22:532–544.
Katahira R, Flotow H, Thomas G, Nosaka AY. 1996. Solutionstructure of the phosphorylated sites of ribosomal protein S6 by1H NMR spectroscopy. Int J Pept Protein Res. 47:282–288.
Klein DJ, Moore PB, Steitz TA. 2004. The roles of ribosomalprotein in the structure assembly, and evolution of the largeribosomal subunit. J Mol Biol. 340:141–177.
Koberna K, Malinsky J, Pliss A, Masata M, Vecerova J,Fialova M, Bednar J, Raska I. 2002. Ribosomal genes infocus: new transcripts label the dense fibrillar components andform clusters indicative of ‘‘Christmas trees’’ in situ. J CellBiol. 157:743–748.
Kruger T, Zentgraf H, Scheer U. 2007. Intranucleolar sites ofribosome biogenesis defined by the localization of earlybinding ribosomal proteins. J Cell Biol. 177:573–578.
Kubota S, Copeland TD, Pomerantz RJ. 1999. Nuclear andnucleolar targeting of human ribosomal protein S25: commonfeatures shared with HIV-1 regulatory proteins. Oncogene.18:1503–1514.
Lecompte O, Ripp R, Thierry JC, Moras D, Poch O. 2002.Comparative analysis of ribosomal proteins in completegenomes: an example of reductive evolution at the domainscale. Nucleic Acids Res. 30:5382–5390.
Leung AKL, Andersen JS, Mann M, Lamond AI. 2003. Bioinfor-matic analysis of the nucleolus. Biochem J. 376:553–569.
Lipsius E, Walter K, Leicher T, Phlippen W, Bisotti MA,Kruppa J. 2005. Evolutionary conservation of nuclear andnucleolar targeting sequences in yeast ribosomal protein S6A.Biochem Biophys Res Commun. 333:1353–1360.
Mayer C, Grummt I. 2006. Ribosome biogenesis and cell growth:mTOR coordinates transcription by all three classes of nuclearRNA polymerases. Oncogene. 25:6384–6391.
Mears JA, Connone JJ, Stagg SM, Gutell RR, Agrawal RK,Harvey SC. 2002. Modeling a minimal ribosome based oncomparative sequence analysis. J Mol Biol. 321:215–234.
Milkereit P, Kuhn H, Gas N, Tschochner H. 2003. The pre-ribosomal network. Nucleic Acids Res. 31:799–804.
Noller HF. 1991. Ribosomal RNA and translation. Annu RevBiochem. 60:191–227.
Quaye IKE, Toku S, Tanaka T. 1996. Sequence requirement fornucleolar localization of rat ribosomal protein L31. Eur J CellBiol. 69:151–155.
Ramakrishnan V, Moore PB. 2001. Atomic structure at last: theribosome in 2000. Curr Opin Struct Biol. 11:144–154.
Raska I, Shaw PJ, Cmarko D. 2006. Structure and function of thenucleolus in the spotlight. Curr Opin Cell Biol. 18:325–334.
Reguly T, Breitkretz A, Boucher R, et al. (20 co-authors). 2006.Comprehensive curation and analysis of global interactionnetworks in Saccharomyces cerevisiae. J Biol. 5:11.
RosoriusO,FriesB, StauberRH,HirschmannN,BevecD,Hauber J.2000. Human ribosomal protein L5 contains defined nuclearlocalization and export signals. J Biol Chem. 275:12061–12068.
Russo G, Ricciarelli G, Pietropaolo C. 1997. Different domainscooperate to target the human ribosomal L7a protein to thenucleus and to the nucleoli. J Biol Chem. 272:5229–5235.
Ruvinsky I, Sharon N, Lerer T, Cohen H, Stolovich-Rain M,Nir T, Dor Y, Zisman P, Meyuhas O. 2005. Ribosomal
Nucleolar Accumulation of Ribosomal Protein S6 589
at Medical C
ollege of Wisconsin L
ibraries-Serials on July 3, 2015http://m
be.oxfordjournals.org/D
ownloaded from

protein S6 phosphorylation is a determinant of cell size andglucose homeostasis. Genes Dev. 19:2199–2211.
Ruvinsky I, Meyuhas O. 2006. Ribosomal protein phosphoryla-tion: from protein synthesis to cell size. Trends Biochem Sci.31:342–348.
Scheer U, Hock R. 1999. Structure and function of the nucleolus.Curr Opin Cell Biol. 11:385–390.
Scherl A, Coute Y, Deon C, Calle A, Kindbeiter K, Sanchez JC,Grecio A, Hochstrasser D, Diat JJ. 2002. Functionalproteomic analysis of human nucleolus. Mol Biol Cell.13:4100–4109.
Schmidt C, Lipsius E, Kruppa J. 1995. Nuclear and nucleolartargeting of human ribosomal protein S6. Mol Biol Cell.6:1875–1885.
Schneider TD, Stephens RM. 1990. Sequence logos: a new way todisplay consensus sequences.NucleicAcidsRes. 18:6097–6100.
Schuwirth BS, Borovinskaya MA, Hau CW, Zhang W, Vila-Sanjurjo A, Holton JM, Cate JH. 2005. Structures of thebacterial ribosome at 3.5 A resolution. Science. 310:793–795.
Shu-Nu C, Lin CH, Lin A. 2000. An acidic amino acid clusterregulates the nucleolar localization and ribosome assembly ofhuman ribosomal protein L22. FEBS Lett. 484:22–28.
Takahashi N, Yanagida M, Fujiyama S, Hayano T, Isobe T.2003. Proteomic snapshot analyses of preribosomal ribonu-cleoprotein complexes formed at various stages of ribosomebiogenesis in yeast and mammalian cells. Mass SpectromRev. 22:287–317.
Timmers AC, Stuger R, Schaap PJ, van’t Riet J, Raue HA. 1999.Nuclear and nucleolar localization of Saccharomyces cerevi-
siae ribosomal proteins S22 and S25. FEBS Lett.452:335–340.
Tschochner H, Hurt E. 2003. Pre-ribosomes on the road from thenucleolus to the cytoplasm. Trends Cell Biol. 13:255–263.
Uchiumi T, Kikuchi M, Ogata K. 1986. Cross-linking study onprotein neighborhoods at the subunit interface of rat liverribosomes with 2-iminothiolane. J Biol Chem. 261:9663–9667.
Venema J, Tollervey D. 1999. Ribosome synthesis in Saccha-romyces cerevisiae. Annu Rev Genet. 33:261–311.
Volarevic S, Thomas G. 2001. Role of S6 phosphorylation andS6 kinase in cell growth. Prog Nucleic Acid Res Mol Biol.65:101–127.
Wettenhall RE, Cohen P. 1982. Isolation and characterization ofcyclic AMP-dependent phosphorylation sites from rat liverribosomal protein S6. FEBS Lett. 140:263–269.
Wool IG, Chan YL, Gluck A. 1995. Structure and evolution ofmammalian ribosomal proteins. Biochem Cell Biol.73:933–947.
Wool IG, Stoffler G. 1974. Structure and function of eukaryoticribosomes. In: Nomura M, Tissieres A, Lengyel P, editors.Ribosomes. Cold Spring Harbor (NY): Cold Spring HarborLaboratory. p. 417–460.
Wool IG, Stoffler G. 1976. Determination of the size of the poolof free ribosomal proteins in rat liver cytoplasm. J Mol Biol.108:201–218.
Manolo Gouy, Associate Editor
Accepted December 28, 2007
590 Kundu-Michalik et al.
at Medical C
ollege of Wisconsin L
ibraries-Serials on July 3, 2015http://m
be.oxfordjournals.org/D
ownloaded from





























