Embed Size (px)
Citation preview

Nox2-derived reactive oxygen species mediateneurovascular dysregulation in the aging mousebrain
Laibaik Park, Josef Anrather, Helene Girouard, Ping Zhou and Costantino Iadecola
Division of Neurobiology, Department of Neurology and Neuroscience, Weill Medical College of CornellUniversity, New York, New York, USA
Aging is associated with cerebrovascular dysregulation, which may underlie the increasedsusceptibility to ischemic stroke and vascular cognitive impairment occurring in the elderindividuals. Although it has long been known that oxidative stress is responsible for thecerebrovascular dysfunction, the enzymatic system(s) generating the reactive oxygen species(ROS) have not been identified. In this study, we investigated whether the superoxide-producingenzyme NADPH oxidase is involved in alterations of neurovascular regulation induced by aging.Cerebral blood flow (CBF) was recorded by laser-Doppler flowmetry in anesthetized C57BL/6 miceequipped with a cranial window (age = 3, 12, and 24 months). In 12-month-old mice, the CBFincreases evoked by whisker stimulation or by the endothelium-dependent vasodilators acetyl-choline and bradykinin were attenuated by 42, 36, and 53%, respectively (P < 0.05). In contrast,responses to the nitric oxide donor S-nitroso-D-penicillamine or adenosine were not attenuated(P > 0.05). These cerebrovascular effects were associated with increased production of ROS inneurons and cerebral blood vessels, assessed by hydroethidine microfluorography. The cerebro-vascular impairment present in 12-month-old mice was reversed by the ROS scavenger Mn (III)tetrakis (4-benzoic acid) porphyrin chloride or by the NADPH oxidase peptide inhibitor gp91ds-tat,and was not observed in mice lacking the Nox2 subunit of NADPH oxidase. These findings establishNox2 as a critical source of the neurovascular oxidative stress mediating the deleteriouscerebrovascular effects associated with increasing age.Journal of Cerebral Blood Flow & Metabolism (2007) 27, 1908–1918; doi:10.1038/sj.jcbfm.9600491; published online11 April 2007
Keywords: cerebral blood flow; endothelium-dependent vasodilation; functional hyperemia; hydroethidine; laser-Doppler flowmetry
Introduction
Aging has profound cerebrovascular effects that arethought to underlie the increased incidence ofischemic stroke and vascular cognitive impairmentaffecting older individuals (Farkas and Luiten, 2001;Rothwell et al, 2005). Aging is associated with areduction in resting cerebral blood flow (CBF) anda dysfunction of the mechanisms regulating thecerebral circulation (Faraci and Heistad, 1998; Farkasand Luiten, 2001). For example, the cerebrovascular
relaxation induced by endothelium-dependent va-sodilators, such as acetylcholine, is impaired inlarge and small cerebral arteries of older animals(Mayhan et al, 1990; Paterno et al, 1994) (fora review, see Faraci and Heistad, 1998). Further-more, the increases in CBF induced by activation ofcentral cholinergic pathways, hypercapnia, hypoxia,or hypotension are attenuated (Hoffman et al, 1981,1984, 1982; Sato et al, 2002). These cerebrovascularalterations reduce cerebrovascular reserves andincrease the susceptibility of the brain to vascularinsufficiency and ischemic injury (Farkas andLuiten, 2001). However, less is known about theeffects of aging on the increase in CBF induced bysynaptic activity, a critical homeostatic responsethat matches the energetic needs of the active brainwith the delivery of nutrients through blood flow(Iadecola, 2004).
There is increasing evidence that the impairmentin cerebrovascular function induced by aging is
Received 25 January 2007; revised 26 February 2007; accepted 27February 2007; published online 11 April 2007
Correspondence: Dr C Iadecola, Division of Neurobiology,Department of Neurology and Neuroscience, Weill MedicalCollege of Cornell University, 411 East 69th Street; KB410, NewYork, New York 10021, USA.E-mail: [email protected]
This work was supported by grants from the National Institutes of
Health (NS38252 and HL18974).
Journal of Cerebral Blood Flow & Metabolism (2007) 27, 1908–1918& 2007 ISCBFM All rights reserved 0271-678X/07 $30.00
www.jcbfm.com

mediated by oxidative stress, but the cellularsources of reactive oxygen species (ROS) and theenzymatic systems generating them have not beendefined (Faraci and Heistad, 1998; Farkas andLuiten, 2001). There are several potential sourcesof ROS in brain, including xanthine oxidase,mitochondrial enzymes, and enzymes involved innitric oxide synthesis or arachidonic acid metabo-lism (Faraci, 2006). However, NADPH oxidase hasrecently emerged as a major source of ROS inneurons, glia, and cerebral blood vessels (Abramovet al, 2005; Infanger et al, 2006; Kazama et al, 2004;Miller et al, 2006). NADPH oxidase is a multiunitenzyme initially discovered in neutrophils (Bedardand Krause 2007). It is composed of membrane-bound (p22phox and gp91phox) and cytoplasmicsubunits (p40phox, p47phox, and p67phox). The cataly-tic subunit of the enzyme gp91phox, also termed‘Nox2,’ is present in several homologs (Nox1through Nox5) (Bedard and Krause, 2007). Activa-tion of the enzyme depends on phosphorylation ofp47phox, which, in concert with the small GTPaseRac1, leads to assembly of the cytoplasmic andmembrane-bound subunits and production of super-oxide (Bedard and Krause, 2007). Although the roleof NADPH oxidase in the cerebrovascular dysfunc-tion induced by hypertension, diabetes, andamyloid-b is well established (Kazama et al, 2004;Mayhan et al, 2006; Miller et al, 2006; Park et al,2005), it is unknown whether this enzyme is alsoinvolved in the cerebrovascular alterations inducedby aging.
In this study, we examined the effect of aging onthe neurovascular and endothelial mechanismsregulating the cerebral microcirculation. We foundthat aging, in addition to its deleterious effects onendothelium-dependent vasodilation, also disruptsthe increase in CBF induced by synaptic activityin the mouse neocortex. Furthermore, we found thatNADPH oxidase-derived ROS, originating fromneurons and blood vessels, can account in full forthe cerebrovascular disturbances observed in oldermice. The findings expand our understanding of thecerebrovascular effects of aging and identify NADPHoxidase as a key source of ROS mediating thealterations in critical homeostatic mechanismscontrolling the cerebral microcirculation.
Materials and methods
All experimental procedures were approved by theInstitutional Animal Care and Use Committee of WeillCornell Medical College.
Mice
Experiments were performed in male C57BL/6 and Nox2(gp91phox)-deficient mice aged 3, 12, or 24 months. Nox2-deficient mice (Pollock et al, 1995) were obtained from our
own colony and were congenic with the C57BL/6 strain(Park et al, 2005).
General Surgical Procedures
Procedures for surgical preparation of the mice have beendescribed previously in detail (Niwa et al, 2001; Park et al,2005) and are only summarized here. Mice were anesthe-tized with isoflurane (induction: 5%; maintenance: 1% to2%). One of the femoral arteries was cannulated forrecording of arterial pressure and collection of bloodsamples. Mice were incubated and artificially ventilatedwith oxygen–nitrogen mixture adjusted to provide anarterial pO2 (paO2) of 120 to 140 mm Hg (SupplementaryTable 1). Rectal temperature was maintained at 371C usinga thermostatically controlled rectal probe connected to aheating pad. After surgery, isoflurane was graduallydiscontinued and anesthesia was maintained with ur-ethane (750 mg/kg; intraperitoneally) and a-chloralose(50 mg/kg; intraperitoneally). Throughout the experiment,the level of anesthesia was monitored by testing cornealreflexes and motor responses to tail pinch.
Monitoring of Cerebral Blood Flow
A small craniotomy (2� 2 mm) was performed to exposethe somatosensory cortex. The dura was removed, and thesite was superfused with a modified Ringer’s solution(371C; pH 7.3 to 7.4) (see Iadecola, 1992 for composition).Cerebral blood flow was continuously monitored at thesite of superfusion with a laser-Doppler probe (Vasamedic,St Paul, MN, USA) positioned stereotaxically on theneocortical surface and connected to a computerized dataacquisition system. Cerebral blood flow values wereexpressed as percent increase relative to the resting level.Zero values for CBF were obtained after the heart wasstopped by an overdose of isoflurane at the end of theexperiment.
Detection of Reactive Oxygen Species
Reactive oxygen species production was assessed usinghydroethidine as a marker, as described previously (Parket al, 2005). Hydroethidine is quickly taken up by cellsand reacts with superoxide to form a stable fluorescentproduct (2-hydroxyethidium), which binds to DNA.Hydroethidine (2mmol/L; Molecular Probes, Eugene, OR,USA) was topically superfused on the somatosensorycortex for 60 mins (Park et al, 2005). The brain was thenremoved and frozen. Brain sections (thickness, 20 mm)were cut through cortex underlying the cranial windowusing a cryostat and collected at 100mm intervals. Sectionswere examined under an Eclipse E800 fluorescencemicroscope (Nikon, Melville, NY, USA) equipped with acustom filter set designed to maximize 2-hydroxyethidiumfluorescence. Images were acquired with a computer-controlled digital monochrome camera (Coolsnap, RoperScientific, Trenton, NJ, USA) using the same fluorescencesettings in all cases. The analysis of ROS production in the
Aging and Nox2-derived ROSL Park et al
1909
Journal of Cerebral Blood Flow & Metabolism (2007) 27, 1908–1918

different conditions studied (see Experimental Protocol)was performed in a masked manner using the IPLabsoftware (Scanalytics, Fairfax, VA, USA). After back-ground subtraction of the camera dark current, pixelintensities of the fluorescent signal were quantified.Fluorescent intensities of all sections (15 to 20 per animal)were added, divided by the total number of pixelsanalyzed, and expressed as relative fluorescence units(RFU) (Park et al, 2005).
Immunocytochemistry and Confocal Microscopy
At the end of the hydroethidine superfusion, mice weredeeply anesthetized with 5% isoflurane and perfusedtranscardially with heparin sulfate (1000 U/mL) followedby 4% paraformaldehyde. Brains were removed andfrozen, and the brain area underlying the cranial windowwas sectioned in a cryostat (thickness, 14 mm). To identifyhydroethidine-positive cells, sections were processed forimmunocytochemistry for the neuronal marker NeuN(1:100, Chemicon International, Temecula, CA, USA), theastrocytic marker glial fibrillary acidic protein (GFAP,1:1000, Sigma, St Louis, MO, USA) or the endothelial cellmarker CD31 (1:100, BD Biosciences, San Diego, CA,USA). Sections were washed and incubated with cyaninedye (Cy5)-conjugated goat anti-mouse immunoglobulin G(for NeuN and GFAP; Jackson ImmunoResearch, WestGrove, PA) and goat anti-rat immunoglobulin G (for CD31;Jackson ImmunoResearch) secondary antibodies. Thespecificity of the labeling was established by omittingthe primary antibody or by preadsorption with theantigen. Images were sequentially acquired using aconfocal laser scanning microscope (Leica, Nussloch,Germany). Ethidium and Cy5 signals were pseudocoloredred and green, respectively.
Experimental Protocol
Cerebral blood flow recordings and ROS measurementswere started after arterial pressure and blood gases were ina steady state (Supplementary Table 1). All pharmacolo-gical agents were dissolved in a modified Ringer’ssolution, unless otherwise indicated.
Effect of Aging on Cerebrovascular Responses in WildType and Nox-2-Null Mice: The cranial window wassuperfused with Ringer’s solution, and the CBF increasesevoked by whisker stimulation (functional hyperemia)and endothelium-dependent and -independent vasodila-tors were tested. Cerebral blood flow responses to whiskerstimulation were recorded while gently stroking thewhiskers with a cotton-tipped applicator for 60 secs.The endothelium-dependent vasodilators acetylcholine(10mmol/L; Sigma), bradykinin (50 mmol/L; Sigma), orthe calcium ionophore A23187 (3mmol/L; Sigma) weretopically superfused for 3 to 5 mins and the evoked CBFincreases recorded. Cerebral blood flow responses to thenitric oxide donor S-nitroso-D-penicillamine (SNAP;50 mmol/L; Sigma) or to adenosine (400 mmol/L; Sigma),agents that produce vasodilation by acting directly on
vascular smooth muscles (Faraci and Heistad, 1998), werealso tested. The increase in CBF produced by hypercapniawas examined by introducing 5% CO2 in the ventilator toincrease arterial pCO2 up to 50 to 60 mm Hg. Once a stableincrease in CBF was obtained, pCO2 was returned tonormocapnia. Responses were tested in C57BL6 mice aged3, 12, or 24 months, and in Nox2 null mice aged 3 or 12months.
Effect of MnTBAP or gp91ds-tat: In some experiments,the effect of the ROS scavenger manganic (I–II)meso-tetrakis (4-benzoic acid) porphyrin (MnTBAP, 100 mmol/L;Porphyrin Products, Logna, UT, USA) was studied. Thecranial window was superfused with Ringer’s solutionand the CBF increases evoked by whisker stimulation,acetylcholine, bradykinin, A23187, SNAP, adenosine, orhypercapnia were tested. Next, the superfusion solutionwas switched to Ringer containing MnTBAP (100 mmol/L)and CBF responses were tested again 30 mins later. Inother experiments, we examined the effect of the NADPHoxidase peptide inhibitor gp91ds-tat (Rey et al, 2001) asdescribed previously (Kazama et al, 2004; Park et al,2005). Cerebrovascular responses were assessed beforeand 30 to 40 mins after superfusion with gp91ds-tat(1mmol/L; YGRKKRRQRRRCSTRIRRQL-NH2) or its scram-bled control (1mmol/L; YGRKKRRQRRRCLRITRQSR-NH2;Bio �Synthesis, Lewisville, TX, USA).
Reactive Oxygen Species Production: The experimentalprotocol for these studies was identical to that in whichCBF responses were examined. The cranial window wassuperfused for 60 mins with Ringer’s solution containinghydroethidine with or without MnTBAP, gp91ds-tat, orthe scrambled peptide. Nox2-deficient mice were alsotreated with the SOD inhibitor diethyldithiocarbamate(10 mmmol/L; topical application) to test their ability togenerate ROS independently of NADPH oxidase. At theend of superfusion, brains were removed and processedfor ROS assessment as described above.
Data Analysis
Data in text and figures are expressed as means7s.d. Two-group comparisons were analyzed by the two-tailed t-testfor dependent or independent samples, as appropriate.Multiple comparisons were evaluated by the analysis ofvariance and Tukey’s test. Statistical significance wasconsidered for P < 0.05.
Results
Aging has Profound Effects on CerebrovascularRegulation
To examine the effect of aging on cerebrovascularreactivity, we compared selected CBF responses inmice aged 3, 12, and 24 months. The increases inCBF evoked by whisker stimulation and by super-fusion with acetylcholine (10 mmol/L) or bradykinin(50 mmol/L) were markedly attenuated starting at 12
Aging and Nox2-derived ROSL Park et al
1910
Journal of Cerebral Blood Flow & Metabolism (2007) 27, 1908–1918
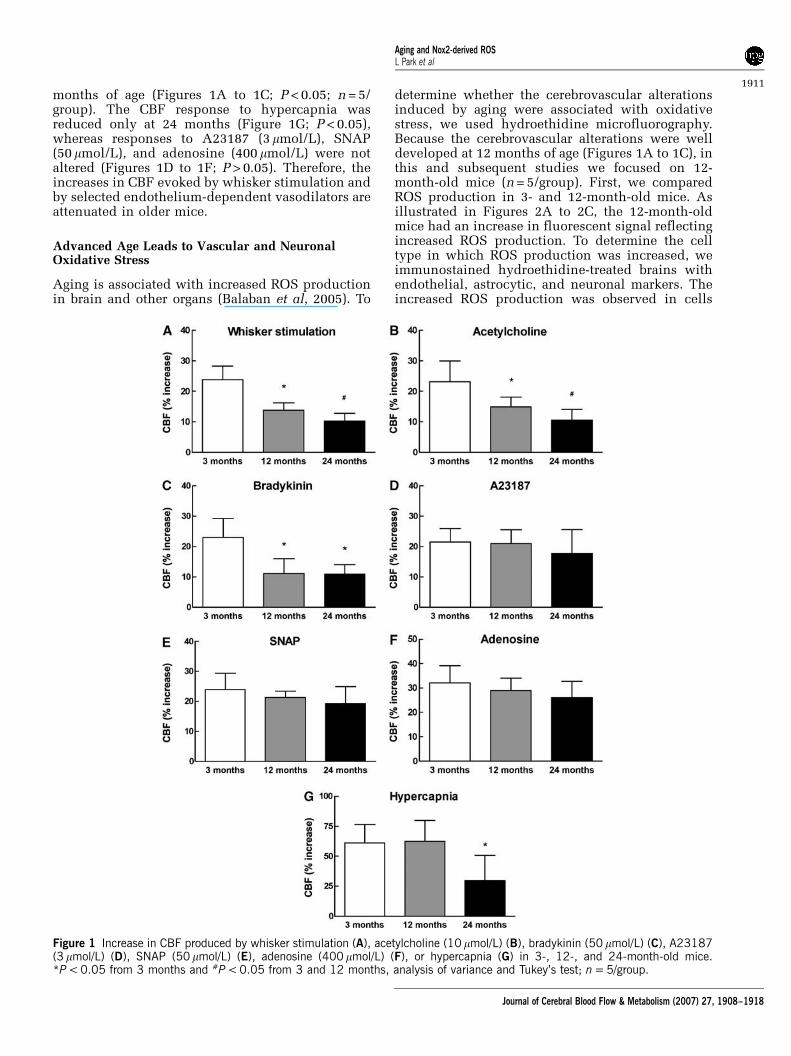
months of age (Figures 1A to 1C; P < 0.05; n = 5/group). The CBF response to hypercapnia wasreduced only at 24 months (Figure 1G; P < 0.05),whereas responses to A23187 (3 mmol/L), SNAP(50 mmol/L), and adenosine (400 mmol/L) were notaltered (Figures 1D to 1F; P > 0.05). Therefore, theincreases in CBF evoked by whisker stimulation andby selected endothelium-dependent vasodilators areattenuated in older mice.
Advanced Age Leads to Vascular and NeuronalOxidative Stress
Aging is associated with increased ROS productionin brain and other organs (Balaban et al, 2005). To
determine whether the cerebrovascular alterationsinduced by aging were associated with oxidativestress, we used hydroethidine microfluorography.Because the cerebrovascular alterations were welldeveloped at 12 months of age (Figures 1A to 1C), inthis and subsequent studies we focused on 12-month-old mice (n = 5/group). First, we comparedROS production in 3- and 12-month-old mice. Asillustrated in Figures 2A to 2C, the 12-month-oldmice had an increase in fluorescent signal reflectingincreased ROS production. To determine the celltype in which ROS production was increased, weimmunostained hydroethidine-treated brains withendothelial, astrocytic, and neuronal markers. Theincreased ROS production was observed in cells
Figure 1 Increase in CBF produced by whisker stimulation (A), acetylcholine (10 mmol/L) (B), bradykinin (50 mmol/L) (C), A23187(3 mmol/L) (D), SNAP (50mmol/L) (E), adenosine (400 mmol/L) (F), or hypercapnia (G) in 3-, 12-, and 24-month-old mice.*P < 0.05 from 3 months and #P < 0.05 from 3 and 12 months, analysis of variance and Tukey’s test; n = 5/group.
Aging and Nox2-derived ROSL Park et al
1911
Journal of Cerebral Blood Flow & Metabolism (2007) 27, 1908–1918
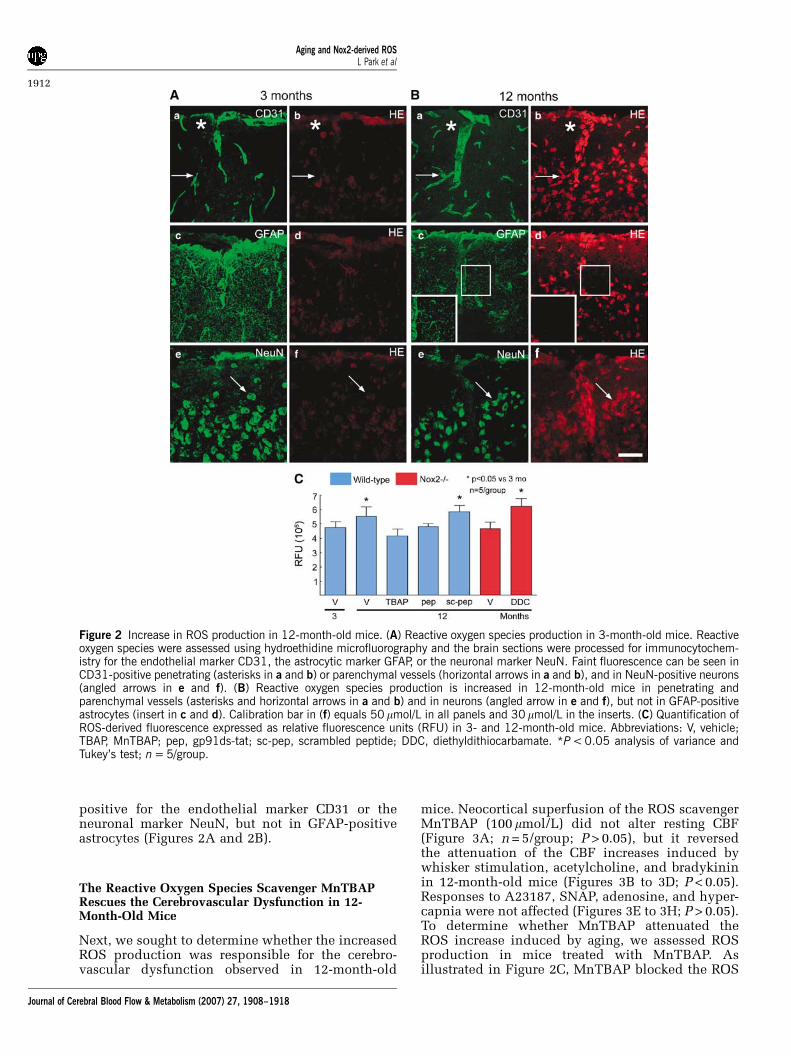
positive for the endothelial marker CD31 or theneuronal marker NeuN, but not in GFAP-positiveastrocytes (Figures 2A and 2B).
The Reactive Oxygen Species Scavenger MnTBAPRescues the Cerebrovascular Dysfunction in 12-Month-Old Mice
Next, we sought to determine whether the increasedROS production was responsible for the cerebro-vascular dysfunction observed in 12-month-old
mice. Neocortical superfusion of the ROS scavengerMnTBAP (100 mmol/L) did not alter resting CBF(Figure 3A; n = 5/group; P > 0.05), but it reversedthe attenuation of the CBF increases induced bywhisker stimulation, acetylcholine, and bradykininin 12-month-old mice (Figures 3B to 3D; P < 0.05).Responses to A23187, SNAP, adenosine, and hyper-capnia were not affected (Figures 3E to 3H; P > 0.05).To determine whether MnTBAP attenuated theROS increase induced by aging, we assessed ROSproduction in mice treated with MnTBAP. Asillustrated in Figure 2C, MnTBAP blocked the ROS
Figure 2 Increase in ROS production in 12-month-old mice. (A) Reactive oxygen species production in 3-month-old mice. Reactiveoxygen species were assessed using hydroethidine microfluorography and the brain sections were processed for immunocytochem-istry for the endothelial marker CD31, the astrocytic marker GFAP, or the neuronal marker NeuN. Faint fluorescence can be seen inCD31-positive penetrating (asterisks in a and b) or parenchymal vessels (horizontal arrows in a and b), and in NeuN-positive neurons(angled arrows in e and f). (B) Reactive oxygen species production is increased in 12-month-old mice in penetrating andparenchymal vessels (asterisks and horizontal arrows in a and b) and in neurons (angled arrow in e and f), but not in GFAP-positiveastrocytes (insert in c and d). Calibration bar in (f) equals 50 mmol/L in all panels and 30 mmol/L in the inserts. (C) Quantification ofROS-derived fluorescence expressed as relative fluorescence units (RFU) in 3- and 12-month-old mice. Abbreviations: V, vehicle;TBAP, MnTBAP; pep, gp91ds-tat; sc-pep, scrambled peptide; DDC, diethyldithiocarbamate. *P < 0.05 analysis of variance andTukey’s test; n = 5/group.
Aging and Nox2-derived ROSL Park et al
1912
Journal of Cerebral Blood Flow & Metabolism (2007) 27, 1908–1918
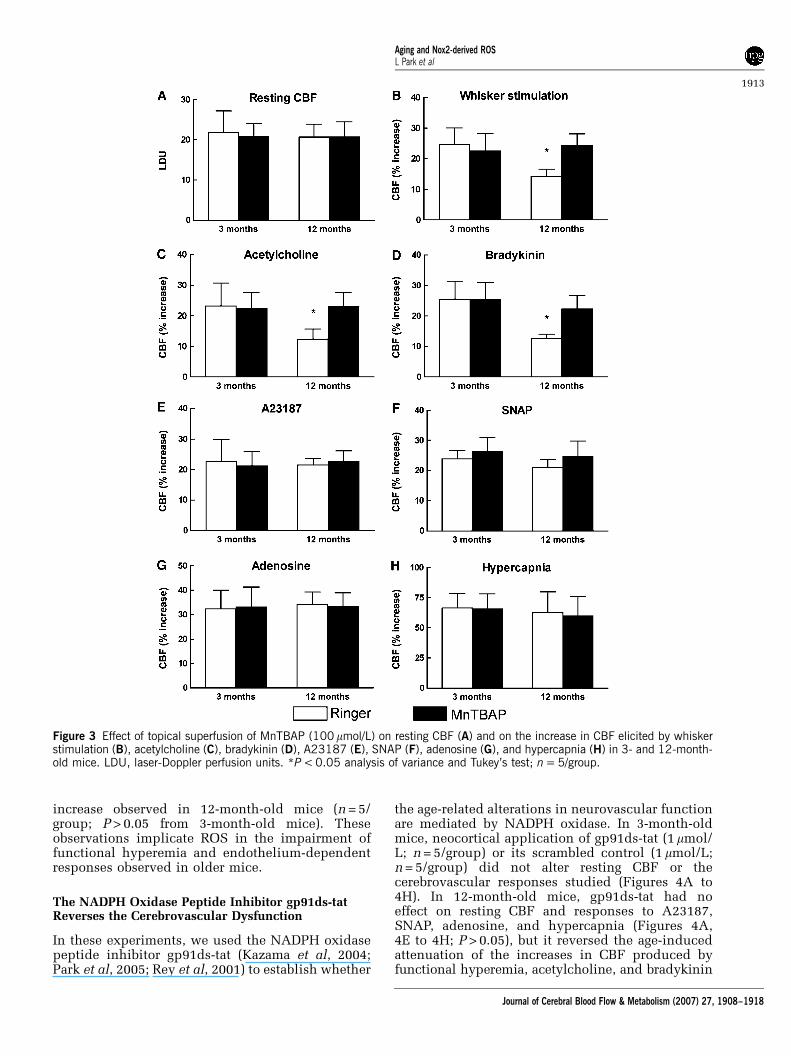
increase observed in 12-month-old mice (n = 5/group; P > 0.05 from 3-month-old mice). Theseobservations implicate ROS in the impairment offunctional hyperemia and endothelium-dependentresponses observed in older mice.
The NADPH Oxidase Peptide Inhibitor gp91ds-tatReverses the Cerebrovascular Dysfunction
In these experiments, we used the NADPH oxidasepeptide inhibitor gp91ds-tat (Kazama et al, 2004;Park et al, 2005; Rey et al, 2001) to establish whether
the age-related alterations in neurovascular functionare mediated by NADPH oxidase. In 3-month-oldmice, neocortical application of gp91ds-tat (1 mmol/L; n = 5/group) or its scrambled control (1 mmol/L;n = 5/group) did not alter resting CBF or thecerebrovascular responses studied (Figures 4A to4H). In 12-month-old mice, gp91ds-tat had noeffect on resting CBF and responses to A23187,SNAP, adenosine, and hypercapnia (Figures 4A,4E to 4H; P > 0.05), but it reversed the age-inducedattenuation of the increases in CBF produced byfunctional hyperemia, acetylcholine, and bradykinin
Figure 3 Effect of topical superfusion of MnTBAP (100 mmol/L) on resting CBF (A) and on the increase in CBF elicited by whiskerstimulation (B), acetylcholine (C), bradykinin (D), A23187 (E), SNAP (F), adenosine (G), and hypercapnia (H) in 3- and 12-month-old mice. LDU, laser-Doppler perfusion units. *P < 0.05 analysis of variance and Tukey’s test; n = 5/group.
Aging and Nox2-derived ROSL Park et al
1913
Journal of Cerebral Blood Flow & Metabolism (2007) 27, 1908–1918
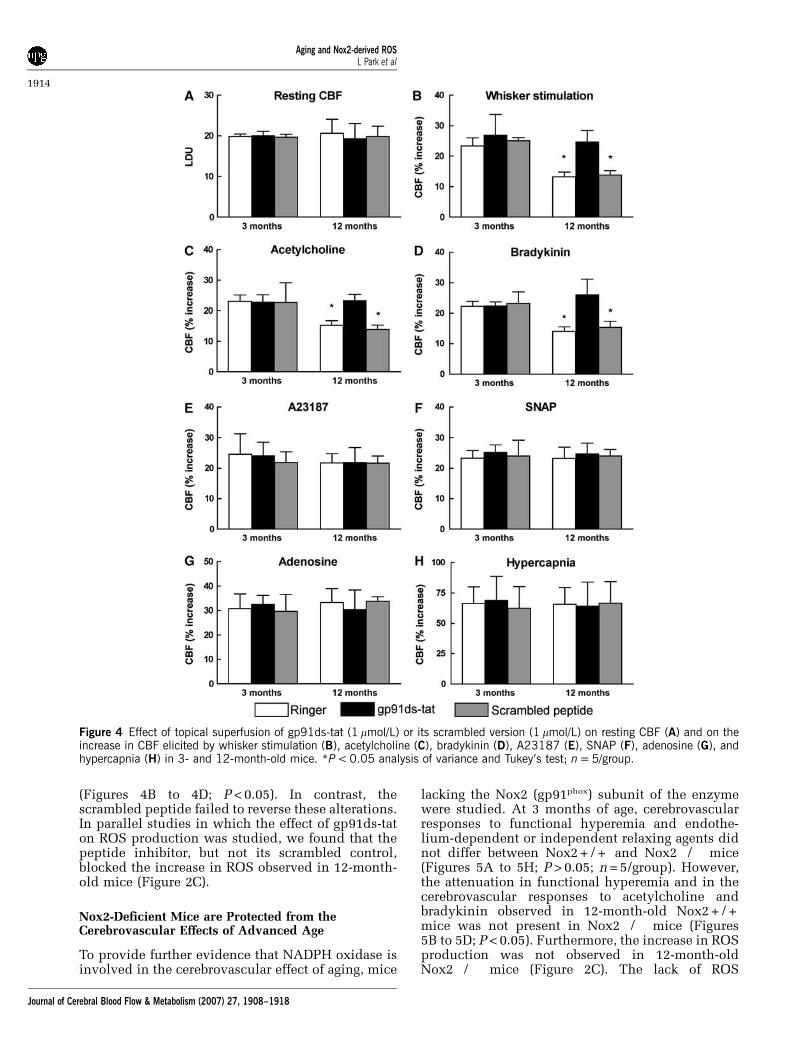
(Figures 4B to 4D; P < 0.05). In contrast, thescrambled peptide failed to reverse these alterations.In parallel studies in which the effect of gp91ds-taton ROS production was studied, we found that thepeptide inhibitor, but not its scrambled control,blocked the increase in ROS observed in 12-month-old mice (Figure 2C).
Nox2-Deficient Mice are Protected from theCerebrovascular Effects of Advanced Age
To provide further evidence that NADPH oxidase isinvolved in the cerebrovascular effect of aging, mice
lacking the Nox2 (gp91phox) subunit of the enzymewere studied. At 3 months of age, cerebrovascularresponses to functional hyperemia and endothe-lium-dependent or independent relaxing agents didnot differ between Nox2 + / + and Nox2�/� mice(Figures 5A to 5H; P > 0.05; n = 5/group). However,the attenuation in functional hyperemia and in thecerebrovascular responses to acetylcholine andbradykinin observed in 12-month-old Nox2 + / +mice was not present in Nox2�/� mice (Figures5B to 5D; P < 0.05). Furthermore, the increase in ROSproduction was not observed in 12-month-oldNox2�/� mice (Figure 2C). The lack of ROS
Figure 4 Effect of topical superfusion of gp91ds-tat (1 mmol/L) or its scrambled version (1 mmol/L) on resting CBF (A) and on theincrease in CBF elicited by whisker stimulation (B), acetylcholine (C), bradykinin (D), A23187 (E), SNAP (F), adenosine (G), andhypercapnia (H) in 3- and 12-month-old mice. *P < 0.05 analysis of variance and Tukey’s test; n = 5/group.
Aging and Nox2-derived ROSL Park et al
1914
Journal of Cerebral Blood Flow & Metabolism (2007) 27, 1908–1918
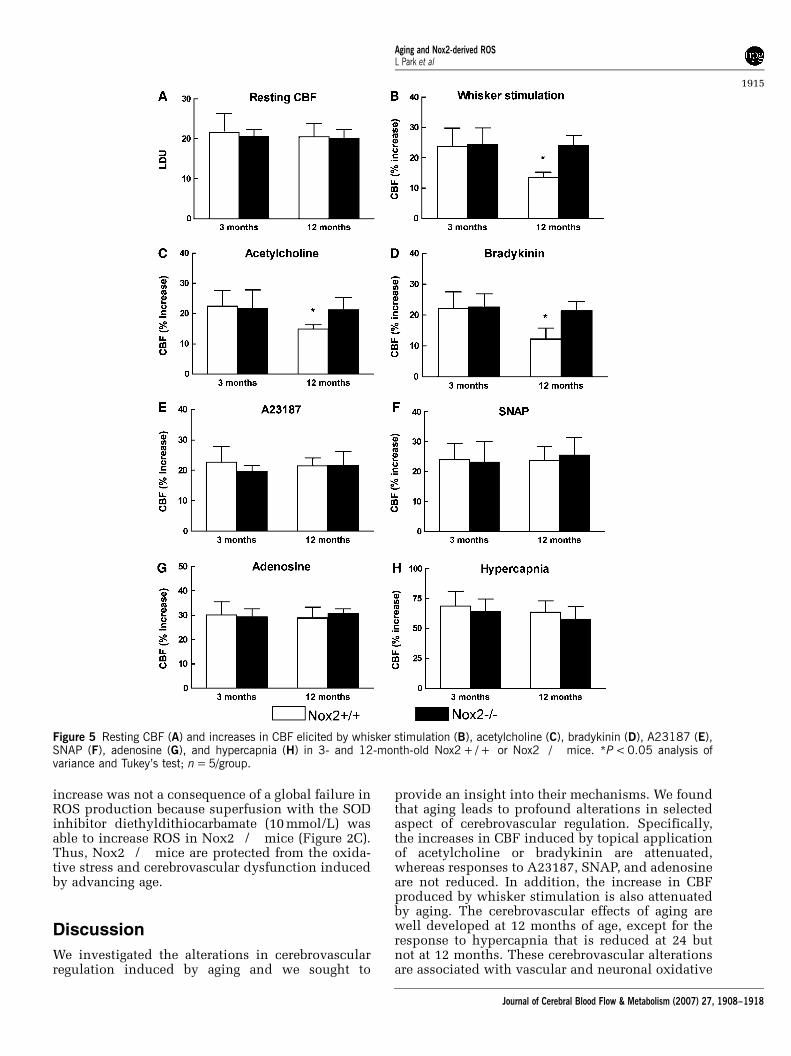
increase was not a consequence of a global failure inROS production because superfusion with the SODinhibitor diethyldithiocarbamate (10 mmol/L) wasable to increase ROS in Nox2�/� mice (Figure 2C).Thus, Nox2�/� mice are protected from the oxida-tive stress and cerebrovascular dysfunction inducedby advancing age.
Discussion
We investigated the alterations in cerebrovascularregulation induced by aging and we sought to
provide an insight into their mechanisms. We foundthat aging leads to profound alterations in selectedaspect of cerebrovascular regulation. Specifically,the increases in CBF induced by topical applicationof acetylcholine or bradykinin are attenuated,whereas responses to A23187, SNAP, and adenosineare not reduced. In addition, the increase in CBFproduced by whisker stimulation is also attenuatedby aging. The cerebrovascular effects of aging arewell developed at 12 months of age, except for theresponse to hypercapnia that is reduced at 24 butnot at 12 months. These cerebrovascular alterationsare associated with vascular and neuronal oxidative
Figure 5 Resting CBF (A) and increases in CBF elicited by whisker stimulation (B), acetylcholine (C), bradykinin (D), A23187 (E),SNAP (F), adenosine (G), and hypercapnia (H) in 3- and 12-month-old Nox2 + / + or Nox2�/� mice. *P < 0.05 analysis ofvariance and Tukey’s test; n = 5/group.
Aging and Nox2-derived ROSL Park et al
1915
Journal of Cerebral Blood Flow & Metabolism (2007) 27, 1908–1918

stress and are reversed by a ROS scavenger. The age-induced alterations in vascular reactivity are abro-gated by a peptide inhibitor of NADPH oxidase, amajor source of ROS in vessels and neurons, and areabsent in mice lacking the Nox2 subunit of NADPHoxidase. These observations provide the first demon-stration that aging impairs not only endothelium-dependent relaxation but also neurovascularcoupling, and that these cerebrovascular alterationsare mediated by Nox2-derived ROS.
The findings of this study cannot be attributed todifferences in arterial pressure or blood gases,factors that have profound effects on CBF (Faraciand Heistad, 1998), because these variables werecarefully controlled and did not differ among thegroups of mice studied. Furthermore, the observa-tions of this study cannot be attributed to age-relateddifferences in the sensitivity to anesthesia leadingto a generalized depression in cerebrovascularreactivity, because only selected CBF responses areattenuated in older mice. Therefore, the reduction incerebrovascular reactivity observed in older mice isnot a result of instability of the preparation orartifacts resulting from anesthesia.
Cerebral blood flow responses to whisker stimula-tion and acetylcholine were attenuated more in 24-than in 12-month-old mice, whereas the response tobradykinin was attenuated equally at 12 and 24months. Conversly, the CBF response to hypercapniawas attenuated only in 24-month-old mice andresponses to A23187, SNAP, and adenosine werenot reduced even at 24 months of age. Thesedifferences in the temporal profile of the attenuationsuggest that the susceptibility to the effects of agingis not identical for all vascular responses of thecerebrovascular bed. Interestingly, the temporaldifferences in the attenuation are not related to themechanism of the vasodilation. For example, theincrease in CBF evoked by bradykinin and A23187is mediated by endothelial COX-1 reaction products(Niwa et al, 2001); yet, the attenuation of theresponse to bradykinin is maximal at 12 months ofage, whereas the response to A23187 is not attenu-ated even at 24 months. One possible explanationfor this discrepancy is that the CBF response tobradykinin depends on activation of bradykininreceptors, whereas the response to A23187 isreceptor-independent (Faraci and Heistad, 1998).Therefore, the attenuation of the response tobradykinin could reflect an age-related disruptionof bradykinin receptors, as described in the heart(Kintsurashvili et al, 2005). Similarly, a reduction inmuscarinic receptors could contribute to the at-tenuation of the CBF response to acetylcholine.However, in the case of acetylcholine, a reduction inthe synthesis and/or bioavailability of nitric oxidecould also play a role (van der Loo et al, 2000).
One of the new findings of this study is that agingattenuates the increase in CBF evoked by whiskerstimulation. Functional hyperemia is essential forthe normal workings of the brain and its impairment
leads to brain dysfunction, as observed in modelsof hypertension and Alzheimer’s disease (Kazamaet al, 2004; Niwa et al, 2000). Reduced hyperemiaduring brain activation limits the delivery of energysubstrates and oxygen to the active brain andimpairs the removal of metabolic byproducts. Theimpairment in functional hyperemia occurs inconcert with alterations of other key regulatorymechanisms of the cerebral circulation, such asresponses to endothelium-dependent vasodilators,hypoxia, hypercapnia, and hypotension (Hoffmanet al, 1981, 1984, 1982; Sato et al, 2002). Thesealterations act synergistically to increase the sus-ceptibility of the brain to vascular insufficiency andischemia, and are likely to play a role in theincreased incidence in ischemic stroke and vascularcognitive impairment observed with advancing age(Farkas and Luiten, 2001).
Aging increases the stiffness of cerebral bloodvessels, and leads to thinning and loss of endothe-lial cells as well as tortuosity and narrowing ofarterioles and capillaries (Hajdu et al, 1990; Hutch-ins et al, 1996). Although these morphologicalchanges have been implicated in the cerebrovascularalterations observed with increasing age they areunlikely to play a major role in our model. This isbecause the cerebrovascular responses tested werenot uniformly disrupted. Rather, responses to theendothelium-dependent vasodilator A23187 andresponses mediated by agents that act directly onvascular smooth muscles, such as adenosine andSNAP, were not attenuated. Therefore, our datasuggest that the altered reactivity is mediated bydysfunction of the vessels rather than mechanicalfactors deriving from structural alterations.
Reactive oxygen species have long been impli-cated in the cerebrovascular dysfunction induced byaging (Faraci and Heistad, 1998; Farkas and Luiten,2001), but their cellular and enzymatic sources havenot been defined. We found that 12-month-old miceexhibit an increase in ROS production in thecerebral cortex. Using double-label immunocyto-chemistry with specific cell-type markers, we foundthat the cellular source of the radicals is primarilyneurons and cerebral blood vessels. Surprisingly,increased ROS production was not detected inastrocytes in 12-month-old mice. The reasons forthis finding remain unclear, but astrocytes areendowed with greater antioxidant defenses thanneurons and, consequently, are more resistant tooxidative stress (Makar et al, 1994; Raps et al, 1989;Savolainen, 1978). However, we cannot rule outthe possibility that astrocytic ROS production isincreased in mice older than 12 months. Indeed,experiments in cultured astrocytes indicate thatwith increasing age the astrocytic antioxidantdefenses become insufficient, leading to develop-ment of oxidative stress (Papadopoulos et al, 1998).
The cerebrovascular alterations observed in12-month-old mice were completely reversed bythe ROS scavenger MnTBAP. This finding is in
Aging and Nox2-derived ROSL Park et al
1916
Journal of Cerebral Blood Flow & Metabolism (2007) 27, 1908–1918

agreement with reports by others (Brown et al, 2006;Didion et al, 2006) and indicates that oxidativestress is involved in the attenuation of cerebro-vascular responses induced by aging. We then investi-gated the enzymatic source of ROS focusing onNADPH oxidase, a superoxide-generating enzymethat has been implicated in the cerebrovasculardysfunction induced by other conditions associatedwith oxidative stress (Fang et al, 2006; Kazama et al,2004; Miller et al, 2006; Park et al, 2005). We foundthat inhibition of NADPH oxidase attenuates ROSproduction and normalizes the cerebrovascularalterations observed in 12-month-old mice. Further-more, mice lacking Nox2 were protected from theoxidative stress and the resulting alterations incerebrovascular function. These findings provideevidence that Nox2-containing NADPH oxidase isthe source of the ROS mediating the cerebrovasculardysfunction observed in 12-month-old mice.
Recent studies have examined the role of NADPHoxidase in the oxidative stress observed in the heartand systemic blood vessels during aging. However,these studies have been largely inconclusive. Somereports have described an increase in NADPHoxidase subunits in systemic blood vessels (Adleret al, 2003; Hamilton et al, 2001; Oudot et al, 2006),whereas others have not (Bachschmid et al, 2004;Csiszar et al, 2002; Newaz et al, 2006). Other studieshave used non-specific pharmacological inhibitors,such as DPI or apocyanin, to assess the contributionof NADPH oxidase to ROS production and/orcardiovascular dysfunction (Adler et al, 2003; Csiszaret al, 2002; Hamilton et al, 2001). In this study, usingboth a peptide inhibitor and Nox2-null mice wehave been able to provide, for the first time, converg-ing evidence that a Nox2-containing NADPH oxidaseis a key factor in the cerebrovascular dysfunctionproduced by aging. However, we cannot rule out thatNADPH oxidase-derived ROS serve to ‘prime’ oxida-tive stress by activating other ROS-generatingsystems, such as mitochondrial enzymes or xanthineoxidase (Brandes, 2005; Newaz et al, 2006).
In conclusion, we have showed that aging, inaddition to its well-established effects on endothe-lium-dependent vasodilation, induces profoundchanges in neurovascular coupling. These alterationsare mediated by ROS derived from a Nox2-containingNADPH oxidase in neurons and cerebral bloodvessels. The data identify, for the first time, Nox2as a key source of ROS mediating the alterationsin vascular reactivity induced by advancing age.Modulation of Nox2-derived ROS may provide a newstrategy to counteract the deleterious consequencesof the cerebrovascular dysfunction induced by aging.
Acknowledgements
CI is the recipient of a Javits Award from NIH/NINDS. We thank Dr Carrie T Drake for her helpfulcomments.
References
Abramov AY, Jacobson J, Wientjes F, Hothersall J, CanevariL, Duchen MR (2005) Expression and modulation of anNADPH oxidase in mammalian astrocytes. J Neurosci25:9176–84
Adler A, Messina E, Sherman B, Wang Z, Huang H, LinkeA, Hintze TH (2003) NAD(P)H oxidase-generatedsuperoxide anion accounts for reduced control ofmyocardial O2 consumption by NO in old Fischer344 rats. Am J Physiol Heart Circ Physiol 285:H1015–22
Bachschmid M, van der Loo B, Schuler K, Labugger R,Thurau S, Eto M, Kilo J, Holz R, Luscher TF, Ullrich V(2004) Oxidative stress-associated vascular aging isindependent of the protein kinase C/NAD(P)H oxidasepathway. Arch Gerontol Geriatr 38:181–90
Balaban RS, Nemoto S, Finkel T (2005) Mitochondria,oxidants, and aging. Cell 120:483–95
Bedard K, Krause K (2007) The NOX Family of ROS-generating NADPH oxidases: physiology and patho-physiology. Physiol Rev 87:245–313
Brandes RP (2005) Triggering mitochondrial radicalrelease: a new function for NADPH oxidases. Hyperten-sion 45:847–8
Brown KA, Chu Y, Lund DD, Heistad DD, Faraci FM (2006)Gene transfer of extracellular superoxide dismutaseprotects against vascular dysfunction with aging. Am JPhysiol Heart Circ Physiol 290:H2600–5
Csiszar A, Ungvari Z, Edwards JG, Kaminski P, Wolin MS,Koller A, Kaley G (2002) Aging-induced phenotypicchanges and oxidative stress impair coronary arteriolarfunction. Circ Res 90:1159–66
Didion SP, Kinzenbaw DA, Schrader LI, Faraci FM (2006)Heterozygous CuZn superoxide dismutase deficiencyproduces a vascular phenotype with aging. Hyper-tension 48:1072–9
Fang Q, Sun H, Arrick DM, Mayhan WG (2006) Inhibitionof NADPH oxidase improves impaired reactivity of pialarterioles during chronic exposure to nicotine. J ApplPhysiol 100:631–6
Faraci FM, Heistad DD (1998) Regulation of the cerebralcirculation: role of endothelium and potassiumchannels. Physiol Rev 78:53–97
Faraci FM (2006) Reactive oxygen species: influence oncerebral vascular tone. J Appl Physiol 100:739–43
Farkas E, Luiten PG (2001) Cerebral microvascularpathologyin aging and Alzheimer’s disease. ProgNeurobiol 64:575–611
Hajdu MA, Heistad DD, Siems JE, Baumbach GL (1990)Effects of aging on mechanics and composition ofcerebral arterioles in rats. Circ Res 66:1747–54
Hamilton CA, Brosnan MJ, McIntyre M, Graham D,Dominiczak AF (2001) Superoxide excess in hyper-tension and aging: a common cause of endothelialdysfunction. Hypertension 37:529–34
Hoffman WE, Albrecht RF, Miletich DJ (1981) Theinfluence of aging and hypertension on cerebralautoregulation. Brain Res 214:196–9
Hoffman WE, Albrecht RF, Miletich DJ (1984) The role ofadenosine in CBF increases during hypoxia in youngversus aged rats. Stroke 15:124–9
Hoffman WE, Miletich DJ, Albrecht RF (1982) Cerebro-vascular and cerebral metabolic responses of aged ratsto changes in arterial PCO2. Neurobiol Aging 3:141–3
Hutchins PM, Lynch CD, Cooney PT, Curseen KA (1996)The micro circulation in experimental hypertensionand aging. Cardiovasc Res 32:772–80
Aging and Nox2-derived ROSL Park et al
1917
Journal of Cerebral Blood Flow & Metabolism (2007) 27, 1908–1918

Iadecola C (1992) Nitric oxide participates in the cerebrovasodilation elicited from cerebellar fastigial nucleus.Am J Physiol 263:R1156–61
Iadecola C (2004) Neurovascular regulation in the normalbrain and in Alzheimer’s disease. Nat Rev Neurosci5:347–60
Infanger DW, Sharma RV, Davisson RL (2006) NADPHoxidases of the brain: distribution, regulation, andfunction. Antioxid Redox Signal 8:1583–96
Kazama K, Anrather J, Zhou P, Girouard H, Frys K, MilnerTA, Iadecola C (2004) Angiotensin II impairs neurovas-cular coupling in neocortex through NADPH oxidase-derived radicals. Circ Res 95:1019–26
Kintsurashvili E, Duka A, Ignjacev I, Pattakos G, Gavras I,Gavras H (2005) Age-related changes of bradykinin B1and B2 receptors in rat heart. Am J Physiol Heart CircPhysiol 289:H202–5
Makar TK, Nedergaard M, Preuss A, Gelbard , Perumal ,Cooper AJ (1994) Vitamin E, ascorbate, glutathione,glutathione disulfide, and enzymes of glutathionemetabolism in cultures of chick astrocytes and neurons:evidence that astrocytes play an important role inantioxidative processes in the brain. J Neurochem62:45–53
Mayhan WG, Arrick DM, Sharpe GM, Patel KP, Sun H(2006) Inhibition of NAD(P)H oxidase alleviates im-paired NOS-dependent responses of pial arterioles intype 1 diabetes mellitus. Microcirculation 13:567–75
Mayhan WG, Faraci FM, Baumbach GL, Heistad DD (1990)Effects of aging on responses of cerebral arterioles. Am JPhysiol 258:H1138–43
Miller AA, Drummond GR, Sobey CG (2006) Novelisoforms of NADPH-oxidase in cerebral vascular con-trol. Pharmacol Ther 111:928–48
Newaz MA, Yousefipour Z, Oyekan A (2006) Oxidativestress-associated vascular aging is xanthine oxidase-dependent but not NAD(P)H oxidase-dependent. JCardiovasc Pharmacol 48:88–94
Niwa K, Haensel C, Ross ME, Iadecola C (2001) Cyclo-oxygenase-1 participates in selected vasodilatorresponses of the cerebral circulation. Circ Res 88:600–8
Niwa K, Younkin L, Ebeling C, Turner SK, Westaway D,Younkin S, Ashe KH, Carlson GA, Iadecola C (2000)Abeta 1 to 40-related reduction in functional hyper-emia in mouse neocortex during somatosensory activa-tion. Proc Natl Acad Sci USA 97:9735–40
Oudot A, Martin C, Busseuil D, Vergely C, Demaison L,Rochette L (2006) NADPH oxidases are in part
responsible for increased cardiovascular superoxideproduction during aging. Free Radic Biol Med 40:2214–2222
Papadopoulos MC, Koumenis IL, Yuan TY, Giffard RG(1998) Increasing vulnerability of astrocytes to oxida-tive injury with age despite constant antioxidantdefenses. Neuroscience 82:915–25
Park L, Anrather J, Zhou P, Frys K, Pitstick R, Younkin S,Carlson GA, Iadecola C (2005) NADPH-oxidase-derivedreactive oxygen species mediate the cerebrovasculardysfunction induced by the amyloid beta peptide.J Neurosci 25:1769–77
Paterno R, Faraci FM, Heistad DD (1994) Age-relatedchanges in release of endothelium-derived relaxingfactor from the carotid artery. Stroke 25:2457–60;discussion 2461–2452
Pollock JD, Williams DA, Gifford MA, Li LL, Du X,Fisherman J, Orkin SH, Doerschuk CM, Dinauer MC(1995) Mouse model of X-linked chronic granuloma-tous disease, an inherited defect in phagocyte super-oxide production. Nat Genet 9:202–9
Raps SP, Lai JC, Hertz L, Cooper AJ (1989) Glutathione ispresent in high concentrations in cultured astrocytesbut not in cultured neurons. Brain Res 493:398–401
Rey FE, Cifuentes ME, Kiarash A, Quinn MT, Pagano PJ(2001) Novel competitive inhibitor of NAD(P)H oxidaseassembly attenuates vascular O(2)(-) and systolic bloodpressure in mice. Circ Res 89:408–14
Rothwell PM, Coull AJ, Silver LE, Fairhead JF, Giles MF,Lovelock CE, Redgrave JN, Bull LM, Welch SJ,Cuthbertson FC, Binney LE, Gutnikov SA, Anslow P,Banning AP, Mant D, Mehta Z (2005) Population-basedstudy of event-rate, incidence, case fatality, andmortality for all acute vascular events in all arterialterritories (Oxford Vascular Study). Lancet 366:1773–1783
Sato A, Sato Y, Uchida S (2002) Regulation of cerebralcortical blood flow by the basal forebrain cholinergicfibers and aging. Auton Neurosci 96:13–9
Savolainen H (1978) Superoxide dismutase and gluta-thione peroxidase activities in rat brain. Res CommunChem Pathol Pharmacol 21:173–6
van der Loo B, Labugger R, Skepper JN, Bachschmid M,Kilo J, Powell JM, Palacios-Callender M, ErusalimskyJD, Quaschning T, Malinski T, Gygi D, Ullrich V,Luscher TF (2000) Enhanced peroxynitrite formationis associated with vascular aging. J Exp Med 192:1731–1744
Supplementary Information accompanies the paper on the Journal of Cerebral Blood Flow & Metabolism website (http://www.nature.com/jcbfm)
Aging and Nox2-derived ROSL Park et al
1918
Journal of Cerebral Blood Flow & Metabolism (2007) 27, 1908–1918





























