Embed Size (px)
Citation preview

Microbiol Immunol 2010; 54: 1–10doi:10.1111/j.1348-0421.2009.00179.x
ORIGINAL ARTICLE
Non-opsonic phagocytosis of homologous non-toxigenicand toxigenic Corynebacterium diphtheriae strainsby human U-937 macrophagesCıntia Silva dos Santos1,3, Louisy Sanches dos Santos3, Monica Cristina de Souza3,Fernanda dos Santos Dourado3, Alexandre Alves de Souza de Oliveira Dias2, Priscila Soares Sabbadini3,Gabriela Andrade Pereira3, Maulori Curie Cabral1, Raphael Hirata Junior3
and Ana Luıza de Mattos-Guaraldi3
1Institute of Microbiology Professor Paulo de Goes, Federal University of Rio de Janeiro (IMPPG/UFRJ), Centro de Ciencias da Saude, Bloco I, CidadeUniversitaria - Ilha do Fundao, Rio de Janeiro, CEP: 21.941-590, 2National Institute for Quality Control in Health (INCQS/FIOCRUZ), Av. Brasil,4365 - Manguinhos, Rio de Janeiro, CEP: 21040-360 and 3Faculty of Medical Sciences, Rio de Janeiro State University (FCM/UERJ), Av. 28 deSetembro, 87 – Fundos, 3o andar. Vila Isabel, Rio de Janeiro, CEP: 20.551-030, Brazil
ABSTRACTAs interactions between bacteria and macrophages dictate the outcome of most infectious diseases,analyses of molecular mechanisms of non-opsonic phagocytosis should lead to new approaches forthe prevention of diphtheria and systemic Corynebacterium diphtheriae infections. The present studyaimed to evaluate human macrophage–bacteria interactions in the absence of opsonin antibodies and theinfluence of the tox gene on this process. Homologous C. diphtheriae tox+and tox– strains were evaluatedfor adhesion, entering and survival within U-937 human macrophages at different incubation periods.Higher numbers of viable bacteria associated with and internalized by macrophages were demonstratedfor the tox+ strain. However, viable intracellular bacteria were detected at T-24 hr only for the tox– strain.Cytoskeletal inhibitors, cytochalasin E, genistein and colchicine, inhibited intracellular viability of bothstrains at different levels. Bacterial replication was evidenced at T-24 hr in supernatants of monolayersinfected with the tox– strain. Host cell death and nuclear alterations were evidenced by the Trypan blueexclusion assay and DAPI fluorescence microscopy. ELISA of histone-associated DNA fragments alloweddetection of apoptosis and necrosis induced by tox+ and tox– strains at T-1 hr and T-3 hr. In conclusion,human macrophages in the absence of opsonins may not be promptly effective at killing diphtheria bacilli.The presence of the tox gene influences the susceptibility of C. diphtheriae to human macrophages andthe outcome of non-opsonic phagocytosis. C. diphtheriae strains exhibit strategies to survive withinmacrophages and to exert apoptosis and necrosis in human phagocytic cells, independent of the toxgene.
Key words apoptosis, Corynebacterium diphtheriae, cytoskeletal rearrangement, necrosis, non-opsonic phagocytosis,U-937 human macrophage cell line.
CorrespondenceProfessor Dr Ana Luıza de Mattos Guaraldi, Disciplina de Microbiologia e Imunologia, Faculdade de Ciencias Medicas, UERJ, Av. 28 deSetembro, 87 – Fundos, 3o andar. Vila Isabel, RJ, Brazil. CEP: 20.551-030.Tel: +55(021)2587-6380; fax: +55 (021)2587-6476; email: [email protected]
Received 13 March 2009; revised 27 July 2009; 27 August 2009; accepted 6 September 2009.
List of Abbreviations: ATCC, American Type Culture Collection; C. diphtheriae Corynebacterium diphtheriae; CFU, colony forming units; CytE,cytochalasin E; DAPI, 4,6-diamidino-2-phenylindoldihydrochloride; DMSO, dimethylsulfoxide; DT, diphtheria toxin; FITC, fluorescein isothiocyanate;FAS test, FITC-labeled phalloidin assay; MOI, multiplicity of infection; PBS-D, Dulbecco’s phosphate-buffered saline; PKC, protein kinase C; PMA,phorbol 12-myristate 13-acetate; PMN, polymophonuclear leukocyte; S. agalactiae Streptococcus agalactiae; tox gene, diphtheria toxin gene.
c© 2009 The Societies and Blackwell Publishing Asia Pty Ltd 1

C. S. dos Santos et al.
Diphtheria has been the cause of many deadly epidemicsworldwide (1–3). The fact that specific epidemic clonesare responsible for severe outbreaks of diphtheria ac-counting for thousands of deaths (4) makes the argu-ment that Corynebacterium diphtheriae virulence factors,other than toxin, are important for their potential tocause human disease (5). The occurrence of diphtheriaamong immunized persons also points out the impor-tance of other microbial factors as well (6). C. diphtheriaestrains have also been reported to cause systemic diseases(7, 8) independent of toxin production, which make theapparent increase in non-toxigenic biotypes a subject ofconcern (9).
The ability to induce C. diphtheriae uptake by non-phagocytic cells and to migrate across epithelial cell bar-riers may be important in the early stages of respiratoryand/or systemic infections. Investigations have shown dif-ferences in the ability of C. diphtheriae strains to enter andsurvive within respiratory epithelial cells (10, 11). Nev-ertheless, mechanisms allowing C. diphtheriae to reachdeeper tissues need further investigation.
The propensity of some Gram-positive bacteria to causeinvasive infections may be partially related to inappropri-ate opsonization due to the lack of type-specific antibod-ies (12). Non-opsonic phagocytosis mediated by receptorsthat recognize adhesins on microbial surfaces has attractedincreasing interest as a potential host defense mechanismagainst extracellular pathogens and as a means of survivalin the host for intracellular pathogens (13). Infectiousagents enter macrophages via heterogeneous pathwaysand modify vacuolar maturation in a manner that favorstheir survival (14). Important differences in the molec-ular mechanisms underlying phagocytosis are now beingappreciated. These include differences in the cytoskeletalelements that mediate internalization, and differences ininflammatory responses. U-937 human cells have beenrecognized as appropriate model systems for the study ofbacteria-macrophage interaction (15).
Macrophage function can be modulated by recent andchronic infection, environmental exposure, drug therapy,cytokines and gene transfer (16). Bacterial exotoxins, suchas DT, can powerfully activate macrophages. Studies deal-ing with the influence of bacterial exotoxins on the phago-cytic activity of human macrophages in culture showedthat phagocytosis was significantly impaired after pro-longed incubation with DT (17, 18).
As interactions between bacteria and macrophages dic-tate the outcome of infectious diseases, the interaction ofC. diphtheriae with macrophages should also be examined.The aim of the present study was to investigate the abilityof homologous toxigenic and non-toxigenic C. diphtheriaestrains to be phagocytosed by U-937 human macrophagesin the absence of opsonins. The participation of cytoskele-
ton rearrangement in this phenomenon and the type ofcell death occurring in infected macrophages were alsoevaluated.
MATERIALS AND METHODS
Bacterial strains and growth conditions
Non-toxigenic ATCC 27010 (C7 s (–) tox– [NCTC 11397])C. diphtheriae subsp. mitis type strain and the homologoustoxigenic ATCC 27012 (tox+) strain from the AmericanType Culture Collection (ATCC, Manassas, VA) were usedin experiments. Stock cultures in 10% skim milk with 25%glycerol added were maintained at –70◦C and recoveredas required by cultivation in Trypticase Soy Broth (Difco,Brazil). Aliquots (500 μl) of log-phase bacterial cultureswith optical density of 0.2 at λ = 660 nm (MOI of 100)were used in all experiments (10).
Chemicals
All reagents were purchased from Sigma Chemical Co. (StLouis, MO, USA) unless otherwise noted.
Macrophages
The human macrophage-like cell line U-937 was main-tained and differentiated by using PMA as previouslydescribed (19, 20). A stock culture of U-937 cells wasmaintained in a monocyte-like, non-adherent form inDMEM added to 10% heat-inactivated FCS, 100 IU/mlpenicillin G and 25 μg/ml gentamycin. Aliquots of U-937 cells in antibiotic-free medium were supplementedwith 250 ng/ml PMA and plated on 24-well tissue-cultureplates (5.0 × 105 cells/well) and maintained in a humid-ified atmosphere containing 5% CO2 at 37◦C for 48 hr.Bacterial adherence and intracellular viability experimentswere carried out with U-937 monolayers grown to ap-proximately 95% confluence in the presence of serum-freeDMEM. Preliminary experiments showed that microor-ganisms were able to survive and proliferate in DMEMwithout macrophages until 24 hr incubation.
Bacterial adherence and intracellularviability experiments
Bacterial adherence and intracellular viability assays werecarried out as previously described (10, 21, 22). For eachof period of incubation examined (0.5, 1, 2, 3, 4 and 24 hr),supernatants and cell lysates were diluted and plated ontoColumbia Agar Base (Difco) to determine the number ofviable bacteria (CFU/ml).
As maximum interaction occurred at approximately3 hr post-infection of U-937 cells for the non-toxigenicATCC 27010 strain, this incubation period was chosen
2 c© 2009 The Societies and Blackwell Publishing Asia Pty Ltd

C. diphtheriae and U-937 cell interaction
for the following investigations concerning mechanismsof bacterial adherence and internalization within U-937macrophages.
Microscopy assays
The microscopy assays were carried out as previously de-scribed (21).
Trypan blue dye exclusion assay
Host cell monolayers were stained with 0.4% Trypan bluesolution diluted at a ratio of 10:1 in PBS-D and the numberof dead (stained) cells was observed under bright fieldmicroscopy (23, 24). Studies dealing with the influenceof exogenous DT (1:40) on C. diphtheriae and U-937 cellsurvival during non-opsonic phagocytosis were based onmethods previously described (17, 18).
Fluorescent DNA staining
DAPI staining was carried out as previously described(25). Briefly, prior to staining, the cells were fixed with4% paraformaldehyde for 30 min at room temperatureand washed with PBS. DAPI (1 μg/ml) was added to thefixed cells for 30 min, after which they were examined byfluorescence microscopy.
FITC-labeled phalloidin assay
Infected semi-confluent U-937 monolayers (3 hr) wereevaluated by the fluorescent actin staining test as previ-ously described (10, 26).
Treatment of U-937 cells with cytochalasin E,colchicine and genistein
Final concentrations of inhibitors and solvents were usedas follows: 5 μM cytochalasin E (CytE), inhibitor of mi-crofilament polymerization in DMSO; 100 μM colchicine,inhibitor of microtubule polymerization, in 70% ethanol;100 μM genistein, tyrosine kinase inhibitor, in DMSO(10, 27, 28). Monolayers were pre-incubated with each in-hibitor separately at 37◦C for 30 min and infected at a MOIof 100 (∼5 × 106 CFU/ml) as described above. Inhibitionof bacterial uptake was confirmed by the absence of bac-terial growth in a gentamycin exclusion test. All inhibitorswere tested for possible effects on cell and bacterial viabil-ity by the Trypan blue exclusion test and CFU counting,respectively, of treated and untreated samples (24, 29).
Apoptosis and necrosis detection by ELISA
Histone-associated DNA fragments (mono- and oligonu-cleosomes) in the cytoplasms (apoptosis) and in the su-pernatants (necrosis) of infected U-937 macrophages were
determined by the Cell Death Detection ELISA PLUS ELISAkit (Roche Diagnostics, Indianapolis, IN). Briefly, the su-pernatants of infected monolayers (1 hr and 3 hr incu-bation) and the lysates from adherent cells were placedin a streptavidin-coated microplate and incubated witha mixture of anti-histone-biotin and anti-DNA peroxi-dase. After addition of peroxidase substrate (ABTS), theabsorbance was measured at 405 nm and background at490 nm (30).
Statistical analysis
Bacterial plate counts were expressed as the mean ± stan-dard deviation (SD; error bars) from three independentexperiments carried out in triplicate. Data for each exper-iment were analyzed by the Tukey test (one-way ANOVA)with Graph Prism. Differences were considered significantat P < 0.05.
RESULTS
Non-opsonic interaction of C. diphtheriaestrains with U-937 macrophages
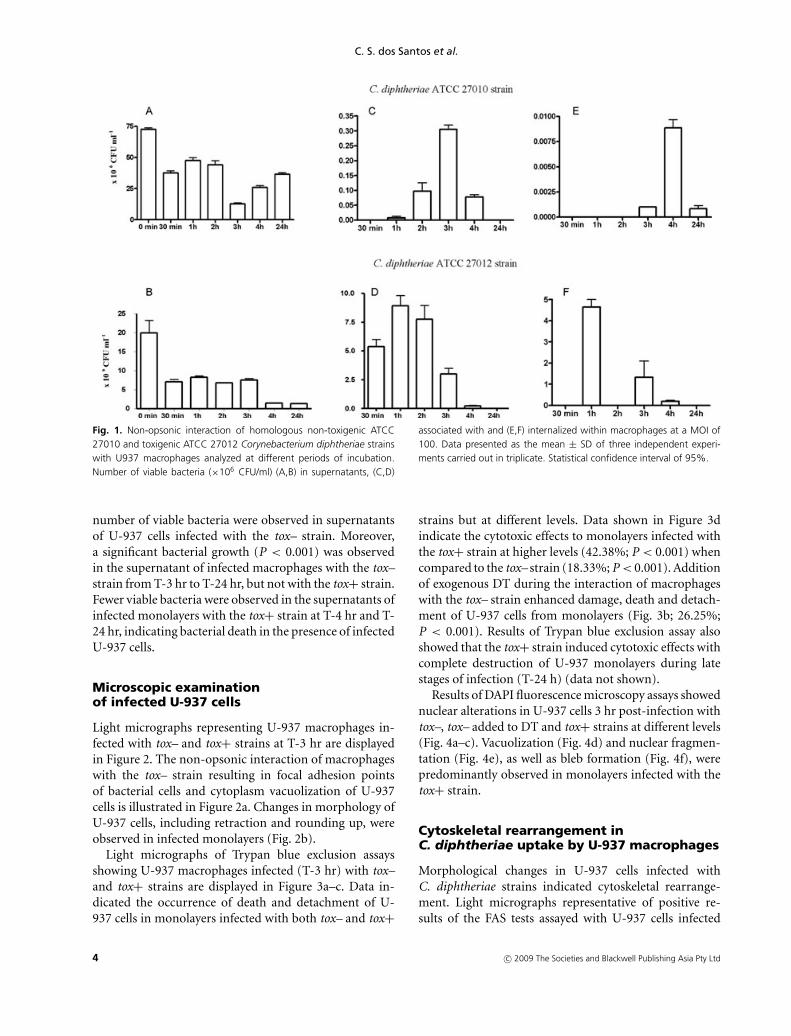
Numbers of viable bacteria associated to and internalizedby U-937 macrophages at specified periods of infection arepresented in Figure 1. The tox– strain showed different lev-els of adherence in all phases of experiments (P < 0.001).A small number of viable associated bacteria were detectedat T-1 hr (0.023 × 106 UFC/ml). The number of adherentbacteria increased significantly in the period from T-1 hrto T-3 hr post-infection (P < 0.001). The highest numberof viable cell-associated bacteria (0.305 × 106 CFU/ml)was observed at T-3 hr (P < 0.001). For the tox+ strain,higher numbers of associated bacteria were observed atT-1 hr to T-2 hr (P > 0.05).
Results of the gentamycin exclusion assays showed theoccurrence of non-opsonic internalization of both tox–and tox+ strains by U-937 cells at different levels. Forthe tox+ strain, the maximum internalization values wereobserved at T-1 hr (4.667 × 106 CFU/ml) and for the tox-strain at T-4 hr (0.009 × 106 CFU/ml).
C. diphtheriae tox- was found to be capable of survivingin the presence of U-937 macrophages for a long periodof time (T-24 hr). The tox– strain also showed the abil-ity to survive in the intracellular environment at T-24 hr(0.001 × 106 CFU/ml).
Viable bacteria in supernatants of U-937monolayers
The results displayed in Figure 1 show that viable bacteriawere observed in supernatants of infected U-937 mono-layers with both strains, but at different levels. A higher
c© 2009 The Societies and Blackwell Publishing Asia Pty Ltd 3
C. S. dos Santos et al.
Fig. 1. Non-opsonic interaction of homologous non-toxigenic ATCC27010 and toxigenic ATCC 27012 Corynebacterium diphtheriae strainswith U937 macrophages analyzed at different periods of incubation.Number of viable bacteria (×106 CFU/ml) (A,B) in supernatants, (C,D)
associated with and (E,F) internalized within macrophages at a MOI of100. Data presented as the mean ± SD of three independent experi-ments carried out in triplicate. Statistical confidence interval of 95%.
number of viable bacteria were observed in supernatantsof U-937 cells infected with the tox– strain. Moreover,a significant bacterial growth (P < 0.001) was observedin the supernatant of infected macrophages with the tox–strain from T-3 hr to T-24 hr, but not with the tox+ strain.Fewer viable bacteria were observed in the supernatants ofinfected monolayers with the tox+ strain at T-4 hr and T-24 hr, indicating bacterial death in the presence of infectedU-937 cells.
Microscopic examinationof infected U-937 cells
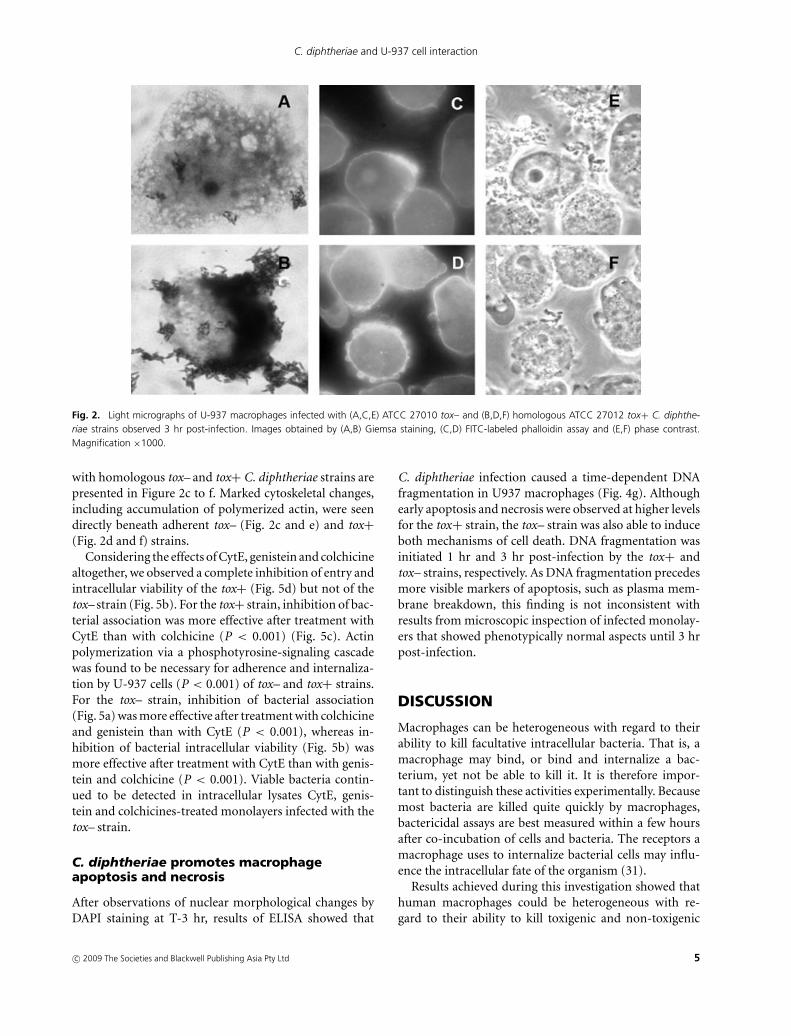
Light micrographs representing U-937 macrophages in-fected with tox– and tox+ strains at T-3 hr are displayedin Figure 2. The non-opsonic interaction of macrophageswith the tox– strain resulting in focal adhesion pointsof bacterial cells and cytoplasm vacuolization of U-937cells is illustrated in Figure 2a. Changes in morphology ofU-937 cells, including retraction and rounding up, wereobserved in infected monolayers (Fig. 2b).
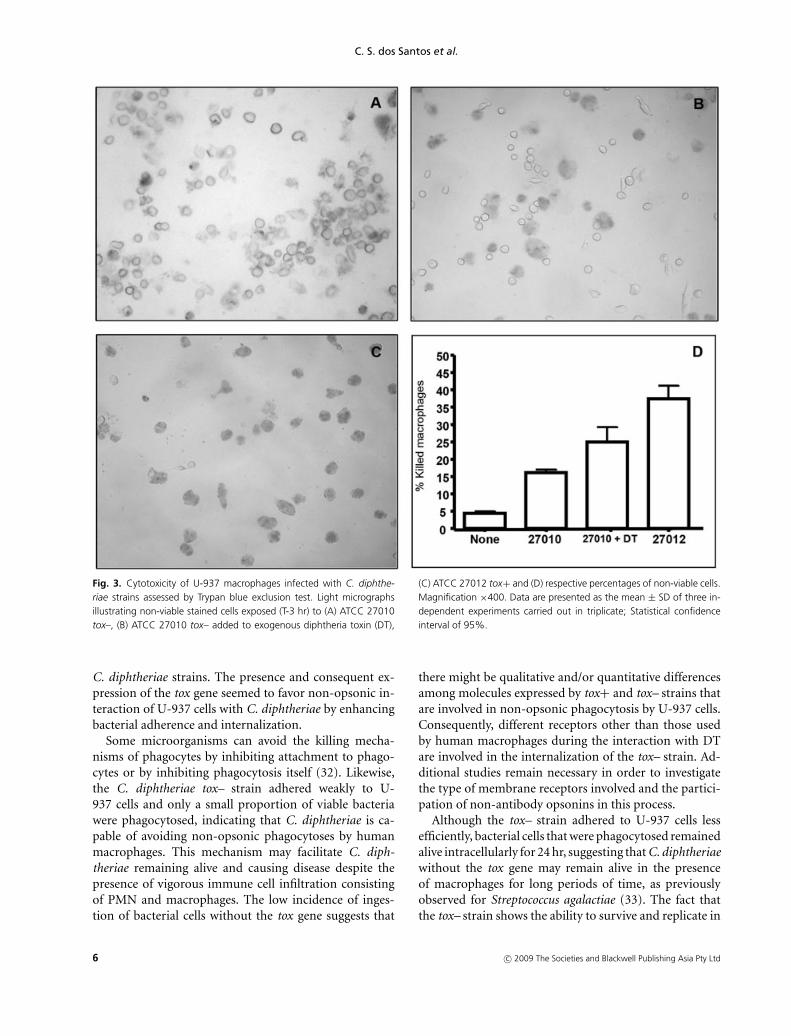
Light micrographs of Trypan blue exclusion assaysshowing U-937 macrophages infected (T-3 hr) with tox–and tox+ strains are displayed in Figure 3a–c. Data in-dicated the occurrence of death and detachment of U-937 cells in monolayers infected with both tox– and tox+
strains but at different levels. Data shown in Figure 3dindicate the cytotoxic effects to monolayers infected withthe tox+ strain at higher levels (42.38%; P < 0.001) whencompared to the tox– strain (18.33%; P < 0.001). Additionof exogenous DT during the interaction of macrophageswith the tox– strain enhanced damage, death and detach-ment of U-937 cells from monolayers (Fig. 3b; 26.25%;P < 0.001). Results of Trypan blue exclusion assay alsoshowed that the tox+ strain induced cytotoxic effects withcomplete destruction of U-937 monolayers during latestages of infection (T-24 h) (data not shown).
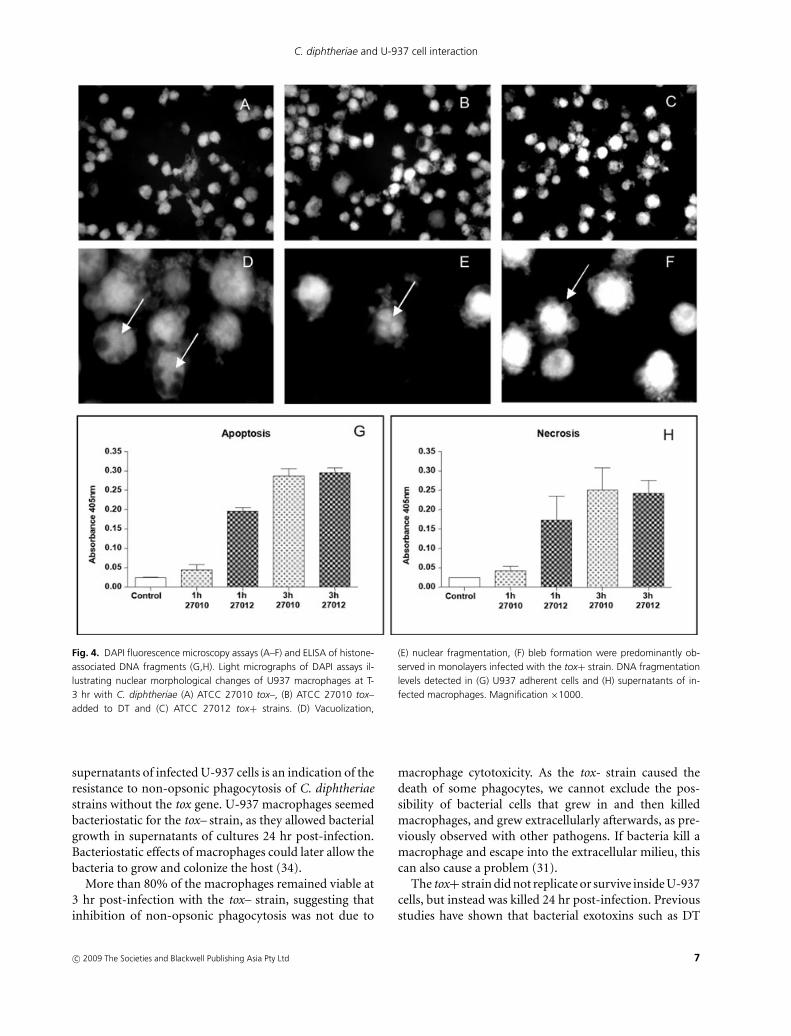
Results of DAPI fluorescence microscopy assays showednuclear alterations in U-937 cells 3 hr post-infection withtox–, tox– added to DT and tox+ strains at different levels(Fig. 4a–c). Vacuolization (Fig. 4d) and nuclear fragmen-tation (Fig. 4e), as well as bleb formation (Fig. 4f), werepredominantly observed in monolayers infected with thetox+ strain.
Cytoskeletal rearrangement inC. diphtheriae uptake by U-937 macrophages
Morphological changes in U-937 cells infected withC. diphtheriae strains indicated cytoskeletal rearrange-ment. Light micrographs representative of positive re-sults of the FAS tests assayed with U-937 cells infected
4 c© 2009 The Societies and Blackwell Publishing Asia Pty Ltd
C. diphtheriae and U-937 cell interaction
Fig. 2. Light micrographs of U-937 macrophages infected with (A,C,E) ATCC 27010 tox– and (B,D,F) homologous ATCC 27012 tox+ C. diphthe-riae strains observed 3 hr post-infection. Images obtained by (A,B) Giemsa staining, (C,D) FITC-labeled phalloidin assay and (E,F) phase contrast.Magnification ×1000.
with homologous tox– and tox+ C. diphtheriae strains arepresented in Figure 2c to f. Marked cytoskeletal changes,including accumulation of polymerized actin, were seendirectly beneath adherent tox– (Fig. 2c and e) and tox+(Fig. 2d and f) strains.
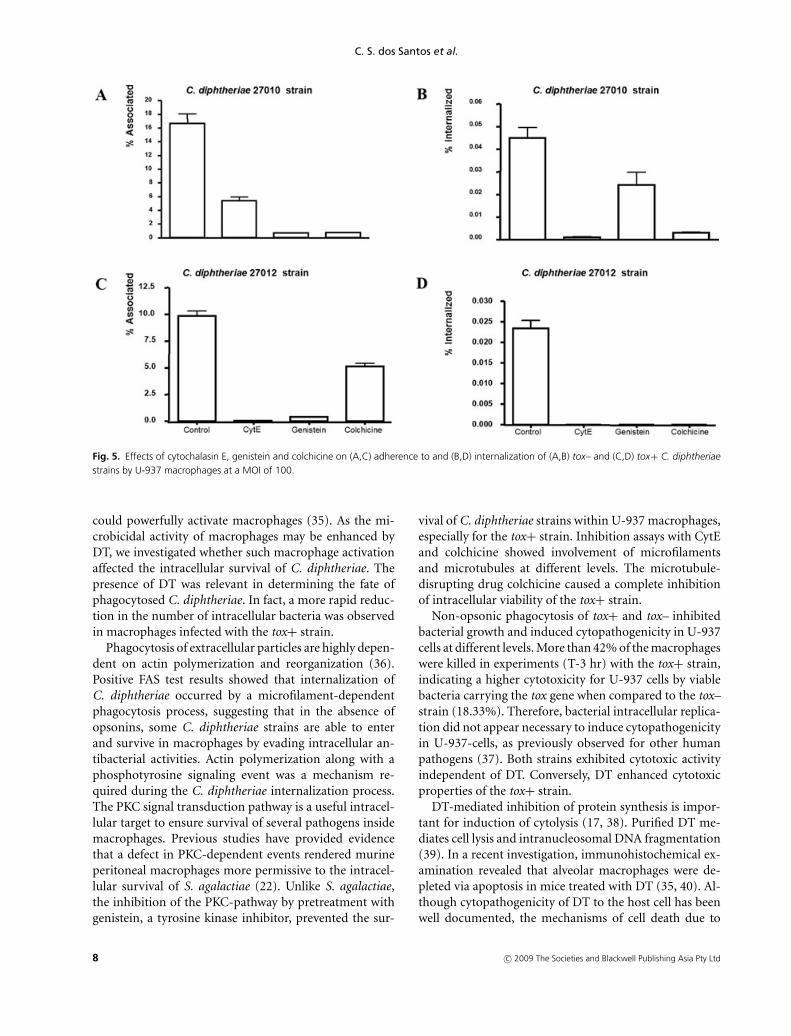
Considering the effects of CytE, genistein and colchicinealtogether, we observed a complete inhibition of entry andintracellular viability of the tox+ (Fig. 5d) but not of thetox– strain (Fig. 5b). For the tox+ strain, inhibition of bac-terial association was more effective after treatment withCytE than with colchicine (P < 0.001) (Fig. 5c). Actinpolymerization via a phosphotyrosine-signaling cascadewas found to be necessary for adherence and internaliza-tion by U-937 cells (P < 0.001) of tox– and tox+ strains.For the tox– strain, inhibition of bacterial association(Fig. 5a) was more effective after treatment with colchicineand genistein than with CytE (P < 0.001), whereas in-hibition of bacterial intracellular viability (Fig. 5b) wasmore effective after treatment with CytE than with genis-tein and colchicine (P < 0.001). Viable bacteria contin-ued to be detected in intracellular lysates CytE, genis-tein and colchicines-treated monolayers infected with thetox– strain.
C. diphtheriae promotes macrophageapoptosis and necrosis
After observations of nuclear morphological changes byDAPI staining at T-3 hr, results of ELISA showed that
C. diphtheriae infection caused a time-dependent DNAfragmentation in U937 macrophages (Fig. 4g). Althoughearly apoptosis and necrosis were observed at higher levelsfor the tox+ strain, the tox– strain was also able to induceboth mechanisms of cell death. DNA fragmentation wasinitiated 1 hr and 3 hr post-infection by the tox+ andtox– strains, respectively. As DNA fragmentation precedesmore visible markers of apoptosis, such as plasma mem-brane breakdown, this finding is not inconsistent withresults from microscopic inspection of infected monolay-ers that showed phenotypically normal aspects until 3 hrpost-infection.
DISCUSSION
Macrophages can be heterogeneous with regard to theirability to kill facultative intracellular bacteria. That is, amacrophage may bind, or bind and internalize a bac-terium, yet not be able to kill it. It is therefore impor-tant to distinguish these activities experimentally. Becausemost bacteria are killed quite quickly by macrophages,bactericidal assays are best measured within a few hoursafter co-incubation of cells and bacteria. The receptors amacrophage uses to internalize bacterial cells may influ-ence the intracellular fate of the organism (31).
Results achieved during this investigation showed thathuman macrophages could be heterogeneous with re-gard to their ability to kill toxigenic and non-toxigenic
c© 2009 The Societies and Blackwell Publishing Asia Pty Ltd 5
C. S. dos Santos et al.
Fig. 3. Cytotoxicity of U-937 macrophages infected with C. diphthe-riae strains assessed by Trypan blue exclusion test. Light micrographsillustrating non-viable stained cells exposed (T-3 hr) to (A) ATCC 27010tox–, (B) ATCC 27010 tox– added to exogenous diphtheria toxin (DT),
(C) ATCC 27012 tox+ and (D) respective percentages of non-viable cells.Magnification ×400. Data are presented as the mean ± SD of three in-dependent experiments carried out in triplicate; Statistical confidenceinterval of 95%.
C. diphtheriae strains. The presence and consequent ex-pression of the tox gene seemed to favor non-opsonic in-teraction of U-937 cells with C. diphtheriae by enhancingbacterial adherence and internalization.
Some microorganisms can avoid the killing mecha-nisms of phagocytes by inhibiting attachment to phago-cytes or by inhibiting phagocytosis itself (32). Likewise,the C. diphtheriae tox– strain adhered weakly to U-937 cells and only a small proportion of viable bacteriawere phagocytosed, indicating that C. diphtheriae is ca-pable of avoiding non-opsonic phagocytoses by humanmacrophages. This mechanism may facilitate C. diph-theriae remaining alive and causing disease despite thepresence of vigorous immune cell infiltration consistingof PMN and macrophages. The low incidence of inges-tion of bacterial cells without the tox gene suggests that
there might be qualitative and/or quantitative differencesamong molecules expressed by tox+ and tox– strains thatare involved in non-opsonic phagocytosis by U-937 cells.Consequently, different receptors other than those usedby human macrophages during the interaction with DTare involved in the internalization of the tox– strain. Ad-ditional studies remain necessary in order to investigatethe type of membrane receptors involved and the partici-pation of non-antibody opsonins in this process.
Although the tox– strain adhered to U-937 cells lessefficiently, bacterial cells that were phagocytosed remainedalive intracellularly for 24 hr, suggesting that C. diphtheriaewithout the tox gene may remain alive in the presenceof macrophages for long periods of time, as previouslyobserved for Streptococcus agalactiae (33). The fact thatthe tox– strain shows the ability to survive and replicate in
6 c© 2009 The Societies and Blackwell Publishing Asia Pty Ltd
C. diphtheriae and U-937 cell interaction
Fig. 4. DAPI fluorescence microscopy assays (A–F) and ELISA of histone-associated DNA fragments (G,H). Light micrographs of DAPI assays il-lustrating nuclear morphological changes of U937 macrophages at T-3 hr with C. diphtheriae (A) ATCC 27010 tox–, (B) ATCC 27010 tox–added to DT and (C) ATCC 27012 tox+ strains. (D) Vacuolization,
(E) nuclear fragmentation, (F) bleb formation were predominantly ob-served in monolayers infected with the tox+ strain. DNA fragmentationlevels detected in (G) U937 adherent cells and (H) supernatants of in-fected macrophages. Magnification ×1000.
supernatants of infected U-937 cells is an indication of theresistance to non-opsonic phagocytosis of C. diphtheriaestrains without the tox gene. U-937 macrophages seemedbacteriostatic for the tox– strain, as they allowed bacterialgrowth in supernatants of cultures 24 hr post-infection.Bacteriostatic effects of macrophages could later allow thebacteria to grow and colonize the host (34).
More than 80% of the macrophages remained viable at3 hr post-infection with the tox– strain, suggesting thatinhibition of non-opsonic phagocytosis was not due to
macrophage cytotoxicity. As the tox- strain caused thedeath of some phagocytes, we cannot exclude the pos-sibility of bacterial cells that grew in and then killedmacrophages, and grew extracellularly afterwards, as pre-viously observed with other pathogens. If bacteria kill amacrophage and escape into the extracellular milieu, thiscan also cause a problem (31).
The tox+ strain did not replicate or survive inside U-937cells, but instead was killed 24 hr post-infection. Previousstudies have shown that bacterial exotoxins such as DT
c© 2009 The Societies and Blackwell Publishing Asia Pty Ltd 7
C. S. dos Santos et al.
Fig. 5. Effects of cytochalasin E, genistein and colchicine on (A,C) adherence to and (B,D) internalization of (A,B) tox– and (C,D) tox+ C. diphtheriaestrains by U-937 macrophages at a MOI of 100.
could powerfully activate macrophages (35). As the mi-crobicidal activity of macrophages may be enhanced byDT, we investigated whether such macrophage activationaffected the intracellular survival of C. diphtheriae. Thepresence of DT was relevant in determining the fate ofphagocytosed C. diphtheriae. In fact, a more rapid reduc-tion in the number of intracellular bacteria was observedin macrophages infected with the tox+ strain.
Phagocytosis of extracellular particles are highly depen-dent on actin polymerization and reorganization (36).Positive FAS test results showed that internalization ofC. diphtheriae occurred by a microfilament-dependentphagocytosis process, suggesting that in the absence ofopsonins, some C. diphtheriae strains are able to enterand survive in macrophages by evading intracellular an-tibacterial activities. Actin polymerization along with aphosphotyrosine signaling event was a mechanism re-quired during the C. diphtheriae internalization process.The PKC signal transduction pathway is a useful intracel-lular target to ensure survival of several pathogens insidemacrophages. Previous studies have provided evidencethat a defect in PKC-dependent events rendered murineperitoneal macrophages more permissive to the intracel-lular survival of S. agalactiae (22). Unlike S. agalactiae,the inhibition of the PKC-pathway by pretreatment withgenistein, a tyrosine kinase inhibitor, prevented the sur-
vival of C. diphtheriae strains within U-937 macrophages,especially for the tox+ strain. Inhibition assays with CytEand colchicine showed involvement of microfilamentsand microtubules at different levels. The microtubule-disrupting drug colchicine caused a complete inhibitionof intracellular viability of the tox+ strain.
Non-opsonic phagocytosis of tox+ and tox– inhibitedbacterial growth and induced cytopathogenicity in U-937cells at different levels. More than 42% of the macrophageswere killed in experiments (T-3 hr) with the tox+ strain,indicating a higher cytotoxicity for U-937 cells by viablebacteria carrying the tox gene when compared to the tox–strain (18.33%). Therefore, bacterial intracellular replica-tion did not appear necessary to induce cytopathogenicityin U-937-cells, as previously observed for other humanpathogens (37). Both strains exhibited cytotoxic activityindependent of DT. Conversely, DT enhanced cytotoxicproperties of the tox+ strain.
DT-mediated inhibition of protein synthesis is impor-tant for induction of cytolysis (17, 38). Purified DT me-diates cell lysis and intranucleosomal DNA fragmentation(39). In a recent investigation, immunohistochemical ex-amination revealed that alveolar macrophages were de-pleted via apoptosis in mice treated with DT (35, 40). Al-though cytopathogenicity of DT to the host cell has beenwell documented, the mechanisms of cell death due to
8 c© 2009 The Societies and Blackwell Publishing Asia Pty Ltd

C. diphtheriae and U-937 cell interaction
infection by C. diphtheriae are not well understood. In thepresent study, we documented the induction of apoptosisand necrosis by C. diphtheriae.
According to the literature, the macrophage’s death maybe partially due to the activation of the apoptosis machin-ery (37, 39–42). The microtubule cytoskeleton is reformedduring the execution phase of apoptosis in many cell typesand under different apoptotic stimuli. The microtubulecytoskeleton plays an important role in preserving plasmamembrane integrity during apoptosis (43). Cytoskeletonrearrangements during C. diphtheriae infection seemedto stimulate death of U-937 macrophages, especially dueto the fact that the tox– strain was also capable of causingnecrosis and apoptosis. Data indicate that the mechanismsof cell death due to infection by C. diphtheriae are not ex-clusively associated with DT production or the presence ofthe tox gene. There are at least two virulence mechanismsby which C. diphtheriae induces cell death by necrosisand/or apoptosis.
The discovery that macrophages can partially phago-cytose C. diphtheriae in the absence of immune serumsuggests that there is a potential role for antibody-independent mechanisms in resistance to infection. Thecurrent demonstration that C. diphtheriae strains survivedwithin U-937 cells indicates that human macrophages, inthe absence of opsonins, may be not effective in killingC. diphtheriae. Adherence and intracellular survival ofC. diphtheriae within human macrophages can be in-volved in throat colonization contributing to the failure ofbacterial eradication and asymptomatic carriage. In addi-tion, intracellular adherence and survival within profes-sional phagocytes may help to account for the virulence ofC. diphtheriae in endovascular infections and for the abil-ity of this microorganism, independent of toxin produc-tion, to establish metastatic foci of infection. Moreover, in-tracellular localization of C. diphtheriae in macrophagescould protect microorganisms from the more effectivemicrobicidal activity of PMN, and from the action ofantimicrobial agents. Data led to the hypothesis thatC. diphtheriae may infect heart valves partially throughphagocyte-facilitated invasion also known as the ‘Trojanhorse’ mechanism (44). This fact may help to explain theaggressive nature of the disease leading to valve replace-ment and/or death of patients with endocarditis despiteantimicrobial therapy in many situations.
ACKNOWLEDGMENTS
This work was supported by grants from CNPq, CAPES,FAPERJ, SR-2/UERJ and Programa de Nucleo de Ex-celencia (PRONEX) of the Brazilian Ministry of Scienceand Technology.
REFERENCES
1. Gilbert L. (1997) Infections with Corynebacterium diphtheriae –changing epidemiology and clinical manifestations. Report of thethird international meeting of the European Laboratory WorkingGroup on Diphtheria (ELWGD), Institute Pasteur, Paris 7–8 June1996. Commun Dis Intell 21: 161–4.
2. Galazka A.M. (2000) The changing epidemiology of diphtheria inthe vaccine era. J Infect Dis 181: S2–S9.
3. Khan N., Shastri J., Argal U., Doctor B. (2007) Resurgence ofdiphtheria in the vaccination era. Indian J Med Microbiol 25: 434–9.
4. Efstratiou A., George R.C. (1996) Microbiology and epidemiologyof diphtheria. Rev Med Microbiol 7: 31–42.
5. Mattos-Guaraldi A.L., Moreira L.O., Damasco P.V., Hirata Jr. R.(2003) Diphtheria remains a threat to health in the developingworld – An overview. Mem Inst Oswaldo Cruz 98: 987–93.
6. Mattos-Guaraldi A.L., Formiga L.C.D., Pereira G.A. (2000) Cellsurface components and adhesion in Corynebacterium diphtheriae.Microb Infect 2: 1507–12.
7. Patey O., Bimet F., Riegel P., Halioua B., Emond J.P., Estrangin E.,Dellion S., Alonso J.M., Kiredjian M., Dublanchet A., Lafaix C.(1997) Clinical and molecular study of Corynebacteriumdiphtheriae systemic infections in France. Coryne Study Group. JClin Microbiol 35: 441–5.
8. Mattos-Guaraldi A.L., Formiga L.C.D. (1998) Bacteriologicalproperties of a sucrose-fermenting Corynebacterium diphtheriaestrain isolated from a case of endocarditis. Curr Microbiol 37:156–8.
9. Belko J., Wessel D.L., Malley R. (2000) Endocarditis caused byCorynebacterium diphtheriae: case report and review of theliterature. Pediatr Infect Dis J 19: 159–63.
10. Hirata Jr. R., Napoleao F., Monteiro-Leal L.H., Andrade A.F.B.,Nagao P.E., Formiga L.C.D., Fonseca L.S., Mattos-Guaraldi A.L.(2002) Intracellular viability of toxigenic Corynebacteriumdiphtheriae strains in HEp-2 cells. FEMS Microbiol Lett 215: 115–9.
11. Bertuccini L., Badassarri L., von Hunolstein C. (2004)Internalization of non-toxigenic Corynebacterium diphtheriae bycultured human respiratory epithelial cells. Microbial Pathog 37:111–8.
12. Schuchat A. (1999) Group B Streptococcus. Lancet 353: 51–6.13. Ofek I., Goldhar J., Keisari Y., Sharon N. (1995) Non-opsonic
phagocytosis of microorganisms. Annu Rev Microbiol 49: 239–76.14. Aderem A., Underhill D.M. (1999) Mechanisms of phagocytosis in
macrophages. Ann Rev Immunol 17: 593–623.15. Iseberg R.R. (1991) Discrimination between intracellular uptake
and surface adhesion of bacterial pathogens. Science 252: 934–8.16. Gordon S.B., Read R.C. (2002) Macrophage defences against
respiratory tract infections. The immunology of childhoodrespiratory infections. Br Med Bull 61: 45–6.
17. Saelinger C., Bonventre P.F., Imhoff J. (1975) Interaction of toxin ofCorynebacterium diphtheriae with phagocytes from susceptible andresistant species. J Infect Dis 13: 431–8.
18. D’Onofrio C., Paradisi F. (1983) Studies dealing with the influenceof bacterial exotoxins and endotoxins on the phagocytic activity ofhuman macrophages in culture. Infect 11: 137–43.
19. Sundstrom C., Nilsson K. (1976) Establishment andcharacterization of a human histiocytic lymphoma cell line(U-937). Int J Cancer 17: 565–77.
20. Todd R.F., Liu D.Y. (1986) Mononuclear phagocyte activation,activation-associated antigens. Fed Proc 45: 2829–36.
21. Hirata Jr. R., Souza S.M.S., Rocha-de-Souza C.M., Andrade A.F.B.,Monteiro-Leal L.H., Formiga L.C.D., Mattos-Guaraldi A.L. (2004)Patterns of adherence to HEp-2 cells and actin polymerisation by
c© 2009 The Societies and Blackwell Publishing Asia Pty Ltd 9

C. S. dos Santos et al.
toxigenic Corynebacterium diphtheriae strains. Microb Pathog 36:125–30.
22. Monteiro G.C., Hirata Jr. R., Andrade A.F., Mattos-Guaraldi A.L.,Nagao P.E. (2004) Surface carbohydrates as recognitiondeterminants in non-opsonic interactions and intracellular viabilityof group B Streptococcus strains in murine macrophages. Int J MolMed 13: 175–80.
23. Ordway D., Viveiros M., Leandro C., Bettencourt R., Almeida J.,Martins M., Kristiansen J. E., Molnar J., Amaral L. (2003). Clinicalconcentrations of thioridazine kill intracellular multidrug-resistantMycobacterium tuberculosis. Antimicrob Agents Chemother 47:917–22.
24. Fellows M.D., O’Donovan M.R. (2007) Cytotoxicity in culturedmammalian cells is a function of the method used to estimate it.Mutagen 22: 275–80.
25. Ghosh S., Pal S., Das S., Dasgupta S.K., Majumdar S. (1998)Lipoarabinomannan induced cytotoxic effects in humanmononuclear cells. FEMS Immunol Med Microbiol 21: 181–8.
26. Knutton S., Baldwin T., Williams P.H., McNeish A.S. (1989) Actinaccumulation at sites of bacterial adhesion to tissue culture cells:basis of a new diagnostic test for enteropathogenic andenterohemorrhagic Escherichia coli. Infect Immun 57: 1290–98.
27. Rosenshine I., Ruschkowski S., Finlay B.B. (1994) Inhibitors ofcytoskeletal function and signal transduction to study bacterialinvasion. Methods Enzymol 236: 467–76.
28. Su B., Johansson S., Fallman M., Patarroyo M., Granstrom M.,Normark S. (1999) Signal transduction-mediated adherence andentry of Helicobacter pylori into cultured cells. Gastroenterol 117:595–604.
29. Baldassarri L., Bertuccini L., Ammendolia M.G., Cocconcelli P.,Arciola C.R., Montanaro L., Creti R., Orefici G. (2004)Receptor-mediated endocytosis of biofilm-forming Enterococcusfaecalis by rat peritoneal macrophages. Indian J Med Res119(Suppl): 131–5.
30. Joo N.E., Watanabe T., Chen C., Chekenya M., Stallcup W.B., KapilaY.L. (2008) NG2, a novel proapoptotic receptor, opposes integrinalfa-4 mediate anoikis through PKC alfa-dependent suppression ofFAK phosphorylation. Cell Death Differ 15: 899–907.
31. Hilbi H., Zychlinsky A., Sansonetti P.J. (1997) Macrophageapoptosis in microbial infections. Parasitol 115: S79–S87.
32. Wood E.T., Ala G.E., Dutro S.M., Totten P.A. (2001) Haemophilusducreyi inhibits phagocytosis by U-937 cells, a humanmacrophage-like cell line. Infect Immun 69: 4726–33.
33. Cornacchione P., Scaringi L., Fettucciari K., Rosati E., Sabatini R.,Orefici G., von Hunolstein C., Modesti A., Modica A., Minelli F.,Marconi P. (1998) Group B streptococci persist insidemacrophages. Immunol 93: 86–95.
34. Campbell P.A., Canono B.P., Drevets D.A. (1994) Measurement ofbacterial ingestion and killing by macrophages. Curr ProtocolsImmunol Supplem 12: 14.6.13.
35. Mookerjee B.K., Kanegasaki S., Kato I. (1982) Effects of diphtheriatoxin and other exotoxins on oxidant generation by human andmurine phagocytes. Dev Comp Immunol 6: 161–170.
36. Diakonova M., Bokoch G., Swanson J.A. (2002) Dynamics ofcytoskeletal proteins during Fc gamma receptor-mediatedphagocytosis in macrophages. Mol Biol Cell 13: 402–11.
37. Gao L.Y., Kwaik Y.A. (1999) Apoptosis in macrophages and alveolarepithelial cells during early stages of infection by Legionellapneumophila and its role in cytopathogenicity. Infect Immun 67:862–70.
38. Morimoto H., Bonavida B. (1992) Diphtheria toxin- andPseudomonas A toxin-mediated apoptosis. ADP ribosylation ofelongation factor-2 is required for DNA fragmentation and cell lysisand synergy with tumor necrosis factor-alpha. J Immunol 149:2089–94.
39. Thorburn A., Thorburn J., Frankel A.E. (2004) Induction ofapoptosis by tumor cell-targeted toxins. Apoptosis 9: 19–25.
40. Miyake Y., Kaise H., Isono K., Koseki H., Kohno K., Tanaka M.(2007) Protective role of macrophages in noninflammatory lunginjury caused by selective ablation of alveolar epithelial type II cells.J Immunol 15: 5001–9.
41. Leek M.D., Sivaloganathan S., Devaraj S.K., Zamiri I., GriffithsG.D., Green M.A. (1990) Diphtheria with a difference – a rareCorynebacterium fatality with associated apoptotic cell death.Histopathol 16: 187–9.
42. Ruckdeschel K., Roggenkamp A., Lafont V., Mangeat P.,Heesemann J., Rouot B. (1997) Interaction of Yersinia enterocoliticawith macrophages leads to macrophage cell death throughapoptosis. Infect Immun 65: 4813–21.
43. Sanchez-Alcazar J.A., Rodriguez-Hernandez A., Cordero M.D.,Fernandez-Ayala D.J., Brea-Calvo G., Garcia K., Navas P. (2007)The apoptotic microtubule network preserves plasma membraneintegrity during the execution phase of apoptosis. Apoptosis 12:1195–1208.
44. Drevets D.A. (1999) Dissemination of Listeria monocytogenes byinfected phagocytes. Infect Immun 67: 3512–7.
10 c© 2009 The Societies and Blackwell Publishing Asia Pty Ltd





























