Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Neurotrophins in Mesial Temporal Lobe EpilepsyWith and Without Psychiatric Comorbidities
Ludmyla Kandratavicius, PhD, Mariana Raquel Monteiro, MSc,Joao Alberto Assirati, Jr., MD, PhD, Carlos Gilberto Carlotti, Jr., MD, PhD,
Jaime Eduardo Hallak, MD, PhD, and Joao Pereira Leite, MD, PhD
AbstractDespite the strong association between epilepsy and psychiatric
comorbidities, data on clinicopathologic correlations are scant. Wepreviously reported differential mossy fiber sprouting (MFS) in mesialtemporal lobe epilepsy (MTLE) patients with psychosis (MTLE + P)and major depression (MTLE + D). Because neurotrophins (NTs) canpromote MFS, here, we investigated MFS, neuronal density and im-munoreactivity for the NT nerve growth factor (NGF), brain-derivedneurotrophic factor (BDNF) and neurotrophin 3 (NT3) in hippocampiof 14 MTLE patients without a psychiatric history, 13 MTLE + D, 13MTLE + P, and 10 control necropsies. Mossy fiber sprouting correlatedwith granular layer NGF immunoreactivity and seizure frequency. Pa-tients with secondarily generalized seizures exhibited less NGF immu-noreactivity versus patients with complex partial seizures. There wasgreater NT immunoreactivity in MTLE versus control groups but lesserNT immunoreactivity in MTLE + P versus MTLE patients; these find-ings correlated with neuropsychologic scores. Patients with MTLE + Dtaking fluoxetine showed greater BDNF immunoreactivity than those nottaking fluoxetine; MTLE + P patients taking haloperidol had decreasedneuronal density and immunoreactivity for NGF and BDNF in specificsubfields versus those not taking haloperidol. There were no differencesin NT3 immunoreactivity among the groups. These findings support aclose association between MFS and NT expression in the hippocampi of
MTLE patients and suggest that distinct structural and neurochemicalmilieu may contribute to the genesis or maintenance of psychiatriccomorbidities in MTLE.
Key Words: Brain-derived neurotrophic factor, Depression, Nervegrowth factor, Neurotrophin, Psychiatric comorbidity, Psychosis,Temporal lobe epilepsy.
INTRODUCTIONTemporal lobe epilepsy (TLE) is the most common cause
of intractable epilepsy in adults. The mesial subtype (MTLE)usually shows hippocampal sclerosis (HS), neuronal loss,gliosis, and mossy fiber sprouting (MFS) (1Y5). Psychiatriccomorbidities are very frequent in TLE patients (6Y11), al-though the precise nature of this association is still a matterof debate (12, 13). We have recently shown neuropathologicdata suggesting that there is a structural basis for psychiatricmanifestations in MTLE patients, such as increased MFS inpatients with a history of major depression and decreasedMFS in interictal psychosis (4).
Neurotrophins (NTs) are low-molecular-weight growthfactors that act as extracellular ligands and affect differen-tiation, maintenance, and survival of cells. Among these endo-genous proteins, nerve growth factor (NGF), brain-derivedneurotrophic factor (BDNF), and neurotrophin 3 (NT3) wereshown to promoteMFS in different animal and in vitromodels ofTLE (14Y16). Neurotrophins are upregulated in the dentate gyrusof TLE patients (17), and in animal models, seizure activity resultsin a transient increase of NGF and BDNF in hippocampal andneocortical neurons (14, 18Y21). Although unaltered in the kin-dling model (15), NT3 expression is increased in the subchronicphase of the kainate model (22). Moreover, prolonged NT3intracerebral infusion triggers sprouting and might play anantiepileptogenic role (15). In major depression and schizo-phrenia, decreased NT expression has been documented (23Y27),whereas antidepressant and antipsychotic treatment might restoreNT plasma levels (23, 28, 29). Interestingly, chronic treatmentwith antidepressants, antipsychotics, and benzodiazepine in naiverats does not alter BDNF or NGF hippocampal levels (30, 31).
Although growth factor involvement in TLE and neuro-psychiatric disorders seems unequivocal, the detailed expressionpatterns and functions of NTs within the human hippocampalformation remain largely unknown. Here, we characterized NT
1029J Neuropathol Exp Neurol ! Volume 72, Number 11, November 2013
J Neuropathol Exp NeurolCopyright ! 2013 by the American Association of Neuropathologists, Inc.
Vol. 72, No. 11November 2013pp. 1029Y1042
From the Department of Neurosciences and Behavior, School of Medicine ofRibeirao Preto, University of Sao Paulo, Sao Paulo, Brazil (LK, MRM,JEH, JPL); Center for Interdisciplinary Research on Applied Neurosci-ences (NAPNA), University of Sao Paulo, Sao Paulo, Brazil (LK, JEH,JPL); National Institute of Science and Technology in TranslationalMedicine (INCT-TM-CNPq), Brazil (JEH); Department of Surgery,School of Medicine of Ribeirao Preto, University of Sao Paulo, Sao Paulo,Brazil (JAA Jr., CGC Jr.).
Send correspondence and reprint requests to: Joao Pereira Leite, MD, PhD,Department of Neurosciences and Behavior, School ofMedicine of RibeiraoPreto, University of Sao Paulo, Av Bandeirantes 3900, CEP 14049-900,Ribeirao Preto, Sao Paulo, Brazil; E-mail: [email protected]
This work was supported by the Fundacao de Apoio a Pesquisa do Estado deSao Paulo-Fapesp (07/56721-7 to Ludmyla Kandratavicius, CInAPCeProject 05/56447-7 to Joao Pereira Leite) and Conselho Nacional deDesenvolvimento Cientifico e Tecnologico-CNPq.
The authors declare that they have no conflict of interest. The funders had norole in study design, data collection and analysis, decision to publish, orpreparation of the manuscript.
Supplemental digital content is available for this article. Direct URL citationsappear in the printed text and are provided in the HTML and PDF versionsof this article on the journal’s Web site (www.jneuropath.com).
Copyright © 2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

immunoreactivity in subfields of the mesial temporal structures,including the hippocampus, subicular complex, and entorhinalcortex in a new cohort of MTLE patients with and without co-morbid major depression and psychosis. We also investigatedthe possible correlation between NT immunoreactivity andMFSand further clinical characteristics.
MATERIALS AND METHODS
PatientsWe analyzed the hippocampal formation from MTLE
specimens freshly collected in the operating room and non-epileptic controls from necropsy, collected between 3.5 and6 hours after death. A less than 24-hour postmortem time limitallows comparison of necropsy tissue with freshly collectedsurgical specimens for their protein levels, cell morphology,and tissue integrity (32, 33). Tissue collection and processingwere conducted according to protocol approved by our in-stitution’s research ethics board.
Mesial temporal lobe epilepsy specimens were derivedfrom 40 MTLE patients who underwent a standard en blocanterior temporal resection (including 3Y4 cm of the hippo-campus) for medically intractable seizures. All had clinical
neuropathologic confirmation of HS. They were divided into3 groups: 14 MTLE patients without any history of psychi-atric disorder (MTLE group); 13 MTLE patients with interic-tal psychosis (MTLE + P group); and 13 MTLE patients with adiagnosis of major depression (MTLE + D group). Accordingto Blumcke HS categories (34), 17 patients had severe HS, 16classical HS, 5 CA1 HS, and 2 CA4 HS. No differences wereseen among MTLE groups with respect to HS categories (Table,Supplemental Digital Content 1, http://links.lww.com/NEN/A508).For comparison, 10 nonepi leptic control hippocampi fromnecropsies were processed and analyzed in the same manneras the surgical cases. Underlying diseases causing death werecardiomyopathy, pulmonary infarct, or renal-hepatic failure,with no history of hypoxic episodes before death, seizures,or neurologic diseases. Furthermore, there was no evidence ofpathologic abnormalities of the mesial temporal structures onpostmortem examination. Mesial temporal lobe epilepsy andcontrol specimens were collected between 1996 and 2006.Clinical characteristics of all groups are shown in the Table.
Clinical Features of MTLE PatientsAll patients were referred for presurgical assessment be-
cause of drug-resistant seizures, as defined byBerg (35). Patients
TABLE. Demographic and Clinical DataMTLE MTLE + D MTLE + P Controls Statistics
Male (n) 8 5 7 4 No difference
Female (n) 6 8 6 6
IPI present (n) 9 8 7 na No difference
IPI absent (n) 5 5 6 na
Febrile seizure present in IPI (n) 5 3 5 na No difference
Febrile seizure absent in IPI (n) 4 3 1 na
Febrile seizure unknown (n) 0 2 1 na
Age of first seizure, years 3.0 T 2.3 4.3 T 5.5 6.8 T 8.4 na No difference
Age when seizures became recurrent or age of onset, years 10.2 T 6.4 13.0 T 8.1 13.8 T 8.4 na No difference
Seizure type: CPS (n) 6 1 4 na No difference
Seizure type: SGS (n) 8 12 9 na tr
Seizure frequency (monthly) 12.7 T 10.1 17.2 T 15.6 11.7 T 10.1 na No difference
Right HS (n) 6 7 5 na No difference
Left HS (n) 6 4 6 na
Bilateral HS (n) 2 2 2 na
Right handedness (n) 12 11 13 na No difference
Left handedness (n) 1 2 0 na
Bilateral handedness (n) 1 0 0 na
Memory in verbal tasks: average or above (n) 3 7 2 na No difference
Memory in verbal tasks: below average (n) 11 6 11 na
Memory in nonverbal tasks: average or above (n) 9 7 3 na No difference
Memory in nonverbal tasks: below average (n) 5 6 10 na tr
Full-scale IQ 82.2 T 7.9 84.3 T 9.6 77.2 T 7.3 na No difference
Years at school 6.2 T 3.0 5.1 T 3.6 4.2 T 2.6 na No difference
Age at surgery (or at death for controls), years 36.1 T 4.7 37.1 T 5.4 38.0 T 6.0 48.1 T 18.9 No difference
Duration of epilepsy, years 25.9 T 6.4 24.1 T 8.9 24.1 T 8.6 na No difference
Collected side: right (n) 7 8 6 6 No difference
Collected side: left (n) 7 5 7 4
Values are indicated as mean T SD when applicable. There was a statistical trend (tr: 0.05 Q p e 0.07), suggesting a higher proportion of MTLE + D patients with SGS than MTLEand a higher proportion of MTLE + P patients with worse performance in nonverbal memory tasks than MTLE.
CPS, complex partial seizure; HS, hippocampal sclerosis; IPI, initial precipitant injury; MTLE, mesial temporal lobe epilepsy; MTLE + D, MTLE patients with a diagnosis of majordepression; MTLE + P, MTLE patients with interictal psychosis; na, not applicable; SGS, secondarily generalized seizures.
Kandratavicius et al J Neuropathol Exp Neurol ! Volume 72, Number 11, November 2013
! 2013 American Association of Neuropathologists, Inc.1030
Copyright © 2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

were evaluated at the Ribeirao Preto Epilepsy Surgery Programusing standardized protocols approved by the institution’sethics committee, and written consent was obtained from eachpatient. Presurgical investigation at the Epilepsy MonitoringUnit included detailed clinical history, neurologic examination,interictal and ictal scalp/sphenoidal electroencephalography,neuropsychology evaluation, and intracarotid amobarbitalmemory and language procedure whenever deemed clinicallynecessary.
Mesial temporal lobe epilepsy was defined followingEngel criteria (36): 1) seizure semiology consistent with MTLE,usually with epigastric/autonomic/psychic auras, followed bycomplex partial seizures (CPSs); 2) presurgical investigationconfirming seizure onset zone in the temporal lobe; 3) anteriorand mesial temporal interictal spikes on electroencephalogra-phy; 4) no lesions other than unilateral or bilateral hippocampalatrophy on high-resolution magnetic resonance imaging scans(reduced hippocampal dimensions and increased T2 signal);5) clinical histopathologic examination compatible with HS;and 6) no evidence of dual pathology identifiable by any of theassessment methods described (clinical, electrophysiology,neuroimaging, and histopathology). Exclusion criteria werefocal neurologic abnormalities on physical examination, gen-eralized or extratemporal electroencephalography spikes, andmarked cognitive impairment indicating dysfunction beyondthe temporal regions.
Information regarding antecedent of an initial precipi-tant injury, febrile seizures, seizure types, drug regimen, andestimated monthly frequency (within the 2 years before sur-gery) were retrospectively collected from medical recordsfor each patient. Psychiatric evaluations were conducted inall MTLE patients. Each diagnosis of major depression wasindependently established during the presurgical evaluationby 2 psychiatrists with experience in psychiatric disordersassociated with epilepsy using the guidelines of the Diagnosticand Statistical Manual of Mental Disorders, Fourth Edition.Once a consensus on the classification of psychotic syndromesassociated with epilepsy is lacking, and neither Diagnosticand Statistical Manual of Mental Disorders, Fourth Edition,nor International Classification of Disease, 10th Revision, hasaddressed this issue specifically, the diagnosis of psychosisassociated with MTLE was established according to Sachdev(37), that is, patients with interictal psychosis did not experi-ence the following: psychotic disorder temporally associatedwith seizures, changes in antiepileptic medications, epilepticstatus, delirium, and psychosis for paradoxical normalization.This group was defined by a prolonged psychotic state thatwas not related to the epileptic seizures. Typically, the psy-chotic states closely resemble schizophrenia, with paranoidideas that might become systematized, ideas of influence, andauditory hallucinations often of a menacing quality. The pointsof difference are common religious coloring of the paranoidideas, tendency of the affect to remain warm and appropriate,and no typical deterioration to the hebephrenic state (9). Pa-tients had no history of psychiatric disorders (before seizureonset) or of substance dependence at any time. Global IQ wascalculated after neuropsychologic tests (complete WAIS-III orWAIS-R protocol).
Tissue Collection and ImmunohistochemicalProcessing
Specimens were segmented into 1-cm blocks transverselyoriented to the hippocampal long axis. Blocks were placed in neo-Timm fixative solution (0.1% sodium sulfide [Sigma, St. Louis,MO] in Millonig buffered glutaraldehyde [Vetec, Rio de Janeiro,Brazil]) or bufferedparaformaldehyde (Sigma).After 48 to96hours,the specimens were processed for neo-Timm histochemistry andhematoxylin and eosin (Laborclin, Pinhais, Brazil) staining or par-affin embedded for immunohistochemistry.
Cryostat sections were mounted on chrome-alum gelatin-coated slides and air-dried for neo-Timm staining (2). Slideswere processed in batches containing at least 1 MTLE and 2control slides. Slides were immersed in a physical developermaintained at 26-C in a darkroom. Developer consisted of10 mL of a 50% gum arabic (Sigma) solution, 30 mL of anaqueous solution of 1.3 mol/L citric acid (Merck, Darmstadt,Germany), 0.9 mol/L sodium citrate (Merck), 90 mL of anaqueous solution of 0.5 mol/L hydroquinone (Merck), and1.5 mL of a 17% silver nitrate solution (Merck). Light sectionswere developed for 40minutes and dark sections for 50minutes.Slides were washed in distilled water for 5 minutes and runningtap water for 10 minutes. They were subsequently air-dried,ethanol dehydrated, xylene cleared, and coverslipped.
Immunohistochemistry was performed with antibodiesthat identified immunoreactivity for NeuN, a nuclear pro-tein found in the nuclei of mature neurons (1:1000 dilu-tion; Chemicon-Millipore, Billerica, MA) and for the NTs:neurotrophic growth factor (1:100 dilution; Santa Cruz Bio-technology, Santa Cruz, CA), BDNF (1:30 dilution; Chemicon-Millipore), and NT3 (1:30 dilution; Chemicon-Millipore).According to the manufacturers and selected references, theseantibodies show no cross-reactivity with other NTs and/orare capable of neutralizing the biologic activity of its ownisoform only (38Y40). Their Western blot profile in humantissue is shown in Figure, Supplemental Digital Content 2,http://links.lww.com/NEN/A509. Briefly, paraffin-embeddedMTLE and control hippocampi were processed together foreach antibody as described in Kandratavicius et al (5), withovernight incubation at room temperature and developed si-multaneously for 10 minutes in 0.05% 3,3¶-diaminobenzidinetetrahydrochloride (Pierce, Rockford, IL) and 0.01% H2O2
(Merck). After sufficient colorization, reaction was halted bywashing in several rinses of distilled water, dehydrated throughgraded ethanol to xylene (Merck), and coverslipped withKrystalon (EM Science, Gibbstown, NJ). Adjacent sectionswere hematoxylin and eosin stained and examined for tissueintegrity. Control sections without the primary antisera did notreveal staining (data not shown).
Cell Count and Neo-Timm QuantificationMesial temporal lobe epilepsy and control hippocampi
were compared for neuronal density and neo-Timm staining.Neuronal counting was performed based on Lorente de Noclassification (41), including fascia dentata granular andsubgranular cells, polymorphic hilar neurons (limited to a re-gion between stratum granulosum and CA4 pyramidal cells,being at least 50 Km from the stratum granulosum and 100 Km
J Neuropathol Exp Neurol ! Volume 72, Number 11, November 2013 NTs in MTLE With Psychiatric Comorbidities
! 2013 American Association of Neuropathologists, Inc. 1031
Copyright © 2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

from CA4), as well as pyramidal cells in CA4 (the portion ofAmmon horn that permeates the inner part of the dentategyrus), CA3, CA2, CA1, prosubiculum, subiculum, parasu-biculum, and entorhinal cortex layer III. Cell densities (neu-rons per cubic millimeter) were estimated in 8-Km NeuN-stained slices at 400" magnification with a morphometric
grid methodology using Abercrombie correction (42), aspreviously described and well established in the literature forsurgical hippocampal fragments (1, 3Y5, 43Y47). We alsoperformed measurements of granular layer dispersion usingNeuN-stained sections and the straight line tool of ImageJanalysis system (National Institutes of Health, USA, public
FIGURE 1. Neuronal density in human hippocampal formation subfields. (A, B) NeuN-stained hippocampal formation in a mesialtemporal lobe epilepsy (MTLE) patient (A) and a control (B). There is marked neuronal loss in the MTLE dentate gyrus, Ammon,horn and Sommer sector, as well as granular layer (GL) dispersion. (C) Neuronal density values from MTLE (black bars), MTLE +major depression (MTLE + D) (gray bars), MTLE + psychosis (MTLE + P) (light gray bars), and from nonepileptic controls (whitebars) are indicated as mean T SD. **Significant difference between epileptics and the control group (p G 0.001). Neuronal loss wasobserved in GL, hilus, CA4, CA1, prosubiculum, and entorhinal cortex. There is also a statistical trend (tr: 0.05 Q p e 0.07) todecreased neuronal density in MTLE + P CA2 versus control. ENT, entorhinal cortex; HIL, hilus; PAR, parasubiculum; PRO,prosubiculum; SUB, subiculum. Scale bar = (A, B) 2 mm.
Kandratavicius et al J Neuropathol Exp Neurol ! Volume 72, Number 11, November 2013
! 2013 American Association of Neuropathologists, Inc.1032
Copyright © 2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

domain). The bottom was considered as the farthest granularneuron in the subgranular zone and the top the farthest gran-ular neuron invading the molecular layer.
Mossy fiber sprouting was evaluated in neo-TimmYstained sections in the hilus, granular layer, and innermolecular layer, always in the superior blade of the dentategyrus. Each subfield of each specimen had 3 different samplesmeasured and averaged for statistical analysis. Measurementsof gray value and length were estimated using ImageJ, aspreviously described (4). In brief, length measurements ofsprouting consisted of the distance from the outer border ofthe granular layer to the visible limit of the sprouted mossyfibers within the molecular layer, including the sprouted mossyfibers that eventually invaded the outer molecular layer (mea-surement referred as visible sprouting in the molecular layer).Images were collected and digitized with a high-resolutionCCD monochrome camera attached to an Olympus micro-scope. This method was used to obtain digitized images of neo-
Timm and anti-NeuN, -NGF, -BDNF, and -NT3Ystained slides.Uniform luminance was maintained and checked every 10measurements using an optical density standard and a grayvalue scale ranging from 0 (white) to 255 (black).
Semiquantitative Analysis ofImmunohistochemistry
Slides adjacent to those examined for neuronal densitywere analyzed for NT immunoreactivity. In brief, all digitizedimages taken in 20" magnification were analyzed with ImageJsoftware, following the same criteria: 1) the software identifiesthe gray value distribution of a subfield’s digital image (totalarea for each subfield analyzed = 313.7 " 235.3 Km, whichcorresponds, e.g. to approximately one-third of an Ammonhorn subfield); 2) the immunoreactive area, that is,positivelystained pixels, is selected, limited to a threshold range; and 3)the threshold range is presettled based on control group sec-tions to exclude the low-intensity gray value of backgroundstaining from the analysis. A similar approach was used byour group elsewhere (48). Results for granular layer includedgranular cell layer per se and proximal molecular layer.Analyses were conducted by one investigator (LudmylaKandratavicius), blinded to hippocampal pathology andgroup classification.
Data AnalysisData were analyzed using the statistical program PAWS
(version 18.0) and SigmaPlot (version 11.0). Groups werecompared using analysis of variance (ANOVA one-way, withBonferroni post hoc test) or unpaired t test for variables withnormal distribution and Kruskal-Wallis one-way ANOVA onranks (with Dunn post hoc test) or Mann-Whitney U rank sumtest for variables without normal distribution. Fisher Exact testwas applied for comparison of relative frequencies of clinicalvariables between groups. Other statistical tests included Pear-son correlation analyses and analysis of covariance. Statisticalsignificance was set at p G 0.05, and values were presented asmean T SD.
RESULTS
Clinical ProfilesThe 4 patients groups did not show significant differ-
ences in sex, age, or collected side (Table). Clinical variablessuch as the presence of an initial precipitant injury and febrileseizures, age of first seizure and seizure onset, seizure fre-quency and epilepsy duration, HS side, handedness, IQ, yearsat school, and performance in verbal memory tests were ho-mogeneously distributed among MTLE groups. The MTLE +D patients exhibited a trend to increased proportion of patientswith secondarily generalized seizures versus patients withoutpsychiatric comorbidities (0.05 Q p e 0.07), and MTLE + Ppatients exhibited a trend to worse performance in nonverbalmemory tasks.
All epileptic patients were on antiepileptic drugs (carba-mazepine, oxcarbazepine, phenobarbital, and/or phenytoin). Inaddition, patients were also taking benzodiazepines (MTLE
FIGURE 2. Mossy fiber sprouting in human dentate gyrus.(AYD) Representative images from mesial temporal lobe epi-lepsy (MTLE) (A), MTLE + major depression (MTLE + D) (B),MTLE + psychosis (MTLE + P) (C), and control (D) groups,showing aberrant sprouting in the molecular layer of the den-tate gyrus in the MTLE groups. (E) Increased gray value mea-surements in MTLE and MTLE + D versus control in GL (* p G0.05), IML (** p G 0.001), and OML (tr: 0.05 Q p e 0.07). In theIML, the gray value in the MTLE + P group is lower than that inthe MTLE and MTLE + D groups (# p G 0.05). (F) Lengthmeasurements show greater sprouting in MTLE groups versusthe control group (** p G 0.001) and significant differencesamong the 3 MTLE groups (## p G 0.001). Values from MTLE(black bars), MTLE + D (gray bars), MTLE + P (light gray bars),and from nonepileptic controls (white bars) are indicated asmean T SD. GL, granular layer; H, hilus; IML, inner molecularlayer; OML, outer molecular layer. Scale bar = (AYD) 100 Km.
J Neuropathol Exp Neurol ! Volume 72, Number 11, November 2013 NTs in MTLE With Psychiatric Comorbidities
! 2013 American Association of Neuropathologists, Inc. 1033
Copyright © 2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

group, 8 of 14; MTLE + D, 10 of 13; MTLE + P group, 10 of13), fluoxetine (MTLE + D, 4 of 13), and haloperidol (MTLE +P group, 10 of 13). No differences in neuronal density, MFS, orNT immunoreactivity were seen between patients taking or nottaking benzodiazepines. No differences in neuropsychologictests between patients taking or not taking benzodiazepines,fluoxetine, or haloperidol were seen.
Neuropathologic Characterization: NeuronalDensity and MFS
Evaluation of epileptogenic and control hippocampalformation (Fig. 1A, B) showed reduced neuron density in the
granular layer, hilus, CA4, CA1, and prosubiculum of allMTLE groups when compared with control (Fig. 1C). In ad-dition, there was a significant neuron density reduction in theentorhinal cortex of MTLE patients with major depression andinterictal psychosis versus controls and a trend to decreasedneuronal density in CA2 of MTLE + P specimens. The MTLEgroups exhibited increased granular layer width when com-pared with that in controls (ANOVA F3,47 = 8.31, p G 0.0001),with no statistically significant differences between MTLEgroups (ANOVA F2,37 = 0.20, p = 0.817) (Table, SupplementalDigital Content 1, http://links.lww.com/NEN/A508). The MFSgray value and length were increased in MTLE groups versus
FIGURE 3. Nerve growth factor (NGF) expression in the human hippocampal formation. (AYT) There is greater expression inmesial temporal lobe epilepsy (MTLE) granular cells (A, B), in hypertrophic hilar cells (C, D), and in pyramidal cells in Ammon horn(EYL), subicular complex (MYR), and entorhinal cortex (S, T). Cells with glial profiles (arrows) can also be seen particularly in CA1and prosubiculum of MTLE specimens. ENT, entorhinal cortex; GL, granular layer; HIL, hilus; PAR, parasubiculum; PRO,prosubiculum; SUB, subiculum. Scale bar = (AYT) 50 Km.
Kandratavicius et al J Neuropathol Exp Neurol ! Volume 72, Number 11, November 2013
! 2013 American Association of Neuropathologists, Inc.1034
Copyright © 2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
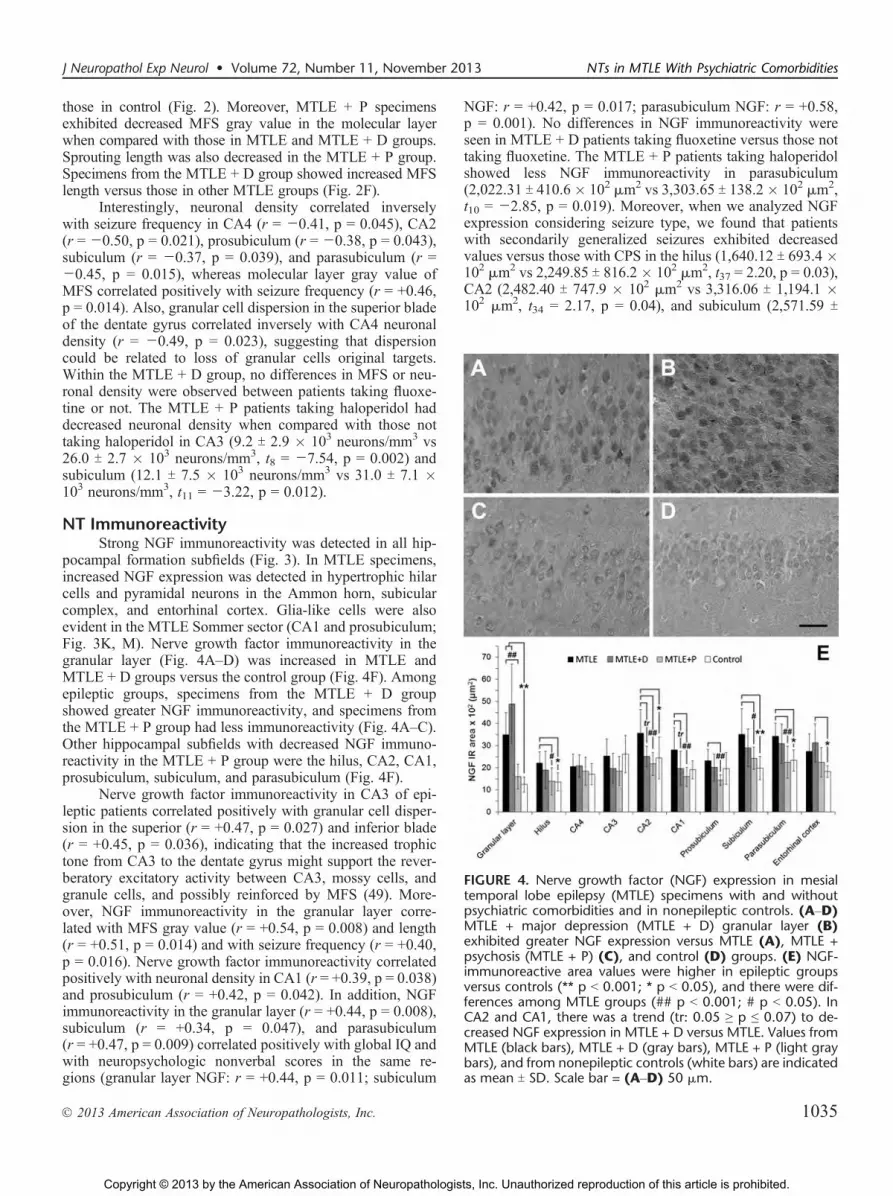
those in control (Fig. 2). Moreover, MTLE + P specimensexhibited decreased MFS gray value in the molecular layerwhen compared with those in MTLE and MTLE + D groups.Sprouting length was also decreased in the MTLE + P group.Specimens from the MTLE + D group showed increased MFSlength versus those in other MTLE groups (Fig. 2F).
Interestingly, neuronal density correlated inverselywith seizure frequency in CA4 (r = j0.41, p = 0.045), CA2(r = j0.50, p = 0.021), prosubiculum (r = j0.38, p = 0.043),subiculum (r = j0.37, p = 0.039), and parasubiculum (r =j0.45, p = 0.015), whereas molecular layer gray value ofMFS correlated positively with seizure frequency (r = +0.46,p = 0.014). Also, granular cell dispersion in the superior bladeof the dentate gyrus correlated inversely with CA4 neuronaldensity (r = j0.49, p = 0.023), suggesting that dispersioncould be related to loss of granular cells original targets.Within the MTLE + D group, no differences in MFS or neu-ronal density were observed between patients taking fluoxe-tine or not. The MTLE + P patients taking haloperidol haddecreased neuronal density when compared with those nottaking haloperidol in CA3 (9.2 T 2.9 " 103 neurons/mm3 vs26.0 T 2.7 " 103 neurons/mm3, t8 = j7.54, p = 0.002) andsubiculum (12.1 T 7.5 " 103 neurons/mm3 vs 31.0 T 7.1 "103 neurons/mm3, t11 = j3.22, p = 0.012).
NT ImmunoreactivityStrong NGF immunoreactivity was detected in all hip-
pocampal formation subfields (Fig. 3). In MTLE specimens,increased NGF expression was detected in hypertrophic hilarcells and pyramidal neurons in the Ammon horn, subicularcomplex, and entorhinal cortex. Glia-like cells were alsoevident in the MTLE Sommer sector (CA1 and prosubiculum;Fig. 3K, M). Nerve growth factor immunoreactivity in thegranular layer (Fig. 4AYD) was increased in MTLE andMTLE + D groups versus the control group (Fig. 4F). Amongepileptic groups, specimens from the MTLE + D groupshowed greater NGF immunoreactivity, and specimens fromthe MTLE + P group had less immunoreactivity (Fig. 4AYC).Other hippocampal subfields with decreased NGF immuno-reactivity in the MTLE + P group were the hilus, CA2, CA1,prosubiculum, subiculum, and parasubiculum (Fig. 4F).
Nerve growth factor immunoreactivity in CA3 of epi-leptic patients correlated positively with granular cell disper-sion in the superior (r = +0.47, p = 0.027) and inferior blade(r = +0.45, p = 0.036), indicating that the increased trophictone from CA3 to the dentate gyrus might support the rever-beratory excitatory activity between CA3, mossy cells, andgranule cells, and possibly reinforced by MFS (49). More-over, NGF immunoreactivity in the granular layer corre-lated with MFS gray value (r = +0.54, p = 0.008) and length(r = +0.51, p = 0.014) and with seizure frequency (r = +0.40,p = 0.016). Nerve growth factor immunoreactivity correlatedpositively with neuronal density in CA1 (r = +0.39, p = 0.038)and prosubiculum (r = +0.42, p = 0.042). In addition, NGFimmunoreactivity in the granular layer (r = +0.44, p = 0.008),subiculum (r = +0.34, p = 0.047), and parasubiculum(r = +0.47, p = 0.009) correlated positively with global IQ andwith neuropsychologic nonverbal scores in the same re-gions (granular layer NGF: r = +0.44, p = 0.011; subiculum
NGF: r = +0.42, p = 0.017; parasubiculum NGF: r = +0.58,p = 0.001). No differences in NGF immunoreactivity wereseen in MTLE + D patients taking fluoxetine versus those nottaking fluoxetine. The MTLE + P patients taking haloperidolshowed less NGF immunoreactivity in parasubiculum(2,022.31 T 410.6" 102 Km2 vs 3,303.65 T 138.2" 102 Km2,t10 = j2.85, p = 0.019). Moreover, when we analyzed NGFexpression considering seizure type, we found that patientswith secondarily generalized seizures exhibited decreasedvalues versus those with CPS in the hilus (1,640.12 T 693.4 "102 Km2 vs 2,249.85 T 816.2 " 102 Km2, t37 = 2.20, p = 0.03),CA2 (2,482.40 T 747.9 " 102 Km2 vs 3,316.06 T 1,194.1 "102 Km2, t34 = 2.17, p = 0.04), and subiculum (2,571.59 T
FIGURE 4. Nerve growth factor (NGF) expression in mesialtemporal lobe epilepsy (MTLE) specimens with and withoutpsychiatric comorbidities and in nonepileptic controls. (AYD)MTLE + major depression (MTLE + D) granular layer (B)exhibited greater NGF expression versus MTLE (A), MTLE +psychosis (MTLE + P) (C), and control (D) groups. (E) NGF-immunoreactive area values were higher in epileptic groupsversus controls (** p G 0.001; * p G 0.05), and there were dif-ferences among MTLE groups (## p G 0.001; # p G 0.05). InCA2 and CA1, there was a trend (tr: 0.05 Q p e 0.07) to de-creased NGF expression in MTLE + D versus MTLE. Values fromMTLE (black bars), MTLE + D (gray bars), MTLE + P (light graybars), and from nonepileptic controls (white bars) are indicatedas mean T SD. Scale bar = (AYD) 50 Km.
J Neuropathol Exp Neurol ! Volume 72, Number 11, November 2013 NTs in MTLE With Psychiatric Comorbidities
! 2013 American Association of Neuropathologists, Inc. 1035
Copyright © 2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

666.7" 102 Km2 vs 3,694.60 T 1,190.3" 102 Km2, t38 = 3.52,p = 0.02).
Brain-derived neurotrophic factor immunoreactivity inneurons was detected in all subfields of the hippocampalformation (Fig. 5). Glia-like cells were also immunostained,particularly in MTLE dentate gyrus and Ammon horn (Fig.5AYK). Comparing all groups, the granular layer was thesubfield with major differences among them (Fig. 6AYD).The MTLE and MTLE + D showed increased BDNF-immunoreactive area versus that of the control group (Fig. 6E);and MTLE + P granular layer BDNF immunoreactivity was
less than that of MTLE (Fig. 6E). In the hilus, BDNFin MTLE + P was less than that of controls, and in CA1,greater BDNF immunoreactivity area was seen for MTLEand MTLE + D (Fig. 6E). Brain-derived neurotrophic factorimmunoreactivity correlated positively with neuronal densityof epileptic patients in the hilus (r = +0.65, p = 0.001), CA3(r = +0.56, p = 0.045), and subiculum (r = +0.41, p = 0.035),but not with MFS or with dispersion in the granular layer.Brain-derived neurotrophic factor immunoreactivity in thegranular layer (r = +0.36, p = 0.037) and subiculum (r = +0.42,p = 0.022) also correlated positively with global IQ, but not
FIGURE 5. Brain-derived neurotrophic factor (BDNF) expression in the human hippocampal formation. (AYT) There is greaterimmunoreactivity in mesial temporal lobe epilepsy (MTLE) granular cells and in cells with glial profiles, for example, in dentate gyrusand Ammon horn (arrows, A, C, K, M). ENT, entorhinal cortex; GL, granular layer; HIL, hilus; PAR, parasubiculum; PRO,prosubiculum; SUB, subiculum. Scale bar = (AYT) 50 Km.
Kandratavicius et al J Neuropathol Exp Neurol ! Volume 72, Number 11, November 2013
! 2013 American Association of Neuropathologists, Inc.1036
Copyright © 2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

with other neuropsychologic scores. Brain-derived neurotrophicfactor immunoreactivity correlated inversely with age of seizureonset in the hilus (r = j0.40, p = 0.032) and CA3 (r = j0.51,p = 0.020). Regarding MTLE + D patients taking fluoxetine,greater BDNF immunoreactivity in CA1 was seen when com-pared with those not taking fluoxetine (3,920.77 T 161.8 " 102
Km2 vs 2,529.46 T 451.4" 102 Km2, t9 = +4.11, p = 0.009). Incontrast, MTLE + P patients taking haloperidol showeddecreased BDNF immunoreactivity in the hilus (1,038.60 T440.6" 102 Km2 vs 1,840.75 T 207.3 " 102 Km2, t11 =j2.96,p = 0.016) and CA4 (1,451.76 T 638.9" 102 Km2 vs 3,100.54 T284.0 " 102 Km2, t11 = j4.22, p = 0.002) versus those nottaking haloperidol.
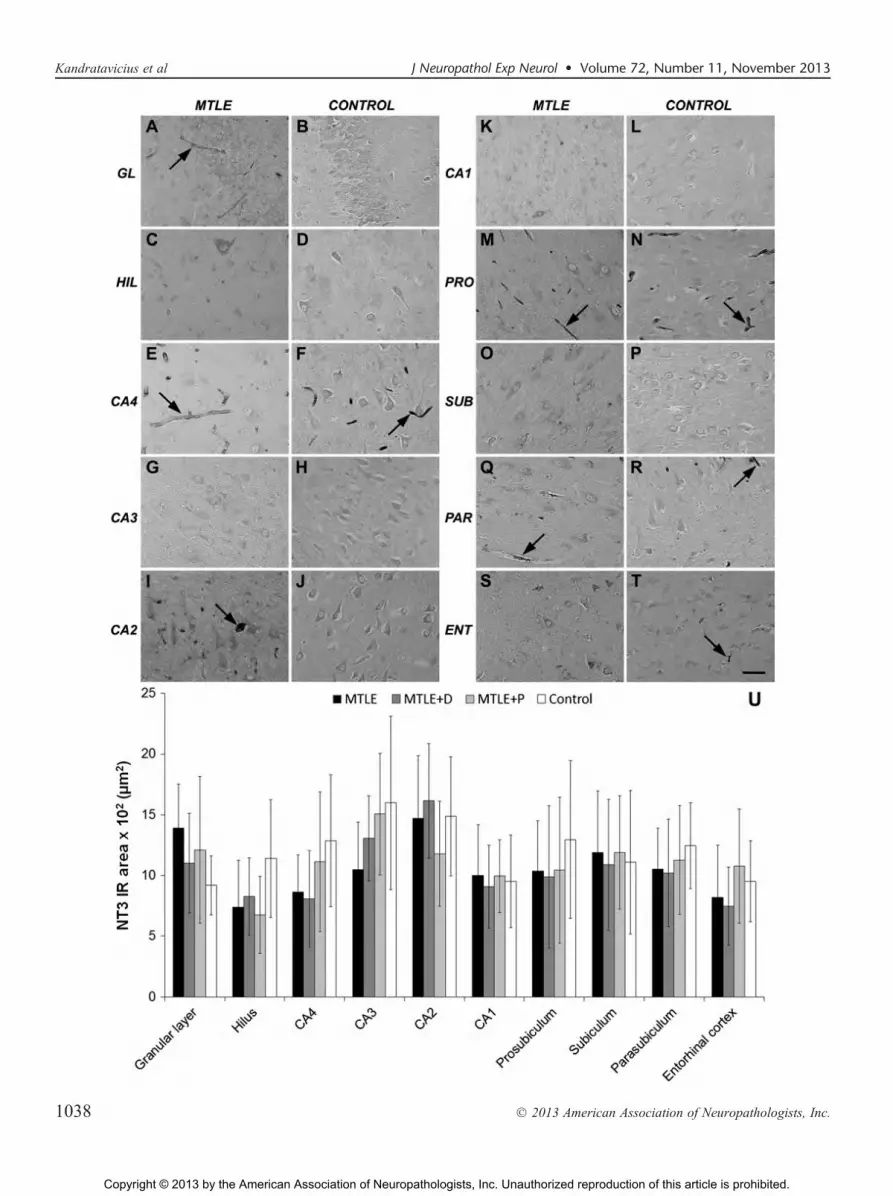
Neurotrophin 3 expression was detected in neuronsand glia, as well as in blood vessels through all hippocampalformation subfields (Fig. 7AYT). No differences in NT3-immunoreactive area (Fig. 7U) or in the qualitative assess-ment of how strong NT3 staining was in blood vessels (datanot shown) were found among groups in any subfield. Neu-rotrophin 3 and MFS correlated inversely in CA4 (r = j0.64,p = 0.003), CA1 (r =j0.58, p = 0.005), subiculum (r =j0.52,p = 0.008), and entorhinal cortex (r = j0.59, p = 0.013). Nodifferences in NT3 expression between patients taking or notfluoxetine or haloperidol were seen.
Given the differences in neuronal densities (Fig. 1),an important question was whether the statistical differencesin NT immunoreactivity for the 4 groups could be accountedfor by changes in neuronal densities. Therefore, we performedan analysis of covariance comparing groups and neuronaldensities with NGF, BDNF, and NT3-immunoractive area.Differential BDNF expression in the hilus, where a positivecorrelation was seen between neuronal density and BDNFexpression, lost its statistical significance after neuron countcorrection. Differences between groups in other subfieldsand for other NTs remained significant after cell count cor-rection, indicating that the 4 patient categories showed sig-nificant differences in NT expression levels that were notinfluenced by changes in neuronal densities.
DISCUSSIONAn extensive number of neuropathologic studies have
been done on hippocampi from MTLE patients, whereas fewerhave examined extrahippocampal tissue (50). Postmortemstudies with specimens from patients with schizophrenia andmajor depression have also shown several abnormalities in thehippocampal formation (12, 13). Despite the strong associationbetween epilepsy and psychiatric comorbidities, neuropatho-logic data are scant. In another series of patients, we previouslyreported decreased MFS in MTLE patients with psychosis andincreased MFS in MTLE patients with major depression (4);this result was confirmed in the present series. Another im-portant replicated finding was decreased neuronal density inthe entorhinal cortex of MTLE patients with psychiatriccomorbidities (4). More severe neuron loss in the entorhinalcortex would contribute to disrupted communication betweenthe hippocampal formation and neocortical and limbic sites,which are known to have a profound effect in modulation ofthe psychopathologic state (12).
NTs, Epilepsy, and Psychopathologic StatesNeurotrophins are part of a set of molecules that can
influence MFS development. Indeed, a positive correlation be-tween MFS progression and increased NGF concentration,but not increased BDNF concentration, was demonstrated inthe kainate model of epilepsy (22), which is similar to thecorrelations we found in this MTLE series. In particular, astudy on transgenic mice suggests that BDNF is not essential toMFS because hippocampal slices of a BDNF-deficient (-/-)mouse display MFS as well as the wild type (51). With res-pect to NT3, a negative correlation with MFS was found, in
FIGURE 6. Brain-derived neurotrophic factor (BDNF) expres-sion in mesial temporal lobe epilepsy (MTLE) specimens withand without psychiatric comorbidities and in nonepilepticcontrols. (AYD) In MTLE (A) and MTLE + major depression(MTLE + D) (B), there is greater BDNF expression in the gran-ular layer versus MTLE + psychosis (MTLE + P) (C) and control(D) groups. (E) BDNF-immunoreactive area values are higherin the dentate gyrus and CA1 of epileptic groups and less in thehilus of MTLE + P versus controls (** p G 0.001; * p G 0.05). Theonly subfield with differences among epileptic groups was thegranular layer, where MTLE + P specimens showed decreasedBDNF expression versus MTLE without psychiatriccomorbidities (# p G 0.05). Values from MTLE (black bars),MTLE + D (gray bars), MTLE + P (light gray bars), and fromnonepileptic controls (white bars) are indicated as mean T SD.Scale bar = (AYD) 50 Km.
J Neuropathol Exp Neurol ! Volume 72, Number 11, November 2013 NTs in MTLE With Psychiatric Comorbidities
! 2013 American Association of Neuropathologists, Inc. 1037
Copyright © 2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
Kandratavicius et al J Neuropathol Exp Neurol ! Volume 72, Number 11, November 2013
! 2013 American Association of Neuropathologists, Inc.1038
Copyright © 2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

contrast to animal model results (22), and measurements ofNT3 mRNA in the granular layer in epileptic patients (52). Ithas been suggested that NT3 can trigger sprouting and in-hibit epileptogenesis (15), and we hypothesize that, in humanMTLE, where a seizure-prone circuit is already established,decreased NT3 input to the dentate gyrus (e.g. from the ento-rhinal cortex) would contribute in the chronic phase to sustainhyperexcitability and to halt further progression of sprouting.Indeed, we found a positive correlation between seizure fre-quency, MFS, and granular layer NGF, in agreement withearlier work in animal models (53, 54). Neurotrophin 3 ex-pression in non-neuronal tissue such as blood vessels has beenverified by others, and one of the proposed mechanisms wouldresult in increased nitric oxide production via endothelial nitricoxide synthase (55). Although increased neuronal nitric oxideproduction in MTLE may be inferred (56), we did not iden-tify qualitative differences in NT3 expression in hippo-campal blood vessels between control and epileptic patients,suggesting that NT3-mediated endothelial nitric oxide produc-tion is unaltered.
The association between seizure facilitation and increasedNGF expression has been shown in animal models of epilepsy(14, 53) and is in agreement with our result of positive corre-lation between seizure frequency and NGF expression in thegranular layer. On the other hand, we found that patients withsecondarily generalized seizures exhibited decreased NGF ex-pression in the hilus, CA2, and subiculum versus patientswith CPS. In a recent study, Alapirtti et al (57) comparedblood samples of 3 TLE patients with secondarily generalizedseizures and 11 patients with CPS, suggesting that the moresevere the seizure type (i.e. secondarily generalized seizures),the stronger the inflammatory response after an acute seizure. Inour series, a molecule related to seizure facilitation was founddownregulated in patients with more severe seizures;however,when NGF is not able to interact with their receptors, seizuredevelopment is halted (58). Inasmuch as we found distincthippocampal subfields related to seizure frequency and seizuretype, differential modulation and action of NGF are feasible.Future studies with NTs receptors will be able to clarify thisfinding.
Despite NGF upregulation found in most hippocampalformation subfields of patients with epilepsy, specimens withMTLE and psychosis showed decreased NGF immunoreactiv-ity in the granular layer, hilus, CA2, CA1, and all subicularcomplex when compared with MTLE without psychiatriccomorbidities. An equivalent result was seen for BDNF im-munoreactivity in the granular layer. There is a significant andstable association between schizophrenia and memory impair-ment (59), although its basis is unknown (60). Most studiescomprise verbal memory deficits, but poor performance ofauditory memory tasks and visuospatial delayed recognitionhave also been reported in patients with schizophrenia and their
relatives (61) and in healthy subjects injected with ketamine(62). Diminished hippocampal NGF and BDNF immunoreac-tivity might be related to the lower cognitive performance trendseen in the MTLE + P group because NGF and BDNF, butnot NT3 (63), are required for optimal cholinergic neurotrans-mission (64). In turn, low cholinergic function results in poormemory performance, as seen in schizophrenia and Alzheimerdisease (65, 66). In accordance with our findings of positivecorrelations between NGF immunoreactivity and IQ and non-verbal memory scores, it has been shown in rodents that in-creases in hippocampal and neocortical NGF reverse deficits inlearning and memory in spatial navigation and object recog-nition tasks (67). More importantly, in conditions of gluta-matergic hypofunction (as it occurs in schizophrenia andanimal models of schizophrenia based on N-methyl-d-aspartateantagonism [68Y70]), there is a decrease in BDNF expression(71), as we found in the MTLE + P group versus the MTLEgroup. Likewise, rats injected with ketamine show decreasedMFS induced by electroconvulsive seizures and decreasedBDNF expression (72). Low BDNF levels have been observedin the plasma of schizophrenic patients (25, 73, 74) and seemindependent of medication despite the high variability betweenthe studies (75). Other studies have also shown associationbetween BDNF polymorphisms and schizophrenia and cogni-tive deficits (76Y79), but not in cases of febrile seizures (80)or TLE (81). Decreased BDNF expression has been describedin postmortem hippocampus of schizophrenic patients (24, 27),and, in contrast, there are also reports of increased BDNF ex-pression (82, 83). The 2 latter studies speculate that the re-sults would be related to a deficient BDNF secretion leading tointraneuronal BDNF accumulation. In view of reciprocalBDNF/glutamate modulation (84), normalization of BDNFlevels in psychosis would counteract glutamatergic hypo-function. Based on the increased BDNF expression found inepilepsy, a comorbid psychotic state would display mildersymptoms than in schizophrenia, as is indeed depicted ininterictal psychosis (9).
Drugs and NT ExpressionDose-dependent fluoxetine-induced neuroprotection has
been described in animal models of epilepsy, ischemia, andParkinson disease (85Y87). In our series, we did not observedifferences in neuronal density but found increased BDNFimmunoreactivity in CA1, the same region reported asprotected, and with increased BDNF levels in fluoxetine-treated ischemic gerbils (85). A similar result has also beenfound in the human dentate gyrus (88). Interestingly, the CA1region is one of the most affected subfields by neuronal loss inMTLE. In our series, most BDNF immunoreactivity detectedin CA1 was in glial cells, and it has been shown that astrocytesare particularly affected by fluoxetine and paroxetine (but nottricyclics) treatment and respond with BDNF upregulation (89).
FIGURE 7. Neurotrophin 3 (NT3) expression in mesial temporal lobe epilepsy (MTLE) specimens with and without psychiatriccomorbidities and in nonepileptic controls. (AYT) There was faint to moderate NT3 expression in neuronal and glial cells and faintto strong staining in blood vessels (arrows) in all groups and subfields of the hippocampal formation. (U) No differences in NT3-immunoreactive area were found among groups. ENT, entorhinal cortex; GL, granular layer; HIL, hilus; PAR, parasubiculum; PRO,prosubiculum; SUB, subiculum. Scale bar = (AYT) 50 Km.
J Neuropathol Exp Neurol ! Volume 72, Number 11, November 2013 NTs in MTLE With Psychiatric Comorbidities
! 2013 American Association of Neuropathologists, Inc. 1039
Copyright © 2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

The effects of typical and atypical antipsychotics on cell pro-liferation and apoptosis have been investigated repeatedly, withcontroversial results. Haloperidol treatment in animal modelshas been shown to increase hippocampal neurogenesis (90), orto have no effect (91), to promote survival of hippocampalstem cells (92) and to induce apoptosis in cortical neurons (93).In our series, significant neuronal loss was seen in CA3 andsubiculum of MTLE + P patients taking haloperidol versusthose not taking haloperidol in the same group, in agreementwith evidence that antipsychotics can reduce brain tissue vol-ume (94). Our results also showed decreased BDNF and NGFhippocampal expression in MTLE + P patients taking halo-peridol, in accordance with animal model studies showing de-creased BDNF hippocampal levels (95) and NGF corticallevels (96).
CONCLUSIONSIt is important to acknowledge some limitations inherent
to our findings in this study. Even with a relatively small samplesize, the present results suggest that further studies exploringMTLE and related comorbidities are worthwhile. Although wecould not perform stereologic counts because of limited tissuesource from surgery, our neuron density numbers are in agree-ment with recent hippocampal stereologic counts performed inMTLE specimens (97). In fact, because all MTLE surgicalspecimens were freshly collected and submitted to identicalprocessing, differences among them are particularly relevant.
In summary, the present results provide the first demon-stration of differential NT expression in human MTLE hippo-campal formation with and without psychiatric comorbidities,supporting the close association between MFS and NTs. It alsoindicates that the use of haloperidol in MTLE might relate toincreased neuronal loss and decreased NT expression. Our re-sults are in agreement with most studies done with postmortemmajor depression, schizophrenia specimens, and animal modelsthat have independently provided important pathophysiologichallmarks. The relatively high prevalence of psychiatric symp-toms in MTLE patients suggests mechanisms and/or substratesshared in these conditions. Clearly, different psychopathologicstates in MTLE rely on distinct structural and neurochemicalmilieu. In the static concept of chronic MTLE, we are not ableto define which exact variable might contribute to the genesis orto the maintenance of a particular psychiatric comorbidity, butit is hoped that future research on the morphologic and bio-chemical abnormalities in this scenario will delineate the mol-ecules that may become targets for new treatments.
ACKNOWLEDGMENTSThe authors thank Renata Caldo Scandiuzzi, Renato
Meirelles e Silva, and Jose Eduardo Peixoto-Santos for theexcellent technical support in neo-Timm histochemistry, pano-ramic NeuN image acquisition, and Western blot, respectively.
REFERENCES1. Mathern GW, Babb TL, Pretorius JK, et al. Reactive synaptogenesis and
neuron densities for neuropeptide Y, somatostatin, and glutamate decar-boxylase immunoreactivity in the epileptogenic human fascia dentata.J Neurosci 1995;15:3990Y4004
2. TL, Kupfer WR, Pretorius JK, et al. Synaptic reorganization by mossyfibers in human epileptic fascia dentata. Neuroscience 1991;42:351Y63
3. Babb TL, Brown WJ, Pretorius J, et al. Temporal lobe volumetric celldensities in temporal lobe epilepsy. Epilepsia 1984;25:729Y40
4. Kandratavicius L, Hallak JE, Young LT, et al. Differential aberrantsprouting in temporal lobe epilepsy with psychiatric co-morbidities.Psychiatry Res 2012;195:144Y50
5. Kandratavicius L, Rosa-Neto P, Monteiro MR, et al. Distinct increasedmetabotropic glutamate receptor type 5 (mGluR5) in temporal lobeepilepsy with and without hippocampal sclerosis. Hippocampus 2013:Epub Ahead of Print; doi: 10.1002/hipo.22160
6. Sundram F, Cannon M, Doherty CP, et al. Neuroanatomical correlatesof psychosis in temporal lobe epilepsy: Voxel-based morphometry study.Br J Psychiatry 2010;197:482Y92
7. Taylor DC. Factors influencing the occurrence of schizophrenia-likepsychosis in patients with temporal lobe epilepsy. Psychol Med 1975;5:249Y54
8. Roberts GW, Done DJ, Bruton C, et al. A ‘‘mock up’’ of schizophrenia:Temporal lobe epilepsy and schizophrenia-like psychosis. Biol Psychia-try 1990;28:127Y43
9. Beard AW, Slater E. The schizophrenic-like psychoses of epilepsy. ProcR Soc Med 1962;55:311Y16
10. Kanner AM. Is major depression a neurologic disorder with psychiatricsymptoms? Epilepsy Behav 2004;5:636Y44
11. Craddock N, Owen MJ. Molecular genetics and the relationship be-tween epilepsy and psychosis. Br J Psychiatry 2010;197:75Y76
12. Kandratavicius L, Lopes-Aguiar C, Bueno-Junior LS, et al. Psychiatriccomorbidities in temporal lobe epilepsy: Possible relationships betweenpsychotic disorders and involvement of limbic circuits. Rev Bras Psiq2012;34:454Y66
13. Kandratavicius L, Ruggiero RN, Hallak JE, et al. Pathophysiologyof mood disorders in temporal lobe epilepsy. Rev Bras Psiq 2012;34:S233Y59
14. Adams B, Sazgar M, Osehobo P, et al. Nerve growth factor accele-rates seizure development, enhances mossy fiber sprouting, and attenu-ates seizure-induced decreases in neuronal density in the kindling modelof epilepsy. J Neurosci 1997;17:5288Y96
15. Xu B,Michalski B, Racine RJ, et al. Continuous infusion of neurotrophin-3triggers sprouting, decreases the levels of TrkA and TrkC, and inhi-bits epileptogenesis and activity-dependent axonal growth in adult rats.Neuroscience 2002;115:1295Y308
16. Scharfman HE, Goodman JH, Sollas AL. Actions of brain-derivedneurotrophic factor in slices from rats with spontaneous seizuresand mossy fiber sprouting in the dentate gyrus. J Neurosci 1999;19:5619Y31
17. Mathern GW, Babb TL, Micevych PE, et al. Granule cell mRNAlevels for BDNF, NGF, and NT-3 correlate with neuron losses orsupragranular mossy fiber sprouting in the chronically damaged andepileptic human hippocampus. Mol Chem Neuropathol 1997;30:53Y76
18. Isackson PJ, Huntsman MM, Murray KD, et al. BDNF mRNA expres-sion is increased in adult rat forebrain after limbic seizures: Tem-poral patterns of induction distinct from NGF. Neuron 1991;6:937Y48
19. Binder DK, Croll SD, Gall CM, et al. BDNF and epilepsy: Too muchof a good thing? Trends Neurosci 2001;24:47Y53
20. Kokaia Z, Kelly ME, Elmer E, et al. Seizure-induced differential ex-pression of messenger RNAs for neurotrophins and their receptorsin genetically fast and slow kindling rats. Neuroscience 1996;75:197Y207
21. Vezzani A, Ravizza T, Moneta D, et al. Brain-derived neurotro-phic factor immunoreactivity in the limbic system of rats after acuteseizures and during spontaneous convulsions: Temporal evolutionof changes as compared to neuropeptide Y. Neuroscience 1999;90:1445Y61
22. Shetty AK, Zaman V, Shetty GA. Hippocampal neurotrophin levelsin a kainate model of temporal lobe epilepsy: A lack of correla-tion between brain-derived neurotrophic factor content and progres-sion of aberrant dentate mossy fiber sprouting. J Neurochem 2003;87:147Y59
23. Shimizu E, Hashimoto K, Okamura N, et al. Alterations of serumlevels of brain-derived neurotrophic factor (BDNF) in depressed patientswith or without antidepressants. Biol Psychiatry 2003;54:70Y75
24. Durany N, Michel T, Zochling R, et al. Brain-derived neurotrophicfactor and neurotrophin 3 in schizophrenic psychoses. Schizophr Res2001;52:79Y86
Kandratavicius et al J Neuropathol Exp Neurol ! Volume 72, Number 11, November 2013
! 2013 American Association of Neuropathologists, Inc.1040
Copyright © 2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

25. Toyooka K, Asama K, Watanabe Y, et al. Decreased levels ofbrain-derived neurotrophic factor in serum of chronic schizophrenic pa-tients. Psychiatry Res 2002;110:249Y57
26. Vargas HE, Gama CS, Andreazza AC, et al. Decreased serum neurotro-phin 3 in chronically medicated schizophrenic males. Neurosci Lett 2008;440:197Y201
27. Knable MB, Barci BM, Webster MJ, et al. Molecular abnormalitiesof the hippocampus in severe psychiatric illness: Postmortem findingsfrom the Stanley Neuropathology Consortium. Mol Psychiatry 2004;9:609Y20
28. Pedrini M, Chendo I, Grande I, et al. Serum brain-derived neuro-trophic factor and clozapine daily dose in patients with schizophrenia:A positive correlation. Neurosci Lett 2011;491:207Y10
29. Gama CS, Andreazza AC, Kunz M, et al. Serum levels of brain-derivedneurotrophic factor in patients with schizophrenia and bipolar disorder.Neurosci Lett 2007;420:45Y48
30. Balu DT, Hoshaw BA, Malberg JE, et al. Differential regulation ofcentral BDNF protein levels by antidepressant and non-antidepressantdrug treatments. Brain Res 2008;1211:37Y43
31. Valvassori SS, Stertz L, Andreazza AC, et al. Lack of effect of antipsy-chotics on BNDF and NGF levels in hippocampus of Wistar rats. MetabBrain Dis 2008;23:213Y19
32. Gittins R, Harrison PJ. Neuronal density, size and shape in the humananterior cingulate cortex: A comparison of Nissl and NeuN staining.Brain Res Bull 2004;63:155Y60
33. Stan AD, Ghose S, Gao XM, et al. Human postmortem tissue: Whatquality markers matter? Brain Res 2006;1123:1Y11
34. Blumcke I, Coras R, Miyata H, et al. Defining clinico-neuropathologicalsubtypes of mesial temporal lobe epilepsy with hippocampal sclerosis.Brain Pathol 2012;22:402Y11
35. Berg AT. Identification of pharmacoresistant epilepsy. Neurol Clin 2009;27:1003Y13
36. Engel J Jr. Surgery for seizures. N Engl J Med 1996;334:647Y5237. Sachdev P. Schizophrenia-like psychosis and epilepsy: The status of
the association. Am J Psychiatry 1998;155:325Y3638. Zochodne DW, Cheng C. Neurotrophins and other growth factors in
the regenerative milieu of proximal nerve stump tips. J Anat 2000;196:279Y83
39. Bechade C, Mallecourt C, Sedel F, et al. Motoneuron-derivedneurotrophin-3 is a survival factor for PAX2-expressing spinal in-terneurons. J Neurosci 2002;22:8779Y84
40. Calinescu AA, Liu T, Wang MM, et al. Transsynaptic activityYdependentregulation of axon branching and neurotrophin expression in vivo.J Neurosci 2011;31:12708Y15
41. Lorente de No R. Studies on the structure of the cerebral cortex II.Continuation of study of the ammonic system. J Psychol Neurol 1934;46:113Y77
42. Abercrombie M. Estimation of nuclear population from microtome sec-tions. Anat Rec 1946;94:239Y47
43. Mathern GW, Adelson PD, Cahan LD, et al. Hippocampal neuron da-mage in human epilepsy: Meyer’s hypothesis revisited. Prog Brain Res2002;135:237Y51
44. Mathern GW, Pretorius JK, Mendoza D, et al. Hippocampal N-methyl-d-aspartate receptor subunit mRNA levels in temporal lobe epilepsy pa-tients. Ann Neurol 1999;46:343Y58
45. Mathern GW, Mendoza D, Lozada A, et al. Hippocampal GABA andglutamate transporter immunoreactivity in patients with temporal lobeepilepsy. Neurology 1999;52:453Y72
46. Mathern GW, Pretorius JK, Kornblum HI, et al. Human hippocampalAMPA and NMDA mRNA levels in temporal lobe epilepsy patients.Brain 1997;120:1937Y59
47. Mathern GW, Babb TL, Vickrey BG, et al. The clinical-pathogenicmechanisms of hippocampal neuron loss and surgical outcomes in tem-poral lobe epilepsy. Brain 1995;118:105Y18
48. Peixoto-Santos JE, Galvis-Alonso OY, Velasco TR, et al. Increasedmetallothionein I/II expression in patients with temporal lobe epilepsy.Plos One 2012;7:e44709
49. Scharfman HE. The CA3 ‘‘backprojection’’ to the dentate gyrus. ProgBrain Res 2007;163:627Y37
50. Thom M. Hippocampal sclerosis: Progress since Sommer. Brain Pathol2009;19:565Y72
51. Bender R, Heimrich B, Meyer M, et al. Hippocampal mossy fibersprouting is not impaired in brain-derived neurotrophic factor-deficientmice. Exp Brain Res 1998;120:399Y402
52. Mathern GW, Babb TL, Leite JP, et al. The pathogenic and progres-sive features of chronic human hippocampal epilepsy. Epilepsy Res1996;26:151Y61
53. Gall CM, Isackson PJ. Limbic seizures increase neuronal produc-tion of messenger RNA for nerve growth factor. Science 1989;245:758Y61
54. Rocamora N, Palacios JM, Mengod G. Limbic seizures induce a differ-ential regulation of the expression of nerve growth factor, brain-derivedneurotrophic factor and neurotrophin-3, in the rat hippocampus. BrainRes Mol Brain Res 1992;13:27Y33
55. Meuchel LW, Thompson MA, Cassivi SD, et al. Neurotrophins inducenitric oxide generation in human pulmonary artery endothelial cells.Cardiovasc Res 2011;91:668Y76
56. Leite JP, Chimelli L, Terra-Bustamante VC, et al. Loss and sproutingof nitric oxide synthase neurons in the human epileptic hippocampus.Epilepsia 2002;43(Suppl 5):235Y42
57. Alapirtti T, Waris M, Fallah M, et al. C-reactive protein and seizuresin focal epilepsy: A video-electroencephalographic study. Epilepsia2012;53:790Y96
58. Rashid K, Van der Zee CE, Ross GM, et al. A nerve growth fac-tor peptide retards seizure development and inhibits neuronal sprout-ing in a rat model of epilepsy. Proc Natl Acad Sci U S A 1995;92:9495Y99
59. Aleman A, Hijman R, de Haan EH, et al. Memory impairment inschizophrenia: A meta-analysis. Am J Psychiatry 1999;156:1358Y66
60. Harrison PJ. The neuropathology of schizophrenia. A critical review ofthe data and their interpretation. Brain 1999;122(Pt 4):593Y624
61. Robles O, Blaxton T, Adami H, et al. Nonverbal delayed recognition inthe relatives of schizophrenia patients with or without schizophreniaspectrum. Biol Psychiatry 2008;63:498Y504
62. Newcomer JW, Farber NB, Jevtovic-Todorovic V, et al. Ketamine-induced NMDA receptor hypofunction as a model of memory impair-ment and psychosis. Neuropsychopharmacology 1999;20:106Y18
63. da Penha Berzaghi M, Cooper J, Castren E, et al. Cholinergic regulationof brain-derived neurotrophic factor (BDNF) and nerve growth factor(NGF) but not neurotrophin-3 (NT-3) mRNA levels in the developingrat hippocampus. J Neurosci 1993;13:3818Y26
64. Knipper M, da Penha Berzaghi M, Blochl A, et al. Positive feedbackbetween acetylcholine and the neurotrophins nerve growth factor andbrain-derived neurotrophic factor in the rat hippocampus. Eur J Neurosci1994;6:668Y71
65. Buckingham SD, Jones AK, Brown LA, et al. Nicotinic acetylcholinereceptor signalling: Roles in Alzheimer’s disease and amyloid neuro-protection. Pharmacol Rev 2009;61:39Y61
66. D’Hoedt D, Bertrand D. Nicotinic acetylcholine receptors: An over-view on drug discovery. Expert Opin Ther Targets 2009;13:395Y411
67. Rispoli V, Marra R, Costa N, et al. Choline pivaloyl ester enhancesbrain expression of both nerve growth factor and high-affinity receptorTrkA, and reverses memory and cognitive deficits, in rats with excito-toxic lesion of nucleus basalis magnocellularis. Behav Brain Res 2008;190:22Y32
68. Coyle JT. Glutamate and schizophrenia: Beyond the dopamine hypoth-esis. Cell Mol Neurobiol 2006;26:365Y84
69. Javitt DC, Zukin SR. Recent advances in the phencyclidine model ofschizophrenia. Am J Psychiatry 1991;148:1301Y8
70. Rujescu D, Bender A, Keck M, et al. A pharmacological model forpsychosis based on N-methyl-d-aspartate receptor hypofunction: Molec-ular, cellular, functional and behavioral abnormalities. Biol Psychiatry2006;59:721Y29
71. Castren E, da Penha Berzaghi M, Lindholm D, et al. Differential effectsof MK-801 on brain-derived neurotrophic factor mRNA levels in differ-ent regions of the rat brain. Exp Neurol 1993;122:244Y52
72. Chen AC, Shin KH, Duman RS, et al. ECS-Induced mossy fiber sprou-ting and BDNF expression are attenuated by ketamine pretreatment.J Ect 2001;17:27Y32
73. Grillo RW, Ottoni GL, Leke R, et al. Reduced serum BDNF levelsin schizophrenic patients on clozapine or typical antipsychotics. J PsychiatrRes 2007;41:31Y35
J Neuropathol Exp Neurol ! Volume 72, Number 11, November 2013 NTs in MTLE With Psychiatric Comorbidities
! 2013 American Association of Neuropathologists, Inc. 1041
Copyright © 2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

74. Pirildar S, Gonul AS, Taneli F, et al. Low serum levels of brain-derivedneurotrophic factor in patients with schizophrenia do not elevate afterantipsychotic treatment. Prog Neuropsychopharmacol Biol Psychiatry2004;28:709Y13
75. Green MJ, Matheson SL, Shepherd A, et al. Brain-derived neuro-trophic factor levels in schizophrenia: A systematic review withmeta-analysis. Mol Psychiatry 2011;16:960Y72
76. Egan MF, Kojima M, Callicott JH, et al. The BDNF val66met poly-morphism affects activity-dependent secretion of BDNF and humanmemory and hippocampal function. Cell 2003;112:257Y69
77. Numata S, Ueno S, Iga J, et al. Brain-derived neurotrophic factor (BDNF)Val66Met polymorphism in schizophrenia is associated with age atonset and symptoms. Neurosci Lett 2006;401:1Y5
78. Gratacos M, Gonzalez JR, Mercader JM, et al. Brain-derived neuro-trophic factor Val66Met and psychiatric disorders: Meta-analysis ofcase-control studies confirm association to substance-related disorders,eating disorders, and schizophrenia. Biol Psychiatry 2007;61:911Y22
79. Forlenza OV, Diniz BS, Teixeira AL, et al. Effect of brain-derivedneurotrophic factor Val66Met polymorphism and serum levels on theprogression of mild cognitive impairment. World J Biol Psychiatry 2010;11:774Y80
80. Chou IC, Tsai CH, Lee CC, et al. Brain-derived neurotrophic factor(BDNF) Val66Met polymorphisms in febrile seizures. Epilepsy Res2004;60:27Y29
81. Bragatti JA, Schenkel LC, Torres CM, et al. No major clinical impactof Val66Met BDNF gene polymorphism on temporal lobe epilepsy.Epilepsy Res 2010;88:108Y11
82. Iritani S, Niizato K, Nawa H, et al. Immunohistochemical study ofbrain-derived neurotrophic factor and its receptor, TrkB, in the hippo-campal formation of schizophrenic brains. Prog NeuropsychopharmacolBiol Psychiatry 2003;27:801Y7
83. Takahashi M, Shirakawa O, Toyooka K, et al. Abnormal expres-sion of brain-derived neurotrophic factor and its receptor in thecorticolimbic system of schizophrenic patients. Mol Psychiatry 2000;5:293Y300
84. Mattson MP. Glutamate and neurotrophic factors in neuronal plasticityand disease. Ann N Y Acad Sci 2008;1144:97Y112
85. Kim do H, Li H, Yoo KY, et al. Effects of fluoxetine on ischemiccells and expressions in BDNF and some antioxidants in the gerbil
hippocampal CA1 region induced by transient ischemia. Exp Neurol2007;204:748Y58
86. Jin Y, Lim CM, Kim SW, et al. Fluoxetine attenuates kainic acidYinducedneuronal cell death in the mouse hippocampus. Brain Res 2009;1281:108Y16
87. Chung YC, Kim SR, Park JY, et al. Fluoxetine prevents MPTP-inducedloss of dopaminergic neurons by inhibiting microglial activation. Neu-ropharmacology 2011;60:963Y74
88. Chen B, Dowlatshahi D, MacQueen GM, et al. Increased hippo-campal BDNF immunoreactivity in subjects treated with antidepres-sant medication. Biol Psychiatry 2001;50:260Y65
89. Allaman I, Fiumelli H, Magistretti PJ, et al. Fluoxetine regulates theexpression of neurotrophic/growth factors and glucose metabolism inastrocytes. Psychopharmacology (Berl) 2011;216:75Y84
90. Dawirs RR, Hildebrandt K, Teuchert-Noodt G. Adult treatment withhaloperidol increases dentate granule cell proliferation in the gerbilhippocampus. J Neural Transm 1998;105:317Y27
91. Halim ND, Weickert CS, McClintock BW, et al. Effects of chronichaloperidol and clozapine treatment on neurogenesis in the adult rathippocampus. Neuropsychopharmacology 2004;29:1063Y69
92. Keilhoff G, Grecksch G, Bernstein HG, et al. Risperidone and haloperi-dol promote survival of stem cells in the rat hippocampus. EurArch Psychiatry Clin Neurosci 2010;260:151Y62
93. Pillai A, Dhandapani KM, Pillai BA, et al. Erythropoietin preventshaloperidol treatment-induced neuronal apoptosis through regulationof BDNF. Neuropsychopharmacology 2008;33:1942Y51
94. Moncrieff J, Leo J. A systematic review of the effects of antipsychoticdrugs on brain volume. Psychol Med 2010;40:1409Y22
95. Angelucci F, Mathe AA, Aloe L. Brain-derived neurotrophic factorand tyrosine kinase receptor TrkB in rat brain are significantly alteredafter haloperidol and risperidone administration. J Neurosci Res 2000;60:783Y94
96. Terry AV Jr, Gearhart DA, Warner S, et al. Protracted effects ofchronic oral haloperidol and risperidone on nerve growth factor, cho-linergic neurons, and spatial reference learning in rats. Neuroscience2007;150:413Y24
97. Alonso-Nanclares L, Kastanauskaite A, Rodriguez JR, et al. A stereo-logical study of synapse number in the epileptic human hippocampus.Front Neuroanat 2011;5:8
Kandratavicius et al J Neuropathol Exp Neurol ! Volume 72, Number 11, November 2013
! 2013 American Association of Neuropathologists, Inc.1042
Copyright © 2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.





























