Embed Size (px)
Citation preview

PII S0361-9230(00)00455-X
Neurochemical phenotype of sympathetic nervous
system outflow from brain to white fat
Haifei Shi and Timothy J. Bartness*
Department of Biology, Neurobiology and Behavior Program, Georgia State University, Atlanta, GA, USA
[Received 3 July 2000; Revised 20 September 2000; Accepted 29 November 2000]
ABSTRACT: The sympathetic innervation of white adipose tis-sue (WAT) appears to be a dominant mechanism triggeringlipolysis. The purpose of this study was to determine the neu-rochemical phenotype of neurons comprising the sympatheticoutflow from brain to WAT. This was accomplished by injectingSiberian hamster WAT with a viral retrograde transneuronaltract tracer, the pseudorabies virus (PRV), in combination withimmunocytochemical characterization of several neurotrans-mitters or their synthetic enzymes in the brain. Catecholamin-ergic (tyrosine hydroxylase [TH] and dopamine-b-hydroxylase[DBH] immunoreactivity) and peptidergic (arginine vasopressin[AVP] and oxytocin [OXY] immunoreactivity) neurons were partof this outflow, but the percentage of double-labeled cells wassmall, consistent with previous studies. Brainstem PRV 1 TH-or PRV 1 DBH-labeled cells were in previously identified nor-adrenergic areas (A5, A6, and subcoeruleus, rostroventrolateralmedulla [RVL], some reticular nuclei). Forebrain double labelingwas greatest in the paraventricular (TH, AVP, OXY) and supra-chiasmatic (AVP) nuclei, both implicated in the central controlof lipolysis. Differences between the PRV double labeling re-ported here for WAT versus that of other sympathetic peripheraltargets were PRV 1 DBH in A5 and RVL, and PRV 1 TH in RVLand in the lateral paragigantocellular and lateral reticular nuclei.Collectively, these results begin to identify the neurochemicalidentity of the sympathetic outflow from brain to WAT. © 2001Elsevier Science Inc.
KEY WORDS: Siberian hamsters, Tyrosine hydroxylase, Dopa-mine beta-hydroxylase, Arginine vasopressin, Oxytocin, Pseu-dorabies virus, Tract tracing, Suprachiasmatic nucleus, Para-ventricular nucleus, Brainstem, Lipolysis.
INTRODUCTION
Energy is primarily stored as lipid in white adipose tissue (WAT)by mammals. When circulating or stored metabolic fuels cannotsatisfy energy needs, lipid is mobilized from WAT through theprocess of lipolysis. Briefly, catecholamines (i.e., norepinephrineand epinephrine) bind to membrane-bound adrenergic receptors onfat cells (adipocytes) triggering the lipolytic cascade that ulti-mately results in the breakdown of triglycerides into free fattyacids and glycerol [17]. It now appears that a dominant means bywhich WAT lipolysis is triggered is by its sympathetic nervoussystem (SNS) innervation, rather than solely by adrenal medullary-released catecholamines (for review see: [3,4]). Thus, adrenal
demedullation of laboratory rats does not block lipid mobilizationdue to cold exposure [14], 2-deoxy-D-glucose-induced glucoseutilization blockade [26], or electrical stimulation of the medialhypothalamus [25], as measured by circulating concentration offree fatty acids or glycerol. In contrast, SNS denervation of WATblocks fasting-induced lipolysis [6].We previously defined the SNS postganglionic innervation of
epididymal and inguinal WAT (EWAT and IWAT, respectively)in Siberian hamsters (Phodopus sungorus) and laboratory ratsusing anterograde and retrograde fluorescent tract tracers [28].Furthermore, we defined the central nervous system (CNS) originsof the SNS outflow from brain to WAT recently using a viral trans-neuronal retrograde tract tracer, the pseudorabies virus (PRV; [1]).Specifically, injections of PRV into EWAT and IWAT labeled ahierarchical chain of functionally connected neurons at sites acrossthe neuroaxis (i.e., spinal cord, brainstem, midbrain, and forebrain;[1]). In addition, the pattern of labeled neurons after injection ofPRV into IWAT and EWAT is more similar than different at allbut the pre- and postganglionic levels of the neuroaxis [1,28].Although the general SNS outflow from brain to WAT is
known, we do not know the neurochemical phenotype of thechains of neurons that comprise these projections. Once identified,receptor agonists or antagonists can be applied at the appropriatepoints in the outflow circuit to affect the sympathetic drive onWAT experimentally and thus stimulate or inhibit lipolysis. There-fore, the purpose of the present study was to begin to define theneurochemical phenotypes of the neurons comprising the sympatheticoutflow from brain to WAT. This was accomplished by using thePRV technique to trace the SNS outflow from brain to WAT com-bined with immunocytochemical labeling of potential neurotransmit-ters or their enzymes of synthesis in Siberian hamsters.
MATERIALS AND METHODS
Animals
Thirty-two adult male Siberian hamsters were obtained fromour breeding colony. Our colony of hamsters was established in1988, supplied with the stock provided by Dr. Bruce Goldman(University of Connecticut) that originated from stock in thecolony of Dr. Klaus Hoffman (Clinical Research Unit for Repro-ductive Medicine, Germany). In 1990, our colony was interbredwith second-generation wild-trapped hamsters provided by Dr.
* Address for correspondence: Dr. Timothy J. Bartness, Department of Biology, Georgia State University, 24 Peachtree Center Rd NE, Atlanta, GA30303-3083, USA. Fax: 11-(404)-651-2509; E-mail: [email protected]
Brain Research Bulletin, Vol. 54, No. 4, pp. 375–385, 2001Copyright © 2001 Elsevier Science Inc.Printed in the USA. All rights reserved
0361-9230/01/$–see front matter
375

Katherine Wynne-Edwards (Queen’s University). Finally, the col-ony was interbred with the hamsters supplied by Dr. G. RobertLynch (University of Colorado) in 1995. These latter hamsterswere derived from the original Hoffman colony, but had beenisolated for about 20 years. The adult hamsters were housed ingroups of 10 to 12 in polyvinyl cages (48 3 27 3 15 cm) untilused in the present experiment and kept in a long summer-like day(light/dark cycle, 16h:8h, with lights on at 0300 h). Animals weregiven Purina Rodent Chow (#5001) and water ad libitum. Allexperimental procedures were approved by Georgia State Univer-sity Institutional Animal Care and Use Committee in accordancewith Public Health Service guidelines.
Virus Injections and Tissue Harvesting
Siberian hamsters were anesthetized with sodium pentobarbital(50 mg/kg, intraperitoneal [i.p.]). The IWAT or EWAT pads of theright side were exposed and injected with the attenuated strainBartha’s K strain of the PRV (courtesy of Dr. Teryl Frey, GeorgiaState University). PRV (120 nl of 1.0 3 108 plaque forming unitsper milliliter) was injected into five areas (600 nl total) across thelength of the IWAT (n 5 13) or the EWAT (n 5 19) pads. Allinjections were done in the morning between 1000 and 1200 h.After the PRV injections, the animals were singly housed inpolyvinyl cages (23 3 26 3 20 cm).No colchicine treatment was given to these animals to enhance
cellular staining because neurotransmitter or synthetic enzymeimmunocytochemical staining can be obtained in hamsters withoutit [5,19,27]. We previously found infected neurons in the rostralhypothalamus and other rostral forebrain areas on the sixth dayafter PRV injections into the IWAT and EWAT pads [1]; there-fore, all hamsters were sacrificed 6 days after injection in thepresent study. The animals were deeply anesthetized with sodiumpentobarbital (80 mg/kg, i.p.) and perfused transcardially with 100ml of 0.9% NaCl and then with 150 ml of 4% paraformaldehydesolution. All perfusions were done between 1000 and 1200 h.
Immunocytochemical Procedures
Brains were removed and suspended in 4% paraformaldehydesolution overnight, and then incubated in 4% paraformaldehydecontaining 25% sucrose solution for 24 h. Brain slices (40 mm) weremade in the coronal plane using a freezing microtome and were keptin 0.1 M sodium phosphate-buffered saline (PBS) containing 0.1%sodium azide at 4°C until processed for immunocytochemistry.Brainstem sections were screened initially for viral infec-
tions. If infected, then the remaining brain and spinal cord weresectioned coronally and processed fully. Double-immunostain-ing methods were used to colocalize PRV with neurotransmit-ters (arginine vasopressin [AVP], oxytocin [OXY], somatosta-tin [SS], substance P [SP], neuropeptide Y [NPY], corticotropinreleasing hormone [CRH], and hypocretin [HC]), or neurotrans-mitter synthetic enzymes (tyrosine hydroxylase [TH], dopa-mine-b-hydroxylase [DBH], phenylethanolamine-N-methyl-transferase [PNMT]). TH and SP immunoreactivity wereanalyzed throughout the whole brain; DBH, PNMT, and NPYimmunoreactivity were analyzed at the levels of the brainstemand midbrain, whereas AVP, OXY, SS, CRH, and HC immu-noreactivity were analyzed only at the level of the hypothala-mus. Fluorescein isothiocyanate (FITC) was used to visualizePRV-labeled neurons, and rhodamine was used to visualize TH,AVP, OXY, and NPY immunoreactivity. 3,39-diaminobenzi-dine tetrahydrochloride (DAB) was used to visualize DBH,PNMT, SS, SP, CRH, and HC immunoreactivity because theneed for increased sensitivity could not be met using fluores-cence markers. Briefly, all brain sections were incubated with
the PRV primary antibody (1:10,000) and a neurotransmitter ora synthetic enzyme primary antibody in 0.1 M PBS containing2% normal goat serum and 0.3% Triton X-100 overnight atroom temperature. The primary antibodies for TH, DBH,PNMT, AVP, OXY, NPY, SS, and SP were obtained commer-cially (Incstar, Stillwater, MN, USA: mouse anti-TH [1:1,000],rabbit anti-DBH [1:1,000], rabbit anti-PNMT [1:1,000], rabbitanti-AVP [1:5,000], rabbit anti-OXY [1:2,000], rabbit anti-NPY [1:2,000], and rabbit anti-SS [1:1,000]; Accurate Chem.,Westbury, NY, USA: rabbit anti-SP [1:1,000]). The primaryantibodies for CRH and HC were gifts (rabbit anti-CRH[1:2,000] from Dr. Jean Rivier, Salk Institute, La Jolla, CA,USA; and rabbit anti-HC [1:5,000] from Dr. Anthony van denPol, Yale University, New Haven, CT, USA). Primary antibodyspecificity was tested by using antisera-free serum for eachneurotransmitter or enzyme in some sections. In addition, othersections were pre-incubated with the same antigen used togenerate each neurotransmitter or synthetic enzyme antibodyexcept CRH and HC. Note that both tests of antibody specificity(i.e., incubation without the primary antibody and pre-adsorp-tion of the primary antibody with its antigen) yield no immu-noreactivity.After the incubation with the PRV primary antibody, the
brain sections were rinsed with 0.02 M PBS three times, andthen were incubated with a goat anti-pig FITC-conjugated sec-ondary antibody and a secondary antibody for 2 h at roomtemperature. The secondary antibodies were diluted to 1:200 in0.1 M PBS solution with 2% normal goat serum and werepurchased commercially (Jackson ImmunoResearch Laborato-ries, West Grove, PA, USA: FITC-conjugated to goat anti-pigfor PRV; rhodamine-conjugated to goat anti-mouse for TH;rhodamine-conjugated to goat anti-rabbit for AVP, OXY, andNPY; biotinylated goat anti-rabbit for DBH, PNMT, SS, SP,CRH, and HC). The sections for PRV, and TH, AVP, OXY andNPY double labeling were then rinsed three times with 0.02 MPBS and were mounted onto slides. The sections for PRV, andDBH, PNMT, SS, SP, CRH, and HC double-labeling wereincubated in a 1:200 solution of the avidin-biotin horseradishperoxidase (HRP) complex (Vectastain ABC Elite Kit; VectorLaboratories, Burlingame, CA, USA) for 1 h at room temper-ature. The HRP was made visible with DAB and 6% H2O2.After drying in the dark at room temperature, all slides werecovered with 0.1 M n-propyl gallete (3,4,5-trihydroxybenzoicacid n-propyl ester; Sigma Chemical Co., St. Louis, MO, USA)in 1:1 Tris buffer-glycerol solution for preservation of FITCand rhodamine immunofluorescence. Fluorescent staining wasvisualized using fluorescence microscopy and the DAB chroma-gen product was visualized using light microscopy.
Histological Quantification of Labeled Neurons
The exact position of PRV-, neurotransmitters or their syn-thetic enzymes-, and double-labeled neurons on each brainsection was localized and quantified using computer-assistedimage analysis (Image Tracer software; Translational Technol-ogy, Phoenix, AZ, USA) and a stage mounted position trans-ducer system (MD3 Microscope Digitizer; Minnesota Datamet-rics Corporation, Shoreview, MN, USA). Briefly, according toour previously published methodology [1,2], camera lucidapictures of brain sections stained with crestyl violet were drawnand scanned into the computer. There were 13 forebrain sec-tions with bregma measurements from 0.20 mm to 24.30 mm;7 midbrain sections with bregma measurements from 25.20mm to 28.80 mm; and 15 brainstem sections with bregmameasurements from 210.04 mm to 214.30 mm; all bregma
376 SHI AND BARTNESS

measurements are relative to a standard rat stereotaxic atlas[18]. A digital image of each drawing was projected onto thecomputer screen with the Image Tracer program. The image ofeach brain section was registered with the same section on themicroscope stage using the stage transducers. For all 13 fore-
brain sections, 7 midbrain sections, and 15 brainstem sectionsfrom all animals, the positions of PRV-labeled neurons andneurotransmitter or its synthetic enzyme 1 PRV-labeled neu-rons were visualized with the microscope and marked in theexact position (60.5 mm) on the computerized image of that
TABLE 1
QUANTITATIVE SUMMARY OF NEUROTRANSMITTER/ENZYME AND VIRALLY INFECTED NEURONS IN BRAINSTEM, MIDBRAIN AND FOREBRAIN
No. of PRV-Labeled Cells No. of TH 1 PRV Cells % TH 1 PRV Cells
Brainstem
RVL 52.61 6 12.01 2.72 6 0.83 5.756 1.43
LPGi 52.00 6 13.86 1.39 6 0.64 2.116 0.77
LRt 15.72 6 4.90 0.83 6 0.33 3.436 1.40
PCRt 9.56 6 2.75 0.506 0.26 6.256 3.30
Sol 49.61 6 15.29 0.226 0.13 0.626 0.50
A5 6.726 1.15 2.616 0.87 32.196 8.44
Facial nucleus 13.066 2.91 2.50 6 0.83 21.196 7.15
LC 16.28 6 4.40 0.67 6 0.45 7.296 5.55
SubC 3.61 6 0.90 1.396 0.41 38.526 9.51
No. of PRV-Labeled Cells No. of DBH 1 PRV Cells % DBH 1 PRV Cells
RVL 17.28 6 4.65 0.22 6 0.10 2.136 1.42
LPGi 23.78 6 4.97 0.17 6 0.09 0.886 0.55
Gi 6.44 6 2.05 0.176 0.09 6.026 3.42
Sol 88.17 6 19.99 0.61 6 0.36 1.426 1.07
A5 12.67 6 2.78 1.286 0.39 24.586 9.00
LC 5.286 1.49 0.066 0.06 2.786 2.78
MVe 27.116 8.79 0.22 6 0.17 1.496 1.08
No. of PRV-Labeled Cells No. of TH 1 PRV Cells % TH 1 PRV Cells
Midbrain
DR 4.67 6 2.40 0.176 0.12 0.586 0.40
PnO 9.17 6 4.08 2.286 1.33 15.756 6.60
LDTg 4.22 6 1.94 0.396 0.20 6.846 4.43
PO/SO 5.11 6 1.88 1.446 0.62 17.556 6.80
No. of PRV-Labeled Cells No. of TH 1 PRV Cells % TH 1 PRV Cells
Forebrain
PVN 27.11 6 8.44 0.61 6 0.22 2.626 1.13
LH 7.72 6 2.25 0.066 0.06 0.226 0.22
DA 7.67 6 4.75 0.336 0.28 1.036 0.75
DMN 2.00 6 1.03 0.336 0.20 14.816 8.18
PH 0.72 6 0.32 0.286 0.18 14.446 8.05
MPA 16.00 6 4.32 0.11 6 0.08 1.176 0.94
ZI 3.78 6 1.29 0.336 0.16 5.286 2.96
No. of PRV-Labeled Cells No. of AVP 1 PRV Cells % AVP 1 PRV Cells
PVN 35.61 6 7.54 0.44 6 0.25 1.076 0.56
MPA 15.89 6 3.54 0.72 6 0.48 2.656 1.56
SCN 3.78 6 1.26 0.226 0.17 1.856 1.27
No. of PRV-Labeled Cells No. of OXY 1 PRV Cells % OXY 1 PRV Cells
PVN 44.11 6 11.65 0.89 6 0.28 3.496 1.23
DA 4.94 6 1.64 0.066 0.06 0.376 0.37
PRV, pseudorabies virus; TH, tyrosine hydroxylase; RVL, rostroventrolateral reticular nucleus; LPGi, lateral paragigantocellular nucleus; LRt, lateralreticular nucleus; PCRt, parvocellular reticular nucleus; Sol, nucleus of the solitary tract; A5, A5 noradrenaline cell group; LC, locus coeruleus; SubC,subcoeruleus; DBH, dopamine b-hydroxylase; Gi, gigantocellular reticular nucleus; MVe, medial vestibular nucleus; DR, dorsal raphe nucleus; PnO,pontine reticular nucleus; LDTg, laterodorsal tegmental nucleus; PO/SO, superior olive and periolivary nucleus; PVN, paraventricular nucleus; LH, lateralhypothalamus; DA, dorsal hypothalamic area; DMN, dorsomedial hypothalamic nucleus; PH, posterior hypothalamic nucleus; MPA, medial preoptic area;ZI, zona incerta; SCN, suprachiasmatic nucleus.
NEUROCHEMICAL CODING OF SNS OUTFLOW 377

section. The number of marked neurotransmitter 1 PRV orsynthetic enzyme 1 PRV neurons was counted in each nucleusthroughout its complete rostral-caudal extent for each animal.Quantification and distribution of single-labeled neurons forthese neurotransmitters or their synthetic enzymes are reportedelsewhere [21].
Statistical Analysis
A quantitative analysis of the colocalization of PRV and theneurotransmitters or synthetic enzymes was made. The percentageof PRV infected neurons showing neurotransmitter or syntheticenzyme immunoreactivity was analyzed with a two-way analysis
of variance (WAT pad 3 brain area; SigmaStat 2.03, JandelScientific, San Rafael, CA, USA) within each of the three basicdivisions of the neuroaxis (i.e., brainstem, midbrain, and fore-brain).
RESULTS
One IWAT- and one EWAT-injected hamster died duringthe 6-day interval after the PRV injection. There was no sign ofsickness in the surviving animals. Of the 12 surviving IWAT-injected animals, 9 became infected by the PRV, and of the 18surviving EWAT-injected animals, 12 were infected. The spinalcord of each infected animal was examined for lysis of the
FIG. 1. Schematic diagrams showing the location of tyrosine hydroxylase1 pseudorabies virus(PRV) cells in the sections of the brainstem of Siberian hamsters injected with PRV intoepididymal or inguinal white adipose tissue (data are combined). Bregma measurements are at theupper right corner of each section. Each ● represents one double-labeled cell. Abbreviations: CC,central canal; C1, C1 adrenaline cell group; LC, locus coeruleus; LPGi, lateral paragigantocel-lular nucleus; LRt, lateral reticular nucleus; MVe, medial vestibular nucleus; PCRt, parvocellularreticular nucleus; RVL, rostroventrolateral reticular nucleus; Sol, nucleus of the solitary tract;SubC, subcoeruleus nucleus; 4V, fourth ventricle.
378 SHI AND BARTNESS

infected neurons. In addition, spread of the virus to the adjacentmusculature was assessed by the presence of alpha motor neu-ron infections in the ventral horn of the spinal cord. Finally,non-controlled spread of the virus was assessed by the presenceof infections contralateral to the injection side in the spinalcord. Cases with lysis, alpha motor neuron labeling or contralat-eral spinal cord infections were excluded from the study. Nei-ther lysis nor alpha motor neuron labeling was seen in any ofthe infected animals. No contralateral spinal cord infectionswere detected in the IWAT-injected animals, but three EWAT-injected animals had contralateral infections and the data fromthese animals were discarded. The remaining EWAT (n 5 9)and all IWAT (n 5 9) cases had infected neurons located in awell-defined cluster in the intermediolateral cell group and thecentral autonomic nucleus of the spinal cord ipsilateral to theinjection site (data not shown). All subsequent analyses weredone only on these specifically infected hamsters.There were no double-labeled neurons for SP, NPY, CRH,
and HC after PRV injection into IWAT or EWAT. Immunocy-tochemical staining of PNMT and SS was not intense enoughfor confident colocalization with PRV-infected neurons. There-fore, we only report on the colocalization of TH-, DBH-, AVP-and OXY-labeled neurons with colocalization of PRV. Overall,
there were no significant differences for the percentage ofneurons double-labeled for TH1, DBH1, AVP1, orOXY1PRV at all levels of neuroaxis between EWAT- andIWAT-injected hamsters. Therefore, these data were combinedfor the schematic representations of the co-localized neuronsand the tables that follow.
Brainstem
Both TH1 and DBH1 PRV cells were found at all levels ofthe brainstem, and their staining patterns closely paralleled oneanother. In the more caudal sections of the brainstem (bregma 5
213.68 mm to 211.80 mm), TH1 PRV cells were found in thelateral paragigantocellular nucleus (LPGi), lateral reticular nucleus(LRt), rostroventrolateral reticular nucleus (RVL), parvocellularreticular nucleus (PCRt), as well as in the nucleus of the solitarytract (Sol; Table 1; Fig. 1A–D). DBH1 PRV cells were seen inthe RVL, LPGi, the gigantocellular reticular nucleus (Gi), and theSol (Table 1; Fig. 2A–B). At the level of the facial nucleus(bregma 5 211.30 mm to 210.04 mm), the majority of theTH 1 PRV and DBH 1 PRV cells were found in the A5 noradren-aline cell group (Fig. 3). TH1 PRV cells also were found in thefacial nucleus. In these more rostral sections of the brainstem, a
FIG. 2. Schematic diagrams showing the location of dopamine-b-hydroxylase1 pseudorabiesvirus (PRV) cells in the sections of the brainstem of Siberian hamsters injected with PRV intoepididymal or inguinal white adipose tissue (data are combined). Bregma measurements areat the upper right corner of each section. Each ● represents one double-labeled cell. Abbre-viations: A5, noradrenergic area 5; C1, C1 adrenaline cell group; Gi, gigantocellular reticularnucleus; LPGi, lateral paragigantocellular nucleus; MVe, medial vestibular nucleus; ROb,raphe obscurus nucleus; RVL, rostroventrolateral reticular nucleus; Sol, nucleus of the solitarytract; 7n, 7th nerve (facial nerve).
NEUROCHEMICAL CODING OF SNS OUTFLOW 379
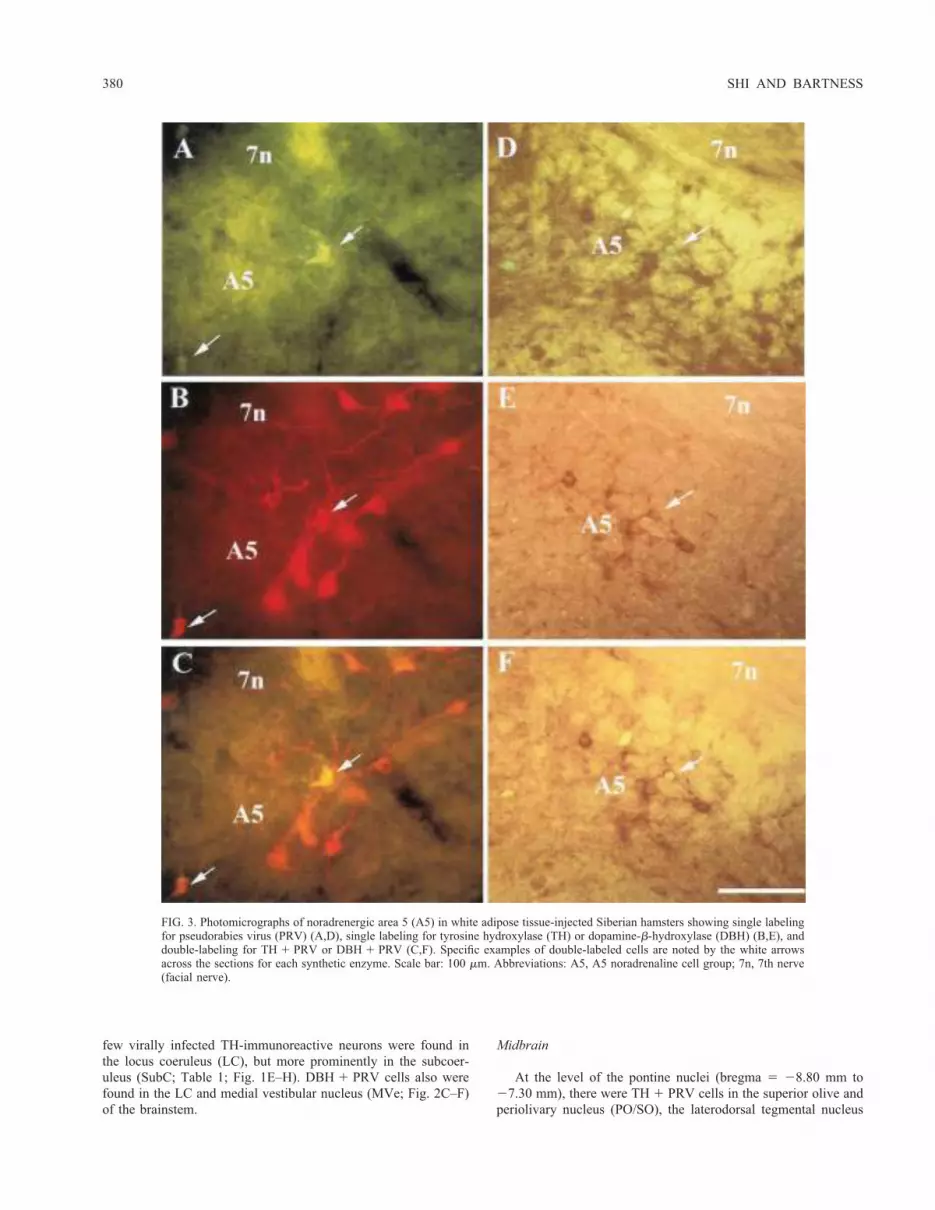
few virally infected TH-immunoreactive neurons were found inthe locus coeruleus (LC), but more prominently in the subcoer-uleus (SubC; Table 1; Fig. 1E–H). DBH1 PRV cells also werefound in the LC and medial vestibular nucleus (MVe; Fig. 2C–F)of the brainstem.
Midbrain
At the level of the pontine nuclei (bregma 5 28.80 mm to27.30 mm), there were TH1 PRV cells in the superior olive andperiolivary nucleus (PO/SO), the laterodorsal tegmental nucleus
FIG. 3. Photomicrographs of noradrenergic area 5 (A5) in white adipose tissue-injected Siberian hamsters showing single labelingfor pseudorabies virus (PRV) (A,D), single labeling for tyrosine hydroxylase (TH) or dopamine-b-hydroxylase (DBH) (B,E), anddouble-labeling for TH 1 PRV or DBH 1 PRV (C,F). Specific examples of double-labeled cells are noted by the white arrowsacross the sections for each synthetic enzyme. Scale bar: 100 mm. Abbreviations: A5, A5 noradrenaline cell group; 7n, 7th nerve(facial nerve).
380 SHI AND BARTNESS

FIG. 4. Schematic diagrams showing the location of tyrosine hydroxylase 1 pseudorabies virus(PRV) cells in the midbrain (A–C) and forebrain (D–G) of Siberian hamsters injected with PRVinto epididymal or inguinal white adipose tissue (data are combined). Bregma measurements are atthe upper right corner of each section. Each ● represents one double-labeled cell. Abbreviations:Aq, aqueduct of sylvius; CG, central gray; DA, dorsal hypothalamic area; DMN, dorsomedialhypothalamic nucleus; DR, dorsal raphe nucleus; f, fornix; LDTg, laterodorsal tegmental nucleus;MPA, medial preoptic area; opt, optic tract; PH, posterior hypothalamic nucleus; PnO, pontinereticular nucleus; PO/SO, superior olive and periolivary nucleus; PVN, paraventricular nucleus;SCN, suprachiasmatic nucleus; VMH, ventromedial hypothalamic nucleus; ZI, zona incerta; 3V,third ventricle.
NEUROCHEMICAL CODING OF SNS OUTFLOW 381

(LDTg), dorsal raphe nucleus (DR), and in the oral part of thepontine reticular nucleus (PnO; Table 1; Fig. 4A–C). There wereno DBH1 PRV cells in the midbrain.
Forebrain
The neurons showing both TH and PRV immunoreactivity were inseveral regions of the hypothalamus, including the medial preopticarea (MPA), the paraventricular nucleus (PVN), the lateral hypothal-amus (LH), dorsal hypothalamic area (DA), dorsomedial hypotha-lamic nucleus (DMN), and the posterior hypothalamic nucleus (PH).TH1 PRV cells also were in nonhypothalamic forebrain structuressuch as the zona incerta (ZI; Table 1; Fig. 4D–G). In the PVN,TH1 PRV cells only were in the medial parvocellular portion.
Finally, there were double-labeled neurons in the medialparvocellular region of the PVN for both AVP and OXY (Table1; Fig. 5). In addition, the AVP 1 PRV cells were in the MPAand in the dorsomedial aspect of the suprachiasmatic nucleus(SCN; Table 1; Fig. 6A–C). The OXY 1 PRV cells also were inthe DA (Table 1; Fig. 6D,E).
DISCUSSION
We previously defined the origins of the SNS outflow frombrain to WAT using the PRV transneuronal tract tracing technique[1]. In the present study, we combined the viral labeling of theSNS outflow to EWAT and to IWAT with immunocytochemicalcharacterization of the neurochemical phenotype of some of these
FIG. 5. Photomicrographs of paraventricular nucleus in white adipose tissue-injected Siberian hamsters showing single labeling forpseudorabies virus (PRV) (A,D), single labeling for oxytocin (OXY) or arginine vasopressin (AVP) (B,E), and double-labeling forOXY 1 PRV or AVP 1 PRV (C,F). Specific examples of double-labeled cells are noted by the white arrows across the sections for eachneuropeptide. Scale bar: 100 mm. Abbreviation: 3V, third ventricle.
382 SHI AND BARTNESS

neurons across the neuroaxis. Of the neurotransmitters and en-zymes of synthesis tested here, we found evidence of cat-echolaminergic neurons in this circuitry (TH and DBH, but notPNMT immunoreactivity), as well as some peptidergic neurons(i.e., AVP, OXY immunoreactivity, but not SS, SP, NPY, CRH, orHC immunoreactivity). Consistent with previous studies, the ab-solute numbers of neurotransmitters or synthetic enzymes1 PRVcells represented a small number of the total number of infectedcells (ranging from 0 to ;40% of all PRV-labeled cells—seebelow for a discussion of what may underlie this relatively lowlevel of double-labeling). Finally, there were no statistically sig-nificant differences in the pattern of double-labeled cells afterIWAT or EWAT virus injections; therefore the results will bediscussed more generally without reference to the injected fat pads.
The double labeling of TH and DBH containing cells withPRV in the brainstem was most notable in previously identifiedpopulations of noradrenergic neurons such as the A5 and A6(locus coeruleus) noradrenergic cell groups, as well as withinthe subcoeruleus area [12,23]. In addition to these well-estab-lished brainstem components of the SNS outflow to peripheraltissues, other sympathetic-related bulbar sites had significantcatecholaminergic-viral labeling. These included the RVL,LPGi, and some reticular nuclei (i.e., LRt, PCRt), as indicatedby TH-labeling. Of the catecholaminergic double-labeled cellsin the brainstem, the A5 and SubC sites showed more doublelabeling than the other nuclei. There also were cells that clearlywere noradrenergic (i.e., DBH-labeled neurons) in these areas,including the Gi and Sol nuclei, and the A5 region. In addition
FIG. 6. Schematic diagrams showing the location of arginine vasopressin (AVP) 1 pseudorabies virus(PRV) (A–C) and oxytocin (OXY)1 PRV (D,E) cells in the forebrain of Siberian hamsters injected withPRV into epididymal or inguinal white adipose tissue (data are combined). Bregma measurements are atthe upper right corner of each section. Each ● represents one double-labeled cell. Abbreviations: DA,dorsal hypothalamic area; LV, lateral ventricle; MPA, medial preoptic area; MPO, medial preopticnucleus; PVN, paraventricular nucleus; SCN, suprachiasmatic nucleus; SON, supraoptic nucleus; ZI, zonaincerta; 3V, third ventricle.
NEUROCHEMICAL CODING OF SNS OUTFLOW 383

to their apparent role in the SNS innervation of WAT, thesecatecholaminergic bulbar sites are also part of the sympatheticoutflow circuitry that regulates cardiovascular [8] and thermo-genic [15,16] functions and they also have been described usingthe PRV tract tracing method [2,13].Of the several forebrain structures showing both viral label-
ing and neurotransmitter or synthetic enzyme immunoreactiv-ity, three seem noteworthy. First, previous studies have de-scribed monosynaptic descending premotor vasopressinergicand oxytocinergic projections from the PVN to sympatheticpreganglionics in the intermediolateral horn of the spinal cord[10,20]. Therefore, it is likely that many of the cells labeledwith AVP 1 PRV and OXY 1 PRV after virus injections intoWAT are part of these chemically defined projections. Indeed,it seems more than a coincidence that there is approximatelythree times more OXY-labeled than AVP-labeled neurons pro-jecting to the spinal cord [20] and that we found nearly the samerelative difference in the percentage of OXY 1 PRV toAVP 1 PRV double-labeled cells. Second, the MPA appears toplay a significant role in lipid mobilization (for review see: [4]),possessing a noradrenergic circuit that triggers lipid mobiliza-tion when activated by the cold [7] that seems likely to includesome or all of the TH 1 PRV cells we observed here. Third, theSCN also showed AVP 1 PRV cells in the present study, andthese cells likely play a role in the central control of peripherallipolysis (for review see: [3,4]), given that coronal microknife
cuts just caudal to the SCN block lipid mobilization resultingfrom glucoprivation, fasting and exercise [9,26].We have compared the results of our neurochemical pheno-
typing of the SNS outflow from brain to WAT with the resultsof similar neurochemical phenotyping studies of the sympa-thetic outflow from brain to the adrenal gland [24], stellateganglion (heart; [13]), pancreas [11], or wall of the tail artery[22]; Table 2) in rats. There are several notable differencesbetween the results of these studies and the present investiga-tion. Siberian hamsters have DBH 1 PRV in the A5 and in theRVL, but rats do not. Siberian hamsters have TH 1 PRV inseveral brainstem nuclei (e.g., RVL, LPGi, LRt, and facialnucleus), but rats do not. Whether these differences representalterations in the neurochemical circuitry specific to these sym-pathetic target tissues and/or to species tested is not known.The relatively small number of double-labeled neurons in
the present study is consistent with the other PRV studiesattempting to chemically code the SNS projections from thebrain to peripheral tissues discussed above [11,13,22,24]. Onepossibility for the low number of double-labeled neurons maybe that infection by the virus inhibits neurotransmitters or theirsynthetic enzymes.Collectively, the results of the present study begin to identify
the neurochemical identity of the CNS origins of the SNS outflowfrom brain to WAT specifically, and should lead toward a deeperunderstanding of the sympathetic projections to the peripherygenerally.
ACKNOWLEDGEMENTS
This research was supported, in part, by National Institute of MentalHealth Research Scientist Development Award KO2-MH00841 and Na-tional Institute Health Research Grant RO1-DK35254 to TJB.
REFERENCES
1. Bamshad, M.; Aoki, V. T.; Adkison, M. G.; Warren, W. S.; Bartness,T. J. Central nervous system origins of the sympathetic nervous systemoutflow to white adipose tissue. Am. J. Physiol. 275:R291–R299;1998.
2. Bamshad, M.; Song, C. K.; Bartness, T. J. CNS origins of the sym-pathetic nervous system outflow to brown adipose tissue. Am. J.Physiol. 276:R1569–R1578; 1999.
3. Bartness, T. J.; Bamshad, M. Innervation of mammalian white adiposetissue: Implications for the regulation of total body fat. Am. J. Physiol.275:R1399–R1411; 1998.
4. Bartness, T. J.; Demas, G. E.; Song, C. K. Central nervous systeminnervation of white adipose tissue. In: Klaus, S., ed. Adipose tissue.Georgetown, TX: Landes Bioscience; 2001:201–229.
5. Bittman, E. L.; Bartness, T. J.; Goldman, B. D.; DeVries, G. J.Suprachiasmatic and paraventricular control of photoperiodism in Si-berian hamsters. Am. J. Physiol. 260:R90–R101; 1991.
6. Cantu, R. C.; Goodman, H. M. Effects of denervation and fasting onwhite adipose tissue. Am. J. Physiol. 212:207–212; 1967.
7. Coimbra, C. C.; Migliorini, R. H. Cold-induced free fatty acid mobi-lization is impaired in rats with lesions in preoptic area. Neurosci. Lett.88:1–5; 1988.
8. Yang, Z.; Coote, J. H. Influence of the hypothalamic paraventricularnucleus on cardiovascular neurones in the rostral ventrolateral medullaof the rat. J. Physiol. (Lond.) 513:521–530; 1999.
9. Gross, J.; Migliorini, R. H. Further evidence for a central regulation offree fatty acid mobilization in the rat. Am. J. Physiol. 232:E165–E171;1977.
10. Hallbeck, M.; Bolmqvist, A. Spinal cord-projecting vasopressinergicneurons in the rat paraventricular hypothalamus. J. Comp. Neurol.411:201–211; 1999.
11. Jansen, A. S. P.; Hoffman, J. L.; Loewy, A. D. CNS sites involved insympathetic and parasympathetic control of the pancreas: A viraltracing study. Brain Res. 766:29–38; 1997.
TABLE 2
PERCENTAGE OF TYROSINE HYDROXYLASE (TH)1. DOPAMINE-B-HYDROXYLASE (DBH)1. ARGININE VASOPRESSIN (AVP)1, ANDOXYTOCIN (OXY)1 PSEUDORABIES VIRUS (PRV) NEURONS AFTERVIRUS INJECTION INTO SEVERAL SYMPATHETIC TARGET SITES
WATAdrenal24
GlandStellae13
Ganglion Pancreas11Tail22
Artery Wall
Brainstem
TH 1 PRV
A5 32.19 70 40 58 39
LC 7.29 0 65 99 65
SubC 38.52 0 20 62 97
RVL 5.75 0 – – –
LPGi 2.11 0 – – –
LRt 3.43 0 – – –
Facial nucleus 21.19 0 – – –
DBH 1 PRV
A5 24.58 – – – –
RVL 2.13 – – – –
Forebrain
TH 1 PRV
PVN 2.62 ,10 0 – –
MPA 1.17 – – – –
AVP 1 PRV
PVN 1.07 1 2 2/rat 0
MPA 2.65 – – – –
OXY 1 PRV
PVN 3.49 1 10 3/rat 8
WAT, white adipose tissue; A5, noradrenergic area 5; LC, locus coer-uleus; SubC, subcoeruleus; RVL, rostroventroateral medulla; LPGi, lateralparagigantocellular nucleus; LRt, lateral reticular nucleus; RVL, rostroven-trolateral medulla; –, not tested; 0, no double–labeled cells were found;2/rat, 2 double–labeled cells were found per rat; 3/rat, 3 double–labeledcells were found per rat.
384 SHI AND BARTNESS

12. Jansen, A. S. P.; Nguyen, X. V.; Karpitskiy, V.; Mettenleiter, T. C.;Loewy, A. D. Central command neurons of the sympathetic nervoussystem—Basis of the fight-or-flight response. Science 270:644–646;1995.
13. Jansen, A. S. P.; Wessendorf, M. W.; Loewy, A. D. Transneuronallabeling of CNS neuropeptide and monoamine neurons after pseudo-rabies virus injections into the stellate ganglion. Brain Res. 683:1–24;1995.
14. Migliorini, R. H.; Garofalo, M. A. R.; Kettelhut, I. C. Increasedsympathetic activity in rat white adipose tissue during prolongedfasting. Am. J. Physiol. 272:R656–R661; 1997.
15. Morrison, S. F. Differential control of sympathetic outflow to brownadipose tissue and to the splanchnic bed by the raphe pallidus (RPa)and the rostral ventrolateral medulla (RVLM). Soc. Neurosci. 23:839;1997.
16. Morrison, S. F. RVLM and raphe differentially regulate sympatheticoutflows to splanchnic and brown adipose tissue. Am. J. Physiol.276:R962–R973; 1999.
17. Newsholme, E. A.; Leech, A. R. Biochemistry for the medical sci-ences. Chichester: John Wiley; 1983.
18. Paxinos, G.; Watson, C. The rat brain in stereotaxic coordinates.Orlando, FL: Academic Press; 1986.
19. Reuss, S.; Burger, K. Substance P-like immunoreactivity in the hypo-thalamic suprachiasmatic nucleus of Phodopus sungorus—Relation todaytime, photoperiod, sex and age. Brain Res. 638:189–195; 1994.
20. Sawchenko, P. E.; Swanson, L. W. Immunohistochemical identifica-tion of neurons in the paraventricular nucleus of the hypothalamus thatproject to the medulla or to the spinal cord in the rat. J. Comp. Neurol.205:260–272; 1982.
21. Shi, H.; Bartness, T. J. Catecholaminergic enzyme, vasopressin and
oxytocin distribution in Siberian hamster brain. Brain Res. Bull. 53:
833–843; 2000.22. Smith, J. E.; Jansen, A. S. P.; Gilbey, M. P.; Loewy, A. D. CNS cell
groups projecting to sympathetic outflow of tail artery: Neural
circuits involved in heat loss in the rat. Brain Res. 786:153–164;
1998.
23. Strack, A. M.; Sawyer, W. B.; Hughes, J. H.; Platt, K. B.; Loewy,
A. D. A general pattern of CNS innervation of the sympathetic outflow
demonstrated by transneuronal pseudorabies viral infections. BrainRes. 491:156–162; 1989.
24. Strack, A. M.; Sawyer, W. B.; Platt, K. B.; Loewy, A. D. CNS cellgroups regulating the sympathetic outflow to adrenal gland as revealedby transneuronal cell body labeling with pseudorabies virus. BrainRes. 491:274–296; 1989.
25. Takahashi, A.; Shimazu, T. Hypothalamic regulation of lipid metab-olism in the rat: Effect of hypothalamic stimulation on lipolysis. J.Auton. Nerv. Syst. 4:195–205; 1981.
26. Teixeira, V. L.; Antunes-Rodrigues, J.; Migliorini, R. H. Evidence forcenters in the central nervous system that selectively regulate fatmobilization in the rat. J. Lipid Res. 14:672–677; 1973.
27. Vincent, S. R. Distributions of tyrosine hydroxylase-, dopamine-b-hydroxylase-, and phenylethanolamine-N-methyltransferase-immuno-reactive neurons in the brain of the hamster (Mesocricetus auratus).J. Comp. Neurol. 238:584–599; 1988.
28. Youngstrom, T. G.; Bartness, T. J. Catecholaminergic innervation ofwhite adipose tissue in the Siberian hamster. Am. J. Physiol. 268:R744–R751; 1995.
NEUROCHEMICAL CODING OF SNS OUTFLOW 385





























