Embed Size (px)
Citation preview

Nematoda do Talude da
Bacia de Campos(Rio de Janeiro . Brasil)
PernambucoFevereiro, 2009
CATÁLOGO DE GÊNEROS

Coordenação do Catálogo Verônica da Fonsêca-Genevois,
André Morgado Esteves, Maria CristinadaSilva e Alessandra Prates Botelho
Revisão CientíficaTânia Nara Campinas Bezerra e
Gustavo Fonseca
Projeto Gráfico e diagramaçãoLuciana Tosta e Fast design
Não é permitido a reprodução e transmissão total ou parcial, sejam quais forem os meios utilizados,sem a devida autorização e citação da fonte.
Impresso no Brasil em fevereiro de 2009 pela:Fast Design - Prog. Visual, Editora e Grafica Rápida LTDA.
e-mail: [email protected]: 100 exemplares
CapaNatália Franca
FotosCapa - Pseudocela Filipjev, 1927Fotos das pranchas dos Gêneros:
Alessandra Prates BotelhoPranchas dos Gêneros:
Luciana Tosta
Ficha catalografica
Patrocínio

Colaboradores:
Alexandre Lacerda de Larrazábal Filho
Graduando em Ciências Biológicas-UFPE
Betânia Cristina Guilherme
Prof. Adjunto 1-UFCG, Campus Cuité
Francisco José Victor de Castro
Prof. Adjunto 1-UFCG, Campus Cuité
Geruso Vieira de Miranda Júnior
Mestre em Biologia Animal-UFPE
Giovanni Amadeus Paiva dos Santos
Doutor em Ciências Biológicas-UFPE
Grácia Maria Bártholo
Doutor em Oceanografia-UFPE
Lídia Lins Pereira
Bacharel em Ciências Biológicas-UFPE
Luciana Davina Tosta Sobral
Bacharel em Ciências Biológicas -UFBA
Coordenação
Verônica Gomes da Fonsêca-Genevois
Prof. Associado II
Departamento de Zoologia - UFPE
André Morgado Esteves
Prof. Adjunto I
Departamento de Zoologia - UFPE
Maria Cristina da Silva
Mestre em Biologia Animal-UFPE
Alessandra Prates Botelho
Mestre em Biologia Animal-UFPE
EQUIPE DE TRABALHO

Mariana da Fonseca Cavalcanti
Bacharel em Ciências Biológicas -UFPE
Neyvan Renato Rodrigues da Silva
Professor CEFET-RN
Orane Falcão de Souza Alves
Professor Adjunto – UFBA
Patrícia Fernandes Neres
Mestre em Biologia Animal-UFPE
Paulo de Barros Passos Filho
Graduando em Ciências Biológicas-UFRPE
Rita de Cássia Coelho de Lima
Mestre em Biologia Animal-UFPE
Taciana Kramer de Oliveira Pinto
Prof. Adjunto 1-UFAL,Campus Penedo
Tatiana Nunes Cavalcante Alves
Mestranda em Biologia Animal-UFPE
Verônica Silva de Oliveira
Licenciada em Ciências Biológicas
Virág Venekey
Doutor em Oceanografia-UFPE
Viviane Freire Lira
Mestre em Biologia Animal-UFPE
Wilson Saraiva
Graduando em Ciências Biológicas-UFPE

SUMÁRIO
Apresentação
Prefácio
Introdução................................................................................13
Aréa de Estudo......................................................................... 18
Material e Métodos ...................................................................19
Resultados ............................................................................... 22
Lista Taxonômica..................................................................... 22
Ordem Enoplida ...................................................................... 35
Ordem Chromadorida.............................................................. 67
Ordem Desmodorida ..............................................................105
Ordem Desmoscolecida .......................................................... 121
Ordem Monhysterida ..............................................................133
Ordem Araeolaimida .............................................................. 153
Ordem Plectida ....................................................................... 167
Abreviações ............................................................................ 181
Glossário ................................................................................183
Referências ............................................................................. 187

7
Prefácio
Compared to other areas of science, Nematology is a young subject with limited number of
devotees. Even younger and smaller is the group of marine nematologists, and so is the
history of the marine nematologist core in Brazil.
As nematodes are the predominant group of meiobenthic animals, often researchers start
with meiofauna to come to nematodes. This is what happened with VERONICA DA FONSECA-
GENEVOIS: she went to Paris (France) in 1985 and was introduced to the wonders of
meiofauna by Dr. JEANNE RENAUD-MORNANT at the ‘Muséum National d’Histoire Naturelle’
in Paris. After her ‘Doctorat d`État en Oceanographie - Meiofauna of the Loire Estuary -
at the University of Nantes, France in 1987 promoted by Dr. FRANÇOIS OTTMAN she returned
to Brazil and started the Meiofauna Laboratory of the Zoology focusing on meiofauna
research at the ‘Universidade Federal de Pernambuco’ in Recife. The first student was
CLELIA MÁRCIA CAVALCANTI DA ROCHA, currently a professor at the Federal Rural University
of Pernambuco, working on Tardigrades. Later, one of her PhD students, TANIA NARA
BEZERRA, studying the Meiofauna of Olinda Isthmus became fascinated by the numbers
and diversity of nematodes and went to Europe to enrol for the master programme “Post-
graduate International Nematology Course” (PINC) at Ghent University (Belgium). She
did the first steps towards the start of nematological research in Recife which leaded to a
close cooperation and friendship between the nematology group of the Ghent University
and of Recife. Many more students followed the steps of TANIA NARA (1999) and graduated
as Master of Science in Nematology at Ghent University: Virág Venekey (2002), GIOVANNI
SANTOS, Maria CRISTINA DA SILVA and NEYVAN RENATO RODRIGUES DA SILVA (2003), all from
Recife and GUSTAVO FONSECA (2004) from the laboratorium of SERGIO NETTO in Santa Catarina
State and TATIANA FABRICIO (2006), a student from ANDRÉ MORGADO ESTEVES, from the Fed-
eral University of Rio de Janeiro.
In the meantime Prof. VERONICA as well as GRACIA MARIA BARTHOLO, FRANCISCO VICTOR DE
CASTRO and ANDRÉ MORGADO ESTEVES came on a temporary visit to Ghent University to be-
come acquainted with some techniques and to update the literature.
Actually, among the PINC-alumni, the group in Recife is certainly top-ranked in the estab-
lishment of a critical mass. And of course more talented and enthusiastic master (VIVIANE

8
LIRA, RITA DE CASSIA COELHO LIMA, LUCIANA DAVINA TOSTA SOBRAL, TATIANA NUNES CAVALCANTI
ALVES, GERUSO VIERA DE MIRANDA JUNIOR, MARIANA DA FONSECA CAVALCANTI, VERONICA SILVA DE
OLIVEIRA, MONICA MARINHO VERÇOSA) and PhD students (NEYVAN RENATO RODRIGUES DA SILVA,
BETANIA CRISTINA GUILHERME, MARIA CRISTINA DA SILVA, PATRICIA FERNANDES NERES, ALESSANDRA
PRATES BOTELHO) are daily involved in several projects in Recife .
After the examples set by her students Prof. VERONICA became herself attracted by nema-
todes around 2OO2 and promoted nematodes to colleagues, this resulted in a poster
‘Biodiversity of Free-living Marine nematodes from Tropical Brazilian Ecosystems’ pre-
senting the marine nematological laboratories in Brazil at the Russian conference in
Vladivostok, Russia.
A perfect and valuable example of the international cooperation between the Nematology
groups of different Universities all over the world : the University of California, Riverside
(Paul DE LEY, as sponsor), the Ghent University (MAGDA VINCX, ANN VANREUSEL, TOM MOENS,
NIC SMOL) and the Federal University of Pernambuco, is demonstrated in 2003-O4 by the
contribution of TANIA NARA BEZERRA and GUSTAVO FONSECA in the transfer of the hard copy
files (erected by NIC SMOL) of the description of all nematodes species per genus into elec-
tronic files and in this way making a substantial part of the NEMYS- digital library
(www.nemys.ugent.be) providing descriptions and literature of all marine nematodes.
Prof. VERONICA is not only an inspiration for her students and colleagues, she is also a
very good manager, successful in searching sponsors for the development of a nemato-
logical laboratory and of projects creating the possibility for youngsters of getting involved
in nematological research. A milestone was certainly the involvement of PETROBRAS, the
Brazilian Petrol and Gas Company, in sampling campaigns and later also in the financial
support of projects such as Deep Sea Water in Campos Basin, which resulted in the expan-
sion of the nematological laboratory through purchasing the necessary equipment such as
microscopes, computer facilities and the creation of an experimental laboratory.
PETROBRAS also opened the international doors for teaching and research aid and coop-
eration in taxonomy: in the period 2004-2008 WILFRIDA DECRAEMER and NIC SMOL, both
from Ghent University visited the laboratory in Recife and added value to nematological
expertise.
The dedication of the meiofauna team of Recife, including PAULO JORGE PARREIRA DOS SANTOS,
LILIA DE SOUSA-SANTOS, VERONICA, ANDRÉ, ADRIANE and CLÉLIA was proved by the successful

9
organisation of the ‘Thirtheenth International Meiofauna Conference’ (THIRIMCO) held
in Recife in 2007, another milestone to mark the efforts on the international platform.
Nowadays Dr. ANDRÉ MORGADO ESTEVES became the co-worker and professor at VERONICA’S
lab. and some of the students already finished their PhD, such as Dr. TANIA NARA BEZERRA
(now a member of the Marine Section at Ghent University), Dr. FRANCISCO VICTOR DE CASTRO,
teaching in Paraíba, DR. GIOVANNI SANTOS, now being a post-doc at Ghent University work-
ing with TOM MOENS; and Dr. VIRÁG VENEKEY; the latter lecturing at the Federal University
of Belem and focusing on estuarine nematodes , Dr. TACIANA KRAMER DE OLIVEIRA PINTO at
Alagoas.at the Northeast of Brazil .In the meantime the list of publications is growing
proving a continuous interest of the whole team.
After a decade of raised nematode interest in Recife it is the right moment for the publica-
tion of a Catalogue of deep sea nematodes found in Campos Basin, studying since 2003
that allowed the biggest taxonomic list in the word.
And this is not the end... new projects are waiting, opening new challenges for the whole
team in discovering multifaceted subjects.
Congratulations Prof. VERONICA DA FONSECA-GENEVOIS, Prof. André Morgado Esteves and
your team, up to a brilliant future!
NIC SMOL
Coordenadora do curso de Nematologia
Universidade de Gent - Bélgica

Apresentação
O projeto de Caracterização ambiental de Águas Profundas da Bacia de Campos foi
criado e desenvolvido entre 2001 e 2007 pelo CENPES/PETROBRAS, em parceria com
oito universidades brasileiras. Este projeto teve como objetivo caracterizar as águas e
sedimentos da região oceânica da Bacia de Campos, entre as profundidades de 700m e
2000m, quanto a características físicas, químicas e biológicas.
Esta iniciativa pode ser considerada pioneira na costa brasileira, no que diz respeito
à avaliação de ambientes profundos, devido a algumas características particulares como a
abrangência regional, a amostragem em grandes profundidades, o viés de
multidisciplinaridade nas avaliações de água e sedimento, as coletas simultâneas de
parâmetros que permitiram a integração dos dados gerados e grande investimento na
identificação taxonômica dos organismos coletados.
O projeto é considerado pela empresa como sendo de alta relevância devido ao foco
na geração de informações ambientais na área dos cinco mega-campos petrolíferos
conhecidos na Bacia de Campos (até as descobertas na camada pré-sal), que respondem a
cerca de 80% da produção brasileira em 2008. Isso representa um considerável aumento
no nível de conhecimento sobre o ambiente marinho profundo desta importante área de
exploração e produção de óleo e gás.
Um dos objetivos específicos deste projeto foi a capacitação de pesquisadores
brasileiros no que diz respeito ao conhecimento de grupos biológicos de grande importância
ecológica em ambientes profundos, porém pouco estudados no Brasil. O Filo Nematoda,
objeto deste catálogo, é um destes grupos, sendo considerado excelente indicador da
qualidade ambiental.
O aumento na capacitação e formação de pesquisadores especialistas na chamada
nematofauna se deu na forma de uma parceria, iniciada em 2002, entre o Centro de
Pesquisas da Petrobras (CENPES) e a Universidade Federal de Pernambuco (UFPE). O
processo passou por diversos níveis de desenvolvimento científico, desde análises ecológicas
até refinamentos relacionados à identificação taxonômica e biologia molecular.
Durante este período, foram formados um doutor, três mestres e um aluno de
iniciação científica. Três teses, cinco dissertações e duas monografias encontram-se em

12
andamento. Esses dados permitem concluir que a disseminação de conhecimento
proporcionada por este projeto permitiu que em seis anos fosse, no mínimo, triplicado o
número pesquisadores aptos à avaliação destes organismos. Vale ressaltar que, uma vez
que outras universidades no país incorporaram a seus quadros estes profissionais
especializados, a ampliação dos núcleos existentes para avaliação da nematofauna
aumentou ainda mais a disseminação deste conhecimento.
Também merece destaque a intensificação das parcerias internacionais com a
Universidade de Gent (Bélgica) e a Universidade da Califórnia (EUA), que permitiram que
alunos brasileiros pudessem visitar centros de referência, bem como se aperfeiçoar junto
a alguns dos maiores especialistas mundiais no assunto que, por sua vez, também realizaram
cursos e visitas técnicas à UFPE.
Este catálogo é apenas um dos frutos desta profícua parceria entre a Petrobras e a
Universidade Federal de Pernambuco e visa aumentar e disseminar o conhecimento sobre
estes pequenos organismos de crescente importância ecológica.
Msc. Ana Paula da Costa Falcão
Bióloga Marinha
CENPES - PETROBRAS

13
INTRODUÇÃO
Ainda que alguns poucos autores continuem a utilizar o termo Aschelminthes para
agrupar diferentes táxons, existe a concordância, na quase totalidade da literatura, de que
o táxon Aschelminthes representa um grupo polifilético e, que seus grupos componentes
(entre eles, os Nematoda), são, na verdade, Filos distintos. Isso está relacionado diretamente
ao fato de que a condição pseudocelomada (= blastocelomada), assim como outras
características, não representaria verdadeiras sinapomorfias dos Aschelminthes, mas sim
que a mesma tenha surgido, várias vezes ao longo da evolução desses grupos, ou seja,
representaria uma homoplasia por convergência relacionada ao modo de vida desses
animais (Ruppert, 1991).
Outro tema que é objeto de grande discussão, ainda muito atual, na literatura
zoológica, é o posicionamento dos Nematoda (e de outros táxons relacionados ao antigo
Filo Aschelminthes) dentro dos Bilateria. Uma visão inicial, com base em observações
morfológicas, considera os Nematoda (e outros “asquelmintos”) relacionados à linha dos
protostômios acelomados (Hyman, 1951). No entanto, Aguinaldo et al. (1997), com base
em análise do gene DNAr 18S, propuseram que os Nematoda estariam relacionados aos
artrópodes e outros grupos, reunindo-os em um grupo monofilético denominado
Ecdysozoa, nome este relacionado ao fato de que todos esses animais sofrem muda. Embora
alguns estudos subseqüentes venham tentando corroborar essa hipótese, existe inúmeras
críticas a mesma, desde o próprio uso desse gene até as diferenças existentes entre esses
táxons, no que diz respeito, a embriogênese e ao próprio processo fisiológico da muda (De
Ley & Blaxter, 2002; Brusca & Brusca, 2003).
Os Nematoda fazem parte de uma das grandes estórias de sucesso do reino animal,
como um dos grupos de invertebrados mais ricos em densidade e diversidade no meio
aquático. Após um século de pesquisas taxonômicas só são conhecidas 10% de suas espécies
com estimativas que variam entre 0,1 a 100 milhões (Meldal et al., 2006). O número de
indivíduos presentes em qualquer habitat sedimentar marinho é normalmente de ordem
ou magnitude maior do que para qualquer outro grande táxon (Platt & Warwick, 1980).
Uma das explicações para este padrão tem sido associada, sobretudo, ao status trófico do
Filo, onde a variedade na morfologia bucal permitiu o aproveitamento de inúmeros recursos
tróficos (Bouwman, 1983).

14
MEIOFAUNANEMATOFAUNA
Em mar profundo, os Nematoda prevalecem em número de espécies e de indivíduos
(80 a 99% da abundância total de metazoários) quando os Foraminifera não são considerados
(Heip et al., 1985). Situações ambientais muito específicas são responsáveis pela diminuição
de densidade e de riqueza, tais como: o aporte sazonal de fitodetritos (Rice & Lambshead,
1994), a interação não equilibrada entre produtividade e distúrbio (Huston, 1994). A relação
densidade/riqueza é dependente da granulometria dos sedimentos e que, a diversidade dos
Nematoda, é maior em ambientes de águas profundas do que naqueles de águas rasas com as
mesmas características sedimentares (Heip et al., 1982).
Estudos que descrevem a composição taxonômica deste grupo e/ou abordam as suas
relações com parâmetros ambientais de mar profundo são raros e limitados a poucos locais,
como o Mediterrâneo (Vitiello, 1976; Vivier, 1978b), Baía de Biscay (Dinet & Vivier, 1979), a
área entre a Carolina do Norte e Venezuela (Tietjen, 1971; Tietjen, 1976; Tietjen, 1984; Tietjen,
1989), a Escócia (Thistle & Sherman, 1985), a Noruega (Jensen, 1988) e o Atlântico Sudeste
(Soltwedel, 1993). Entretanto, mais raros ainda, são aqueles referentes ao Atlântico Sudoeste,
podendo-se contar apenas com o de Netto et al. (2005), referente à Bacia de Campos. Os
demais estudos ainda apresentam seus resultados restritos a apresentações em reuniões
científicas e trabalhos acadêmicos, no entanto, esses mesmos estudos ainda não publicados, já
vêem impulsionando a formação de novos taxonomistas no Brasil (ver referências em
THIRIMCO, 2007). Com isso, sabe-se que um volume considerável de publicações está por
vir, incluindo, as descrições das novas espécies pertencentes aos gêneros mais abundantes da
Bacia de Campos e que constam deste catálogo.
Para melhor visualização das áreas de mar profundo prospectadas até o presente,
apresenta-se a Figura 1.
Figura 1: Mapa Mundi da distribuição de Nematoda em mar profundo.

15
Segundo Lorenzen (1994) os caracteres morfológicos mais comuns usados na
classificação de Nematoda são:
1 – cutícula
2 - cabeça, setas,
3 - anfídio,
4 - cavidade bucal,
Tipos cutículas: A. lisa; B. estriada; C. fortemente estriada; D. desmen; E. anelada com ponteslongitudinais; F. anelada com pontes longitudinais em forma de “V”; G. heterogênia; H. pontuada.
Tipos de anfídio: A. bolha; B. espiral; C. circular; D. loop; E. oval; F. bolso; G. criptoespiral; H. alongadolongitudinalmente.
Tipos de boca: A. diminuta; B. sem dente; C. com dente dorsal; D. com dente dorsal e subventral; E. complacas; F. em duas partes separadas por fileiras de odontias; G. com mandíbula; H. tubular.

16
5 - órgãos reprodutores
A denominada “Escola Inglesa da Nematologia Marinha” impulsionou a
identificação, ao nível de Gênero dos Nematoda marinhos com a criação de excelentes
chaves pictoriais (Platt & Warwick, 1983, 1988; Warwick et al, 1998).
Para a identificação ao nível específico, a morfometria é um procedimento
considerado fundamental, ainda que hajam várias exceções descritas na bibliografia
especializada que não constam do esquema abaixo.
CARACTERES MORFOMÉTRICOS L: comprimento total do corpo mbd: diâmetro máximo do corpo ph: comprimento da faringe t: comprimento da cauda S: comprimento da espícula Sc: comprimento da seta cefálica PROPORÇÕES Índices de De Man (1880): a: comprimento total do corpo/diâmetro máximo. b: comprimento total do corpo/comprimento total da faringe. c: comprimento total do corpo/ comprimento total da cauda.
S
Sc

17
A aplicação da técnica molecular, aos estudos de sistemática do Filo Nematoda, foi
intensificada no final do século XX (De Ley & Blaxter, 2002). Blaxter (2001) afirmou que
antes das análises filogenéticas moleculares, o estudo sistemático do filo era limitado pelo:
(1) número de espécies e diversidade morfológica, (2) limitações das análises de sua
morfologia por microscopia óptica, (3) a inevitável especialização da sistemática desses
animais e (4) a dificuldade de formar sistematas. De Ley & Blaxter (2002) defendem que
“a ontogênese e ultra-estrutura dos nematóides são fracamente entendidas e que tais
dificuldades têm levado a criação de múltiplas classificações, pelo menos, parcialmente
conflitantes”. Meldal et al. , (2006) advertem que “a falta de critérios objetivos para acessar
a homologia entre caracteres morfológicos, tem dificultado a reconstrução da filogenia do
filo Nematoda”. Assim, tem-se hoje a tendência de associar as tradicionais características
morfológicas às análises moleculares, criando-se a Taxonomia Integrativa (Fonseca et al.,
2008; Dayrat, 2008).
Desta forma, os recentes estudos filogenéticos, assim como estudos com DNA e
genômica estão trazendo grandes modificações na taxonomia de Nematoda, tal como o
sistema proposto por De Ley & Blaxter (2002).

18
ÁREA DE ESTUDO
A Bacia de Campos (21o30’W e 23o30’S) encontra-se na região Sudeste do Brasil
(Figura 1), apresentando largura que varia de 120km a 150km e profundidades máximas
entre 2.400 e 3.000 metros (Soares-Gomes et al., 1999). É considerada relativamente
jovem em termos de idade geológica, possuindo por volta de 140 – 130 milhões de anos
(Defeo, 2002). O talude dessa região é coberto por sedimentos finos terrígenos e por uma
fração arenosa, além de uma predominância de foraminíferos nos sedimentos (Soares-
Gomes et al., 1999).
A área de estudo está sujeita à influência das Correntes do Brasil, que até os 200m
de profundidade corre paralela à costa e transporta a massa de água conhecida como Água
Tropical. Abaixo dos 300m de profundidade há uma inversão na direção da corrente,
transportando grande parte de massas de água provenientes da Água Central do Atlântico
Sul (Metzler et al., 1997).
As águas brasileiras do Atlântico Sul apresentam concentração de nutrientes,
biomassa e níveis de produtividade baixos. Estes fatos estão associados com a presença de
massas de AT oligotrófica, com águas superficiais quentes e alta salinidade transportada
pela CB da isóbata de 200m (Signorini, 1978 apud Metzler et al., 1997). Contudo,
incrementos na biomassa planctônica e na produtividade têm sido observados
freqüentemente devido à entrada de uma massa de água profunda (ACAS) fria e rica em
nutrientes entre 200 e 600m de profundidade. Essa massa de água origina-se em uma
convergência subtropical próximo aos 300S (Brandini, 1991 apud Metzler et al., 1997).
A área também é sujeita a influência das massas de água de Contorno Intermediário
(CCI), de Contorno Profundo (CCP), Água Intermediária Antártica (AIA) e Água Profunda
do Atlântico Norte (APAN) (PETROBRAS, 2001 e 2003).

19
MATERIAL E MÉTODOS
Amostras de sedimento destinadas ao estudo da nematofauna na Bacia de Campos
(Figura 2) foram coletadas com Box-corer da Ocean Instruments, do tipo USNEL SPADE
CORER modificada para coleta de 0,25 m2 de sedimento, sub-dividida em 25 testemunhos
(Figura 3).
Figura 2: Bacia de Campos (Rio de Janeiro, Brasil) com localização das estações de coleta na área Sul e
Norte (fonte: CENPES-PETROBRAS, 2004).
Figura 3: Box-corer tipo colméia (A) com sua divisão interna em várias células e estratificação sedimentar
(B) (fonte: CENPES-PETROBRAS, 2004).
A
B

20
Três testemunhos de cada lançamento da Box-corer foram utilizados para as coleta
de meiofauna nos estratos de 0-2 cm, 2-5 cm. Em cada testemunho, as amostras foram
tomadas com uma seringa de plástico de 2,0 cm de diâmetro interno, inserida no sentido
horizontal ao diâmetro do corer. Na campanha OCEANPROF II uma amostragem
suplementar foi realizada em 5 pontos de cada área da Bacia de Campos. Assim, trinta
amostras foram coletadas com testemunho de 10,0 x 10,0 cm, mantendo-se a mesma
estratificação sedimentológica.
Cinco isóbatas foram escolhidas para amostragem do Bentos (750 m, 1.050 m, 1.350
m, 1.650 m e 1.950 m) nas áreas Sul e Norte da Bacia de Campos. O material foi coletado
em 45 estações oceanográficas distribuídas em 9 transectos. Cada amostra foi transferida
para um recipiente plástico de 100 ml e fixadas em formol 10% tamponado com bórax
Em laboratório foi utilizada a metodologia de Boisseau (1957) para extração da
fauna, sendo as amostras lavadas em água corrente e retidas em peneiras com aberturas
de malhas de 0,044 mm e 0,5 mm. O material filtrado neste intervalo foi colocado em
placa de Petri para centrifugação manual, sendo o sobrenadante vertido em placas de
Dolffus, composta de 200 quadrados de 0,25 cm² cada um.
Em microscópio esterioscópico todos os nematódeos contidos em cada amostra
foram contados, retirados com auxílio de estilete e estocados para confecção de lâminas
permanentes após a diafanização. Para as amostras de maior volume, 100 nematódeos
foram retirados aleatoriamente. A densidade dos gêneros foi calculada através da área da
seringa utilizada na coleta, a fim de padronizar os resultados para a medida internacional
da meiofauna (ind./10 cm2).
O processo de diafanização seguiu o método de De Grisse (1969) que consiste na
introdução dos indivíduos em três soluções, contendo: 1 - 99% formoldeído a 4% mais 1%
glicerina (em dessecador, por vinte e quatro horas); 2 - 95% etanol mais 5% glicerina
(introduzida a cada duas horas por 24 horas a temperatura ambiente); 3 - 50% etanol
mais 50% glicerina (por duas horas a temperatura ambiente).
Para montagem de lâminas permanentes cinco indivíduos foram colocados em gota
de glicerina centralizada em um círculo de parafina. Sobre o círculo, uma lamínula foi
adicionada, sendo o conjunto levado ao aquecimento para fixação. Este procedimento
seguiu os passos descritos por Cobb (1917). As lâminas foram inicialmente levadas ao

21
microscópio óptico para a identificação dos animais ao nível de gênero, utilizando-se as
chaves pictoriais propostas por Platt & Warwick (1983), Platt & Warwick (1988), Warwick
et al. (1998). A classificação da nematofauna baseou-se em Lorenzen (1994) e em De Ley
et al., 2006.
Os gêneros identificados dizem respeito às campanhas de RONCADOR (apenas os
nematódeos encontrados na macrofauna), de ALBACAR (amostras coletadas em Barra-
cuda e Caratinga), de OCEANPROF I e de OCEANPROF II (incluindo as trinta amostras
suplementares).
Constam deste catálogo os gêneros representados por machos na comunidade
meiofaunística. Aqueles gêneros identificados através de fêmeas estão apenas contidos
na lista taxonômica, destacados com asterisco.

22
RESULTADOS
Lista Taxonômica
Usando-se a classificação de Lorenzen (1994), feita com base na morfologia, a lista da
nematofauna da Bacia de Campos apresenta 192 Gêneros, 40 Famílias, 5 Ordens e 1 Classe,
podendo assim ser considerada como a maior lista de Nematoda de mar profundo ao nível
mundial (tanto ao nível de gênero como de família).
Em função do surgimento de uma nova proposta de classificação (De Ley et al., 2006),
baseada em estudos moleculares, fez-se um ajuste dessa lista a esse novo sistema de
classificação, passando a mesma a ser composta por 189 Gêneros, pertencentes a 43
Famílias e 9 Ordens.
Esses resultados mostram que mesmo com a necessidade de ajustes dos táxons, em função
da aplicação de novas técnicas no estudo da sistemática dos Nematoda, poder-se afirmar
que a composição da nematofauna da Bacia de Campos continua sendo a mais rica já
registrada em literatura.
A lista taxonômica contém 189 gêneros, distribuídos em 43 famílias pertencentes a 9 ordens,
sendo Enoplida, a mais abrangente, com 13 famílias, Plectida com 7 familias, Monhysterida
com 6 famílias, Desmodorida com 5 famílias, Chromadorida com 4 famílias, Araeolaimida
com 3 famílias, Triplonchida e Desmoscolecida com 2 famílias cada e Rhabditida com
apenas 1 família.
Chromadoridae e Xyalidae apresentaram o maior número de gêneros, cada uma delas
com 18 gêneros.

23
FILO NEMATODA Potts, 1932
CLASSE ENOPLEA Inglis, 1983
SUBCLASSE ENOPLIA Pearse, 1942
ORDEM ENOPLIDA Filipjev, 1929
Subordem Enoplina Chitwood & Chitwood, 1937
Superfamília Enoploidea Dujardin, 1845
Família Enoplidae Dujardin, 1845
Enoplus Dujardin, 1845*
Família Thoracostomopsidae Filipjev, 1927
Enoplolaimus De Man, 1893*
Enoploides Ssaweljev, 1912*
Epacanthion Wieser, 1953
Fenestrolaimus Filipjev, 1927*
Mesacanthion Filipjev, 1927
Paramesacanthion Wieser, 1953*
Trileptium Cobb, 1933*
Família Anoplostomatidae Gerlach & Riemann, 1974
Anoplostoma Bütschli, 1874*
Família Phanodermatidae Filipjev, 1927
Crenopharynx Filipjev, 1934*
Micoletzkyia Ditlevsen, 1926
Phanoderma Bastian, 1865*
Phanodermella Kreis, 1928*
Phanodermopsis Ditlevsen, 1926*
Família Anticomidae Filipjev, 1918
Anticoma Bastian, 1865*
Cephalanticoma Platonova, 1976
Odontanticoma Platonova, 1976*
Paranticoma Micoletzky, 1930*

24
Subordem Trefusina Siddiqi, 1983
Superfamília Trefusioidea Gerlach, 1966
Família Simpliconematidae Blome & Schrage, 1985
Simpliconema Blome & Schrage, 1985*
Família Trefusiidae Gerlach, 1966
Halanonchus Cobb, 1920*
Trefusia De Man, 1883*
Subordem Oncholaimina De Coninck, 1965
Superfamília Oncholaimoidea Filipjev, 1916
Família Oncholaimidae Filipjev, 1916
Adoncholaimus Filipjev, 1918*
Meyersia Hopper, 1967
Oncholaimus Dujardin, 1845
Metoncholaimus Filipjev, 1918*
Prooncholaimus Micoletzky, 1924*
Viscosia De Man, 1890
Família Enchelidiidae Filipjev, 1918
Bathyeurystomina Lambshead & Platt, 1979*
Calyptronema Marion, 1870*
Eurystomina Filipjev, 1921
Pareurystomina Micoletzky, 1930*
Polygastrophora De Man, 1922*
Symplicostoma Bastian, 1865*
Subordem Tripyloidina De Coninck, 1965
Superfamília Tripyloidoidea Filipjev, 1928
Família Tripyloididae Filipjev, 1928
Tripyloides De Man, 1888*

25
Subordem Ironina Siddiqi, 1983
Superfamília lronoidea De Man, 1876
Família lronidae De Man, 1876
Dolicholaimus De Man, 1888*
Syringolaimus De Man, 1888
Thalassironus De Man, 1889
Família Leptosomatidae Filipjev, 1916
Cylicolaimus De Man, 1889
Leptosomatum Bastian, 1865
Metacylicolaimus Stekhoven, 1946
Platycoma Cobb, 1894*
Platycomopsis Ditlevsen, 1926*
Thoracostoma Marion, 1870*
Família Oxystominidae Chitwood, 1953
Halalaimus De Man, 1888
Litinium Cobb, 1920
Nemanema Cobb, 1920
Oxystomina Filipjev, 1921
Thalassoalaimus De Man, 1893
Wieseria Gerlach, 1956*
ORDEM TRIPLONCHIDA Cobb, 1920
Subordem Tobrilina Tsalolikhin, 1976
Superfamília Prismatolaimoidea Micoletzky, 1922
Família Prismatolaimidae Micoletzky, 1922
Pandolaimus Allgén, 1929*
Família Triodontolaimidae Thorne, 1935
Triodontolaimus De Coninck, 1965*

26
CLASSE CHROMADOREA
SUBCLASSE CHROMODORIA
ORDEM CHROMADORIDA Chitwood, 1933
Subordem Chromadorina Filipjev, 1929
Superfamília Chromadoroidea Filipjev, 1917
Família Chromadoridae Filipjev, 1917
Acantholaimus Allgén, 1933
Actinonema Cobb, 1920
Chromadora Bastian, 1865*
Chromadorella Filipjev, 1918*
Chromadorita Filipjev, 1922
Dichromadora Kreis, 1929
Endeolophos Boucher, 1976
Euchromadora De Man, 1886
Graphonema Cobb, 1898*
Hypodontolaimus De Man, 1886*
Innocuonema Inglis, 1969*
Neochromadora Micoletzky, 1924
Parachromadorita Blome, 1974*
Parapinnanema Inglis, 1969*
Prochromadorella Micoletzky, 1924*
Rhips Cobb, 1920*
Spiliphera Bastian, 1865*
Trochamus Boucher & Bovée, 1972*
Família Neotonchidae Wieser & Hopper, 1966
Comesa Gerlach, 1956
Filitonchus Platt, 1982*
Nannolaimus Cobb, 1920
Gomphionchus Platt, 1982*
Neotonchus Cobb, 1933*
Família Cyatholaimidae Filipjev, 1918
Acanthonchus Cobb, 1920*
Cyatholaimus Bastian, 1865*

27
Longicyatholaimus Micoletzky, 1924
Marylynnia Hopper, 1977
Metacyatholaimus Stekhoven, 1942
Minolaimus Vitiello, 1970
Nannolaimoides Ott, 1972*
Paracanthonchus Micoletzky, 1922*
Paracyatholaimus Micoletzky, 1924
Paracyatholaimoides Gerlach, 1953*
Paralongicyatholaimus Stekhoven, 1942*
Pomponema Cobb, 1970*
Família Selachinematidae Cobb, 1915
Cheironchus Cobb, 1917
Choanolaimus De Man, 1880
Gammanema Cobb, 1920
Halichoanolaimus De Man, 1886
Latronema Wieser, 1954
Richtersia Steiner, 1916*
Synonchiella Cobb, 1920*
ORDEM DESMODORIDA De Coninck, 1965
Subordem Desmodorina De Coninck, 1965
Superfamília Desmodoroidea Filipjev, 1922
Família Desmodoridae Filipjev, 1922
Chromaspirinia Filipjev, 1918*
Desmodora De Man, 1889
Eubostrichus Greef, 1869*
Metachromadora Filipjev, 1918*
Molgolaimus Ditlevsen, 1921*
Onyx Cobb, 1891*
Persperia Wieser & Hopper, 1967*
Polysigma Cobb, 1920*
Pseudochromadora Daday, 1889*
Pseudonchus Cobb, 1920
Spirinia Gerlach, 1963

28
Família Draconematidae Filipjev, 1918
Draconema Cobb, 1913*
Eudraconema Busseau, 1993*
Superfamília Microlaimoidea Filipjev, 1922
Família Microlaimidae Micoletzky, 1922
Aponema Jensen, 1978*
Bolbolaimus Cobb, 1920
Microlaimus De Man, 1880
Spirobolbolaimus Soetaert & Vincx, 1988
Família Aponchiidae Gerlach, 1963
Synonema Coo, 1920
Família Monoposthiidae Filipjev, 1934
Nudora Cobb, 1920
Rhinema Cobb, 1920
ORDEM DESMOSCOLECIDA Filipjev, 1929
Superfamília Desmoscolecoidea Shipley, 1896
Família Desmoscolecidae Shipley, 1896
Calligyrus Lorenzen, 1969*
Desmogerlachia Freudenhammer, 1975*
Desmolorenzenia Freudenhammer, 1975
Desmoscolex Claparéde, 1863
Greeffiella Cobb, 1922
Pareudesmoscolex Weischer, 1962*
Quadricoma Filipjev, 1922
Tricoma Cobb, 1893
Família Cyartonematidae Tchesunov, 1990
Cyartonema Cobb, 1920*
Southernia Allgén, 1929*

29
ORDEM MONHYSTERIDA Filipjev, 1929
Subordem Monhysterina De Coninck & Schuurmans Stekhoven, 1933
Superfamília Monhysteroidea De Man, 1876
Família Monhyteridae Chitwood, 1951
Gammarinema Kinne & Gerlach, 1953*
Geomonhystera Andrássy, 1971*
Thalassomonhystera Jacobs, 1987*
Família Sphaerolaimidae Filipjev, 1918
Doliolaimus Lorenzen, 1966*
Metasphaerolaimus Gourbault e Boucher,1981
Sphaerolaimus Bastian, 1865
Subsphaerolaimus Lorenzen, 1978*
Superfamília Sphaerolaimiodea Filipjev, 1918
Família Xyalidae Chitwood, 1951
Ammotheristus Lorenzen, 1977*
Amphimonhystrella Timm, 1961*
Cobbia De Man, 1907*
Daptonema Cobb, 1920
Elzalia Gerlach, 1957
Gnomoxyala Lorenzen, 1977*
Gonionchus Cobb, 1920*
Linhystera Juario, 1974 *
Manganonema Bussau, 1993*
Metadesmolaimus Stekhoven, 1935*
Omicronema Cobb, 1920*
Paramonohystera Steiner, 1916*
Promonhystera Wieser, 1956*
ProrhynchonemaGourbault, 1982*
Pseudosteineria Wieser, 1956*
Scaptrella Cobb, 1917 *
Stylotheristus Lorenzen, 1977*
Theristus Bastian, 1865

30
Subordem Leptolaimina Andrássy, 1974
Superfamília Siphonolaimoidea Filipjev, 1918
Família Siphonolaimidae Filipjev, 1918
Siphonolaimus De Man, 1893*
Família Linhomoeidae Filipjev, 1922
Desmolaimus De Man, 1880*
Didelta Cobb, 1920
Disconema Filipjev, 1918
Eumopholaimus Schultz, 1932*
Linhomoeus Bastian, 1865*
Megadesmolaimus Wieser, 1954*
Metalinhomoeus De Man, 1907
Paralinhomoeus De Man, 1907 *
Terschellingia De Man, 1888
Família Bodonematidae Jensen, 1991
Bodonema Jensen, 1991**
ORDEM ARAEOLAIMIDA De Coninck & Schuurmans Stekhoven, 1933
Superfamília Axonolaimoidea Filipjev, 1918
Família Comesomatidae Filipjev, 1918
Cervonema Wieser, 1954
Comesoma Bastian, 1865*
Dorylaimopsis Ditlevsen, 1918*
Hopperia Vitiello, 1969
Laimella Cobb, 1920
Paracomesoma Hope e Murphy, 1972*
Paramesonchium Hopper, 1967*
Pierrickia Vitiello, 1970
Sabatieria Rouville, 1903
Família Coninckiidae Lorenzen, 1981
Coninckia Gerlach, 1956

31
Família Diplopeltidae Filipjev, 1918
Araeolaimus De Man, 1888*
Campylaimus Cobb, 1920
Diplopeltula Gerlach, 1950
Diplopeltis Cobb in Stiles & Hassal, 1905*
Southerniella Allgén, 1932*
ORDEM PLECTIDA Malakhov, 1982
Subordem Leptolaimina Chitwood, 1933
Superfamília Leptolaimoidea Orley, 1880
Família Leptolaimidae Orley, 1880
Antomicron Cobb, 1920
Camacolaimus De Man, 1889
Halaphanolaimus Southern, 1914*
Leptolaimus De Man, 1876*
Procamacolaimus Gerlach, 1954*
Família Aegialoalaimidae Lorenzen, 1981
Aegialoalaimus De Man, 1907
Família Diplopeltoididae Tchesunov, 1990
Diplopeltoides Gerlach, 1962
Família Paramicrolaimidae Lorenzen, 1981
Paramicrolaimus Wieser, 1954*
Superfamília Ceramonematoidea Cobb, 1933
Família Tarvaiidae Lorenzen, 1981
Tarvaia Allgén, 1934*
Família Ceramonematidae Cobb, 1933
Ceramonema Cobb, 1920
Metadasynemella De Conick, 1942
Pselionema Cobb, 1933

Superfamília Haliplectoidea Chitwood, 1951
Família Haliplectidae Chitwood, 1933
Haliplectus Cobb, 1913 *
Setoplectus Vitiello, 1971*
ORDEM RHABIDITIDA Chitwood, 1933
Subordem Rhabditina Chitwood, 1933
Superfamília Rhabditoidea Örley, 1880
Família Rhabditidae Örley, 1880
Rhabditis Dujardin, 1845*
* Gêneros onde só foram encontrados individuos fêmeas ou juvenis.
** A posição filetica desta família ainda é incerta, por isso ela é considerada a parti neste
trabalho.
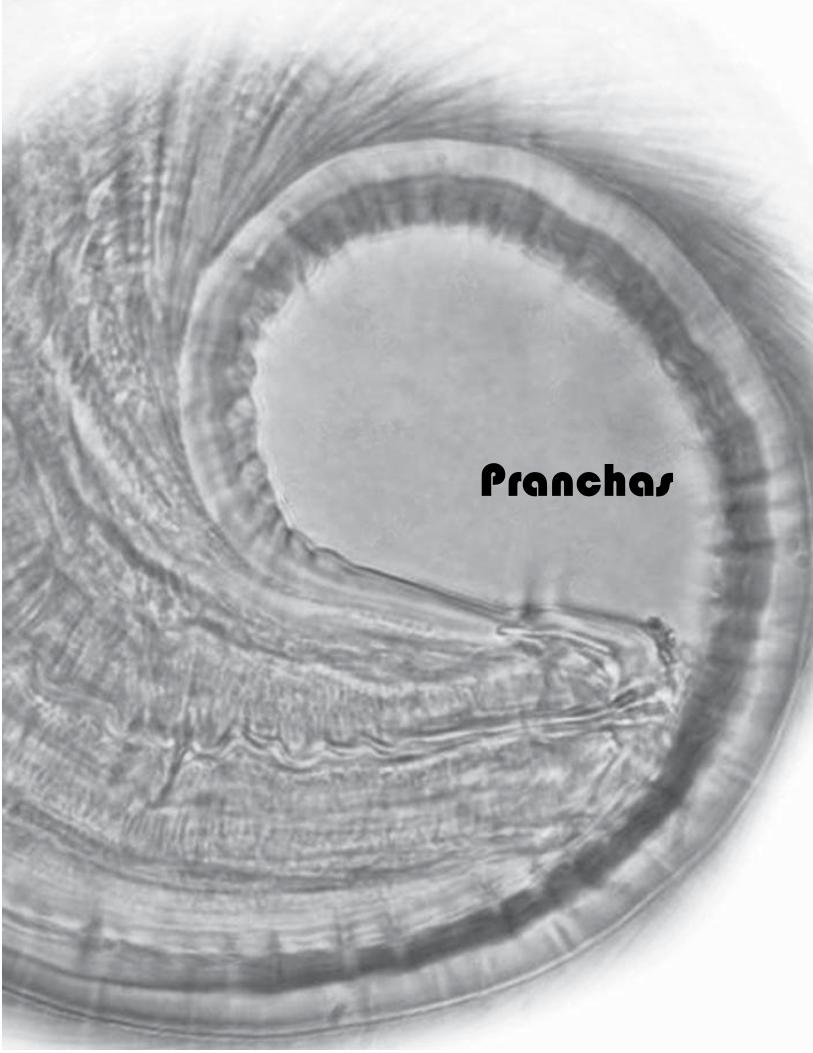
33
Pranchas

OR
DE
M E
NO
PL
IDA
FILIPJEV
, 19
29
A Ordem Enoplida Filipjev,1929 caracteriza-se por apresentar indivíduos
com cutícula lisa ou fracamente estriada, nunca pontuada; anfídio não
espiral com fóvea anfidial em forma de bolso; seis sensilas labiais
internas em forma de papilas ou setas curtas; seis labiais externas e quatro
setas cefálicas em dois ou três círculos; região labial frequentemente
dividida em três lábios; cavidade bucal presente ou ausente, quando
presente: pequena ou grande, com ou sem forte esclerotização, com ou
sem dentes; faringe cilíndrica, ligada à parede da cavidade bucal e/ou à
parede do corpo, em alguns casos levemente alargada na base; glândulas
ventrosublaterais e dorsal abrem-se em setores faringeais ou diretamente
posterior à cavidade bucal; metanemes presentes ou ausentes; fêmeas
didélficas, com ovários opostos antidromous refletidos; machos com
dois testículos opostos; glândulas caudais, quando presentes, abrem-se
em um poro único.
Enoplida integra os maiores nematódeos, que, quando adultos chegam a
medir 3 mm. A maioria é marinha, existindo pouquissímas espécies de
água doce. De acordo com Lorenzen (1994), cuja classificação é baseada
nos metanemes, a Subclasse Enoplia apresenta 2 Ordens: Enoplida e
Trefusiida. Enoplida congrega duas Subordens: Enoplina e Tripyloidina.
Porém, estudos moleculares recentes (De Ley & Blaxter, 2002; 2004)
mostram uma nova classificação, com sete subordens na Ordem Enoplida
e três subordens na Ordem Triplonchida, desconsiderando a Ordem
Trefusiida. (Figura 4).
CARACTERÍSTICAS GERAIS
Figura 4: Parte anterior do corpo de nematódeos típicos de cada subordem de Enoplida
(Smoll & Coomans, 2006) que ocorrem na Bacia de Campos: A= Enoplina; B= Trefusiina;
C= Oncholaimina; D= Ironina; E= Tripyloidina.

36
Epacanthion
Platt, H. M.; Warwick, R. M. (1983) Free-living
Marine Nematodes – Part I British Enoplids. Syn-
opses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 307p.
Greenslade, P.; Nicholas, W. L. (1991) Some
Thoracostomopsidae (Nematoda: Enoplida) from
Australia, including descriptions of two new gen-
era and diagnostic Keys. Invertebr. Taxon., 4, 1031–
1052.
Smol, N.; Coomans, A. (2006) Order Enoplida. In:
Eyualem-Abebe, Traunspurger, W. & Andrássy, I.
(eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 225–292). CABI Publishing, UK.
Cutícula usualmente lisa. Cabeça afunilada ou
em forma de cone. Arranjo cefálico 6+6+4.
Seis setas labiais internas, longas e inseridas
na base dos lábios, seis setas labiais externas
longas e quatro setas cefálicas curtas. Setas
labiais externas e cefálicas inseridas na metade
ou na porção superior da cápsula cefálica.
Setas cervicais freqüentemente presentes.
Lábios altos, marcados por um lobo acessório
com estriações amplas. Estriação semi-lunar
presente. Mandíbulas consistindo de dois
tubos laterais fortemente cuticularizados
com uma lacuna central, conectados
anteriormente por uma fina barra curva.
Anfídio em forma de bolso. Faringe
relativamente longa e cilíndrica. Cárdia
piriforme. Fêmeas didélficas, com ovários
opostos refletidos do lado esquerdo do
intestino. Machos com dois testículos do lado
esquerdo do intestino. Espículas levemente
curvadas. Gubernáculo simples. Um
suplemento pré-cloacal presente. Três
glândulas caudais dispostas antes da cloaca.
Cauda cônica.
1050 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Wieser, 1953
Indivíduos que podem medir de 2 a 4 mm.
As espículas podem se apresentar em
diferentes comprimentos de acordo com a
espécie sendo usualmente longas (e” 2,5
diâmetros anais). A revisão mais recente
que inclui este gênero foi feita por
Greenslade & Nicholas em (1991).
Comentários
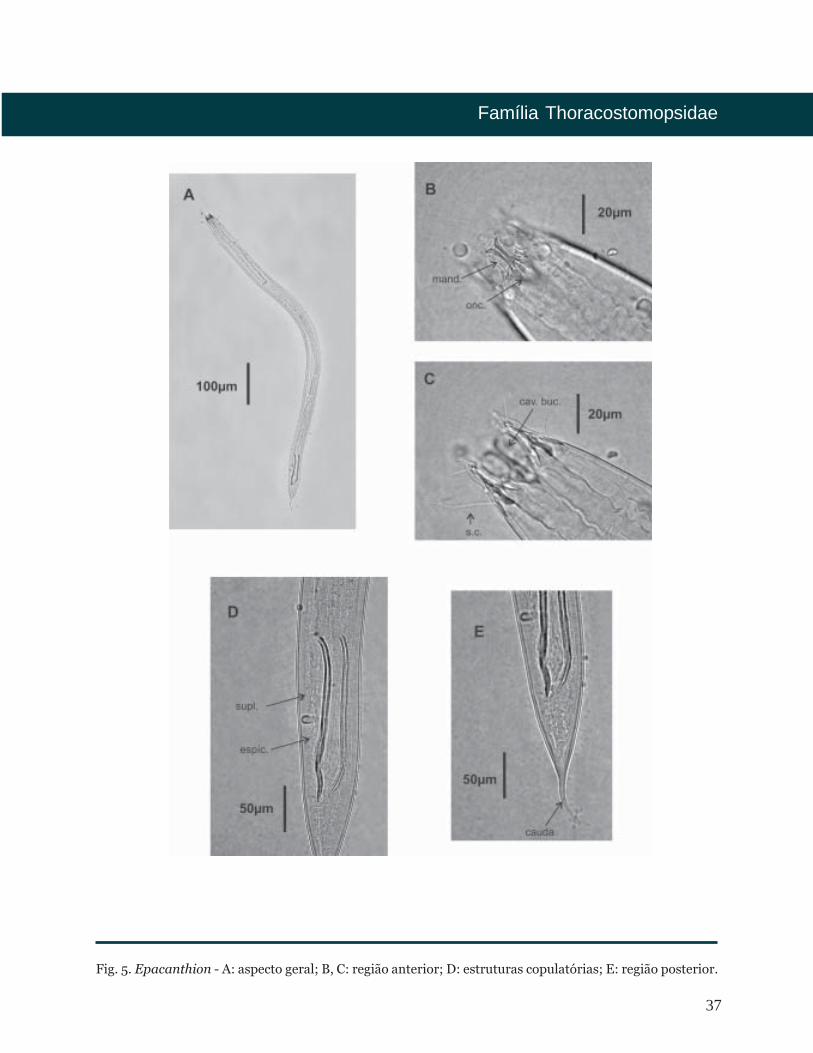
37
Fig. 5. Epacanthion - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Thoracostomopsidae

38
Mesacanthion
Warwick, R. M. (1970) Fourteen new species of
marine nematodes from Exe estuary. Bull. Br. Mus.
nat. Hist. (Zool.) 19: pp137–177.
Platt, H. M.; Warwick, R. M. (1983) Free-living
Marine Nematodes – Part I British Enoplids. Syn-
opses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 307p.
Smol, N.; Coomans, A. (2006) Order Enoplida. In:
Eyualem-Abebe, Traunspurger, W. & Andrássy, I.
(eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 225–292). CABI Publishing, UK.
Cutícula lisa ou com leve estriação. Região
anterior esclerotizada formando uma
cápsula cefálica. Arranjo cefálico em três
círculos diferentes: seis setas labiais
internas, seis setas labiais externas e quatro
setas cefálicas posicionadas na região an-
terior da cápsula cefálica. Setas cefálicas
mais longas que as labiais. Inúmeras setas
subcefálicas na extremidade anterior do
corpo. Cavidade bucal contém lábios
relativamente baixos, sem estriações
internas; três mandíbulas em forma de
gancho; e uma onchia dorsal. Anfídio em
forma de bolso. Fêmeas com ovários
refletidos pareados. Machos com
testículos pareados, opostos e estendidos.
Espículas geralmente pequenas e em
algumas espécies aberta distalmente.
Gubernáculo presente ou ausente, com
ou sem apófise. Suplemento pré-cloacal
presente ou ausente. Machos de algumas
espécies com dois pares de setas pós-
cloacais. Cauda cônica afilada na
extremidade posterior.
1050, 1350 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Filipjev, 1927
Mesacanthion possui um arranjo de setas
na região anterior similar ao
Enoplolaimus. No entanto, em
Mesacanthion as setas cefálicas estão na
porção anterior da cápsula cefálica,
enquanto em Enoplolaimus estão na parte
posterior.
Comentários
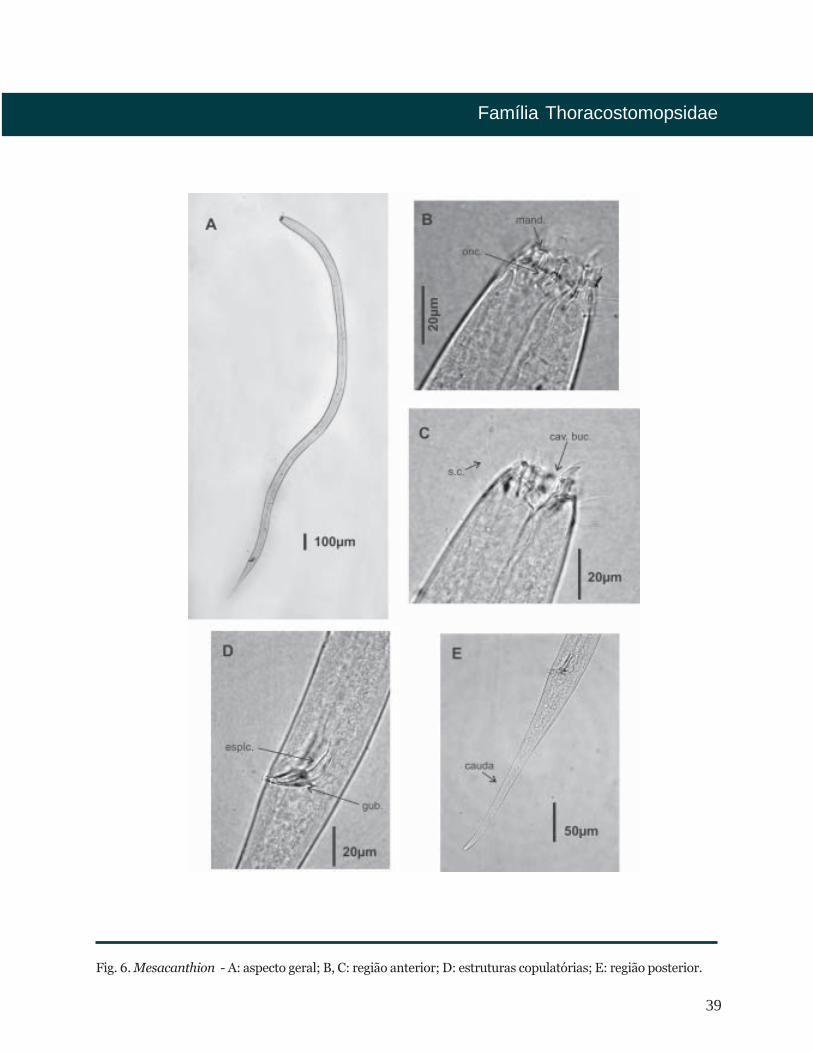
39
Fig. 6. Mesacanthion - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Thoracostomopsidae

40
Micoletzkya
Wieser, W. (1953) Free-living marine nematodes I.
Enoploidea. Lunds Universitets Arsskrift. N. F. Avd.
2. Bd 49. Nr 6.
Platt, H. M.; Warwick, R. M. (1983) Free-living
Marine Nematodes – Part I British Enoplids. Syn-
opses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 307p.
Cutícula lisa. Cápsula cefálica
fracamente desenvolvida. Arranjo cefálico
6+10. Seis setas labiais externas e quatro
cefálicas em um mesmo círculo. Setas
somáticas ausentes. Anfídio em forma de
bolso após as setas cefálicas. Cavidade
bucal pequena e simples. Faringe cilíndrica
e sem bulbo basal. Poro excretor abre-se
imediatamente após a cavidade bucal.
Fêmeas didélficas com ovários opostos
e refletidos. Machos com dois testículos
opostos. Espículas longas.
Gubernáculo simples e sem apófise.
Um único suplemento pré-cloacal
presente. Cauda longa, cônico-cilíndrica.
750, 1050, 1350 e 1950 metros.
O comprimento total dos machos deste
gênero pode variar de 3 a 9 mm.
Comentários
Ditlevsen, 1926
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia

41
Fig. 7. Micoletzkya - A: aspecto geral; B: região anterior; C: estruturas copulatórias; D: região posterior.
Família Phanodermatidae

42
Cephalanticoma
Inglis, W. G. (1964) The Marine Enoplida (Nema-
toda): a comparative study of the head. Bull. Brit.
Mus. nat. Hist. (Zoology): II (4): 266–376.
Platt, H. M.; Warwick, R. M. (1983) Free-living
Marine Nematodes – Part I British Enoplids. Syn-
opses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 307p.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
167p.
Smol, N.; Coomans, A. (2006) Order Enoplida. In:
Eyualem-Abebe, Traunspurger, W. & Andrássy, I.
(eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 225–292). CABI Publishing, UK.
Cutícula lisa. Metanemes de forma
variável. Cápsula cefálica bem
desenvolvida com a musculatura faringeal
inserida na mesma. Seis papilas labiais
internas, seis setas labiais externas e quatro
cefálicas. Grupo de setas cervicais situadas
lateralmente no corpo. Cavidade bucal
pequena com três dentes na porção ante-
rior e circundada por tecido faringeal.
Anfídio em forma de bolso. Contorno da
faringe liso. Sistema excretor-secretor
posicionado à esquerda da faringe. Poro
excretor-secretor bem visível e posterior às
setas cervicais. Sistema reprodutivo das
fêmeas didélfico com ovários antidromous
opostos refletidos. Machos, geralmente com
dois testículos ou com um único testículo
anterior. Espículas curvadas com ala ven-
tral. Gubernáculo com peças laterais
acessórias e sem apófise. Um único
suplemento pré-cloacal tubular
presente. Duas fileiras de setas ventrais en-
tre o suplemento e a abertura do ânus.
Glândulas caudais restritas à região caudal.
Cauda filiforme com setas na região termi-
nal.
750 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Inglis (1964) não considera a presença da
cápsula cefálica como uma forte
característica para o gênero, pois em
alguns espécimes pode não estar
claramente visível.
Comentários
Platonova, 1976
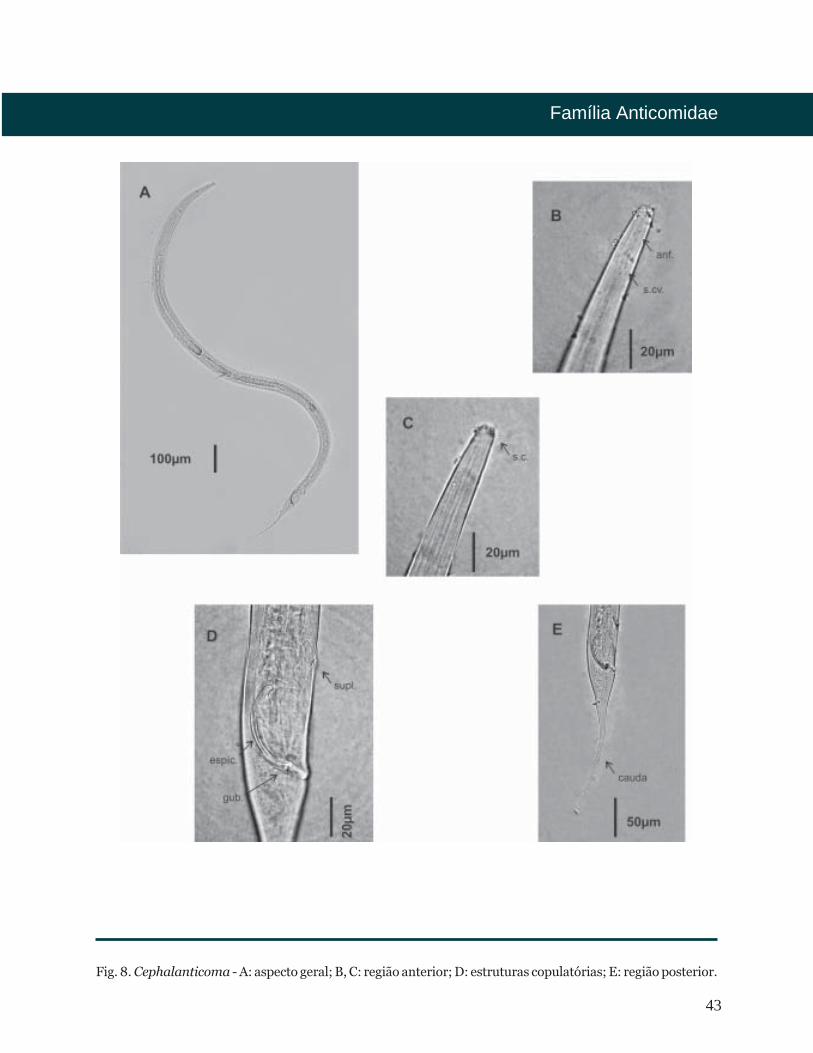
43
Fig. 8. Cephalanticoma - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Anticomidae

44
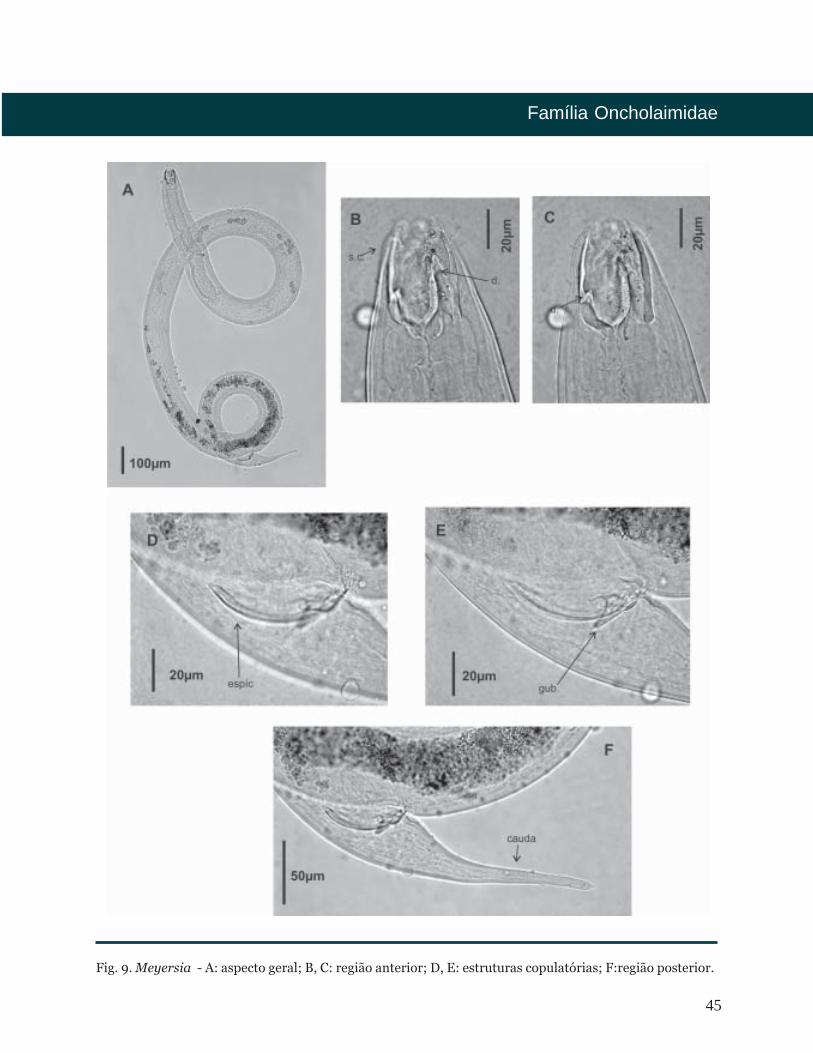
Meyersia
Hopper, B. E. (1967) Free-living marine nematodes
from Biscayne Bay, Florida, II. Oncholaimidae: de-
scriptions of five new species and of one new genus
(Meyersia). Mar. Biol. I (2): 145–151.
Platt, H. M.; Warwick, R. M. (1983) Free-living
Marine Nematodes – Part I British Enoplids. Syn-
opses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 307p.
Smol, N.; Coomans, A. (2006) Order Enoplida. In:
Eyualem-Abebe, Traunspurger, W. & Andrássy, I.
(eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 225–292). CABI Publishing, UK.
Cutícula lisa. Arranjo cefálico em dois
círculos de seis setas labiais internas e dez
setas cefálicas. Anfídio em forma de
bolso. Cavidade bucal grande, fortemente
cuticularizada, com três dentes. Dois
dentes grandes de mesmo diâmetro e
comprimento na região subventral, e um
dente menor dorsal. Fêmeas didélficas
com ovários refletidos pareados.
Sistema demaniano bem
desenvolvido. Machos com testículos
opostos e estendidos. Espículas simples.
Gubernáculo presente. Glândulas
caudais anterior ao ânus. Cauda cônico-
clíndrica. Espinerete visível.
1650 metros.
De Man, 1890
Bibliografia
Distribuição Batimétrica
Diagnose Distribuição Geográfica
O número e a posição dos dentes, a
estrutura do sistema reprodutor da fêmea
e o nível de desenvolvimento do sistema
demaniano são importantes
características diagnósticas para o gênero.
Comentários
45
Fig. 9. Meyersia - A: aspecto geral; B, C: região anterior; D, E: estruturas copulatórias; F:região posterior.
Família Oncholaimidae

46
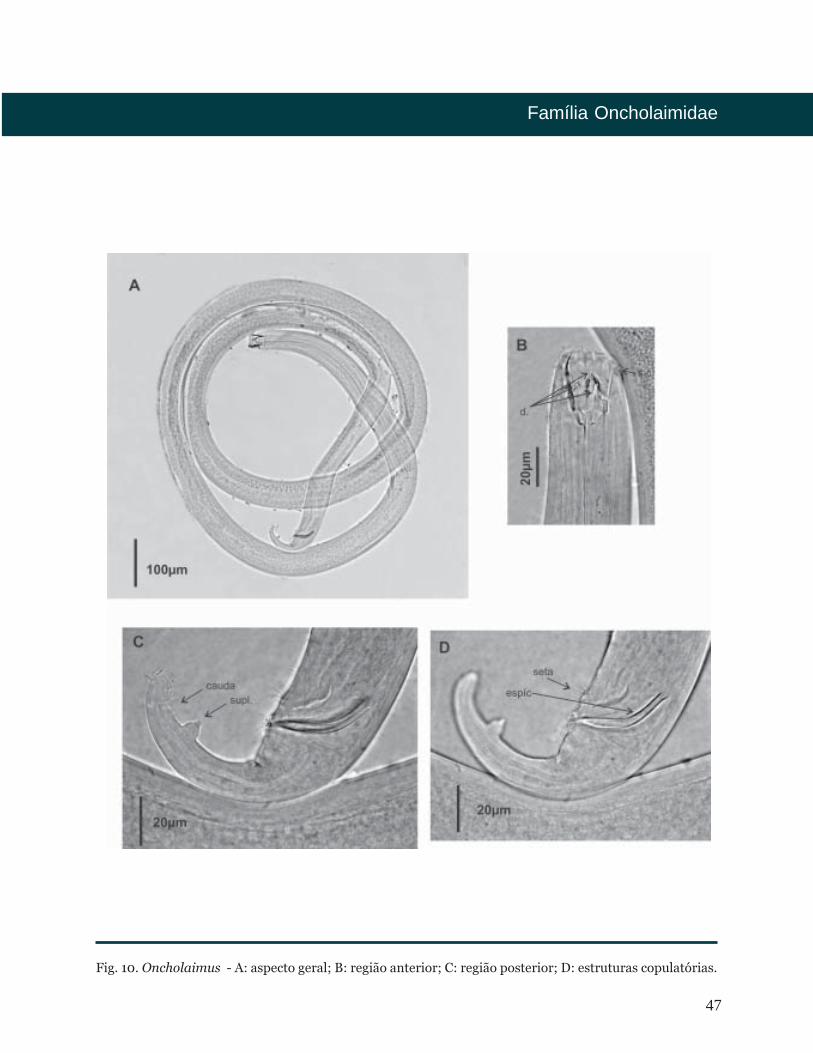
Oncholaimus
De Coninck, L. A.; Shuurmans Stekhoven, J. H.
(1933) The freeliving marine nemas of the Belgian
Coast II Mém. Mus. r. Hist. nat. Belg. 58: 1–163.
Kreis, H. A. (1934) Oncholaiminae Filipjev, 1916 Eine
monongraphische Studie Capita zoo. 4 (5): 1–271.
Platt, H. M.; Warwick, R. M. (1983) Free-living
Marine Nematodes – Part I British Enoplids. Syn-
opses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 307p.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
167p.
Smol, N.; Coomans, A. (2006) Order Enoplida. In:
Eyualem-Abebe, Traunspurger, W. & Andrássy, I.
(eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 225–292). CABI Publishing, UK.
Cutícula lisa. Setas labiais externas e
cefálicas em um único círculo. Setas
cervicais presentes ou ausentes. Anfídio
em forma de bolso. Cavidade bucal
fortemente cuticularizada com um dente
esquerdo grande subventral e dois dentes
menores, sendo um dorsal e outro
subventral. Fêmeas monodélficas-
prodélficas com ovários refletidos e
antidromous. Sistema demaniano
presente. Machos com dois testículos
opostos à direita do intestino. Espículas
curtas de forma simples, levemente
curvadas ventralmente. Gubernáculo
ausente. Suplementos pré-cloacais,
em forma de papilas e setas que circundam
a cloaca. Um proeminente suplemento
pós-cloacal localizado na região caudal.
Três glândulas caudais. Cauda curta e
usualmente curvada. Espinerete visível.
750, 1050, 1350 e 1950 metros.
Distribuição Geográfica
As espécies de Oncholaimus são facilmente
separadas em dois grupos em relação à
forma da cauda: indivíduos com a cauda
curta e ventralmente curvada e aqueles
com cauda longa e reta.
Comentários
Dujardin, 1845
Distribuição Batimétrica
Bibliografia
Diagnose
47
Fig. 10. Oncholaimus - A: aspecto geral; B: região anterior; C: região posterior; D: estruturas copulatórias.
Família Oncholaimidae

48
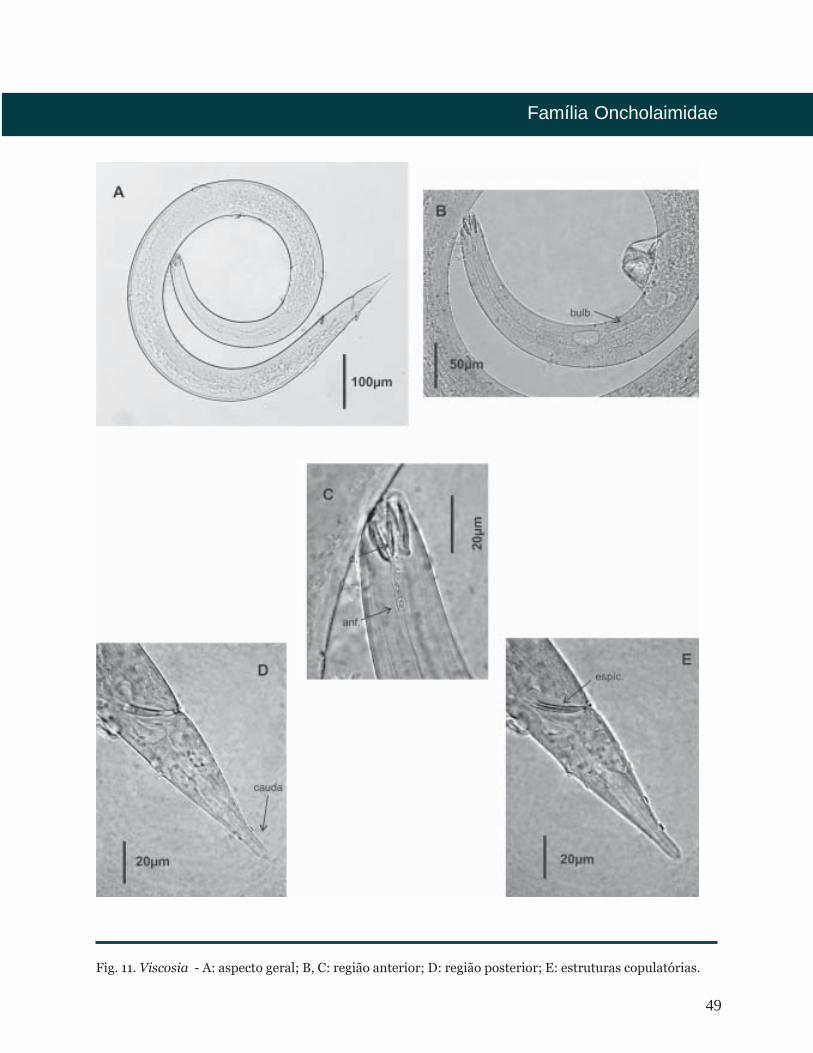
Viscosia
Shuurmans Stekhoven, J. H. (1950) The freeliving
marine nemas of the Mediterraneam. I. The Bay of
Villefranche. Mém. Inst. r. Sci. nat. Bel. 2 Sér. 37:
1–220.
Platt, H. M.; Warwick, R. M. (1983) Free-living
Marine Nematodes – Part I British Enoplids. Syn-
opses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 307p.
Smol, N., & Coomans, A. (2006) Order Enoplida.
In: Eyualem-Abebe, Traunspurger, W. & Andrássy,
I. (eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 225-292). CABI Publishing, UK.
Cutícula lisa. Região anterior formada por
seis lábios com seis papilas labiais, seis
setas labiais externas e quatro cefálicas.
Anfídio em forma de bolso. Cavidade
bucal cuticularizada com um grande
dente subventral e outros dois dentes
menores, um dorsal e outro subventral.
Faringe cilíndrica sem bulbo basal. Fêmeas
com ovários pareados. Sistema
demaniano presente. Machos
geralmente com dois testículos opostos.
Espículas estreitas, relativamente curtas.
Gubernáculo geralmente ausente. Cauda
cônico-cilíndrica.
750, 1050, 1350, 1650 e 1950 metros.
Distribuição Geográfica
A ausência de gubernáculo é uma
característica genérica, porém existem
exceções com em V. shuurmansi.
Comentários
De Man, 1889
Distribuição Batimétrica
Bibliografia
Diagnose
49
Fig. 11. Viscosia - A: aspecto geral; B, C: região anterior; D: região posterior; E: estruturas copulatórias.
Família Oncholaimidae

50
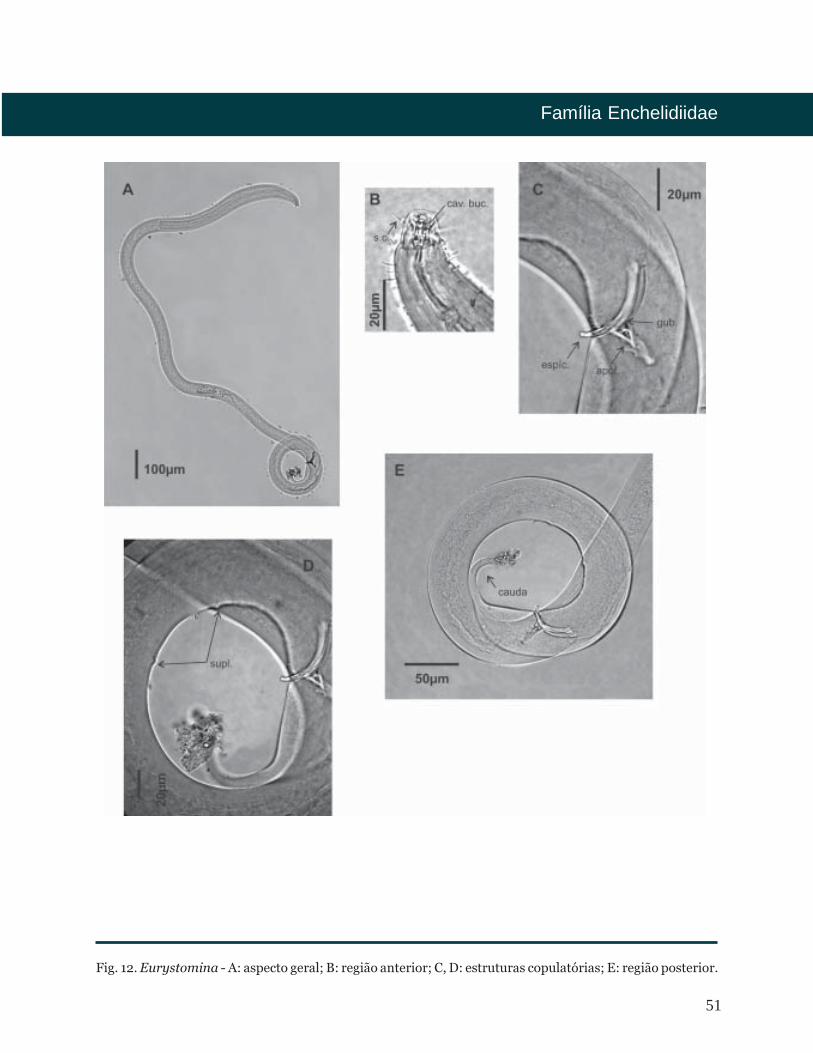
Eurystomina
Wieser, W. (1953) Free-living marine nematodes.
I. Enoploidea. Chile reports 10. Lunds Univenztets
Arsskrift 49, 155 pp.
Platt, H. M.; Warwick, R. M. (1983) Free-living
Marine Nematodes – Part I British Enoplids. Syn-
opses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 307p.
Smol, N.; Coomans, A. (2006) Order Enoplida. In:
Eyualem-Abebe, Traunspurger, W. & Andrássy, I.
(eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 22–292). CABI Publishing, UK.
Cutícula lisa. Região cefálica com um
círculo de seis papilas labiais internas e um
segundo círculo formado por dez setas.
Cavidade bucal em forma de V, contendo
na região anterior de 1 a 5 fileiras de
dentículos. Na porção posterior um grande
dente subventral e dois dentes menores:
um dorsal e outro subventral. Anfídio em
forma de bolso com posição variável.
Ocelo presente em algumas espécies.
Faringe sem bulbo terminal. Posição do
poro secretor-excretor variável. Fêmeas
com dois ovários opostos e refletidos.
Machos com dois testículos opostos.
Espículas arqueadas. Gubernáculo
perpendicular às espículas, com ou sem
apófise. Dois suplementos pré-cloacais
cuticularizados em forma de asa. Três
glândulas caudais posterior ao ânus. Cauda
cônico-cilíndrica com a região cilíndrica
afilada.
750, 1050, 1350, 1650 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Filipjev, 1921
Smol & Coomans (2006) citam que os
suplementos pré-cloacais podem
apresentar a forma de copo com asas
laterais presentes ou ausentes.
Comentários
51
Fig. 12. Eurystomina - A: aspecto geral; B: região anterior; C, D: estruturas copulatórias; E: região posterior.
Família Enchelidiidae

52
Syringolaimus
Timm, R. W. (1963) Marine nematodes of the sub-
order Monhysterina from the Arabian Sea at
Karachi. Proc. helminth. Soc. Wash., 30 (1): 34–49.
Gourbault, N.; Vincx, M. (1985) Deux especes
nouvelles d’Ironidae marins; observations sur lês
spermatozoides flagelles des Nematodes. Bull. Mus.
natn. Hist. nat., Paris, 4e ser., 7, sect. A, (1): 109–
118.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
167p.
Cutícula levemente ou fortemente estriada.
Setas labiais e cefálicas papiliformes
geralmente de difícil visualização ao
microscópio óptico. Cavidade bucal tubu-
lar e alongada. Geralmente três dentes
proeminentes na porção anterior da
cavidade bucal. Dentes podem ser inteiros,
bífidos ou em forma de quela. Anfídio em
forma de bolso. Região basal da faringe
apresenta um verdadeiro bulbo de formato
oval ou redondo. Fêmeas didélficas com
ovários refletidos, raramente
estendidos. Machos, geralmente com dois
testículos opostos. Espículas estreitas
apresentando um capitulum e
geralmente um velum. Gubernáculo
presente. Três glândulas caudais na porção
anterior da cauda. Cauda longa, cônica ou
cônico-cilíndrica, com a porção posterior
podendo ser filiforme. Espinerete
presente ou não.
750, 1050, 1350, 1650 e 1950 metros.
Distribuição Geográfica
S. renaudae Gourbault & Vincx, 1985 foi
um dos primeiros casos registrados na
literatura relacionados à presença de
espermatozóides flagelados em Nematoda.
Comentários
De Man, 1888
Distribuição Batimétrica
Bibliografia
Diagnose
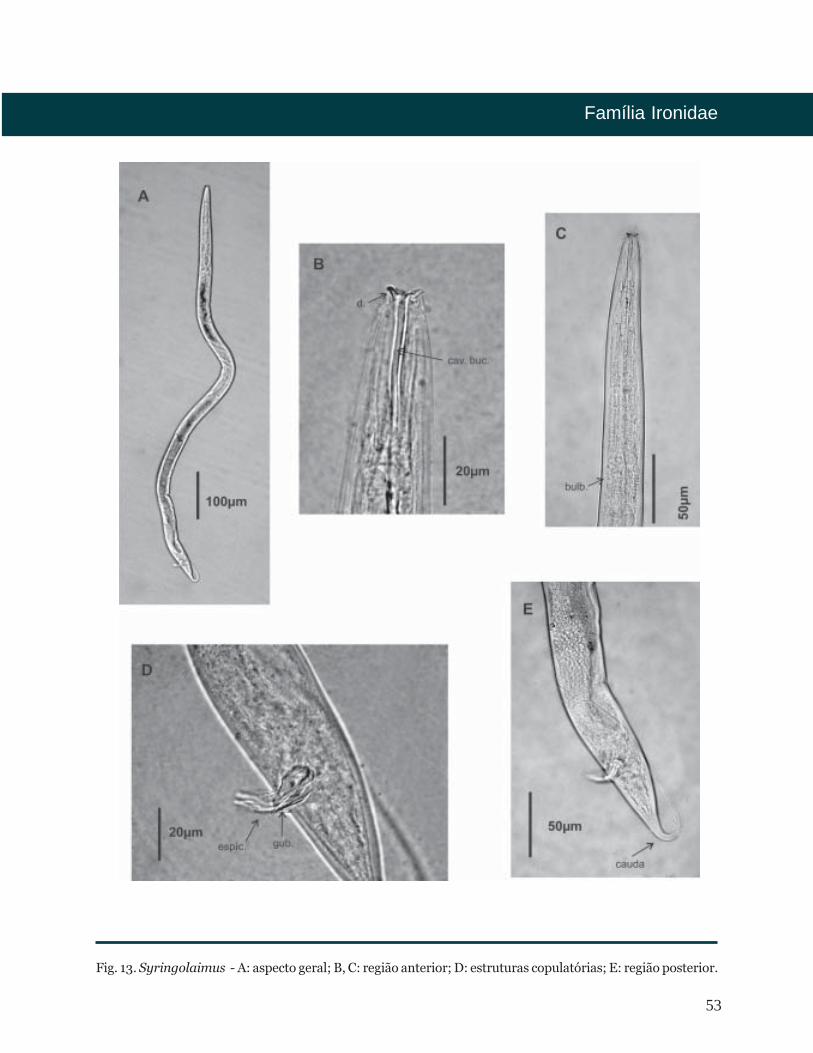
53
Fig. 13. Syringolaimus - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Ironidae

54
Cylicolaimus
Vitiello, P. (1970) Nematodes libres marins des
vases profondes du golfe du Lion. I. Enoplida.
Tethys, 2(1): 139–210.
Platt, H. M.; Warwick, R. M. (1983) Free-living
Marine Nematodes – Part I British Enoplids. Syn-
opses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 307p.
Smol, N.; Coomans, A. (2006) Order Enoplida. In:
Eyualem-Abebe, Traunspurger, W. & Andrássy, I.
(eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 225–292). CABI Publishing, UK.
Cutícula lisa. Cápsula cefálica bem
definida com arranjo de seis papilas labiais
internas, seguidas por outro círculo de dez
setas cefálicas. Cavidade bucal grande e
tubular com um complicado arranjo de
dentes e outras estruturas. Um círculo de
pequenos dentes na porção anterior da
boca e grandes dentes em diferentes
posições de acordo com a espécie, além de
placas triangulares laterais. Anfídio em
forma de bolso. Faringe com contorno liso.
Poro excretor abrindo-se na cavidade
bucal. Fêmeas didélficas com ovários
opostos e refletidos. Machos com dois
testículos opostos. Espículas curvadas.
Gubernáculo com apófise dorso-cau-
dal. Suplementos pré-cloacais e
espinhos subventrais presentes. Cauda
curta e arredondada.
750 e 1650 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Neste gênero são encontrados indivíduos
longos, medindo de 23 a 30 mm.
Comentários
De Man, 1889
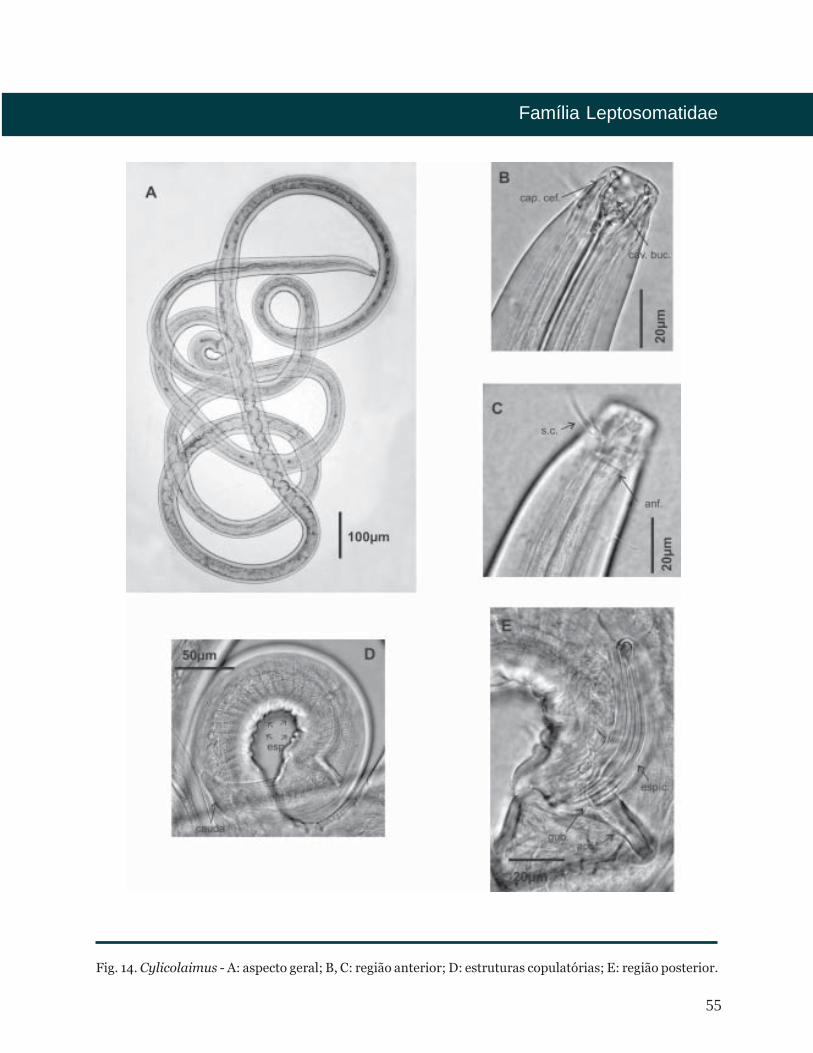
55
Família Leptosomatidae
Fig. 14. Cylicolaimus - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.

56
Halalaimus
Timm, R. W. (1962) A survey of the marine nema-
todes of Chesapeake Bay, Maryland Contr. Chesa-
peake biol. Lab. no 95: 13-70, 13 pls.
Platt, H. M.; Warwick, R. M. (1983) Free-living
Marine Nematodes – Part I British Enoplids. Syn-
opses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 307p.
Keppner, E. J. (1992) Eleven new species of free liv-
ing marine nematodes of the genus Halalaimus De
Man, 1888 (Nematoda, Enoplida) from Florida with
keys to the species. Gul. Res. Resp. Vol 8 (4): 333–
362.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
167p.
Cutícula frequentemente lisa ou levemente
estriada. Estriações podem ser melhor
visualizadas na cauda. Seis setas labiais
externas e quatro setas cefálicas localizadas
no mesmo círculo ou em círculo separados.
Setas cefálicas relativamente menores que
as labiais. Cavidade bucal diminuta ou
ausente. Anfídio estreito e alongado,
posicionado na região anterior do corpo.
Fêmea didélfica com ovários opostos,
antidromous, refletidos, usualmente
ventrais ao intestino. Machos com dois
testículos opostos. Espículas lisas com o
capitulum curvado ventralmente.
Gubernáculo pequeno, restrito a região
posterior da espícula e fortemente
cuticularizado. Seta pré-cloacal com
poro presente ou ausente. Três glândulas
caudais presentes. Cauda cônica-cilíndrica
com a segunda parte filiforme e alongada.
Ala caudal presente ou ausente.
750, 1050, 1350, 1650 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Keppner (1992) criou uma chave de
identificação para as espécies,
considerando a divisão do gênero em 4
grupos. Esta divisão é baseada na presença
ou ausência da ala caudal, da seta pré-cloa-
cal e do poro pré-cloacal.
Comentários
De Man, 1888

57
Fig. 15. Halalaimus - A: aspecto geral; B, C: região anterior; D: região posterior; E: estruturas copulatórias.
Família Oxystominidae

58
Litinium
Cobb, N. A. (1920) One hundred new nemas. (Type
species of 100, new genera). Contr. Sci. Nematol.
(Cobb), (9): 217–343.
Platt, H. M.; Warwick, R. M. (1983) Free-living
Marine Nematodes – Part I British Enoplids. Syn-
opses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 307p.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
167p.
Smol, N.; Coomans, A. (2006) Order Enoplida. In:
Eyualem-Abebe, Traunspurger, W. & Andrássy, I.
(eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 225–292). CABI Publishing, UK.
Cutícula lisa. Setas cefálicas após o
anfídio. Cavidade bucal pequena ou
ausente. Anfídio apresenta dimorfismo
sexual: machos com anfídio em forma de
ferradura e fêmeas com anfídio circular
contornado por uma linha em forma de
coração aberto na região posterior. Cárdia
indistinta. Intestino formado por paredes
grossas com lúmen pouco visível. Fêmeas
didélficas com ovários opostos,
reflexos e antidromous. Machos com
dois testículos opostos. Espículas curtas.
Gubernáculo curto e sem apófise.
Glândulas caudais posicionadas em frente
ao ânus. Cauda conóide. Não apresenta
setas caudais.
750, 1050, 1350, 1650 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Nematódeos medindo de 1 a 2 mm.
Geralmente a cavidade bucal destes
indivíduos é pequena ou ausente, no
entanto em L. parmatum Wieser, 1954 a
boca é cilíndrica e fracamente
cuticularizada.
Comentários
Cobb, 1920

59
Fig. 16. Litinium - A: aspecto geral; B, C: região anterior; D: região posterior; E:estruturas copulatórias.
Família Oxystominidae

60
Nemanema
Gerlach, S. A. (1967) Freilebende Meeres-
Nematoden von den Sarso-Inseln (Rotes Meer) 3.
Beitrag der Arbeitsgruppe Litoralforschung. „Me-
teor” ForschErgbn., (D) (2): 19–43.
Platt, H. M.; Warwick, R. M. (1983) Free-living
Marine Nematodes – Part I British Enoplids. Syn-
opses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 307p.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
167p.
Cutícula lisa. Diferenciação lateral
medindo metade da largura do corpo. Seis
setas cefálicas e quatro setas cervicais
presentes. Cavidade bucal ausente.
Anfídio oval com uma estrutura curvada
que se inicia da região anterior através de
duas projeções que se estendem até a
região posterior. Cárdia grande, conóide,
com largura correspondente à metade da
largura do corpo na base do pescoço.
Intestino com largura de 2/3 da largura
do corpo. Poro do sistema secretor-
excretor alongado e a uma curta distância
da constricção da cárdia. Fêmeas
didélficas, com um ovário anterior
rudimentar e um posterior desenvolvido.
Machos com um testículo anterior ventral.
Espículas curvas. Gubernáculo curto.
Glândulas caudais arranjadas em
seqüência, no lado ventral em frente ao
ânus. A anterior estende-se a partir do
ânus a uma distância de três a seis vezes o
tamanho da cauda. Cauda curta e conóide,
mais arqueada no macho que na fêmea.
750, 1050, 1350 e 1650 metros.
Indivíduos longos, medindo de 1 a 6mm.
Este gênero foi por muito tempo subgênero
de Oxystomina, diferenciando-se deste
pela forma da cauda.
Comentários
Cobb, 1920
Distribuição GeográficaDiagnose
Distribuição Batimétrica
Bibliografia

61
Fig. 17. Nemanema - A: aspecto geral; B: região anterior; C: estruturas copulatórias; D: região posterior.
Família Oxystominidae

62
Oxystomina
Timm, R. W. (1967) Some estuarine nematodes from
the Sunderbans. Proc. Pak. Acad. Sci. 4 (1): 1–13.
Tchesunov, A. B. (1978) New free-living nematodes
in the Caspian Sea. Zool. Zh. 57 (4): 505–511.
Platt, H. M.; Warwick, R. M. (1983) Free-living
Marine Nematodes – Part I British Enoplids. Syn-
opses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 307p.
Smol, N.; Coomans, A. (2006) Order Enoplida. In:
Eyualem-Abebe, Traunspurger, W. & Andrássy, I.
(eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 225–292). CABI Publishing, UK.
Cutícula lisa. Diferenciação lateral
corresponde a mais de 1/3 do diâmetro do
corpo. Células ovais visualizadas ao longo
do copo. Setas distribuídas em três círculos
de 6+6+4: seis papilas labiais internas
indistintas, seis setas labiais externas e
quatro setas cefálicas posicionadas
posteriormente devido à região do pescoço
ser bastante alongada. Cavidade bucal
ausente ou muito pequena. Anfídio
tipicamente oval, com uma estrutura curva
em torno da margem anterior que se
estende posteriormente em duas projeções,
algumas vezes maior nos machos do que nas
fêmeas. Faringe estreita na porção anterior,
expandindo-se gradualmente até a porção
posterior. Cárdia presente ou ausente.
Poro secretor-excretor usualmente
visualizado. Fêmeas com apenas o ovário
posterior desenvolvido e algumas vezes
apresentam apenas um resquício do ovário
anterior. Machos com um ou dois testículos.
Espículas curvas. Gubernáculo quase
sempre presente. Glândulas caudais
contidas na cauda ou posicionadas em
frente ao ânus. Cauda usualmente clavada.
750, 1050, 1350 e 1650 metros.
Distribuição Geográfica
São indivíduos que medem de 1,5 a 3 mm.
As espécies são separadas principalmente
pelo comprimento e posição do anfídio,
além das setas cefálicas e cervicais, contudo
estas últimas podem estar reduzidas e
Filipjev, 1921
Distribuição Batimétrica
Bibliografia
Diagnose
freqüentemente podem não ser
visualizadas em algumas espécies. A cauda,
mesmo sendo geralmente clavada pode ser
filiforme com em O. chitwoodi Timm, 1967.
Comentários

63
Fig. 18. Oxystomina - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Oxystominidae

64
Thalassoalaimus
Wieser, W. (1953) Free-living marine nematodes.
I. Enoploidea. Chile reports 10. Lunds Univenztets
Arsskrift 49, 155 pp.
Vitiello, P. (1970) Nématodes libres marins des
vases profondes du Golfe de Lion. 1 Enoplida. Téthys
2 (1) : 139–210, 23 pls.
Smol, N., & Coomans, A. (2006) Order Enoplida.
In: Eyualem-Abebe, Traunspurger, W. & Andrássy,
I. (eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 225-292). CABI Publishing, UK.
Cutícula lisa. Região anterior com doze
setas cefálicas em dois círculos
separados. Quatro setas cervicais poste-
rior ao anfídio. Cavidade bucal
pequena. Dimorfismo sexual
relacionado ao anfídio que apresenta a
forma de ferradura nas fêmeas e de bolso
nos machos. Faringe longa. Poro do
sistema secretor-excretor visível.
Fêmeas, geralmente com um ovário pos-
terior. Machos com testículos pareados
e refletidos. Espículas curvadas.
Gubernáculo curto. Suplementos
pré-cloacais geralmente
papiliformes. Glândulas caudais
longas, posterior ao ânus. Cauda
cônica com a região terminal
fortemente cuticularizada (cápsula
caudal).
750, 1050, 1350 e 1650 metros.
Distribuição Geográfica
Em sua maioria, é composto por espécies
marinhas, apresentando apenas uma única
espécie de água doce.
Comentários
De Man, 1893
Distribuição Batimétrica
Bibliografia
Diagnose
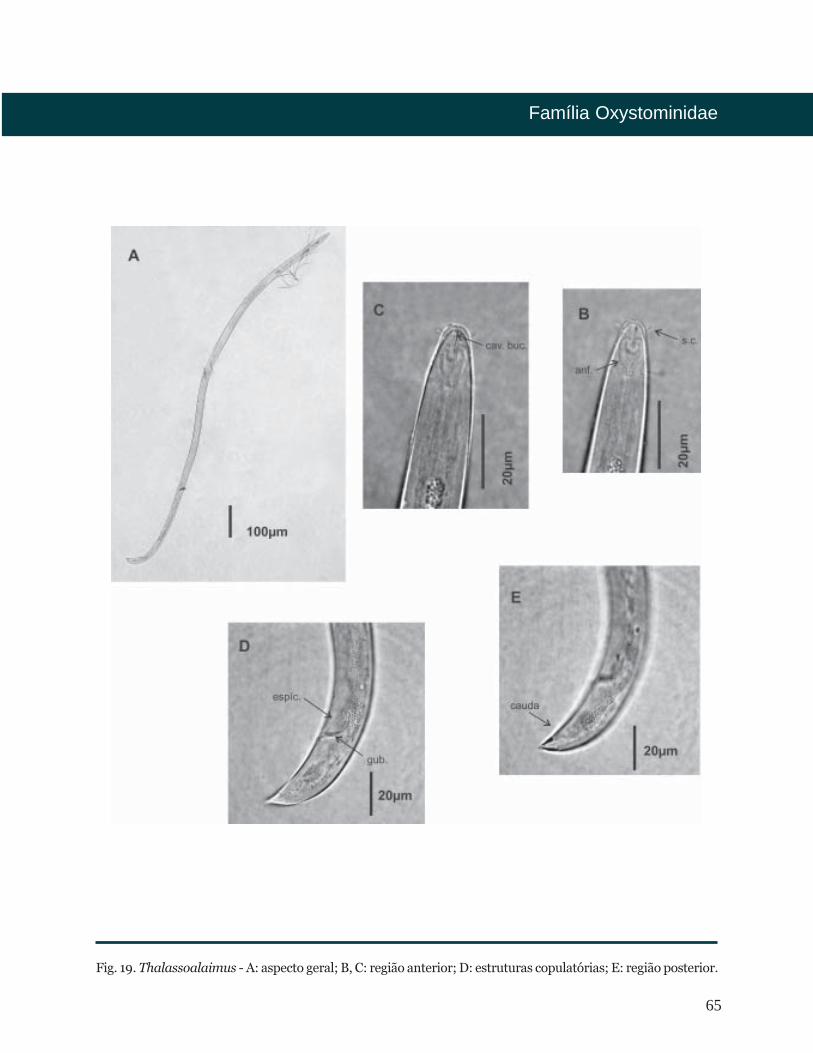
65
Fig. 19. Thalassoalaimus - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Oxystominidae

OR
DE
M C
HR
OM
AD
OR
IDA
CHITWOOD
, 19
33
A Ordem Chromadorida é caracterizada por apresentar cutícula
com ornamentações que podem variar desde pontuações até estriações
transversais, frequentemente com diferenciação lateral; anfídio com
diferentes formas (espiral, multiesperal, loops simples ou alongados);
setas cefálicas em três círculos (6 + 6 + 4) ou em dois círculos separados
(6 + 10); cavidade bucal com 12 rugas, um dente dorsal e dois dentes
menores subventrais; faringe cilíndrica com um bulbo posterior
fortemente muscular e esclerotizado; glândula ventral com ducto
terminal cuticularizado; fêmeas didélficas com ovários opostos
ou não; machos com um ou dois testículos, espículas pareadas,
gubernáculo e suplementos pré-cloacais; glândulas caudais e
espinerete presentes.
É composta essencialmente por Nematoda marinhos (Decraemer &
Smol, 2006). De acordo com Lorenzen (1994), a Ordem Chromadorida
possui apenas a Subordem Chromadorina com a única Superfamília
Chromadoroidea. Dentro desta Superfamília, seis Famílias são incluídas:
Chromadoridae, Ethmolaimidae, Neotonchidae, Achromadoridae,
Cyatholaimidae and Selachinematidae. Na classificação proposta por De Ley
& Blaxter (2002), a Família Selachinematidae não foi incluída (Figura 5).
CARACTERÍSTICAS GERAIS
A B C D
Figura 20: Porção anterior do corpo de gêneros representantes das Famílias de
Chromadorida (Decraemer & Smoll, 2006)que ocorrem na Bacia de Campos: A=
Chromadoridae; B= Neotonchidae; C= Cyatholaimidae; D= Selachinematidae.

68
Acantholaimus
Gourbault, N.; Vincx, M. (1985) Nématodes
abyssaux (Campagne Walda du N/O “Jean Char-
cot”) VI. Le genre Acantholaimus: espèces
nouvelles et étude de l’appareil reproducteur à
apermatozoïdes géants. Bull. Mus. natn. Hist. nat.,
Paris, 4e sér., 7, section A, no 3: 611–632.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Muthumbi. A. W.; Vincx, M. Acantholaimus
(chromadoridae:nematoda) from Indian Ocean:
description of seven espécies. Hydrobiologia
346:59–76.
Ornamentação da cutícula de forma
variável: lisa, levemente pontuada e até
fortemente pontuada. Diferenciação lateral
quando presente, formada por duas fileiras
de pontos maiores longitudinais. Arranjo
cefálico formado por seis setas labiais
internas, seis labiais externas e quatro
cefálicas. Comprimento das setas da região
cefálica variável. Setas somáticas
presentes. Cavidade bucal pequena ou
grande com três ou mais dentes sólidos.
Anfídio circular ou crípticoespiral.
Faringe com bulbo basal presente ou
ausente. Cárdia curta. Fêmeas didélficas
com ovários opostos e refletidos.
Ovário anterior à direita do intestino e o
posterior à esquerda. Machos com um
único testículo estendido anterior.
Espículas curvadas, achatadas e infladas
na porção proximal, com uma apófise
ventral nesta região. Gubernáculo
presente ou ausente. Cauda cônica na
porção anterior e filiforme na região pos-
terior.
750, 1050, 1350, 1650 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Allgén, 1933
Acantholaimus é um gênero típico de
sedimentos dos mares profundos em todo
o mundo.
Comentários
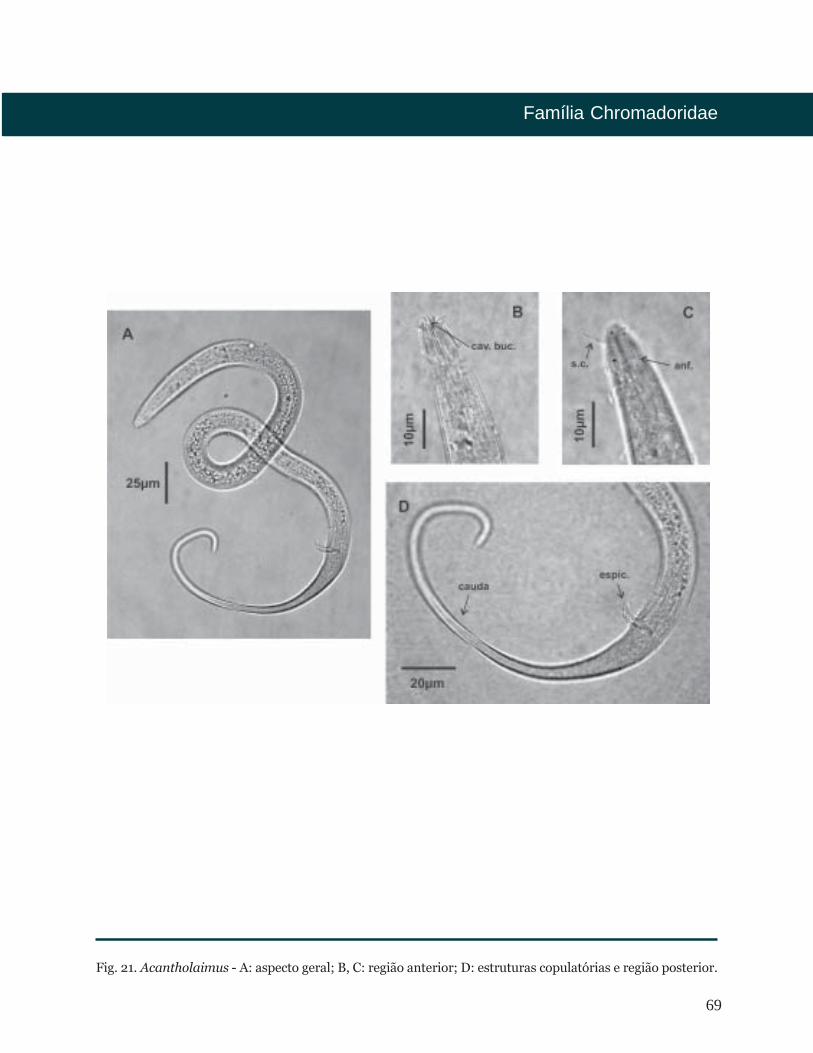
69
Fig. 21. Acantholaimus - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias e região posterior.
Família Chromadoridae

70
Actinonema
Timm, R. W. (1961) The marine nematodes of the
Bay of Bengal. Proc. Pakist. Acad. Sci. 1: 1–88.
Boucher, G. (1976). Nématodes dês sables fins
infralittoraux de La Pierre Noire (Manche
occidentale). II. Chromadorida. Bull. Mus. Hist.
Nat. Paris, 3e ser, No. 352, Zool., 245, 25-61.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Jensen, P. (1991) Nine new and less known nema-
tode species from the deep-sea benthos of the Nor-
wegian Sea. Hydrobiologia, 222, 57–76.
Ornamentação cuticular heterogênea e
complexa: pontuações irregulares ao redor
do anfídio, transversais homogênea na
porção anterior da faringe e com
diferenciação lateral a partir de
aproximadamente 60% do comprimento
total da faringe. Arranjo cefálico em dois
círculos, o primeiro com seis setas labiais
internas e o segundo de dez setas cefálicas.
Anfídio oval com contorno duplo e
posicionado transversalmente na região
cefálica. Cavidade bucal com rugas
esclerotizadas; um dente dorsal de
tamanho mediano e dois dentes
subventrais diminutos. Faringe cilíndrica
com bulbo basal. Fêmeas didélficas com
ovários opostos e refletidos. Machos
com um testículo estendido anterior em
posição variável ao intestino. Espículas
delgadas e curvadas, fortemente
esclerotizadas. Gubernáculo com
telamons em forma de “L”. Cauda cônica.
750, 1050, 1350, 1650 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Cobb, 1920
Boucher (1976) descrevendo A. celtica
mostrou através de desenhos a variação do
padrão cuticular ao longo do corpo do
indivíduo.
Comentários
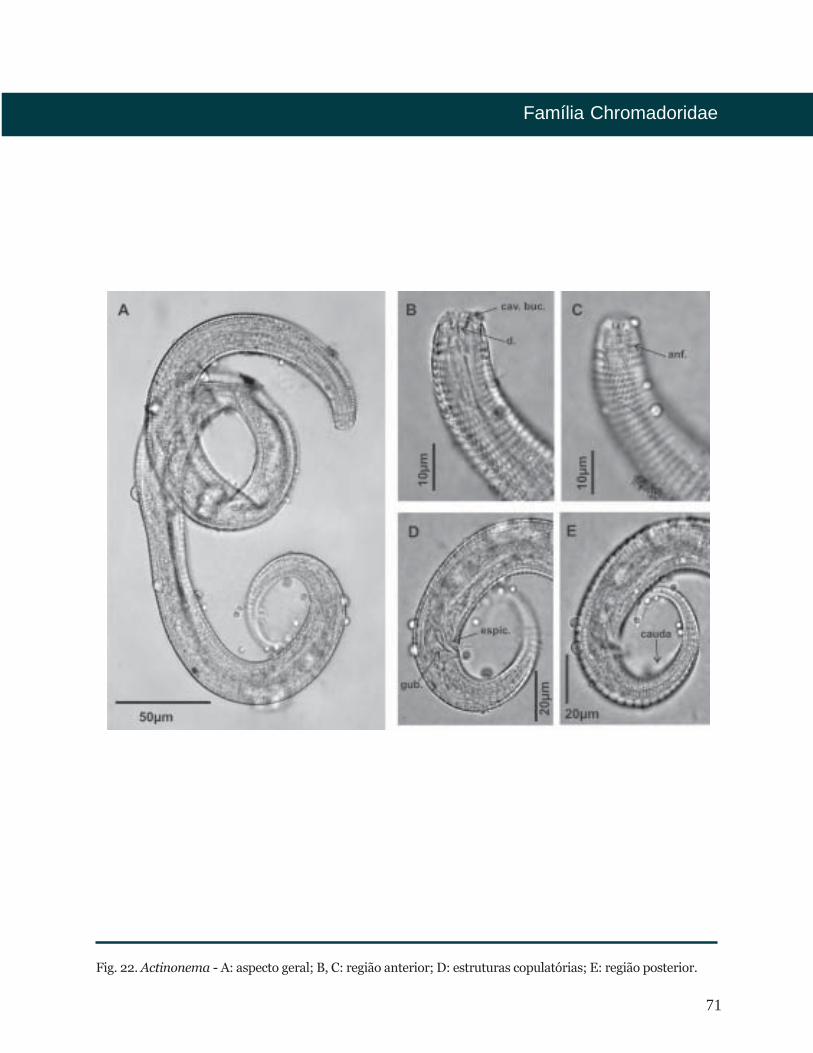
71
Família Chromadoridae
Fig. 22. Actinonema - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.

72
Chomadorita
Pastor de Ward, C. T. (1984) Tres especies nuevas
de nematodes marinos de vida libre
(Chromadoridae y Comesomatidae) para la Ria
Deseado, Santa Cruz, Argentina. Physis (Buenos
Aires), Secc. A, 42 (102): 39–48.
Lorenzen, S. (1986) Chromadorita ceratoserolis sp.
n. (Chromadoridae), a free-living marine nematode
epibiotically on the Isopod Ceratosolis trilobitoides
from Antarctica. Polar Biol., 6, 247–260.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Cutícula homogênea, com pontuações fortes
e diferenciação lateral presente ou ausente.
Arranjo cefálico de 6+6+4. Setas somáticas
presentes. Anfídio oval ou com um loop oval
horizontal. Cavidade bucal com doze rugas
bem desenvolvidas, dentes ocos, sendo um
dorsal e um ou dois subventrais. Raramente
possui um único dente dorsal de difícil
visualização. Faringe fortemente muscular
circundando a cavidade bucal, bulbo basal
presente ou ausente. Sistema secretor-
excretor visível. Posição do poro secretor-
excretor variável. Fêmeas didélficas com
ovários opostos e refletidos. Ovário an-
terior localizado à direita do intestino e o pos-
terior à esquerda. Machos com um único
testículo estendido à direita do intestino.
Espículas curvadas. Gubernáculo sem
apófise. Suplementos pré-cloacais
presentes ou ausentes. Três glândulas
caudais visíveis. Cauda cônica.
750, 1050, 1350, 1650 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Filipjev, 1922
Platt e Warwick (1988) citam que podem
existir confusões entre Chromadorita e
Innocuonema, devido às similaridades
morfológicas. Ambos possuem espécies sem
suplementos pré-cloacais, pontuações
da cutícula levemente alongada, e ponta da
cauda dorsalmente curvada.
Comentários
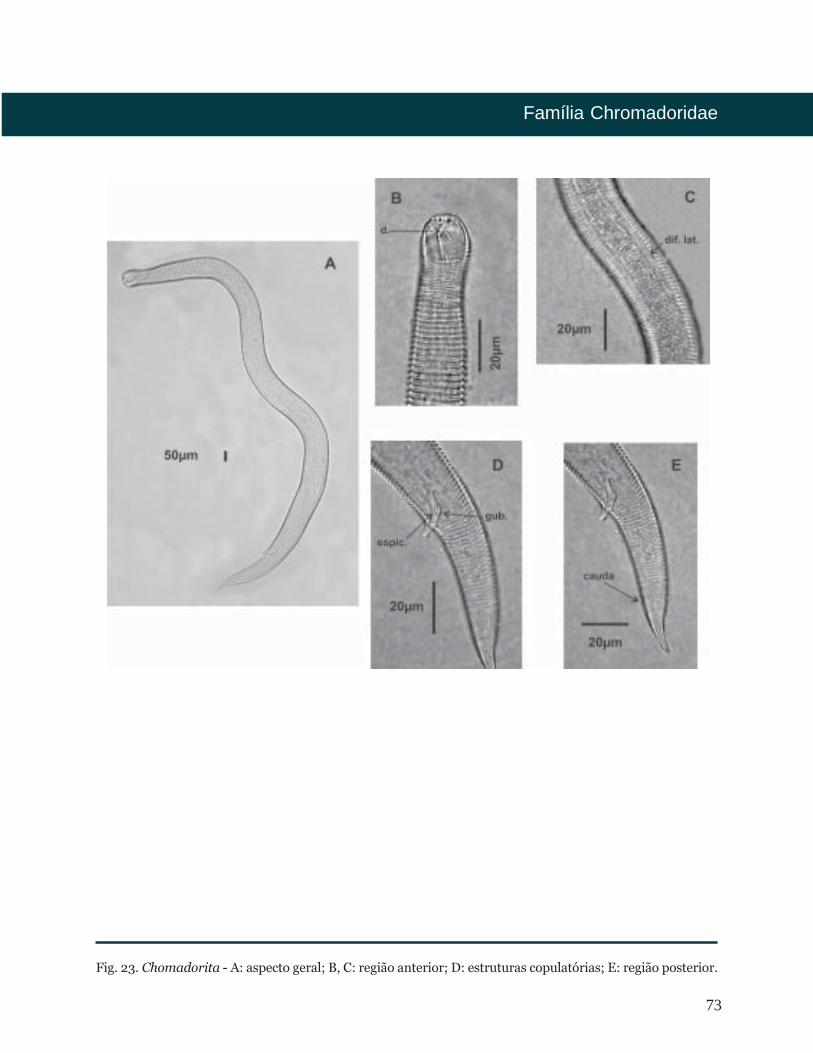
73
Fig. 23. Chomadorita - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Chromadoridae

74
Dichromadora
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Muthumbi, A. W.; Vincx, M. (1998) Chromadoridae
(Chromadorida: Nematoda) from the Indian Ocean:
Description of new and known species.
Hydrobiologia 364: 119–153.
Cutícula homogênea, com pontuações
fortes e diferenciação lateral com duas
fileiras longitudinais de pontos mais lar-
gos. Arranjo cefálico de seis setas cefálicas
pequenas e quatro setas longas em dois
círculos diferentes. Setas somáticas
presentes. Cavidade bucal com 3 dentes
ocos, sendo um dente dorsal, dois
subventrais menores e vários dentículos.
Anfídio oval, horizontal, na base das setas
cefálicas. Faringe com bulbo terminal
evidente. Sistema secretor-excretor visível.
Posição do poro secretor-excretor variável
de acordo com a espécie. Fêmeas
didélficas com ovários opostos e
refletidos. Machos com um único
testículo estendido anterior. Espículas
geralmente curvadas. Gubernáculo com
formas variadas. Suplementos pré-
cloacais em forma de copo presentes ou
ausentes. Espinerete visível. Cauda
cônica com a porção terminal estreita.
750, 1050 e 1350 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Kreis, 1929
Este gênero é morfologicamente muito
próximo de Chromadorita, sendo um
gênero principalmente marinho, mas
possui também representantes estuarinos
e de água doce.
Comentários
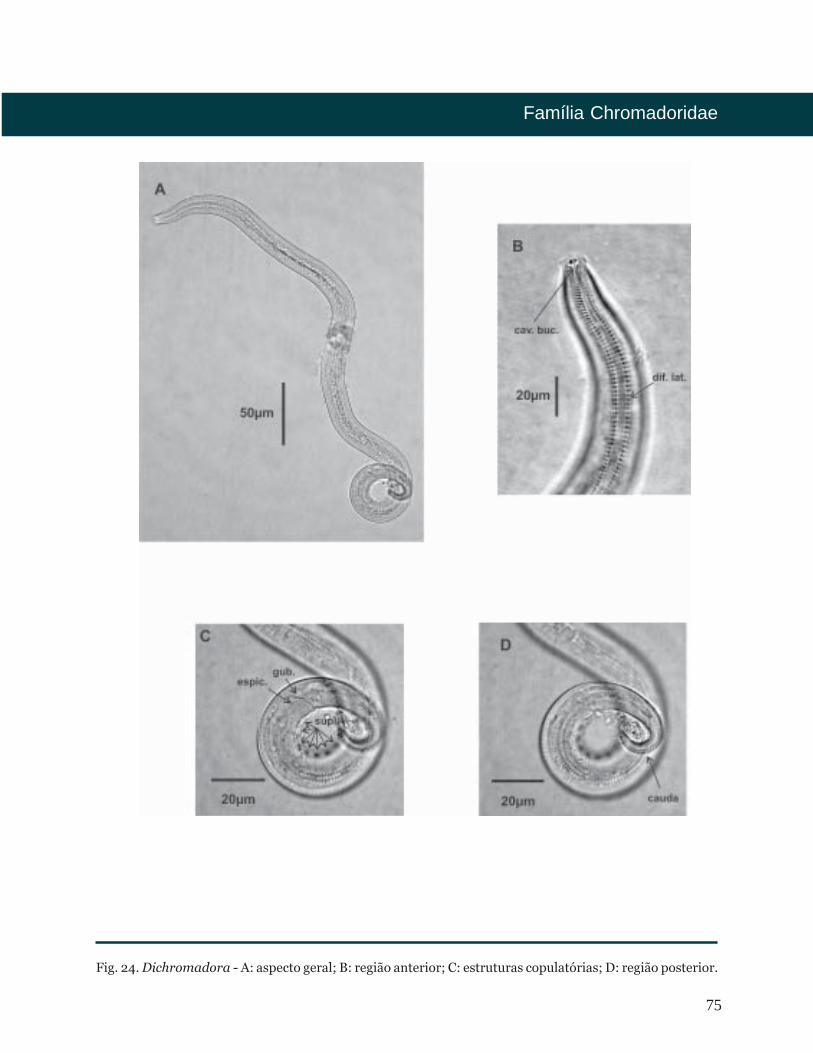
75
Fig. 24. Dichromadora - A: aspecto geral; B: região anterior; C: estruturas copulatórias; D: região posterior.
Família Chromadoridae

76
Endeolophos
Wieser, W. (1954) Free-living marine nematodes.
II. Chromadoroidea. (Reports ... 17.) Acta. Univ.
lund. N. F., Avd. 2, 50 (16): 148 p.
Blome, D. (1985) Interstitiella fauna von Galapagos,
XXXV. Chromadoridae (Nematoda). Microfauna
Marina, 2, 271–329.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
167p.
Cutícula heterogênea com região anterior
formada por anéis transversais de pontos.
Diferenciação lateral presente, porém sem
pontuações e ornamentações. Arranjo
cefálico de seis setas labiais internas, seis
labiais externas e quatro cefálicas. Setas
somáticas presentes ou ausentes. Anfídio
estreito e transversal. Cavidade bucal com
um dente dorsal de tamanho mediano e um
(ou dois) dente subventral muito pequeno.
Faringe com um bulbo posterior
fracamente desenvolvido. Fêmeas
didélficas com ovários opostos. Ma-
chos com um testículo estendido anterior.
Espículas pequenas. Gubernáculo com
ou sem apófise. Suplementos pré-
cloacais ausentes. Espinerete visível.
Cauda cônica.
750, 1050, 1650 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Boucher, 1976
A descrição original cita a ausência de
suplementos pré-cloacais, no entanto
em E. fossiferus a presença ou ausência é
duvidosa.
Comentários

77
Fig. 25. Endeolophos - A: aspecto geral; B, C, D: região anterior; E: estruturas copulatórias; F:região posterior.
Família Chromadoridae

78
Euchromadora
Cobb, N. A. (1914) Antarctic marine free-living
nematodes of the Shackleton expedition. Contr. Sci.
Nematol. (1): 1–33.
Shuurmans Stekhoven, J. H. (jr.) (1950) The
freeliving marine nemas of the Mediterranean. I.
The Bay of Villefranche. Mém. Inst. r. Sci. nat. Belg.
2 Sér. 37: 1–220.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Cutícula complexa e com pontuações
hexagonais ou ovóides que aparecem ao
longo de todo o comprimento com uma
discreta diferenciação no meio do corpo.
Arranjo cefálico com seis setas labiais e
quatro cefálicas em dois círculos
diferentes. Setas somáticas presentes ou
ausentes. Anfídio de difícil visualização
e de formato alongado transversalmente
sem projeção da fóvea anfideal.
Cavidade bucal com um sólido dente dor-
sal e vários dentículos. Faringe com
porção posterior levemente alargada,
porém sem bulbo basal definido. Fêmeas
didélficas com ovários opostos e
refletidos. Machos com um único
testículo estendido anterior. Espículas
delgadas e curvadas. Gubernáculo com
estruturas laterais telamons.
Suplementos pré-cloacais ausentes.
Espinerete curto. Cauda cônica e
relativamente curta.
750, 1350 e 1650 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
de Man, 1886
As características utilizadas para
diferenciar as espécies são o formato da
espícula e do gubernáculo.
Comentários
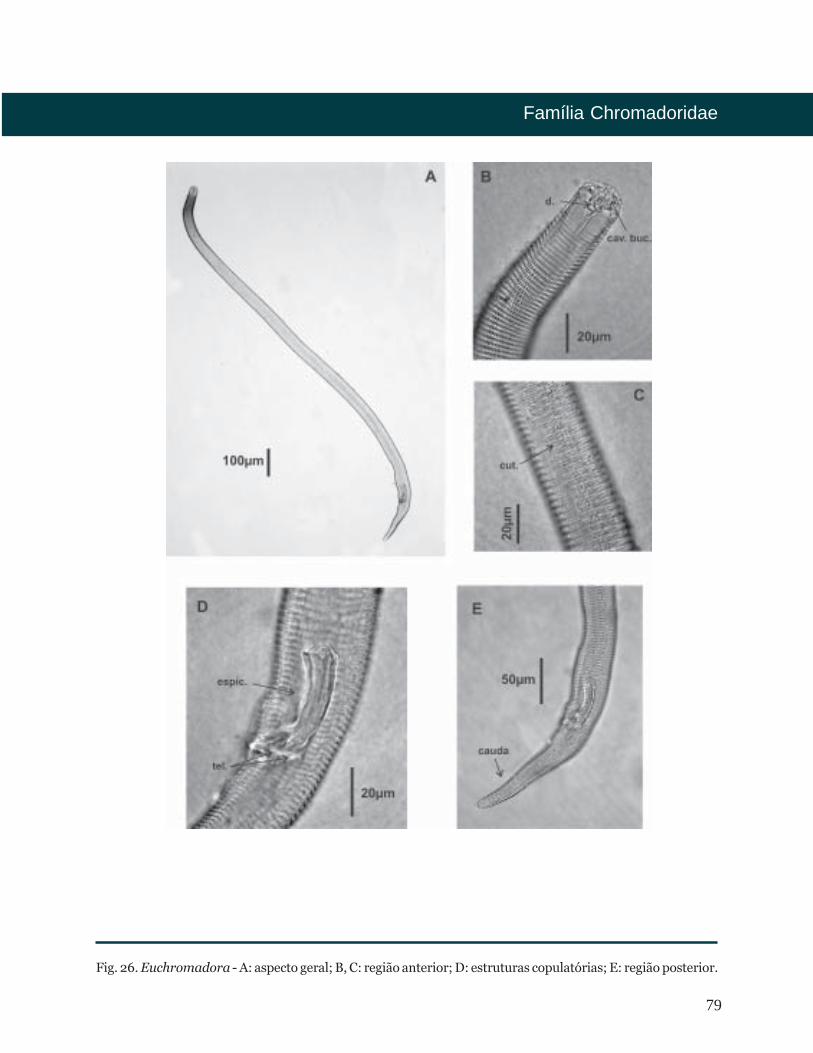
79
Fig. 26. Euchromadora - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Chromadoridae

80
Neochromadora
Cobb N. A. (1930) Marine free-living nemas. Sci-
ent. Rep. Australas. Antarct. Exped. (1911–14), Ser:
C, Zool. & Bot. 6 (7): 28 p.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Decraemer, W., & Smol, N. (2006) Orders
Chromadorida, Desmodorida and Desmoscolecida.
In: Eyualem-Abebe, Traunspurger, W. & Andrássy,
I. (eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 497-573). CABI Publishing, UK.
Cutícula heterogênea com ornamentação
complexa. Diferenciação lateral de duas ou
três fileiras longitudinais de pontuações
maiores que as observadas no restante do
corpo. Arranjo cefálico formado por seis setas
labiais internas e quatro cefálicas em dois
círculos diferentes. Setas somáticas
presentes. Cavidade bucal com um dente
dorsal oco e dois dentes ventrosublaterais.
Faringe alargada na porção anterior,
especialmente na região de inserção do dente
dorsal e posteriormente com um bulbo ter-
minal bem desenvolvido. Cárdia alongada.
Sistema secretor-excretor frequentemente
visível. Posição do poro secretor-excretor
variável de acordo com a espécie. Fêmeas
didélficas com ovários opostos,
estendidos ou refletidos. Machos com
um único testículo anterior estendido.
Numerosos suplementos pré-cloacais
em forma de copo. Espículas curvadas,
fortemente esclerotizada. Gubernáculo
curto e sem apófise. Três glândulas caudais
visíveis. Espinerete longo. Cauda cônica.
750, 1050, 1350, 1650 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Micoletzky, 1924
Neochromadora é um gênero
essencialmente marinho, com apenas uma
espécie de água doce e ocasionalmente em
águas salobras.
Comentários

81
Fig. 27. Neochromadora - A: aspecto geral; B, C,D: região anterior; E, F: estruturas copulatórias; G: região posterior.
Família Chromadoridae

82
Comesa
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Insti-
tuted. 167p.
Warwick, R. M.; Platt, H. M., Sommerfield, P.
(1998) Free-living Marine Nematodes Part III –
Monhysterids. Synopses of the British Fauna (New
Series) Eds: Doris M. Kermack e R.S.K. Barnes. 53,
296p.
Muthumbi, A. W.; Vanreusel, A. (2006) Order
Araeolaimida. In: Eyualem-Abebe, Traunspurger,
W. & Andrássy, I. (eds) Freshwater Nematodes:
Ecology and Taxonomy (pp. 604–610). CABI Pub-
lishing, UK.
Corpo longo e robusto. Cutícula com
pontuações e poros cuticulares laterais.
Diferenciação lateral presente ou ausente.
Extremidade anterior redonda. Seis setas
labiais externas curtas e quatro cefálicas
presentes. Anfídio espiral. Cavidade
bucal bem definida com um distinto dente
dorsal e projeções ventrais. Faringe com
bulbo muscular posterior. Fêmeas
didélficas com ovários de mesmo
tamanho, opostos e refletidos. Vulva
situada na região mediana do corpo. Ma-
chos com dois testículos, dispostos em
lados opostos do intestino. O testículo
posterior é menor que o anterior.
Espículas e gubernáculo curtos.
Apresentam de 8 a 14 suplementos pré-
cloacais em forma de copo. Cauda
cônica, com ponta arredondada.
750 e 1050 metros.
Gerlach, 1956
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
Animais que medem de 0,6 a 1,5 mm.
Todas as espécies contidas neste gênero
são marinhas.
Comentários

83
Fig. 28 - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Neotonchidae

84
Nannolaimus
Gerlach, S. A. (1957) Die Nematodenfauna des
Sandstrandes an der Küste Von Mittelbrasilien
(Brasilianische Meeres-Nematoden IV) Mitt. Zool.
Mus. Berl. 33 (2): 411–459.
Platt, H. M. (1982) Revision of the Ethmolaimidae
(Nematoda: Chromadorida). Bulletin of the British
Museum (Natural History). Zoology series, vol. 43,
No 4: 185–252.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Cutícula com pontuações fortes. Arranjo
cefálico de seis setas labiais internas, seis
labiais externas e quatro cefálicas
relativamente longas em três círculos
separados. Setas somáticas presentes.
Anfídio multiespiral. Cavidade bucal
pobremente desenvolvida com um dente
dorsal diminuto. Faringe com bulbo basal.
Sistema excretor-secretor de difícil
visualização. Poro excretor-secretor
também de difícil visualização.
Suplementos pré-cloacais em forma
de copo. Espículas curtas, curvadas e com
a região proximal cefalada. Gubernáculo
sem apófise, curto e paralelo às
espículas. Cauda cônica.
1950 metros.
Cobb, 1920
Bibliografia
Distribuição Batimétrica
Diagnose Distribuição Geográfica
Este gênero já pertenceu a Família
Cyatholaimidae. A separação ocorreu
devido ao arranjo cefálico de Nannolaimus
que é distribuído em três círculos
diferentes, ao contrario do que ocorre em
Cyatholaimidae que apresenta os dois
últimos círculos fusionados.
Comentários

85
Fig. 29. Nannolaimus - A: aspecto geral; B, C, D: região anterior; E: estruturas copulatórias; F:região posterior.
Família Neotonchidae

86
Longicyatholaimus
Vitiello, P. (1970) Nématodes libres marins des
vases profondes du Golfu du Lion II. Chromadorida.
Téthys 2: 449–500.
Hopper, B. E. (1972) Free-living marine nematodes
from Biscayne Bay, Florida. IV. Cyatholaimidae: on
the occurrence of Marylynnia n.gen. and
Longicyatholaimus Micoletzky, 1924 in Biscayne
Bay, with a description of L. longicaudatus (de Man,
1876) from the type locality. Zool. Anz. 189 (1/2)
64–88.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
167p.
Cutícula com fileiras de pontos transversais
e com diferenciação lateral. Pontuações
maiores na região anterior. Poros
epidermais presentes. Arranjo cefálico de
seis papilas labiais internas e dez cefálicas,
sendo as labiais internas maiores que as
cefálicas. Setas somáticas presentes.
Anfídio multiespiral. Cavidade bucal
pequena com um dente dorsal e
possivelmente com dois dentículos
subventrais. Faringe cilíndrica, sem bulbo
basal. Cárdia pequena. Sistema secretor-
excretor visível. Posição do poro secretor-
excretor variável de acordo com a espécie.
Fêmeas didélficas com ovários opostos
e refletidos. Machos com dois testículos
opostos e estendidos. Suplementos pré-
cloacais presentes. Espículas curvadas
com a região distal encoberta pela porção
terminal do gubernáculo. Gubernáculo
pequeno e sem apófise, com estruturas
adicionais similares a dentes na porção dis-
tal. Cauda filiforme.
750, 1050, 1350 e 1950 metros.
Micoletzky, 1924
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
As estruturas acessórias e denteadas
presentes na porção distal do
gubernáculo também estão presentes em
outros gêneros da família. A presença dos
dois dentes subventrais na cavidade bucal
é discutida por alguns autores.
Comentários
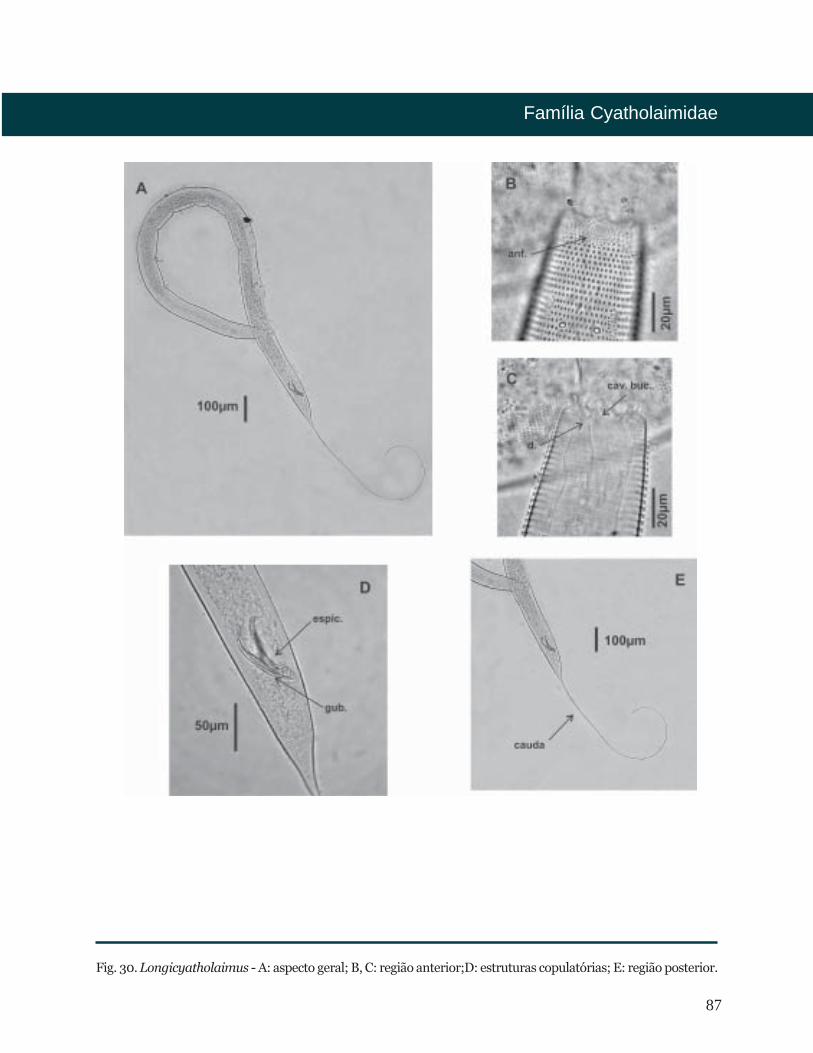
87
Fig. 30. Longicyatholaimus - A: aspecto geral; B, C: região anterior;D: estruturas copulatórias; E: região posterior.
Família Cyatholaimidae

88
Marylynnia
Warwick, R. M. (1971) The Cyatholaimidae (Nema-
toda, Chromadoroidea) off the coast of
Northumberland. Cah. Biol. mar. 12 (1): 95–100.
Hopper, B. E. (1972) Free-living marine nematodes
from Biscayne Bay, Florida. IV. Cyatholaimidae: on
the occurrence of Marylynnia n.gen. and
Longicyatholaimus Micoletzky, 1924 in Biscayne
Bay, with a description of L. longicaudatus (de Man,
1876) from the type locality. Zool. Anz. 189 (1/2)
64–88.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
167p.
Cutícula com fileiras de pontos transversais
ligados a poros. Diferenciação lateral de
pontos maiores e mais espaçados entre si.
Os poros na cutícula podem ser de dois tipos:
tipo 1- simples e arredondados ou ovais
longitudinalmente, os últimos entre dois
pontos e referem-se a pontuações laterais
modificadas (LMPs) e tipo 2- estendendo-
se até a parte cônica da cauda. Arranjo
cefálico de seis setas labiais internas e dez
cefálicas. Setas somáticas presentes.
Anfídio multiespiral. Cavidade bucal com
um proeminente dente dorsal e dois pares
de dentes subventrais. Faringe longa e sem
bulbo basal. Sistema secretor-excretor
visível. Posição do poro secretor-excretor
variável. Fêmeas didélficas com ovários
opostos e refletidos. Machos com dois
testículos opostos ou um único anterior.
Suplementos pré-cloacais em forma de
copo, sem cuticularização. Espículas
curvadas com a porção proximal mais
cuticularizada que o restante. Velum bem
desenvolvido. Gubernáculo sem apófise,
alargado suavemente na porção distal com
vários dentes. Cauda cônico-cilíndrica.
750, 1050, 1350 e 1950 metros.
Hopper, 1977
Distribuição GeográficaDiagnose
Distribuição Batimétrica
Bibliografia
Geralmente os machos apresentam
espículas de comprimento e forma
semelhantes, no entanto M. complexa possui
as espículas de comprimento diferentes.
Comentários
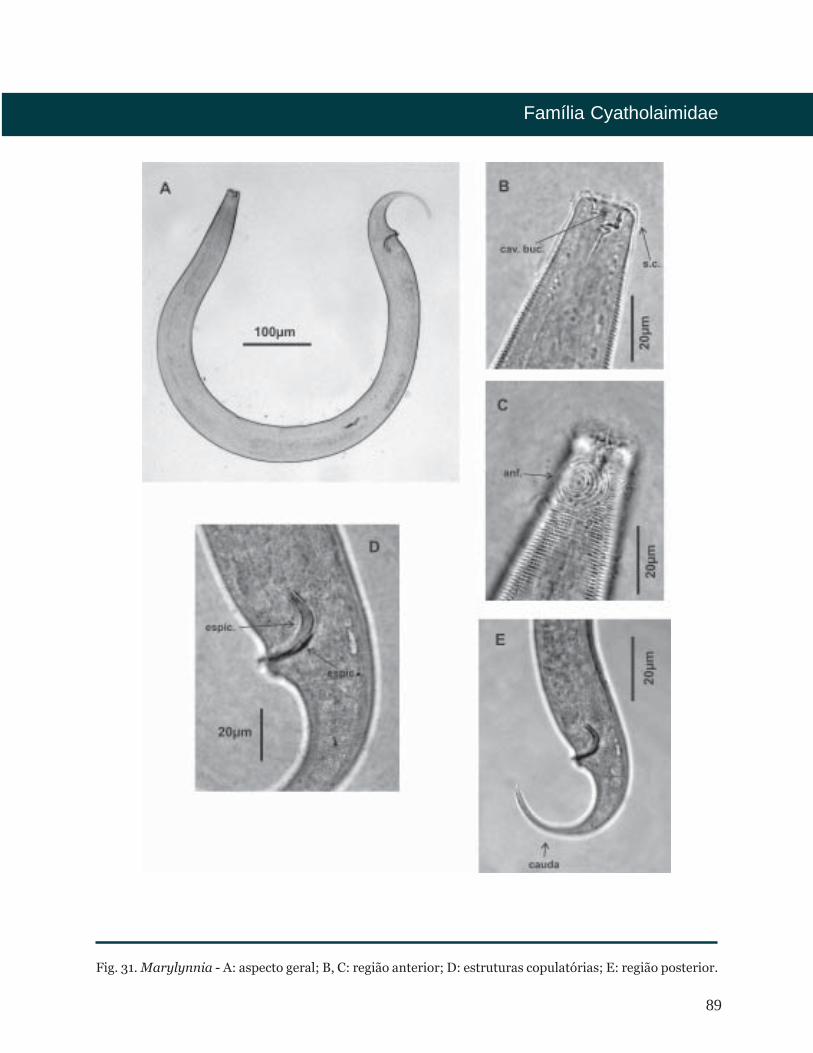
89
Fig. 31. Marylynnia - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Cyatholaimidae

90
Minolaimus
Vitiello, P., (1970) Nematodes libres Marins des
vases profondes du golgo du Lion. II.
Chromadorida. Tethys 2(2): 449–500.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
167p.
Cutícula levemente pontuada com
diferenciação lateral que consiste de
pontuações maiores. Região anterior
atenuada. Arranjo cefálico formado por
seis setas labiais externas e quatro
cefálicas curtas em círculos separados.
Anfídio multiespiral posterior as setas
cefálicas. Cavidade bucal pequena e sem
dentes. Cárdia pequena. Faringe
cilíndrica, levemente alargada na região
posterior, porém sem bulbo basal
definido. Poro secretor-excretor visível
com posição variável de acordo com a
espécie. Fêmeas com dois ovários
opostos antidrosmous. Machos com
dois testículos. Região pré-anal com
numerosos suplementos em forma de
papilas proeminentes. Espículas
curvadas e fortemente esclerotizadas.
Gubernáculo pequeno e sem apófise
dorsal. Três glândulas caudais.
Espinerete nem sempre visível. Cauda
filiforme.
750, 1050, 1350 e 1950 metros.
Distribuição Geográfica
Vitiello, 1970
Distribuição Batimétrica
Bibliografia
Diagnose
Uma das principais características de
Minolaimus são a posição e o número de
voltas do anfídio.
Comentários

91
Fig. 32. Minolaimus - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Cyatholaimidae

92
Paracyatholaimus
Kreis, H. A. (1963) Marine Nematoda. Copenhagen
& Reykjavik, Ejnar Munskgaard – The Zoology of
Iceland 2 (14): 1–68.
Lorenzen, S. (1972) Die Nematodenfauna im
verklappungsgebiet für industrieabwasser
nordwestlich von Helgoland II. Desmodorida und
Chromadorida. Zool. Anz. 187 (5/6): 283–302.
Pastor de Ward, C. T. (1985) Free-living marine
nematodes of the Deseado River Estuary
(Chromadoroidea: Chromadorida, Ethmolaimidae,
Cyatholaimidae and Choniolaimidae) Santa Cruz,
Argentina, 5. Centro Nacional Patagonica. Publ.
esp., 6, 1–83.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Cutícula com pontuações, de difícil
visualização e sem diferenciação lateral.
Arranjo cefálico de seis setas labiais
internas no primeiro círculo e dez setas
em um segundo círculo. Anfídio
multiespiral. Cavidade bucal com um
grande dente dorsal e dois menores
subventrais no lado oposto da boca. Vulva
posicionada no meio do corpo. Fêmeas
didélficas com ovários opostos,
antidrosmous e refletidos, o anterior
à direita e o posterior à esquerda do
intestino. Machos com um único testículo
anterior, estendido e à esquerda do
intestino. Suplementos pré-cloacais
setosos. Espículas curvadas.
Gubernáculo pequeno e simples,
fortemente alargado na porção terminal
e arredondado, apófise presente ou
ausente. Três glândulas caudais.
Espinerete visível. Cauda cônica.
1050 e 1650 metros.
Distribuição Geográfica
Micoletzky, 1924
Distribuição Batimétrica
Bibliografia
Diagnose
Na literatura, é relatado o fenômeno de
viviparidade para este gênero, como na
espécie P. chilensis.
Comentários

93
Fig. 33. Paracyatholaimus - A: aspecto geral; B: região anterior; C: região posterior; D: estruturas copulatórias.
Família Cyatholaimidae

94
Cheironchus
Allgén, C. A. (1959) Freeliving marine nematodes.Further zool. Results Swed. Antarct. Exped. 1901-1903, 5 (3): 293 p.
Vitiello, P. (1970) Nematodes libres marins desvases profondes du Golfe du Lion. II.Chromadorida. Tethys 2(2): 449–500.
Platt, H. M.; Warwick, R. M. (1988) Free-livingmarine Nematodes – Part II Chromadorids. Syn-opses of the British Fauna (New Series). Ed. Dorism. Kermarck and R. S. K. Barnes. 38, 502p.
Cutícula anelada com fileiras de pontos e
diferenciação lateral presente. Arranjo
cefálico de 6+6+4 em três círculos
separados, sendo o último círculo com
setas de maior comprimento. Setas
somáticas usualmente presentes. Cavidade
bucal com mandíbulas reduzidas. Dentes
presentes ou ausentes. Anfídio
multiespiral. Faringe fortemente muscu-
lar, com bulbos nas duas extremidades
bem desenvolvidos. Fêmeas didélficas,
com ovários opostos e refletidos.
Vulva geralmente, um pouco acima da
região mediana do corpo. Machos,
geralmente com dois testículos. Espículas
curtas e curvadas. Gubernáculo delgado,
curto e sem apófise. Suplementos pré-
cloacais em forma de copo. Cauda cônica,
curta e com espinerete visível.
1050 metros.
Distribuição Geográfica
Cobb, 1917
Distribuição Batimétrica
Bibliografia
Diagnose
Fêmeas grávidas pertencentes a este
gênero conseguem carregar até dez ovos no
útero que podem medir até 8,5 µm cada.
Comentários
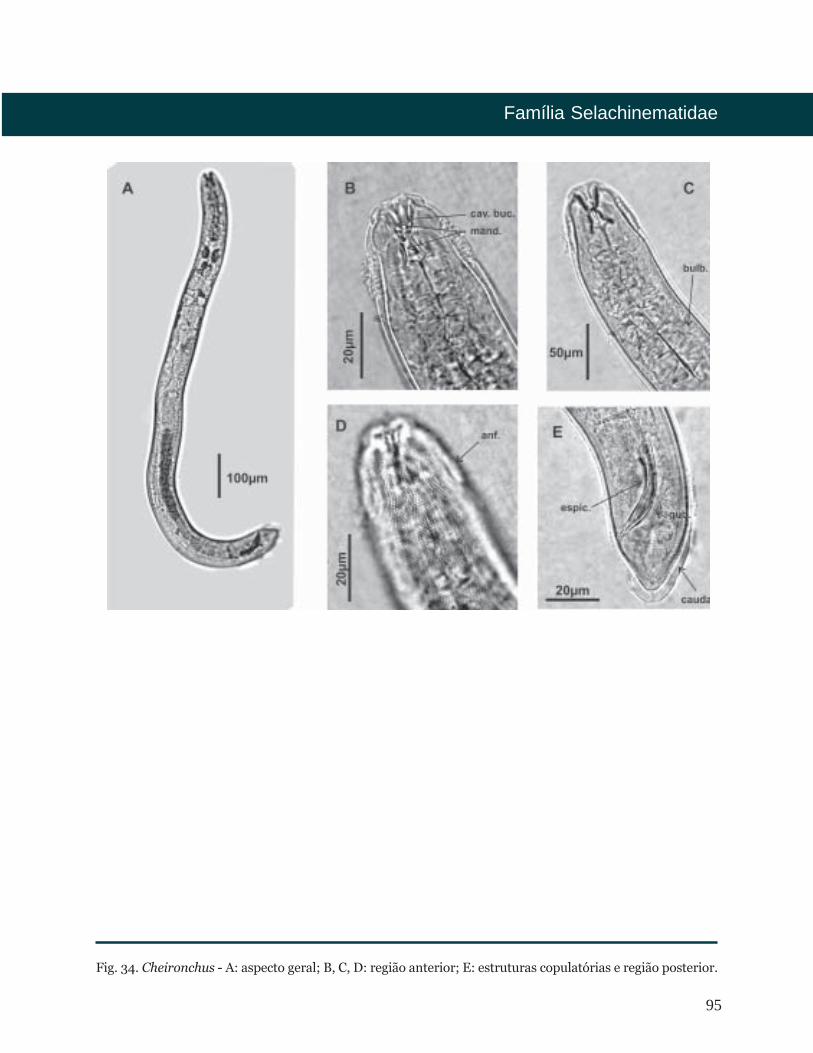
95
Fig. 34. Cheironchus - A: aspecto geral; B, C, D: região anterior; E: estruturas copulatórias e região posterior.
Família Selachinematidae

96
Choanolaimus
Gerlach, S. A. (1952) Nematoden aus dem
Küstengrundwasser. Abh. math.-naturw. Kl. Akad.
Wiss. Mainz. Nr. 6: 315–372.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Cutícula formada por anelações com fileiras
transversais de pontos. Diferenciação lat-
eral com pontuações maiores e mais
distantes entre si. Poros glandulares
presentes. Arranjo cefálico 6+6+4
papiliforme. Cavidade dividida em duas
partes: uma porção anterior alargada, com
rugas fortemente desenvolvidas; e uma
região posterior mais estreita, tubular e com
colunas longitudinais. Entre as duas partes
encontram-se fileiras de dentículos de
diversos tamanhos. Anfídio multiespiral.
Faringe levemente alargada na região an-
terior, sem bulbo basal. Fêmeas didélficas
com ovários refletidos. Machos com um
testículo estendido anterior. Espículas de
comprimento e forma variável de acordo
com a espécie. Gubernáculo curto,
simples e delgado, geralmente paralelo às
espículas. Suplementos pré-cloacais
em forma de papilas ligadas a delicados
poros. Três glândulas caudais. Cauda
extremamente curta com extremidade
arredondada e com espinerete visível.
1050 e 1350 metros.
Distribuição Geográfica
de Man, 1880
Distribuição Batimétrica
Bibliografia
Diagnose
As espécies deste gênero são predadores,
inclusive de outros Nematoda. Em algumas
das espécies, os autores descrevem a
ausência das glândulas caudais.
Comentários
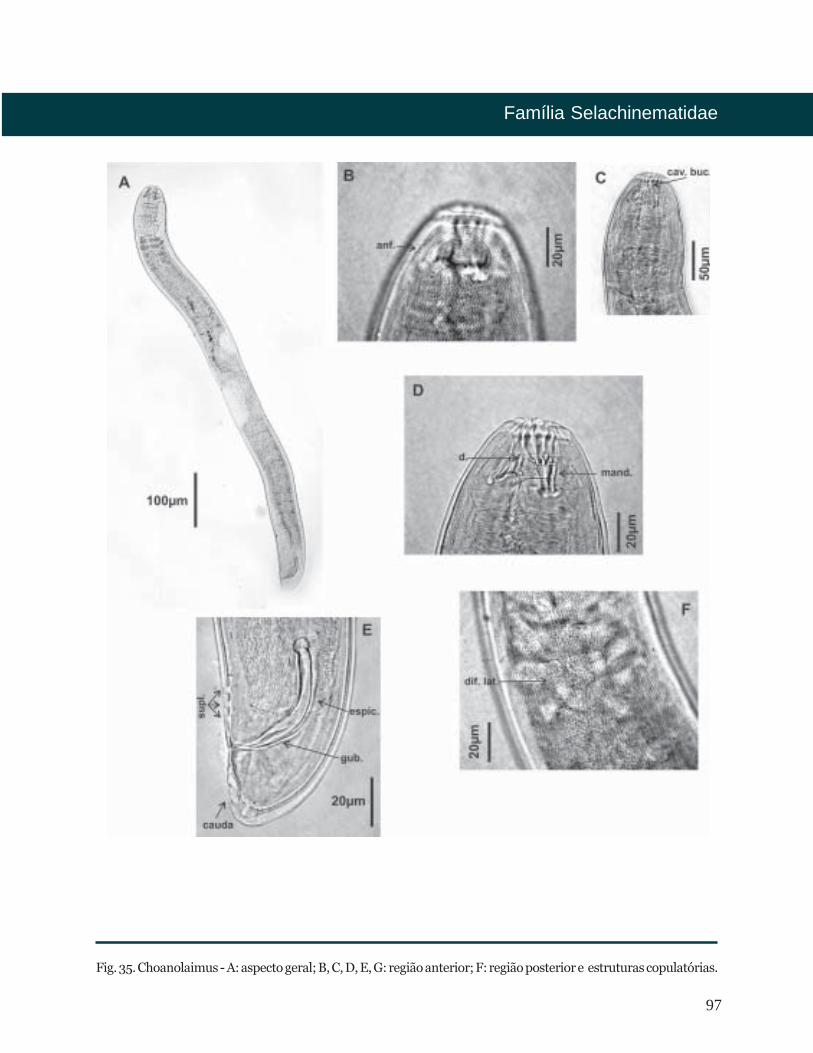
97
Fig. 35. Choanolaimus - A: aspecto geral; B, C, D, E, G: região anterior; F: região posterior e estruturas copulatórias.
Família Selachinematidae

98
Gammanema
Murphy, D. G. (1964) Free-living marine nema-
todes, I. Southerniella youngi, Dagda phinneyi and
Gammanema smithi, new species. Proc. helminth.
Soc. Wash. 31 (2): 190–198.
Bibliografia: Murphy, D. G. (1964) Free-living
marine nematodes, I. Southerniella youngi, Dagda
phinneyi and Gammanema smithi, new species.
Proc. helminth. Soc. Wash. 31 (2): 190–198.
Murphy, D. G. (1965) Chilean marine nematodes.
Veröff. Inst. Meeresforsch. Bremerh. 9: 173–203.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Cutícula formada por fileiras de pontos, sem
diferenciação lateral. Seis papilas labiais
cônicas, seis setas labiais externas curtas e
quatro setas cefálicas que podem ou não estar
no mesmo círculo das labiais externas. Setas
somáticas presentes ou ausentes. Anfídio
espiral. Cavidade bucal dividida em duas
partes: uma porção anterior alargada com
uma linha cuticular, finas estriações
longitudinais e 12 rugas fortemente
desenvolvidas; e uma região posterior, mais
estreita, tubular e com colunas longitudinais.
Cárdia bem definida. Faringe geralmente
curta com um suave alargamento na porção
anterior e sem bulbo basal na região poste-
rior. Fêmeas didélficas com ovários
opostos e estendidos. Machos com dois
testículos estendidos anteriormente em
posição variável de acordo com a espécie.
Suplementos pequenos e em forma de
copo. Espículas delgadas e levemente
curvadas. Gubernáculo geralmente
pequeno e sem apófise. Três glândulas
caudais visíveis. Cauda relativamente curta
de forma variável: cônica, redonda e
cilíndrica. Espinerete curto e estreito.
750, 1350 e 1650 metros.
Distribuição Geográfica
Cobb, 1920
Distribuição Batimétrica
Bibliografia
Diagnose
Apesar da maioria dos machos apresentarem
dois testículos estendidos anteriormente, a
espécie G. smithi possui dois testículos opostos.
Comentários
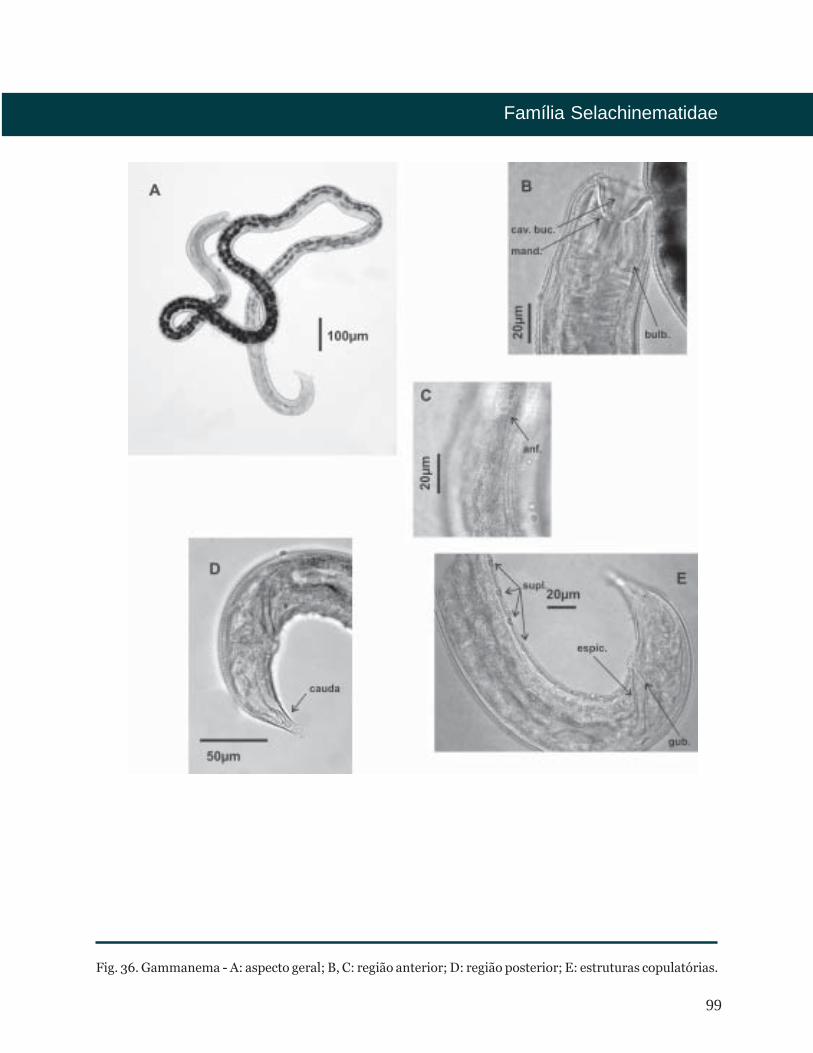
99
Fig. 36. Gammanema - A: aspecto geral; B, C: região anterior; D: região posterior; E: estruturas copulatórias.
Família Selachinematidae

100
Halichoanolaimus
Gerlach, S. A. (1957) Marine nematoden aus dem
Mangrove-Gebiet Von Cananéia. Abh. math.-
naturw. Kl. Akad. Wiss. Mainz. 5: 129–176.
Timm, R. W. (1961) The marine nematodes of the
Bay of Bengal. Proc. Pakist. Acad. Sci., Vol. I, No. 1:
25–88.
Inglis, W. G. (1968) Interstitial nematodes from St.
Vincent’s Bay, New Caledonia. Expéd. française. sur
les recifs coralliens de la Nouvelle Calédonie, Paris,
1967. (Paris, Editions de la Fondation Singer-
Polignac). 2: 29–74.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
Cutícula com fileiras de pontos transversais.
Pontuações maiores na região anterior e
espaçadas na cauda. Diferenciação lateral
presente ou ausente. Arranjo cefálico em três
círculos separados. Setas somáticas presentes.
Anfídio multiespiral. Cavidade bucal
esclerotizada, com duas porções: uma porção
anterior alargada com linha cuticular, finas
estriações longitudinais e 12 rugas fortemente
desenvolvidas; e uma posterior, estreita, tubu-
lar com colunas longitudinais. Entre as duas
porções encontram-se uma fileira de
dentículos em forma de blocos. Faringe curta,
levemente alargada na porção posterior, sem
bulbo basal. Sistema secretor-excretor visível.
Posição do poro secretor-excretor variável de
acordo com a espécie. Fêmeas didélficas com
ovários opostos e refletidos. Vulva, um
pouco acima da região mediana do corpo. Ma-
chos, com dois testículos. Suplementos pré-
cloacais setosos. Espículas curvadas. Ala
ventral das espículas presente ou ausente.
Gubernáculo delgado, longo ou curto e sem
apófise. Três glândulas caudais.
Espinerente visível. Cauda alongada com
uma porção anterior cônica e a região poste-
rior cilíndrica.
750, 1050, 1350, 1650 e 1950 metros.
Distribuição Geográfica
de Man, 1886
Distribuição Batimétrica
Bibliografia
Diagnose
O tipo e a distribuição das mandíbulas e dos
dentes diferenciam Halichoanoalaimus dos
outros membros da família.
Comentários
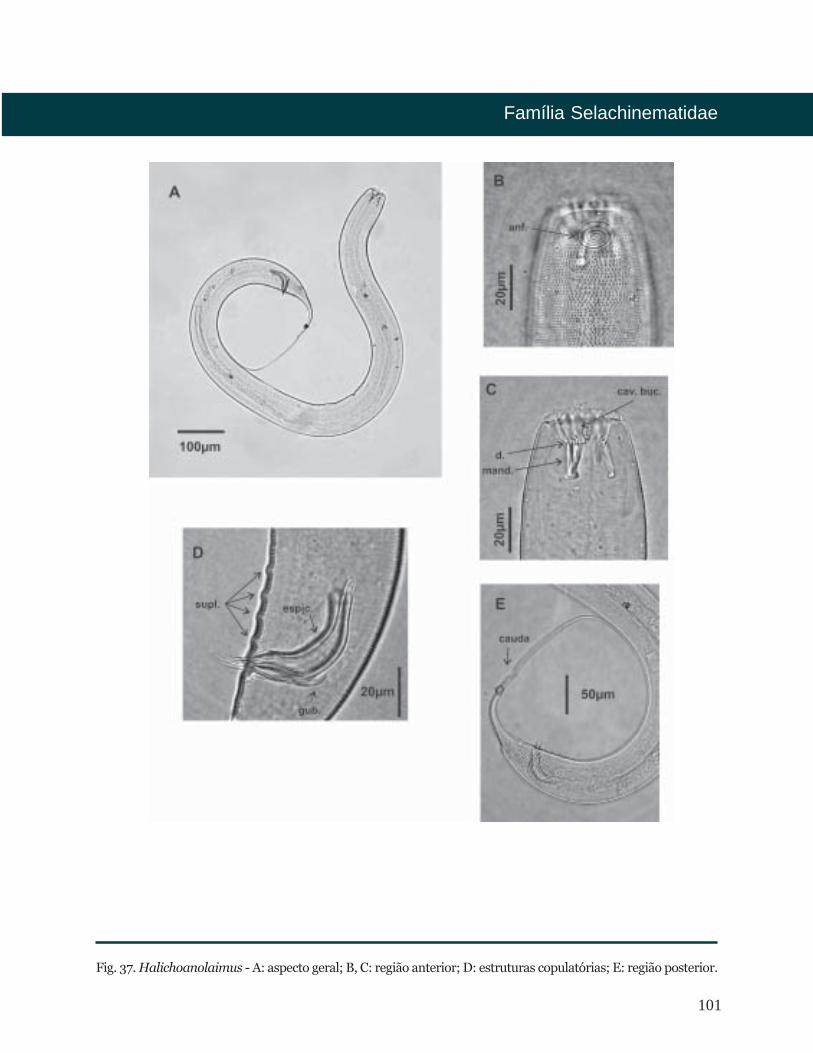
101
Fig. 37. Halichoanolaimus - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Selachinematidae

102
Latronema
Wieser, W. (1954) Free-living marine nematodes.
II. Chromadoidea. (Reports...17.) Acat. Univ. lund.
, N. F., Avd. 2, 50 (16): 148 p.
Gerlach, S. A (1953) Die Nematodenbesiedlung des
Sandstrandes und des Küstengrundwassers an der
italienische n Küste. I. Systematischer Teil. Archo.
Zool. Ital. 37: 517–640.
Gerlach, S. A. (1964) Revision der Choniolaiminae
und Selachinematinae (freilebende Meeres-
Nematoden). Mitt. hamburg. zool. Mus. Inst.
(Kosswig-Festschrift): 23–50.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Cutícula com 12-50 fileiras de pontos
longitudinais. Arranjo cefálico formado
por dez setas cefálicas separadas
simetricamente. Setas somáticas
presentes ou ausentes. Anfídio
multiespiral. Cavidade bucal com
mandíbulas e dividida em duas porções.
Porção anterior com doze apófises
arranjadas em três grupos. Cada apófise
é diferenciada em dois dentes que
marcam a borda entre a porção anterior e
posterior da cavidade bucal. Porção pos-
terior fracamente desenvolvida. Faringe
geralmente curta, fortemente muscular e
sem bulbo basal. Fêmeas didélficas com
ovários antidrosmous refletidos.
Machos com dois testículos opostos.
Suplementos pré-cloacais em forma
de copo. Espículas levemente curvadas
e abertas na porção proximal.
Gubernáculo sem apófise. Três
glândulas caudais visíveis. Espinerete
curto. Cauda curta e cônica.
1650 metros.
Distribuição Geográfica
Wieser, 1954
Distribuição Batimétrica
Bibliografia
Diagnose
A cutícula de Latronema é uma
característica comum ao gênero, exceto
pela espécie L. annulatun que apresenta a
cutícula com fileiras de 12 a 18 anelações.
Comentários
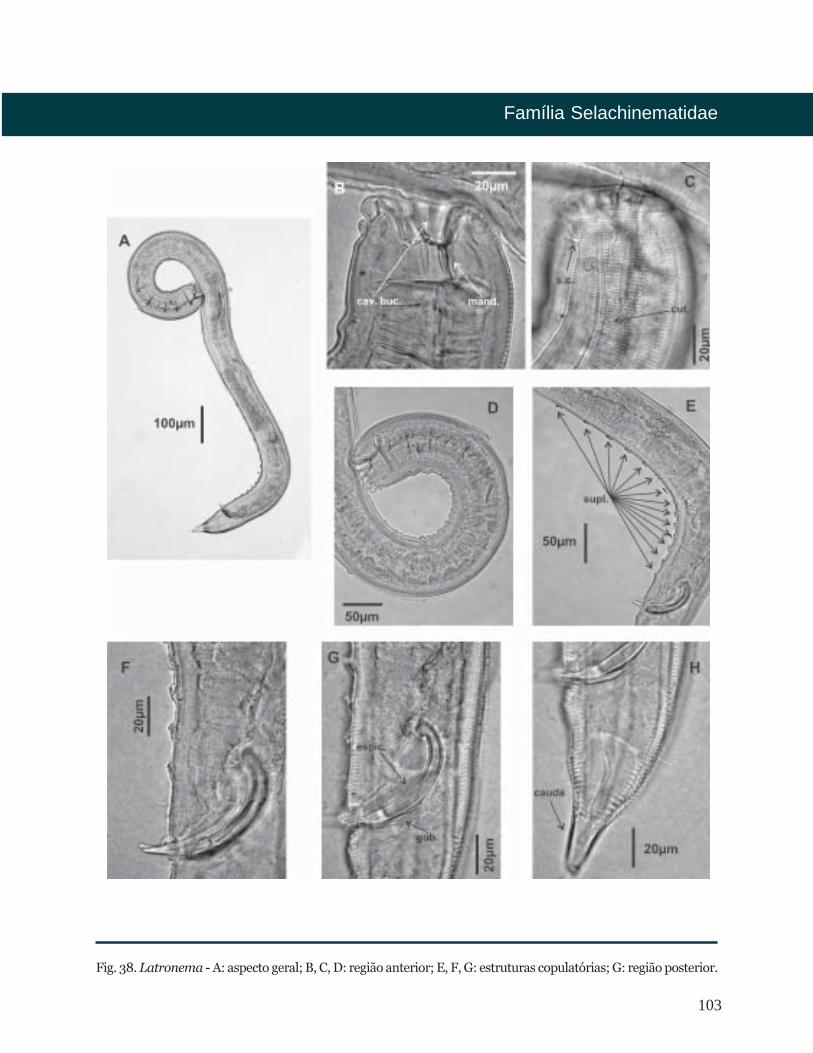
103
Fig. 38. Latronema - A: aspecto geral; B, C, D: região anterior; E, F, G: estruturas copulatórias; G: região posterior.
Família Selachinematidae

OR
DE
M D
ES
MO
DO
RID
A DE CONINCK
, 19
65
Indivíduos pertencentes à Ordem Desmodorida são caracterizados
por apresentar cutícula somente com estriações tranversais e nunca com
pontuações; setas cefálicas quase sempre em três círculos separados (6 +
6 + 4); cavidade bucal frequentemente com um dente dorsal distinto e
dentes menores subventrais; faringe geralmente com um bulbo termi-
nal; machos com um ou dois testículos; suplementos pré-cloacais
tubulares ou papilosos.
Os desmodorídeos são animais essencialmente marinhos, exceto
pelo gênero Prodesmodora Micoletzky, 1923, com 40% de espécies
encontradas em água doce.
A posição de Desmodorida como Ordem foi confirmada com a
classificação filogenética de De Ley & Blaxter (2002). Dentro de
Desmodorida estão inseridas as Famílias: Desmodoridae,
Epsilonematidae, Draconematidae, Microlaimidae, Aponchiidae,
Monoposthiidae (Figura 6).
CARACTERÍSTICAS GERAIS
Figura 39: Porção anterior do corpo de gêneros representantes das Famílias de
Desmodorida (Decraemer & Smoll, 2006) que ocorrem na Bacia de Campos: A=
Desmodoridae; B= Draconematidae; C= Microlaimidae; D= Aponchiidae; E=
Monoposthiidae.

106
Desmodora
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Jensen, P. (1991) Nine new and less known nema-
tode species from the deep-sea benthos of the Nor-
wegian Sea. Hydrobiologia, 222, 57–76.
Soetaert, K.; Heip, C. (1995) Nematodes assem-
blages of deep sea and shelf break sites in the North
Atlantic and Mediterranean Sea. Marine Ecology
Progress Series, 125, 171–183.
Verschelde, D.; Gourbault, N.; Vincx, M. (1998)
Revision of Desmodora with descriptions of new
desmodorids (Nematoda) from hydrothermal vents
of the Pacific. J. Mar. Biol. Ass. U.K., 78, 75–112.
Decraemer, W., & Smol, N. (2006) Orders
Chromadorida, Desmodorida and Desmoscolecida.
In: Eyualem-Abebe, Traunspurger, W. & Andrássy,
I. (eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 497–573). CABI Publishing, UK.
Cutícula com estriações transversais.
Cápsula cefálica presente, sem
estriações. Arranjo cefálico de seis papilas
ou setas labiais internas, seis papilas ou
setas labiais externas e quatro setas
cefálicas em três círculos separados, sendo
o último círculo sempre de setas maiores.
Algumas espécies apresentam setas sub-
cefálicas. Cavidade bucal, normalmente,
com um dente dorsal. Anfídio variando
de criptoespiral a multiespiral, raramente
em loop, circundado com duas linhas,
inserido na cápsula cefálica. Bulbo
faringeal presente. Fêmeas didélficas,
com ovários opostos refletidos. Ma-
chos com um testículo anterior presente.
Espículas de formato variável de acordo
com a espécie. Gubernáculo simples e
sem apófise dorsal. Algumas espécies
apresentam suplementos pré-cloacais,
algumas outras pré e pós-cloacais. Cauda
cônica e sem estriações na porção terminal.
750, 1050, 1350 e 1950 metros.
De Man, 1889
Diagnose
Bibliografia
Distribuição Batimétrica
Distribuição Geográfica
Animais que medem de 1 a 10 mm. Algumas
espécies possuem, além do dente dorsal, um
pequeno dente subventral e até fileiras de
dentículos, como: D. pilosa Ditlevsen, 1926.
Desmodorídeos são quase sempre
mencionados em estudos de mar profundo,
mesmo que sejam encontrados poucos
indivíduos.
Comentários

107
Fig. 40. Desmodora - A: aspecto geral; B: região anterior; C: estruturas copulatórias; D: região posterior.
Família Desmodoridae

108
Molgolaimus
Jensen, P. (1978) Revision of Microlaimidae, erec-
tion of Molgolaimidae fam. n., and remarks on the
systematic position of Paramicrolaimus (Nema-
toda, Desmodorida). Zoologica Scripta. 7: 159–173.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Insti-
tuted. 167p.
Cutícula finamente estriada. Arranjo cefálico
6+6+4. Primeiro e segundo círculo de sensilas
não visível. Apenas o terceiro círculo, setiforme,
é visualizado. Cavidade bucal fracamente
esclerotizada, estreita e com dentes
pequenos. Anfídeo circular, posterior à
constrição cefálica. Faringe cilíndrica
terminando em um bulbo esférico muscular e
com lúmen fracamente esclerotizado. Cárdia
longa. Poro secretor-excretor geralmente
localizado anteriormente ao anel nervoso e
raramente posterior a este. Fêmeas didélficas
com ovários opostos refletidos. Machos
com um único testículo anterior, não refletido
e com o ramo posterior reduzido ou ausente.
Espículas de formato variável de acordo com
a espécie. Gubernáculo com ou sem
apófise. Suplementos pré-cloacais
geralmente presentes. Cauda variando de curta
e cônica a finamente alongada com porção pos-
terior cilíndrica.
750, 1050, 1350, 1650 e 1950 metros.
Indivíduos deste gênero podem medir de 0,8 a
1,5 mm. De acordo com Lorenzen (1981),
Molgolaimus pertence à família Desmodoridae
por possuir ovários refletidos, além de algumas
outras características dessa família. Nas
fêmeas, é observada a presença de espermateca
em algumas espécies. Na revisão de
Microlaimidae, Jensen (1978) retirou várias
espécies de Molgolaimus e distribuiu em
Microlaimus, devido à presença de um único
testículo anterior e de ovários refletidos.
Comentários
Ditlevsen, 1921
Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
Diagnose

109
Fig. 41. Molgolaimus - A: aspecto geral; B, C: região anterior; D: região posterior e estruturas copulatórias.
Família Desmodoridae

110
Pseudonchus
De Coninck, L. A. P. (1942) De Symmetrie-
verhoudingen ann het Vooreinde der (Vrijlevende)
Nematoden. Natuurw. Tijdschr. 24 (2–4): 29–68.
Decraemer, W., & Smol, N. (2006) Orders
Chromadorida, Desmodorida and Desmoscolecida.
In: Eyualem-Abebe, Traunspurger, W. & Andrássy,
I. (eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 497-573). CABI Publishing, UK.
Cutícula com finas estrias transversais. Ala
ventral delgada presente na cutícula. Arranjo
cefálico 6+6+4. Primeiro círculo papiliforme,
segundo e terceiro setiformes, sendo as setas
do terceiro círculo mais longas. Setas
subcefálicas presentes. Cavidade bucal
bilateral ampla e tubular, dividida em
duas seções, com feixes de dentes
ventrosublaterais no nível da subdivisão e
também na seção anterior. Anfídio em
forma de um loop simples ou um
criptoespiral, circular. Fêmeas didélficas,
com ovários refletidos. Machos com um
testículo. Espículas curvadas com porção
proximal arredondada. Gubernáculo
curto, paralelo as espículas. Suplementos
pré-cloacais, quando presentes, no mesmo
nível da ala ventral. Três glândulas caudais.
Cauda cônica. Medem de 1,5 a 3mm.
1350, 1650 e 1950 metros.
Cobb, 1920
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
Pseudonchus é caracterizado pelo fato do
setor dorsal da cavidade bucal (cheilostoma,
protostoma e telostoma) ter desaparecido,
de forma que os dois setores sub-laterais
restantes estendem-se até à região dorsal
para preencher o vazio, tornando-se
laterais. Como resultado, a simetria trira-
dial da cavidade bucal é substituída por uma
simetria bilateral, presente tanto nos lábios
(cheilostoma), como na cavidade bucal
propriamente dita (protostoma).
Comentários
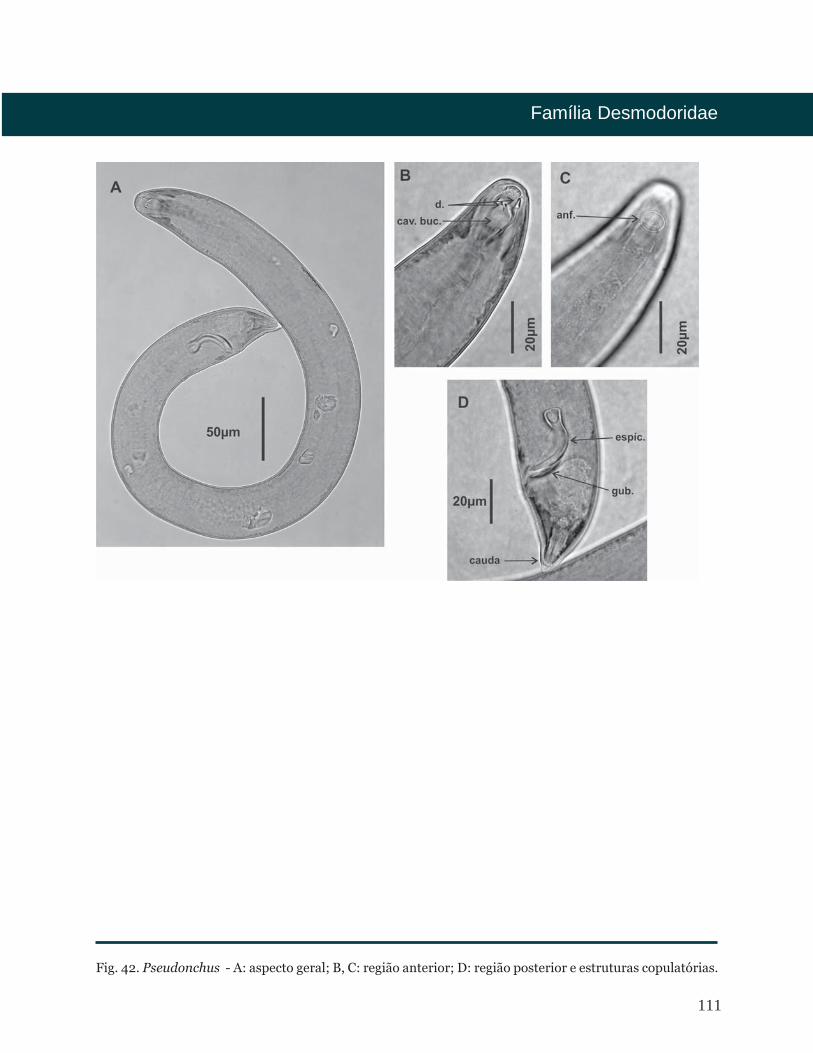
111
Fig. 42. Pseudonchus - A: aspecto geral; B, C: região anterior; D: região posterior e estruturas copulatórias.
Família Desmodoridae

112
Spirinia
Luc, M.; De Coninck, L. A. (1959) Nématodes libres
de la région de Roscoff. Archs. Zool. Exp. gén. 98:
103–165.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Decraemer, W., & Smol, N. (2006) Orders
Chromadorida, Desmodorida and Desmoscolecida.
In: Eyualem-Abebe, Traunspurger, W. & Andrássy,
I. (eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 497-573). CABI Publishing, UK.
Cutícula finamente estriada. Cápsula
cefálica ausente. Arranjo cefálico 6+6+4.
Os dois primeiros círculos papiliformes e
o terceiro círculo setiforme. Setas
somáticas arranjadas em oito feixes.
Cavidade bucal relativamente pequena ou
de um tamanho mediano, com um dente
pequeno dorsal, dois dentes
ventrosublaterais menores presentes ou
ausentes. Anfídio em simples espiral,
usualmente na porção anterior da cabeça,
circundado pelas anelações cuticulares.
Faringe com bulbo terminal curto e
redondo. Fêmeas didélficas com ovários
refletidos. Machos com um único
testículo estendido. Espículas com ve-
lum e capitulum proeminentes.
Gubernáculo simples, sem apófise,
paralelo às espículas. Suplementos
ausentes. Cauda, usualmente, cônica.
750, 1050, 1350, 1650 e 1950 metros.
Gerlach, 1963
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
Indivíduos que medem de 2 a 4 mm.
Spirinia schneideri Villot, 1875 apresenta
dimorfismo sexual na cauda. A fêmea
possui cauda cônica, no entanto, o macho
possui cauda subhemisférica, com uma
ponta terminal.
Comentários
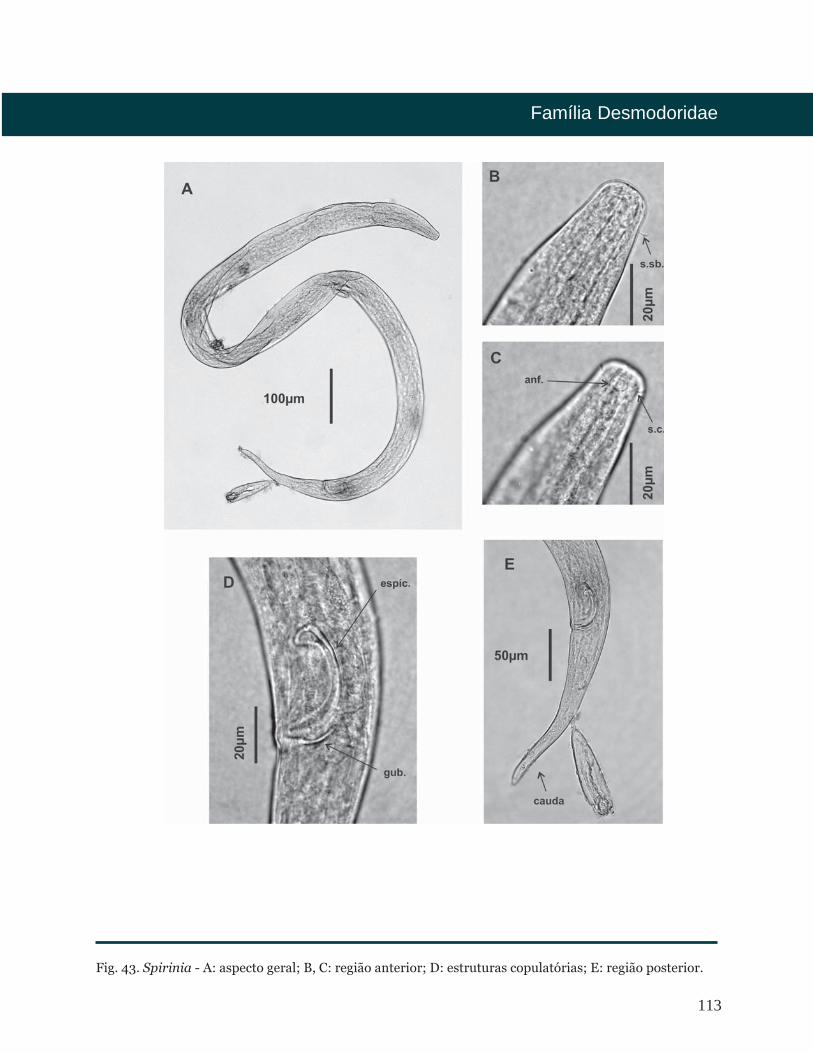
113
Fig. 43. Spirinia - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Desmodoridae

114
Bolbolaimus
Jensen, P. (1978) Revision of Microlaimidae, Erec-
tion of Molgolaimidae fam. n., and Remaks on the
Systematic Position of Paramicrolaimus (Nema-
toda, Desmodorida). Zoologica Scripta. Vol. 7, 159–
173.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Insti-
tuted. 167p.
Cutícula fortemente estriada. Cápsula
cefálica ausente. Arranjo cefálico 6+6+4,
sendo seis papilas labiais internas, seis
setas labiais externas e quatro cefálicas
posteriores. Cavidade bucal esclerotizada,
com um sólido dente dorsal e pequenos
dentes subventrais. Em algumas espécies
existe uma porção adicional na região
bucal que contém outro dente posterior.
Anfídeo circular circundado parcial ou
completamente pelas estriações
cuticulares. Bulbo levemente desenvolvido
na região periférica anterior da cavidade
bucal. Bulbo esôfágico e cárdia presentes.
Fêmeas didélficas com ovários
opostos estendidos. Machos com dois
testículos. Espículas curvadas.
Gubernáculo simples, sem apófise,
paralelo às espículas. Algumas espécies
apresentam suplementos pré-cloacais
tubulares. Cauda cônica.
750 e 1350 metros.
Cobb, 1920
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
Indivíduos presentes neste gênero podem
medir de 1 a 2 mm. As principais
características diferenciais para
Bolbolaimus são: presença de bulbo mus-
cular na região anterior e uma forte
cuticularização da cavidade bucal.
Comentários
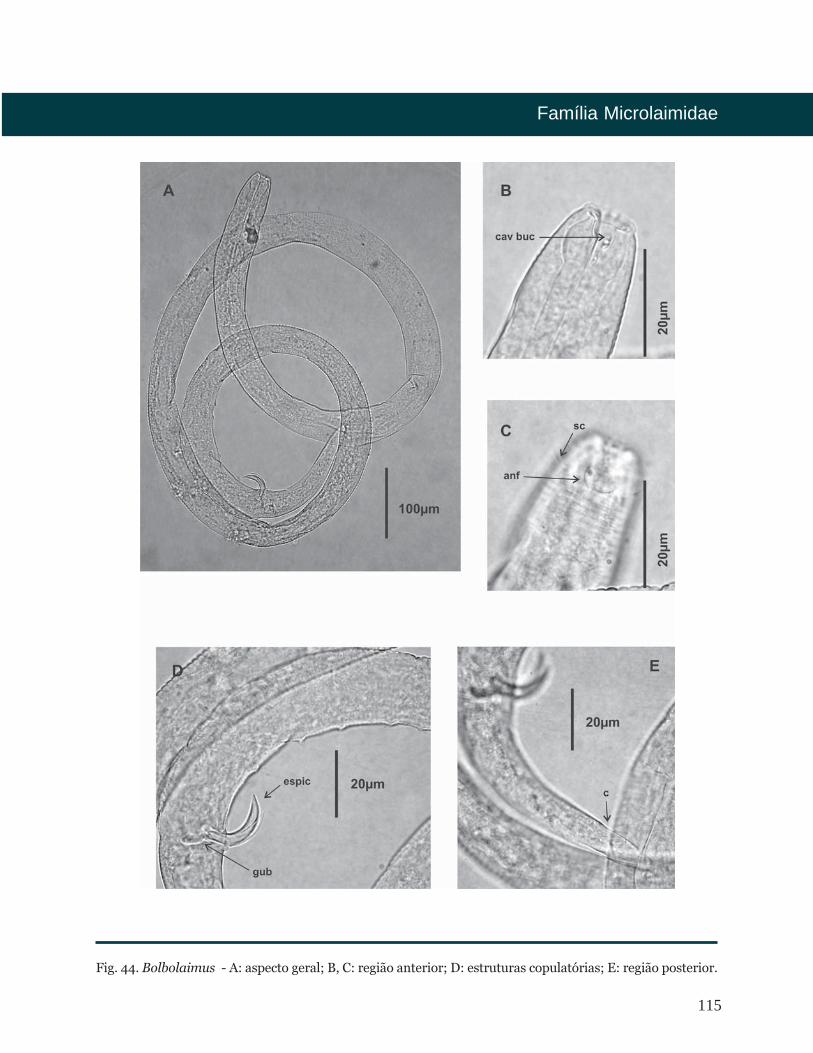
115
Fig. 44. Bolbolaimus - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Microlaimidae

116
Microlaimus
Jensen, P. (1978) Revision of Microlaimidae, erec-
tion of Molgolaimidae fam. n., and remarks on the
systematic position of Paramicrolaimus (Nema-
toda, Desmodorida). Zoologica Scripta. 7: 159–173.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Insti-
tuted. 167p.
Cutícula com anelações, porém em alguns
casos pontuada ou com barras
longitudinais. Arranjo cefálico 6+6+4. As
seis sensilas labiais externas, usualmente
setosas, seguidas por quatro setas cefálicas,
usualmente mais longas que as labiais
externas. Setas somáticas curtas,
ocasionalmente, presentes. Cavidade bucal
cuticularizada. Anfídio transversalmente
circular ou longitudinalmente oval, ou
ainda com tendência à espiralização.
Faringe algumas vezes envolvendo a
cavidade bucal. Bulbo esofágico oval ou
piriforme. Fêmeas didélficas com
ovários opostos estendidos. Machos
com um ou dois testículos. Espículas
geralmente curtas. Gubernáculo sem
apófise. Suplementos pré-cloacais
ausentes. Cauda cônico-cilíndrica.
750, 1050, 1350, 1650 e 1950 metros.
De Man, 1880
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
Indivíduos que medem de 0,5 a 2 mm. Na
revisão de Microlaimidae, Jensen (1978)
elevou a Subfamília Molgolaiminae ao
nível de Família, tornando-se então
Molgolaimidae.
Comentários

117
Fig. 45. Microlaimus - A: aspecto geral; B, C: região anterior; D: região posterior ; E: estruturas copulatórias.
Família Microlaimidae

118
Spirobolbolaimus
Soetaert, K. & Vincx, M. (1988) Spirobolbolaimus
bathyalis, gen. nov., sp. nov. (Nematoda,
Microlaimidae) from the Mediterranean (Calvi).
Hydrobiologia 164 : 33–38.
Gourbault, N. & Vincx, M. (1990) Chromadorida
(Nematoda) from Guadeloupe and Polynesia with
evidence of intersexuality. Zoologica Scripta 19(1):
31–37.
Cutícula estriada. Cápsula cefálica
presente. Arranjo cefálico 6+6+4. Seis
papilas labiais internas, seis setas labiais
externas e quatro papilas ou setas cefálicas
presentes. Sensilas somáticas
papiliformes, dispostas em quatro feixes
submedianos. Fileiras de setas cervicais na
região do pescoço. Cavidade bucal com três
fortes dentes, sendo um dorsal e um par
de dentes subventrais. Anfídio
multiespiral, fortemente esclerotizado.
Faringe com um bulbo na porção anterior
e outro posterior. Fêmeas didélficas, com
ovários opostos estendidos. Machos
com dois testículos opostos e estendidos.
Espículas fortemente esclerotizadas,
com capitulum estreito. Gubernáculo
sem apófise dorsal. Cauda cônica.
1350 metros.
Soetaert & Vincx, 1988
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
Indivíduos que podem medir de 0,7 a 1,7
mm. Neste gênero foi encontrado o
fenômeno de intersexualidade na espécie
Spirobolbolaimus boucherorum
Gourbault & Vincx (1990).
Comentários

119
Fig. 46. Spirobolbolaimus - A: aspecto geral; B, C: região anterior; D: região posterior; E: estruturas copulatórias.
Família Microlaimidae

OR
DE
M D
ES
MO
SC
OL
EC
IDA
FILIPJEV
, 19
29
CARACTERÍSTICAS GERAIS
Figura 47: Porção anterior do corpo de gêneros representantes das Famílias de
Desmoscolecida (Decraemer & Smoll, 2006) que ocorrem na Bacia de Campos: A=
Desmoscolecidae; B= Cyartonematidae.
A Ordem Desmoscolecida apresenta corpo conspicuamente
anelado; anéis sólidos presentes ou ausentes; cutícula plana ou
ornamentada com escalas, verrugas ou espinhos; cabeça com duas setas
cefálicas subdorsais ou subventrais; anfídio vesicular, circular ou oval;
setas somáticas tubulares, freqüentemente abertas ou diferenciadas na
ponta; esôfago levemente alargado posteriormente, não sobrepondo-se
ao intestino; ocelo pode estar presente, normalmente, posterior à base
do esôfago; células intestinais usualmente com glóbulos grandes e pálidos;
fêmeas usualmente com ováriosopostos estendidos; machos com
um ou dois testículos estendidos; espículas pareadas, de mesmo
comprimento e usualmente com região proximal cefalada; gubernáculo
usualmente presente; três glândulas caudais; espinerete terminal.
Na nova classificação proposta por De Ley & Blaxter (2002),
Desmoscolecida é novamente reconhecida como um grupo separado de
Chromadorida. Esta classificação não está de acordo com Lorenzen (1981,
1994) que coloca Desmoscolecida como Subordem de Chromadorida.
É composta pelas Famílias: Desmoscolecidae, Meyliidae e
Cyartonematidae. São essencialmente marinhos, porém também são
encontrados em salt marches, terrestres e água doce. (Figura 6).

122
Desmoscolex
Cutícula com 12 a 44 desmens ovais
simetricamente separados por claras zonas
cuticulares de igual largura ou mais finas que
os desmens. Interzonas com 1 ou 5 anéis
descobertos ou cobertos com feixes
transversais de espinhos ou poros. Arranjo
cefálico de 6+6+4, sendo os dois primeiros
círculos de muito difícil visualização. As
quatro setas cefálicas inseridas em pedúnculos
na porção anterior da cabeça. Setas somáticas
alternam pares de espinhos subdorsais e
subventrais. Anfídio circular ou oval (como
uma bolha), nunca espiral, com um corpus
gelatum vesicular. Boca pequena. Faringe
curta e cilíndrica, estendendo-se até a altura
do segundo ou terceiro anel, nas espécies com
17–18 anéis. Porção posterior do intestino em
saco fechado (excepcionalmente ausente),
pós-cloacal. Fêmeas didélficas com ovários
opostos estendidos e uma espermateca em
cada lado. Machos com um único testículo.
Espículas de formas variadas de acordo com
a espécie. Gubernáculo curto. Último
desmen da cauda com espinho dorsal.
750, 1050, 1350, 1650 e 1950 metros.
Claparède, 1863
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Timm, R. W. (1970) A Revision of the Nematode
Order Desmoscolecida Filipjev, 1929. University of
California Publications in Zoology, Berkeley and
Los Angeles. Volume 93, pp 1–115.
Decraemer, W. (1985) Revision and phylogenetic
systematics of the Desmoscolecida (Nematoda).
Hydrobiologia 120, 259–283.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Lorenzen, S. (1994) The phylogenetic systematics of
freeliving Nematodes. The Ray Society Instituted. 167p.
Decraemer, W., & Smol, N. (2006) Orders
Chromadorida, Desmodorida and Desmoscolecida.
In: Eyualem-Abebe, Traunspurger, W. & Andrássy,
I. (eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 497–573). CABI Publishing, UK.
Bibliografia
Indivíduos deste gênero medem de 0,1 a 0,5
mm. De acordo com Platt & Warwick (1988),
os espécimes considerados típicos deste gênero
apresentam 17 desmens ao longo de seu
comprimento total. O número de desmens
presentes no animal é bem menor que o
número de anelações observadas no corpo.
Comentários
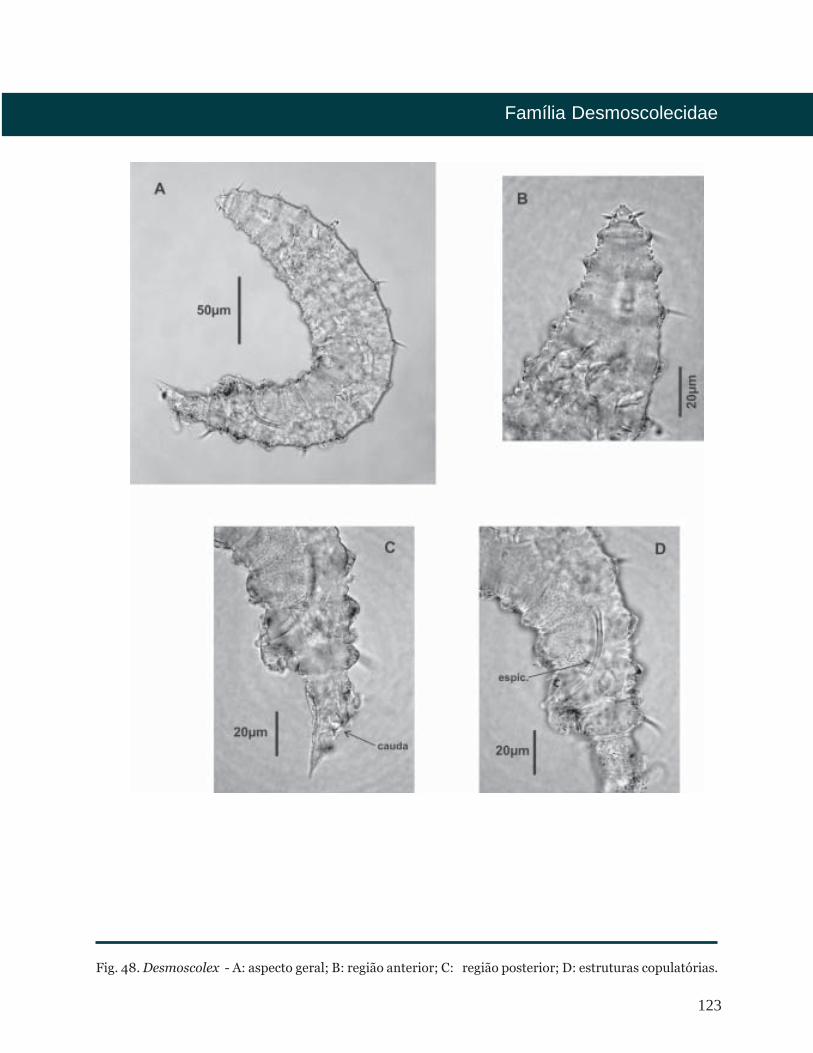
123
Fig. 48. Desmoscolex - A: aspecto geral; B: região anterior; C: região posterior; D: estruturas copulatórias.
Família Desmoscolecidae

124
Subgênero Desmolorenzenia
Corpo pequeno, afunilando-se nas
extremidades, composto de 18 desmens
assimétricos, completos ou parcialmente
cobertos, separados por interzonas mais
estreitas ou de igual largura. Porção visível
da cutícula apresentando, às vezes, uma leve
anelação. Apresenta desmen duplo, a
partir do qual a direção dos desmens muda.
Esse desmen duplo não é separado por
uma interzona larga e é coberto com uma
camada contínua de secreção e material
agregado. Arranjo cefálico 6+6+4, os dois
primeiros círculos são de muito difícil
visualização. As quatro setas cefálicas
inseridas em pedúnculos, na porção ante-
rior da cabeça. Setas somáticas apresentam
o mesmo padrão de distribuição dos
Desmoscolecidae. Anfídio circular ou oval.
Cavidade bucal curta, com paredes grossas.
Faringe curta e cilíndrica até o nível do anel
nervoso afunilando-se depois do mesmo.
Intestino anteriormente com células
glandulares finas (porção ventricular) logo
expandindo-se em uma porção cilíndrica
com inclusões globulares pequenas e
grandes. Porção posterior do intestino em
saco fechado, pós-cloacal. Fêmea didélfica
com ovários opostos. Vulva de difícil
visualização. Macho com um único
testículo. Espículas e gubernáculo curtos.
Último desmen da cauda com espinho
dorsal.
750, 1050, 1350 e 1950 metros.
Freudenhammer, 1975
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Indivíduos pequenos que podem medir de
0,2 a 0,3 mm.
Comentários
Timm, R. W. (1970) A Revision of the Nematode
Order Desmoscolecida Filipjev, 1929. University of
California Publications in Zoology, Berkeley and
Los Angeles. Volume 93, pp 1–115.
Decraemer, W. (1985) Revision and phylogenetic
systematics of the Desmoscolecida (Nematoda).
Hydrobiologia 120, 259–283.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Bibliografia

125
Fig. 49. Subgênero Desmolorenzenia - A: aspecto geral; B: região anterior; C: estruturas copulatórias;
D: região posterior.
Família Desmoscolecidae

126
Greefiella
Timm, R. W. (1970) A Revision of the Nematode
Order Desmoscolecida Filipjev, 1929. University of
California Publications in Zoology, Berkeley and
Los Angeles. Volume 93, pp 1–115.
Timm, R. W. (1978) Marine nametodes of the or-
der Desmoscolecida from McMurdo Sound, Ant-
arctica. Biology of the Antarctic seas VI. Antarctic
Research Series, 26: 225–236.
Decraemer, W. (1985) Revision and phylogenetic
systematics of the Desmoscolecida (Nematoda).
Hydrobiologia 120, 259–283.
Decraemer, W., & Smol, N. (2006) Orders
Chromadorida, Desmodorida and Desmoscolecida.
In: Eyualem-Abebe, Traunspurger, W. & Andrássy,
I. (eds) Freshwater Nematodes: Ecology and Tax-
onomy (pp. 497–573). CABI Publishing, UK.
Indivíduos pequenos com anelações
simples presentes no corpo, nunca
desmens. Cada anelação adornada por
numerosos espinhos maiores que o
diâmetro de cada anel. Cabeça
arredondada. Inúmeras setas presentes na
cabeça. Setas subdorsais e subventrais
presentes entre as fileiras de espinhos nas
anelações. Cavidade bucal ausente ou de
difícil viaualização. Anfídio grande e
elíptico, mais largo que a cabeça. Ocelo
presente. Fêmea didélfica com ovários
opostos. Macho com um único testículo
estendido. Gubernáculo ausente.
Espículas retas e estreitas. Três glândulas
caudais presentes. Cauda cônica.
750, 1050 e 1050 metros.
Cobb, 1922
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
Timm (1978) cita a presença de
aproximadamente 80 setas na região
cefálica.
Comentários
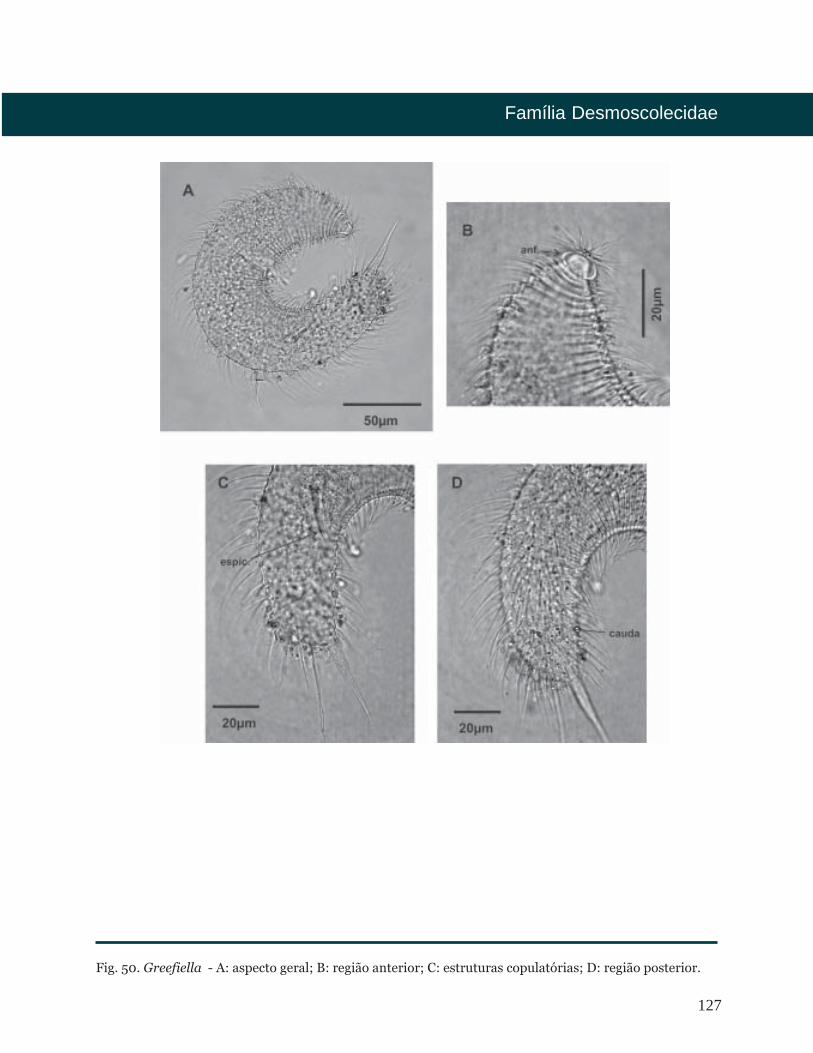
127
Fig. 50. Greefiella - A: aspecto geral; B: região anterior; C: estruturas copulatórias; D: região posterior.
Família Desmoscolecidae

128
Tricoma
Timm, R. W. (1970) A Revision of the Nematode
Order Desmoscolecida Filipjev, 1929. University of
California Publications in Zoology, Berkeley and
Los Angeles. Volume 93, pp 1–115.
Decraemer, W. (1985) Revision and phylogenetic
systematics of the Desmoscolecida (Nematoda).
Hydrobiologia 120, 259–283.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Cutícula com 29 a 240 desmens ovais
simétricos, muito próximos, separados uns
dos outros por interzonas de um anel cu-
ticular. Não apresenta anel de inversão.
Cabeça aproximadamente triangular com
espessas paredes cuticulares. Anfídio ve-
sicular, corpus gelatum sem estrutura
espiral. Setas somáticas seguem o padrão
tricomóide (setas sub-ventrais
usualmente mais numerosas que as sub-
dorsais, em par ou não; anel final sem
setas). Região labial com seis lábios
separados, portando uma ou duas coroas
de seis papilas. Faringe mais ou menos
cilíndrica. Fêmeas didélficas, com
ovários opostos e estendidos. Machos
com dois testículos. Espículas curvadas.
Gubernáculo presente. Anel final
normalmente cilíndrico, coberto
anteriormente por uma faixa de desmen,
afunilando-se posteriormente ao nível do
espinerete. Poucas espécies possuem o
espinerete em forma de espinho.
1050 e 1350 metros.
Cobb, 1894
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
Indivíduos que medem de 0,2 a 1,3 mm. O
número de desmens presentes no animal,
corresponde ao número de anelações
observadas no corpo. Um Tricoma típico
possui de 60 a 80 desmens.
Comentários

129
Fig. 51. Tricoma - A: aspecto geral; B: região anterior; C: estruturas copulatórias e região posterior.
Família Desmoscolecidae

130
Subgênero Quadricoma
Timm, R. W. (1970) A Revision of the Nematode
Order Desmoscolecida Filipjev, 1929. University of
California Publications in Zoology, Berkeley and
Los Angeles. Volume 93, pp 1–115.
Decraemer, W. (1985) Revision and phylogenetic
systematics of the Desmoscolecida (Nematoda).
Hydrobiologia 120, 259–283.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Cutícula com 33 a 66 desmens
assimétricos, mais amplos em sua borda
anterior quando situados na porção ante-
rior e se tornam opostos no restante do
corpo. Possui um anel a partir do qual
ocorre a inversão. Visualmente, não há
espaços entre os desmens devido à
proximidade entre eles. Setas subdorsais
e subventrais presentes. Cabeça ampla, tri-
angular com pedúnculos de inserção das
setas cefálicas. Anfídio circular que cobre
quase totalmente a cabeça. Cavidade bucal
pequena e estreita. Ocelo presente.
Fêmeas didélficas, com ovários
opostos e estendidos. Machos com dois
testículos. Espículas longas e delgadas.
Gubernáculo presente ou ausente.
Cauda cônica com região terminal curta.
750, 1050, 1350 e 1950 metros.
Filipjev, 1922
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
Animais que medem de 0,2 a 1,4 mm. Este
sub-gênero de Tricoma, é definido pelo
conjunto das características: forma dos
anéis, presença de anel de inversão, forma
do espinerete e forma da cabeça.
Comentários

131
Fig. 52. Subgênero Quadricoma - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E:
região posterior.
Família Desmoscolecidae

OR
DE
M M
ON
HY
ST
ER
IDA
FILIPJEV
, 19
29A Ordem Monhysterida é caracterizada por representantes com cutícula
finamente estriada ou com anelações; arranjo cefálico em três círculos
separados (6 + 6 + 4) ou somente dois círculos (6 + 10); anfídio,
geralmente, circular; fêmeas monodélficas ou didélficas com ovários
estendidos; machos com dois testículos ou somente um anterior;
suplementos pré-cloacais raramente presentes; glândulas caudais,
abrindo-se separadamente ou através de um único poro.
Esta Ordem é representada por nematódeos principalmente marinhos, mas
existem alguns indivíduos livres terrestres (Coomans & Eyualem-Abebe, 2006).
São animais pequenos ou de comprimento médio com o corpo afunilado nas
extremidades. De acordo com Lorenzen (1994) a Ordem Monhysterida era
composta por 4 Superfamílias: Monhysteroidea, Sphaerolaimoidea,
Siphonolaimoidea e Axonolaimoidea. Esta classificação era baseada somente
no sistema reprodutor das fêmeas que apresentam ovários estendidos. Após
as recentes análises moleculares, De Ley & Blaxter (2001, 2004), retiraram a
Superfamília Axonolaimoidea. As três superfamílias incluem as Famílias
Monhysteridae, Sphaerolaimidae, Xyalidae e Linhomoeidae (figura 7). Em De
Ley et al. (2006) a ordem encontra-se dividida em duas Subordens:
Monhysterina, com as Superfamílias: Monhysteroidea (Família
Monhysteridae) e Sphaerolaimoidea (Famílias Sphaerolaimidae e Xyalidae) e
Linhomoeina com a Superfamília Siphonoloimoidea (Famílias
Siphonolaimidae, Linhomoidae e Fusivermidae).
CARACTERÍSTICAS GERAIS
Figura 53: Porção anterior do corpo de gêneros representantes das Famílias de Monhysterida,
segundo Coomans & Eyualem-Abebe (2006) que ocorrem na Bacia de Campos: A=
Monhysteridae; B= Sphaerolaimidae; C= Xyalidae; D= Linhomoeidae, E= Siphonolaimidae.

134
Metasphaerolaimus
Gourbault, N.; Boucher, B. (1981) Nematodes
abyssaux (Champgne Walda N/O “Jean Charcot”).
III Une sous-familie et six especes nouvelles de
Sphaerolaimidae. Bulletin Museeu Nationale
Histoire Natural, Paris, 4e s., 3, sect A, (4): 1035-
1052.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Insti-
tuted. 167p.
Warwick, R. M.; Platt, H. M., Sommerfield, P.
(1998) Free-living Marine Nematodes Part III –
Monhysterids. Synopses of the British Fauna (New
Series) Eds: Doris M. Kermack e R.S.K. Barnes. 53,
296p.
Cutícula levemente estriada. Arranjo
cefálico de seis papilas labiais, seis setas
cefálicas curtas e quatro longas, além de oito
grupos de setas subcefálicas. Poucas setas
cervicais. Cavidade bucal com seis
mandíbulas em forma de “H”, presas
anteriormente e articuladas posteriormente
em placas sólidas. Anfídio circular
situado posterior à cavidade bucal, sendo
maior nos machos. Lumen faringeal
espesso e cuticularizado. Fêmeas
monodélficas com ovário posicionado
anteriormente. Machos com um testículo
anterior. Espícula reta e curta.
Gubernáculo presente ou ausente. Três
glândulas caudais visiveis. Cauda cônica.
750, 1050, 1350, 1650 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Gourbault & Boucher, 1981
Na descrição original feita por Gourbault
& Boucher (1981) é relatado o fato de este
gênero possuir um dimorfismo sexual
relacionado ao anfídio circular que se
apresenta maior nos machos. Fato este
registrado na espécie de M. cancellatus
Gourbault & Boucher, 1981.
Comentários

135
Fig. 54. Metasphaerolaimus - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E:
região posterior.
Família Sphaerolaimidae

136
Daptonema
Warwick, R. M.; Platt, H. M., Sommerfield, P.
(1998) Free-living Marine Nematodes Part III –
Monhysterids. Synopses of the British Fauna (New
Series) Eds: Doris M. Kermack e R.S.K. Barnes. 53,
296p.
Coomans, A., & Eyualem-Abebe. (2006) Order
Monhysterida. In: Eyualem-Abebe, Traunspurger,
W. & Andrássy, I. (eds) Freshwater Nematodes:
Ecology and Taxonomy (pp. 574-603). CABI Pub-
lishing, UK.
Cutícula estriada. Arranjo cefálico 6+10 ou
12. Seis papilas labiais internas, seis setas
labiais externas e quatro cefálicas, em um
único círculo onde as cefálicas são iguais ou
menores que as labiais. Freqüentemente
ocorrem setas cefálicas adicionais. Setas
somáticas, presentes. Cheilostoma
amplo, em forma de cúpula. Cavidade
bucal simples, em forma de funil e sem
dentes. Anfídio circular. Fêmeas
monodélficas com ovário estendido,
situado ao lado esquerdo do intestino.
Machos usualmente com dois testículos,
sendo o anterior ao lado esquerdo e poste-
rior ao lado direito do intestino. Espículas
curtas, cerca de um diâmetro cloacal e
dobradas em forma de “L”. Gubernáculo
freqüentemente com peças-guia laterais e
raramente com apófise dorso-caudal.
Cauda cilíndrica e com setas terminais.
750, 1050, 1350, 1650 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Cobb, 1920
Indivíduos que podem medir de 0,5 a 2
mm. Daptonema é um gênero muito
próximo de Theristus, sendo separado
deste somente pela forma da cauda e a
presença de setas terminais.
Comentários
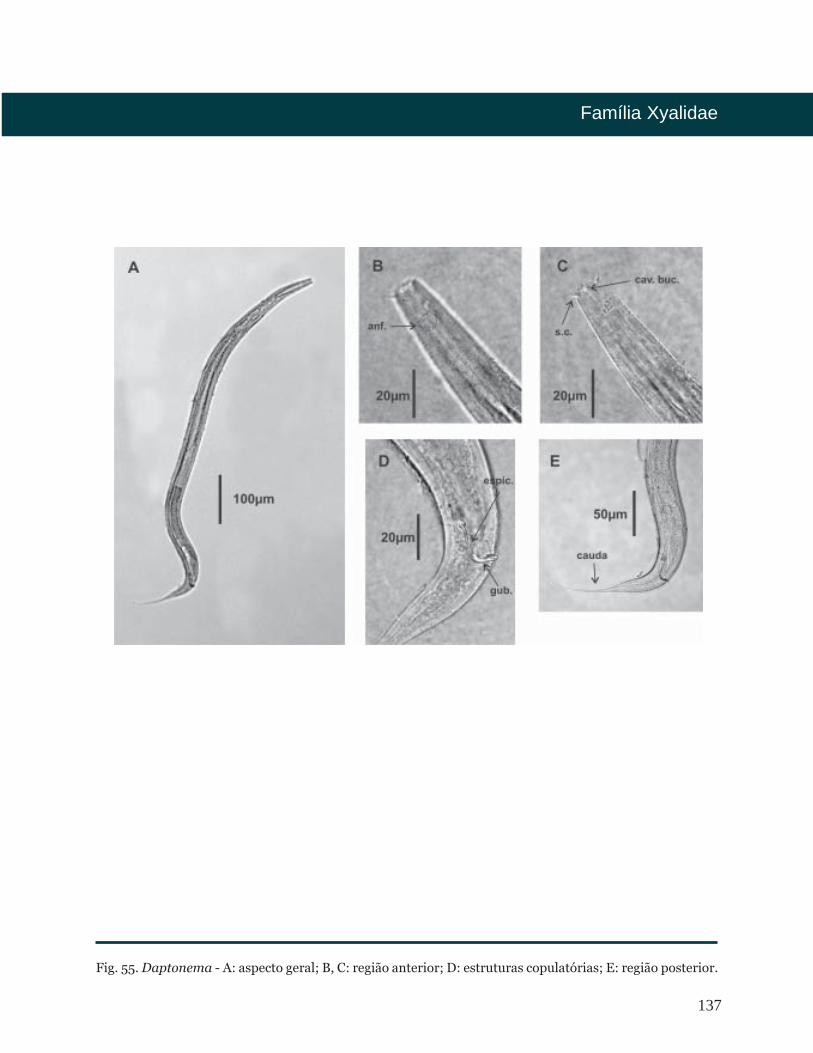
137
Fig. 55. Daptonema - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Xyalidae

138
Elzalia
Timm, R. W. (1961) The marine nematodes of the
Bay of Bengal. Proc. Pakist. Acas. Sci. 1: 1-88.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
167p.
Zhang, Y.; Zhang, Z. N. (2006) Two new species of
the genus Elzalia (Nematoda: Monysterida:
Xyalidae) from the Yellow Sea, China. J. Mar. Biol.
Ass. U. K. 86, 1047–1056.
Cutícula com estriações bem
desenvolvidas. Arranjo cefálico 6+10. Seis
papilas labiais internas, seguidas por seis
setas labiais externas e quatro setas
cefálicas num único círculo. Cavidade
bucal profunda, cilíndrica e fortemente
esclerotizada. Anfídio levemente oval,
atingindo até 90% do diâmetro da região
correspondente do corpo, possivelmente
com uma abertura transversal indistinta.
Faringe alargada, sem bulbo basal. Cárdia
não proeminente. Fêmeas monodélficas
com ovário prodélfico. Machos com um
único testículo anterior. Espículas
alongadas. Gubernáculo simples e
pequeno. Cauda com três setas terminais.
750, 1050, 1350, 1650 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Gerlach, 1957
Indivíduos presentes neste gênero podem
medir de 0,5 a 1 mm. O gubernáculo de
E. gerlachi e E. striatitenuis apresenta uma
estrutura complexa, com a presença de
algumas estruturas acessórias.
Comentários
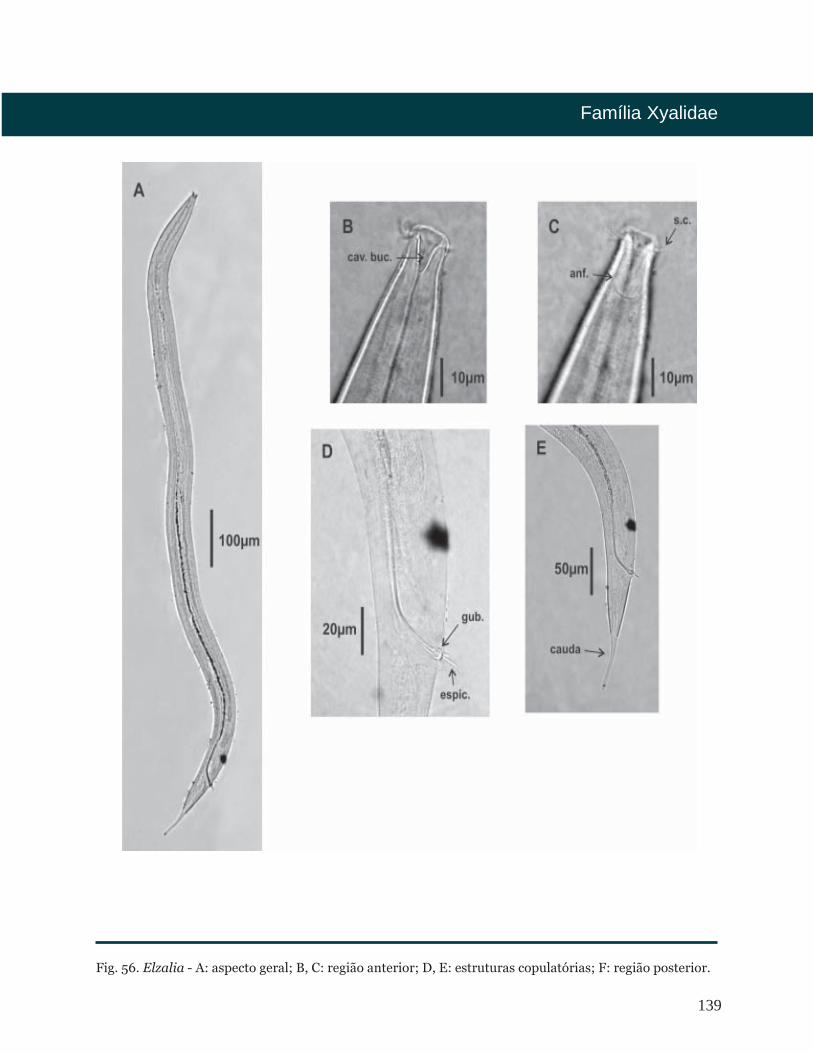
139
Fig. 56. Elzalia - A: aspecto geral; B, C: região anterior; D, E: estruturas copulatórias; F: região posterior.
Família Xyalidae

140
Theristus
Filipjev, I. (1922) Encore sur Nématodes libres de
la Mer Noire. Trudy stavropol “sel”. Khor. Inst. 1:
83–184.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
167p.
Warwick, R. M.; Platt, H. M., Sommerfield, P.
(1998) Free-living Marine Nematodes Part III –
Monhysterids. Synopses of the British Fauna (New
Series) Eds: Doris M. Kermack e R.S.K. Barnes. 53,
296p.
Coomans, A., & Eyualem-Abebe. (2006) Order
Monhysterida. In: Eyualem-Abebe, Traunspurger,
W. & Andrássy, I. (eds) Freshwater Nematodes:
Ecology and Taxonomy (pp. 574-603). CABI Pub-
lishing, UK.
Cutícula estriada. Arranjo cefálico 6+10 ou
6+14. Cavidade bucal simples e sem
dentes. Anfídio circular. Faringe muscu-
lar, cilíndrica. Cárdia visível. Fêmeas
monodélficas ou didélficas, com
ovário(s) estendido(s). Machos com um ou
dois testículos. Espículas curtas.
Gubernáculo com apófise.
Suplementos pré-cloacais presente em
algumas espécies. Cauda cônica e sem setas
terminais.
750, 1050, 1350, 1650, e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Bastian, 1865
Podem medir de 1 a 2 mm. Theristus é
um gênero bastante similar a
Daptonema, exceto pelo tipo de cauda e
a ausência das setas terminais. As fêmeas
de Theristus littoralis Filipjev, 1922
apresentam dois ovários e os machos um
testículo.
Comentários
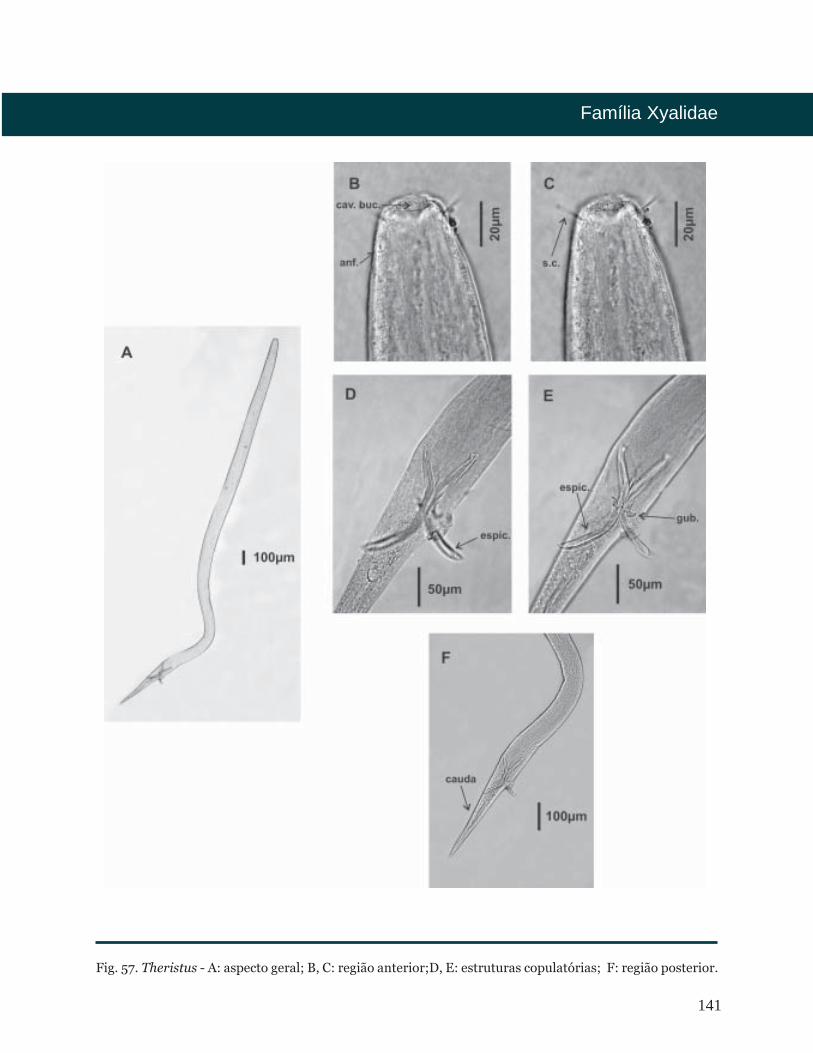
141
Fig. 57. Theristus - A: aspecto geral; B, C: região anterior;D, E: estruturas copulatórias; F: região posterior.
Família Xyalidae

142
Didelta
Cobb, N. A. (1920) One hundred new nemas (type
species of 100 new genera). Contr. Sci. Nematol.
(Cobb) 9: 217-343.
Wieser, W. (1956) Freeliving marine nematodes.
III. Axonolaimoidea and Monhysteroidea. Acta
Univ. lund. (N. F. 2) 52 (13): 1–115.
Warwick, R. M.; Platt, H. M., Sommerfield, P.
(1998) Free-living Marine Nematodes Part III –
Monhysterids. Synopses of the British Fauna (New
Series) Eds: Doris M. Kermack e R.S.K. Barnes. 53,
296p.
Cutícula finamente estriada. Arranjo
cefálico de 6+10 ou 12. Seis papilas labiais
internas muito pequenas ou não visíveis,
seguidas por dez setas cefálicas num
mesmo círculo. Quatro setas médias
cervicais presentes. Cavidade bucal
pequena com paredes finas. Anfídio oval
sobre uma placa esclerotizada. Faringe
cônico-cilíndrica, que se alarga
progressivamente sem formar bulbo.
Cárdia alongada presente. Fêmeas
didélficas. Machos com dois testículos.
Espículas curvadas. Gubernáculo com
apófise dorsal. Suplementos pré-
cloacais freqüentemente ausentes. Cauda
cônica-cilíndrica.
1350 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Cobb, 1920
Indivíduos que podem medir de 2 a 4 mm.
Didelta scutata Wieser, 1956 apresenta 5
a 6 suplementos minúsculos. Embora na
descrição original de Cobb (1920)
mencione que “não há cárdia”,
posteriormente afirma que “o intestino é
separado do esôfago por uma coluna com
1/3 do diâmetro da base do pescoço”.
Comentários
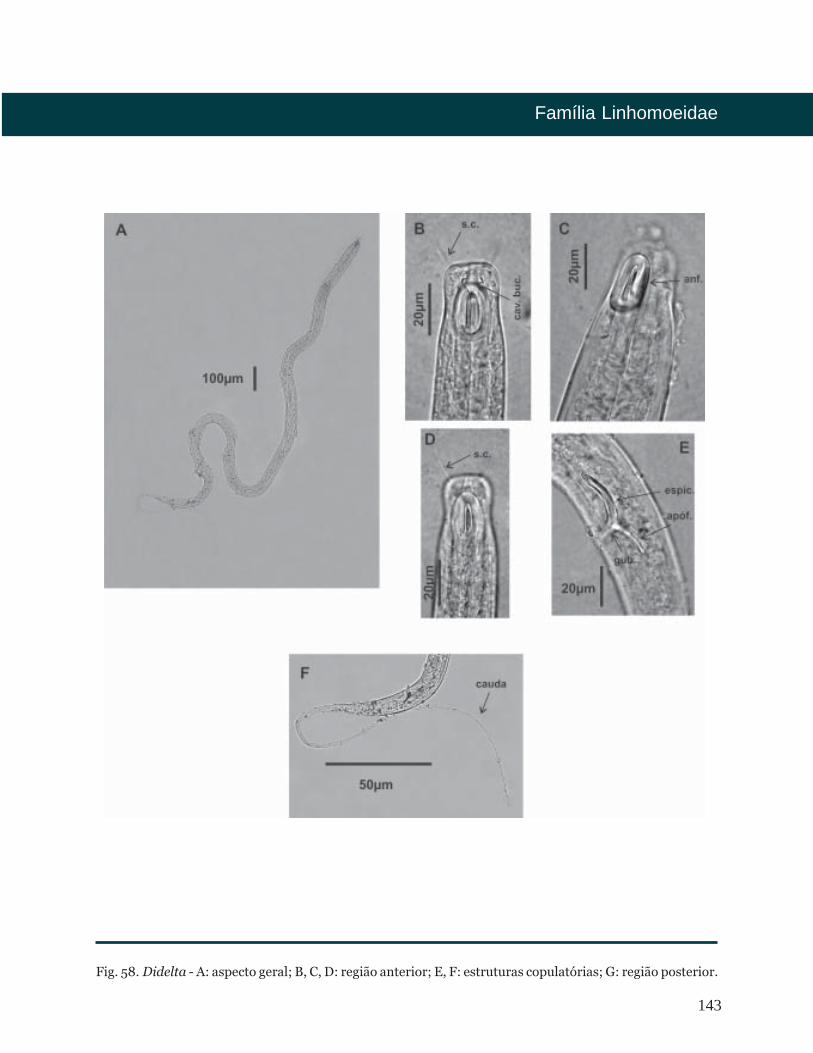
143
Fig. 58. Didelta - A: aspecto geral; B, C, D: região anterior; E, F: estruturas copulatórias; G: região posterior.
Família Linhomoeidae

144
Disconema
Gerlach, S. A. (1963) Freilebende
Meeresnematoden von den Malediven II Kieler
Meereifimehung 19, 67-103;
Vitiello, P. (1969) Linhomoeidae (Nematoda) des
vases profondes du Golfe du Lion. Téthys 1 (2):
493–527.
Warwick, R. M.; Platt, H. M., Sommerfield, P.
(1998) Free-living Marine Nematodes Part III –
Monhysterids. Synopses of the British Fauna (New
Series) Eds: Doris M. Kermack e R.S.K. Barnes. 53,
296p.
Cutícula geralmente estriada e raramente
lisa. Arranjo cefálico: (6)+10 ou 12. Papilas
labiais de difícil visualização. Setas labiais
externas e cefálicas em um mesmo círculo.
Cavidade bucal geralmente ausente,
quando presente é muito pequena e sem
cuticularização. Anfídio curvado,
alongado ou oval. Faringe alarga-se
posteriormente, sem forma bulbo. Sistema
secretor-excretor visível. Cárdia
estendida e destacada do intestino. Fêmeas
geralmente didélficas com ovários
estendidos. Machos com dois testículos
opostos. Usualmente a gônada anterior
posicionada do lado direito ou esquerdo do
intestino e o ramo posterior do lado oposto.
Espículas curvas. Gubernáculo
pequeno e com apófise. Cauda cônica ou
cônico-cilíndrica.
750 metros.
Filipjev, 1918
Bibliografia
Distribuição Batimétrica
Diagnose Distribuição Geográfica
Indivíduos que podem medir de 1 a 4 mm.
De acordo com Vitiello (1969), o
gubernáculo em Disconema é tão
pequeno que se torna de difícil
visualização. Este gênero é exclusivamente
marinho.
Comentários

145
Fig. 59. Disconema - A: aspecto geral; B: região anterior; C: estruturas copulatórias; D:região posterior.
Família Linhomoeidae

146
Metalinhomoeus
Schuurmans Stekhoven, J. H. (1950) The freeliving
marine nemas of the Mediterranean. I The Bay of
Villefranche. Mém. Inst. R. Sci. nat. Belg. 2 Sér. 37:
1–220.
Timm, R. W. (1967) New marine nematodes of the
Family Linhomoeidae from East Pakistan. Proc.
Pak. Acad. Sci. Vol. 4, no 1.
Vitiello, P. (1969) Linhomoeidae (Nematoda) des
vases profondes du Golfe du Lion. Téthys 1 (2):
403–527.
Warwick, R. M.; Platt, H. M., Sommerfield, P.
(1998) Free-living Marine Nematodes Part III –
Monhysterids. Synopses of the British Fauna (New
Series) Eds: Doris M. Kermack e R.S.K. Barnes. 53,
296p.
Cutícula levemente estriada. Região
cefálica com seis papilas labiais internas e
quatro setas cefálicas. Algumas espécies
com 4 setas subcefálicas. Cavidade bucal
pequena e sem dentes. Anfídio circular ou
em forma de loop. Faringe com um bulbo
basal posterior. Cárdia longa e destacada
do intestino. Sistema secretor-excretor
visível. Poro secretor-excretor geralmente
entre o bulbo e o anel nervoso. Fêmeas
didélficas com ovários refletidos. Ma-
chos com um único testículo refletido.
Espícula curvada. Gubernáculo com ou
sem apófise. Cauda cônica ou cônico-
alongada.
750, 1050 e 1350 metros.
De Man, 1907
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
A cavidade bucal pode apresentar paredes
laterais cuticularizadas como em M.
biratus Vitiello, 1969. Na descrição origi-
nal, a faringe apresenta um bulbo basal, no
entanto a espécie M. effilatus Stekhoven,
1950 não possui um bulbo basal
verdadeiro.
Comentários
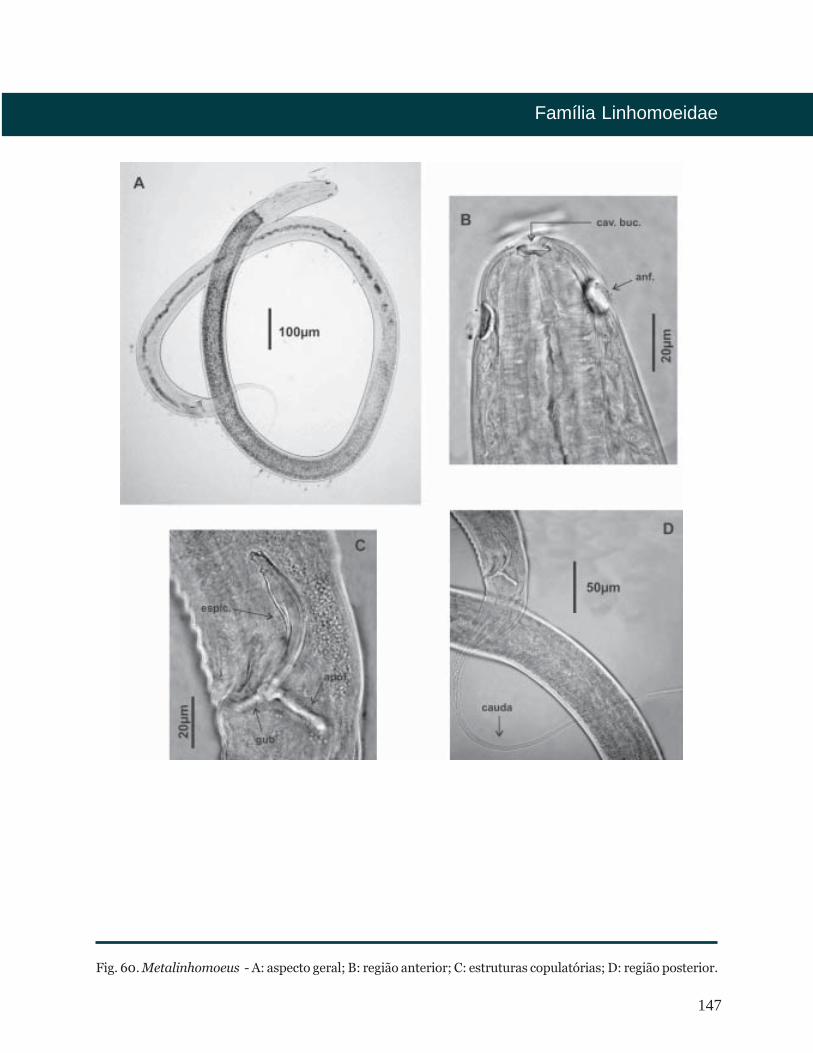
147
Fig. 60. Metalinhomoeus - A: aspecto geral; B: região anterior; C: estruturas copulatórias; D: região posterior.
Família Linhomoeidae

148
Terschellingia
Wieser, W. (1956) Free-living marine nematodes.
III. Axonolaimoidea and Monhysteroidea (Re-
ports…26). Acta Univ. lund., N. F., Avd. 2, 52 (13):
115 p.
Ott, J. A. (1972) Twelve new species of nematodes
from an intertidal sandflat in North Carolina. Int.
Rev. ges. Hydrobiol. Hydrog. 57: 463–469.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
167p.
Warwick, R. M.; Platt, H. M., Sommerfield, P.
(1998) Free-living Marine Nematodes Part III –
Monhysterids. Synopses of the British Fauna (New
Series) Eds: Doris M. Kermack e R.S.K. Barnes. 53,
296p.
Cutícula com leves estriações transversais.
Arranjo cefálico composto de seis setas
labiais cônicas e quatro cefálicas em dois
círculos separados. Quatro setas
subcefálicas situadas na região cervical.
Setas somáticas geralmente ausentes,
porém presentes na região ventral da cauda
dos machos de algumas espécies. Cavidade
bucal pequena ou ausente. Anfídio circu-
lar posicionado no mesmo nível das setas
cefálicas. Faringe com um bulbo terminal.
Cárdia proeminente. Fêmeas geralmente
didélficas com ovários estendidos,
raramente monodélficas. Machos
geralmente com dois testículos estendidos
e raramente com um. Espícula curvada
com uma lamela central presente na
porção proximal. Gubernáculo com
apófise. Cauda cônico-cilíndrica, em
algumas espécies, filiforme.
750, 1050 e 1350 metros.
De Man, 1888
Distribuição GeográficaDiagnose
Distribuição Batimétrica
Bibliografia
Neste gênero existe uma grande variedade
em relação ao tipo de cauda encontrada nas
espécies. Nas descrições ocorre uma
variação desde cônica como em T.
brevicauda Ott, 1972, até longa e filiforme
como em T. longicaudata De Man, 1888.
Comentários
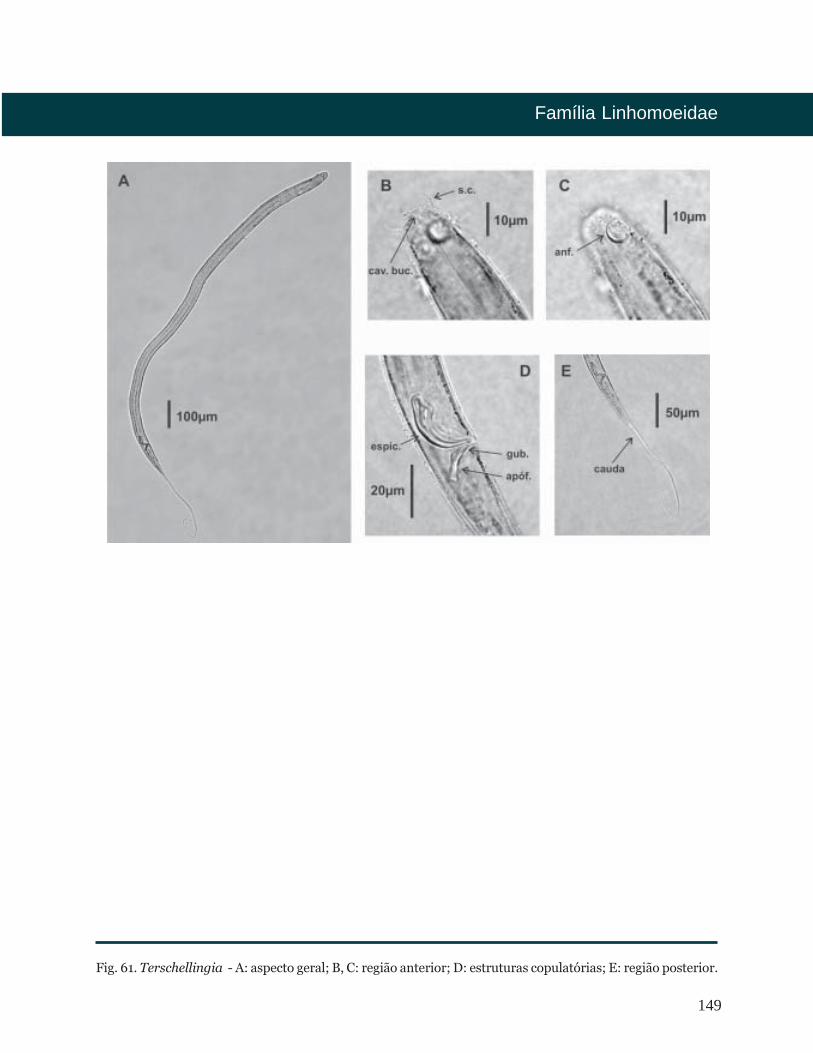
149
Fig. 61. Terschellingia - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Linhomoeidae

150
Bodonema
Jensen, P. (1991) Bodonematidae fam. n. (Nema-
toda, Monhysterida) accommodating Bodonema
vossi gen. et p. n. from the deep-sea benthos of the
Norwegian Sea. – Sarsia 76: 11–15.
Warwick, R. M.; Platt, H. M., Sommerfield, P.
(1998) Free-living Marine Nematodes Part III –
Monhysterids. Synopses of the British Fauna (New
Series) Eds: Doris M. Kermack e R.S.K. Barnes. 53,
296p.
Corpo alargado atenuando-se anteriormente.
Cutícula lisa. Setas cefálicas em três círculos,
com o segundo e a terceiro círculo de setas com
o mesmo comprimento. Anfídio em espiral
única, curvando-se ventralmente. Abertura
bucal pequena, sem dentes, que se continua
numa porção tubular esclerotizada. Faringe
diferenciada em três regiões: 1) região ante-
rior cilíndrica, com lúmen esclerotizado e
paredes musculares pouco desenvolvidas; 2)
região média extensa, fortemente
muscularizada, com vários bulbos e lúmen
esclerotizado; 3) região posterior estreita
com a musculatura pouco desenvolvida isth-
mus, lúmen pouco esclerotizado, com um
bulbo piriforme terminal. Fêmeas didélficas
com ovários opostos e estendidos, o ante-
rior mais desenvolvido que o posterior. Ma-
chos com dois testículos opostos e estendidos.
Espículas simples. Gubernáculo com
apófise. Cauda, cônica, curta, com abertura
subterminal na borda dorsal que se liga a
ductos das glândulas caudais que
aparentemente são atrofiados.
1050 e 1650 metros.
Jensen, 1991
Distribuição GeográficaDiagnose
Distribuição Batimétrica
Bibliografia
Indivíduos que medem de 1,2 a 1,6 mm. Este
gênero é principalmente caracterizado pela
faringe diferenciada em três regiões, não
sendo encontrada em outros Nematoda.
Comentários

151
Fig. 62. Bodonema - A: aspecto geral; B, C, D: região anterior; E: região posterior; F: estruturas copulatórias.
Família Bodonematidae

OR
DE
M A
RA
EO
LA
IMID
A DE CONINK
, 19
65
Araeolaimida foi elevada a Ordem por De Ley & Blaxter (2004) e
possui representantes com as seguintes características: cutícula lisa,
estriada ou pontuada; anfídio predominantemente espiral, em forma
de arco e raramente em forma de poro; sensilas labiais papiliformes, não
visualizadas, setas labiais externas e quatro setas cefálicas geralmente
em dois círculos separados, quatro setas cefálicas são maiores ou do
mesmo comprimento das seis labiais anteriores; cavidade bucal com seis
ou três dentes; faringe sem bulbo basal; fêmeas didélficas com ovários
estendidos, raramente monodélficas; machos com dois testículos ou
um único anterior e papilas pré-anais presentes ou ausentes.
Muthumbi & Vanreusel (2006) também considream o status de
ordem dentro da Superclasse Chromadoria e da Classe Chromadorea.
Araeolaimida incluem quatro Famílias: Axonolaimidae, Comesomatidae,
Diplopeltidae e Coninckiidae, originalmente inseridas na Superfamília
Axonolaimoidea (Figura 8). De Ley et al (2006) manteve a posição de
Ordem, contendo uma Superfamília: Axonolaimoidea e as famílias citadas
anteriormente.
CARACTERÍSTICAS GERAIS
Figura 63: Porção anterior do corpo de gêneros representantes das Famílias de
Araeolaimida, segundo Muthumni & Vanreusel (2006) que ocorrem na Bacia de Cam-
pos: A= Comesomatidae; B= Diplopeltidae; C= Coninckiidae.

154
Cervonema
Wieser, W. (1954) Free-living marine nematodes.
II Chromadoroidea. (Reports...17). Acata. Univ.
Lund., N. F., Avd. 2, 50 (16): 148 p.
Jensen, P. (1979) Revision of Comesomatidae
(Nematoda). Zoologica Scripta. Vol. 8, 81–105.
Platt, H. M. (1985) The freeliving marine nema-
todes genus Sabatieria (Nematoda:
Comesomatidae). Taxonomic revision and pictorial
keys. Zoological Journal of the Linnean Society, 83
27–78.
Warwick, R. M.; Platt, H. M., Sommerfield, P.
(1998) Free-living Marine Nematodes Part III –
Monhysterids. Synopses of the British Fauna (New
Series) Eds: Doris M. Kermack e R.S.K. Barnes. 53,
296p.
Cutícula levemente pontuada ou estriada.
Seis setas labiais externas de mesmo
comprimento das quatro setas cefálicas.
Cavidade bucal pequena e sem dentes.
Região anterior ao anfídio alongada.
Anfídio multiespiral, situado a mais de
duas vezes o diâmetro da cabeça em
relação à região anterior. Faringe
cilíndrica, anteriormente circunda a maior
parte da cavidade bucal e, posteriormente
alarga-se formando uma dilatação
enrugada. Cárdia pequena. Fêmeas
didélficas com ovários opostos e
estendidos. Machos com dois testículos
opostos e estendidos. Espículas curtas e
retas. Gubernáculo normalmente sem
apófise dorsal. Suplementos pré-
cloacais ausentes. Cauda alongada.
750, 1050, 1350, 1650 e 1950 metros.
Wieser, 1954
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
Animais que medem entre 1 e 1,5 mm. Na
espécie C. allometricum Wieser, 1954 o
gubernáculo apresenta uma apófise
dorsal. A região anterior alongada,
presente em Cervonema é uma
característica única para o gênero e não é
encontrada em outros indivíduos da
Família Comesomatidae.
Comentários
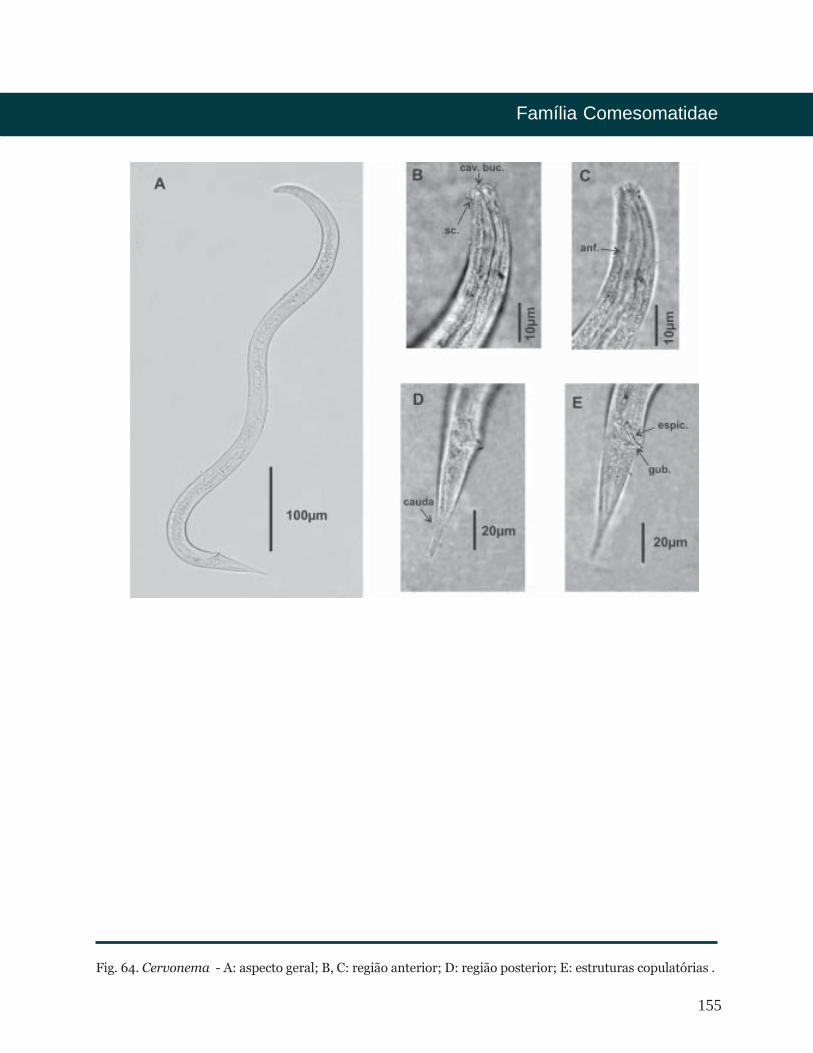
155
Fig. 64. Cervonema - A: aspecto geral; B, C: região anterior; D: região posterior; E: estruturas copulatórias .
Família Comesomatidae

156
Hopperia
Warwick, R. M. (1973) Free-living marine nema-
todes from the Indian Ocean. Bull. British Mus. Nat.
Hist., 25(3): 85–117.
Pastor de Ward, C. T. (1982) Tres especies nuevas
de nematodes marinos de vida libre
(Chromadoridae y Comesomatidae) para la Ria
Deseado, Santa Cruz, Argentina. Physis (Buenos
Aires) Secc. A, 42 (102): 39–48.
Jensen, P. (1992) Description of the marine nema-
tode Hopperia australis sp. n. (Comesomatidae) from
mangroves in Darwin, Australia, with a pictorial key
to Hopperia species. Cach. Biol. Mar. 33: 495–500.
Warwick, R. M.; Platt, H. M., Sommerfield, P. (1998)
Free-living Marine Nematodes Part III – Monhysterids.
Synopses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 53, 296p.
Cutícula pontuada com diferenciação lateral,
sem formar linhas longitudinais. Pontuações
da diferenciação lateral, ligeiramente
maiores, menos abundantes e arranjadas ao
acaso. Arranjo cefálico formado por seis
papilas labiais internas, seis papilas labiais
externas e quatro setas cefálicas em três
círculos diferentes. Margem anterior do
anfídio multiespiral no mesmo nível das
setas cefálicas. Cavidade bucal dividida em
duas regiões: 1) região anterior com três
projeções bem cuticularizadas em forma de
dentes; e 2) região posterior cilíndrica, de
paredes esclerotizadas, envolvida por
músculos protáteis longitudinais.
Raramente, seis dentes presentes.
Raramente, faringe com um bulbo distinto.
Fêmeas didélficas, com ovários
refletidos ou não. Machos com dois
testículos, opostos e estendidos. Espícula
curvada com lobos de tecido glandular.
Gubernáculo com apófise dorso-caudal.
Suplementos pré-cloacais na forma de
ductos indistintos, poros ou papilas. Cauda
longa, cônica, com a porção posterior
filiforme.
750, 1350 e 1650 metros.
Vitiello, 1969
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
H. americana Pastor di Ward, 1984 e H.
australis Jensen, 1992 apresentam ovários
estendidos e H. muscatensis Warwick,
1973 não parecem ter ovários refletidos.
Comentários
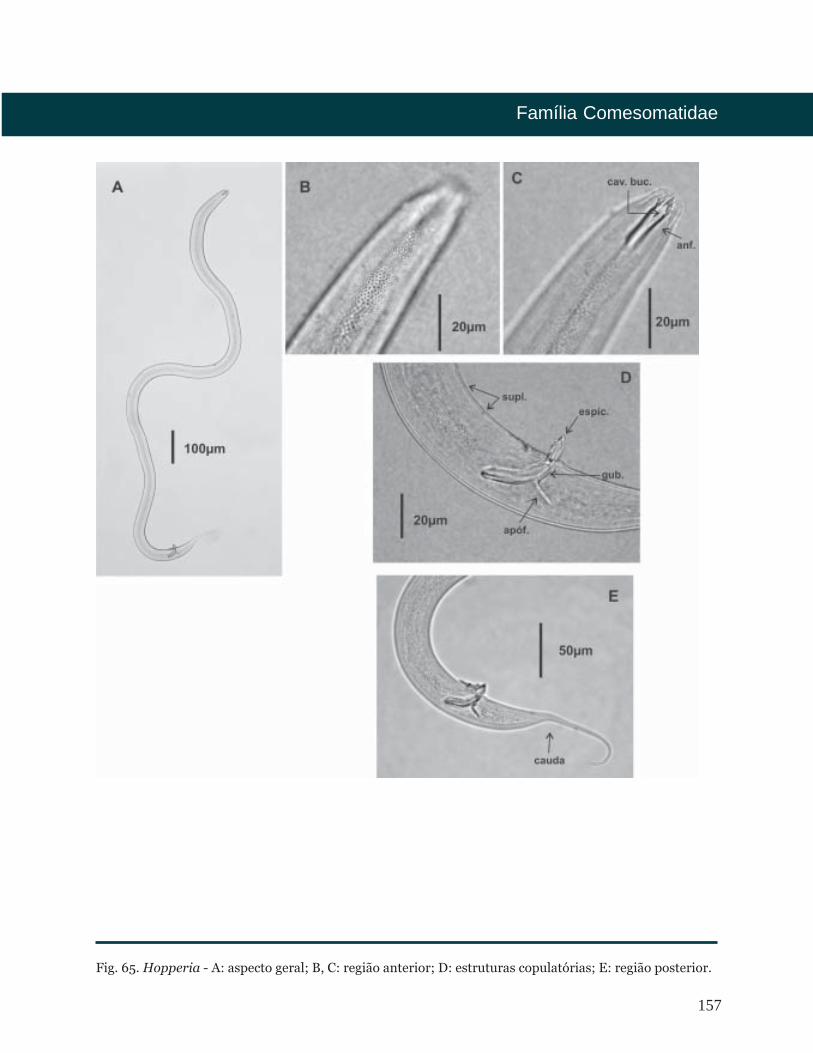
157
Fig. 65. Hopperia - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Comesomatidae

158
Laimella
Hopper, B. E. (1967) Free-living marine nematodes
from Biscayne Bay, Florida, I. Comesomatidae: the
male of Laimella longicauda Cobb, 1920, and de-
scription of Arctajania n. g. Marine Biology, vol. 1,
n. 2: 140–144.
Jensen, P. (1979) Revision of Comesomatidae
(Nematoda). Zoologica Scripta. Vol. 8, 81–105.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Cutícula levemente estriada e sem
pontuações. Arranjo das setas cefálicas em
três círculos (6+6+4); segundo e terceiro
círculos muito próximos; setas do segundo
círculo mais curtas que as cefálicas.
Cavidade bucal pequena e fracamente
cuticularizada, algumas vezes apresenta
três dentes pequenos. Anfídio
multiespiral. Cárdia pequena. Fêmeas
didélficas com ovários opostos e
estendidos. Machos com dois testículos
opostos e estendidos. Espículas curtas.
Gubernáculo com apófise. Suple-
mentos pré-cloacais papiliformes
diminutos ou ausentes. Glândulas caudais
visíveis. Cauda alongada, com a porção
anterior cônica e a posterior filiforme.
750 e 1050 metros.
Cobb, 1920
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
As glândulas caudais dos indivíduos
pertencentes a este gênero, estão restritas
a porção cônica da cauda.
Comentários
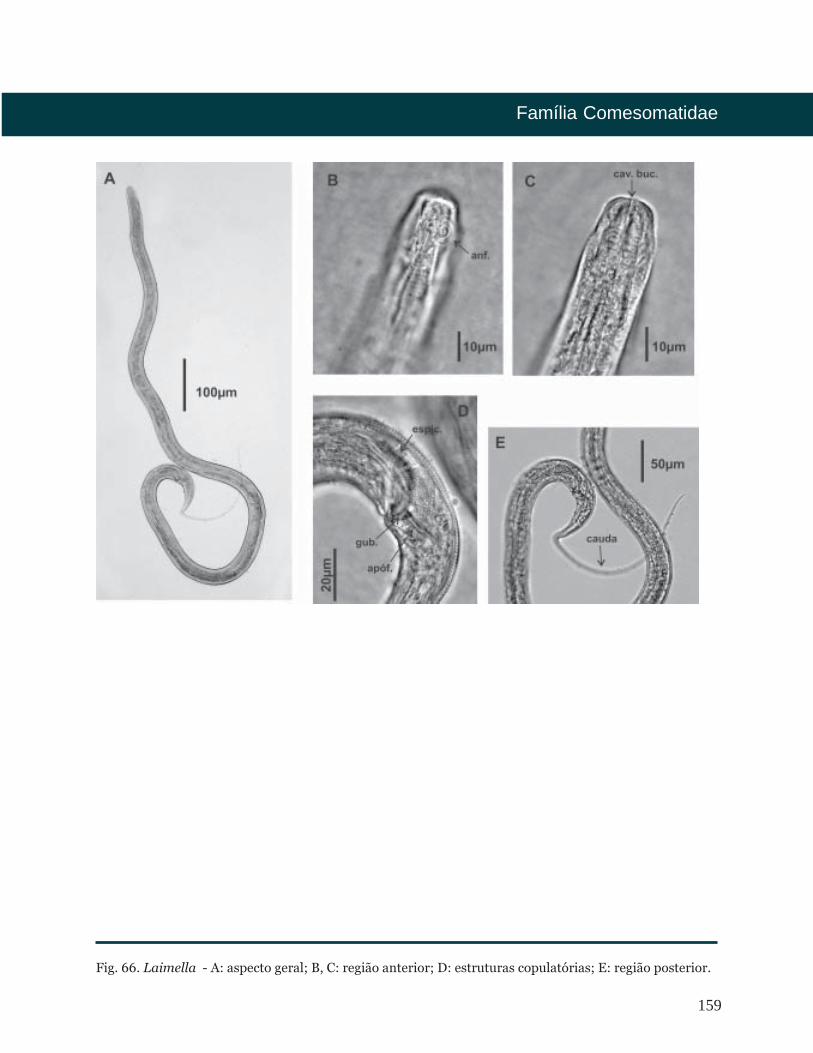
159
Fig. 66. Laimella - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Comesomatidae

160
Pierrickia
Vitiello, P., (1970) Nematodes libres marins dês
vases Profondes du golfe du lion. I I. Chromadorida.
Tethys. 2 (2): 449-500.
Gourbault, N. (1980) Nématodes abyssaux
(Campagne Walda du N/O “J Charcot”) II Espéces
et genre nouveaux de Comesomatidae. Bulletin du
Muséum National d’Histoire Naturelle, 2(A), 3
737–749.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Cutícula pontuada com diferenciação lat-
eral de pontos irregulares sem formar uma
linha longitudinal. Arranjo das setas
cefálicas em dois círculos (6+10)
compostos por setas de mesmo
comprimento. Anfídio multiespiral, pos-
terior ao círculo de setas cefálicas.
Cavidade bucal pequena. Faringe alarga-
se posteriormente sem formar um bulbo
basal. Cárdia pequena. Fêmeas
didélficas com dois ovários opostos,
pouco visíveis. Machos com dois testículos.
Espículas curtas e curvadas.
Gubernáculo com apófise dorsal.
Ausência de suplementos pré ou pós-
cloacais. Cauda cônico-cilíndrica com
setas terminais.
750, 1050, 1350, 1650 e 1950 metros.
Vitiello, 1970
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
Indivíduos podem medir de 1 a 2 mm. Este
gênero pode apresentar dimosfismo sexual
como na espécie P. vitielloi, Gourbault
(1980).
Comentários
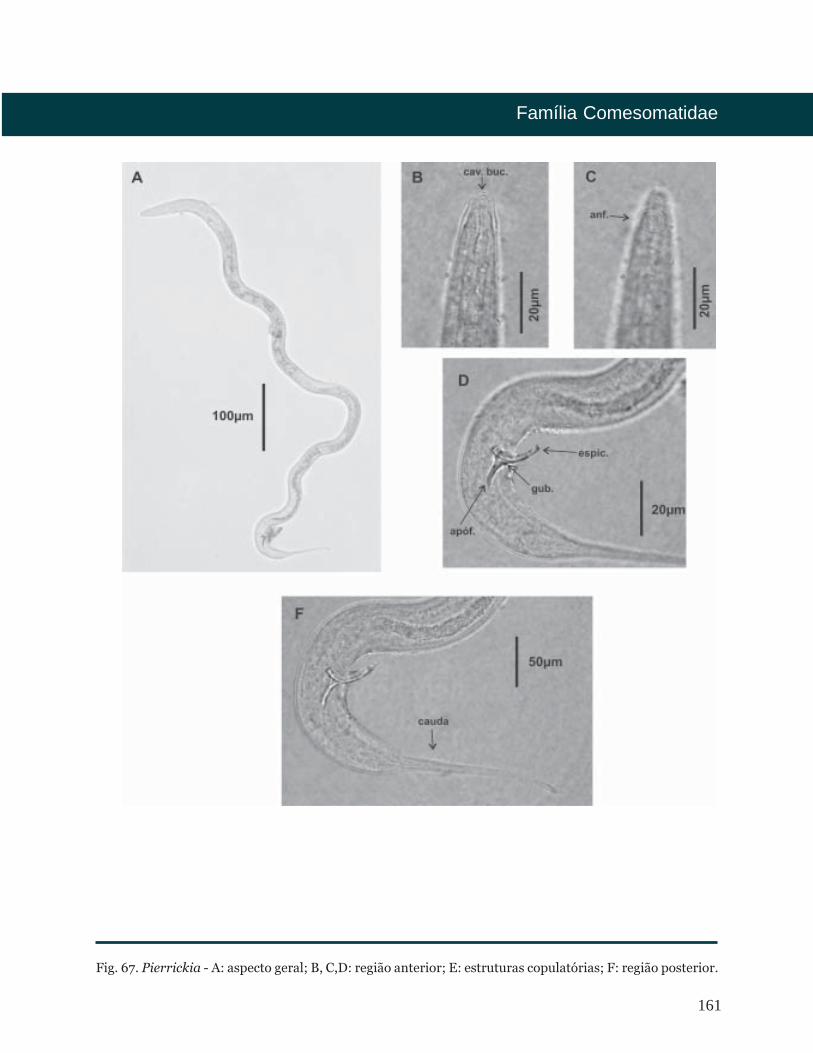
161
Fig. 67. Pierrickia - A: aspecto geral; B, C,D: região anterior; E: estruturas copulatórias; F: região posterior.
Família Comesomatidae

162
Sabatieria
Jensen, P. (1979) Revision of Comesomatidae
(Nematoda). Zoologica Scripta. Vol. 8, 81–105.
Platt, H. M. (1985) The freeliving marine nema-
todes genus Sabatieria (Nematoda:
Comesomatidae). Taxonomic revision and pictorial
keys. Zoological Journal of the Linnean Society, 83
27–78.
Warwick, R. M.; Platt, H. M., Sommerfield, P.
(1998) Free-living Marine Nematodes Part III –
Monhysterids. Synopses of the British Fauna (New
Series) Eds: Doris M. Kermack e R.S.K. Barnes. 53,
296p.
Cutícula pontuada. Diferenciação lateral
presente, porém sem fileiras longitudinais
de pontuações. Setas labiais internas nem
sempre visíveis. Setas labiais externas e
cefálicas em dois círculos separados, sendo
o segundo círculo de setas maiores que o
primeiro. Cavidade bucal em forma de
copo sem dentes. Anfídio multiespiral
localizado logo após as setas cefálicas.
Fêmeas didélficas com dois ovários
estendidos ou refletidos. Machos com
dois testículos, o posterior pode ser
refletido ou reduzido. Espícula de
comprimento e forma variável.
Gubernáculo com apófise dorsal
estreita. Suplementos pré-cloacais
usualmente presentes e geralmente
diminutos. Três glândulas caudais
presentes. Cauda cônico-cilíndrica.
750, 1050, 1350, 1650 e 1950 metros.
Rouville, 1903.
Diagnose
Bibliografia
Distribuição Batimétrica
Distribuição Geográfica
O gênero Sabatieria é dividido em cinco
sub-grupos (Platt, 1985): praedatrix,
armata, pulchra, celtica e ornata. Os
grupos foram nominados de acordo com a
espécie mais representativa dos mesmos.
Comentários
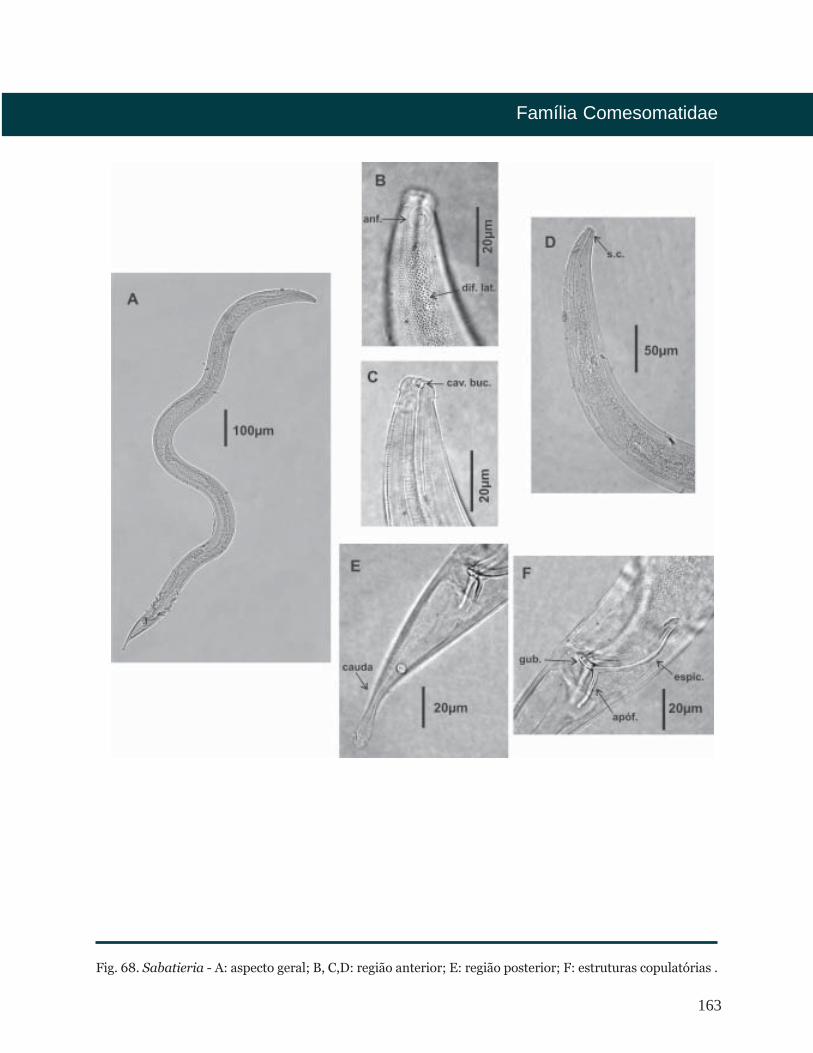
163
Fig. 68. Sabatieria - A: aspecto geral; B, C,D: região anterior; E: região posterior; F: estruturas copulatórias .
Família Comesomatidae

164
Diplopeltula
Vitiello, P. (1972) Sur quelques espèces de
Diplopeltula (Nematoda, Araeolaimida). Vie Milieu
(B) 21: 535–544.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Insti-
tuted. 167p.
Warwick, R. M.; Platt, H. M.; Sommerfield, P.
(1998) Free-living Marine Nematodes Part III –
Monhysterids. Synopses of the British Fauna (New
Series) Eds: Doris M. Kermack and R.S.K. Barnes.
53, 296p.
Cutícula estriada. Quatro setas cefálicas
sempre proeminentes, seis setas labiais
internas muito pequenas ou não
visualizadas. Anfídio em forma de “U”
invertido, sobre placa cuticularizada.
Cavidade bucal pequena ou ausente.
Quando presente, terminal ou subtermi-
nal. Dentes nunca visualizados. Ocelo
ausente. Faringe, normalmente, sem bulbo
terminal posterior. Fêmeas didélficas.
Machos com dois testículos. Suplemen-
tos pré-cloacais ausentes. Cauda cônica.
750, 1050, 1350, 1650 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Gerlach, 1950
Indivíduos que medem de 1 a 2 mm. A
faringe da maioria das espécies não forma
um bulbo verdadeiro, porém em D.
bulbosa Vitiello, 1972 o bulbo terminal é
bem desenvolvido.
Comentários
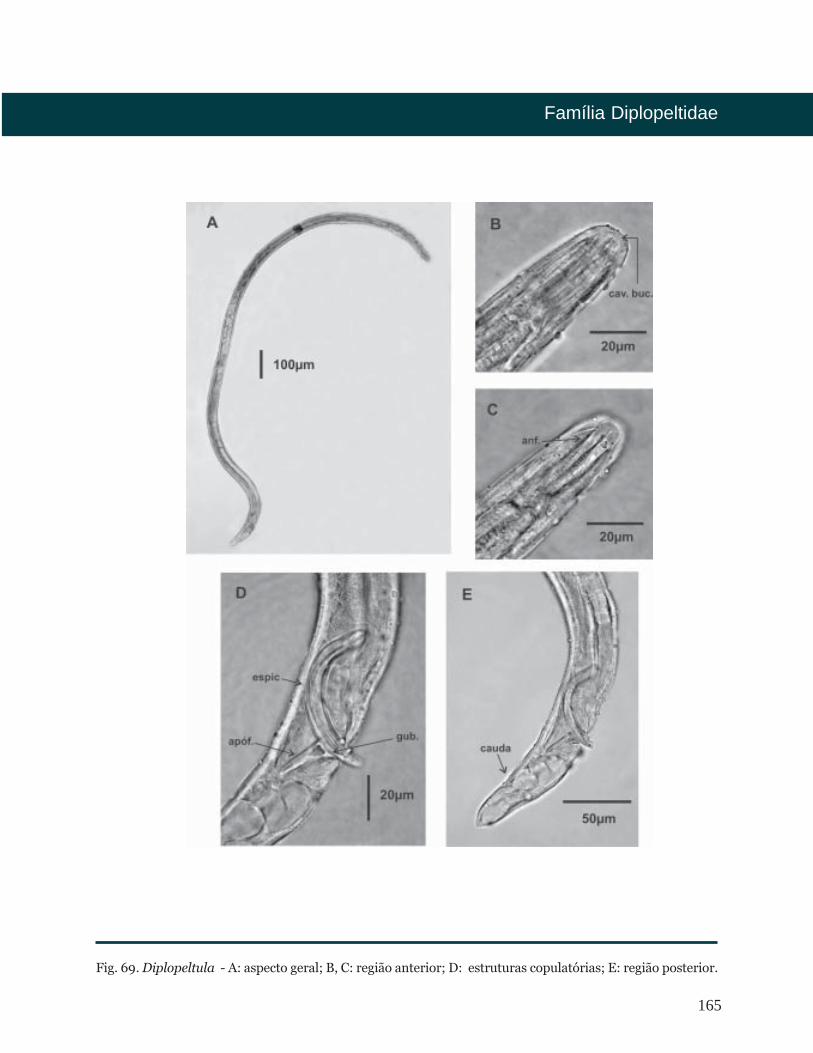
165
Fig. 69. Diplopeltula - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Diplopeltidae

OR
DE
M P
LE
CT
IDA
MALAKHOV
, 19
82
A Ordem Plectida foi criada para incluir famílias que estavam na Ordem
Araeolaimida Andrássy, 1976 e Chromadorida Lorenzen, 1981. As
características dos representantes desta Ordem são: cutícula anelada,
geralmente com poros, glândulas e setas somáticas; diferenciação lateral
presente ou ausente; sensilas labiais internas e externas usualmente
papiliformes; quatro cefálicas setiformes; glândula ventral e o poro
excretor usualmente presentes; boca cilíndrica; fêmeas didélficas,
algumas com ovários refletidos; machos com dois testículos opostos
ou estendidos; espículas simétricas; gubernáculo quase sempre
presente; cauda pode estar reduzida em alguns Gêneros.
Neste grupo estão incluídos Nematoda aquáticos (marinhos, estuarinos
e de água doce). Segundo Holovachov, (2004) e Holovachov & Bostrom,
(2004), a Ordem Plectida é composta pelas Superfamílias Leptolaimoidea,
Plectoidea, Camacolaimoidea, Ceramonematoidea e Haliplectoidea. São
Famílias pertencentes a esta Ordem: Ohridiidae, Leptolaimidae,
Aphanolaimidae, Chronogastridae, Plectidae e Metateratocephalidae
(Figura 9). De ley et al, (2006) adicionaram as famílias: Rhadinematidae,
Aegialoalaimidae, Diplopeltoididae, Paramicrolaimidae, Odontolaimidae,
Tarvaiidae, Ceramonematidae, Tubolaimoididae, Peresianidae,
Haliplectidae e Aulolaimidae à ordem.
CARACTERÍSTICAS GERAIS
Figura 70: Porção anterior do corpo de gêneros representantes das Famílias de Plectida(
Holovachov & De Ley, 2006) que ocorrem na Bacia de Campos: A= Leptolaimidae; B=
Ceramonematidae; C= Aegialoalaimidae, D= Diplopeltoididae, E= Paramicrolaimidae, F=
Tarvaiidae, G= Haliplectidae.

168
Antomicron
Cobb, N. A. (1920) One hundred new nemas (type
species of 100 new genera). Contribution to a sci-
ence of Nematology (Baltimore). 9: 217–343.
Lorenzen, S. (1966) Diagnosen einiger freilebender
Nematoden von der schleswigholsteinischen
Westküste. Veröff. Inst. Meeresforsch. Bremerh.
10: 31–48.
Vitiello, P. (1971) Espèces nouvelles de
Leptolaimidae (Nematoda) et description Du genre
Leptolaimoides n. gen. Cah. Biol. Mar. XII: 419–432.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Insti-
tuted. 167p.
Warwick, R. M.; Platt, H. M., Sommerfield, P. (1998)
Free-living Marine Nematodes Part III – Monhysterids.
Synopses of the British Fauna (New Series) Eds: Doris
M. Kermack e R.S.K. Barnes. 53, 296p.
Cutícula fortemente estriada, estriações
espaçadas. Extremidade anterior do corpo
sem anelações. Arranjo cefálico com seis
sensilas labiais anteriores, geralmente
diminutas, e quatro setas cefálicas
posteriores. Anfídio em forma de um loop
alongado. Cavidade bucal diminuta não
cuticularizada. Faringe com um bulbo
posterior. Fêmeas didélficas. Machos
com um ou dois testículos. Espículas
curvadas. Gubernáculo simples.
Possuem de 3 a 10 suplementos pré-
cloacais tubulares, fortemente
esclerotizados. Três glândulas caudais.
Cauda cônico-cilíndrica.
750 e 1350 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Örley, 1880
Indivíduos que podem medir de 0,5 a 1,5
mm. Neste gênero, os suplementos pré-
cloacais podem estar associados a
glândulas e podem se estender até a região
do bulbo terminal da faringe.
Comentários
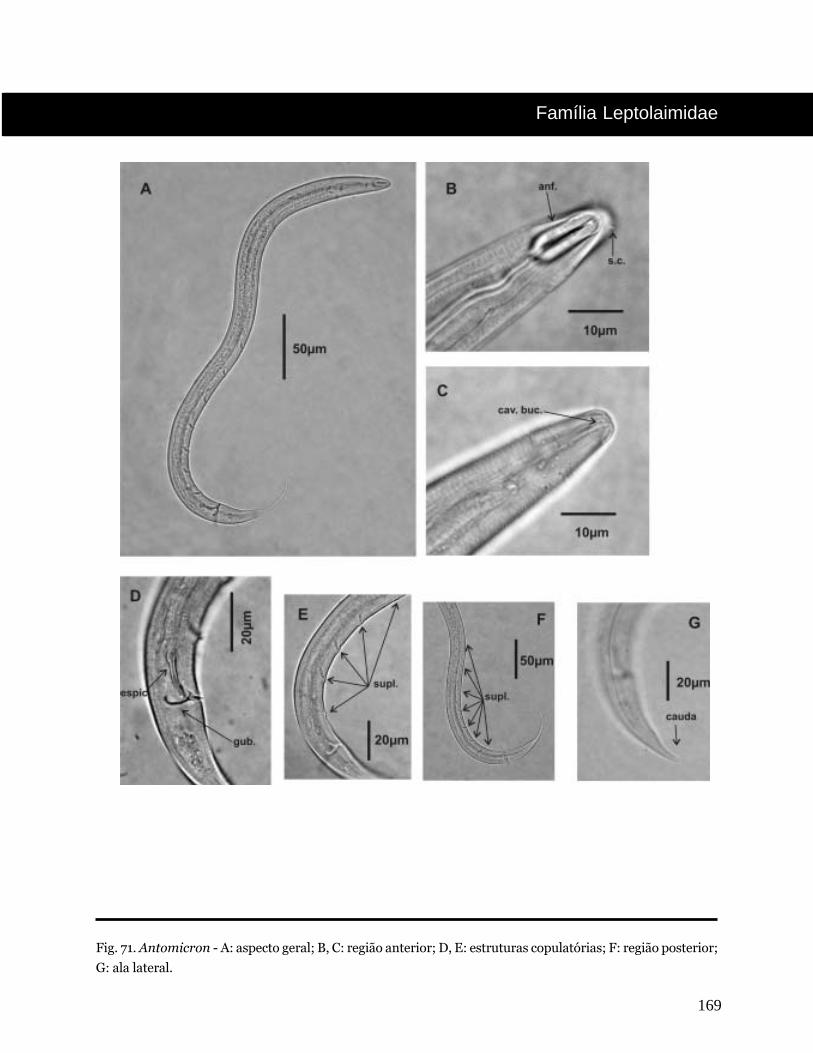
169
Fig. 71. Antomicron - A: aspecto geral; B, C: região anterior; D, E: estruturas copulatórias; F: região posterior;
G: ala lateral.
Família Leptolaimidae

170
Camacolaimus
De Coninck, L. A.; Schuurmans Stekhoven. (1933)
The freeliving marine nemas of the Belgian coast.
II Mém. Mus. R. Hist. nat. Belg. 58: 1-163.
Wieser, W.; Hopper, B. (1967) Marine nematodes
of the east coast of North America. I. Florida. Bull.
Mus. comp. Zool. Harv. 135 (5): 239-344.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Warwick, R. M.; Platt, H. M., Sommerfield, P.
(1998) Free-living Marine Nematodes Part III –
Monhysterids. Synopses of the British Fauna (New
Series) Eds: Doris M. Kermack e R.S.K. Barnes. 53,
296p.
Cutícula fortemente estriada. Estriações
começam logo após as setas cefálicas e se
estendem até antes do espinerete. Linhas
longitudinais laterais presentes. Lábios
indistintos com seis papilas cônicas.
Quatro setas cefálicas curtas. Cavidade
bucal com um longo dente dorsal em
forma de estilete. Anfídio espiral.
Sistema excretor-secretor visível. Fêmeas
com didélficas, raramente monodélfica.
Machos com dois testículos. Espícula
delgada e levemente curvada.
Gubernáculo pequeno. Suplementos
pré-cloacais ausentes. Suplementos
pós-cloacais, formados por setas
grossas. Três glândulas caudais. Cauda
cônica com espinarete terminal.
750, 1050, 1350 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
De Man, 1889
Indivíduos que medem de 1 a 3 mm. Nas
descrições é relatado que a cavidade bucal
é composta de um longo dente dorsal em
forma de estilete, no entanto alguns
autores afirmam que não é um dente de
fácil visualização.
Comentários

171
Família Leptolaimidae
Fig. 73. Camacolaimus - A: aspecto geral; B: região anterior; C: estruturas copulatórias; D: região posterior.

172
Diplopeltoides
Jensen, P. (1991) Nine new and less known nema-
tode species from the deep-sea benthos of the Nor-
wegian Sea. Hydrobiologia, 222: 57–76.
Lorenzen, S. (1994) The phylogenetic systematics
of freeliving Nematodes. The Ray Society Instituted.
167p.
Warwick, R. M.; Platt, H. M., Sommerfield, P.
(1998) Free-living Marine Nematodes Part III –
Monhysterids. Synopses of the British Fauna (New
Series) Eds: Doris M. Kermack and R.S.K. Barnes.
53, 296p.
Cutícula estriada. Quatro setas cefálicas.
Cavidade bucal ausente. Anfídio em loop.
Faringe com isthmus estreito, um bulbo
pouco desenvolvido na porção anterior e
um bulbo bem desenvolvido
posteriormente. Fêmeas didélficas com
ovários opostos. Machos com dois
testículos. Espículas curtas, curvadas,
finas e com a porção proximal
arredondada. Gubernáculo pequeno e
sem apófise. Suplementos pré-
cloacais ausentes. Três glândulas caudais.
Cauda cônica.
750, 1050, 1350, 1650 e 1950 metros.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Gerlach, 1962
Indivíduos que medem cerca de 0,5 mm.
Em algumas espécies é visualizada uma
esclerotização na região anterior lateral
ao anfídio como ocorre em D. linkei Jensen
(1991).
Comentários

173
Fig. 73. Diplopeltoides - A: aspecto geral; B: região anterior; C: estruturas copulatórias e região posterior.
Família Diplopeltoididae

174
Ceramonema
Furstenberg, J. P.; Vincx, M. (1993) Five new spe-
cies of the family Ceramonematidae (Nematoda:
Order Chromadorida) from South Africa.
Nematologia 39: 142–145.
Tchesunov, A. V.; Miljutina, M. A. (2002) A rewiew
of the family Ceramonematidae (marine free-living
nematodes), with descriptions of nine species from
the White Sea. Zoosyst. Rossica vol. II. Zoological
Institute. St. Petersburg.
Cutícula consiste de 70 a 320 espaçadas e
espessas anelações iguais ou desiguais.
Quando desiguais, a largura aumenta
gradualmente da primeira anelação
subcefálica até perto da cárdia, então a
largura da anelação diminuiu e, novamente
volta a aumentar gradualmente até a anelação
anal. Vacualização intracuticular pode estar
presente. Zigapófises anelares presentes ou
ausentes. Região labial não destacada. Setas
labiais externas e cefálicas distribuídas em
dois círculos separados. Anfídio em forma
de loop, alongado e arredondado. Fêmeas
didélficas, com ovários refletidos. Ma-
chos com dois testículos opostos. O testículo
anterior estendido e o posterior refletido,
dispostos de ambos os lados do intestino. O
testículo anterior é maior que o posterior.
Espículas curtas. Gubernáculo simples.
Cauda cônica com anelações que se estreitam
gradualmente até a porção terminal.
Diagnose
Distribuição Batimétrica
Distribuição Geográfica
Bibliografia
Cobb, 1920
Indivíduos que medem de 0,5 a 2 mm. De
acordo com Tchesunov & Miljutina (2002),
várias descrições das espécies não incluíam
detalhamento sobre os órgãos internos dos
ceramonematídeos devido à complexidade
e a refração cuticular quando observado
em microscópio óptico que dificulta a
visualização.
Comentários
750 e 1050 metros.
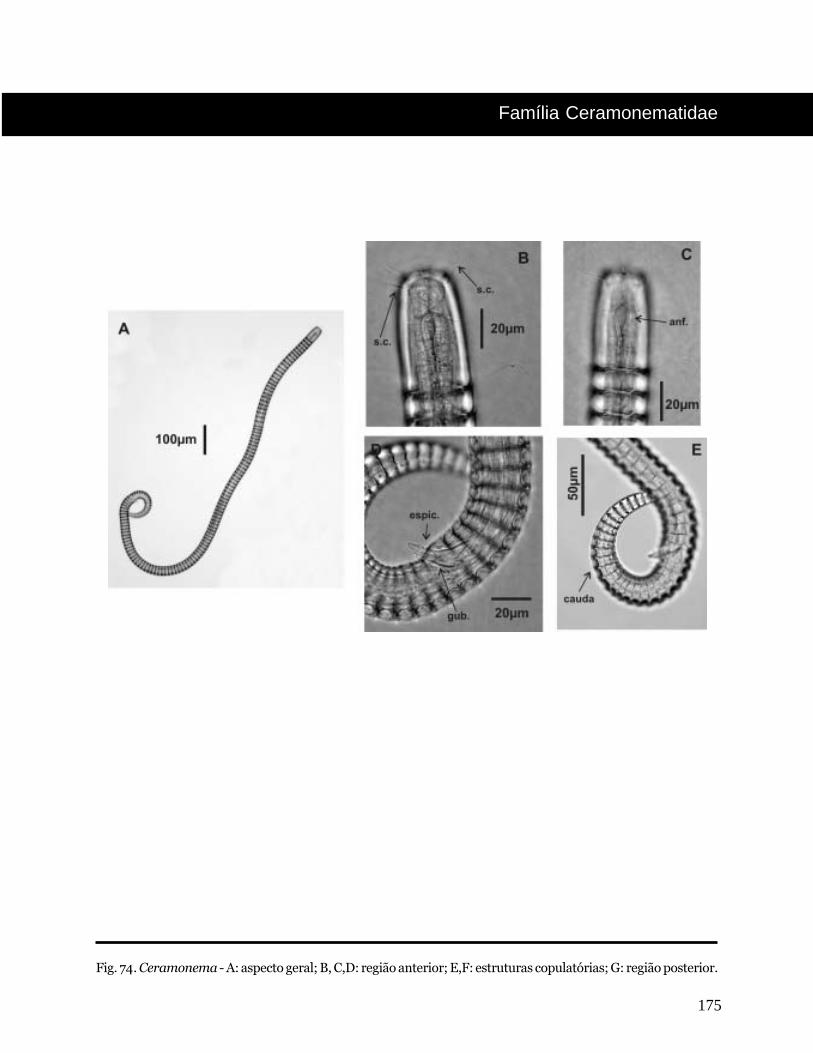
175
Fig. 74. Ceramonema - A: aspecto geral; B, C,D: região anterior; E,F: estruturas copulatórias; G: região posterior.
Família Ceramonematidae

176
Metadasynemella
De Coninck, L. A. P. (1942) Sur quelques espèces
de Nématodes libres (Ceramonematidae Cobb,
1933), avec quelques remarques de systématique.
Bull. Mus. r. Hist. nat. Belg. 18 (22): 1–37.
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Tchesunov, A. V.; Miljutina, M. A. 2002. A rewiew
of the family Ceramonematidae (marine free-living
nematodes), with descriptions of nine species from
the White Sea. Zoosyst. Rossica vol. II. Zoological
Institute. St. Petersburg.
Corpo curto, raramente alongado.
Anelações cuticulares em número variável
(até 175), espaçadas desiguais ao longo do
corpo. Zigapófises ausentes ou
pobremente desenvolvidas. Vacualização
intracuticular presente ou ausente.
Região labial destacada da cápsula
cefálica. Cápsula cefálica usualmente
com cristas transversais e longitudinais.
Todas as setas inseridas na região labial.
Setas labiais externas e cefálicas dispostas
em dois círculos muito próximos ou unidas
em um único círculo. Anfídio em loop.
Fêmeas didélficas, com ovários
refletidos. Machos com dois testículos
opostos. O testículo anterior estendido e o
posterior refletido, dispostos de ambos os
lados do intestino. O testículo anterior é
maior que o posterior. Espícula curta.
Gubernáculo simples e levemente
curvado. Cauda cônica.
750, 1050, 1350, 1650 e 1950 metros.
De Coninck, 1942
Bibliografia
Distribuição Batimétrica
Diagnose Distribuição Geográfica
Indivíduos que medem cerca de 0,5 mm.
De acordo com Tchesunov & Miljutina
(2002), os órgãos internos de deste gênero
são pobremente descritos devido à
complexidade cuticular.
Comentários
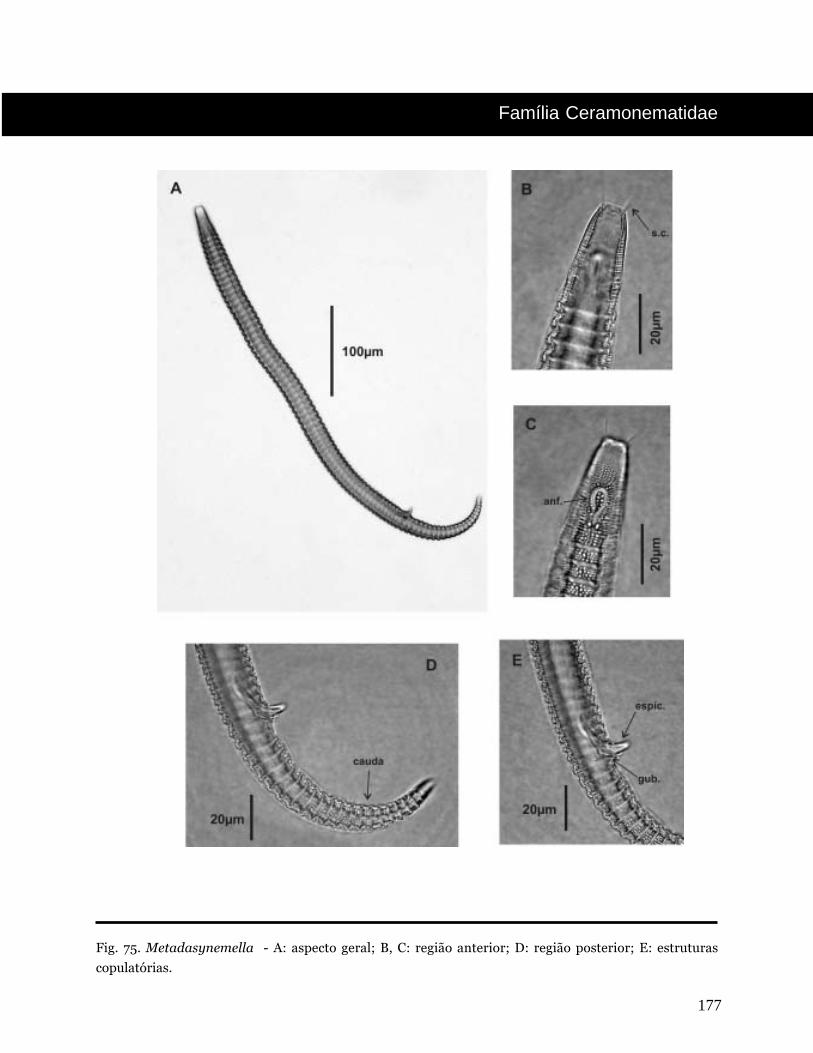
177
Fig. 75. Metadasynemella - A: aspecto geral; B, C: região anterior; D: região posterior; E: estruturas
copulatórias.
Família Ceramonematidae

178
Pselionema
Platt, H. M.; Warwick, R. M. (1988) Free-living
marine Nematodes – Part II Chromadorids. Syn-
opses of the British Fauna (New Series). Ed. Doris
m. Kermarck and R. S. K. Barnes. 38, 502p.
Tchesunov, A. V.; Miljutina, M. A. 2002. A rewiew
of the family Ceramonematidae (marine free-living
nematodes), with descriptions of nine species from
the White Sea. Zoosyst. Rossica vol. II. Zoological
Institute. St. Petersburg.
Cutícula consistindo de 70 a 210 anelações
espessas e espaçaadas. Zigapófises mais
ou menos desenvolvidas. Região labial
não destacada. Quatro setas cefálicas
presentes. Anfídio em loop alongado.
Cavidade bucal ausente. Fêmeas
didélficas, com ovários refletidos.
Machos com dois testículos opostos. O
testículo anterior estendido e o posterior
recurvado, dispostos de ambos os lados
do intestino. O testículo anterior é maior
que o posterior. Espículas e
gubernáculo curtos. Cauda cônica.
750, 1050, 1350, 1650 e 1950 metros.
Cobb, 1933
Diagnose Distribuição Geográfica
Distribuição Batimétrica
Bibliografia
Indivíduos componentes deste gênero
medem de 0,4 a 1,5 mm. Assim como em
Ceramonema e Metadasynemella, este
gênero também não possui descrições
detalhadas relacionadas aos órgãos
internos (Tchesunov & Miljutina, 2002).
Comentários
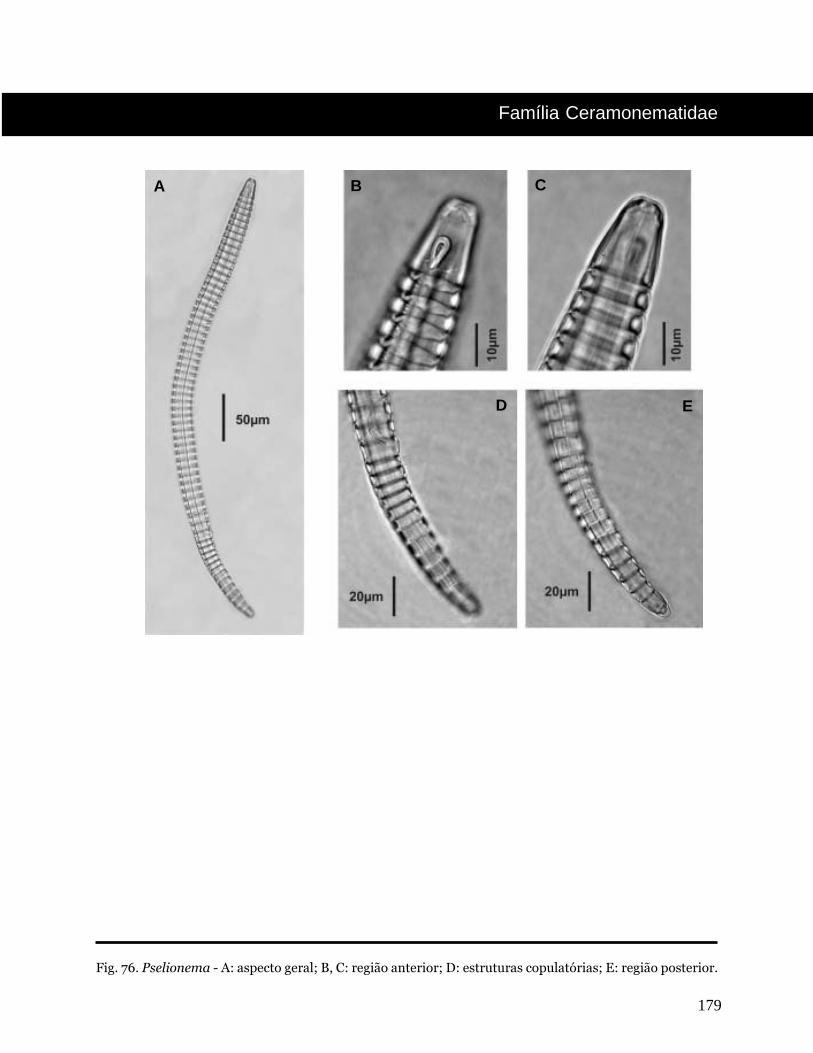
179
Fig. 76. Pselionema - A: aspecto geral; B, C: região anterior; D: estruturas copulatórias; E: região posterior.
Família Ceramonematidae
A B C
D E

181
Abreviações
Anf. = anfídio
apóf .= apófise
bulb. = bulbo da faringe
cauda = cauda
cáp. cef. = cápsula cefálica
cav. buc.= cavidade bucal
cut. = cutícula
d. = dente
dif. lat. = diferenciação lateral
esp.= espinhos
espíc. = espícula
gub. = gubernáculo
mand.= mandíbula
onc.= onchia
pap.= papila
s.= seta
s.c. = Seta cefálica
s.cv. = Seta cervical
s.sb. = seta subcefálica
supl. = suplemento

183
Glossário
Anel nervoso: anel de tecido nervoso que circunda a faringe.
Anfídio: órgãos sensoriais em par, situados na lateral.
Anfídio criptoespiral: anfidio com abertura circular e espiral levemente visivel.
Apófise: uma porção separada de uma estrutura principal, principalmente do gubernáculo.
Cápsula caudal: linha espessa de origem cuticular presente na região terminal da cauda.
Cápsula cefálica: modificação da camada interna da cutícula no ponto em que a porção
anterior da faringe adere-se a parede do corpo. Em geral, é fortemente esclerotizada.
Capitulum: extremidade anterior da esp’icula de forma arrendondada.
Cárdia: estrutura muscular presente na porção posterior da faringe conectando a região
anterior do intestino.
Cheilostoma: cavidade labial da boca, delimitada anteriormente pela abertura da cavidade
oral e posteriormente pelo protostoma.
Corpus gelatum: substância gelatinosa presente na cavidade do anfídio.
Cuticularizado: derivado da cutícula, mas pode ser usado de forma geral para qualquer
estrutura que pareça ser rígida ou quando é relativamente reforçado pelo tecido que o
circunda.
Desmens: anéis formados por secreções e partículas exteriores, como grãos de areia, por
exemplo, podendo estar associados a fileiras de poros ou espinhos ou separados por
interzonas, sem ornamentações de espinhos ou poros.
Espícula: órgão copulatório, usualmente em par, presente na cloaca dos machos.
Espinerete: estrutura de forma pontiaguda presente na porção terminal da cauda, serve
como um plug controlando a excressao das glândulas caudais pelo poro.

184
Estriação semi-lunar: estriações presentes na região labial principalmente em alguns
Thoracostomopsídeos.
Fovea anfideal: cavidade presente no anfídio.
Gubernáculo: peça cuticularizada situada na regiao dorsal da espícula ou envolvendo a
extremidade posterior.
Isthmus: Uma secção estreita da faringe.
Interzonas cuticulares: claras anelações entre os anéis (desmens).
Metanemes: receptores filamentosos que se estendem lateralmente na região epidermal.
Ocelo: estrutura sensitiva pigmentada, posicionada geralmente na região do pescoço.
Ovários antidromous: acontece quando a área da porção germinativa e a da região de
crescimento do ovário estão curvadas.
Ovários prodélficos: os dois ovários são direcionados anteriormente.
Ovários refletidos: ovários curvados nas extremidades.
Placas periorais: placas que circundam a abertura da cavidade bucal.
Protostoma: é a região mediana da cavidade bucal, é delimitada anteriormente pelo
cheilostoma e posteriormente pelo telostoma.
Sistema Reprodutor Anfidélfico: sistema reprodutor com dois ovários opostos.
Sistema Reprodutor Didélfico: sistema reprodutor com dois ovários.
Sistema demaniano: sistema de vasos que conectam o intestino ao útero e posteriormente
com o ambiente exterior.
Sistema Reprodutor Monodélfico: sistema com apenas um único ovário.
Suplementos pré-cloacais: nos machos, órgãos de secreção e de conexão ou junção na
cópula, que estão em posição ventromediana e derivados da cutícula.

185
Telamon: peças acessórias localizadas lateralmente à espícula.
Telostoma: uma válvula curta que conecta a boca a faringe.
Tricomóide: padrão de distribuição das setas somáticas em que as setas sub-ventrais,
usualmente mais numerosas que as sub-dorsais podem estar em par ou não, além do anel
final não possuir setas.
Vacualização intracuticular: diminutos vacúolos ou vesículas que podem estar circundando
toda a anelação cuticular, ou podem estar concentrados em pequenas áreas situadas abaixo
das pontes longitudinais presentes na cutícula de integrantes da Família Ceramonematidae.
Velum: delicada membrana cuticular no lado interno da espícula.
Zigapófises: delgados picos ou montes que se sobrepõem as anelações cuticulares presentes
na Família Ceramonematidae.

186

187
REFERÊNCIAS
Aguinaldo, A.M; Turbeville, J.M.; Linford, L.S.; Rivera, M.C.; Garey, J.R.; Raff, R.A. &
Lake, J.A. 1997. Evidence for a clade of nematodes, arthropods and other moulting ani-
mals. Nature, 387 : 489–493.
Allgén, C.A. 1932. Weitere Beiträge zur Kenntnis der marinen Nematodenfauna der
Campbellinsel. Nyt Mag. Naturvid, 70: 97-198.
Allgén, C. A. 1942 Die freilebenden Nematoden des Mittelmeeres Zool. Jb. (Systematik)
Band 76, Heft 1/2: 1-102.
Allgén, C. A. 1952 Über einige Desmoscoleciden aus den schwedischen Südpolar-Expedi-
tion 1901 bis 1903. Zoologischer Anzeiger Bd. 148, Heft 3/ 4.
Bastian, H. C. 1865 Monograph of the Anguillulidae, or Free Nematoids, Marine, Land,
and Freshwater; with Descriptions of 100 New Species. The Transactions of the Linnean
Society of London. Volume XXV. Part II: 73-184
Blaxter, M.L. 2001. Molecular analysis of nematode evolution. Chapter 1, pp. 1-24. In:
M.W. Kennedy & W. Harnett (eds) Parasitic nematodes – molecular biology, biochemis-
try and immunology. CABI Publishing, 486 pp.
Blome, D. & Schrage, M. 1985. Freilebende Nematodes aus der Antarktis.
Veröffentlichungen des Instituts für Meeresforschung in Bremerhaver, 21: 71-96
Boisseau, J. P. 1957. Technique pour l’étude quantitative de la faune interstitielle des sables.
In: R. P. Higgiens & H. Thiel 1988. Introduction to the study of meiofauna. Washington:
D.C. Smithsonian Institution Press, 488 pp.
Bouwman, L. A. 1983. Systematic, ecology and feeding biology of estuarine nematodes.
Tese de Doutorado, Wageningen Universiteit, Holanda, 173 pp.
Brusca, R.C. & Brusca, G.J. 2003. Inveterbrates. 2º ed.. Sinauer Associates, Inc. 895 pp.
Cattrijsse, A. & Vincx, M. 2001. Biodiversity of the benthos and the avifauna of the Bel-
gian coastal waters: summary of data collected between 1970 and 1998. Sustainable
Management of the North Sea. Federal Office for Scientific, Technical and Cultural Af-
fairs: Brussel, Belgium. 48 pp

188
Cobb, N. A. 1914. Antarctic marine free-living nematodes of the Shackleton expedition
Contributions to Science of Nematology, 1: 1-33.
Cobb, N. A. 1917. Notes on nemas. Contributions to Science of Nematology. 5: 117-128.
Coomans, A.1989. The freeliving nematofauna of Belgium: a review [Overzicht van de
vrijlevende nematofauna van België (Nematoda)], In: K. Wouters & L.Baert (ed.) Pro-
ceedings of the Symposium “Invertebrates of Belgium”. pp. 43-56
Dayrat, B. 2005. Towards integrative taxonomy. Biological Journal of the Linnean Soci-
ety 85: 407–415.
De Coninck, L.A.P. 1942. Sur quelques espèces nouvelles de nématodes libres
(Ceramonematinae Cobb, 1933), avec quelques remarques de systématique. Bulletin du
Musée Royal d’Histoire Naturalle de Belgique, 18(22): 1-37.
De Grisse, A. T. 1969. Redescription ou modification de quelques techniques utilisés dans
l’ étude des nématodes phytoparasitaires. Mededelingen Rijksfakulteit
Landbouwwetenschappen Gent. 34: 351-369.
De Ley, P. & Blaxter, M.L. 2002. Systematic position and filogeny. Chapter 1, pp 1-30. In:
D.L. Lee (ed.) The biology of nematodes. Taylor and Francis, London,
De Ley, P.; Decraemer, W. & Abebe, E. 2006. Introduction: Summary of Present Knowl-
edge and Research Addressing the Ecology and Taxonomy of Freshwater Nematodes. Chap-
ter 1, pp. 3-30. In: E. Abebe; W. Traunspurger & I. Andrássy (eds) Freshwater Nema-
todes: Ecology and Taxonomy. CABI Publishing, UK. 576 pp.
De Man, J. G. 1890 Quatrième note sur les nématodes libres de la mer du Nord et de la
Manche. Memoires Societe Zoologique de France, 3: 169-194.
De Man, J. G. 1922 Neue freilebende Nematoden aus der Zuidersee. Tijdschr.
ned.dierk.Vereen., 18: 124-134.
Defeo, R. 2002. Bacia de Campos, apenas 26 anos ou mais? (www.clickmacae.com.br -
acessado em 05/10/2007).
Dinet, A. & Vivier, M. H. 1979. Le meiobenthos abyssal du Golfe de Gascogne. II. Les
peuplements de Nematodes et leur divesité specifique. Cahiers de Biologie Marine, 20:
109-123.

189
Filipjev, I. N. 1918 Free-living marine nematodes of the Sevastopol area Transactions of
the Zoological Laboratory and the Sevastopol Biological Station of the Russian Academy
of Sciences Series II No 4 (Issue I & II) (Translated from Russian).
Filipjev, I., 1922. Encore sur les Nematodes libres de la Mer Noire. Trudy Stravropol. Sel.
Khoz. Inst. 1: 83–184
Fonseca, G.; Derycke, S. & Moens, T. 2008. Integrative taxonomy in two free-living Nema-
tode species complexes. Biological Journal of the Linnean Society, 94: 737-753.
Gerlach, S. A. & Riemann, F. 1973. The Bremerhaven Checklist of Aquatic Nematodes, A
Catalog of Nematoda Adenophorea Exclunding the Dorylaimida. Veröffentlichungen des
Instituts für Meeresforschung in Bremerhaver. Kommissionsverlag Franz Leuwer, Bremen.
Suppl. 4, Heft 1.
Gerlach, S. A. & Riemann, F. 1974. The Bremerhaven Checklist of Aquatic Nematodes, A
Catalog of Nematoda Adenophorea Exclunding the Dorylaimida. Veröffentlichungen des
Instituts für Meeresforschung in Bremerhaver. Kommissionsverlag Franz Leuwer, Bremen.
Suppl. 4, Heft 2.
Gerlach, S. A. 1952 Nematoden aus dem Küstengrundwasser Abh. math.-naturw. Kl. Akad.
Wiss. Mainz, 6: 315-372.
Greeff, R. 1869 Untersuchungen über einige merkwürdige Thiergruppen des Arthropoden-
und Wurm-Typus. Arch. f. Naturgesch, 35: 71-121.
Heip, C.; Vincx, M. & Vranken, G. 1985. The ecology of marine nematodes. Oceanography
and Marine Biology Annual Review, 23: 399-489.
Heip, C.; Vincx, M.; Smol, N. & Vranken, G. 1982. The systematics and ecology of free-
living marine nematodes. Helminthological Abstracts Series B, Plant Nematology. 51 (1):
1-31.
Herman, P. M. J. & Heip, C. 198. On the use of Meiofauna in ecological monitoring: Who
needs taxonomy? Marine Pollution Bulletin, 19 (12): 665-668.
Huston, M. A. 1994. Biological diversity: The Coexistence of Species on Changing Land-
scape. Cambridge University Press, Cambridge, 708 pp.
Hyman. L.H. 1951. Acanthocephala, Aschelminthes, and Entoprocta. pp. 151–170. In: The
Invertebrates. vol. 3, McGraw-Hill, New York.

190
Jensen, P. 1988. Nematoda assemblages in the deep-sea benthos of Norwegian Sea. Deep-
Sea Research, 35 (7): 1173-1184.
Lorenzen, S. 1969 Desmoscoleciden (eine Gruppe freilebender Meeresnematoden) aus
Küstensalzwiesen. Veröffentlichungen des Instituts für Meeresforschung in Bremerhaver,
12: 231-265.
Lorenzen, S. 1994. The phylogenetic systematics of free-living nematodes. The Ray Soci-
ety Institute, London, 755 pp.
Mawson, P. M. 1958. Free-living nematodes. Section 3: Enoploidea from subantarctic sta-
tions B.A.N.Z. Antarctic Research Expedition Reports, 6 (14): 307-358.
Meldal, B.H.M.; Debenham, N.J.; De Ley, P.; Tandingan De Lay, I.; Vanfleteren, J.R.;
Vierstraete, A.R.; Bert, W.; Borgonie, G.; Moens, T.; Tyler, P.A.; Austen, M.C.; Blaxter,
M.L.; Rogers, A.D. & Lambshead, P.J.D. 2006. An improved molecular phylogeny of the
Nematoda with special emphasis on marine taxa. Molecular Phylogenetics Evolution, 42:
622–636.
Metzler, P. M.; Glibert, P. M.; Gaeta, S. & Luddam, J. M. 1997. New and regendrated pro-
duction in the South Atlantic off Brazil. Pergamon, Deep-Sea Research I, 44 (3): 363-384.
Müller, Y. 2004. Faune et flore du littoral du Nord, du Pas-de-Calais et de la Belgique:
inventaire. [Coastal fauna and flora of the Nord, Pas-de-Calais and Belgium: inventory].
Commission Régionale de Biologie Région Nord Pas-de-Calais: France. 307 pp.
Netto, Q. A.; Galluci, F. & Fonseca, G. F. C. 2005 Meiofaunal communites of continental
slope and deep-sea sites off SE Brazil. Deep - Sea Research I, 52: 845-099.
Paladian, G. & Andriescu, I. 1963 Contributions à l’étude des Desmoscolecidae (Nema-
toda) des eaux roumaines de la Mer Noire. Trav. Mus. Hist. nat. Gr. Antipa 4: 167-173.
Panceri, P. 1876. Osservazioni intorno a nuove forme di vermi nematodi marini. Atti R.
Accad. Scienze fis. mat., 7 (10): 1-10.
Pastor C.T. 1979. Free living marine nematodos (Subclass Adenophorea) of the ría Deseado
(Santa Cruz, Argentina). Systematic Contributions I. Annales De La Societe Royale
Zoologique De Belgique. 1 2: 29 45.
Pastor de Ward, C.T. 1984. Nematodes marinos de la Ria Deseado (Leptolaimina:
Leptolaimidae, Haliplectidae) Santa Cruz, Argentina. Physis, Secc. A, 42(103): 87 92.

191
Pastor de Ward, C.T. 1985. Free living marine nematodes of the Deseado river estuary
(Chromadoroidea: Chromadoridae, Ethmolaimidae, Cyatholaimidae and Choniolaimidae)
Santa Cruz, Argentina. Especial del Centro Nacional Patagónico, nº. 6: 1 83, CENPAT,
Chubut, Argentina.
Pastor De Ward, C.T. 1994. Nematodes marinos de la ria Deseado (Oncholaimoidea:
Enchelidiidae), Santa Cruz, Argentina. Physis (BsAs), Secc.A, 49(116 117): 27 39.
Pastor de Ward, C.T. 1998. New free living marine nematodos from Deseado river estuary
(Ironoidea: Oxystominidae) Santa Cruz, Argentina. XV. Physis (Buenos Aires), Secc. A,
56: 1-6.
Pastor de Ward, C.T. 2004. Two new species of Hopperia (Nematoda, Comesomatidae)
and Metachromadora (Nematoda, Desmodoridae) from Patagonia, Chubut, Argentina.
Zootaxa, 542: 1-15.
PETROBRAS, 2001. Monitoramento ambiental da atividade de produção de Petróleo na
Bacia de Campos. Etapa de pré-monitoramento- Relatório Final. 222p.
PETROBRAS, 2003 Monitoramento ambiental da atividade de perfuração de Poços de
Petróleo e Gás na Bacia de Campos. Caracterização e Monitoramento Pós - Atividade
Relatório Final. 189p.
Platt, H. M. & R. M.Warwick, 1980. The significance of free-living nematodes to the lit-
toral ecosystem. 729–759. In: J. H. Price, D. E. G. Irvine & W. F. Farnham (eds). The
Shore Environment. Vol. 2. Ecosystems. Academic Press, Cambridge, London.
Platt, H. E. & Warwick, R. M. 1983. Free-living Marine Nematodes. Part I. British Enoplids.
Synopses of the British Fauna (New Series). Cambridge University Press, Cambridge, 307
pp.
Platt, H. M. & Warwick, R. M. 1988. Free-living Marine Nematodes. Part II. British
Chromadorids. Synopses of the British Fauna (New Series). Linnean Society of London
and the Estuarine & Brackish- Water Sciences Association, Brill. 502 pp.
Reinhard, W. 1881 Uber Echinoderes und Desmoscolex der Umgebung von Odessa
Zoologischer Anzeiger, 4: 588-592.
Rice, A. L. & Lambshead, P. J. D. 1994. Patch dynamics in the deep-sea benthos: the role
of a heterogeneous supply of organic matter. Aquatic ecology: scale, pattern and process

192
(eds P. S. Giller, Hildrew, A. G. & D. G. Raffaelli). 469–499. In: 34th Symposium of The
British Ecological Society, Blackwell Scientific Publications, Oxford.
Ruppert, E.E. 1991. Introduction to the aschelminth phyla: A consideration of mesoderm,
body cavities, and cuticle. Chapter 1, pp 1-17. In: F.W. Harrison & E.E. Ruppert (eds.)
Microscopic Anatomy of Invertebrates, vol. 4, Aschelminthes. Wiley-Liss, New York,
438pp.
Schepotieff, A. 1907 Zur Systematik der Nematoideen. Zoologischer Anzeiger, 31: 132-
161.
Schuurmans Stekhoven, J. H. Jr. 1942 The free living nematodes of the Mediterranean.
III. The Balearic Islands Zoölogische Mededeelingen, 23: 229-262.
Schuurmans Stekhoven, J.H. 1942. Les nématodes libres marins du Bassin de Chasse
d’Ostende [The free-living nematods of the Sluice Dock at Ostend]. Med. K. Belgique Institut
Naturelles Wet. 18(12): 1-26.
Soares - Gomes, A; Abreu, C. M. R. C.; Absher, T. M. & Figueiredo, A. G. 1999. Abiotic
features and the abundance of macrozoobenthos of continental margin sediments of East
Brazil. Marine Ecology Progress Series, 127: 113-119.
Soetaert, K.; Vincx, M.; Wittoeck, J.; Tulkens, M. & Van Gansbeke, D. 1994. Spatial pat-
terns of Westerschelde meiobenthos. Estuarine, Coastal and Shelf Science, 39: 367-388.
Soltwedel, T. 1993. Meiobenthos und biogene Sedimentkomponenten im tropischen Ost-
Atlantik. Tese de Doutorado Hamburg University, 178 pp.
Somerfield, P.J.; Cochrane, S.J.; Dahle, S. & Pearson, T.H. 2006. Free-living nematodes
and macrobenthos in a high-latitude glacial fjord. Journal of Experimental Marine Biol-
ogy Ecology, 330: 284-296
Stauffer, H. 1924 Die Lokomotion der Nematoden. Beiträge zur Kausalmorphologie der
Fadenwürmer Zool.Jb. (Syst.), 49 (1-2): 1-118.
Steiner, G. 1916. Neue und wenig bekannte Nematoden von der Westküste Afrikas.
Zoologischer Anzeiger, 47: 322-351
THIRIMCO, 2007 - 13° Congresso Internacional de Meiofauna – Recife, Brasil. 29 de Julho
a 03 de Agosto.

193
Thistle, D. & Sherman, K. 1985. The nematode fauna of a deep-sea site exposed to strong
near bottom currents on the Scotian Rise (Western Atlantic). Deep-Sea Research, 32: 1077–
1088.
Tietjen, J. H. 1971. Ecology and distribution of deep-sea meiobenthos of North Carolina.
Deep-Sea Research, 18: 941-957.
Tietjen, J. H. 1976. Distribution and species diversity of the deep-sea nematodes off North
Carolina. Deep-Sea Research I, 23: 755–768.
Tietjen, J. H. 1984. Distribution and species diversity of deep sea nematodes in the Ven-
ezuela basin. Deep-Sea Research I, 31: 119–132.
Tietjen, J. H. 1989. Ecology of deep-sea nematodes from the Puerto Rico Trench area and
Hatteras Abyssal Plain. Deep-Sea Research I. 36: 1579–1594.
Timm, R. W. 1978 Marine nematodes of the order Desmoscolecida from McMurdo Sound,
Antarctica. In: D. L. Pawson (ed.), Biology of the Antarctic Seas 6. Antarctic Research
Series. 26: 225–236.
Travisi, A. & Vidakovic, J. 1997. Nematofauna in the Adriatic Sea: review and check-list of
free-living nematode species. Helgoländer Meeresuntersuchungen, 51: 503-519
Vanreusel, A. & Vincx, M. 1989. Free-living marine nematodes from the Southern Bight of
the North Sea. III. Species of the Monoposthiidae, Filipjev, 1934. Cahiers De Biologie
Marine, 30: 69-83
Vincx, M. 1989. Seasonal fluctuations and production of nematode communities in the
Belgian coastal zone of the North Sea, In: K. Wouters & L. Baert (eds.). Proceedings of the
Symposium “Invertebrates of Belgium”. pp 57-66.
Vitiello, P. 1976. Peuplement de Nématodes marins des fonds envasés de Provence. Ibid.
52 (2): 283-311.
Vivier, M.H. 1978. Influence d’un déversement industriel profond sur la nématofaune (Can-
yon de cassidaigne, Mediterranée). Tethys, 8: 307-321.
Warwick, R. M.; Platt, H. M. & Sommerfield, P. J. 1998. Free-living Marine Nematodes.
Part III. British Monhysterids. Synopses of the British Fauna (New Series). Shrewsbury:
Field Studies Council. 296 pp.

194
Wieser, W. 1954. Free-living marine nematodes II. Chromadoroidea. Acta Univ. Lund., N.
F., Avd. 2, 50 (16): 148p.





























